Embed Size (px)
Citation preview

RESEARCH ARTICLE
Genetic polymorphisms of methylenetetrahydrofolatereductase (MTHFR) gene Ala222Val and susceptibilityto ovary cancer: a systematic review and meta-analysis
Xinwei Pan & Yan Lu & Ying Long & Desheng Yao
Received: 26 September 2013 /Accepted: 27 September 2013# International Society of Oncology and BioMarkers (ISOBM) 2013
Abstract Many studies have reported the role of methy-lenetetrahydrofolate reductase (MTHFR) gene Ala222Valpolymorphism with ovary cancer risk, but the resultsremained controversial. To derive a more precise estimationof the relationship, a meta-analysis was performed. Oddsratios (ORs) with 95 % confidence intervals (CIs) wereestimated to assess the association between MTHFRAla222Val polymorphism and ovary cancer risk. A totalof 8 studies including 3,723 cases and 4,001 controls werealso involved in this meta-analysis. When all the eligiblestudies were pooled into this meta-analysis, no significantassociation between ovary cancer risk and MTHFRAla222Val polymorphism was found in all genetic models[codominant model: OR=0.980, 95 % CI=0.756–1.270,Ph=0.088, P=0.877; dominant model: OR=1.022, 95 %CI=0.864–1.208, P h=0.033, P =0.803; recessive model:OR = 1.050, 95 % CI = 0.803–1.373, P h = 0.032,P =0.723; allele comparison model: OR=1.028, 95 %CI=0.898–1.178, P h=0.012, P =0.685]. In the stratifiedanalysis by ethnicity, no evidence of any associations ofthis polymorphism with ovary cancer was found in theCaucasian populations. Our meta-analysis supports thatthe MTHFR Ala222Val polymorphism is not contributedto the risk of ovary cancer from currently available evidence.
Keywords MTHFR . Polymorphism . Ovarian cancer .
Meta-analysis
Introduction
Ovarian cancer is one of the most common gynecologicalmalignancies with high mortality, and it is difficult to makean early diagnosis [1, 2]. It is also one of the leading causes ofmalignant deaths in women in the world [3]. Despite thepublic health importance of ovarian cancer, its etiology re-mains unclear [4, 5]. Many studies suggest that the geneticfactors play an important role in the etiology of ovarian cancer[6, 7]. Besides, examination of genetic polymorphisms mayexplain individual differences in the risk of ovarian cancer [8].One-carbon metabolism, also referred as folate-mediated one-carbon metabolism, which plays a critical role in DNA syn-thesis and methylation, has an impact on both genetic andepigenetic pro-carcinogenic processes [9]. Folate plays animportant role as a donor of one-carbon unit for nucleotidesynthesis and DNA methylation [10]. Low folate levels werereported to be associated with uracil disincorporation, chro-mosomal DNA damage, DNA strand breaks, impaired DNArepair, and DNA hypomethylation [11]. Methylenetetra-hydrofolate reductase (MTHFR), which located on the shortarm of chromosome 1 (1p36.3), plays a key role in thefolate metabolism [12]. MTHFR mediates the irreversibleconversion of 5,10-methylenetetrahydrofolate (5,10-MTHF)to 5-methyltetrahydrofolate (5-MTHF) [10]. Several singlenucleotide polymorphisms (SNPs) in the MTHFR genehave been identified. Among which the most commonlystudied are C677T in exon 4 and A1298C in exon7[13–15]. These two polymorphisms were shown to beassociated with reduced enzyme activity, leading to anincrease amount of 5,10-MTHF for thymidylate and purinesynthesis and a decrease amount of 5-MTHF for DNA meth-ylation [14–16].
X. Pan :Y. Lu :Y. Long :D. Yao (*)Department of Gynecologic Oncology, The Affiliated TumorHospital of Guangxi Medical University, Nanning 530021, Chinae-mail: [email protected]
Tumor Biol.DOI 10.1007/s13277-013-1283-0
In recent years, several studies have been conducted toevaluate the association between MTHFR Ala222Val poly-morphisms and ovarian cancer risk, with inconclusive results[17–20]. Therefore, to derive a more precise estimation of theassociation between MTHFR Ala222Val polymorphism andovarian cancer risk, a meta-analysis was performed.
Materials and methods
Publication search
Eligible articles were identified by a search of the MEDLINEbibliographical database for the period up to September 2013
Table 1 Characteristics of the eligible studies in present meta-analysis
Author Year Ethnicity Genotype method Sample size Case Control PWHE
Case Control Ala/Ala Ala/Val Val/Val Ala/Ala Ala/Val Val/Val
Jakubowska et al. [26] 2007 Caucasians PCR-RFLP 146 290 73 56 15 128 134 18 0.03
Wu et al. [27] 2007 Asian PCR-RFLP 81 80 17 40 24 32 35 13 0.52
Terry (NEC) et al. [17] 2010 Caucasians PCR-RFLP 1120 1,160 427 492 140 499 488 138 0.27
Terry (NHS) et al. [17] 2010 Caucasians PCR-RFLP 158 496 71 72 10 210 217 55 0.93
Terry (MAY) et al. [17] 2010 Caucasians PCR-RFLP 364 412 164 167 33 193 168 51 0.13
Webb et al.[19] 2011 Caucasians PCR-RFLP 1638 1278 744 709 185 571 568 139 0.90
Prasad et al. [18] 2011 Caucasians PCR-RFLP 80 125 72 3 5 116 8 1 0.06
Pawlik et al. [29] 2011 Caucasians PCR-RFLP 136 160 67 55 13 63 79 18 0.36
PHWE P value of Hardy–Weinberg equilibrium, PCR-RFLP PCR restriction fragment length polymorphism
NOTE: Weights are from random effects analysis
Overall (I-squared = 61.0%, p = 0.012)
ID
Webb et al
Prasad et al
Terry(NEC) et al
Study
Wu et al
Pawlik et al
Terry(NHS) et al
Jakubowska et al
Terry(MAY) et al
1.08 (0.79, 1.47)
OR (95% CI)
1.02 (0.80, 1.31)
8.06 (0.92, 70.35)
1.19 (0.91, 1.55)
3.48 (1.42, 8.51)
0.68 (0.31, 1.50)
0.54 (0.26, 1.11)
1.46 (0.70, 3.07)
0.76 (0.47, 1.24)
100.00
Weight
22.13
1.88
21.55
%
8.14
9.56
10.64
10.34
15.75
1.08 (0.79, 1.47)
OR (95% CI)
1.02 (0.80, 1.31)
8.06 (0.92, 70.35)
1.19 (0.91, 1.55)
3.48 (1.42, 8.51)
0.68 (0.31, 1.50)
0.54 (0.26, 1.11)
1.46 (0.70, 3.07)
0.76 (0.47, 1.24)
100.00
Weight
22.13
1.88
21.55
%
8.14
9.56
10.64
10.34
15.75
1.0142 1 70.3
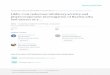
Fig. 1 Forest plot for the association between the MTHFR C677T polymorphism and ovarian cancer risk (codominant model)
Tumor Biol.
(last search: September 30, 2009) using combinations of thefollowing keywords: “ovarian carcinoma,” “ovarian cancer,”“ovarian tumors,” “ovary carcinoma,” “ovary cancer,” “ovarytumors,” “methylenetetrahydrofolate reductase,” “MTHFR,”“polymorphism,” “polymorphisms,” “mutation,” and “muta-tions”, without any restrictions on sample size and language.All the searched studies were retrieved, and their referenceswere checked as well for other relevant articles. Review articleswere also searched to find additional eligible studies. Whenmore than one of the same patient population was included inseveral publications, only the most recent or complete studywas used in this meta-analysis.
Inclusion criteria and exclusion criteria
The following criteria were used to select the eligible studies:(1) case–control studies, (2) evaluation of the MTHFRAla222Val polymorphism and ovarian cancer risk, (3) identifi-cation of ovarian cancer was confirmed histologically or path-ologically, (4) sufficient reported genotypic frequencies in bothcase and control populations for estimating an odds ratio (OR)with a 95 % confidence interval (CI), and (5) the genotype
distribution among the control population was consistent withHardy–Weinberg Equilibrium (HWE). The major reasons forexclusion of studies were (1) case only studies, (2) reviewpapers, and (3) containing overlapping data.
Data extraction
Information was carefully extracted from all eligible publica-tions independently by two investigators according to theinclusion criteria listed above. The following data were col-lected from each study: first author’s surname, year of publi-cation, country, ethnicity, genotyping method, sample size,and numbers of genotype Ala222Val for both cases and con-trols, respectively. Ethnicity was categorized as “Caucasian”and “Asian.” When a study did not state which ethnic groupswere included or if it was impossible to separate participantsaccording to phenotype, the sample was termed as “mixedpopulation.”We did not define a minimum number of patientsfor including a study in our meta-analysis. Disagreementswere resolved by discussion between the two authors. If thesetwo authors could not reach a consensus, another author wasconsulted to resolve the disagreement.
NOTE: Weights are from random effects analysis
Overall (I-squared = 54.2%, p = 0.033)
Pawlik et al
Jakubowska et al
Prasad et al
Terry(NEC) et al
Terry(MAY) et al
Webb et al
Terry(NHS) et al
Wu et al
ID
Study
1.02 (0.86, 1.21)
0.66 (0.41, 1.05)
0.82 (0.55, 1.22)
1.43 (0.53, 3.88)
1.18 (1.00, 1.40)
1.07 (0.81, 1.43)
0.97 (0.84, 1.12)
0.89 (0.62, 1.28)
2.51 (1.25, 5.04)
OR (95% CI)
100.00
8.96
10.76
2.57
21.92
15.67
23.24
12.08
4.80
Weight
%
1.02 (0.86, 1.21)
0.66 (0.41, 1.05)
0.82 (0.55, 1.22)
1.43 (0.53, 3.88)
1.18 (1.00, 1.40)
1.07 (0.81, 1.43)
0.97 (0.84, 1.12)
0.89 (0.62, 1.28)
2.51 (1.25, 5.04)
OR (95% CI)
100.00
8.96
10.76
2.57
21.92
15.67
23.24
12.08
4.80
Weight
%
1.198 1 5.04
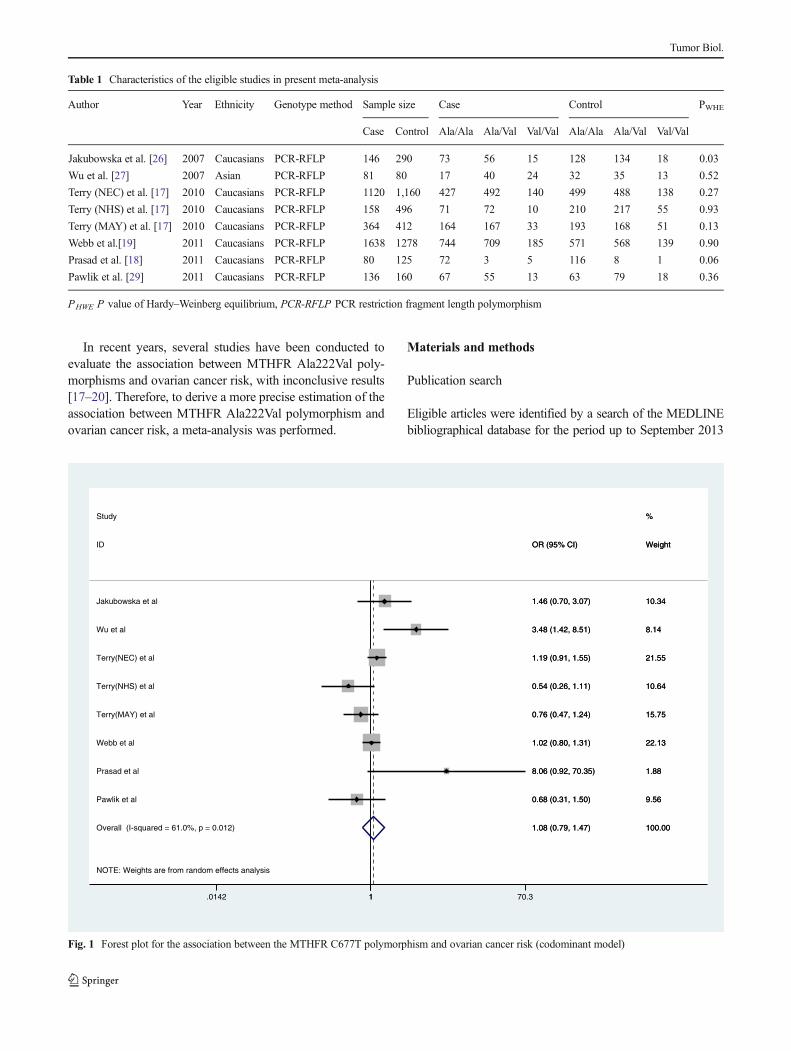
Fig. 2 Forest plot for the association between the MTHFR C677T polymorphism and ovarian cancer risk (dominant model)
Tumor Biol.
Statistical analysis
The strength of the association between MTHFR Ala222Valpolymorphism and risk of ovary cancer was assessed bycalculating the crude ORs with their 95 % CIs. We assessedthe association between MTHFR Ala222Val polymorphismand risk of ovary cancer using the allele model (val-alleleversus Ala-allele), the codominant model (Val/Val versusAla/Ala), the dominant model (Val/Val+Val/Aa versus Ala/Ala), and the recessive model (Val/Val versus Val/Aa+Ala/Ala). Subgroup analyses were performed by ethnicity, and theethnicity was defined mainly as Caucasians and Asians. Inconsideration of the possibility of heterogeneity across thestudies, a statistical test for heterogeneity was performedbased on the Q statistic [21]. If the P value of the Q testwas more than 0.10, it indicated a lack of heterogeneity amongthe included studies. Moreover, the summary OR estimate ofeach study was calculated by the fixed-effects model (theMantel–Haenszel method) [22]. Otherwise, the random-effects model (the DerSimonian and Laird method) was used[23]. The potential publication bias was firstly estimated byvisual inspection of the funnel plot, and the symmetry of the
funnel plot was further assessed using Egger’s linear regres-sion test (P <0.05 as statistically significant) [24]. All statisti-cal tests were performed with the STATA software version12.0 (Stata Corporation, College Station, TX, USA).
Results
Characteristics of included studies
Based on our search criteria, 17 individual records werefound; 8 full-text publications were preliminarily identifiedfor further detailed evaluation after excluding 9 records[25–33]. According to the exclusion criteria, two publicationswere excluded including one for lack of available data [25]and one for case study only [28]. One paper reported threeindividual case–control studies, and the data from this paperwere extracted as three individual case–control studies [29].At last, 8 individual case–control studies with 3,379 cases and4,078 controls were included into this meta-analysis [26, 27,29–32]. The detailed characteristics of these studies are sum-marized in Table 1. The number of cases varied from 80 to 1,
NOTE: Weights are from random effects analysis
Overall (I-squared = 54.3%, p = 0.032)
Webb et al
Terry(NHS) et al
Terry(MAY) et al
Prasad et al
ID
Terry(NEC) et al
Pawlik et al
Study
Jakubowska et al
Wu et al
1.05 (0.80, 1.37)
1.04 (0.83, 1.32)
0.54 (0.27, 1.09)
0.71 (0.44, 1.12)
8.27 (0.95, 72.12)
OR (95% CI)
1.09 (0.85, 1.40)
0.84 (0.40, 1.79)
1.69 (0.83, 3.47)
2.17 (1.01, 4.65)
100.00
23.57
9.72
15.48
1.46
Weight
22.90
8.80
%
9.41
8.66
1.05 (0.80, 1.37)
1.04 (0.83, 1.32)
0.54 (0.27, 1.09)
0.71 (0.44, 1.12)
8.27 (0.95, 72.12)
OR (95% CI)
1.09 (0.85, 1.40)
0.84 (0.40, 1.79)
1.69 (0.83, 3.47)
2.17 (1.01, 4.65)
100.00
23.57
9.72
15.48
1.46
Weight
22.90
8.80
%
9.41
8.66
1.0139 1 72.1
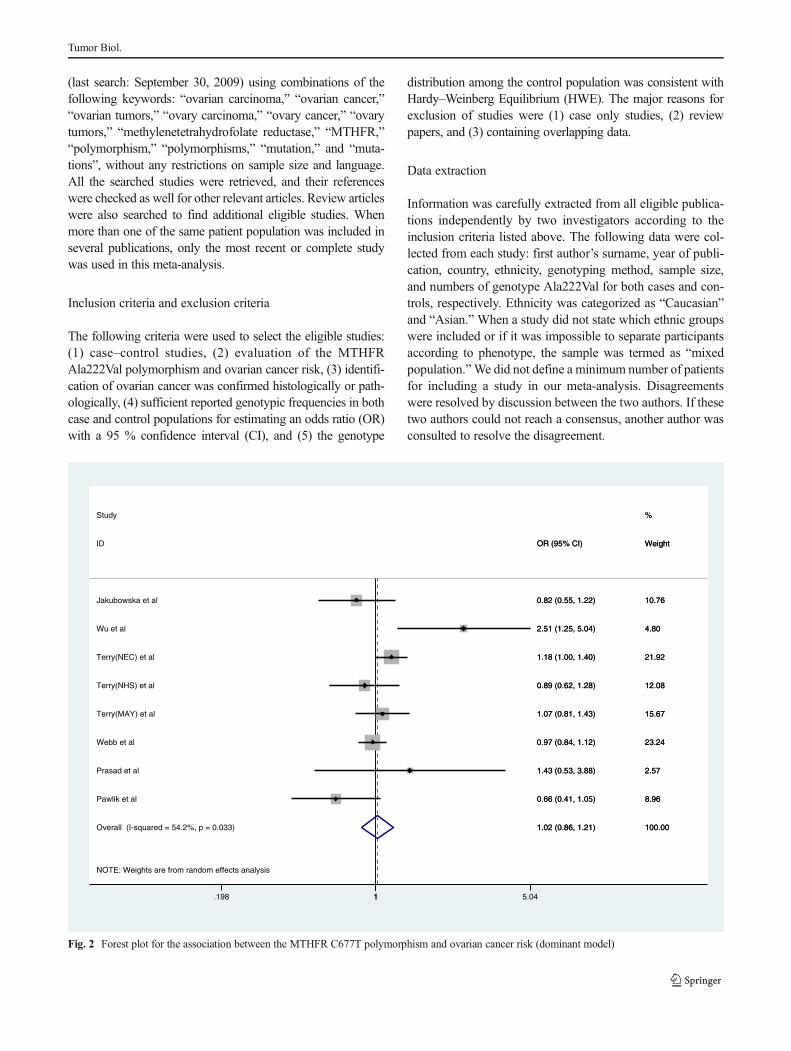
Fig. 3 Forest plot for the association between the MTHFR C677T polymorphism and ovarian cancer risk (recessive model)
Tumor Biol.

363, with a mean of 422, and the number of controls variedfrom 80 to 1,414, with a mean of 510. There were sevenstudies with confirmation of HWE, and one study with depar-tures from HWE.
Quantitative synthesis
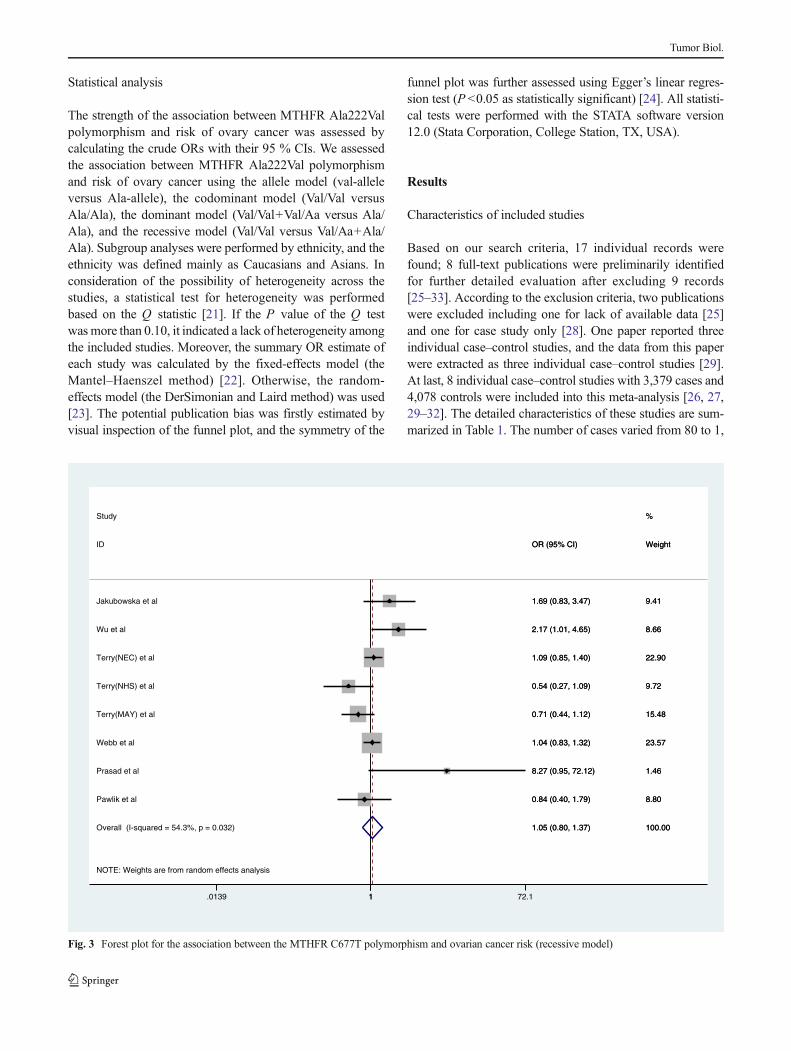
When all the eligible studies were pooled into thismeta-analysis,there was no significant association between ovary cancer riskand MTHFR Ala222Val polymorphism in all genetic models[codominant model: OR=0.980, 95 % CI=0.756–1.270,Ph=0.088, P=0.877, Fig. 1; dominant model: OR=1.022,95 % CI=0.864–1.208, Ph=0.033, P=0.803, Fig. 2; recessivemodel: OR=1.050, 95 % CI=0.803–1.373, P h=0.032,P=0.723, Fig. 3; allele comparison model: OR=1.028, 95 %CI=0.898–1.178, Ph=0.012, P=0.685, Fig. 4]. In subgroupanalyses stratified by ethnicity, MTHFR Ala222Val polymor-phism was also not associated with ovary cancer risk inCaucasian populations under codominant model [OR=2.80,95 % CI=1.09–7.21, Ph=0.87, P=0.03], dominant model[OR=0.994, 95 % CI=0.871–1.134, Ph=0.180, P=0.928],
recessive model [OR=0.980, 95 % CI=0.760–1.265, Ph=0.072,P=0.879], and allele comparison model [OR=0.991, 95 %CI=0.891–1.102, Ph=0.127, P=0.865]. Because there wasonly one case–control study related to the association of thispolymorphism and ovary cancer susceptibility, no impor-tant result can be gained from this meta-analysis. Afterexcluding the studies in which the genotype distributionsdeviated from HWE, all of the results remained stable.
Sensitive analysis
We tested the inclusion criteria of this meta-analysis by asensitivity analysis. Sensitivity analysis was conducted todetermine whether modification of the inclusion criteria ofthis meta-analysis affected the results. A single study involvedin the meta-analysis was deleted each time to reflect theinfluence of individual data. The ORs were calculated bymeans of a random-effects model. The correspondingsynthetic results were not materially altered by omittingeach study, which suggested that the results of the meta-analyses were stable.
NOTE: Weights are from random effects analysis
Overall (I-squared = 60.9%, p = 0.012)
ID
Prasad et al
Terry(NHS) et al
Webb et al
Terry(NEC) et al
Terry(MAY) et al
Jakubowska et al
Pawlik et al
Wu et al
Study
1.03 (0.90, 1.18)
OR (95% CI)
2.12 (0.91, 4.96)
0.84 (0.63, 1.11)
0.99 (0.89, 1.11)
1.12 (0.99, 1.26)
0.97 (0.78, 1.20)
0.98 (0.72, 1.33)
0.76 (0.54, 1.08)
1.93 (1.24, 3.01)
100.00
Weight
2.31
12.25
21.71
20.88
15.58
10.90
9.55
6.81
%
1.03 (0.90, 1.18)
OR (95% CI)
2.12 (0.91, 4.96)
0.84 (0.63, 1.11)
0.99 (0.89, 1.11)
1.12 (0.99, 1.26)
0.97 (0.78, 1.20)
0.98 (0.72, 1.33)
0.76 (0.54, 1.08)
1.93 (1.24, 3.01)
100.00
Weight
2.31
12.25
21.71
20.88
15.58
10.90
9.55
6.81
%
1.201 1 4.96
Fig. 4 Forest plot for the association between the MTHFR C677T polymorphism and ovarian cancer risk (allele comparison model)
Tumor Biol.

Publication bias
Both Begg’s funnel plot and Egger’s test were performed toassess the publication bias of literatures. Begg’s funnel plotsdid not reveal any evidence of obvious asymmetry inMTHFRAla222Val. The Egger’s test results suggested no evidence ofpublication bias in the meta-analysis of MTHFR Ala222Val,indicating that our results were statistically robust.
Discussion
SNP is the most common form of human genetic variation andmay contribute to an individual’s susceptibility to cancer;however, the underlying molecular mechanism is unknown.Previous studies suggested that some variants may affecteither the expression or activity levels of enzymes and there-fore may be mechanistically associated with cancer risk[34–36]. MTHFR is a key enzyme in the folate metabolismpathway. The most commonly found polymorphisms of theMTHFR gene are C677T and A1298C. The C677T polymor-phism which positioned in exon 4 leads to an alanine to valineconversion at codon 222. These polymorphisms, which resultin lowered blood levels of folate, have been linked to a varietyof cancers, including ovarian cancer [26, 37–39]. Due to thedifferent role of MTHFR genetic polymorphism on abnormalDNA methylation and DNA synthesis, it has been hypothe-sized that MTHFR Ala222Val polymorphism is associatedwith risk of ovarian cancer, and many reports have beenpublished but no clear consensus has been reached. This ledus to undertake the present meta-analysis, which could quan-tify the synthesis of all the available data and might help us toexplore a more robust estimate of the role of this polymor-phism with ovarian carcinogenesis.
The present meta-analysis included 2,620 cases and 3,225controls, concerning the MTHFR Ala222Val polymorphismand ovary cancer risk. In this present work, we found that thevariant genotypes of the MTHFR Ala222Val polymorphismwere significantly associated with ovary cancer risk.Simultaneously, the same results were presented in the strati-fied analysis by ethnicity. However, 10 studies involving 3,230 cases and 3,582 controls described Lys751Gln genotypesand were also involved in this meta-analysis. Overall, nosignificant associations were found between this polymor-phism and ovary cancer susceptibility. In the stratified analysisby ethnicity and sources of controls, we got the same resultsamong different descent populations, hospital-based studies,and population-based studies in all genetic models.
Some limitations of this meta-analysis should be acknowl-edged. Firstly, the number of cases and controls in the includ-ed studies was not enough. Secondly, our result was based onunadjusted estimates, while a more precise analysis should beconducted and adjusted by other factors like diet habit,
smoking, drinking status, environmental factors and so on.Thirdly, in the subgroup analyses by ethnicity, a relativelylimited study number made it impossible to perform ethnicsubgroup analysis of Asian populations. Thus, additionalstudies are warranted to evaluate the effect of this functionalpolymorphism on ovary cancer risk in different ethnicities,especially in Asians. In addition, our analysis did not considerthe possibility of gene–gene or SNP–SNP interactions or thepossibility of linkage disequilibrium between polymorphisms.
In summary, this meta-analysis provided evidence that theMTHFR Ala222Val polymorphism contributes to the devel-opment of ovary cancer. In order to verify our findings, largerand well-designed studies are needed to further evaluate theassociation between these two common polymorphisms of theMTHFR gene and ovary cancer risk.
Conflicts of interest None
References
1. Clarke-Pearson DL. Clinical practice. Screening for ovarian cancer.N Engl J Med. 2009;361:170–7.
2. Jemal A, Siegel R, Xu J, et al. Cancer statistics, 2010. CA Cancer JClin. 2010;60:277–300.
3. Jemal A, Siegel R,Ward E, et al. Cancer statistics. CA Cancer J Clin.2008;58:71–96.
4. Cannistra SA. Cancer of the ovary. N Engl J Med. 2004;351:2519–29.5. Pennington KP, Swisher EM. Hereditary ovarian cancer: beyond the
usual suspects. Gynecol Oncol. 2012;124:347–53.6. Diaz-Padilla I, Amir E, Marsh S, et al. Genetic polymorphisms as
predictive and prognostic biomarkers in gynecological cancers: asystematic review. Gynecol Oncol. 2012;124:354–65.
7. Khanra K, Panda K, Bhattacharya C, et al. Association of twopolymorphisms of DNA polymerase Beta in exon-9 and exon-11with ovarian carcinoma in India. Asian Pac J Cancer Prev. 2012;13:1321–4.
8. Bhurgri Y, Shaheen Y, Kayani N, et al. Incidence, trends and mor-phology of ovarian cancer in Karachi (1995–2002). Asian Pac JCancer Prev. 2011;12:1567–71.
9. Stern LL, Mason JB, Selhub J, et al. Genomic DNA hypomethyla-tion, a characteristic of most cancers, is present in peripheral leuko-cytes of individuals who are homozygous for the C677T polymor-phism in the methylenetetrahydrofolate reductase gene. CancerEpidemiol Biomarkers Prev. 2000;9:849–53.
10. Choi SW, Mason JB. Folate and carcinogenesis: an integratedscheme. J Nutr. 2000;130:129–32.
11. Duthie SJ. Folic acid deficiency and cancer: mechanisms of DNAinstability. Br Med Bull. 1999;55:578–92.
12. Goyette P, Pai A, Milos R, et al. Gene structure of human and mousemethylenetetrahydrofolate reductase (MTHFR). Mamm Genome.1998;9:652–6.
13. Angeline T, Jeyaraj N, Granito S, et al. Prevalence of MTHFR genepolymorphisms (C677T and A1298C) among Tamilians. Exp MolPathol. 2004;77:85–8.
14. Weisberg IS, Jacques PF, Selhub J, et al. The 129XA-C polymor-phism in methylenetetrahydrofolate reductase (MTHFR): in vitro ex-pression and association with homocysteine. Atherosclerosis. 2001;156:409–15.
Tumor Biol.

15. Weisberg I, Tran P, Christensen B, et al. A second genetic polymor-phism in methylenetetrahydrofolate reductase (MTHFR) associatedwith decreased enzyme activity. Mol Genet Metab. 1998;64:169–72.
16. Frosst P, Blom HJ, Milos R, et al. A candidate genetic risk factor forvascular disease: a common mutation in methylenete-trahydrofolatereductase. Nat Genet. 1995;10:111–3.
17. Terry KL, Tworoger SS, Goode EL, et al. MTHFR polymorphisms inrelation to ovarian cancer risk. Gynecol Oncol. 2010;119:319–24.
18. Prasad VV, Wilkhoo H. Association of the functional polymorphismC677T in the methylenetetrahydrofolate reductase gene with colo-rectal, thyroid, breast, ovarian, and cervical cancers. Onkologie.2011;34:422–6.
19. Webb PM, Ibiebele TI, Hughes MC, et al. Folate and relatedmicronutrients, folate-metabolising genes and risk of ovarian cancer.Eur J Clin Nutr. 2011;65:1133–40.
20. Pawlik P, Mostowska A, Lianeri M, et al. Folate and choline metab-olism gene variants in relation to ovarian cancer risk in the Polishpopulation. Mol Biol Rep. 2012;39:5553–60.
21. Cochran WG. The combination of estimates from different experi-ments. Biometrics. 1954;10:101–29.
22. Mantel N, Haenszel W. Statistical aspects of the analysis of data fromretrospective studies of disease. J Natl Canc Inst. 1959;22:719–48.
23. DerSimonian R, Laird N.Meta-analysis in clinical trials. Control ClinTrials. 1986;7:177–88.
24. Egger M, Davey Smith G, Schneider M, Minder C. Bias in meta-analysis detected by a simple, graphical test. BMJ. 1997;315:629–34.
25. Gershoni-Baruch R, Dagan E, Israeli D, et al. Association of theC677T polymorphism in the MTHFR gene with breast and/or ovar-ian cancer risk in Jewish women. Eur J Cancer. 2000;36:2313–6.
26. Jakubowska A, Gronwald J, Menkiszak J, et al. Methylenetetrahydro-folate reductase polymorphisms modify BRCA1-associated breast andovarian cancer risks. Breast Cancer Res Treat. 2007;104:299–308.
27. Wu Y, Zhang JY, Zuo WJ. Association between genetic polymor-phisms of methylenetetrahydrofolate reductase C677Tand susceptibil-ity to ovarian cancer. Xian Dai Fu Chan Ke Jin Zhan. 2007;16:811–3.
28. Magnowski P, Seremak-Mrozikiewicz A, Nowak-Markwitz E, et al.No association between MTHFR 677C > T polymorphism and
ovarian cancer risk in BRCA1 mutation carriers in Wielkopolskaregion. Ginekol Pol. 2010;81:506–10.
29. Pawlik P, Mostowska A, Lianeri M, et al. Folate and choline metab-olism gene variants in relation to ovarian cancer risk in the Polishpopulation. Mol Biol Rep. 2011;39:5553–60.
30. Momparler RL, Bovenzi V. DNA methylation and cancer. J CellPhysiol. 2000;183:145–54.
31. Shen H, Wang L, Spitz MR, Hong WK, Mao L, Wei Q. A novelpolymorphismin human cytosine DNA-methyltransferase-3B pro-moter is associated with an increased risk of lung cancer. CancerRes. 2002;62:4992–5.
32. Skoog T et al. A common functional polymorphism (C3A substitu-tion at position-863) in the promoter region of the tumor necrosisfactor (TNF-) gene associated with reduced circulating levels of TNF.Hum Mol Genet. 1999;8:1443–9.
33. Galbiatti AL, Ruiz MT, Maniglia JV, et al. Head and neck cancer:genetic polymorphisms and folate metabolism. Braz JOtorhinolaryngol. 2012;78:132–9.
34. Zhao P, Lin F, Li Z, et al. Folate intake, methylenetetrahydrofolatereductase polymorphisms, and risk of esophageal cancer. Asian Pac JCancer Prev. 2011;12:2019–23.
35. ZhangY, Chen GQ, Ji Y, et al. Quantitative assessment of the effect ofMTHFR polymorphisms on the risk of lung carcinoma. Mol BiolRep. 2012;39:6203–11.
36. Skoog T, et al. A common functional polymorphism (C→A substi-tution at position-863) in the promoter region of the tumor necrosisfactor-alpha (TNF-alpha) gene associated with reduced circulatinglevels of TNF-alpha. Hum Mol Genet. 1999;8:1443–1449.
37. Galbiatti AL, Ruiz MT, Maniglia JV, et al. Head and neckcancer: genetic polymorphisms and folate metabolism. Braz JOtorhinolaryngol. 2012;78:132–139.
38. Zhao P, Lin F, Li Z, et al. Folate intake, methylenetetrahydrofolatereductase polymorphisms, and risk of esophageal cancer. Asian Pac JCancer Prev. 2011;12:2019–2023.
39. ZhangY, Chen GQ, Ji Y, et al. Quantitative assessment of the effect ofMTHFR polymorphisms on the risk of lung carcinoma. Mol BiolRep. 2012;39:6203–6211.
Tumor Biol.