Embed Size (px)
Citation preview

J. Comp. Path. 2011, Vol. 145, 68e76 Available online at www.sciencedirect.com
Cor
002
doi
www.elsevier.com/locate/jcpa
Gastritis in Alaskan Racing Sled Dogs
J. W. Ritchey*, M. S. Davis†, M. A. Breshears*, M. D. Willard‡,K. K. Williamson†, C. M. Royer†, M. E. Paytonx and A. S. Cragun*
*Department of Veterinary Pathobiology, †Department of Physiological Sciences, College of Veterinary Medicine, Oklahoma
State University, Stillwater, OK 74078, ‡Department of Small Animal Clinical Sciences, College of Veterinary Medicine,
Texas A&M University, College Station, TX 77843 and xDepartment of Statistics, College of Arts & Sciences,
Oklahoma State University, USA
resp
1-99
:10.1
Summary
Alaskan racing sled dogs are a well-established model of exercise-induced gastric disease. The aim of this studywas to define the temporal development of microscopical gastric lesions during long distance racing. Twogroups of dogs were examined: group I comprised conditioned dogs that were exercising and group II wereconditioned dogs not exercising. The gastric mucosa was examined endoscopically and sampled for routine his-topathology and microscopical scoring, immunohistochemistry (IHC) and detection of apoptotic epithelialcells. Overall, group I dogs exhibited more significant epithelial lesions, including ulcers, compared withdogs in group II. Group II dogs exhibited the most severe mucosal inflammatory infiltrates. Although theintensity of inflammation differed, the nature of the inflammation was similar between groups, consisting ofdiffuse lymphocytic infiltration and a unique interface-type infiltrate that obscured the basement membranezone and was accompanied by intraepithelial infiltration of lymphocytes. IHC confirmed the presence ofCD3+ T and CD79+ B lymphocytes within the mucosal infiltrates; however, most of the intraepithelial andinterface infiltrates were CD3+ T cells. Spiral-shaped bacterial organisms were seen in the gastric tissues; how-ever, their presence did not correlate with either the severity of epithelial lesions, inflammation or the pattern ofinterface inflammation. The number of apoptotic epithelial cells was widely variable and not significantly dif-ferent between groups. These findings confirm previous observations that gastric ulcers develop in conditioneddogs under racing stress. The unique nature of the interface-type gastric inflammation is similar to that of hu-man lymphocytic gastritis and may suggest an immune-mediated mechanism for the changes seen in Alaskanracing sled dogs.
� 2010 Elsevier Ltd. All rights reserved.
Keywords: dog; exercise stress; inflammation; stomach
Introduction
Strenuous exercise has been associated with gastricdisease in man (Moses, 1990; Choi et al., 2001),horses (Murray et al., 1996; Vatistas et al., 1999) anddogs (Davis et al., 2003a); however, the mechanismof exercise-induced gastric disease is unknown.Various causes have been postulated, including tran-sient splanchnic ischaemia due to redistribution ofblood flow during exercise, impact trauma from con-cussion during running and a physiological stress re-sponse, either through direct action of circulatingglucocorticoids or indirectly through immunological
ondence to: J.W. Ritchey (e-mail: [email protected]).
75/$ - see front matter
016/j.jcpa.2010.11.008
alterations and proliferation of pathogenic bacteria.However, none of these hypotheses have been eitherconfirmed or refuted.
Alaskan racing sled dogs appear useful for studyingthe pathogenesis of exercise-induced gastric disease.Sustained strenuous exercise in Alaskan racing sleddogs has been associated with morphological andphysiological changes in the gastric mucosa (Daviset al., 2003a, 2005, 2006; Royer et al., 2005). Gastriculcers and haemorrhages can be demonstrated in48e60% of dogs performing multiday exercise(Davis et al., 2003a,b). These gastric lesions areusually subclinical and have not been correlatedwith a negative effect on performance. Surprisingly,the prevalence and severity of the gastric lesions
� 2010 Elsevier Ltd. All rights reserved.

Gastritis in Alaskan Racing Sled Dogs 69
have also been unrelated to the distance run orduration of exercise. Gastric lesions occur within thefirst day of sustained exercise; however, there is noevidence that the lesions become more severe overthe course of that challenge (Davis et al., 2006). A pre-liminary survey of histological lesions in gastric biop-sies of dogs completing a 1,000-mile race reportedspiral-shaped bacteria in many sections, but foundno association between the presence of the bacteriaand the severity of endoscopic lesions (Davis et al.,2003a). Microscopical lesions were also noted in thegastric biopsies collected from the preliminary study;however, detailed characterization of the lesions andcorrelation with either endoscopic findings or thepresence of spiral-shaped bacteria were beyond thescope of that report.
The aim of the present study was to characterizethe microscopical appearance of the gastric mucosain Alaskan racing sled dogs at periods of rest and dur-ing the development of exercise-induced gastric dis-ease.
Table 1
Summary of microscopical evaluation*
Variable Score
Lesion 1
Surface epithelial lesions 0 None
1 Mild attenuation, vacuolation
2 Marked attenuation,degeneration, discrete ulcers
3 Marked ulceration with exudation
Lesion 2
Inflammation 0 No inflammation (normal)
1 Mild infiltration
2 Moderate infiltration
3 Severe infiltration
Lesion 3
Interface inflammatory lesion 0 Not present
1 Present
Lesion 4
Helical bacterial colonies 0 Not present
1 Present
*Adapted from Day et al., 2008.
Materials and Methods
Dogs and Experimental Procedures
Young adult (48.2� 22 months of age), intact male(n¼ 24) and female (n¼ 16) Alaskan racing sleddogs were used. The Oklahoma State University In-stitutional Animal Care and Use Committee ap-proved all procedures according to the principlesoutlined in the National Institutes of Health (NIH)Guide for the Care and Use of Laboratory Animals.Informed consent was obtained from the owners ofthe dogs before the start of the studies.
Forty endurance-trained racing sled dogs were as-signed to either endurance-conditioned and exercised(group I, n¼ 34) or endurance-conditioned and res-ted control (group II, n¼ 6) groups. All 34 group Idogs rested for at least 3 days prior to the start ofthe study. Exercise consisted of teams of 12e18 dogspulling a lightly-laden sled and musher overa 50-mile groomed-track located in Cantwell, Alaska,USA. Teams traveled the 50 miles in approximately6 h, were fed and rested for 6 h, and then repeatedthe 50-mile run in approximately 6 h. At the conclu-sion of each 100-mile segment, six dogs were removedfrom the exercise group for further examination. Theremaining dogs were fed and rested for 6 h and thenresumed running the next 100-mile segment. Thesix group II dogs received food and water at intervalssimilar to the exercised dogs, but did not participatein the racing exercise. At the end of the fifth100-mile segment, all remaining exercised dogs(n¼ 10) and the control dogs were examined.
Dogs sequentially removed from the exercise groupat 100-mile intervals and all remaining exercised orrested dogs (group II) at the end of 500 miles were al-lowed to eat and drink ad libitum for 12 h and werethen fasted for 12 h. Dogs were anaesthetized and en-doscopic examination was performed as previouslydescribed (Davis et al., 2003a). Results of gastroscopyof the dogs in this study have been previously reported(Davis et al., 2006). Gastric biopsies (n¼ 3e4) wereobtained from endoscopically normal areas of the gas-tric body and were fixed in 10% neutral buffered for-malin. Following endoscopic examination, the dogsrecovered uneventfully.
Histopathology, Immunohistochemistry and TdT Labelling
Formalin-fixed gastric biopsy tissues were embeddedin paraffin wax, sectioned (5 mm) and stained withhaematoxylin and eosin (HE). Examination was per-formed independently by two board-certified veteri-nary pathologists blinded to treatment groups. Thebiopsies were scored usingmodification of a histopath-ological standardization scheme for endoscopic bi-opsy samples (Day et al., 2008). The standardizationscheme provides pictorial templates for normal gas-tric mucosa and mucosa exhibiting mild, moderateor severe inflammatory and epithelial lesions. Eachtype of lesion (i.e. inflammatory or epithelial) wasgiven an individual score that correlated with a rangeof normal to severe lesions (lesions 1 and 2, Table 1).In addition, the biopsies were evaluated for a uniqueinterface pattern of infiltrating inflammatory cells (ei-ther present or absent, lesion 3, Table 1) and for the

70 J.W. Ritchey et al.
presence of helical bacteria within the biopsy sample(lesion 4, Table 1). For each of the four gastric lesions,the lesion scores were compared statistically betweeneach of the two groups of dogs. In addition, the scorefor lesion 4 was evaluated statistically for correlationbetween bacterial infection and the severity of epithe-lial lesions (lesion 1), inflammation (lesion 2) or infil-trating pattern (lesion 3).
Immunohistochemistry (IHC) was performed todetermine the phenotype of infiltrating lymphocytes.Formalin-fixed, paraffin wax-embedded tissue sam-ples were dewaxed and rehydrated before epitope re-trieval (citrate buffer, pH 6.0). Endogenousperoxidase activity was blocked with 3% H2O2 andnon-specific immunoreactivity blocked with 10%normal goat serum. Labelling for CD3 proceededwith application of polyclonal rabbit anti-humanCD3 (A0452, DakoCytomation, Carpinteria, Califor-nia) followed by biotinylated goat anti-rabbit immu-noglobulin (Ig) G (E0432, DakoCytomation).Labelling for CD79 proceeded with sequential appli-cation of monoclonal murine anti-human CD79(M7051, DakoCytomation) followed by polyclonalrabbit anti-murine IgG (Z0259, DakoCytomation)and biotinylated goat anti-rabbit IgG (EO432,DakoCytomation). The primary antibodies wereapplied at a dilution of 1 in 1,000 (for 1 h) andsecondary antibodies applied at a dilution of 1 in500 (for 20 min). After application of the biotinylatedantibodies, horseradish peroxidaseestreptavidin con-jugate (P0397, DakoCytomation) was applied ata dilution of 1 in 1,000 for 20 min followed by devel-opment with 3,30-diaminobenzidine (DAB, K4006,DakoCytomation). Sections were counterstainedwith Mayer’s haematoxylin.
Apoptotic cells were detected by end-labellingDNA fragments with biotinylated nucleotides usinga commercially available terminal deoxynucleotidyltransferase kit (TACS� TdT Kit, R&D Systems,Minneapolis, Minnesota). The paraffin-wax embed-ded tissue samples were dewaxed and rehydratedand processed according to the manufacturer’s in-structions. The tissues were counterstained withmethyl green. Apoptotic cells were identified on thebasis of histochemical (brown colouration of the nu-clei) and morphological (cell condensation) criteria.Apoptotic cells were counted in 10 randomly selectedmicroscopical high power, fields (40�microscope ob-jective).
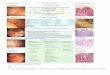
Fig. 1. Gastric mucosa from an Alaskan racing sled dog (group Idog from 300 miles). Epithelial lesions include vacuolationand attenuation of epithelial cells with occasional ulcers(arrow) that are shallow, overlaid by fibrin andneutrophilsand surrounded by attenuated epithelial cells (arrow-heads). Ulcers were only seen in dogs of group I. HE.Bar, 200 mm.
Statistical Analyses
Analysis of variance was used to compare treatmentlevels for various response variables with the useof PROC MIXED in PC SAS Version 9.1 (SAS
Institute, Cary, North Carolina). Pathologist to pa-thologist variability was accounted for by the use ofa RANDOM statement (i.e. blocked for the effect ofpathologist). Planned contrasts were performed tocompare non-zero distances with controls (no dis-tance) and to assess the effect of training and running.Means and standard errors are reported for each dis-tance and they are compared with pair-wise t-tests.The correlation between categories 4 and 1, 2 and 3was further investigated with a contingency tableand a chi square test. P values are reported for allcomparisons.
Results
Microscopical Lesions and Scoring
Epithelial Lesions. Epithelial lesions were present inall dogs and differed only in severity.Mild lesions con-sisted primarily of vacuolation of the basal cytoplasmwith apical displacement of the nucleus. In moder-ately affected samples, the columnar epithelium wasreplaced by either flattened or cuboidal epithelialcells, suggesting either compensatory attenuation sec-ondary to cell loss or re-epithelialization to covera previous defect. Four dogs from group 1 (i.e. oneeach from 100, 200, 300 and 500 mile distances) ex-hibited ulcers characterized by abrupt loss of the sur-face epithelium with local accumulation of fibrin andcellular debris (Fig. 1). Ulcers were not seen in sam-ples from the dogs of group II. Overall, lesion 1 (i.e.

Table 2
Summary of microscopical lesion scores (mean ± standard error of the mean)
Dog group Lesion 1 (Surface epithelium) Lesion 2 (Inflammation) Lesion 3 (Interface infiltrate) Lesion 4 (Presence of helical bacteria)
Group I dogs (distance run)
100 miles 1.08� 0.20 1.21� 0.17 0.58� 0.15 0.83� 0.17200 miles 1.21� 0.15 1.17� 0.14 0.58� 0.15 0.67� 0.21
300 miles 1.20� 0.21 1.03� 0.15 0.17� 0.11 0.67� 0.21
400 miles 0.75� 0.11 0.72� 0.09 0.50� 0.15 0.83� 0.17
500 miles 0.83� 0.10 0.78� 0.09 0.15� 0.08 0.70� 0.15
Group II dogs 0.54� 0.13 1.29� 0.21 0.58� 0.15 0.33� 0.21
Gastritis in Alaskan Racing Sled Dogs 71
epithelial lesion) scores (Tables 2 and 3) weresignificantly higher in the dogs of group I comparedwith those in group II (P¼ 0.0039).
Inflammatory Lesions. There were two distinct pat-terns of inflammatory cell infiltration that typicallyoccurred simultaneously. One lesion (lesion 2,Tables 2 and 3) included patchy to disseminatedinfiltrates of lymphocytes with fewer neutrophils,macrophages and plasma cells that were confinedto the lamina propria (Fig. 2A). In severely inflamedsections the lymphocytes formed prominent follicularaggregates, some with germinal centres. The appear-ance and distribution of the inflammation within thegastric mucosa were similar in both groups, but theseverity differed (Table 2). Within the group Idogs, the microscopical inflammatory scores for le-sion 2 decreased over the course of increased mile-age. Group II dogs exhibited the highestinflammatory scores, which were significantlygreater than the scores obtained from dogs of groupI (P¼ 0.0496).
The other inflammatory pattern (lesion 3, Tables 2and 3) consisted of an interface distribution ofinflammatory cells in the superficial lamina propria(Fig. 2B and C). The cellular infiltrates were domi-nated by lymphocytes and were focused at the base-ment membrane zone (BMZ) of the surfaceepithelium and neck epithelium of the gastric glands.The inflammatory infiltrate effaced the BMZ, withadditional lymphocytes forming prominent intraepi-thelial infiltrates. This pattern was present in samplesfrom both groups of dogs, but was not found in everydog of either group. Therefore, the interface pattern
Table
Summary of probabilities from contrasts be
Label Lesion 1 (Surface epithelium) Lesion 2 (Inflam
Group I versus Group II 0.0039* 0.0496*
*Significant P values.
of inflammation was not unique to either group(P¼ 0.2078).
Bacterial Infection. Spiral-shaped bacterial organisms(lesion 4, Tables 2 and 3) were noted along the surfaceof the mucosa, within the gastric pits and occasionallywithin apparent intracellular vacuoles of parietal cellslocated at the isthmus (Fig. 3). These organisms wereseen in samples from both groups and were more con-sistently seen in tissues from dogs of group I; however,this was not significant (P¼ 0.0516). The presence ofbacteria (lesion 4) did not significantly correlate withthe severity of epithelial lesions (lesion 1, P¼ 0.4730),the severity of inflammation (lesion 2, P¼ 0.2163) orthe pattern of interface inflammation (lesion 3,P¼ 0.2448).
Immunohistochemistry
The lymphocytic infiltrates within the lamina propriawere a mixture of CD3+ T lymphocytes (predomi-nating) and CD79+ B lymphocytes. The T cellswere dispersed whereas the B cells were concentratedmostly in follicular aggregates. The lymphocytic infil-trates obscuring the BMZ and the intraepithelial infil-trates were nearly all T cells (Fig. 4).
Apoptosis
The endurance-trained and exercised (group I) dogsdid not have an increase in apoptotic cells over thecourse of the 500-mile run (Table 4, P¼ 0.1377).The group I dogs exhibited higher numbers of apo-ptotic cells compared with dogs of group II; however,
3
tween groups of Alaskan racing sled dogs
mation) Lesion 3 (Interface inflammation) Lesion 4 (Helical bacteria)
0.2078 0.0516

Fig. 2. (A) Gastric mucosa from a group I (300 mile) Alaskan racing sled dog. There is severe inflammation of the lamina propriacharacterized by diffuse infiltration with lymphocytes and fewer neutrophils, macrophages and plasma cells. HE. Bar, 500 mm.(B) Gastric mucosa from a group I dog at 300 miles. (C) Gastric mucosa from a group I dog at 100 miles. In both sections(B) and (C), a lymphocytic infiltrate obscures the basement membrane (arrowheads) and forms nests of intraepithelial infiltrates(arrows). This ‘interface’ pattern was not unique to either group of dogs. HE. Bar (B and C), 100 mm.
72 J.W. Ritchey et al.
the difference was not statistically significant(P¼ 0.0544).
Discussion
Gastritis, gastric ulcers and haemorrhage have beenidentified as important causes of decreased perfor-mance and even acute death of dogs competing inthe 1,100 mile Iditarod sled race (Dennis et al.,2008). Alaskan sled dogs appear to be a useful modelto study the pathogenesis of exercise-induced gastritisand this study focused on the microscopical lesionspresent in conditioned dogs under exercise stress.Because the microscopical lesions of ulceration andhaemorrhage are predictable, and because secondarylesions associated with ulceration may obscure thelesion(s) associated with ulcer development, wechose to evaluate biopsy samples obtained from
endoscopically normal regions of the stomach,regardless of whether or not there were lesions in othergastric regions.
The original intent of this study was to documentthe temporal development of gastric disease in dogsover the course of exertion associated with long dis-tance racing. We were surprised to find that gastricinflammation did not appear to increase in eitherprevalence or intensity over time and distance in theexercised animals. Although not statistically signifi-cant, the trend was for the inflammatory gastric le-sions to improve over time and distance in the dogsof group I. These findings are similar to thosereported previously for racing sled dogs in which en-doscopic lesions and mucosal barrier alterationswere recognized at the onset of exercise, but did notprogress in severity over time and distance (Daviset al., 2006).

Fig. 3. Gastric mucosa from an Alaskan racing sled dog. Helicalbacteria are present along the surface of the mucosalepithelial cells, within gastric pits (arrow) and withinapparent intracellular vacuoles of parietal cells at the isth-mus region (arrowheads). HE. Bar, 10 mm.
Gastritis in Alaskan Racing Sled Dogs 73
Group I dogs had the highest epithelial lesionscores and these were the only dogs with gastric ulcer-ation. The microscopical scores for dogs in group Iwere significantly greater than those for dogs in group
Fig. 4. Gastricmucosa from anAlaskan racing sled dog. Lymphocytesmentmembrane, and the intraepithelial infiltrates (arrows) areto the follicular aggregates as seen in Fig. 2A. IHC. Bar, 200 m
II (P¼ 0.0039), indicating a probable effect of exer-cise on lesion development.
The endurance trained (group II) dogs exhibitedsignificantly higher gastric inflammation scores(P¼ 0.0496). This difference is most likely related tothe decline in severity scores with the increase in rac-ing time and distance in group I dogs, especially thosethat covered 400 and 500 miles. A similar decline inmicroscopical score associated with increasing exer-cise time and distance was also seen in the evaluationof lesion 1. It is unknown whether this apparent ‘anti-inflammatory effect’ of exercise is a true physiologicalmechanism or is secondary to an unintentionallyskewed distribution of the sample population.
Microscopical gastritis is not uncommon in dogs(Lidbury et al., 2009); however, the distribution ofthe inflammatory cells in the gastric tissues of thesedogs appears unique. The most intense inflammationwas centred on the BMZ of the surface and neck re-gion epithelium and accompanied by intraepithelialinfiltrates of T lymphocytes. The inflammation oftenobscured the BMZ, resulting in an ‘interface-type’ le-sion with an intraepithelial component similar to thelymphocytic gastritis syndrome described in man(Haot et al., 1988). As with the dogs reported here,the endoscopic appearance of the stomach in humanpatients with lymphocytic gastritis can be normal(M€uller et al., 2001). Lymphocytic gastritis in peopleis most often associated with either Helicobacter pylori
infection (Niemel€a et al., 1995) or autoimmune
disseminated in the lamina propria (arrowheads) or along the base-primarily CD3+T cells. B cells (CD79+)were primarily restrictedm.

Table 4
Total apoptotic cells in 10 randomly-selected
high-power fields
Dog group Mean apoptotic cells� SEM*
Group I(distance run prior to sampling)
100 miles 32.0� 23.2
200 miles 97.7� 26.4
300 miles 41.5� 19.3400 miles 89.5� 38.1
500 miles 67.0� 30.0
Group II (rested) 12.3� 12.2
*Standard error of the mean. By ANOVA, the overall P value for theeffect of distance was 0.1377. The comparison of group I to group II
dogs has a P value of 0.0544.
74 J.W. Ritchey et al.
(coeliac) disease (Feeley et al., 1998). Organisms con-sistent with Helicobacter spp. were seen in the biopsysamples from the dogs in this study and were seen ina majority of the samples (i.e. 15 of 19) from Alaskansled dogs studied previously (Davis et al., 2003a). Theorganisms were present on the mucosal surface andoccasionally within the glandular lumen. In addition,the organisms were also present within apparent in-tracellular vacuoles of parietal cells restricted to theisthmus. It was not possible to determine whetherthe organisms were present within the cytoplasm,true intracellular vacuoles or within intracellular can-alicular spaces (Hermanns et al., 1995; Ito et al., 2008).The results of this study show no correlation betweenthe presence of spiral-shaped bacteria and the severityof the microscopical inflammation score. Indeed, dogsof group II had the highest inflammation score, butthe lowest scores for lesion 4. There was also nocorrelation between the presence of bacteria andepithelial lesions (lesion 1) or the interface patternof inflammation (lesion 3). Therefore, similar to theconclusions of other studies (Hermanns et al., 1995;Davis et al., 2003a; Wiinberg et al., 2005), we foundno association between the presence of mucosalspiral-shaped bacteria and the severity of epitheliallesions or gastritis.
The cause of chronic gastritis in sled dogs is un-known. The predominantly superficial nature of thegastritis is usually associated with caustic agents(e.g. non-steroidal anti-inflammatory drugs, in-creased gastric acid) and dietary factors. There is ev-idence that one or both may play a role in the gastritisof Alaskan racing sled dogs. Pharmacological inhibi-tion of gastric acid secretion reduced the prevalenceand severity of gastric ulcers in exercised Alaskansled-racing dogs in previous studies, suggesting thatgastric acid plays some role (Davis et al., 2003b).Elevated serum gastrin levels have been associated
with lymphocytic gastritis (Zavros et al., 2002), andphysical training has been shown to increase gastrinlevels after feeding in horses (Furr et al., 1994). Bothsituations could apply to the Alaskan racing sleddogs. To our knowledge, serum gastrin levels havenot yet been evaluated in Alaskan sled dogs.
The diet of Alaskan sled dogs is unique. To meettheir exceptionally high energy demands, the dogsare fed an extremely high-fat diet (i.e. >60%) fre-quently during racing and approximately 38% die-tary fat during conditioning. Strenuous exercise(Kondo et al., 1994) and high dietary fat (Boulbyet al., 1999) can cause delayed gastric emptying,which could produce acid retention. Furthermore,high dietary fat produces gastritis in laboratory ani-mals (Laurila et al., 2001) as well as people eating ke-togenic diets to control epilepsy (Kang et al., 2004).We lack a group of dogs that live in the same environ-ment, eat the same diet, but have never been trainedor raced for a true negative control comparison be-cause such a group essentially does not exist. There-fore, we speculate that the high-fat diet, strenuousexercise and the interface-type gastritis observed inthe gastric samples from these dogs suggest an im-mune-mediated pathogenesis. Once initiated, thistype of injury can be self-perpetuating, which couldexplain the inflammatory lesions in dogs that areweeks removed from racing and/or conditioning.
Maintenance of the gastric epithelial barrier is a dy-namic process of epithelial cell proliferation balancedby cell loss through apoptosis (VonHerbay andRudi,2000). Uncoupling epithelial proliferation and loss isthe basic mechanism of any gastric epithelial barrierlesion, whether unchecked proliferation (neoplasia),atrophy or ulceration. The unique position of the lym-phocytic inflammatory cells at the BMZ and withinintraepithelial regions is consistent with T-cell-medi-ated induction of apoptosis. The microscopical pic-ture in the dogs is similar to the T-cell-mediatedapoptosis known to be important in the pathogenesisof models of Helicobacter gastritis (Ishihara et al., 2001;Souza et al., 2006). In the Alaskan sled dogs, there isa trend for apoptosis to be increased in conditioneddogs under racing stress. Although not statisticallysignificant (P¼ 0.0544), there may be somebiological relevance. Further investigation into therole of apoptosis is needed, including more specificidentification of the phenotype of infiltrating T cellsand evaluation of the specific apoptotic pathways(e.g. mediated by the interaction of Fas/Fas ligandor tumour necrosis factor a with its receptor) thatmay play a role.
In summary, Alaskan racing sled dogs exhibit gas-tritis that appears to be related to endurance training.The microscopical nature of the interface gastritis is

Gastritis in Alaskan Racing Sled Dogs 75
similar to human lymphocytic gastritis and suggestsan immune-mediated mechanism. Gastric ulcerswere acute and only seen in endurance-traineddogs that were raced on the groomed course. Becauseof the short timeframe in which the ulcers can de-velop, it seems more likely that ulcers are caused byepithelial cell loss rather than reduced cell prolifera-tion. The cause of epithelial cell loss is still unknown;however, increased T-cell-mediated apoptosis mayplay a role. Whatever the mechanism, perhaps thebackground inflammation lowers the threshold for ul-cerogenesis that is ultimately precipitated by an as yetunknown mechanism induced by strenuous physicalexertion.
Acknowledgments
The authors thank Mr. R. Swenson of Two Rivers,Alaska, for serving as musher for the study. Thanksare also due to Mr. C. Andrew and Ms. D. Giracellofrom the Oklahoma Animal Disease Diagnostic Labo-ratory for assistance with histology and immunohisto-chemistry. This work was supported by funding fromthe Defense Advanced Research Projects Agency(DARPA) of the US Department of Defense.
References
Boulby P, Moore R, Gowland P, Spiller RC (1999) Fat de-lays emptying but increases forward and backward an-tral flow as assessed by flow-sensitive magneticresonance imaging. Neurogastroenterology and Motility,11, 27e36.
Choi SC, Choi SJ, Kim JA, Kim TH, Nah YH et al. (2001)The role of gastrointestinal endoscopy in long-distancerunners with gastrointestinal symptoms. European
Journal of Gastroenterology and Hepatology, 13, 1089e1094.Davis MS, Willard M, Nelson S, Mandsager RE,
McKiernan BC et al. (2003a) Prevalence of gastriclesions in racing Alaskan sled dogs. Journal of VeterinaryInternal Medicine, 17, 311e314.
Davis MS, Willard M, Nelson S, McCullough S,Mandsager RE et al. (2003b) Efficacy of omeprazolefor the prevention of exercise-induced gastritis in racingAlaskan sled dogs. Journal of Veterinary Internal Medicine,17, 163e166.
Davis MS, Willard M, Williamson KK, Royer CM,Payton ME et al. (2006) The temporal relationship be-tween gastrointestinal protein loss, gastric ulcerationor erosion, and strenuous exercise in racing Alaskansled dogs. Journal of Veterinary Internal Medicine, 20,835e839.
Davis MS, Willard MD, Williamson KK, Steiner JM,Williams DA (2005) Sustained strenuous exercise in-creases intestinal permeability in racing Alaskan sleddogs. Journal of Veterinary Internal Medicine, 19, 34e39.
DayMJ, Bilzer T,Mansell J,Wilcock B,Hall EJ et al. (2008)Histopathological standards for the diagnosis of
gastrointestinal inflammation in endoscopic biopsy sam-ples from the dog and cat: a report from the World SmallAnimal Veterinary Association Gastrointestinal Stan-dardization Group. Journal of Comparative Pathology, 138,S1eS43.
Dennis MM, Nelson SN, Cantor GH,Mosier DA, Blake JEet al. (2008) Assessment of necropsy findings in sled dogsthat died during Iditarod sled dog races: 23 cases(1994e2006). Journal of the American Veterinary Medical
Association, 232, 564e573.Feeley KM, Heneghan NA, Stevens FM, McCarthy CF
(1998) Lymphocytic gastritis and coeliac disease: evi-dence of a positive association. Journal of Clinical Pathol-ogy, 51, 207e210.
Furr M, Taylo L, Kronfeld D (1994) The effects of exercisetraining on serum gastrin responses in the horse. CornellVeterinarian, 84, 41e45.
Haot J, Hamichi L,Wallez N,Mainguet P (1988) Lympho-cytic gastritis: a newly described entity. A retrospectiveendoscopic and histological study. Gut, 29, 1258e1264.
Hermanns W, Kregel K, Breuer W, Lechner J (1995)Helicobacter-like organisms: histopathological examina-tion of gastric biopsies from dogs and cats. Journal ofComparative Pathology, 112, 307e318.
Ishihara S, Fukuda R, Kawashima K, Moriyama N,Suetsugu H et al. (2001) T cell-mediated cytotoxicityvia Fas/Fas ligand signaling inHelicobacter pylori-infectedgastric corpus. Helicobacter, 6, 283e293.
Ito T, Kobayashi D, Uchida K, Takemura T, Nagaoka Set al. (2008)Helicobacter pylori invades the gastric mucosaand translocates to the gastric lymph nodes. LaboratoryInvestigation, 88, 664e681.
Kang HC, Chung DE, Kim DW, Kim HD (2004) Early-and late-onset complications of the ketogenic diet for in-tractable epilepsy. Epilepsia, 45, 1116e1123.
Kondo T, Naruse S, Hayakawa T, Shibata T (1994) Effectof exercise on gastroduodenal functions in untraineddogs. International Journal of SportsMedicine, 15, 186e191.
Laurila A, Cole SP, Merat S, Obonyo M, Palinski W et al.(2001) High fat, high-cholesterol diet increases the inci-dence of gastritis in LDL receptor-negative mice.Arterio-sclerosis, Thrombosis, and Vascular Biology, 21, 991e996.
Lidbury JA, Suchodolski JS, Steiner JM (2009) Gastric his-topathologic abnormalities in dogs: 67 cases(2002e2007). Journal of the American Veterinary Medical
Association, 234, 1147e1153.Moses FM (1990) The effect of exercise on the gastrointes-
tinal tract. Sports Medicine, 9, 159e172.M€uller H, Rappel S, Volkholz H, Stolte M (2001) Lym-
phocytic gastritis: a rare disorder of the gastric mucosa.Der Pathologe, 22, 56e61.
Murray MJ, Schusser GF, Pipers FS, Gross SJ (1996)Factors associated with gastric lesions in thoroughbredracehorses. Equine Veterinary Journal, 28, 368e374.
Niemel€a S, Karttunen T, Kerola T, Karttunen R (1995)Ten year follow up study of lymphocytic gastritis: fur-ther evidence on Helicobacter pylori as a cause of lympho-cytic gastritis and corpus gastritis. Journal of Clinical
Pathology, 48, 1111e1116.

76 J.W. Ritchey et al.
Royer CM, Willard MD, Williamson KK, Steiner JM,Williams DA et al. (2005) Exercise stress, intestinal per-meability, and gastric ulceration in racing Alaskan sleddogs. Equine and Comparative Exercise Physiology, 2, 53e59.
Souza HS, Neves MS, Elia CC, Tortori CJ, Dines I et al.(2006) Distinct patterns of mucosal apoptosis in H. py-
lori-associated gastric ulcer are associated with alteredFasL and perforin cytotoxic pathways. World Journal of
Gastroenterology, 12, 6133e6141.Vatistas NJ, Snyder JR, Carlson G, Johnson B, Arthur RM
et al. (1999) Cross-sectional study of gastric ulcers of thesquamous mucosa in thoroughbred racehorses. EquineVeterinary Journal, 29, S34eS39.
Von Herbay A, Rudi J (2000) Role of apoptosis in gastricepithelial turnover. Microscopy Research and Technique,48, 303e311.
Wiinberg B, Spohr A, Dietz HH, Egelund T, Greiter-Wilke A et al. (2005) Quantitative analysis of inflamma-tory and immune responses in dogs with gastritis andtheir relationship to Helicobacter spp. infection. Journalof Veterinary Internal Medicine, 19, 4e14.
Zavros Y, Rieder G, Ferguson A, Samuelson LC,Merchant JL (2002) Hypergastrinemia in response togastric inflammation suppresses somatostatin. AmericanJournal of Physiology: Gastrointestinal and Liver Physiology,282, G175eG183.
½ R
A
eceived, January 25th, 2010
ccepted, November 12th, 2010
�