Embed Size (px)
Citation preview

Gene transfer and plant transformation
Why?
Genetic manipulation
Crop improvement
Basic research tool
Approaches
Agrobacterium
Direct DNA transfer
Viral vectors
Chloroplast transformation
“Gene targeting” – site-‐specific integration
Outcomes
Positive
Negative
Biosafety

The “Elephant in the Room”
In order to translate to utility in the field, a plant transformation strategy must include the ability to produce normal, fertile, and productive plants
This in turn requires an ability to manipulate, or regenerate, normal plants
Manipulation – introduction of foreign genes into plants
viral vectors
Regeneration
a matter of tissue culture
very species-‐ and cultivar-‐ dependent
culture-‐free transformation
used with Arabidopsis

Gene transfer strategies
How do we find “transformed” plant cells?
selectable markers
in yeast and Chlamydomonas, one uses nutritional markers to complement auxotrophic phenotypes of laboratory strains (leu, trp, his, ura)
in E. coli, one usually uses antibiotic resistance markers (kan, tet, chl, str, amp)
in plants, one uses antibiotic or herbicide resistance markers (kan or other aminoglycosides, one of several herbicides)
selectable markers used in plants are foreign genes - derived from bacteria, usually; they must be specifically tailored for expression in plant cells

Making gene transfer work: expression vectors as with selectable markers, foreign genes must usually be modified so that they can be expressed in plants
most variation in expression vectors is seen in promoters
constitutive or regulated promoters
regulation may be “natural” - promoters derived from plant genes with desirable expression characteristics
deliberately designed - constitutive or basal promoters may be specifically altered to produce a desired expression characteristic
a variation on this theme - plant promoters can be modified to contain sequences recognized by bacterial or eucaryotic
these systems can be “turned on” with the application of a specific inducer (usually benign)
in order for these to work, the plant must also contain the corresponding transcription factor
Other factors to consider
introns may increase expression of some genes, and in some plants
inadvertent RNA processing signals must be removed from AU-rich coding regions
subcellular localization information may be incorporated into expression systems
VP16 activation
domain
tet repressor DNA-binding
domain
“Top10”
gene of interesttet promoter (x7)
VP16 activation
domain
tet repressor DNA-binding
domainTATA
gene of interesttet promoter (x7)
VP16 activation
domain
tet repressor DNA-binding
domain
TATA
+ tetracycline
no transcription
The problem - inefficient expression of Bt genes inplants
From Diehn et al. (Plant Physiol. 117, 1433-1443, 1998)
Genes used in this study:
Northern blot analysis:
-> low levels of Bt mRNA may be due to aberrantpolyadenylation
(note that CHX treatment often increases the levels of RNAsthat are unstable due to translational defects)
-> the shorter Bt RNAs are truncated from their 3’-ends
RT/PCR mapping ofpoly(A) sites
Transcript mapping (RT/PCR productcorresponds to at least one in vivo site)
-> truncated Bt RNAs end in poly(A), at discreet siteslocated within Bt coding sequences

the BT coding region possesses potential plantpolyadenylation signals
Summary - low levels of Bt mRNA in transgenic plants areat least partly due to premature polyadenylation at siteswithin the Bt coding region
This case illustrates a problem that will be associated withexpression of AT-rich genes in plants
Others have overcome this problem by systematicallychanging the A+T content of Bt coding regions

Agrobacterium-mediated gene transfer
the keys
- to make a segment of DNA that contains a selectable marker and a gene of interest to look like a T-DNA
- to get this “T-DNA” into an Agrobacterium cell so that it can be mobilized by the vir genes
- to produce and find transformed plant cells that can be regenerated into normal, fertile plants
requirements
- a transfer cassette bounded by functioning borders
- ways to get this cassette into Agrobacterium
- disarmed Ti plasmids that retain functional vir genes
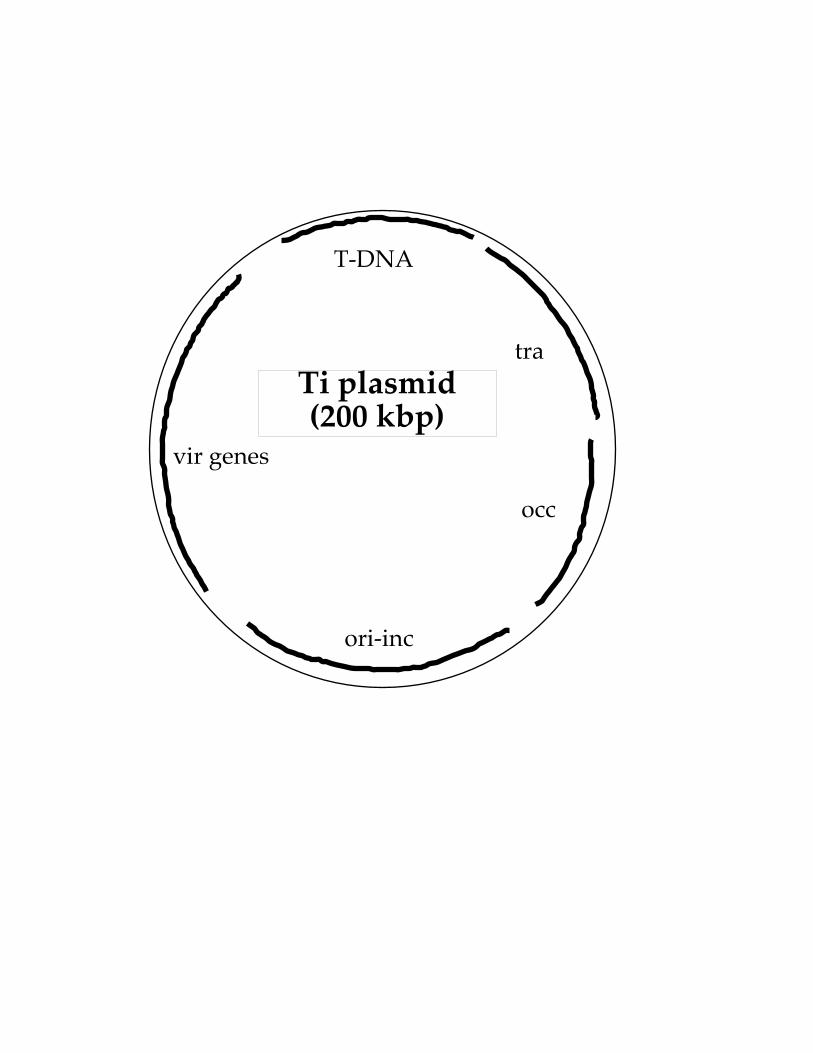
Ti plasmid(200 kbp)
T-DNA
vir genes
ori-inc
occ
tra
VirE2 likely protects T-strands from nucleolytic deg-radation in the plant cell (Yusibov et al., 1994; Rossiet al., 1996). VirF may play a role in stripping proteinsoff the T-strand prior to T-DNA integration (Tzfiraet al., 2004).
Although vir genes were first defined geneticallybecause of their importance in virulence (Koekmanet al., 1979; Garfinkel and Nester, 1980; Holsters et al.,1980; DeGreve et al., 1981; Leemans et al., 1981), nogene within T-DNA is essential for T-DNA transfer.The ability to delete wild-type oncogenes and opinesynthase genes from within T-DNA and replace themwith genes encoding selectable markers and other goihelped initiate the field of plant genetic engineering(Bevan et al., 1983; Fraley et al., 1983; Herrera-Estrellaet al., 1983).
DEVELOPMENT OF BINARY VECTOR SYSTEMS
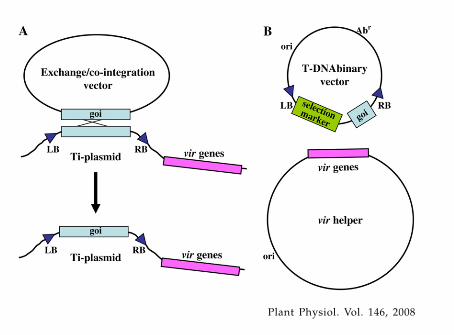
Initial efforts to introduce goi into T-DNA for sub-sequent transfer to plants involved cumbersome ge-netic manipulations to recombine these genes into theT-DNA region of Ti-plasmids (co-integrate or ex-change systems; Garfinkel et al., 1981; Zambryskiet al., 1983; Fraley et al., 1985; Fig. 1A). This was be-cause Ti/Ri-plasmids are very large, low copy numberin Agrobacterium, difficult to isolate and manipulatein vitro, and do not replicate in Escherichia coli, thefavored host for genetic manipulation. T-DNA regionsfrom wild-type Ti-plasmids are generally large and donot contain unique restriction endonuclease sites suit-able for cloning a goi. In addition, scientists wanted toeliminate oncogenes from T-DNA to regenerate nor-mal plants. Opine synthase genes were also generallydeemed superfluous in constructions designed to de-liver goi to plants.
In 1983, two groups made a key conceptual break-through that would allow laboratories that did not
specialize in microbial genetics to use Agrobacteriumfor gene transfer. Hoekema et al. (1983) and deFramond et al. (1983) determined that the vir andT-DNA regions of Ti-plasmids could be split onto twoseparate replicons. As long as both of these repliconsare located within the same Agrobacterium cell, pro-teins encoded by vir genes could act upon T-DNA intrans to mediate its processing and export to the plant.Systems in which T-DNA and vir genes are located onseparate replicons were eventually termed T-DNAbinary systems (Fig. 1B). T-DNA is located on thebinary vector (the non-T-DNA region of this vectorcontaining origin[s] of replication that could functionboth in E. coli and in Agrobacterium tumefaciens, andantibiotic-resistance genes used to select for the pres-ence of the binary vector in bacteria, became known asvector backbone sequences). The replicon containingthe vir genes became known as the vir helper. Strainsharboring this replicon and a T-DNA are considereddisarmed if they do not contain oncogenes that couldbe transferred to a plant.
The utility of binary systems for ease of geneticmanipulation soon became obvious. No longer werecomplex, cumbersome microbial genetic technologiesnecessary to introduce a goi into the T-region of aTi-plasmid. Rather, the goi could easily be clonedinto small T-DNA regions within binary vectors spe-cially suited for this purpose. After characterization andverification of the construction in E. coli, the T-DNAbinary vector could easily be mobilized (by bacterialconjugation or transformation) into an appropriateAgrobacterium strain containing a vir helper region.
Over the past 25 years, both T-DNA binary vectorsand disarmed Agrobacterium strains harboring vir helperplasmids have become more sophisticated and suitedfor specialized purposes. Table I lists many commonlyused T-DNA binary vectors (and vector series). Table IIlists many commonly used disarmed Agrobacterium virhelper strains.
Figure 1. Schematic diagram of co-integration/exchange systems and T-DNA binary vector systemsto introduce genes into plants using Agrobacterium-mediated genetic transformation. A, Co-integration/exchange systems. Genes of interest (goi) are exchangedinto the T-DNA region of a Ti-plasmid (either onco-genic or disarmed) via homologous recombination.Following exchange, the exchange/co-integrationvector can be cured (removed) from the Agrobacte-rium cell; B, T-DNA binary vector systems. Genes ofinterest are maintained within the T-DNA region of abinary vector. Vir proteins encoded by genes on aseparate replicon (vir helper) mediate T-DNA process-ing from the binary vector and T-DNA transfer from thebacterium to the host cell. The selection marker is usedto indicate successful plant transformation. ori, Originof replication; Abr, antibiotic-resistance gene used toselect for the presence of the T-DNA binary vector in E.coli (during the initial stages of gene cassette con-struction) or in Agrobacterium.
Lee and Gelvin
326 Plant Physiol. Vol. 146, 2008

Advantages and disadvantages of Agrobacterium as a gene transfer tool advantages
- technically simple
- yields relatively uncomplicated insertion events (low copy number, minimal rearrangements)
- unlimited size of foreign DNA
- efficient (for most plants)
- adaptable to different cell types, culture procedures (protoplasts, tissue sections, “non-culture” methods)
- transformants are mitotically and meiotically stable
Disadvantages
- host range is limited: not all plants may be susceptible to Agrobacterium
- with susceptible plants, accessible culture/regeneration systems must be adaptable to Agrobacterium-mediated gene transfer

Direct DNA uptake Electroporation
the observation: passing a brief, intense electrical pulse through a suspension of (wall-less) cells in a solution of DNA results in the introduction of significant quantities of DNA into the cell
mechanism: poorly understood
disadvantages:
- can be only used with protoplasts (requires successful regeneration of plants from protoplasts to be used for making transgenic plants)
- frequency of stable transformation is low (0.001 or less)
- in stable transformants, the integrated DNA is extensively rearranged (high copy number, extensive recombination and other alterations); this can lead to some mitotic and meiotic instability
advantages:
- specialized vectors are not needed (expression cassettes can be made in standard E. coli vectors)
- useful for high-efficiency transient expression of foreign genes in plants (for this, no selectable marker is needed)

Coprecipitation
the observation: Ca/DNA coprecipitates, when formed at cell surfaces, can be taken up by cells
in plants, wall-less cells are amenable to coprecipitation; the mechanism of uptake is poorly understood
advantages and disadvantages - as with electroporation

“Biolistics”
the introduction of DNA into cells using microprojectiles
literally, DNA-coated particles are “shot” into target cells
targets may be cells, tissues, whole plants
can be used for stable transformation, transient expression
advantages:
- specialized vectors are not needed (expression cassettes can be made in standard E. coli vectors)
- useful for high-efficiency transient expression of foreign genes in plants (for this, no selectable marker is needed)
- theoretically unlimited “host range” (applicable to all plants)
- may be used with methods that obviate the need for tissue culture/regeneration
disadvantages:
- in stable transformants, the integrated DNA is extensively rearranged (high copy number, extensive recombination and other alterations); this can lead to some mitotic and meiotic instability
- does not completely eliminate the need for tractable tissue culture/regeneration systems
a
T - D N A h o m o l o g o u s r e c o m b i n a t i o n , c o n c a t a m e r f o r m a t i o n , d o u b l e - s t r a n d b r e a k a g e , n u c l e a s e — t r i m m i n g “
d o u b l e - s t r a n d e d p l a s m i d

Viral Vectors Principle – modify viruses to allow expression of foreign genes
The promise of the approach
viruses are natural gene transfer agents (in a sense)
viruses can replicate to high levels
viruses are adapted to utilize host machineries to produce large quantities of viral proteins
Classes of plant viruses:
ssRNA - (+) and (-) sense
dsRNA
ssDNA – ambisense (geminiviruses)
dsDNA - pararetroviruses
[one class of virus that is not known in higher plants - ds DNA viruses that replicate via a semiconservative DNA->DNA means]
July 10, 1996 17:34 Annual Reviews SCHOLT.TXT AR14-16
PLANT VIRUS GENE VECTORS 303
35S promoter of CaMV upstream of the full-length cDNA insert, followed bydirect DNA inoculation, is also being increasingly used for infection of plantswith cloned RNA viruses (6).
Because recent technological advances have circumvented many of the initialdisadvantages that existed for investigations on RNA viruses, the gene vectorpotential can now be tested equally for DNA and RNA viruses. Therefore,the remainder of this review will focus on conceptual strategies (Figure 2),rather than genome composition, for the development of viruses as vehicles forexpression of foreign genes.
BioassaysA major criterion determining whether a virus might be suitable as a genevector is its ability to infect host plants efficiently. This is generally a straight-forward procedure for viruses that can be transmitted mechanically becausetheir genomes are often infectious when derived from appropriately constructedcloned material. However, some viruses are not amenable to mechanical inocu-lation. For example, phloem limited luteoviruses and geminiviruses are recalci-trant to mechanical passage from plant to plant. This disadvantage can be over-come by use of agroinoculation (Figure 1) (28) whereby infectious cloned viralDNA or cDNA is introduced into the host nucleus byAgrobacteriumT-DNA
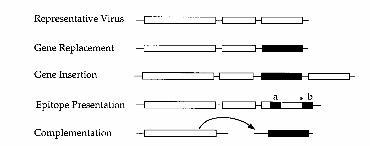
Figure 2 Comparison of strategies used to express foreign genes (black box) from differentviruses as discussed in the text or references therein. White boxes indicate viral genes. The epitopepresentation method (a) involves translational fusion of a small sequence inside the coat proteingene or (b) translational readthrough of an amber stopcodon (∗) at the 3′ end.

July 10, 1996 17:34 Annual Reviews SCHOLT.TXT AR14-16
Annu. Rev. of Phytopathol. 1996. 34:299–323Copyright c© 1996 by Annual Reviews Inc. All rights reserved
PLANT VIRUS GENE VECTORS FORTRANSIENT EXPRESSION OFFOREIGN PROTEINS IN PLANTS
Herman B. Scholthof and Karen-Beth G. ScholthofDepartment of Plant Pathology and Microbiology, Texas A&M University, CollegeStation, Texas 77843
Andrew O. JacksonDepartment of Plant Biology, University of California, Berkeley, California 94720
KEY WORDS: plant biology, biotechnology, gene transfer, autonomous replication, transientexpression, gene replacement, gene insertion, epitope presentation, complemen-tation, foreign gene, reporter gene
ABSTRACT
The development of plant virus gene vectors for expression of foreign genesin plants provides attractive biotechnological tools to complement conventionalbreeding and transgenic methodology. The benefits of virus-based transient RNAand DNA replicons versus transgenic gene expression include rapid and conve-nient engineering coupled with flexibility for expeditious application in variousplant species. These characteristics are especially advantageous when very highlevels of gene expression are desired within a short time, although instability ofthe foreign gene in the viral genome can present some problems. The strategiesthat have been tested for foreign gene expression in various virus-based vectorsinclude gene replacement, gene insertion, epitope presentation, use of virus con-trolled gene expression cassettes, and complementation. Recent reports of theutililization of virus vectors for foreign gene expression in fundamental researchand biotechnology applications are discussed.
INTRODUCTION
History of Gene TransferThe practice of genetically altering plants to improve agronomic traits has beenan integral part of food production throughout the history of agriculture. In the
2990066-4286/96/0901-0299$8.00

Tobamoviruses
type member - tobacco mosaic virus
single stranded RNA viruses
genomic RNA is message-sense RNA [ (+) strand ]
linear genome of about 6000-6500 nt
genomic RNA is capped and has a non-polyadenylated 3' end
3' end of genomic RNA can assume a tRNA-like structure (can beaminoacylated by tRNA charging enzymes and modified by CCA-adding enzymes)
genomic RNA encodes 3-4 open reading frames
all are encoded by the same RNA strand
RNA genome encodes just 3-4 "transcripts", or subgenomic RNAs
Questions:
how are the possible open reading frames expressed (if they are)?
how does the RNA genome replicate?
how does the virus move from cell to cell, from infected touninfected plant?
RNAs
proteins
126K
183K
CP
30K
CP
30K
126K/183K,replication
126K 183K/54K 30K 17K ¨
¨
¨
¨
OASΩ
(+)
RI (ds)
progeny(+)
5' 3'
5' 3'
5' 3'
3' 5'
5' 3'
5' 3'
subgenomic RNAs (+)

Functions of the open reading frames
126/183 kD protein: probable virus-encoded component of thereplicase (RNA-dependent RNA polymerase); mutants are notviable, sequence shows significant homology with otherknown RNA-dependent RNA polymerases, 126 kD protein hascapping activity
30 kD protein: cell-cell movement protein (short-distancemovement); purified protein binds non-specifically to singlestranded nucleic acids, mutants do not move from cell to cell (tsmutants have been identified and characterized, cloned wt 30kD protein can complement movement mutants), protein islocalized to plasmadesmata, protein can change (increase)plasmadesmatal exclusion limit
coat protein: sole protein component of the virus, a determinantof resistance (mutants affect hypersensitive response) and long-distance movement

Gene expression
three "mRNAs", and thus three "promoters"
promoters are recognized by the viral replicase, and mRNAsynthesis is analogous to genome replication (no dsDNAintermediates)
genomic mRNA has 5' and 3' translational enhancers: 5' enhanceris a probable high-affinity internal ribosome binding site; 3'enhancer is functionally analogous to poly(A) tails, works inconjunction with the 5' enhancer (not with 5' cap structures)
different subgenomic mRNAs are expressed at different times inthe infection process (seems to be determined by the"promoter")
Genome replication (and mRNA synthesis)
two step - synthesis of a genome-length (-) strand intermediate,then synthesis of genomic and subgenomic RNAs
3'-tRNA structure is important for replication ("telomeres"?)
host proteins are probably involved
RNAs
proteins
126K
183K
CP
30K
CP
30K
126K/183K,replication
126K 183K/54K 30K 17K ®
®
®
®
OASΩ
(+)
RI (ds)
progeny(+)
5' 3'
5' 3'
5' 3'
3' 5'
5' 3'
5' 3'
subgenomic RNAs (+)

RNAs
proteins
126K
183K
CP
30K
CP
30K
126K/183K,replication
126K 183K/54K 30K 17K ®
®
®
®
OASΩ
RNAs
proteins
126K
183K
CP
30K
CP
30K
126K/183K,replication
126K 183K/54K 30K 17K ®
®
®
®
OASΩ
“wild-type”
gene replacement
gene addition
RNAs
proteins
126K
183K
YFP
30K
YFG
30K
126K/183K,replication
126K 183K/54K 30K YFG ®
®
®
®
OASΩ
Y F
YFP
® YFP
infiltrated area. Moreover, it is known that TMV MP cancomplement cell-to-cell movement of PVX viruses lacking CP(17), and therefore, in mixed infections, MP expressed fromTMV is expected to provide cell-to-cell movement ability to thePVX construct. Observation of mesophyll protoplasts preparedfrom infiltrated areas revealed that coinfection of TMV withCP-less PVX provided coexpression of GFP and DsRED in only67% of the cells. To increase this value, viral vectors werecoinfiltrated with a construct expressing PVX CP from the 35Spromoter (pICH22066 construct). With this combination, coex-pression of TMV and PVX increased to 82%. This combina-tion, which provides a high level of expression of both genes andhigh level of coexpression, was used for the rest of this study.
Coexpression of IgG HC and LC by Using TMV and PVX Vectors. Thenoncompeting vector technology was then applied for expressionof immunoglobulins, which are natural tetramers composed oftwo identical HCs and two identical LCs. The human tumor-specific mAb A5, which belongs to the IgG1 subclass, was chosen
as a first candidate. The HC- and LC-coding sequences (includ-ing their native human endoplasmic reticulum-targeting signalpeptides) were subcloned into 3 TMV and 3 PVX provectorconstructs. The resulting constructs were agroinfiltrated into N.benthamiana leaves, together with the corresponding 5 provec-tor modules, a source of recombinase, and a construct forexpression of PVX CP (a total of six constructs, including two 5provector modules and two 3 provector modules). Two differentcombinations were tested: TMV-HC PVX-LC and PVX-HC TMV-LC. As a control, HC and LC were expressed separatelyby using TMV vectors. In all cases, slight toxicity symptomsappeared in infected leaves at 5–6 dpi and progressed further.Toxicity symptoms were stronger for the combination PVX-HC TMV-LC.
Expression of the HC and LC was analyzed by SDSPAGErun under reducing conditions followed by Coomassie bluestaining or Western blotting probed with HC- and LC-specificantibodies. Accumulation of the HCs and LCs was analyzed from3 to 11 dpi. For the PVX-HC TMV-LC combination, accu-mulation of the LC reached a maximum at 4 dpi and remainedstable until 11 dpi (Fig. 3A). The level of HC increased until 5–6dpi and remained relatively stable until 11 dpi. For the reversecombination, TMV-HC PVX-LC, accumulation of the LC wasslower, but it increased continuously until 10–11 dpi (Fig. 3B).Expression of the HC showed similar kinetics. At 11 dpi, and forboth combinations, tissue in expressing areas displayed signifi-cant necrotic symptoms. However, the level of expression of bothchains remained high, suggesting their relative stability. For bothcombinations, the LC accumulated at a substantially higher levelcompared with the HC. The difference was stronger for thePVX-HC TMV-LC combination.
HCs and LCs Coexpressed with TMV and PVX Vectors Form AssembledAntibodies with Specific Antigen-Binding Activity. To test whetherthe HC and LC assembled into functional monoclonal antibod-
Fig. 1. Plasmid constructs. (A) TMV-based provectors: 5 module pICH17388;3 module cloning vectors pICH10990 and pICH11599 (differ in the structure ofthe multicloning site); and 3 modules for expression of DsRED (pICH10580),GFP (pICH7410), A5 LC (pICH21910), and A5 HC (pICH21920). (B) PVX-basedprovectors: 5 module pICH21380; and 3 modules for expression of GFP [coatprotein (CP)-containing vector pICH21470, CP-less vector pICH21282], A5 LC(pICH22240), and A5 HC (pICH22250). (C) Integrase module pICH14011 andconstruct for expression of the PVX CP (pICH22066). (D) Constructs for expres-sion of the A5 LC (pICH23660) and the A5 HC (pICH23670) under control of the35S promoter. LB and RB, binary left and right borders, respectively; Pact,Arabidopsis actin 2 promoter; P35S, 35S promoter; Phsp, Arabidopsis heatshock protein hsp 81.1 promoter; T, nos terminator; RdRp, RNA-dependentRNA polymerase; MP, movement protein; int, intron; AttP and AttB, recom-bination sites; MCS, multicloning site; 3TMV and 3PVX, 3 untranslatedregions of TMV and PVX, respectively; SP, signal peptide; NLS, nuclear local-ization signal.
Fig. 2. Coexpression of GFP and DsRED in N. benthamiana leaves by usingviral vectors (6 dpi). Leaf sectors infected with a mixture of two TMV constructsexpressing GFP or DsRED (A) or coinfiltrated with a PVX vector expressing GFPand a TMV vector expressing DsRED (B) are viewed under the UV light of ahand-held lamp. Protoplasts prepared from leaf areas coinfiltrated with TMVconstructs (C and D) or PVX and TMV constructs (E and F) are viewed under blue(C and E) or red (D and F) light.
14702 www.pnas.orgcgidoi10.1073pnas.0606631103 Giritch et al.

Rapid high-yield expression of full-size IgG antibodiesin plants coinfected with noncompeting viral vectorsAnatoli Giritch*, Sylvestre Marillonnet*, Carola Engler*, Gerben van Eldik†, Johan Botterman†, Victor Klimyuk*,and Yuri Gleba*‡
*Icon Genetics GmbH, Weinbergweg 22, 06120 Halle, Germany; and †Bayer BioScience N.V., Technologiepark 38, B-9052 Gent, Belgium
Communicated by Charles J. Arntzen, Arizona State University, Tempe, AZ, August 2, 2006 (received for review May 24, 2006)
Plant viral vectors allow expression of heterologous proteins athigh yields, but so far, they have been unable to express hetero-oligomeric proteins efficiently. We describe here a rapid andindefinitely scalable process for high-level expression of functionalfull-size mAbs of the IgG class in plants. The process relies onsynchronous coinfection and coreplication of two viral vectors,each expressing a separate antibody chain. The two vectors arederived from two different plant viruses that were found to benoncompeting. Unlike vectors derived from the same virus, non-competing vectors effectively coexpress the heavy and light chainsin the same cell throughout the plant body, resulting in yields ofup to 0.5 g of assembled mAbs per kg of fresh-leaf biomass. Thistechnology allows production of gram quantities of mAbs forresearch purposes in just several days, and the same protocol canbe used on an industrial scale in situations requiring rapid re-sponse, such as pandemic or terrorism events.
monoclonal antibody potato virus X tobacco mosaic virus
A lthough the ability of plants to express full-size humanantibodies was discovered 17 years ago (1–3), the idea of
industrial-scale antibody production in plants has been aban-doned by most companies, mostly because of limitations ofexisting expression protocols. Stably transformed (transgenic)plants are able to express correctly folded and functional anti-bodies of both the IgG and IgA classes, but yields are generallyvery low (usually in the range of 1–40 gg of fresh biomass); inaddition, the time necessary to generate the first grams ofresearch antibody material is very long, requiring 2 years(4–8).
Transient expression systems, on the other hand, allow pro-duction of research quantities of antibody material much faster.However, the early versions of transfection systems, such asAgrobacterium-mediated transient expression or viral vector-mediated expression, cannot provide for high-level coexpressionof two or several polypeptides necessary for the assembly ofheterooligomeric proteins, in particular IgG antibodies (9–12).
Recently, we have developed a scalable transient expressiontechnology (magnifection) that is based on replication of viralvectors delivered to multiple parts of a plant body (systemicdelivery) by Agrobacterium (13). Such a technology is in essencean en masse infiltration of whole, mature plants with a dilutedagrobacteria suspension carrying T-DNAs encoding viral repli-cons. The magnifection process allows expression of variousproteins, but, until now, it has been used to express onlysingle-polypeptide proteins or homooligomers (14). Attempts toexpress two or more different polypeptides from one viralreplicon failed because of drastically reduced expression levelsobtained with bicistronic constructs (unpublished results).
Therefore, we decided to explore expression protocols thatinvolve two or more viral replicons. We report here a generalsolution for coexpression of high amounts of two heterologouspolypeptides by using two different viral vectors, each expressinga separate polypeptide. The vectors described here are built onthe backbones of two noncompeting viruses: tobacco mosaicvirus (TMV) and potato virus X (PVX). This expression tech-
nology leads to yields of assembled full-size monoclonal antibodyat levels as high as 0.5 g of mAb per kg of fresh leaf biomass (oneto two orders of magnitude higher than other transient expres-sion systems). The molecules produced are fully functional, andthe first gram of material can be produced in 2 weeks afterinfiltration. Because the ratio of heavy chain (HC) to light chain(LC) expression and other expressionprocessing parametershave not been fully optimized, the protocol likely has thepotential for further yield improvement.
ResultsTwo TMV-Based Replicons Expressing Different Genes Segregate EarlyDuring Cell-to-Cell Movement. Expression of heterooligomericproteins requires expression of two (or more) different proteinswithin one cell. One approach to achieving this goal wouldconsist of using two TMV-based viral vectors, each one express-ing a different protein subunit. To test this strategy, viral vectorsexpressing GFP and red fluorescent protein from Discosoma(DsRED) were inoculated into Nicotiana benthamiana leaves byusing Agrobacterium-mediated delivery (constructs shown in Fig.1). Six days postinfiltration (dpi), infiltrated leaf areas exhibiteda mosaic of either GFP- or DsRED-expressing sectors, suggest-ing early segregation and spatial separation of distinct TMVpopulations and, therefore, a lack of coexpression (Fig. 2A).Confirming this observation, only 4–5% of protoplasts preparedfrom infected leaf tissue coexpressed both reporter genes (Fig.2 C and D).
TMV and PVX Are Able to Replicate Within the Same Cell with HighEfficiency. To test whether viral vectors derived from TMV andPVX could be used to coexpress two proteins of interest, N.benthamiana leaves were coinoculated with a mixture ofagrobacteria carrying a DsRED-containing TMV vector or aGFP-containing PVX vector. At 6 dpi, the infiltrated areasdisplayed a uniform pattern of yellow fluorescence, indicatingthat both genes were coexpressed in a majority of cells (Fig. 2B).Observation of mesophyll protoplasts prepared from the infil-trated areas revealed that GFP and DsRED were coexpressed in95% of the cells (Fig. 2 D and F).
We also made PVX vectors lacking CP, and we found that, aswith TMV vectors, removal of the CP leads to higher level ofexpression of the protein of interest in the infiltrated leaf (resultsnot shown). Because PVX vectors lacking CP are unable to movefrom cell to cell (15, 16), they were inoculated at a high densityof agrobacterial cells (a 101 dilution relative to a saturatedovernight culture) to transfect a majority of plant cells in the
Conflict of interest statement: No conflicts declared.
Freely available online through the PNAS open access option.
Abbreviations: CP, coat protein; dpi, days postinfiltration; DsRED, red fluorescent proteinfrom Discosoma; HC, heavy chain; LC, light chain; MP, movement protein; PVX, potato virusX; RdRp, RNA-dependent RNA polymerase; TMV, tobacco mosaic virus.
See Commentary on page 14645.
‡To whom correspondence should be addressed. E-mail: [email protected].
© 2006 by The National Academy of Sciences of the USA
www.pnas.orgcgidoi10.1073pnas.0606631103 PNAS October 3, 2006 vol. 103 no. 40 14701–14706
APP
LIED
BIO
LOG
ICA
LSC
IEN
CES
SEE
COM
MEN
TARY

Delayed treatment of Ebola virus infection withplant-derived monoclonal antibodies providesprotection in rhesus macaquesGene Garrard Olinger, Jr.a,1,2, James Pettitta,1, Do Kimb, Cara Workingc, Ognian Bohorovb, Barry Bratcherc, Ernie Hiattc,Steven D. Humec, Ashley K. Johnsonc, Josh Mortonc, Michael Paulyb, Kevin J. Whaleyb, Calli M. Leara, Julia E. Bigginsa,Corinne Scullya, Lisa Hensleya,3, and Larry Zeitlinb,2
aDivision of Virology, United States Army Medical Research Institute of Infectious Diseases, Frederick, MD 21702; bMapp Biopharmaceutical, Inc., San Diego,CA 92121; and cKentucky BioProcessing, LLC, Owensboro, KY 42301
Edited by Charles J. Arntzen, Arizona State University, Tempe, AZ, and approved September 14, 2012 (received for review August 7, 2012)
Filovirus infections can cause a severe and often fatal disease inhumans and nonhuman primates, including great apes. Here, threeanti-Ebola virus mouse/human chimeric mAbs (c13C6, h-13F6, andc6D8) were produced in Chinese hamster ovary and in whole plant(Nicotianabenthamiana) cells. In pilot experiments testing amixtureof the three mAbs (MB-003), we found that MB-003 produced inbothmanufacturing systems protected rhesusmacaques from lethalchallenge when administered 1 h postinfection. In a pivotal follow-up experiment, we found significant protection (P< 0.05)whenMB-003 treatment began24or 48h postinfection (four of six survived vs.zero of two controls). In all experiments, surviving animals that re-ceived MB-003 experienced little to no viremia and had few, if any,of the clinical symptoms observed in the controls. The results repre-sent successful postexposure in vivo efficacy by a mAb mixture andsuggest that this immunoprotectant should be further pursued asa postexposure and potential therapeutic for Ebola virus exposure.
passive immunization | therapy
For more than 35 y, a therapy has been sought to treat thesevere lethal disease caused by Ebola virus (EBOV; family
Filoviridae), which are among the most virulent infectious agentsknown, causing acute and frequently fatal hemorrhagic fever inhumans and nonhuman primates (NHP) (1, 2). Outbreaks inhumans occur intermittently, causing localized high morbidity andmortality. Due to its infectiousness, the lack of approved diag-nostics, and the rapidity of modern travel, the potential exists forany outbreak to become an international epidemic. Currently thereare no licensed vaccines or treatments against EBOV infection.Candidate postexposure interventions in advanced developmentinclude siRNA and phosphorodiamidate morpholino oligomers(PMOs) antisense strategies, as well as postexposure immunizationwith a vesicular stomatitis virus (VSV)-based vaccine (3–5). All ofthese interventions have demonstrated reduced mortality in NHPswhen delivered 0.5–1 h postinfection (p.i.), but clinical and some-times severe disease is observed in survivors.Although the majority of the 20 or more mAb products ap-
proved by the FDA are for noninfectious disease indications(6), passive immunization with antibodies has been an effectivemethod to prevent a variety of viral, bacterial, fungal, and parasiticdiseases that cause disease in humans and animals (7, 8). However,the use of antibody therapy for EBOV has been fraught withconflicting data as to the utility of this class of intervention. Al-though convalescent-phase serum has been used in small numbersfor sporadic outbreaks of EBOV and other filovirus infections (9),it is unclear if these treatments were beneficial. Early proof-of-concept efforts with equine and ovid hyperimmune serum pro-tected baboons from a low challenge (<30 LD50 and <1 LD50,respectively) when treatment was initiated less than 1 h p.i (10–12).However, later efforts with macaque models of EBOV diseaseusing a more robust challenge were unsuccessful (13). Somesuccesses with antibodies in rodents (14–16) have been described,but these results are tempered by the lack of translation to themore robust and lethal macaque models of EBOV infection (17).For example, a potent neutralizing human IgG1 mAb, KZ52,
protected guinea pigs but provided no protection in macaques(18, 19). Recently, studies have demonstrated a role for poly-clonal IgG in protection in the macaque model of disease;transfer of concentrated IgG with a high neutralizing titerpurified from convalescent-phase EBOV-exposed rhesus mac-aques demonstrated protection in naïve animals that receivedthe IgG up to 48 h p.i (20). However, the use of purified conva-lescent macaque serum as a medical countermeasure is likely cost-prohibitive and would have significant logistical and regulatoryhurdles. As an approach to develop a mAb-based medical coun-termeasure, Marzi et al. (18) demonstrated that two neutralizingmouse/human chimeric mAbs against EBOV could provide limitedprotection in rhesus macaques (one of three animals survived)when dosing was initiated 24 h before challenge (1,000 pfu ∼1,000LD50). More recently, Qiu et al. (21) found a mixture of neutral-izing murine mAbs could protect cynomolgus macaques whengiven 1 (four of four survived) or 2 d p.i. (two of four survived).Development of the mAb-based EBOV immunoprotectant
MB-003 was built upon prior work characterizing three mousemAbs (13C6, 13F6, and 6D8) directed against three distinct,nonoverlapping EBOV glycoprotein (GP) epitopes (14). Onlyone of these mAbs (13C6) binds to secreted GP (sGP), which hasbeen speculated to act as a decoy for protective antibodies (22).Two of these mAbs (13C6 and 6D8) neutralize virus in thepresence of complement, and one has no neutralizing activity(13F6). These mAbs were shown to individually protect againstlethal challenge prophylactically in mouse models of EBOVinfection and have a therapeutic window of at least 48 h after viralexposure (14). To develop a product that would be appropriatefor human use, the murine mAbs were deimmunized (23) and/orchimerized with human constant regions, yielding c13C6, h-13F6,and c6D8. To evaluate a more cost-effective and scalable alter-native to production in Chinese hamster ovary (CHO) cells, themAbs were produced in a Nicotiana benthamiana-based rapid an-tibody manufacturing platform (RAMP) using magnICON (ICONGenetics) deconstructed viral vectors (24). The RAMP systemallows rapid, scalable production of mAbs in less than a month,and has been used to produce mAbs under cGMP conditions(25). Via the use of a transgenic strain ofN. benthamiana in whichplant-specific glycosyltransferases (α1,3 fucosyltransferase andβ1,2 xylosyltransferase) are inhibited by RNAi (26), the RAMP-
Author contributions: G.G.O., B.B., E.H., S.D.H., A.K.J., J.M., M.P., K.J.W., C.M.L., J.E.B., C.S.,L.H., and L.Z. designed research; J.P., D.K., C.W., O.B., E.H., S.D.H., A.K.J., J.M., M.P., andL.Z. performed research; G.G.O., J.P., O.B., B.B., E.H., S.D.H., A.K.J., J.M., M.P., C.M.L., J.E.B.,C.S., L.H., and L.Z. analyzed data; and G.G.O., J.P., D.K., C.W., O.B., B.B., E.H., S.D.H., A.K.J.,J.M., M.P., K.J.W., C.M.L., J.E.B., C.S., L.H., and L.Z. wrote the paper.
Conflict of interest statement: K.J.W. and L.Z. are owners of Mapp Biopharmaceutical, Inc.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.1G.G.O. and J.D.P contributed equally to this work.2To whom correspondence may be addressed. E-mail: [email protected] or [email protected].
3Present address: Office of the Chief Scientist, US Food and Drug Administration, MedicalScience Countermeasures Initiative, Silver Spring, MD 20993.
18030–18035 | PNAS | October 30, 2012 | vol. 109 | no. 44 www.pnas.org/cgi/doi/10.1073/pnas.1213709109
Adap%ng picorna-‐type viruses for use as vectors
² Picorna-‐type viruses encode polyproteins that are processed by site-‐specific proteases (think TEV protease)
² It is possible to flank foreign genes with protease-‐recogni%on sites and incorporate them into infec%ous viral genomes





























