Embed Size (px)
Citation preview

Funktionelle Bedeutung der Infektion von T-Lymphozyten
durch Respiratorisches Syncytial Virus (RSV)
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie
der Ruhr-Universität Bochum
angefertigt in der
Medizinischen Fakultät
Abteilung für Experimentelle Pneumologie
(Leiter: Prof. Dr. med. A. Bufe)
vorgelegt von
Marion Kauth
aus
Bochum
Bochum 2002

___________________________________________________________________________
Referent: Prof. Dr. A. Bufe Korreferent: Prof. Dr. H. Lübbert

____________________________________________________________________________ I
Inhalt Abkürzungen......................................................................................................................... 1
Zusammenfassung............................................................................................................... 4
Summary ................................................................................................................................ 6
1.Einleitung ............................................................................................................................ 8
1.1 Die verschiedenen Funktionen von T-Zellen bei der Immunantwort ................. 8
1.2 Das TH1-/TH2-Konzept und Interleukin-10 ............................................................... 9
1.3 Respiratorisches Syncytial Virus ............................................................................... 10
1.3.1 Taxonomie und struktureller Aufbau von RSV ....................................................... 11
1.3.2 Infektion verschiedener Zelltypen durch RSV......................................................... 12
1.3.3 Die spezifische T-Zellantwort auf eine RSV-Infektion............................................ 14
1.4 Chronisch obstruktive Lungenerkrankung ................................................................... 15
1.5 Zielsetzung......................................................................................................................... 19
2. Material und Methoden.............................................................................................. 21
2.1 Material............................................................................................................................ 21
2.1.1 Spenderblut............................................................................................................... 21
2.1.2 Virusmaterial ............................................................................................................ 21
2.1.3 Medien und Lösungen .............................................................................................. 21
2.1.4 Antikörper ................................................................................................................ 23
2.1.5 Reagenzien ............................................................................................................... 24
2.2 Methoden ......................................................................................................................... 25
2.2.1 Zellpräparation ......................................................................................................... 25
2.2.1.1 Isolierung mononukleärer Zellen des peripheren Blutes ............................. 25
2.2.1.2 Magnetische Zellsortierung.......................................................................... 25
2.2.1.3 Gewinnung von Alveolarmakrophagen ....................................................... 27
2.2.1.4 Zellzählung und Vitalitätsbestimmung ........................................................ 28

INHALT II
2.2.2 Virusanzucht............................................................................................................. 28
2.2.3 Kultivierung und Stimulation der Zellen.................................................................. 29
2.2.4 Quantitative Zytokinbestimmungen in Kulturüberständen ...................................... 30
2.2.5 Immunfluoreszenzmarkierungen und durchflusszytometrische Zellanalyse ........... 31
2.2.6 Zellanalyse im konfokalen Laser-Rastermikroskop................................................. 33
2.2.7 Isolierung von zellulärer und viraler RNA............................................................... 34
2.2.8 Reverse Transkription der RNA............................................................................... 35
2.2.9 Etablierung einer quantitativen real-time PCR für humanes Interleukin-10............ 36
2.2.9.1 Prinzip der quantitativen real-time PCR ...................................................... 36
2.2.9.2 Festlegung der Primer- und Sonden-Sequenzen .......................................... 36
2.2.9.3 Reaktionsansätze der quantitativen real-time PCR für humanes
Interleukin-10............................................................................................... 38
2.2.9.4 Relative Quantifizierung der IL-10 mRNA Expression .............................. 39
2.2.9.5 Analyse der PCR-Produkte .......................................................................... 40
2.2.9.6 Klonierung der PCR-Produkte ..................................................................... 40
2.2.9.7 Sequenzierung der PCR-Produkte ............................................................... 42
2.2.9.8 Absolute Quantifizierung der IL-10 Kopienzahl ......................................... 43
2.2.10 RSV-Nachweis mittels quantitativer real-time PCR .............................................. 44
2.2.11 Statistische Auswertungen...................................................................................... 44
3. Ergebnisse........................................................................................................................ 45
3.1 Ergebnisse der einstufigen und zweistufigen magnetischen Zellsortierung..... 45
3.1.1 Reinheit der Zellfraktionen nach magnetischer Sortierung...................................... 45
3.1.2 Zellausbeute bei der magnetischen Zellsortierung ................................................... 48
3.2 Phänotypische Charakterisierung der Interaktion zwischen RSV und
T-Zellen ........................................................................................................................... 49
3.2.1 Analyse der Interaktion zwischen RSV und T-Zellen im Durchflusszytometer...... 49
3.2.1.1 Anteil RSV-positiver T-Zellen unter verschiedenen Kulturbedingungen ... 49
3.2.1.2 Expression von ICAM-1 auf RSV-stimulierten T-Zellen............................ 52
3.2.1.3 Expression von CD69 auf RSV-stimulierten T-Zellen ................................ 53
3.2.2 Analyse der Interaktion zwischen RSV und T-Zellen im konfokalen Laser-
Rastermikroskop ...................................................................................................... 53
3.2.3 Nachweis von RSV-RNA in RSV-exponierten T-Zellkulturen ............................... 56

INHALT III 3.3 Nachweis von humanem Interleukin-10 mittels quantitativer real-time
PCR .................................................................................................................................. 58
3.3.1 Auswahl der Primer und Amplifikation der Zielsequenz......................................... 58
3.3.2 Spezifität der quantitativen real-time PCR für humanes Interleukin-10 .................. 60
3.3.3 Sensitivität und Variabilität der quantitativen real-time PCR für humanes
Interleukin-10........................................................................................................... 62 3.4 Untersuchungen zur funktionellen Bedeutung der Interaktion zwischen
RSV und T-Zellen in vitro ........................................................................................... 64
3.4.1 Vitalität der virusstimulierten T-Zell-Kulturen........................................................ 64
3.4.2 IL-10 mRNA-Expression in RSV-stimulierten T-Zellkulturen ............................... 64
3.4.3 IL-10 mRNA-Expression in T-Zellkulturen mit anderen Stimuli............................ 67
3.4.4 IL-10 mRNA-Expression in RSV-stimulierten Monozytenkulturen ....................... 68
3.4.5 IL-10 Proteinkonzentration im Überstand RSV-stimulierter T-Zellkulturen
und Monozytenkulturen ........................................................................................... 69
3.4.6 IL-10 mRNA Expression in Kokulturen aus T-Zellen und Monozyten................... 71
3.4.7 IL-10 Proteinkonzentration im Überstand RSV-stimulierter Kokulturen aus
T-Zellen und Monozyten ......................................................................................... 74
3.4.8 IL-10 Proteinkonzentration im Überstand RSV-stimulierter Kokulturen aus
Alveolarmakrophagen und T-Zellen........................................................................ 76
3.4.9 IFN-γ Proteinkonzentration im Überstand RSV-stimulierter Kokulturen................ 77
3.4.10 Verhältnis von IL-10 und IFN-γ Produktion in RSV-stimulierter Kokulturen ...... 79
4. Diskussion ........................................................................................................................ 82
4.1 Gewinnung hochreiner T-Zellen aus peripherem Blut............................................ 82
4.2 Phänotypische Charakterisierung RSV-stimulierter T-Zellen ................................ 82
4.3 Nachweis einer funktionellen Bedeutung der Interaktion von RSV und T-Zellen
in vitro ..................................................................................................................... 87
4.4 Die Rolle RSV-stimulierter T-Zellen in verschiedenen Kokulturmodellen............ 93
4.5 Möglicher Einfluß RSV-stimulierter T-Zellen auf die virusinduzierte
Immunantwort bei COPD ....................................................................................... 96
5. Literatur ........................................................................................................................... 99

____________________________________________________________________________ 1
Abkürzungen α1-AT alpha1-Antitrypsin
AdV Adenovirus
AM Alveolarmakrophagen
APC Antigen-präsentierende Zelle
ATCC American Type Culture Collection
BAL Bronchoalveoläre Lavage
BLAST Basic Local Alignment Search Tool
bp Basenpaar
BSA Bovines Serumalbumin
CD Cluster of Differentiation
cDNA komplementäre Desoxyribonukleinsäure
CLSM konfokales Laser-Rastermikroskop
COPD Chronisch obstruktive Lungenerkrankung
CV Variationskoeffizient
DMEM Dulbecco’s Modified Eagle Medium
DNA Desoxyribonukleinsäure
EBV Epstein-Barr-Virus
EDTA Ethylendiamintetraacetat
ELISA Enzyme-linked immunosorbent assay
FAM 6-Carboxyfluorescein
FCS Fötales Kälberserum
FITC Fluorescein-Isothiocyanat
GM-CSF Granulozyten-Makrophagen-Kolonie-stimulierender Faktor
GRO-α Growth regulated oncogene-alpha
GUS ß-Glucuronidase
HA Hämagglutinierende Einheiten
HBSS Hanks’ Balanced Salt Solution
HEPES 4-(2-Hydroxyethyl)-piperazin-1-ethan-sulfonsäure
HRV humanes Rhinovirus
ICAM Interzelluläres Adhäsionsmolekül
I.E. Internationale Einheiten

ABKÜRZUNGEN 2 IFN Interferon
IL Interleukin
LB Luria-Bertani
LTB4 Leukotrien-B4
MACS Magnetically activated cell sorting
mAk monoklonaler Antikörper
MCP-1 Monozyten chemotaktisches Protein-1
MHC Haupthistokompatibilitätskomplex
MIP-1α Makrophagen inflammatorisches Protein-1α
MMP Matrixmetalloproteinase
m.o.i. multiplicity of infection
mRNA Boten-Ribonukleinsäure
NCBI National Center for Biotechnology Information
NDV Newcastle Disease Virus
pAk polyklonaler Antikörper
PBMC Mononukleäre Zellen des peripheren Blutes
PBS Phosphat-gepufferte Kochsalzlösung
PCR Polymerase-Kettenreaktion
PE Phycoerythrin
PGE2 Prostaglandin E2
PHA Phytohämagglutinin
PIV Parainfluenzavirus
PMA Phorbol-12-Myristat-13-Acetat
rIL-10 rekombinantes humanes Interleukin-10
RNA Ribonukleinsäure
RSV Respiratorisches Syncytial Virus
RSV-F RSV-Fusionsprotein
SLPI sekretorischer Leukoprotease Inhibitor
TAMRA 6-Carboxytetramethylrhodamin
TC1/2 cytotoxische T-Zelle Typ 1/2
TGF transformierender Wachstumsfaktor
TH1/2 T-Helferzelle Typ 1/2
TIMP tissue inhibitor of matrixmetalloproteinases
Tr1 regulatorische T-Zelle Typ 1

ABKÜRZUNGEN 3 TMB 3,3’,5,5’-Tetramethylbenzidin
TM Schmelztemperatur
TNF Tumornekrosefaktor
TPA Tetradecanoyl-Phorbolazetat
Tris Tris(hydroxymethyl)aminomethan
TZR T-Zellrezeptor
U Unit
x g Vielfaches der Erdbeschleunigung

____________________________________________________________________________ 4
Zusammenfassung
Respiratorisches Syncytial Virus (RSV) wird sehr häufig in humanen Atemwegen gefunden
und gilt als Auslöser einer schweren Entzündung der kleinen Atemwege (Bronchiolitis) im
frühen Kindesalter. Seine Beteiligung an der Pathogenese und der anfallsartigen
Verschlechterung (Exazerbation) der chronisch obstruktiven Lungenerkrankung (COPD) bei
Erwachsenen wird diskutiert. In einem Teilprojekt der Klinischen Forschergruppe
„Bedeutung von Virusinfektionen bei akuten und chronischen Atemwegserkrankungen im
Kindes- und Erwachsenenalter“ wurde der Einfluss einer RSV-Infektion auf die funktionellen
Eigenschaften von Alveolarmakrophagen und kokultivierten T-Zellen in vitro untersucht.
Dabei konnte beobachtet werden, dass nicht nur die Makrophagen, sondern vermutlich auch
die T-Zellen von RSV infiziert wurden. In der vorliegenden Arbeit wird daher die Interaktion
zwischen RSV und T-Zellen zellbiologisch und immunologisch näher charakterisiert. Dabei
sollte überprüft werden, ob es sich tatsächlich um eine Infektion der T-Zellen durch RSV
handelte und inwieweit diese Interaktion darüber hinaus funktionelle Bedeutung hat.
T-Zellen und Monozyten wurden aus dem peripheren Blut freiwilliger erwachsener Spender
mittels Gradientenzentrifugation und magnetischer Zellsortierung isoliert. CD3+ isolierte
RSV-stimulierte T-Zellen wurden phäntotypisch im Durchflusszytometer und konfokalen
Laser-Rastermikroskop (CLSM) analysiert. Die extrazelluläre Analyse im Durch-
flusszytometer ergab eine Zunahme des Anteils RSV-positiver T-Zellen bei steigender
Virusdosis. Die zugehörige Kurve zeigte den Verlauf einer Sättigungskurve. Bei
gleichzeitiger Stimulierung mit antiCD28 oder PHA war der Anteil RSV-positiver Zellen
höher als bei reiner RSV-Stimulierung. Mittels CLSM-Aufnahmen wurde gezeigt, dass die
Bindung von RSV an die T-Zellen räumlich getrennt vom T-Zellrezeptor-Komplex erfolgte.
Diese Beobachtungen legen nahe, dass es sich bei der Bindung um eine spezifische Bindung
an eine Oberflächenstruktur handelt, die je nach Aktivierungszustand der T-Zelle unter-
schiedlich stark exprimiert wird.
Mittels CLS-Mikroskopie wurde RSV 20 Stunden nach der Exposition auch intrazellulär in
T-Zellen nachgewiesen. Die Messung von RSV-RNA in RSV-exponierten T-Zellen im Zeit-
verlauf mittels quantitativer real-time PCR erbrachte keine Hinweise auf eine produktive
Infektion.

ZUSAMMENFASSUNG 5
Für die Untersuchung des Einflusses der RSV-Exposition auf die Funktion der T-Zellen
wurde eine quantitative real-time PCR für das immunsuppressive Zytokin Interleukin(IL)-10
etabliert. Zudem wurde die Reinheit der T-Zellfraktion durch Erweiterung der magnetischen
Zellsortierung um eine vorgeschaltete CD14-Depletion weiter gesteigert. Es zeigte sich, dass
RSV-Stimulierung zu einer signifikanten Zunahme der IL-10 mRNA Expression in CD3+14-
T-Zellen führte. Die Höhe der IL-10 mRNA Expression war signifikant mit dem Anteil RSV-
positiver T-Zellen in den Kulturansätzen korreliert. In den Kulturüberständen wurden die
Konzentrationen von IL-10 und Interferon(IFN)-γ mittels ELISA bestimmt. In Verbindung
mit zusätzlichen Stimuli konnte eine Zunahme der IL-10 Freisetzung nach RSV-Stimulierung
in den T-Zellkulturen gezeigt werden. Die IL-10 mRNA Expression und die IL-10
Proteinkonzentration in den Kulturüberständen korrelierten signifikant miteinander. Das
bedeutet, dass die RSV-induzierte Steigerung der IL-10 mRNA Expression auch potentiell
biologisch wirksam werden konnte.
In Kokulturen aus CD3+14- T-Zellen und CD14+ Monozyten waren die IL-10 mRNA Expres-
sion und die IL-10 Proteinkonzentration im Kulturüberstand nach RSV-Stimulierung bei
gleicher Zellzahl gegenüber reinen CD14+ Kulturen synergistisch erhöht. Diese Zunahme der
IL-10 Konzentration bei RSV-Stimulierung zeigte sich auch in Kokulturen aus
Alveolarmakrophagen und autologen PHA-stimulierten CD3+ T-Zellen. In Verbindung mit
einer relativen Abnahme der IFN-γ Konzentration in den RSV-stimulierten Ansätzen ergab
sich daraus in beiden Kokultursystemen ein signifikant erhöhter IL-10/IFN-γ Quotient.
Aktivierte T-Zellen sind im Rahmen der Entzündungsreaktion der COPD deutlich vermehrt in
der Bronchialschleimhaut zu finden. Daher könnte eine RSV-Infektion der unteren Atemwege
bei einem COPD-Patienten zu einer höheren IL-10 Freisetzung und/oder einem erhöhten
IL-10/IFN-γ Quotienten führen und eine über das regulatorische Maß hinaus verstärkte
Suppression der antiviralen Immunantwort bewirken. Eine dadurch verzögerte Eliminierung
von RSV oder anderen Atemwegsviren könnte den Entzündungsprozess verstärken und eine
Exazerbation auslösen. Diese Zusammenhänge zeigen einen möglichen Mechanismus auf,
durch den eine RSV-Infektion direkt oder indirekt zur Exazerbation einer COPD beitragen
kann.

____________________________________________________________________________ 6
Summary
Respiratory syncytial virus (RSV) is one of the most frequent respiratory viruses found in
humans and known as a causal factor of severe small airway disease in young children. It is
also discussed to play a role in the pathogenesis and worsening (exacerbation) of chronic
obstructive pulmonary disease (COPD). As part of the Clinical Research Group “Significance
of viral infections in acute and chronic respiratory diseases of children and adults” a project
was launched to investigate the influence of RSV-exposition on the functional properties of
alveolar macrophages and co-cultured T cells in vitro. Interestingly, RSV was found not only
to infect the macrophages but probably also the T cells. Investigations reported here were
performed to characterize this interaction in more detail and to reveal wether T cells get really
infected with RSV. Moreover, possible functional consequences of this interaction should be
examined.
T cells and monocytes were isolated from peripheral blood of voluntary adult donors by
density gradient centrifugation and magnetic cell sorting. CD3+ isolated T cells were
phenotypically analysed by flow cytometry and confocal laser scanning microscopy (CLSM)
after stimulation with RSV. Extracellular staining showed higher ratios of RSV-positive
T cells with increasing viral dose. Shape of the corresponding graph was that of a saturation
curve. Simultaneous stimulation with antiCD28 or PHA augmented the ratio of RSV-positive
cells compared to stimulation with RSV solely. As shown by CLSM, binding occurred
independently of TCR complex. Together these findings suggest that RSV attaches
specifically to a structure on the surface of the T cells that is up regulated when the T cell is
activated.
Evidence of intracellular RSV particles was found by CLSM in T cell cultures 20 hours past
infection. RSV-specific RNA was measured in RSV-exposed T cells using a quantitative real-
time PCR. Within 48 hours after inoculation, no replication was detected that corresponded to
a productive infection of the T cells compared to the rise in viral RNA after infection of a
RSV-permissive cell line. However, low-level replication could not be excluded.
A quantitative real-time PCR specific for the immunosuppressive cytokine interleukin(IL)-10
was established for investigating the influence of RSV-exposition on functional properties of

SUMMARY 7 challenged T cells. As a further prerequisite, purity of T cell fractions was improved by
implementing an additional step of CD14+ depletion in the magnetic cell sorting procedure.
RSV-challenge led to a significant rise in IL-10 mRNA expression in CD3+14- T cells. There
was a significant correlation between IL-10 mRNA expression and ratio of RSV-positive
T cells. In culture supernatants concentrations of IL-10 and interferon(IFN)-γ were measured
by ELISA. In the presence of additional stimuli as antiCD28 or PHA it could be shown that
not only mRNA expression but also IL-10 concentration in supernatants was higher in RSV-
challenged samples. Moreover, there was a significant correlation between IL-10 mRNA
expression and IL-10 concentration in culture supernatants indicating that RSV-induced
increase in IL-10 mRNA expression could exert biological activity.
IL-10 mRNA expression and IL-10 concentration in supernatant were synergistically
increased in co-cultures of CD3+14- T cells and CD14+ monocytes as compared to cultures of
pure CD14+ cells after challenge with RSV. This rise in IL-10 concentration was also
observed in PHA-stimulated co-cultures of alveolar macrophages with autologous CD3+
T cells. In both co-culture systems increased IL-10 levels were accompanied by a relative
decrease in IFN-γ concentrations resulting in a significantly elevated IL-10/IFN-γ ratio in
RSV-stimulated cultures.
Activated T cells are numerous in inflammatory bronchial tissue of patients with COPD.
Therefore, a RSV infection of the lower respiratory tract of patients with COPD could lead to
a higher IL-10 response and/or an elevated IL-10/IFN-γ quotient that dampens the antiviral
immunresponse compared to healthy subjects. Slowed down elimination of RSV or other
respiratory viruses could amplify airway inflammation causing exacerbation of established
COPD. Possibly, this could be the mechanism by which RSV infections contribute to
exacerbations of COPD in a direct or indirect manner.

8
1. Einleitung
1.1 Die verschiedenen Funktionen von T-Zellen bei der Immunantwort
Das menschliche Immunsystem umfasst eine unspezifische Komponente, die als angeborene
Immunität und eine antigenspezifische Komponente, die als erworbene oder adaptive Immu-
nität bezeichnet wird. Die adaptive Immunität setzt sich im wesentlichen aus zwei
Bestandteilen zusammen, der zellulären und der humoralen Immunantwort. Letztere wird
hauptsächlich durch Antikörper vermittelt. T-Lymphozyten oder kürzer T-Zellen sind in
verschiedenen Funktionen an adaptiven Immunantworten des menschlichen Immunsystems
direkt oder indirekt beteiligt. Als zytotoxische T-Zellen sind sie direkte Effektorzellen in der
zellulären Immunantwort. Als sogenannte T-Helferzellen aktivieren sie B-Zellen zur
Antikörperproduktion bei einer humoralen Immunantwort, wirken aber auch aktivierend auf
Makrophagen, die wiederum an der zellulären Immunantwort beteiligt sind (Janeway 1997).
Die Antigenspezifität der T-Zellen wird durch membranständige Rezeptorproteine an ihrer
Zelloberfläche bestimmt, die als T-Zellrezeptoren (TZR) bezeichnet werden. Mit dem TZR
eng assoziiert ist das auf der Oberfläche von T-Zellen exprimierte CD3-Molekül, das mit
seinen zytoplasmatischen Domänen die Signalübertragung ins Innere der Zelle nach Bindung
des T-Zellrezeptors an seinen Liganden übernimmt. Im Gegensatz zu den Immunglobulinen
der B-Zellen erkennen T-Zellrezeptoren Antigene nur, wenn sie als kurzes Peptidfragment an
spezialisierte Glykoproteine, sogenannte MHC-Moleküle gebunden auf der Oberfläche einer
anderen Zelle präsentiert werden. Dabei wirken zwei Klassen von MHC-Molekülen mit den
verschiedenen T-Zelltypen zusammen (s. Tab. 1.1). MHC-I-Moleküle präsentieren Peptid-
fragmente von intrazellulären Erregern wie Viren oder Parasiten und werden von allen
kernhaltigen Zellen exprimiert. Der Komplex aus intrazellulärem Fremdantigen und MHC-I
wird durch CD8+ zytotoxische T-Zellen erkannt, die die Abtötung der infizierten Zelle
bewirken. Dagegen präsentieren MHC-II-Moleküle Antigene, die aus dem extrazellulären
Milieu in intrazelluläre Vesikel aufgenommen wurden. MHC-II-Moleküle werden in erster
Linie von follikulären dendritischen Zellen, Makrophagen und B-Zellen exprimiert und
interagieren mit CD4+ T-Helferzellen (Parkin 2001).

EINLEITUNG 9
Tab. 1.1: Übersicht über Beteiligung und Funktion verschiedener T-Zelltypen
bei der zellvermittelten und humoralen Immunantwort
(aus: Janeway 1997, modifiziert)
1.2 Das TH1-/TH2-Konzept und Interleukin-10
Außer durch direkten Zell/Zell-Kontakt erfolgt die Regulation des Immunsystems durch
humorale Faktoren, vor allem Zytokine, die von aktivierten Immunzellen freigesetzt werden.
Über entsprechende Rezeptoren an der Zelloberfläche können diese löslichen Botenstoffe
aktivierend oder inhibierend auf die Zelle selbst oder andere Zellen wirken. Nach dem
Konzept von Mosmann (1989), das im Mausmodell entwickelt wurde, werden CD4+
T-Helferzellen nach ihrem Zytokinprofil in TH1- und TH2-Zellen unterteilt. Demnach
produzieren TH1-Zellen die Zytokine Interleukin(IL)-2, Interferon-γ (IFN-γ) und Tumor-
Nekrose-Faktor-β (TNF-β). Die Ausschüttung dieser Zytokine wirkt in erster Linie auf den
zellulären Arm der Immunantwort ein, da z.B. Makrophagen durch IFN-γ aktiviert werden
(Tab. 1.1 Mitte). TH1-Zellen werden daher auch als proinflammatorische T-Zellen bezeichnet.
T-Helferzellen vom TH2-Typ produzieren IL-4, IL-5, IL-6, IL-10 und IL-13 und unterstützen
damit die humorale Immunantwort, da die freigesetzten Interleukine die B-Zellaktivierung

EINLEITUNG 10
fördern (Tab. 1.1 rechts). Durch Salgame (1991) wurde das TH1-/TH2-Konzept auch für
humane CD4+-Zellen nachgewiesen.
In jüngerer Zeit ist dieses Konzept jedoch in einigen Punkten differenziert und erweitert
worden. Zum einen wurde inzwischen auch für CD8+ T-Zellen die Existenz funktioneller
Subgruppen belegt, die sich ähnlich wie CD4+ T-Zellen in ihrem Zytokinprofil unterschieden
und analog zu diesen als TC1- oder TC2-Zellen bezeichnet werden (Vukmanovic-Stejic 2000).
Zum anderen zeigte sich insbesondere in Bezug auf humanes Interleukin-10 (IL-10), dass eine
klare Abgrenzung zwischen TH1- und TH2-Zelle nicht immer möglich ist. Die Ausschüttung
von IL-10 konnte sowohl für TH1-Klone als auch für TH2-Klone gezeigt werden (del Prete
1993). Darüber hinaus wurden inzwischen CD4+ T-Zellen identifiziert, die viel IL-10, IFN-γ,
TGF-β und IL-5, aber nur wenig IL-2 und kein IL-4 produzieren. Funktionell supprimieren
diese Zellen antigenspezifische Immunantworten, so dass sie als regulatorische Typ I
T-Zellen (Tr1) bezeichnet wurden (Groux 1997; Jonuleit 2000). Ähnliche, durch TGF-β
induzierte regulatorische T-Zellen wurden auch als TH3-Zellen bezeichnet (Weiner 2001). Die
immunsuppressive Wirkung von IL-10 beruht vor allem auf der Verminderung der antigen-
präsentierenden und akzessorischen Kapazität von Monozyten/Makrophagen und dendri-
tischen Zellen durch Inhibierung der Expression von MHC-II-Molekülen und den kostimula-
torischen Molekülen B7.1 (CD80) und B7.2 (CD86) (de Waal Malefyt 1991a; Ding 1993;
Corinti 2001). Darüber hinaus wirkt IL-10 aber auch direkt inhibierend auf die Produktion
verschiedener proinflammatorischer Zytokine wie IL-1α, IL-1β, IL-6, TNF-α und GM-CSF
(de Waal-Malefyt 1991). Ein weiterer Unterschied zum Immunsystem der Maus besteht darin,
dass im humanen System Monozyten als Hauptproduzenten von IL-10 nachgewiesen wurden
(Borish 1998).
1.3 Respiratorisches Syncytial Virus
Respiratorisches Syncytial Virus (RSV) wird sehr häufig in humanen Atemwegen gefunden
und gilt als Auslöser einer schweren Entzündung der kleinen Atemwege (Bronchiolitis) im
frühen Kindesalter. Bis zum Ende des zweiten Lebensjahres beträgt die Durchseuchungsrate
nahezu 100 %, wobei ca. 1 % der Infektionen stationärer Behandlung bedürfen (Collins
2001). Weigl (2001) leitete aus einer Untersuchung in den Jahren 1996 bis 1999 in Kiel und

EINLEITUNG 11
Umgebung für Gesamtdeutschland eine Inzidenzrate von 10.000 RSV-Infektionen pro Jahr
ab, die eine Krankenhausbehandlung der erkrankten Kinder erforderten. Die Bedeutung von
RSV-Infektionen für die Entstehung und Verschlechterung (Exazerbation) chronischer
Lungenerkrankungen bei Erwachsenen wird zur Zeit in verschiedenen klinischen Studien
untersucht und intensiv diskutiert (s. unten).
1.3.1 Taxonomie und struktureller Aufbau von RSV
RSV gehört zur Gattung Pneumovirus der Familie Paramyxoviridae. Zur gleichen Familie,
jedoch jeweils zu anderen Gattungen, gehören die Parainfluenzaviren Typ 1 bis 4, das
Mumps- und das Masernvirus sowie das Newcastle-Disease-Virus (NDV), ein human apatho-
genes Virus, das für verschiedene Geflügelspezies infektiös ist (Modrow 1998).
RSV enthält ein 15,2 Kilobasen (kb) umfassendes Minus-Strang-RNA-Genom, das voll-
ständig sequenziert ist und für 11 Proteine kodiert. Das Nukleoprotein (N), das Phospho-
protein (P) und die RNA-Polymerase (L) bilden zusammen mit der viralen Nukleinsäure das
Nukleokapsid im Innern des Virus (Zhang 2002). Die Virushülle besteht aus einer Lipid-
Doppelschicht mit dem membranassoziierten Matrixprotein (M) und drei Transmembran-
Proteinen, dem Fusionsprotein (F), dem Attachmentprotein (G) und einem kleinen, hydro-
phoben Protein (SH), dessen Funktion noch unbekannt ist. Die übrigen Proteine sind zwei
Proteine M2-1 und M2-2, die regulatorische Funktionen bei der Transkription erfüllen und
zwei sogenannte nicht-strukturelle Proteine, die es nur bei Pneumoviren gibt und deren
Funktion noch nicht vollständig geklärt ist (Hall 2001). Der Aufbau eines RSV-Partikels ist in
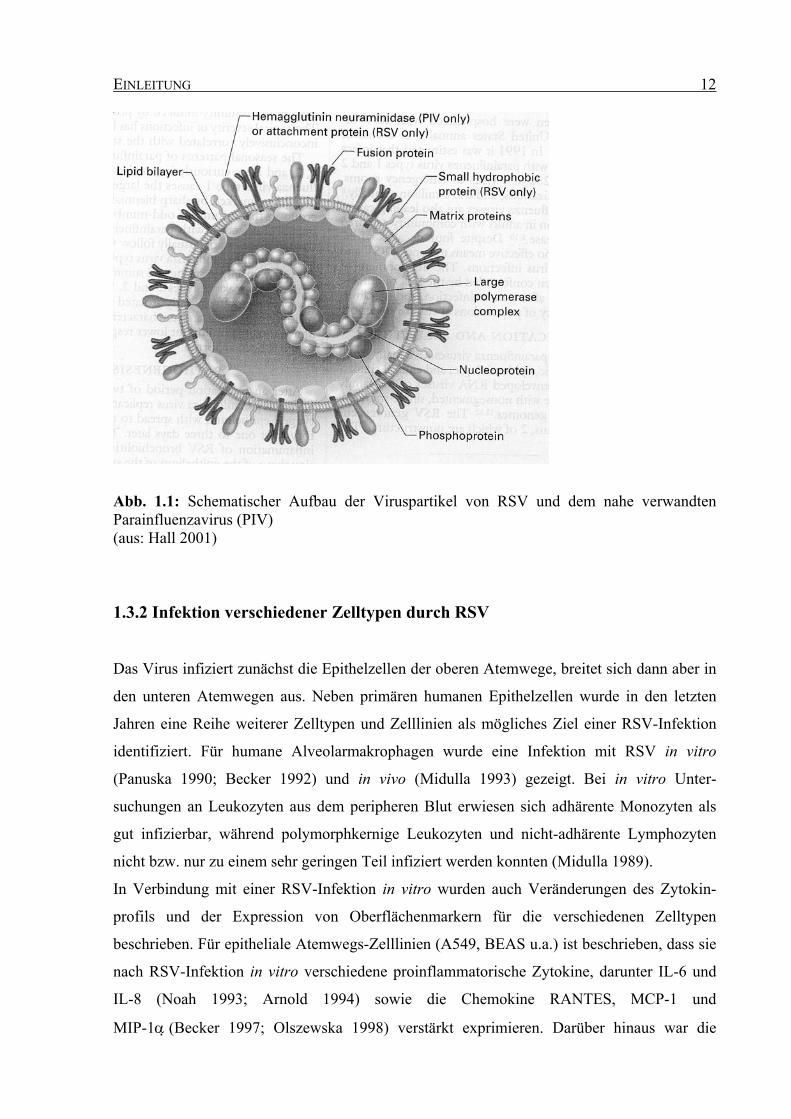
Abbildung 1.1 schematisch dargestellt.
EINLEITUNG 12
Abb. 1.1: Schematischer Aufbau der Viruspartikel von RSV und dem nahe verwandten Parainfluenzavirus (PIV) (aus: Hall 2001)
1.3.2 Infektion verschiedener Zelltypen durch RSV
Das Virus infiziert zunächst die Epithelzellen der oberen Atemwege, breitet sich dann aber in
den unteren Atemwegen aus. Neben primären humanen Epithelzellen wurde in den letzten
Jahren eine Reihe weiterer Zelltypen und Zelllinien als mögliches Ziel einer RSV-Infektion
identifiziert. Für humane Alveolarmakrophagen wurde eine Infektion mit RSV in vitro
(Panuska 1990; Becker 1992) und in vivo (Midulla 1993) gezeigt. Bei in vitro Unter-
suchungen an Leukozyten aus dem peripheren Blut erwiesen sich adhärente Monozyten als
gut infizierbar, während polymorphkernige Leukozyten und nicht-adhärente Lymphozyten
nicht bzw. nur zu einem sehr geringen Teil infiziert werden konnten (Midulla 1989).
In Verbindung mit einer RSV-Infektion in vitro wurden auch Veränderungen des Zytokin-
profils und der Expression von Oberflächenmarkern für die verschiedenen Zelltypen
beschrieben. Für epitheliale Atemwegs-Zelllinien (A549, BEAS u.a.) ist beschrieben, dass sie
nach RSV-Infektion in vitro verschiedene proinflammatorische Zytokine, darunter IL-6 und
IL-8 (Noah 1993; Arnold 1994) sowie die Chemokine RANTES, MCP-1 und
MIP-1α (Becker 1997; Olszewska 1998) verstärkt exprimieren. Darüber hinaus war die

EINLEITUNG 13
Expression von Adhäsionsmolekülen wie ICAM-1 and VCAM-1, die an der Migration von
Leukozyten an den Ort einer Infektion beteiligt sind, auf epithelialen Zellen nach RSV-
Infektion in vitro signifikant erhöht (Arnold 1996; Wang 2000).
Bei Alveolarmakrophagen und Monozyten zeigte sich nach RSV-Infektion in vitro ebenfalls
eine Induktion oder signifikante Zunahme von IL-6, IL-8 und TNF-α (Becker 1991; Arnold
1995). Darüber hinaus wurde in Alveolarmakrophagen auch eine vermehrte Expression von
IL-10 nach RSV-Infektion in vitro nachgewiesen und als Ursache für eine unvollständige
Immunität nach einer RSV-Erstinfektion diskutiert (Panuska 1995). Diaz (1999) untersuchte
den Einfluss einer RSV-Infektion im Vergleich zu einer Adenovirus(AdV)-Infektion in vitro
an unseparierten PBMC und fand erhöhte IL-10 Proteinkonzentrationen in den
Kulturüberständen der RSV-infizierten PBMC, die zusammen mit nur geringfügig
gesteigerten IFN-γ Konzentrationen zu einem erhöhten IL-10/IFN-γ Quotienten führten. Im
Gegensatz dazu wiesen die AdV-infizierten Zellen eine geringere IL-10 Zunahme und eine
stärkere IFN-γ Zunahme auf, woraus sich ein im Vergleich zu den uninfizierten Kontrollen
erniedrigter IL-10/IFN-γ Quotient ergab.
In Bezug auf T-Zellen gibt es bisher nur zwei Berichte zu einer RSV-Infektion in vitro.
Thurau (1998) ermittelte in RSV-infizierten PBMC in vitro die Infektionsraten verschiedener
PBMC-Subpopulationen mittels intrazellulärer durchflusszytometrischer Analysen durch
Doppelmarkierungen mit zelltyp-spezifischen und RSV-spezifischen Antikörpern und fand
durchschnittlich 4 % RSV-positive CD3+ Zellen, wenn eine relativ hohe Anzahl an
Viruspartikeln (multiplicity of infection = m.o.i. von 3) zugesetzt wurde. Bei Differenzierung
zwischen CD8+ und CD4+ Zellen zeigte sich, dass es sich bei den RSV-positiven T-Zellen
überwiegend um CD8+ Zellen handelte. An der Zunahme IL-5 produzierender CD8+ Zellen
nach Stimulierung mit PMA und Ionomycin in den RSV-infizierten PBMC im Vergleich zu
den uninfizierten Kulturen waren die RSV-positiven CD8+ Zellen jedoch nicht beteiligt, so
dass dieser Effekt als APC-vermittelt diskutiert wurde. Diaz (1999) ermittelte in CD4+- oder
CD8+-isolierten PBMC-Subpopulationen nach einer RSV-Infektion in vitro mit m.o.i 1 einen
Anteil RSV-positiver T-Zellen von 6 % bzw. 5 %. Eine zelltyp-spezifische Markierung
erfolgte dabei nicht und die Reinheit der isolierten Subpopulationen wurde mit jeweils 95 %
angegeben. Die in der gleichen Arbeit durchgeführten Zytokinmessungen erfolgten in
unseparierten PBMC und erlauben daher keine Rückschlüsse auf den produzierenden Zelltyp.

EINLEITUNG 14
1.3.3 Die spezifische T-Zellantwort auf eine RSV-Infektion
Die Auswirkung einer RSV-Infektion auf die Induktion RSV-spezifischer T-Zellen ist im
Mausmodell eingehend untersucht worden (Übersicht in Varga 2002). Diese Untersuchungen
standen in Zusammenhang mit den missglückten Impfversuchen mit Formalin-inaktivertem
RSV, die in den 60er Jahren durchgeführt worden waren, aber bei späteren natürlichen
Reinfektionen zu erhöhter Morbidität im Vergleich zu nicht geimpften Kontrollen geführt
hatten. Es konnte gezeigt werden, dass die einzelnen RSV-Proteine sehr verschiedene
Immunantworten erzeugen. So ist das Oberflächenprotein-G von RSV an der Induktion
spezifischer CD4+ T-Zellen mit TH2-Charakteristik beteiligt. Eine Aktivierung von
spezifischen CD8+ T-Zellen konnte in diesem Zusammenhang nicht beobachtet werden.
RSV-F hingegen führt zur klonalen Expansion spezifischer CD4+ T-Zellen mit TH1
Zytokinprofil und CD8+ zytotoxischer T-Zellen (Alwan 1993). Darüber hinaus wurde durch
Depletions- und Transfer-Modelle die besondere Bedeutung CD8+ T-Zellen für die Kontrolle
der RSV-induzierten Immunpathologie in zuvor vakzinierten (Hussel 1997) oder
sensibilisierten Mäusen (Schwarze 1999) eindrücklich belegt. Hussell (1998) zeigte aber
auch, dass die Immunantwort in Abhängigkeit vom genetischen Background der untersuchten
Mäuse differierte.
Einige Teilaspekte sind auch für das humane System bereits untersucht. RSV-spezifische
humane CD4+ T-Zelllinien exprimierten je nach Art des für die Generierung verwendeten
RSV-Antigens unterschiedliche Zytokinprofile. Durch lebendes RSV oder F-Protein
generierte spezifische T-Zelllinien produzierten überwiegend IL-2 und IFN-γ, während
G-Protein-spezifische T-Zellklone IL-4 und IL-10 synthetisierten. Aus Stimulierung mit
Formalin-inaktiviertem RSV hervorgegangene T-Zellen exprimierten ein gemischtes
Zytokinprofil (Jackson 1996). Bei Analysen des humanen Repertoires an RSV-spezifischen
zytotoxischen T-Zellen wurden wie in der Maus keine G-Protein spezifischen CD8+ T-Zellen
entdeckt (Cherrie 1992; Openshaw 1995).
Eine direkte Bindung von RSV an T-Zellen und der möglicherweise daraus entstehende
Einfluss auf die Immunantwort sind außer in dem erwähnten Umfang in den Arbeiten von
Thurau (1998) und Diaz (1999) bisher nicht beschrieben. Allgemein wird angenommen, dass
neben direkten Schäden durch das Virus, bei einer RSV-Infektion der unteren Atemwege
(RSV-LRTI) im Kindesalter auch die durch das Virus ausgelöste Immunantwort entscheidend
an der Pathogenese der Erkrankung beteiligt ist (Simoes 2001). Mehrere Studien belegten,

EINLEITUNG 15
dass eine RSV-Bronchiolitis bei Kleinkindern mit einem erhöhten Risiko bronchial-
obstruktiver Symptome wie Keuchen und verminderter Lungenfunktion in den Folgejahren
einhergeht (Übersicht in Sigurs 2001). Im Alter von 13 Jahren war das Risiko jedoch nicht
mehr signifikant erhöht (Stein 1999). Alle diese Studien sind rein epidemiologisch. Die
molekularen und immunologischen Mechanismen, die hinter den Folgen der RSV-LRTI
stehen, sind aber bis heute nicht abschließend geklärt. Bont (2000) zeigte in einer Studie mit
50 Kindern, die wegen einer RSV-Bronchiolitis stationär behandelt wurden, durch
Zytokinmessungen in Blutkulturen, dass die Entwicklung von wiederkehrendem Keuchen im
Jahr nach der RSV-LRTI mit einer erhöhten IL-10 Produktion verbunden war. Dieser
Unterschied zeigte sich nicht während der akuten Infektion, sondern in der
Rekonvaleszenzphase 3 - 4 Wochen später. Die Höhe der IL-10 Antwort ex vivo korrelierte
zudem mit der Häufigkeit des Keuchens. Allerdings führte im Mausmodell eine RSV-
Infektion nach allergischer Sensibilisierung auch in IL-10 Knockout-Mäusen zu
Atemwegshyperreaktivität (Mäkelä 2002), so dass zumindest in diesem Modell IL-10 nicht
der einzige immunpathologische Faktor zu sein scheint.
1.4 Chronisch obstruktive Lungenerkrankung
Die chronisch obstruktive Lungenerkrankung (COPD) ist gekennzeichnet durch eine
fortschreitende, nicht reversible Verengung der Atemwege in Verbindung mit einem chroni-
schen Entzündungsprozess (Pauwels 2001). COPD ist derzeit die vierthäufigste Todesursache
weltweit und in ihrer Prävalenz weiter steigend (Lopez 1998). Betroffen sind insbesondere
Erwachsene über 45 Jahren. Eine Studie der Weltgesundheitsorganisation (WHO) schätzt,
dass COPD im Jahr 2020 von derzeit Platz 12 auf Platz 5 der weltweit häufigsten Krankheiten
gestiegen sein wird (WHO 2000). Dabei wurden alle Altersgruppen gleichmäßig berück-
sichtigt, so dass der Anteil bei Älteren eigentlich noch größer ist.
Rauchen ist der wichtigste Risikofaktor für die Entwicklung einer COPD. Etwa 15 – 20 %
aller aktiven Raucher entwickeln eine COPD (Vestbo 2002). Zusätzlich können auch andere
Faktoren bei der Induktion einer COPD eine Rolle spielen. Als ein genetischer
Prädispositionsfaktor ist ein erblich bedingter α1-Antitrypsin-Mangel beschrieben, der zu
einer Störung des Proteasen/Antiproteasen-Gleichgewichts führt (Eriksson 1996). Ein

EINLEITUNG 16
α1-Antitrypsin-Mangel liegt aber nur bei 1 – 2 % der COPD-Patienten vor (Mahadeva 1998).
Für einen Polymorphismus in der Promotorregion des TNF-α Gens wurde in einer
taiwanesischen Population eine Assoziation zur COPD gefunden (Huang 1997). Eine
englische Studie zeigte hingegen, dass der gleiche Polymorphismus bei Kaukasiern nicht mit
einem erhöhten Risiko für COPD verbunden war (Higham 2000).
Die Pathogenese der COPD ist erst teilweise geklärt. Generell findet man bei der COPD einen
Entzündungsprozeß, der durch eine erhöhte Anzahl aktivierter Makrophagen und die
Einwanderung von CD8+ T-Lymphozyten und neutrophilen Granulozyten in die
Atemwegsmukosa gekennzeichnet ist (Saetta 1993, 1998; O’Shaughnessy 1997). Es wird
davon ausgegangen, dass ein komplexes Zusammenspiel zwischen Zellen und löslichen
Mediatoren zu den entzündlichen Veränderungen in den kleinen Atemwegen und der daraus
resultierenden Atemwegsobstruktion führt.
Makrophagen und Neutrophile tragen unter anderem durch Freisetzung verschiedener
Chemokine und Proteinasen zur Stimulierung der Mukussekretion und Degradation des
Lungenparenchyms bei (Barnes 2000). Alveolarmakrophagen produzieren nach Aktivierung
durch inflammatorische Zytokine oder andere Agentien (z.B. Zigarettenrauch) verschiedene
Matrixmetalloproteinasen (MMP), insbesondere Collagenase-1 (MMP-1), Gelatinase B
(MMP-9) und Makrophagen-Elastase (MMP-12), aber auch Stromelysin-1 (MMP-3) und
Matrilysin (MMP-7). Dadurch können sie ein breites Spektrum an Komponenten der
extrazellulären Matrix in der Lunge hydrolysieren (Shapiro 1999). Neutrophile Granulozyten
speichern Proteinasen wie die Neutrophilen-Elastase, MMP-8 und MMP-9 in ihren Granula,
so dass bei einer Stimulierung der Zellen, die zur Ausschüttung des Granula-Inhaltes führt,
sofort relativ große Mengen an elastolytischen Enzymen freigesetzt werden (Stockley 1999).
In nicht-pathologischen Prozessen wird die Aktivität dieser Proteasen durch Inhibitoren wie
α1-Antitrypsin und verschiedene TIMPs (tissue inhibitors of matrixmetalloproteinases)
reguliert, so dass die Gewebszerstörung auf ein physiologisch notwendiges Maß, wie z.B. für
die Wanderung der Neutrophilen durch das Bindegewebe, beschränkt bleibt (Stockley 1999).
Bei der COPD kann das Gleichgewicht zwischen Proteasen und Antiproteasen in der Lunge
durch die erhöhte Anzahl und Aktivierung der Makrophagen und Neutrophilen nachhaltig
gestört werden, wodurch eine fortschreitende Gewebsschädigung (Emphysem) entsteht. In
diesem Zusammenhang konnte Finlay (1997) zeigen, dass Makrophagen aus der
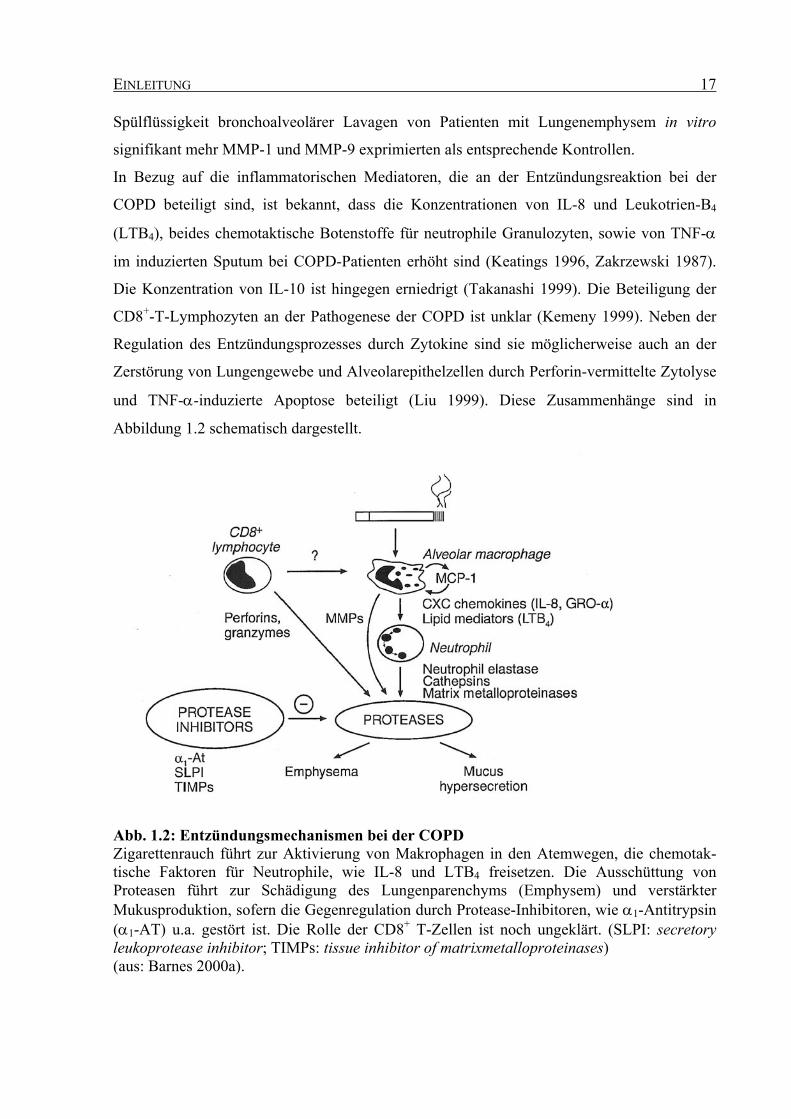
EINLEITUNG 17
Spülflüssigkeit bronchoalveolärer Lavagen von Patienten mit Lungenemphysem in vitro
signifikant mehr MMP-1 und MMP-9 exprimierten als entsprechende Kontrollen.
In Bezug auf die inflammatorischen Mediatoren, die an der Entzündungsreaktion bei der
COPD beteiligt sind, ist bekannt, dass die Konzentrationen von IL-8 und Leukotrien-B4
(LTB4), beides chemotaktische Botenstoffe für neutrophile Granulozyten, sowie von TNF-α
im induzierten Sputum bei COPD-Patienten erhöht sind (Keatings 1996, Zakrzewski 1987).
Die Konzentration von IL-10 ist hingegen erniedrigt (Takanashi 1999). Die Beteiligung der
CD8+-T-Lymphozyten an der Pathogenese der COPD ist unklar (Kemeny 1999). Neben der
Regulation des Entzündungsprozesses durch Zytokine sind sie möglicherweise auch an der
Zerstörung von Lungengewebe und Alveolarepithelzellen durch Perforin-vermittelte Zytolyse
und TNF-α-induzierte Apoptose beteiligt (Liu 1999). Diese Zusammenhänge sind in
Abbildung 1.2 schematisch dargestellt.
Abb. 1.2: Entzündungsmechanismen bei der COPD Zigarettenrauch führt zur Aktivierung von Makrophagen in den Atemwegen, die chemotak-tische Faktoren für Neutrophile, wie IL-8 und LTB4 freisetzen. Die Ausschüttung von Proteasen führt zur Schädigung des Lungenparenchyms (Emphysem) und verstärkter Mukusproduktion, sofern die Gegenregulation durch Protease-Inhibitoren, wie α1-Antitrypsin (α1-AT) u.a. gestört ist. Die Rolle der CD8+ T-Zellen ist noch ungeklärt. (SLPI: secretory leukoprotease inhibitor; TIMPs: tissue inhibitor of matrixmetalloproteinases) (aus: Barnes 2000a).

EINLEITUNG 18
Die klinische Manifestation der COPD ist zunächst durch Husten und Auswurf
gekennzeichnet. Im weiteren Verlauf tritt dann eine Atemwegsobstruktion hinzu, die je nach
Schweregrad der COPD periodisch wiederkehrenden Verschlechterungen (Exazerbationen)
unterliegt. Diese Exazerbationen sind mit einer Verminderung der Lungenfunktion und einer
Verringerung der allgemeinen Lebensqualität verbunden (Pauwels 2001). Während die
Bedeutung atemwegspathogener Bakterien sowohl für die Pathogenese der COPD als auch für
die Exazerbation bereits eingehend beschrieben ist (Sethi 2000, 2000a; Wilson 2000), wird
die Rolle viraler Infektionen bei der COPD zur Zeit intensiv untersucht und diskutiert. Erste
Ergebnisse zeigten einen Zusammenhang zwischen Exazerbationen und dem Nachweis von
Atemwegsviren insbesondere für Rhinovirus, RSV und Influenzavirus in einer Längs-
schnittuntersuchung (Seemungal 2001) und in einer Fall-Kontroll-Studie (Rohde 2002).
Durch Verwendung der Polymerase-Kettenreaktion (PCR) für den Virusnachweis konnten
dabei auch sehr geringe Mengen viraler Nukleinsäuren nachgewiesen werden. Ungeklärt blieb
jedoch, ob der Virusnachweis immer eine akute Infektion anzeigte oder auch Ausdruck einer
persistierenden Infektion sein könnte (Hogg 2001; Borg 2002). Für Adenoviren ist ein
möglicher Zusammenhang zwischen einer persistierenden Infektion und steroid-resistentem
Asthma im Kindesalter (Macek 1994) bzw. COPD bei Erwachsenen (Matsuse 1992) bereits
beschrieben worden. In beiden Fällen wurde Adenovirusprotein in den Atemwegen ohne
Anzeichen für eine akute Adenovirusinfektion bei den Patienten nachgewiesen, nicht jedoch
bei entsprechenden Kontrollpersonen. In ergänzenden in vitro Untersuchungen hierzu wurde
gezeigt, dass Bronchialepithelzellen, die das adenovirale E1A-Gen in ihrem Genom tragen,
vermehrt IL-8 und ICAM-1 produzieren (Hogg 1999).
Für RSV wurde im Tiermodell eine persistierende Infektion mit begleitenden Veränderungen
in der Lungenstruktur und –funktion bei Meerschweinchen in vivo nachgewiesen (Dakhama
1997). In weiteren Tiermodellen zur RSV-Infektion wird überwiegend die virusinduzierte
Entstehung von Asthma bronchiale untersucht. In einem Mausmodell der RSV-Infektion mit
anschließender allergischer Sensibilisierung über die Atemwege konnte gezeigt werden, dass
RSV-Infektion zu einer Atemwegsentzündung mit Atemwegshyperreaktivität (AHR) führt.
Dabei war die Atemwegsentzündung T-Zell vermittelt und die Entstehung der AHR abhängig
von CD8+ T-Zellen als Regulatoren, von IL-5 als Mediator und von eosinophilen Granulo-
zyten als Effektoren (Schwarze 1999a, 1999b).
Durch die steigende Prävalenz der COPD (s. oben) wird diese Erkrankung bei immer mehr
Menschen die Lebensqualität entscheidend verringern und zu einer großen Belastung des

EINLEITUNG 19
Gesundheitswesens führen. Fortschritte im Verständnis der pathogenetischen Zusammen-
hänge und der daraus entstehenden Entwicklung neuer Therapieansätze sind daher dringend
gefordert (Croxton 2002). In diesem Zusammenhang wurde 1998 die Klinische
Forschergruppe „Bedeutung von Virusinfektionen bei akuten und chronischen
Atemwegserkrankungen im Kindes- und Erwachsenenalter“ ins Leben gerufen. In diesem
durch das BMBF-geförderten Verbundprojekt der Ruhr-Universität Bochum werden
klinische, morphologische, zellbiologische und molekulare Untersuchungen zur Aufklärung
der Kausalzusammenhänge zwischen Virusinfekten und dem klinischen Krankheitsbild der
COPD und des Asthma bronchiale durchgeführt. Auch die oben erwähnte klinische COPD-
Studie (Rohde 2002) wurde im Rahmen dieses Projektes initiiert. In einem Teilprojekt mit
zellbiologisch / immunologischem Schwerpunkt wurden RSV-induzierte Veränderungen in
Kokulturen aus humanen Alveolarmakrophagen und autologen T-Zellen in vitro von der
Verfasserin untersucht. Dabei zeigte sich, dass nicht nur Makrophagen, sondern vermutlich
auch T-Zellen von RSV infiziert werden können. Diese Beobachtung war ein entscheidender
Ausgangspunkt dieser Arbeit.
1.5 Zielsetzung
Zielsetzung der hier vorliegenden Arbeit war es, eine in vitro beobachtete direkte Interaktion
zwischen RSV und T-Zellen systematisch zu untersuchen. Durch Analysen im Durchfluss-
zytometer und konfokalen Laser-Rastermikroskop sollte die Interaktion eingehend charak-
terisiert werden. Dazu war es nötig, durch Kombination verschiedener Antikörper eine
Markierung zu finden, die für die konfokale Immunfluoreszenz-Mikroskopie geeignet war
und zudem neben dem Nachweis der RSV-Antigene auch eine eindeutige Identifizierung des
Zelltyps ermöglichte. Insbesondere sollte geklärt werden, ob das Virus auch in die Zellen
eindringt und - sofern dies bestätigt werden konnte - ob eine Replikation von RSV in den
T-Zellen stattfindet.
Darüber hinaus sollte als zentrale Fragestellung untersucht werden, inwieweit dieser Virus-
Zell-Interaktion auch funktionelle Bedeutung zukommt. Dazu war zunächst der Reinheitsgrad
der T-Zellpopulation insbesondere in Bezug auf den Anteil an Monozyten zu verbessern. Im
Anschluss daran sollte eine quantitative real-time PCR für humanes Interleukin-10 als sehr

EINLEITUNG 20
sensitive Meßmethode etabliert werden, um eine mögliche Zunahme der Interleukin-10
Expression als Reaktion der T-Zellen auf den Viruskontakt untersuchen zu können. Parallel
dazu sollten die Konzentration von IL-10 und ggf. weiteren Zytokinen in den Überständen
von RSV-infizierten T-Zellkulturen mittels ELISA bestimmt werden.
Die Untersuchungen der reinen T-Zellkulturen sollten zudem um Kokultur-Experimente
ergänzt werden, bei denen aufgereinigte T-Zellen zum einen mit CD14+ Monozyten aus dem
peripheren Blut und zum anderen mit Alveolarmakrophagen aus der Spülflüssigkeit broncho-
alveolärer Lavagen kultiviert wurden. Die Ergebnisse dieser Experimente sollten vor dem
Hintergrund einer möglichen Bedeutung von RSV-Infektionen für Exazerbationen der COPD
und der noch ungeklärten Rolle der in die Lunge eingewanderten T-Zellen in der Pathogenese
dieser Erkrankung diskutiert werden.

21
2. Material und Methoden
2.1 Material
2.1.1 Spenderblut
Für die Zellisolierung wurde venöses Blut von freiwilligen erwachsenen Spendern gewonnen,
deren Daten unter Einhaltung der Vorschriften des Datenschutzes in einer Spenderdatei der
Abteilung für Experimentelle Pneumologie geführt werden. Die Blutentnahmen sind von der
Ethikkommission der Medizinischen Fakultät der Ruhr-Universität Bochum genehmigt
(Registrier-Nr. 1486).
2.1.2 Virusmaterial
Respiratorisches Syncytiales Virus (RSV), Stamm Long (ATCC VR-26) wurde zur Anzucht
(s. 2.2.2) von der American Type Culture Collection (ATCC, Manassas, Virginia, USA)
bezogen.
Newcastle Disease Virus (NDV) wurde freundlicherweise durch Herrn Prof. R. Zawatzky
(Deutsches Krebsforschungszentrum, Heidelberg) zur Verfügung gestellt (Stock-Konzen-
tration 3200 HA/ml) und bei –70° C gelagert.
2.1.3 Medien und Lösungen
Medium für die Zellkultur:
RPMI 1640 (Life Technologies, Karlsruhe)
+ 1 % v/v L-Glutamin-Lösung, 200 mM (Biochrom, Berlin)
+ 1 % v/v Penicillin-/Streptomycin-Lösung, 10000 U / 10 mg/ml (Biochrom, Berlin)
+ 5 % FCS (Biochrom, Berlin), 30 min. hitzeinaktiviert bei 56 °C

MATERIAL UND METHODEN 22
Medium für die Virusanzucht:
DMEM mit 25 mM HEPES (AppliChem, Darmstadt)
+ 1 % v/v Penicillin-/Neomycin-/Streptomycin-Lösung, 5 / 10 / 5 mg/ml (Gibco, Karlsruhe)
+ 0,5 % FCS (ICN Biomedicals, Eschwege), 30 min. hitzeinaktiviert bei 56 °C
LB-Medium für die Bakterienkultur:
1,0 % w/v Trypton
0,5 % w/v Hefeextrakt
0,5 % w/v NaCl
Zur Herstellung von Agarplatten wurde 1,5 % w/v Agar zugefügt.
Als Antibiotikum-Zusatz wurde Ampicillin (Endkonzentration 100 µg/ml) oder Kanamycin
(Endkonzentration 50 µg/ml) verwendet (s. jeweils Angabe im Text).
Phosphat-gepufferte-Kochsalzlösung (PBS):
137 mM Natriumchlorid (NaCl)
2,7 mM Kaliumchlorid (KCl)
10 mM di-Natriumhydrogenphosphat (Na2HPO4)
2 mM Kaliumdihydrogenphosphat (KH2PO4)
PBS für die magnetische Zellsortierung:
PBS
+ 2 mM EDTA
+ 0,5 % w/v BSA
Puffer für die Agarose-Gelelektrophorese:
Laufpuffer (TBE)
89 mM Tris
89 mM Borsäure
2 mM EDTA

MATERIAL UND METHODEN 23
Ladepuffer
TBE
+ 0,25 % w/v Bromphenolblau
+ 15 % w/v Ficoll
2.1.4 Antikörper
für die magnetische Zellsortierung verwendete monoklonale Antikörper:
antiCD3-MicroBeads, Klon UCHT1, IgG1 (Miltenyi Biotec, Bergisch Gladbach)
antiCD14-MicroBeads, Klon UCHM1, IgG2a (Miltenyi Biotec)
für die Zellkultur verwendete monoklonale Antikörper:
antiCD28, Klon CD28.2, IgG1 (Pharmingen, Heidelberg)
antiICAM-1, Klon 15.2, IgG1 (Dianova, Hamburg)
für die Durchflusszytometrie verwendete Antikörper:
monoklonale Antikörper:
antiCD3-PE, Klon Leu-4/SK7, IgG1 (Becton Dickinson, Heidelberg)
antiCD4-PE, Klon Leu-3a/SK3, IgG1 (Becton Dickinson)
antiCD8-PE, Klon Leu-2a, IgG1 (Becton Dickinson)
antiCD14-FITC, Klon TÜK4, IgG2a (Miltenyi Biotec)
antiCD69-PE, Klon FN50, IgG1 (Dianova)
Isotypkontrollen:
IgG1-PE, Klon X40 (Becton Dickinson)
IgG2a-FITC, Klon 5A7 (Dianova)
polyklonale Antikörper:
antiRSV-FITC Ziegenserum (Dunn Labortechnik, Asbach)
Sofern nicht anders angegeben, handelt es sich bei allen monoklonalen Antikörpern um Anti-
körper aus der Maus.

MATERIAL UND METHODEN 24
2.1.5 Reagenzien
Agarose Biozym, Hessisch Oldenburg
Ampicillin Sigma, Taufkirchen
β-Mercaptoethanol Sigma
Borsäure Merck, Darmstadt
Bromphenolblau Riedel-de Haën, Seelze
BSA Sigma
di-Natriumhydrogenphosphat Merck
EDTA Sigma
Ethidiumbromid Amresco, Solon, Ohio, USA
Ficoll/Isopaque (1,077 g/ml) PAN Biotech, Aidenbach
Hanks’ Balanced Salt Solution (HBSS) Biochrom, Berlin
Hefeextrakt Roth, Karlsruhe
Kaliumchlorid Riedel-de Haën
Kaliumdihydrogenphosphat Merck
Kanamycin Sigma
Natriumazid Merck
Natriumchlorid Merck
Natriumheparin Ratiopharm, Ulm
Paraformaldehyd Merck
Phytohämagglutinin (PHA) Biochrom
Tetramethylbenzidin-(TMB-)Substrat Pharmingen, Heidelberg
Tris Serva, Heidelberg
Trypton Roth

MATERIAL UND METHODEN 25
2.2 Methoden
2.2.1 Zellpräparation
2.2.1.1 Isolierung mononukleärer Zellen des peripheren Blutes
Venöses Blut von erwachsenen Spendern wurde unmittelbar nach der Entnahme mit
20 I.E./ml Natriumheparin versetzt, um die Koagulation zu verhindern. Das heparinisierte
Blut wurde 1:2 mit HBSS verdünnt und mit Ficoll unterschichtet. Die Isolierung der mono-
nukleären Zellen des peripheren Blutes (PBMC) erfolgte über eine Dichtegradienten-
Zentrifugation nach Böyum (1968) unter sterilen Bedingungen. Nach der Zentrifugation
(460 x g, 20 min., 25° C) wurden der Interphasering, der die PBMC enthält, mittels Pasteur-
pipette abgenommen, die Zellen zweimal in HBSS gewaschen (180 x g, 10 min., 4° C) und je
nach weiterer Verwendung in PBS oder Zellkulturmedium resuspendiert.
2.2.1.2 Magnetische Zellsortierung
Die Aufreinigung von T-Zellen aus PBMC erfolgte über das magnetische Zellsortierungs-
system ‚MidiMACS’ (Miltenyi Biotec, Bergisch Gladbach) nach dem Protokoll des Herstel-
lers. Die wesentlichen Bestandteile des Systems sind ein starker Permanentmagnet, Trenn-
säulen mit einer ferromagnetischen Matrix und Mikromagnetpartikel, die an monoklonale
Antikörper gekoppelt werden können (Miltenyi 1990). Zur Vorbereitung der Separation
wurden 100 bis 150 Mio. PBMC in einer Konzentration von 108 Zellen/ml in PBS mit
antiCD3-Antikörpern, an die Mikromagnetpartikel gekoppelt waren (CD3-MicroBeads,
Miltenyi Biotech), 15 min. bei 4° C inkubiert, in PBS gewaschen (180 x g, 10 min., 4° C) und
resuspendiert. Die Zellsuspension wurde unter sterilen Bedingungen auf eine MidiMACS-
Trennsäule mit einer Kapazität von 109 Gesamtzellen und 108 markierten Zellen aufgetragen
und die Sortierung nach dem Protokoll des Herstellers für positive Selektion durchgeführt.
Dabei wurden die mit CD3-MicroBeads markierten Zellen in der Säulenmatrix durch das
umgebende Magnetfeld zurückgehalten, während unmarkierte Zellen durch anschließendes
Spülen der Säule mit PBS als negative Zellfraktion eluiert wurden. Nach Entfernen der Säule

MATERIAL UND METHODEN 26
aus dem Magnetfeld wurde die markierte, CD3-positive Zellfraktion ebenfalls durch Spülen
mit PBS aus der Säule gewonnen. Die so aufgereinigten Zellen werden im folgenden als
CD3+ T-Zellen bezeichnet.
Wurden sowohl CD14+ Monozyten als auch CD3+ T-Zellen in hoher Reinheit für ein Experi-
ment benötigt, so erfolgte eine zweistufige Sortierung mit einer vorgeschalteten Depletion.
Diese Sortierung wurde in einem AutoMACS-System durchgeführt, das nach dem gleichen
Prinzip wie das oben beschriebene MidiMACS-System arbeitet, bei dem aber Proben-
aufnahme und Spülvorgänge automatisiert ablaufen. Zudem kann die Stärke des Magnetfeldes
und die Durchflussgeschwindigkeit bei der Säulenpassage bei diesem Gerät durch Auswahl
aus verschiedenen Sortierprogrammen reguliert werden. Die PBMC wurden in einem ersten
Schritt analog der oben beschriebenen CD3-Markierung mit CD14-MicroBeads markiert und
mit dem Programm ‚sensitive Depletion’ im AutoMACS sortiert. Bei einer Depletion wird
das System so eingestellt, dass auch schwach markierte Zellen in der Säule zurückgehalten
werden, sodass die negative Zellfraktion nur CD14-negative Zellen enthielt. Die positive Zell-
fraktion wurde danach noch einmal mit dem Programm ‚positive Selektion’ sortiert, um
schwach positive Zellen zu entfernen und eine Zellfraktion mit deutlich CD14-positiven
Zellen zu erhalten. Die so gewonnene Zellpopulation wird im folgenden als CD14+ Mono-
zyten bezeichnet. Die CD14-negative Fraktion wurde in einem zweiten Schritt mit
CD3-MicroBeads markiert und anschließend wie oben beschrieben sortiert. Diese zweistufig
sortierten Zellen werden im folgenden als CD3+14- T-Zellen bezeichnet. Das Sortierschema
ist in Abbildung 2.1 im Überblick dargestellt.

MATERIAL UND METHODEN 27
Abb. 2.1: Sortierschema der magnetischen Zellsortierung
Zur Bestimmung der Effektivität der Aufreinigungen wurden die resultierenden Zellfrak-
tionen qualitativ hinsichtlich ihrer Reinheit im Durchflusszytometer (s. 2.2.5) und quantitativ
hinsichtlich der erzielten Zellausbeute durch Zellzählung (s. 2.2.1.4) analysiert.
2.2.1.3 Gewinnung von Alveolarmakrophagen
Alveolarmakrophagen (AM) wurden aus der Spülflüssigkeit bronchoalveolärer Lavagen
(BAL) gewonnen, die in der Abteilung für Pneumologie, Allergologie und Schlafmedizin der
Berufsgenossenschaftlichen Kliniken Bergmannsheil, Bochum zu diagnostischen Zwecken
durchgeführt wurden. Eine schriftliche Einverständniserklärung der Patienten lag jeweils vor.
Die BAL-Zellen wurden in einem Routinelabor der Abteilung durch Zentrifugation (180 x g,
10 min., 4° C) und zweimaliges Waschen in PBS aus der Spülflüssigkeit gewonnen, gezählt
und durch eine Pappenheim-Färbung nach Zelltypen differenziert. Der nicht für routine-
mäßige Analysen benötigte Teil der Zellen stand für in vitro Experimente zur Verfügung. Die
Zellen wurden in Zellkulturmedium (s. 2.1.3) unter Zusatz von Amphotericin B (Endkonzen-
tration 0,25 µg/ml) als Antimykotikum resuspendiert und unter Berücksichtigung des Ergeb-
nisses der Zelldifferenzierung auf eine Konzentration von 106 AM/ml eingestellt. Die Abtren-
nung von Lymphozyten und Granulozyten erfolgte durch Adhärenz der Alveolarmakrophagen
an die Plastikoberfläche der Zellkulturplatten. Nach Inkubation von je 500 µl Zellsuspension

MATERIAL UND METHODEN 28
pro Vertiefung in 24-Loch-Zellkulturplatten (Costar, Bodenheim) für 1h bei 37° C, 5 % CO2
wurden die nicht adhärenten Zellen zusammen mit dem Kulturmedium abgenommen und
verworfen. Die adhärenten Makrophagen wurden zweimal mit frischem Medium in der
Kulturplatte gewaschen und entsprechend dem gewünschten Versuchsansatz weiter inkubiert.
2.2.1.4 Zellzählung und Vitalitätsbestimmung
Zur Bestimmung der Zellzahl wurden 10 µl Zellsuspension (Zellen in PBS oder Medium)
1:10 in Türks-Lösung (Merck, Darmstadt) verdünnt und eine Neubauer-Zählkammer mit
dieser Färbelösung gefüllt. Aus dem Ergebnis der Zählung aller Zellen in den vier Eck-
quadraten unter einem Lichtmikroskop (BH-2, Olympus, Hamburg) konnte die Gesamt-
zellzahl pro Milliliter nach folgender Formel berechnet werden:
Summe Eckquadrate : 4 x 104 (Kammerfaktor) x 10 (Verdünnungsfaktor) = Zellzahl/ml.
Alternativ wurden Zellzählungen automatisiert mit einem Sysmex F-820 Zellzählgerät
(Sysmex Medical Electronics, Norderstedt) nach Anleitung des Herstellers durchgeführt.
Zur Bestimmung der Zellvitalität wurden 50 µl Zellsuspension mit 10 µl Trypanblau-Lösung
(0,5 % w/v in 0,9 % NaCl) 5 min. bei Raumtemperatur inkubiert. In dieser Zeit färbt der
Farbstoff tote Zellen blauviolett an, lebende Zellen jedoch nicht, da die Zellmembran lebender
Zellen nicht permeabel für den Farbstoff ist. Die Vitalität einer Zellfraktion wurde durch
Auszählung von 300 Zellen unter dem Lichtmikroskop ermittelt und als Anteil lebender
Zellen in % angegeben.
2.2.2 Virusanzucht
Die Anzucht des Respiratorischen Syncytial Virus (RSV) erfolgte in HEp-2 Zellen, einer
adhärent wachsenden humanen Larynx-Karzinom-Zelllinie (ATCC CCL-23). Semikonfluent
bewachsene HEp-2 Kulturen wurden mit einer multiplicity of infection (m.o.i.) von 0,1 mit
RSV infiziert, d.h. pro 10 Zellen wurde ein infektiöses Viruspartikel der Kultur zugesetzt. Zur
Virusanlagerung wurden die Zellen zunächst in DMEM (s. 2.1.3) ohne FCS 2h bei 37° C
ohne CO2-Zufuhr inkubiert, sodann wurde das Medium gegen DMEM mit 0,5 % FCS ausge-
tauscht und die Zellen weiter inkubiert. Nach 72h konnten die infizierten Zellen mit einem

MATERIAL UND METHODEN 29
Zellschaber vom Boden der Kulturflaschen gelöst und zusammen mit dem Kulturüberstand
gesammelt und zentrifugiert werden (300 x g, 10 min., Raumtemperatur). Der Überstand
wurde entweder direkt als RSV-Suspension eingesetzt oder für die Aufreinigung der RSV-
Partikel mittels Ultrazentrifugation durch ein 10 %iges Sucrose-Kissen (60.000 x g, 2 h, 8° C)
verwendet. Der Virustiter des Kulturüberstandes oder der aufgereinigten RSV-Suspension
wurde durch immunzytochemische Färbung in limitierenden Verdünnungsexperimenten mit
HEp-2 Zellen nach der Methode von Reed und Münch als 50 % tissue culture infectious dose
(TCID50) bestimmt. Ohne Aufreinigung wurden Titer bis zu einer TCID50 von 6 x 106/ml,
nach Aufreinigung von 1 x 107 bis 2 x 108/ml erzielt. Bis zur Verwendung für die in vitro
Experimente wurde das Virusmaterial portioniert bei –70° C gelagert. Einmal aufgetautes
Material wurde nicht wieder eingefroren.
2.2.3 Kultivierung und Stimulation der Zellen
Für funktionelle Untersuchungen zur Induktion von Zytokinen oder Expression von Oberflä-
chenmarkern wurden die verschiedenen isolierten Zellpopulationen wie PBMC, T-Zellen,
Monozyten und Alveolarmakrophagen in Zellkulturmedium (s. 2.1.3) auf eine Konzentration
von 106 Zellen/ml eingestellt und in 24-Loch-Zellkulturplatten (Costar, Bodenheim) zu je
0,5 Mio. Zellen/Loch bei 37° C, 5 % CO2 inkubiert. Je nach Fragestellung wurden folgende
Stimuli einzeln oder in Kombination eingesetzt:
- RSV in einer m.o.i. von 0,1 bis 1 bei Verwendung von RSV-Partikeln in HEp-2 Zellkultur-
überstand und in einer m.o.i. von 1 bis 6 bei Verwendung von aufgereinigten Viruspartikeln
- NDV 1:1000 verdünnt, d.h. 3,2 hämagglutinierende Einheiten pro ml Zellkultur. Diese
Konzentration hatte sich in Titrationsversuchen unserer Arbeitsgruppe als optimal für die
Stimulation von humanen Zellen erwiesen.
- antiCD28-Antikörper, Endkonzentration 1 µg/ml
- Phytohämagglutinin (PHA), Endkonzentration 2,4 µg/ml

MATERIAL UND METHODEN 30
Als Kontrollen wurden in jedem Experiment Zellen ohne Stimulus in gleicher Weise kulti-
viert. Bei Verwendung von RSV-Partikeln in HEp-2 Zellkulturüberstand wurde den Kontroll-
kulturen Überstand von nicht infizierten HEp-2 Zellen in gleicher Menge zugesetzt. Diese
Ansätze werden im folgenden als Mock-Kontrollen bezeichnet.
Die Inkubationsdauer lag zwischen 0,5 und 48 h und ist daher bei den Ergebnissen jeweils mit
angegeben.
Nach Ablauf der gewünschten Inkubationszeit wurden die Zellen durch mehrmaliges
Aufziehen und Ablassen des Kulturüberstandes mit einer Pipette vom Boden aufgewirbelt
und zusammen mit dem Überstand in ein 1,5 ml Reaktionsgefäß (Eppendorf, Hamburg) über-
führt. Parallelansätze mit gleichen Stimulationsbedingungen wurden dabei vereinigt. Nach
Zentrifugation (180 x g, 10 min., 4° C) wurden die zellfreien Überstände in frische Reakti-
onsgefäße überführt und bis zur Bestimmung der Zytokinkonzentration bei –20° C gelagert.
Die sedimentierten Zellen wurden entweder für eine anschließende Analyse im Durchfluss-
zytometer oder im konfokalen Laser-Rastermikroskop aufbereitet (s. 2.2.5 und 2.2.6) oder für
eine spätere RNA-Isolierung in entsprechendem Puffer lysiert (s. 2.2.7) und bei -70° C
eingefroren.
Alle Arbeiten mit infektiösen RSV-Partikeln wurden gemäß den gesetzlichen Bestimmungen
in Laboratorien der biologischen Sicherheitsstufe L2 durchgeführt.
2.2.4 Quantitative Zytokinbestimmungen in Kulturüberständen
Für die quantitative Bestimmung von Zytokinen in den Kulturüberständen wurde die Methode
des Enzyme-linked-immunosorbent-assay (ELISA) angewendet. Dieses Nachweisverfahren
ermöglicht die quantitative Bestimmung eines Antigens (hier IL-10 oder IFN-γ) unabhängig
von Fremdproteinen in den Kulturüberständen. Für die Bestimmung wurden ‚OptEIA-Sets’
(Pharmingen, Heidelberg) für humanes IL-10 bzw. ein ELISA-System für humanes IFN-γ
(Hoffmann-La Roche, Basel, Schweiz) verwendet, die zwei aufeinander abgestimmte Anti-
körper sowie den rekombinanten Proteinstandard enthielten. Zunächst wurde der erste anti-
genspezifische Antikörper an eine Mikrotiterplatte (Falcon® ‚ProBind’, Becton Dickinson,
Heidelberg) gebunden und freie Bindungsstellen mit BSA-haltigem Puffer abgesättigt.

MATERIAL UND METHODEN 31
Während der folgenden Inkubation mit der zu analysierenden Probe (IL-10: 100 µl Kultur-
überstand pur oder verdünnt, IFN-γ: 100 µl Kulturüberstand 1:5 oder höher verdünnt) erfolgte
die Bindung der Zytokin-Moleküle an den ersten Antikörper. Durch Bindung eines zweiten
Zytokin-spezifischen und Peroxidase-gekoppelten Antikörpers an die gebundenen Zytokin-
Moleküle erfolgte die Umsetzung eines zugefügten TMB-Substrats proportional zur gebun-
denen Zytokin-Menge. Nach Zusatz von 2 N Schwefelsäure zum Abstoppen der Reaktion
wurde die Farbentwicklung in einem Photometer für Mikrotiterplatten (Dynatech MR5000,
Dynex Technologies GmbH, Denkendorf) gemessen. Alle Inkubationen und Waschschritte
wurden gemäß dem Protokoll des Herstellers durchgeführt und sind daher hier nicht im Detail
beschrieben. Die quantitative Auswertung erfolgte unter Verwendung eines speziellen Soft-
wareprogramms für ELISA-Auswertungen (MikroWin Version 3.29, Mikrotek Laborsysteme
GmbH, Overath). Anhand einer Standardkurve aus sieben Standardkonzentrationen (IL-10:
7,8 pg/ml bis 500 pg/ml, IFN-γ: 15,6 pg/ml bis 1000 pg/ml), die bei jedem Test mitgeführt
wurden, wurde der Zytokingehalt der Proben aus den Extinktionswerten ermittelt. Lag das
Ergebnis einer Probe oberhalb des linearen Bereiches der Standardkurve, wurde die Messung
in stärkerer Verdünnung wiederholt. Als Leerwert diente der Verdünnungspuffer für die
Standard- und Probenverdünnungen. Alle Standards und Proben wurden jeweils in Doppel-
bestimmungen gemessen und der Mittelwert als Ergebnis eingesetzt.
2.2.5 Immunfluoreszenzmarkierungen und durchflusszytometrische Zellanalyse
Zur Bestimmung der Reinheit der magnetisch sortierten Zellpopulationen sowie für phäno-
typische Untersuchungen nach RSV-Exposition wurden die Zellen mit fluorochrom-
konjugierten Antikörpern markiert und in einem Durchflusszytometer (FACS Calibur, Becton
Dickinson, Heidelberg) analysiert. Dazu wurden frisch isolierte oder kultivierte Zellen mit
einer Konzentration von 2 x 106 Zellen/ml in PBS mit 0,1 % w/v Natriumazid (PBS-Azid)
resuspendiert und für jede Markierung je 2 x 105 Zellen mit fluorochrom-konjugierten zelltyp-
spezifischen Antikörpern (antiCD3, -CD4, -CD8, -CD14) oder RSV-spezifischen Antikörpern
(s. 2.1.4) für 15 - 30 min. bei 4° C inkubiert. Die für den jeweiligen Antikörper optimale
Konzentration in Bezug auf spezifisches Signal und unspezifische Hintergrundfärbung wurde
unter Berücksichtigung der Herstellerinformationen in Vorversuchen bestimmt. Um
ungebundene Antikörper zu entfernen, wurden die Zellen nach der Inkubation mit dem

MATERIAL UND METHODEN 32
10fachen Volumen an PBS-Azid gewaschen und zentrifugiert (180 x g, 10 min., 4° C). Die
Zellen wurden in PBS mit 1,5 % Paraformaldehyd fixiert und konnten so bis zur Messung im
Durchflusszytometer im Dunkeln bei 4° C bis zu einer Woche ohne Qualitätsverluste gelagert
werden.
Bei der Analyse im Durchflusszytometer werden die Zellen in Suspension durch das Prinzip
der hydrodynamischen Fokussierung in einer Messkapillare einzeln an einem senkrecht zur
Fließrichtung einfallenden Laserstrahl vorbeigeführt. Das Licht des Argon-Lasers (Wellen-
länge 488 nm) regt die Fluoreszenz der markierten Zellen an und wird zudem von den Zellen
vorwärts und seitwärts gestreut. Die drei Parameter Vorwärtsstreulicht, Seitwärtsstreulicht
und Fluoreszenz wurden für jede Zelle simultan durch entsprechende Detektoren erfasst,
elektronisch verstärkt und gespeichert. Aus jeder Probe wurden 10.000 Zellen analysiert. Die
Auswertung der Daten erfolgte durch ein spezielles Softwareprogramm (CELLQuest™,
Version 1.2, Becton Dickinson), das eine eindimensionale Darstellung der einzelnen Para-
meter in Histogrammen oder eine kombinierte Auswertung zweier Parameter in zweidimen-
sionalen Dotplots ermöglicht. Das Vorwärtsstreulicht ist ein relatives Maß für die Größe einer
Zelle, das Seitwärtsstreulicht ein Maß für die Granularität, so dass sich aus der Kombination
dieser beiden Parameter für jeden Zelltyp ein charakteristisches Streulichtmuster ergibt. Dies
ermöglicht im Streulichtdiagramm eine Abgrenzung verschiedener Zellpopulationen
(Lymphozyten, Monozyten, Granulozyten) gegeneinander und gegenüber Thrombozyten und
Zelltrümmern. Durch Setzen entsprechender Auswertefenster kann jede der Zellpopulationen
für die Auswertung der Fluoreszenz auch einzeln analysiert werden. Zur Abgrenzung der
unspezifischen Fluoreszenz bei der Analyse wurden jeweils Isotypkontrollen mitgeführt.
Dabei werden die Zellen mit Antikörpern inkubiert, die den gleichen IgG-Subtyp und die
gleiche Fluorochromkonjugation wie der jeweils spezifische Antikörper aufweisen, deren
Spezifität aber Antigene betrifft, die bei humanen Zellen nicht vorkommen. Bei der
Auswertung der Fluoreszenzmessungen wurden die Zellen als positiv gewertet, deren
Fluoreszenzsignal stärker als das der Isotypkontrollen war. Wiesen auch die Isotypkontrollen
vereinzelt Zellen mit starken Fluoreszenzsignalen auf, so wurde die Grenze zwischen positiv
und negativ so gesetzt, dass bei den Isotypkontrollen > 99 % der Zellen negativ waren. Bei
Analysen nach Kultivierung der Zellen wurden unstimulierte Zellen als Negativkontrollen
mitgeführt.

MATERIAL UND METHODEN 33
2.2.6 Zellanalyse im konfokalen Laser-Rastermikroskop
Für die Analyse der Zellen im konfokalen Laser-Rastermikroskop (CLSM) zur Lokalisation
der RSV-Partikel wurden Zytozentrifugenpräparate aus den Zellkulturen angefertigt. Die
Zellen wurden in einer Konzentration von 2,5 x 105/ml in PBS mit 20 % v/v FCS
resuspendiert und für jedes Präparat 4 x 104 Zellen mittels einer Zytozentrifuge (Cytospin 3,
Shandon, Pittsburgh, USA) auf Glasobjektträger aufgebracht (55 x g, 2 min., Raumtempe-
ratur). Danach wurden die Präparate direkt in PBS mit 10 % w/v Paraformaldehyd 10 min. bei
Raumtemperatur fixiert, zweimal mit PBS gewaschen und bis zur weiteren Bearbeitung in
PBS bei 4° C aufbewahrt (max. 1 Woche). Die auf den Glasobjektträgern fixierten Zellen
wurden im Institut für Pathologie der Ruhr-Universität Bochum mit verschiedenen
Kombinationen fluorochrom-konjugierter Antikörper markiert und anschließend in einem
CLSM (Leica Microsystems, Bensheim) analysiert. Ziel war es, eine Immunfluoreszenz-
Markierung zu finden, die neben dem Nachweis der RSV-Antigene auch eine eindeutige
Identifizierung des Zelltyps ermöglichte. Die ausgetesteten Antikörper-Kombinationen sind in
Tabelle 2.1 aufgeführt. Vor Inkubation mit den Sekundärantikörpern wurden die Proben mit
Serum der jeweiligen Spezies, aus der der Sekundärantikörper stammte, abgesättigt.
Im Gegensatz zu normalen Licht-Mikroskopen ermöglicht ein CLSM eine schichtweise
Durchleuchtung der Präparate. Daher sollte neben der Überprüfung der Ergebnisse aus der
Analyse im Durchflusszytometer mit dieser Methode auch Aufschluss über eine mögliche
intrazelluläre Lokalisation der RSV-Partikel gewonnen werden. Die Auswertung und
fotografische Dokumentation erfolgte mit freundlicher Unterstützung durch Herrn O. Anhenn
vom Institut für Pathologie der Ruhr-Universität Bochum.
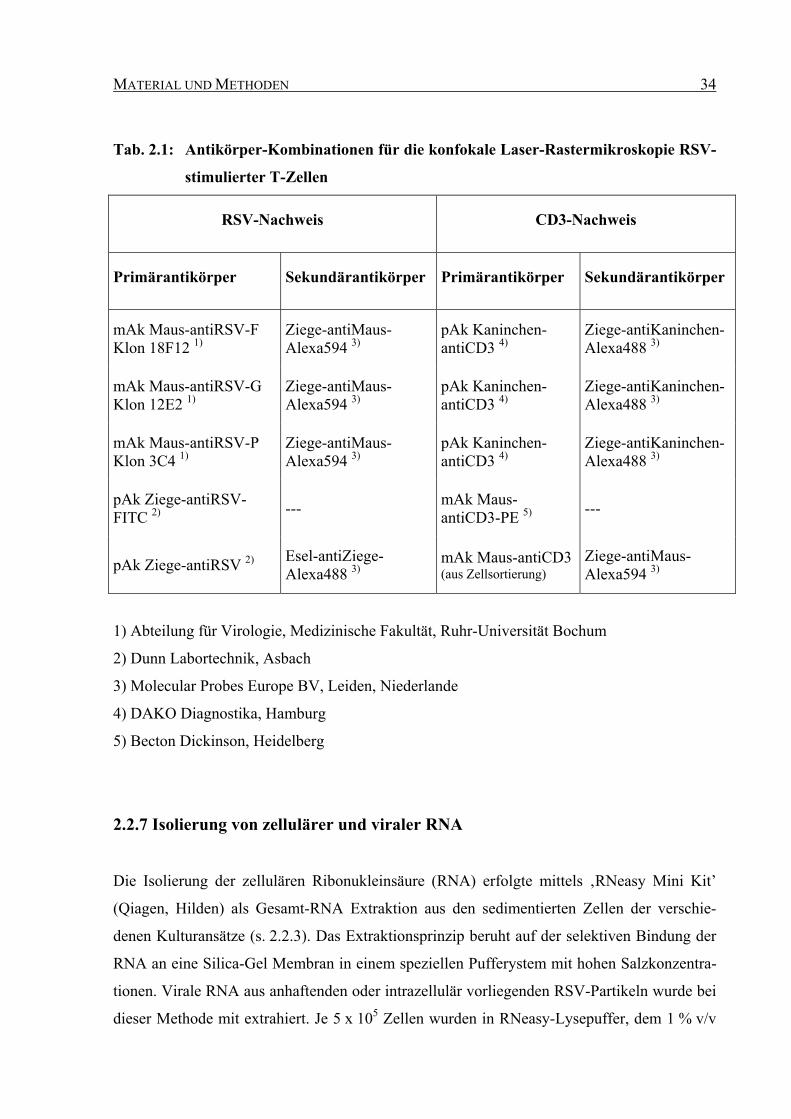
MATERIAL UND METHODEN 34
Tab. 2.1: Antikörper-Kombinationen für die konfokale Laser-Rastermikroskopie RSV-
stimulierter T-Zellen
RSV-Nachweis CD3-Nachweis
Primärantikörper Sekundärantikörper Primärantikörper Sekundärantikörper
mAk Maus-antiRSV-F Klon 18F12 1)
Ziege-antiMaus-Alexa594 3)
pAk Kaninchen-antiCD3 4)
Ziege-antiKaninchen-Alexa488 3)
mAk Maus-antiRSV-G Klon 12E2 1)
Ziege-antiMaus-Alexa594 3)
pAk Kaninchen-antiCD3 4)
Ziege-antiKaninchen-Alexa488 3)
mAk Maus-antiRSV-P Klon 3C4 1)
Ziege-antiMaus-Alexa594 3)
pAk Kaninchen-antiCD3 4)
Ziege-antiKaninchen-Alexa488 3)
pAk Ziege-antiRSV-FITC 2) --- mAk Maus-
antiCD3-PE 5) ---
pAk Ziege-antiRSV 2) Esel-antiZiege-Alexa488 3)
mAk Maus-antiCD3 (aus Zellsortierung)
Ziege-antiMaus-Alexa594 3)
1) Abteilung für Virologie, Medizinische Fakultät, Ruhr-Universität Bochum
2) Dunn Labortechnik, Asbach
3) Molecular Probes Europe BV, Leiden, Niederlande
4) DAKO Diagnostika, Hamburg
5) Becton Dickinson, Heidelberg
2.2.7 Isolierung von zellulärer und viraler RNA
Die Isolierung der zellulären Ribonukleinsäure (RNA) erfolgte mittels ‚RNeasy Mini Kit’
(Qiagen, Hilden) als Gesamt-RNA Extraktion aus den sedimentierten Zellen der verschie-
denen Kulturansätze (s. 2.2.3). Das Extraktionsprinzip beruht auf der selektiven Bindung der
RNA an eine Silica-Gel Membran in einem speziellen Pufferystem mit hohen Salzkonzentra-
tionen. Virale RNA aus anhaftenden oder intrazellulär vorliegenden RSV-Partikeln wurde bei
dieser Methode mit extrahiert. Je 5 x 105 Zellen wurden in RNeasy-Lysepuffer, dem 1 % v/v

MATERIAL UND METHODEN 35
β-Mercaptoethanol als RNase-Inhibitor jeweils frisch zugesetzt wurde, lysiert und bis zur
weiteren Aufarbeitung bei -70° C gelagert. Die aufgetauten Proben wurden zunächst durch
Zentrifugation durch eine Siebmatrix (QiaShredder, Qiagen) homogenisiert und hoch-
molekulare DNA durch die entstehenden Scherkräfte fragmentiert. Dann wurden die Lysate
auf Zentrifugationssäulchen, die die Silica-Gel Membran enthielten, aufgetragen und die
RNA gemäß dem Protokoll des Herstellers in mehreren Wasch- und Zentrifugationsschritten
in einer Kühlzentrifuge (3K30, Sigma, Osterode a. H.) gereinigt. Eine fakultativ im Protokoll
vorgesehene zusätzliche DNase-Behandlung der Proben wurde nicht durchgeführt, da sich in
Vorversuchen gezeigt hatte, dass genomische DNA bei der späteren Polymerase-
Kettenreaktion nicht amplifiziert wurde (s. 2.2.9). Die Elution der gereinigten RNA von der
Säule erfolgte in je 30 µl RNase-freiem Wasser. Die RNA-Extrakte wurden bei -70° C
gelagert.
2.2.8 Reverse Transkription der RNA
Die Umschreibung von RNA in komplementäre DNA (cDNA), als Ausgangsmaterial für die
Polymerase-Kettenreaktion (s. 2.2.9), erfolgte unter Verwendung der Reagenzien eines
‚TaqMan® Reverse Transcription Kit’ (Applied Biosystems, Foster City, Kalifornien, USA).
Bei dieser Methode entstehen durch Anlagerung von Hexanukleotiden verschiedenster
Sequenzen, sogenannter ‚random primer’, an die RNA kurze doppelsträngige Abschnitte, die
als Startpunkt für die Syntheseaktivität der MultiScribe™ Reversen Transkriptase dienen,
einer RNA-abhängigen DNA-Polymerase aus dem Moloney Murine Leukaemia Virus. Durch
Verwendung von random Primern anstelle von sequenzspezifischen Primern konnte die
synthetisierte cDNA als Ausgangsmaterial für verschiedene PCR-Systeme eingesetzt werden.
Der Reaktionsmix enthielt 50 mM KCl, 10 mM Tris-HCl (pH 8,3), 5,5 mM MgCl2, 0,5 mM
dNTP, 2,5 µM random Primer, 10 U RNase Inhibitor, 30 U MultiScribe™ Reverse
Transkriptase und 5 µl RNA in einem Gesamtvolumen von 25 µl. Als Negativkontrolle wurde
jeweils ein Ansatz mit 5 µl sterilem H2O anstelle von RNA mitgeführt. Die Synthese-
Reaktionen wurde in ultradünnnen 0,2 ml Reaktionsgefäßen (Biozym, Hessisch Oldenburg) in
einem programmierbaren Thermocycler (PTC-200, MJ Research, Massachusetts, USA) mit
folgendem Temperaturprofil durchgeführt: 10 min. 25° C für die Anlagerung der Primer,
30 min. 48° C für die cDNA-Synthese und 5 min. 95° C zur Inaktivierung der Reversen
Transkriptase. Die cDNA-Proben wurden bei –20° C gelagert.

MATERIAL UND METHODEN 36
2.2.9 Etablierung einer quantitativen real-time PCR für humanes Interleukin-10
2.2.9.1 Prinzip der quantitativen real-time PCR
Die Polymerase-Kettenreaktion (PCR) dient dazu, DNA mittels einer thermostabilen DNA-
Polymerase und Desoxynukleotiden (dNTP) in vitro spezifisch zu amplifizieren (Mullis
1987). Bei einer konventionellen PCR wird die amplifizierte DNA erst am Ende der Reaktion
analysiert, meist durch eine Agarose-Gelelektrophorese. Rückschlüsse auf die eingesetzte
Menge an Ausgangsmaterial sind dann nicht mehr möglich (Jung 2000). Bei der quantitativen
real-time PCR hingegen wird die zunehmende Menge an amplifizierter DNA in jedem PCR-
Zyklus erfasst. Bei einer quantitativen real-time PCR nach der TaqMan®-Methode (Applied
Biosystems) geschieht dies durch die Verwendung sequenzspezifischer Oligonukleotide als
Sonden, die an die zu quantifizierende Ziel-DNA spezifisch binden und einen fluorochromen
Reporter-Farbstoff sowie ein ‚Quencher’-Molekül enthalten, das die vom Reporter-Farbstoff
emittierte Fluoreszenz kompensiert. Sobald die Ziel-DNA im Verlauf der PCR-Reaktion
amplifiziert wird, wird die Sonde durch die 5’-3’ Nuklease-Aktivität der Polymerase
abgebaut. Reporter- und Quencher-Molekül werden voneinander getrennt und die Fluoreszenz
des Reporter-Farbstoffes wird freigesetzt. Da an jedes Ziel-DNA-Molekül genau ein Sonden-
Molekül bindet, korreliert die freigesetzte Fluoreszenz in jedem PCR-Zyklus mit der Menge
an generierter Ziel-DNA. Die freigesetzte Fluoreszenz wird mittels einer Kamera in einem
entsprechend ausgerüsteten PCR-Gerät für jeden Reaktionsansatz in jedem Zyklus der PCR
gemessen und Software-gestützt ausgewertet (s. 2.2.9.4).
2.2.9.2 Festlegung der Primer- und Sonden-Sequenzen
Die Sequenzen der bei der PCR-Amplifikation als spezifische ‚forward’ und ‚reverse’ Primer
benötigten Oligonukleotide und die Sequenz der Sonde wurden mit dem Software-Programm
Primer Express™, Version 1.0 (Applied Biosystems) festgelegt. Auf Basis der veröffent-
lichten Sequenz für das humane IL-10 Gen (Vieira 1991, NCBI-Accession M57627;
Sanjanwala/de Waal-Malefyt Direct Submission 1994, unpublished, NCBI-Accession
U16720) wurde unter Berücksichtigung der Kriterien für die Verwendung in einer

MATERIAL UND METHODEN 37
quantitativen real-time PCR die für diese Ziel-DNA optimale Primer-/Sonden-Kombination
ausgewählt. Die wesentlichen Kriterien sind eine Schmelztemperatur der Primer zwischen
58° C und 60° C, eine Schmelztemperatur der Sonde zwischen 67° C und 70° C und eine
Länge des amplifizierten DNA-Abschnittes von 50 bis 150 Basenpaaren (bp). Zusätzlich
wurden die Primer-Sequenzen so ausgewählt, dass das Zielfragment Exon-Intron-Grenzen des
humanen IL-10 Gens überspannte. Dadurch wurde die Amplifikation von genomischer DNA,
die in Spuren als Verunreinigung in den RNA-Extrakten enthalten sein kann, unterbunden.
Die Sequenzen der für die hier etablierte IL-10 PCR als Primer und Sonde verwendeten
Oligonukleotide sind in Tab. 2.2 angegeben. Die Spezifität der Sequenzen wurde in einem
NCBI-Datenbank-Abgleich mittels BLAST-Search (Altschul 1990) überprüft. Das erwartete
PCR-Produkt hatte eine Länge von 86 bp.
Die Oligonukleotid-Synthese für die Primer erfolgte durch Applied Biosystems, OligoFactory
(Weiterstadt). Die Synthese der Sonde und die Markierung mit dem Reporter-Farbstoff FAM
(6-Carboxyfluorescein) am 5’-Ende und dem Quencher-Farbstoff TAMRA
(6-Carboxytetramethylrhodamin) am 3’-Ende wurden bei Eurogentec (Seraing, Belgien)
durchgeführt. Für die Amplifikation der ß-Glucuronidase (GUS) als Referenz-Gen für die
relative Quantifizierung (s. 2.2.9.4) wurde eine kommerziell erhältliche Primer-/Sonden-
Kombination (Applied Biosystems) verwendet.
Tab. 2.2: Sequenzen von Primern und Sonde für die quantitative real-time PCR für
humanes IL-10
Bezeichnung Sequenz 5’ → 3’ TM
IL-10 forward Primer 5’ GAG ATC TCC GAG ATG CCT TCA G 3’ 59° C
IL-10 reverse Primer 5’ CAA GGA CTC CTT TAA CAA CAA GTT GT 3’ 58° C
IL-10 Sonde 5’ AGA GTG AAG ACT TTC TTT CAA ATG AAG
GAT CAG C 3’ 67° C
TM = Schmelztemperatur

MATERIAL UND METHODEN 38
2.2.9.3 Reaktionsansätze der quantitativen real-time PCR für humanes Interleukin-10
Als Grundbestandteil der Ansätze für die quantitative real-time PCR wurde eine zweifach
konzentrierte Basislösung (TaqMan® Universal PCR Master Mix, Applied Biosystems)
verwendet, die AmpliTaq Gold® DNA Polymerase, AmpErase® Uracil-N-Glycosylase
(UNG), dATP, dCTP, dGTP und dUTP, einen passiven Referenzfarbstoff (ROX) und opti-
mierte Pufferkomponenten enthielt. AmpliTaq Gold® ist eine thermostabile DNA-Polymerase
aus Thermus aquaticus, die aufgrund einer Modifikation erst nach Erhitzung auf 95° C ihre
Enzym-Aktivität entfaltet. AmpErase® ist ein Schutz vor Reamplifikation kontaminierender
PCR-Produkte aus früheren Amplifikationen, da dUTP-enthaltende DNA-Abschnitte durch
das Enzym hydrolysiert werden, während RNA und dTTP-enthaltende DNA intakt bleiben
(Longo 1990).
Die PCR-Ansätze enthielten 12,5 µl TaqMan® Basislösung, je 300 nM IL-10 forward und
reverse Primer, 20 nM IL-10 Sonde und 5 µl cDNA in einem Gesamtvolumen von 25 µl. Als
Negativ-Kontrollen wurden die H2O-Kontrolle aus der cDNA-Synthese und in mindestens
zwei Ansätzen steriles H2O anstelle von cDNA verwendet. Positiv-Kontrollen enthielten statt
cDNA je 5 µl Plasmid-DNA verschiedener Konzentrationen (s. 2.2.9.8). Die Ansätze für die
Amplifikation der ß-Glucuronidase (GUS) als Referenz-Gen für die relative Quantifizierung
(s. 2.2.9.4) enthielten statt der IL-10 Primer und Sonde jeweils 1,25 µl GUS-spezifischen
Primer-/Sonden-Mix (Applied Biosystems).
Die Amplifikation erfolgte in speziellen, mit Deckelstreifen verschließbaren Reaktionsplatten
(MicroAmp® Optical 96-well Reaction Plate, Applied Biosystems) in einem GeneAmp® 5700
Sequence Detection System. Das PCR-Programm startete mit zwei einleitenden Temperatur-
Schritten: 2 min. 50° C zum Abbau möglicher PCR-Produkt Kontaminationen durch die
Uracil-N-Glycosylase (UNG) und 10 min. 95° C für die Aktivierung der AmpliTagGold®
Polymerase und die Inaktivierung der UNG. Die folgenden 40 PCR-Zyklen umfassten jeweils
die zwei Temperatur-Schritte: 15 sec. 95° C für die Denaturierung der gebildeten DNA-
Doppelstränge und 1 min. 60° C als kombinierten Schritt für die Primer-/Sonden-Anlagerung
und den Elongations-Schritt.

MATERIAL UND METHODEN 39
2.2.9.4 Relative Quantifizierung der IL-10 mRNA Expression
Die Auswertung der PCR-Ergebnisse erfolgte mittels GeneAmp SDS Software, Version 1.0
(Applied Biosystems). Die Rohdaten wurden zunächst gegen den in der TaqMan Basislösung
enthaltenen internen Referenzfarbstoff ROX abgeglichen. Diese normalisierten Reporter-
Signale (Rn) ergeben in einem Diagramm aufgetragen gegen die entsprechende Zykluszahl die
Amplifikationskurve einer Probe. Nachdem manuell ein Schwellenwert festgelegt wurde, bei
dem sich alle Amplifikationskurven in der logarithmischen Phase (= linearer Kurvenverlauf
bei logarithmischer Skalierung der y-Achse) befinden, errechnet das Programm für jede Probe
den sogenannten ‚Ct-Wert’ als den PCR-Zyklus, in dem die Amplifikationskurve dieser Probe
den Schwellenwert erreicht. Dabei korreliert der Ct-Wert mit der Ausgangsmenge an spezi-
fischer cDNA in dieser Probe. Je geringer die Ausgangsmenge an spezifischer cDNA in der
Probe ist, desto später erreicht die Amplifikationskurve den Schwellenwert, d. h. desto höher
ist der Ct-Wert dieser Probe. Bei 100%iger Effizienz des PCR-Systems nimmt der Ct-Wert
einer Probe bei der Halbierung der Ausgangsmenge um einen Zyklus zu. Von jeder Probe
wurde ein Ct-Wert für die IL-10 cDNA und in einem weiteren Ansatz im gleichen PCR-Lauf
ein Ct-Wert für die cDNA der β-Glucuronidase (GUS) als Referenz-Gen bestimmt. GUS
wurde als Referenz-Gen gewählt, da in Vorversuchen in unserer Arbeitsgruppe gezeigt wurde,
dass die Expression dieses Gens von 11 untersuchten Haushalts-Genen bei PBMC unter
verschiedensten Stimulationsbedingungen am konstantesten war (Löseke 2002). Die Diffe-
renz aus CtGUS – CtIL10 bildet den ∆Ct-Wert der Probe. Durch die Verwendung der ∆Ct-Werte
werden die Proben in Bezug auf die eingesetzte RNA-/cDNA-Menge normalisiert, so dass ein
quantitativer Vergleich der IL-10 Expression zwischen verschiedenen Proben möglich ist. Um
den Effekt verschiedener Stimulationen auf die Zellen direkt miteinander vergleichen zu
können, wurden bei einigen Auswertungen zusätzlich ∆∆Ct-Werte aus der Differenz zwischen
dem ∆Ct-Wert der stimulierten und dem ∆Ct-Wert der entsprechenden unstimulierten Probe
(Kalibrator) gebildet. Eine Zunahme der IL-10 Expression durch die Stimulation wird
dadurch relativ zur IL-10 Expression bei Kultivierung ohne spezifischen Stimulus berechnet.
Die Menge an IL-10 Transkript in der Kalibrator-Probe wird gleich 1 gesetzt und die Menge
in allen anderen Proben als x-fach Vielfaches relativ zur Kalibrator-Probe ausgedrückt. Für
PCR-Systeme, die unter Beachtung der Kriterien für die TaqMan®-PCR entwickelt und
optimiert wurden (s. 2.2.9.2) liegt die Effizienz der Reaktion nahe 100% (GeneAmp SDS
Benutzer-Handbuch, S. A-11). Dies vorausgesetzt, errechnet sich die IL-10 mRNA Menge

MATERIAL UND METHODEN 40
einer Probe, normalisiert auf GUS als interne Referenz und relativ zur entsprechenden
Kalibrator-Probe als Potenz von 2 mit dem ∆∆Ct-Wert als Exponenten (= 2∆∆Ct).
2.2.9.5 Analyse der PCR-Produkte
Zur Kontrolle der Spezifität des PCR-Systems wurden die PCR-Produkte in der Etablierungs-
phase mittels Agarose-Gelelektrophorese analysiert. Je 20 µl PCR-Produkt wurden mit 4 µl
Ladepuffer versetzt und in horizontalen Agarose-Gelen (3 % w/v Agarose in TBE-Puffer) bei
75 V 1,5 h elektrophoretisch aufgetrennt. Die Agarose-Gele enthielten 0,7 µg/ml Ethidi-
umbromid, so dass die DNA-Banden unter UV-Licht sichtbar wurden und mit einer Sofort-
bildkamera (DS34, Polaroid, Offenbach) fotografiert werden konnten. Die Agarose-Gel-
elektrophorese gab Aufschluss über die Größe des entstehenden PCR-Produktes und eventuell
vorhandene Nebenprodukte.
Eine eventuelle Entstehung von Primer-Dimeren konnte durch Verwendung von SybrGreen in
den PCR-Reaktionsansätzen anstelle der fluorogenen Sonde überprüft werden. SybrGreen ist
ein fluoreszierender Farbstoff, der in doppelsträngiger DNA sequenzunabhängig interkaliert
und vom GeneAmp® 5700-System ebenfalls detektiert werden kann. Am Ende eines PCR-
Laufs mit SybrGreen wird zusätzlich zu den Amplifikationskurven ein Dissoziationsprotokoll
erstellt, aus dem die Schmelztemperaturen der entstandenen PCR-Produkte abgelesen werden
können. Die Schmelzkurven ergeben sich aus der ersten Ableitung (dF/dT) der Funktion
Fluoreszenzintensität (F) in Abhängigkeit von der Temperatur (T). Entstehen neben dem Ziel-
fragment auch Primer-Dimere, so weisen die Schmelzkurven Nebenmaxima bei niedrigeren
Schmelztemperaturen auf. Die mit der Primer-Express-Software (s. 2.2.9.2) theoretisch
errechnete Schmelztemperatur für das mit diesem PCR-System amplifizierte IL-10 Fragment
lag bei 78,0° C.
Zur Überprüfung der Sequenzspezifität wurden die PCR-Produkte zusätzlich sequenziert
(s. 2.2.9.7).
2.2.9.6 Klonierung der PCR-Produkte
Zur Erstellung eines Standards für die IL-10 PCR und für eine Sequenzierung der PCR-
Produkte wurden größere Mengen der spezifischen Ziel-DNA benötigt, die durch Klonierung

MATERIAL UND METHODEN 41
des PCR-Produktes gewonnen wurden. Dazu mussten die PCR-Produkte zunächst aufge-
reinigt werden. Die Elution aus den PCR-Ansätzen erfolgte unter Verwendung kommerzieller
Fertig-Reagenzien (MinElute Kit, Qiagen) nach dem Protokoll des Herstellers. In den
Reaktionsansätzen enthaltene Proteine, Salze, Primer und dNTPs wurden entfernt und die
gereinigte Ziel-DNA in 10 µl H2O resuspendiert.
Als Klonierungssystem wurden der Vektor pCR®4-TOPO® und Escherichia coli K12, Stamm
TOP10 als Zielzellen eingesetzt und die Klonierung nach dem Protokoll des Herstellers
durchgeführt (TOPO TA Cloning® Kit for Sequencing, Invitrogen, Groningen, Niederlande).
pCR®4-TOPO® ist ein linearisierter Vektor mit überhängenden Thymidinresten an den
3’-Enden und kovalent gebundenem Enzym Topoisomerase I. Aufgrund einer terminalen
Transferase-Aktivität der Taq Polymerase enthalten die PCR-Produkte jeweils einen einzel-
nen überhängenden Adenosinrest am 3’-Ende beider Stränge. Die Bindung der Topoiso-
merase I an den Vektor kann durch Reaktion mit einer 5’-Hydroxylgruppe gelöst werden
(Shuman 1994). Bei der Ligationsreaktion (15 min. bei Raumtemperatur) geschieht dies durch
die 5’-Hydroxylgruppen der PCR-Produkte, so dass das PCR-Produkt durch die Bindung
zwischen seinen überhängenden Adenosinresten und den überhängenden Thymidinresten des
Vektors in die Vektorsequenz eingebunden wird.
Chemisch kompetente E. coli TOP10 Zellen wurden mit dem Ligationsansatz durch Inkuba-
tion auf Eis und Hitzeschock-Behandlung (30 sec, 42° C) transformiert. Die anschließende
Inkubation in SOC-Medium (2 % Trypton, 0,5 % Hefeextrakt, 10 mM NaCl, 2,5 mM MgCl2,
10 mM MgSO4, 20 mM Glukose) für 60 min. bei 37° C diente der Expression des auf dem
Vektor kodierten Kanamycin-Resistenz-Gens in den Zellen, die das Plasmid in sich trugen.
Die Zellsuspension wurde auf LB-Agarplatten, die 50 µg/ml Kanamycin enthielten,
ausplattiert und über Nacht bei 37° C bebrütet. Das Wachstum von Zellen, die einen Vektor
ohne Insert trugen, wurde dadurch vermieden, dass leere Vektoren das für die E. coli Zelle
letale ccdB-Gen kodieren (Bernard 1994), dessen Sequenz bei Insertion eines PCR-Produktes
zerstört wird.
Positive Klone wurden in 7 ml Flüssigkultur (LB-Medium mit 50 µg/ml Kanamycin) über
Nacht im Schüttler bei 37° C vermehrt und danach die Plasmid-DNA aufgereinigt (Quantum
Prep Plasmid Miniprep Kit, Bio-Rad Laboratories, München). Je 1,5 ml der Übernachtkultur
wurden zentrifugiert (13000 x g, 30 sec, Raumtemperatur) und der Überstand verworfen.
Dieser Vorgang wurde zwei- bis dreimal im selben Reaktionsgefäß wiederholt, bis ein Zell-
sediment deutlich sichtbar war. Dann wurden die Zellen in mehreren Schritten gemäß der
Angaben des Herstellers durch Zusatz vorgefertigter Puffer-Lösungen und Zentrifugation

MATERIAL UND METHODEN 42
lysiert, die Plasmid-DNA an eine Filtermembran gebunden, gewaschen und in 100 µl H2O
eluiert. Der DNA-Gehalt des Eluats wurde aus der Messung der Absorption der Lösung bei
260 nm (A260) in einem UV-Spektrophotometer (DU®-70, Beckman Instruments, München)
wie folgt berechnet (Sambrook 2001, Seite A8.21):
A260 x 0,05 x Verdünnung = DNA-Gehalt in µg/µl.
2.2.9.7 Sequenzierung der PCR-Produkte
Die Sequenzierung der Plasmid-DNA/PCR-Produkte erfolgte nach der Didesoxymethode
(Kettenabbruchmethode) von Sanger (1977). Durch den Einbau von Didesoxynukleotiden
(ddNTPs) kommt es bei der Amplifikation zu Strangabbrüchen, so dass Fragmente aller mög-
lichen Längen entstehen. Die vier verwendeten ddNTPs (ddATP, ddCTP, ddGTP, ddTTP)
sind mit unterschiedlichen Fluoreszenzfarbstoffen markiert, die in einer anschließenden
Gelelektrophorese durch einen 488 nm Argon-Laser angeregt werden. Da die einzelnen
Fragmente in der Reihenfolge ihrer Länge den Laserstrahl passieren, kann aus den Signalen
der farbmarkierten Terminatoren die Basensequenz abgeleitet werden.
Je 500 ng Plasmid-DNA wurden nach den Herstellerangaben des verwendeten Sequenzier-Kit
(Big Dye Prism Terminator Kit, Applied Biosystems) in einer zyklischen Sequenzierreaktion
linear amplifiziert. Als Primer wurde jeweils einer der beiden M13-Primer verwendet, für die
Bindungsstellen in den pCR®4-TOPO® Vektor 89 bp vor bzw. 60 bp hinter der Klonie-
rungsstelle für das PCR-Produkt eingebaut sind. Die Primer-Sequenzen und das Temperatur-
programm der Sequenzierreaktionen, die in einem PTC-200 Thermocycler (MJ Research,
Massachusetts, USA) durchgeführt wurde, sind in Tabelle 2.3 angegeben. Die Produkte der
Sequenzierreaktion wurden anschließend gemäß der Angaben im Herstellerprotokoll durch
eine Ethanol/Natriumacetat-Fällung der DNA für die folgende Gelelektrophorese aufge-
reinigt.
Die gelelektrophoretische Analyse und Auswertung der Sequenzier-Proben erfolgte auf einem
ABI PRISM 310 Sequenzierautomaten (Applied Biosystems) in der Abteilung Molekulare
Medizin, Berufsgenossenschaftliches Forschungsinstitut für Arbeitsmedizin, Institut an der
Ruhr-Universität Bochum.

MATERIAL UND METHODEN 43
Tab. 2.3a: Primer-Sequenzen der Sequenzierreaktion
Bezeichnung Sequenz 5’ → 3’
M13 forward Primer 5’ GTA AAA CGA CGG CCA G 3’
M13 reverse Primer 5’ CAG GAA ACA GCT ATG AC 3’
Quelle: Benutzerhandbuch ‘TOPO TA Clonino® Kit for Sequencing’, Invitrogen
Tab. 2.3b: Temperaturprogramm der Sequenzierreaktion Anzahl Zyklen Funktion Temperatur Zeit
1 erstes Denaturieren 96° C 5 min
Denaturieren 96° C 10 sec
Primer-Anlagerung 55° C 5 sec 25
Kettenverlängerung 60° C 4 min
2.2.9.8 Absolute Quantifizierung der IL-10 Kopienzahl
Neben der in 2.2.9.4 beschriebenen relativen Quantifizierung der IL-10 mRNA Expression,
war durch Verwendung von Plasmid-DNA als Standard auch eine absolute Quantifizierung
der in den PCR-Ansätzen enthaltenen IL-10 Kopienzahl möglich. Aus Plasmid-DNA mit
einer Ausgangskonzentration von 200 pg/µl wurden durch dekadische Verdünnungen DNA-
Standardlösungen mit Konzentrationen von 20 pg/µl bis 2 x 10-6 pg/µl erstellt und portioniert
bei –20° C gelagert. Bei jedem PCR-Lauf wurden Ansätze mit je 5 µl der verschiedenen
Standardlösungen anstelle von 5 µl cDNA in Doppelbestimmung mitgeführt. Bei der Aus-
wertung wurde aus diesen Ansätzen Software-gestützt eine Standardkurve erstellt und der
IL-10 Gehalt der Proben durch Interkalieren berechnet. Aus dem molekularen Gewicht des
Plasmids einschließlich Insert ergab sich, dass 100 pg Plasmid-DNA 0,04 fmol IL-10 cDNA
entsprachen, also 24,2 x 106 Kopien. Bei einem Volumen der PCR-Reaktion von 25 µl
entsprach ein PCR-Ergebnis von 1 pg/µl also 6,05 x 106 Kopien in diesem PCR-Ansatz. Unter
Berücksichtigung der für die cDNA-Synthese und die PCR eingesetzten Anteile an RNA bzw.

MATERIAL UND METHODEN 44
cDNA errechnete sich die Kopienzahl/Zelle für einen Kulturansatz damit aus folgender
Gleichung:
Betrag des PCR-Ergebnisses in pg/µl x 6,05 x 106 x 30 Gesamtzellzahl im Kulturansatz
Dieser Umrechnung liegt die Annahme zugrunde, dass RNA-Isolation und cDNA-Synthese
ohne Verluste an IL-10 Transkripten erfolgten.
2.2.10 RSV-Nachweis mittels quantitativer real-time PCR
Analog des IL-10 Nachweises erfolgte in RSV-stimulierten Kulturansätzen auch ein RSV-
Nachweis mittels quantitativer real-time PCR. Dazu wurde ein in unserer Arbeitsgruppe
etabliertes PCR-System benutzt, mit dem ein 76 bp großes Fragment der Sequenz für das
RSV-Fusionsprotein (RSV-F) nachgewiesen wird (Borg 2002). Als Ausgangsmaterial für die
PCR wurden wiederum je 5 µl der cDNA, die mit random Primern synthetisiert worden war
(s. 2.2.8), eingesetzt. Die PCR wurde wie in Abschnitt 2.2.9 beschrieben durchgeführt. Die
Auswertung erfolgte anhand der ermittelten Ct-Werte der Proben. Durch die Erstellung von
Standardkurven aus den Ct-Werten von Plasmid-DNA, die das RSV-F PCR-Produkt enthielt,
war auch eine absolute Quantifizierung möglich. Die Standardkurve für die RSV-F PCR
reichte von 2,4 x 106 bis 24 Kopien/25 µl PCR-Ansatz.
2.2.11 Statistische Auswertungen
Für alle statistischen Auswertungen wurde SPSS 9.0 (SPSS Inc., Chicago, Illinois, USA) als
Softwareprogramm verwendet. Unterschiede zwischen verschiedenen Stimulationsbedin-
gungen sowie zwischen verschiedenen Zelltypen wurden mit dem Wilcoxon-Test für gepaarte
Stichproben untersucht. Für Vergleiche zwischen verschiedenen Spendergruppen wurde der
Mann-Whitney-U-Test verwendet. Korrelationen wurden nach der Spearman-Rangkorrelation
berechnet. Alle Tests wurden zweiseitig durchgeführt. Irrtumswahrscheinlichkeiten von unter
5 % (p < 0,05) wurden als statistisch signifikant angesehen.

45
3. Ergebnisse
3.1 Ergebnisse der einstufigen und zweistufigen magnetischen Zellsortie-
rung
3.1.1 Reinheit der Zellfraktionen nach magnetischer Sortierung
Für die in dieser Arbeit beschriebenen Experimente wurden T-Zellen und Monozyten aus dem
peripheren Blut erwachsener Spender mittels Gradientenzentrifugation und magnetischer
Zellsortierung isoliert. Die Kontrolle der Reinheit der so gewonnenen Zellfraktionen erfolgte
durch phänotypische Untersuchungen mit zelltyp-spezifischen Antikörpern im Durchfluss-
zytometer (s. 2.2.5).
Für die einstufige CD3+ Sortierung ergab sich eine Reinheit von 94,18 ± 3,08 % (N = 8) in
Bezug auf CD3+ T-Zellen in der positiven Fraktion. Bei den übrigen Zellen handelte es sich
überwiegend um CD14+ Monozyten sowie vereinzelt um B-Zellen und NK-Zellen. Für Unter-
suchungen, bei denen die angewandte Methode keine Differenzierung zwischen verschie-
denen Zelltypen erlaubt, wie PCR und ELISA, musste jedoch ausgeschlossen werden, dass
andere Zellen als T-Zellen als Quelle der gemessenen Zytokine in Frage kommen. Daher
wurde die Sortierung für die funktionellen Analysen um eine CD14-Depletion erweitert.
Die CD3+14- sortierten Zellfraktionen enthielten zu 97,65 ± 1,56 % (N = 14) CD3+ T-Zellen.
Der Anteil verunreinigender CD14+ Monozyten konnte dabei von durchschnittlich
3,08 ± 1,95 % in einstufig sortierten CD3+ T-Zellen auf 0,22 ± 0,23 % gesenkt werden.
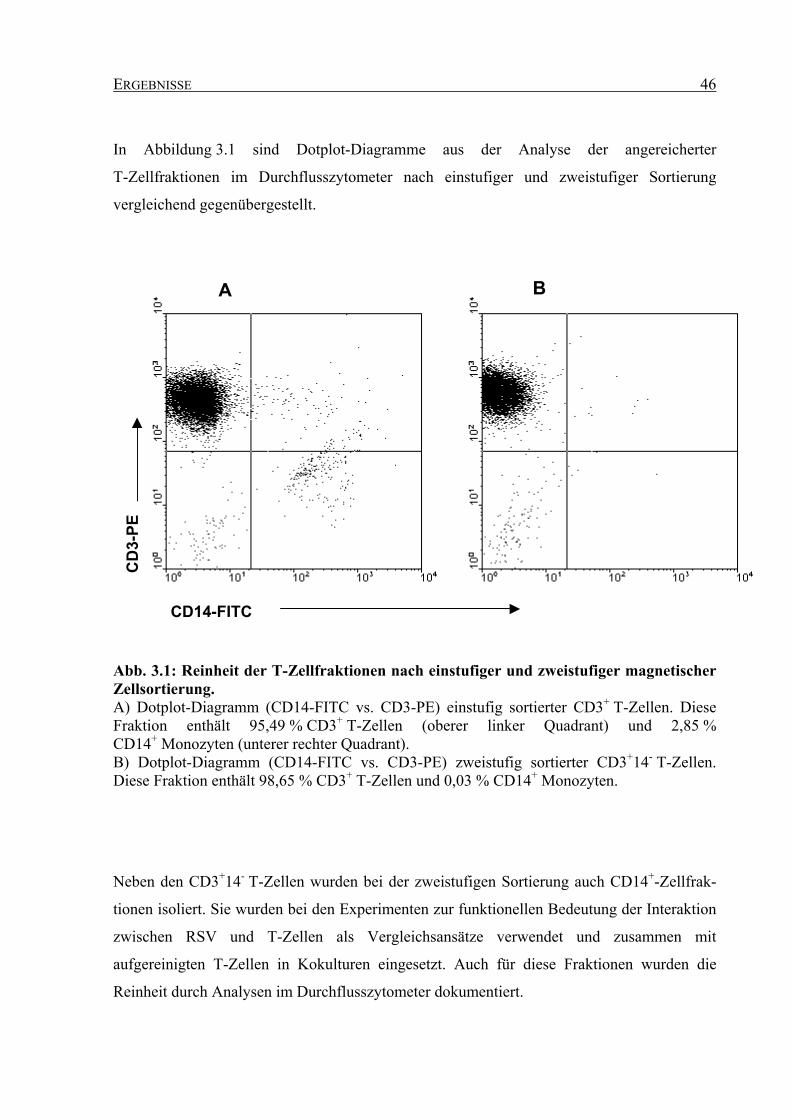
ERGEBNISSE 46
In Abbildung 3.1 sind Dotplot-Diagramme aus der Analyse der angereicherter
T-Zellfraktionen im Durchflusszytometer nach einstufiger und zweistufiger Sortierung
vergleichend gegenübergestellt.
CD14-FITC
CD
3-PE
BA
Abb. 3.1: Reinheit der T-Zellfraktionen nach einstufiger und zweistufiger magnetischer Zellsortierung. A) Dotplot-Diagramm (CD14-FITC vs. CD3-PE) einstufig sortierter CD3+ T-Zellen. Diese Fraktion enthält 95,49 % CD3+ T-Zellen (oberer linker Quadrant) und 2,85 % CD14+ Monozyten (unterer rechter Quadrant). B) Dotplot-Diagramm (CD14-FITC vs. CD3-PE) zweistufig sortierter CD3+14- T-Zellen. Diese Fraktion enthält 98,65 % CD3+ T-Zellen und 0,03 % CD14+ Monozyten.
Neben den CD3+14- T-Zellen wurden bei der zweistufigen Sortierung auch CD14+-Zellfrak-
tionen isoliert. Sie wurden bei den Experimenten zur funktionellen Bedeutung der Interaktion
zwischen RSV und T-Zellen als Vergleichsansätze verwendet und zusammen mit
aufgereinigten T-Zellen in Kokulturen eingesetzt. Auch für diese Fraktionen wurden die
Reinheit durch Analysen im Durchflusszytometer dokumentiert.
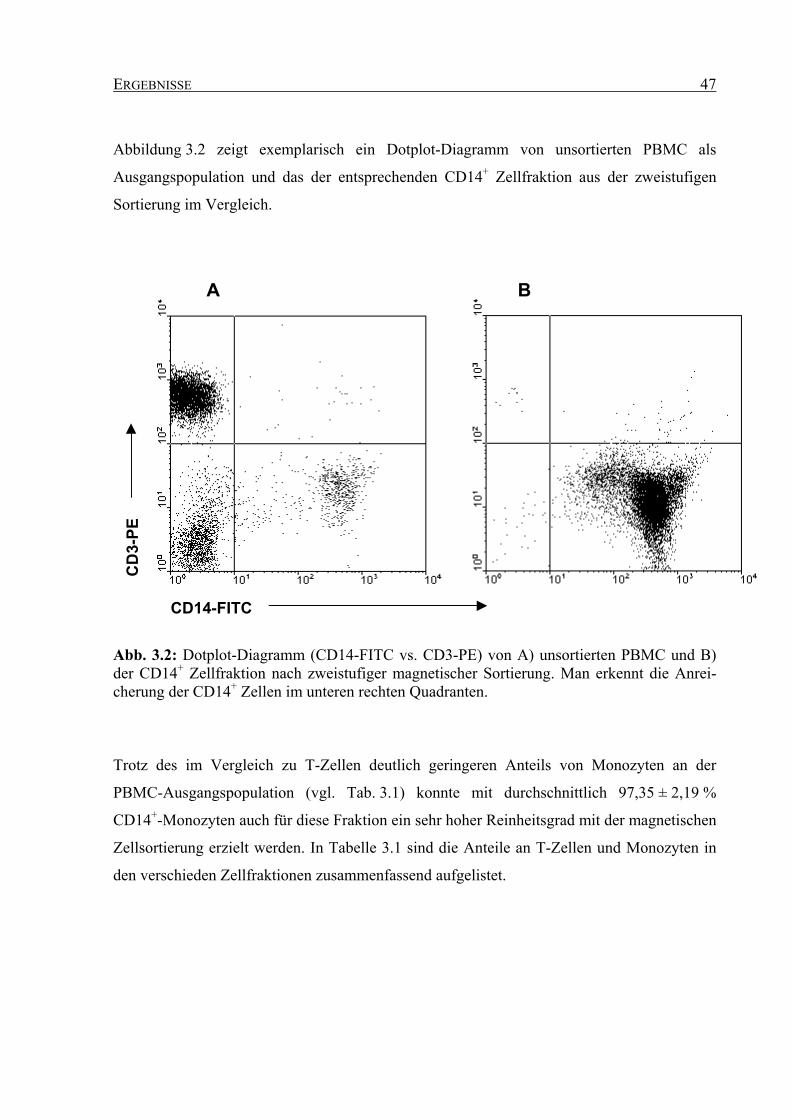
ERGEBNISSE 47
Abbildung 3.2 zeigt exemplarisch ein Dotplot-Diagramm von unsortierten PBMC als
Ausgangspopulation und das der entsprechenden CD14+ Zellfraktion aus der zweistufigen
Sortierung im Vergleich.
BA
CD14-FITC
CD
3-PE
Abb. 3.2: Dotplot-Diagramm (CD14-FITC vs. CD3-PE) von A) unsortierten PBMC und B) der CD14+ Zellfraktion nach zweistufiger magnetischer Sortierung. Man erkennt die Anrei-cherung der CD14+ Zellen im unteren rechten Quadranten.
Trotz des im Vergleich zu T-Zellen deutlich geringeren Anteils von Monozyten an der
PBMC-Ausgangspopulation (vgl. Tab. 3.1) konnte mit durchschnittlich 97,35 ± 2,19 %
CD14+-Monozyten auch für diese Fraktion ein sehr hoher Reinheitsgrad mit der magnetischen
Zellsortierung erzielt werden. In Tabelle 3.1 sind die Anteile an T-Zellen und Monozyten in
den verschieden Zellfraktionen zusammenfassend aufgelistet.

ERGEBNISSE 48
Tab. 3.1: Anteile an CD3+ T-Zellen und CD14+ Monozyten in den Fraktionen der
magnetischen Zellsortierung
Anteil CD3+ [%] Anteil CD14+ [%]
PBMC 69,28 ± 10,21 7,93 ± 4,43
CD3+ Fraktion aus einstufiger Sortierung 94,18 ± 3,08 3,07 ± 1,95
CD3+14- Fraktion aus zweistufiger Sortierung 97,65 ± 1,56 0,22 ± 0,23
CD14+ Fraktion aus zweistufiger Sortierung 0,34 ± 0,52 97,35 ± 2,19
3.1.2 Zellausbeute bei der magnetischen Zellsortierung
Bei jeder Zellisolierung wurden 100 ml heparinisiertes Vollblut pro Spender eingesetzt und
daraus 1 - 2 x 106 PBMC/ml Vollblut gewonnen. Durchschnittlich standen 126 x 106 PBMC
für die anschließende magnetische Zellsortierung zur Verfügung. Die Zellausbeute lag bei der
einstufigen CD3+-Sortierung bei 39,5 ± 12,2 % der eingesetzten PBMC (N = 81). Bei der
zweistufigen Sortierung (N = 14) betrug sie für die CD3+14- Fraktion 21,8 ± 8,5 % der einge-
setzten PBMC und 32,2 ± 11,5 % bezogen auf die CD14-depletierte Fraktion. Die
CD14+-Fraktion umfasste 14,6 ± 6,0 %. Somit standen im allgemeinen 28 x 106 T-Zellen und
19 x 106 CD14+ Monozyten für die nachfolgenden Experimente zur Verfügung.
In Verbindung mit dem im vorangehenden Abschnitt dokumentierten hohen Anrei-
cherungsgrad zeigt dies, dass die in dieser Arbeit beschriebene magnetische Zellsortierung
eine effektive Methode zur Gewinnung sehr reiner CD3+- und CD14+-Zellpopulationen aus
PBMC darstellt.
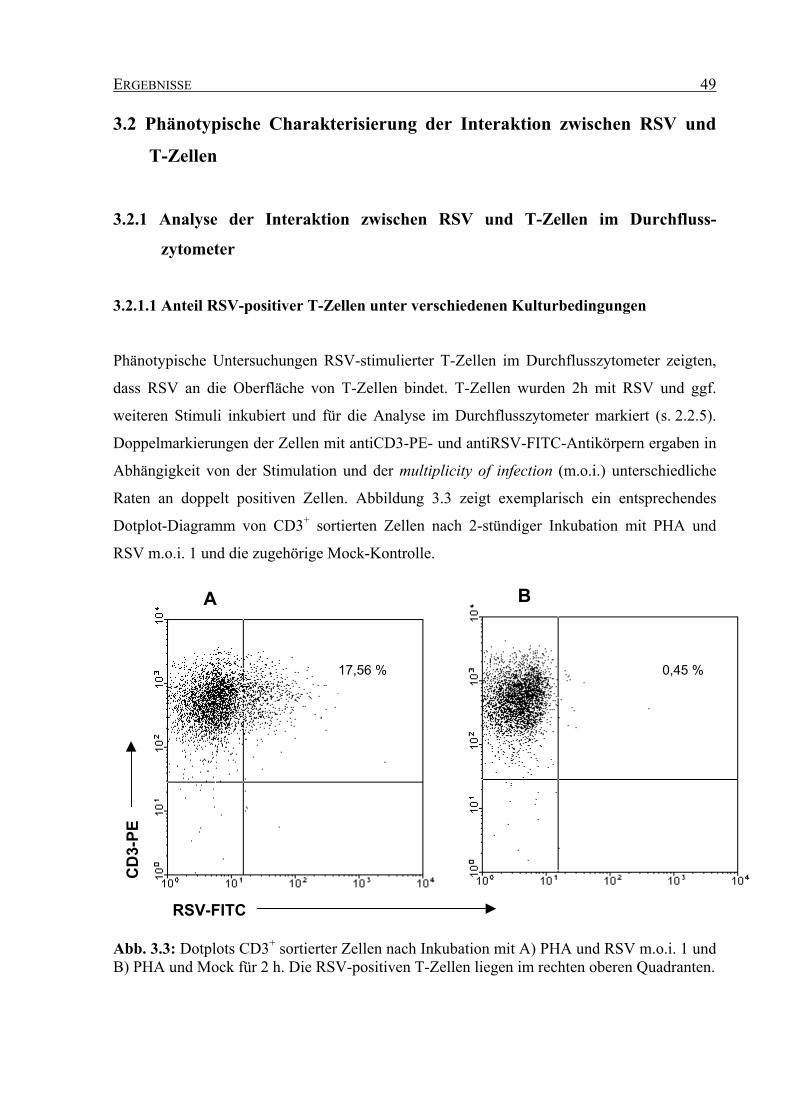
ERGEBNISSE 49
3.2 Phänotypische Charakterisierung der Interaktion zwischen RSV und
T-Zellen
3.2.1 Analyse der Interaktion zwischen RSV und T-Zellen im Durchfluss-
zytometer
3.2.1.1 Anteil RSV-positiver T-Zellen unter verschiedenen Kulturbedingungen
Phänotypische Untersuchungen RSV-stimulierter T-Zellen im Durchflusszytometer zeigten,
dass RSV an die Oberfläche von T-Zellen bindet. T-Zellen wurden 2h mit RSV und ggf.
weiteren Stimuli inkubiert und für die Analyse im Durchflusszytometer markiert (s. 2.2.5).
Doppelmarkierungen der Zellen mit antiCD3-PE- und antiRSV-FITC-Antikörpern ergaben in
Abhängigkeit von der Stimulation und der multiplicity of infection (m.o.i.) unterschiedliche
Raten an doppelt positiven Zellen. Abbildung 3.3 zeigt exemplarisch ein entsprechendes
Dotplot-Diagramm von CD3+ sortierten Zellen nach 2-stündiger Inkubation mit PHA und
RSV m.o.i. 1 und die zugehörige Mock-Kontrolle.
0,45 % 17,56 %
RSV-FITC
BA
CD
3-PE
Abb. 3.3: Dotplots CD3+ sortierter Zellen nach Inkubation mit A) PHA und RSV m.o.i. 1 und B) PHA und Mock für 2 h. Die RSV-positiven T-Zellen liegen im rechten oberen Quadranten.
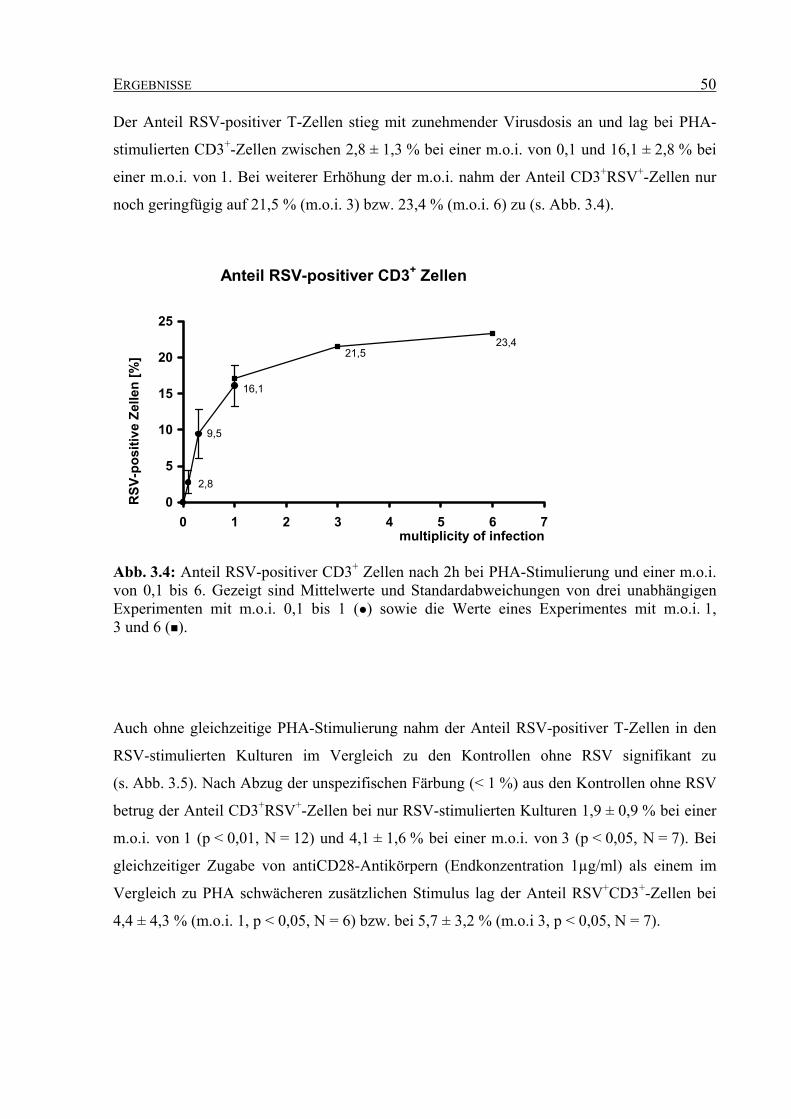
ERGEBNISSE 50
Der Anteil RSV-positiver T-Zellen stieg mit zunehmender Virusdosis an und lag bei PHA-
stimulierten CD3+-Zellen zwischen 2,8 ± 1,3 % bei einer m.o.i. von 0,1 und 16,1 ± 2,8 % bei
einer m.o.i. von 1. Bei weiterer Erhöhung der m.o.i. nahm der Anteil CD3+RSV+-Zellen nur
noch geringfügig auf 21,5 % (m.o.i. 3) bzw. 23,4 % (m.o.i. 6) zu (s. Abb. 3.4).
Anteil RSV-positiver CD3+ Zellen
16,1
2,8
9,5
23,421,5
0
5
10
15
20
25
0 1 2 3 4 5 6 7multiplicity of infection
RSV
-pos
itive
Zel
len
[%]
Abb. 3.4: Anteil RSV-positiver CD3+ Zellen nach 2h bei PHA-Stimulierung und einer m.o.i. von 0,1 bis 6. Gezeigt sind Mittelwerte und Standardabweichungen von drei unabhängigen Experimenten mit m.o.i. 0,1 bis 1 ( ) sowie die Werte eines Experimentes mit m.o.i. 1, 3 und 6 ( ).
Auch ohne gleichzeitige PHA-Stimulierung nahm der Anteil RSV-positiver T-Zellen in den
RSV-stimulierten Kulturen im Vergleich zu den Kontrollen ohne RSV signifikant zu
(s. Abb. 3.5). Nach Abzug der unspezifischen Färbung (< 1 %) aus den Kontrollen ohne RSV
betrug der Anteil CD3+RSV+-Zellen bei nur RSV-stimulierten Kulturen 1,9 ± 0,9 % bei einer
m.o.i. von 1 (p < 0,01, N = 12) und 4,1 ± 1,6 % bei einer m.o.i. von 3 (p < 0,05, N = 7). Bei
gleichzeitiger Zugabe von antiCD28-Antikörpern (Endkonzentration 1µg/ml) als einem im
Vergleich zu PHA schwächeren zusätzlichen Stimulus lag der Anteil RSV+CD3+-Zellen bei
4,4 ± 4,3 % (m.o.i. 1, p < 0,05, N = 6) bzw. bei 5,7 ± 3,2 % (m.o.i 3, p < 0,05, N = 7).
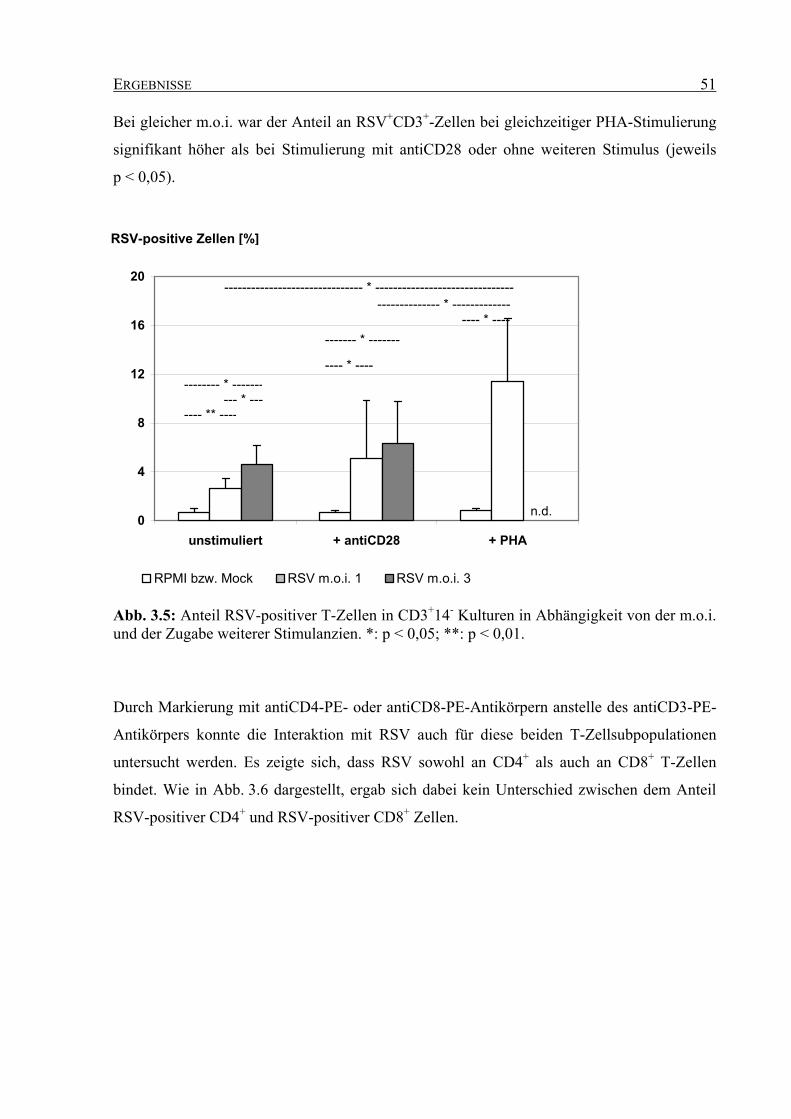
ERGEBNISSE 51
Bei gleicher m.o.i. war der Anteil an RSV+CD3+-Zellen bei gleichzeitiger PHA-Stimulierung
signifikant höher als bei Stimulierung mit antiCD28 oder ohne weiteren Stimulus (jeweils
p < 0,05).
]
RSV-positive Zellen [%0
4
8
12
16
20
unstimuliert + antiCD28 + PHA
RPMI bzw. Mock
---- ** ----
-------- * -------
------- * -------
---- * ----
---- * ----
------------------------------- * --------------------------------------------- * -------------
--- * ---
n.d.
RSV m.o.i. 1 RSV m.o.i. 3
Abb. 3.5: Anteil RSV-positiver T-Zellen in CD3+14- Kulturen in Abhängigkeit von der m.o.i. und der Zugabe weiterer Stimulanzien. *: p < 0,05; **: p < 0,01.
Durch Markierung mit antiCD4-PE- oder antiCD8-PE-Antikörpern anstelle des antiCD3-PE-
Antikörpers konnte die Interaktion mit RSV auch für diese beiden T-Zellsubpopulationen
untersucht werden. Es zeigte sich, dass RSV sowohl an CD4+ als auch an CD8+ T-Zellen
bindet. Wie in Abb. 3.6 dargestellt, ergab sich dabei kein Unterschied zwischen dem Anteil
RSV-positiver CD4+ und RSV-positiver CD8+ Zellen.
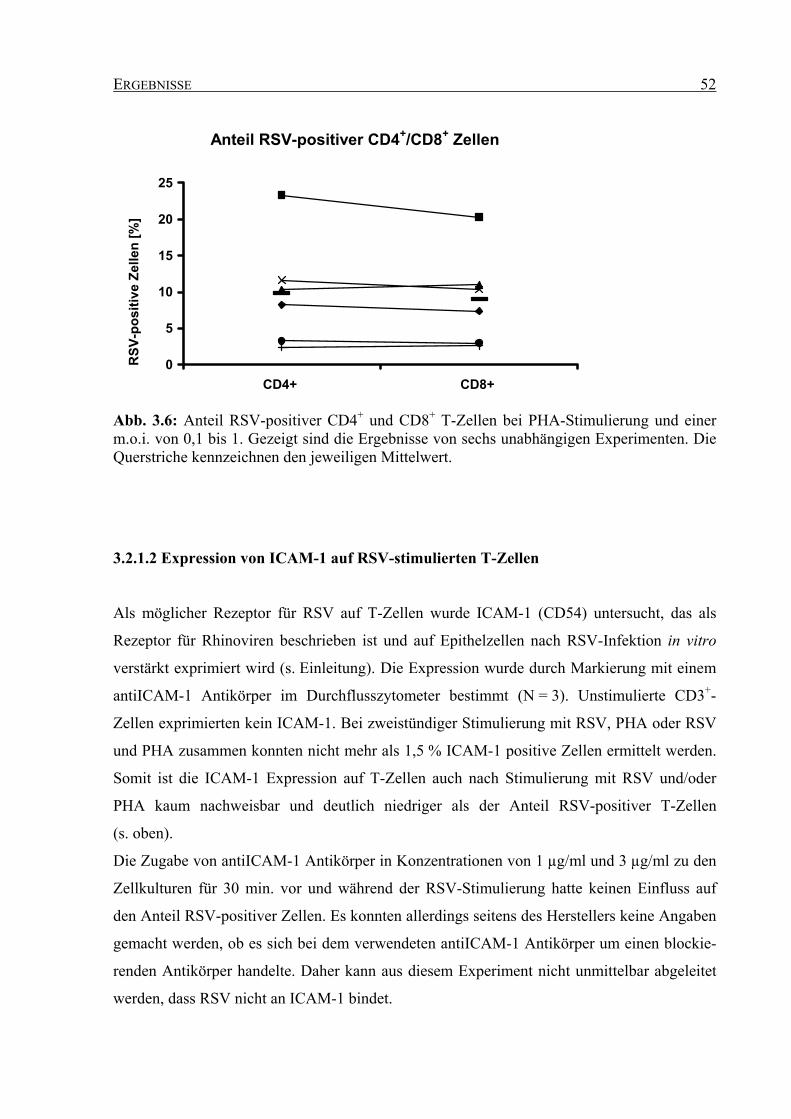
ERGEBNISSE 52
Anteil RSV-positiver CD4+/CD8+ Zellen
0
5
10
15
20
25
CD4+ CD8+
RSV
-pos
itive
Zel
len
[%]
Abb. 3.6: Anteil RSV-positiver CD4+ und CD8+ T-Zellen bei PHA-Stimulierung und einer m.o.i. von 0,1 bis 1. Gezeigt sind die Ergebnisse von sechs unabhängigen Experimenten. Die Querstriche kennzeichnen den jeweiligen Mittelwert.
3.2.1.2 Expression von ICAM-1 auf RSV-stimulierten T-Zellen
Als möglicher Rezeptor für RSV auf T-Zellen wurde ICAM-1 (CD54) untersucht, das als
Rezeptor für Rhinoviren beschrieben ist und auf Epithelzellen nach RSV-Infektion in vitro
verstärkt exprimiert wird (s. Einleitung). Die Expression wurde durch Markierung mit einem
antiICAM-1 Antikörper im Durchflusszytometer bestimmt (N = 3). Unstimulierte CD3+-
Zellen exprimierten kein ICAM-1. Bei zweistündiger Stimulierung mit RSV, PHA oder RSV
und PHA zusammen konnten nicht mehr als 1,5 % ICAM-1 positive Zellen ermittelt werden.
Somit ist die ICAM-1 Expression auf T-Zellen auch nach Stimulierung mit RSV und/oder
PHA kaum nachweisbar und deutlich niedriger als der Anteil RSV-positiver T-Zellen
(s. oben).
Die Zugabe von antiICAM-1 Antikörper in Konzentrationen von 1 µg/ml und 3 µg/ml zu den
Zellkulturen für 30 min. vor und während der RSV-Stimulierung hatte keinen Einfluss auf
den Anteil RSV-positiver Zellen. Es konnten allerdings seitens des Herstellers keine Angaben
gemacht werden, ob es sich bei dem verwendeten antiICAM-1 Antikörper um einen blockie-
renden Antikörper handelte. Daher kann aus diesem Experiment nicht unmittelbar abgeleitet
werden, dass RSV nicht an ICAM-1 bindet.

ERGEBNISSE 53
Diese Ergebnisse, insbesondere die niedrige Expressionsrate von ICAM-1 auf T-Zellen auch
nach Stimulierung, deuten an, dass andere Rezeptoren als ICAM-1 an der Bindung von RSV
an die T-Zellen beteiligt sind.
3.2.1.3 Expression von CD69 auf RSV-stimulierten T-Zellen
Im Rahmen der phänotypischen Untersuchungen RSV-stimulierter T-Zellen wurde auch die
Expression von CD69, einem Marker für die Aktivierung von T-Zellen (Hara 1986),
untersucht (N = 5). Frisch isolierte CD3+14- T-Zellen exprimierten kein CD69. Nach 24-
stündiger Kulturdauer war eine geringe Expression von 1,6 ± 0,8 % auf den T-Zellen
nachweisbar. Durch Zugabe von antiCD28-Antikörpern (Endkonzentration 1µg/ml) nahm die
Expression auf 4,4 ± 2,6 % CD69+ Zellen nach 24h zu. Stimulierung mit RSV hatte keinen
Einfluss auf die CD69-Expression (m.o.i. 1: 1,6 ± 1,0 %; m.o.i. 3: 2,2 ± 1,2 %). RSV-
Stimulierung in Verbindung mit gleichzeitiger antiCD28-Zugabe führte zu keiner höheren
CD69-Expression als antiCD28-Zugabe allein (m.o.i. 1 + antiCD28: 3,6 ± 2,8 %; m.o.i. 3
+ antiCD28: 4,0 ± 2,5 %).
3.2.2 Analyse der Interaktion zwischen RSV und T-Zellen im konfokalen Laser-
Rastermikroskop
In Zusammenarbeit mit dem Institut für Pathologie der Ruhr-Universität Bochum wurde eine
Immunfluoreszenzmarkierung für RSV-stimulierte T-Zellen und Kontrollen etabliert, die eine
Analyse der Proben mit einem konfokalen Laser-Rastermikroskop (CLSM) ermöglichte. Im
Gegensatz zu der Untersuchung RSV-exponierter Zellen im Durchflusszytometer oder einem
Fluoreszenzmikroskop erlaubt diese Methode eine schichtweise Durchleuchtung der Proben
und gibt dadurch genaueren Aufschluss über die Lokalisation der durch Antikörper markier-
ten RSV-Partikel.
Von den getesteten Antikörper-Kombinationen (s. 2.2.6) führten die Markierungen mit mono-
klonalen antiRSV-F, antiRSV-G oder antiRSV-P Antikörpern aus der Maus zu falsch positi-
ven Ergebnissen, da der sekundäre Ziege-antiMaus Antikörper auch die für die Zellsortierung

ERGEBNISSE 54
verwendeten Maus-antiCD3-Antikörper detektierte. Die für die Durchflusszytometrie
eingesetzten antiRSV und antiCD3-Antikörper, die direkt FITC- bzw. PE-gekoppelt waren
und deshalb keine Markierung mit Fluorochrom-konjugierten Sekundärantikörpern erfor-
derten, erwiesen sich als nicht auswertbar im CLSM. Daher wurde die Möglichkeit der
Detektion der für die Zellsortierung verwendeten CD3-Antikörper durch einen sekundären
Ziege-antiMaus-Alexa594 konjugierten Antikörper schließlich gezielt genutzt. In Kombina-
tion mit einem Ziege-antiRSV-Primärantikörper und einem Esel-antiZiege-Alexa488-
Sekundärantikörper ergab sich ein in Bezug auf Spezifität und Signalstärke sehr gutes
Markierungsergebnis (s. Abb. 3.7 und 3.8). Dabei wurde neben dem RSV-Nachweis auch eine
eindeutige Identifizierung des Zelltyps über den Nachweis des CD3-Oberflächenantigens als
T-Zellmarker erzielt.
Die CLSM-Bilder bestätigten die Befunde der Durchflusszytometrie, indem auch hier RSV-
Partikel an der Oberfläche von T-Zellen nachgewiesen werden konnten. Abbildung 3.7 zeigt
eine CLSM-Aufnahme von T-Zellen 2 h nach Stimulation mit PHA und RSV m.o.i. 1 und die
zugehörige Mock-Kontrolle. Die Aufnahmen erlaubten überdies die Aussage, dass die
Bindung von RSV an die T-Zellen räumlich getrennt vom TZR-Komplex erfolgte, da die
CD3-Signale und die RSV-Signale nicht kolokalisiert waren.
Darüber hinaus erbrachten die CSLM-Aufnahmen den Nachweis, dass RSV auch intrazellulär
in den T-Zellen vorlag. Proben, die 20 h nach RSV-Exposition fixiert und markiert wurden,
zeigten vereinzelt T-Zellen mit einem verbreiterten Zytoplasmasaum und darin liegenden
Ansammlungen von RSV-Partikeln (s. Abb. 3.8). In Proben von T-Zellen ohne RSV-
Stimulierung und in Proben, die 2 h nach der Virusexposition präpariert wurden, fanden sich
solche Strukturen nicht.
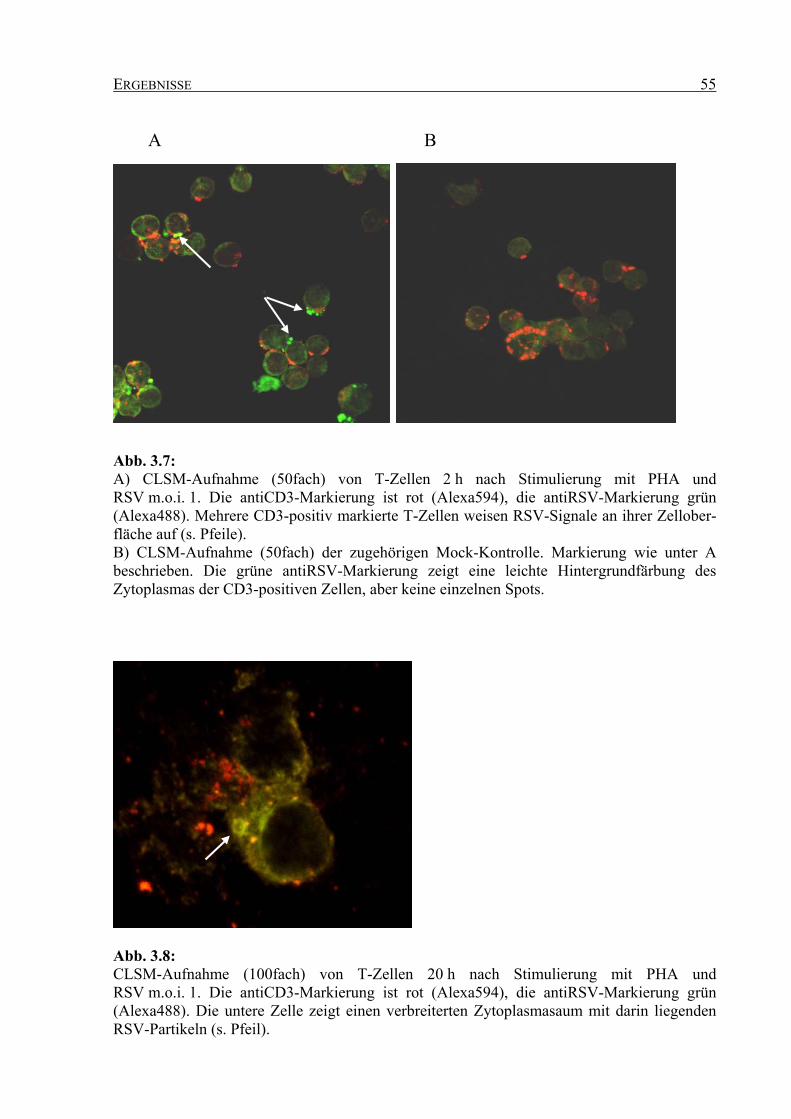
ERGEBNISSE 55
AbbA) RSV(AleflächB) CbescZyto
AbbCLSRSV(AleRSV
A
. 3.7: CLSM-Aufnahme (50fach) von T-Zell m.o.i. 1. Die antiCD3-Markierung ist roxa488). Mehrere CD3-positiv markierte T-e auf (s. Pfeile). LSM-Aufnahme (50fach) der zugehörige
hrieben. Die grüne antiRSV-Markierungplasmas der CD3-positiven Zellen, aber ke
. 3.8: M-Aufnahme (100fach) von T-Zellen m.o.i. 1. Die antiCD3-Markierung ist roxa488). Die untere Zelle zeigt einen verbr-Partikeln (s. Pfeil).
B
en 2 h nach Stimulierung mit PHA und t (Alexa594), die antiRSV-Markierung grün Zellen weisen RSV-Signale an ihrer Zellober-
n Mock-Kontrolle. Markierung wie unter A zeigt eine leichte Hintergrundfärbung des ine einzelnen Spots.
20 h nach Stimulierung mit PHA und t (Alexa594), die antiRSV-Markierung grün eiterten Zytoplasmasaum mit darin liegenden

ERGEBNISSE 56
3.2.3 Nachweis von RSV-RNA in RSV-exponierten T-Zellkulturen
Nachdem RSV-Partikel an der äußeren Zellmembran und intrazellulär bei T-Zellen
nachgewiesen werden konnten, war zu klären, ob auch eine Virus-Replikation in den Zellen
stattfand, also eine produktive Infektion in den Zellen vorlag. Dazu wurde RSV-RNA in
RSV-exponierten T-Zellkulturen im Zeitverlauf mittels einer quantitativen real-time PCR für
das RSV-Fusionsglykoprotein-Gen (RSV-F) nachgewiesen (s. 2.2.10) und die Ergebnisse mit
den Daten infizierter HEp-2 Zellen (Borg 2002), die eine produktive Virusreplikation zeigen,
verglichen.
In sechs T-Zellkulturen wurde 2h, 24h und 48h nach der Infektion die Menge an RSV-F RNA
in Doppelbestimmungen ermittelt. Dabei handelte es sich bei den je drei Infektionen mit
m.o.i. 1 und 3 zweimal um Ansätze, bei denen die Zellen eines Spenders parallel mit m.o.i. 1
und m.o.i. 3 infiziert wurden. Die Ct-Werte geben den PCR-Zyklus an, in dem die
Amplifikationskurve der jeweiligen Probe einen bestimmten Schwellenwert überschreitet. Je
höher die Ausgangsmenge an RSV-F spezifischer cDNA in der Probe ist, desto niedriger ist
der Ct-Wert dieser Probe. Unterscheiden sich die Ct-Werte zweier Proben um den Wert 1
(∆Ct = 1), so enthält die Probe mit dem niedrigeren Ct-Wert doppelt soviel RSV-F spezifische
cDNA wie die andere. (Weitere Einzelheiten zur Berechnung s. unter 2.2.9.4.) Die Ergebnisse
sind in Abbildung 3.9 zusammengefasst.
Aus der Infektion der T-Zellkulturen mit m.o.i. 1 bzw. 3 ergab sich eine von Beginn an hohe
Menge an spezifischer RSV-RNA in diesen Ansätzen, die sich in Ct-Werten zwischen 16
und 17,5 in den 2h-Proben niederschlug. Die Menge an RSV-F RNA in den 24h- und 48h-
Proben unterschied sich mit ∆Ct-Werten von +0,1 bis +1,0 jeweils nur unwesentlich von dem
entsprechenden 2h-Wert. Im Gegensatz dazu war in infizierten HEp-2 Zellkulturen innerhalb
von 24h eine deutliche Zunahme der RSV-F RNA zu verzeichnen. Das Sinken des Ct-Wertes
von 25,2 in der 2h-Kultur auf 19,0 in der 24h-Kultur bedeutet eine ungefähr 70fache
Erhöhung der viralen RNA-Menge.
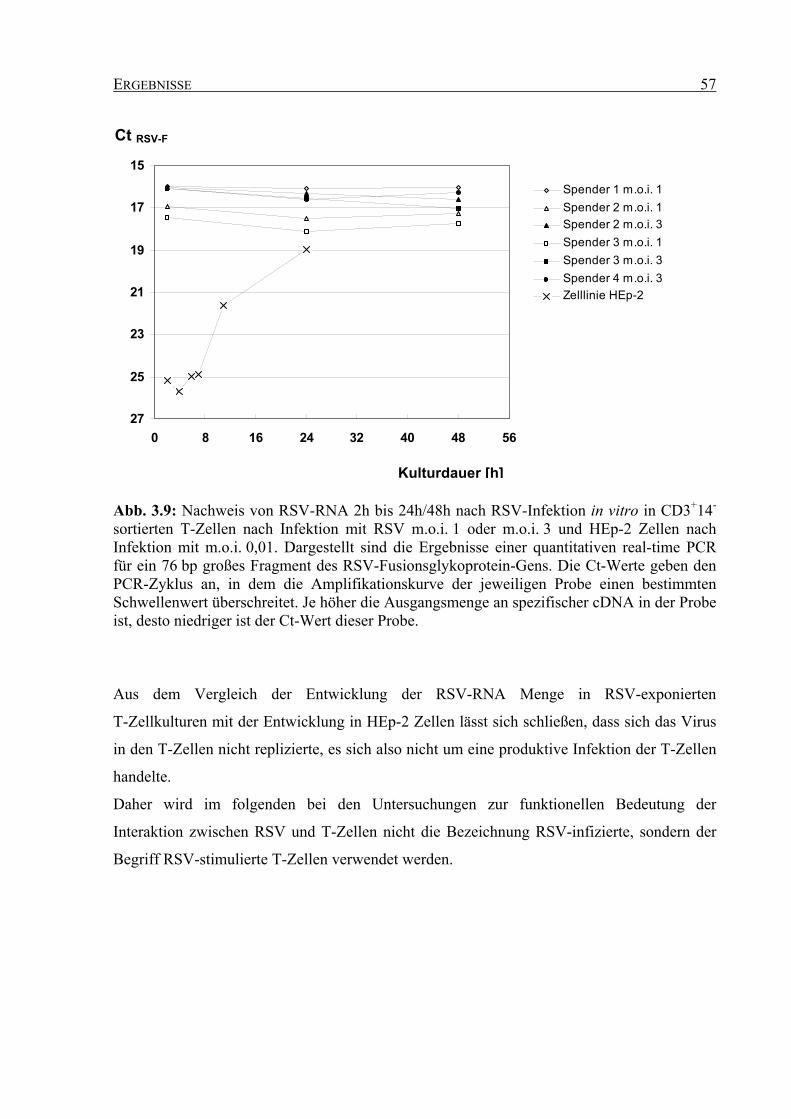
ERGEBNISSE 57
15
17
19
21
23
25
270 8 16 24 32 40 48 56
Spender 1 m.o.i. 1Spender 2 m.o.i. 1Spender 2 m.o.i. 3Spender 3 m.o.i. 1Spender 3 m.o.i. 3Spender 4 m.o.i. 3Zelllinie HEp-2
Ct RSV-F
Abb. 3.9: Nachweis von RSV-RNA 2h bissortierten T-Zellen nach Infektion mit RSInfektion mit m.o.i. 0,01. Dargestellt sind für ein 76 bp großes Fragment des RSV-FuPCR-Zyklus an, in dem die AmplifikatioSchwellenwert überschreitet. Je höher die Aist, desto niedriger ist der Ct-Wert dieser Pr
Aus dem Vergleich der Entwicklung
T-Zellkulturen mit der Entwicklung in HEp
in den T-Zellen nicht replizierte, es sich als
handelte.
Daher wird im folgenden bei den Un
Interaktion zwischen RSV und T-Zellen n
Begriff RSV-stimulierte T-Zellen verwende
Kulturdauer [h]
24h/48h nach RSV-Infektion in vitro in CD3+14- V m.o.i. 1 oder m.o.i. 3 und HEp-2 Zellen nach die Ergebnisse einer quantitativen real-time PCR sionsglykoprotein-Gens. Die Ct-Werte geben den nskurve der jeweiligen Probe einen bestimmten usgangsmenge an spezifischer cDNA in der Probe obe.
der RSV-RNA Menge in RSV-exponierten
-2 Zellen lässt sich schließen, dass sich das Virus
o nicht um eine produktive Infektion der T-Zellen
tersuchungen zur funktionellen Bedeutung der
icht die Bezeichnung RSV-infizierte, sondern der
t werden.

ERGEBNISSE 58
3.3 Nachweis von humanem Interleukin-10 mittels quantitativer real-time
PCR
3.3.1 Auswahl der Primer und Amplifikation der Zielsequenz
Zur Untersuchung des Einflusses der RSV-Stimulation auf die Interleukin-10 Expression von
T-Zellen wurde eine quantitative real-time PCR nach der TaqMan®-Methode (s. 2.2.9.1)
etabliert. Unter Berücksichtigung der Kriterien für den Entwurf von Primern und Sonde bei
dieser Methode (s. 2.2.9.2) wurden die Primer so gewählt, dass das entstehende PCR-Produkt
die Exon-Intron-Grenzen zwischen Exon 1 und Exon 2 des IL-10 Gens überspannte. Es
umfasste insgesamt 86 bp, davon 41 bp von Exon 1 und 45 bp von Exon 2. Genomische DNA
enthält zwischen diesen beiden Exons 865 bp Intron-Sequenz, wodurch eine Amplifikation
von genomischer DNA mit diesem PCR-System ausgeschlossen wurde. Das IL-10 Gen und
die Lage des amplifizierten PCR-Produktes sind in Abbildung 3.10 schematisch dargestellt.
Ein Abgleich der Primer- und Sondensequenzen mit der Sequenzdatenbank des NCBI
(National Center for Biotechnology Information, Bethesda, Maryland, USA) erbrachte außer
für die verschiedenen Einträge der IL-10 mRNA keine Übereinstimmung mit anderen
Bereichen des humanen Genoms.
GAGATCTCCG AGATGCCTTC AGCAGAGTGA AGACTTTCTT TCAAATGAAG 50 GATCAGCTGG ACAACTTGTT GTTAAAGGAG TCCTTG 100
IL-10 PCR-Produkt (86 bp)
Exon 4 Exon 5 Exon 3Exon 2 Exon 1
Abb. 3.10: Lage des IL-10 PCR-Produktes innerhalb des humanen IL-10 Gens.

ERGEBNISSE 59
Die Analyse der PCR-Ergebnisse nach einer quantitativen real-time PCR mittels der
GeneAmp SDS-Software ergab für jede Probe nach Abgleich gegen den im PCR-Ansatz
enthaltenen Referenzfarbstoff normalisierte Reportersignale (Rn), die aufgetragen gegen die
entsprechende Zykluszahl die Amplifikationskurve dieser Probe darstellten. In Abbil-
dung 3.11 sind diese Amplifikationskurven für eine dekadische Verdünnungsreihe von IL-10
Plasmid-DNA mit 100 – 0,0001 pg/PCR-Ansatz entsprechend 24 x 106 bis 24 Kopien/PCR-
Ansatz gezeigt. Aus dem Schnittpunkt mit der Schwellenwert-Linie (zur Festsetzung
s. 2.2.9.4) ergibt sich der jeweilige Ct-Wert der Probe. Die Abbildung zeigt die erwartete
Verschiebung der Kurven zu höheren Ct-Werten mit abnehmender IL-10 Konzentration.
100 pg IL-10 Plasmid-DNA ergaben einen Ct-Wert von 15,67, 0,0001 pg einen Ct-Wert von
34,18. Dabei entsprachen die Abstände zwischen den einzelnen dekadischen Verdünnungs-
schritten insbesondere im Konzentrationsbereich zwischen 101 und 10-3 pg mit ∆Ct-Werten
von 3,28 / 3,48 / 3,32 und 3,38 nahezu exakt dem theoretischen Wert von 3,33 für zwei
Proben mit zehnfachem Konzentrationsunterschied.
Rn
Zykluszahl Abb. 3.11: Amplifikationskurven aus der quantitativen real-time PCR für humanes IL-10. Gezeigt sind die Kurven für eine dekadische Verdünnungsreihe von IL-10 Plasmid-DNA mit Konzentrationen von 102 pg bis 10-4 pg (entsprechend 24 x 106 bis 24 Kopien) pro PCR-Ansatz. Die Ct-Werte der Proben, die der Zykluszahl am Schnittpunkt der Kurve mit der Schwellenwert-Linie (x-x-x) entsprechen, werden mit abnehmender Konzentration höher. (links außen: Amplifikationskurve des Ansatzes mit 102 pg, rechts außen: Amplifika-tionskurve des Ansatzes mit 10-4 pg IL-10 Plasmid-DNA.) Weitere Erläuterungen s. Text und Abschnitt 2.2.9.4.

ERGEBNISSE 60
3.3.2 Spezifität der quantitativen real-time PCR für humanes Interleukin-10
ur Überprüfung der Spezifität wurden die amplifizierten PCR-Produkte gelelektrophoretisch
bb. 3.12: Ergebnis einer Gelelektrophorese von PCR-Produkten aus verschiedenen IL-10
8: von cDNA aus RSV-stimulierten
Z
aufgetrennt. Die Analyse in einem 3 %igen Agarosegel (s. 2.2.9.5) zeigte für jede Probe eine
einzelne scharfe Bande ohne Nebenbanden und in der erwarteten Größe. Abbildung 3.12 zeigt
ein solches Gel mit amplifizierter IL-10 Plasmid-DNA (s. 2.2.9.6) in den Spuren 1 – 3 und
Proben aus RSV-stimulierten CD3+14- - und CD14+-Zellkulturansätzen in den Spuren 5 – 8.
Die PCR-Ansätze in den Spuren 1 – 3 enthielten von links nach rechts 100 pg, 10 pg bzw.
1 pg IL-10 Plasmid als Ausgangsmenge, zeigen jedoch in der Gelektrophorese einheitlich
starke Banden, da Unterschiede in der Ausgangsmenge in der Plateauphase der PCR nicht
mehr detektierbar sind. Die untere Marker-Bande in Spur 4 enthält ein 100 bp-Fragment, die
nächst höhere mit einem Laufstreckenunterschied von 7 mm ein 200 bp-Fragment. Die Lage
der Banden der PCR-Produkte 1 mm unterhalb der 100 bp Marker-Bande stimmt daher gut
mit der erwarteten Größe des PCR-Produktes von 86 bp überein.
Spur-Nr. 6 7 8 1 2 3 4 5
APCR-Läufen in einem 3 %igen Agarosegel. Die Spuren enthalten von links nach rechts folgende Proben: Spur 1 - 3: PCR-Produkte aus der Amplifizierung von 100 pg, 10 pg und 1 pg IL-10
Plasmid-DNA Spur 4: 2 µg DNA-Marker (100 bp-Leiter, MBI) Spur 5 – PCR-Produkte aus der Amplifizierung
+ - +-CD3 14 - und CD14 Zellkulturansätzen zweier verschiedener Spender (Spur 5 + 6: Spender A; Spur 7 + 8: Spender B)
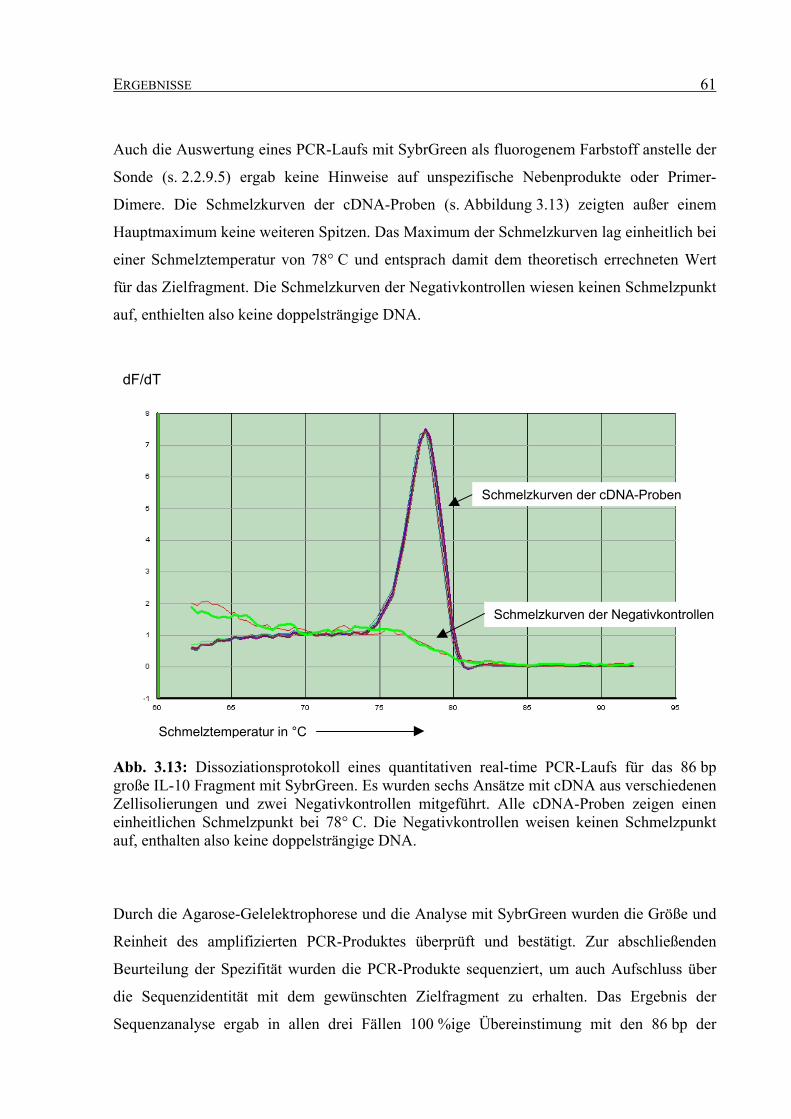
ERGEBNISSE 61
Auch die Auswertung eines PCR-Laufs mit SybrGreen als fluorogenem Farbstoff anstelle der
bb.
urch
Sonde (s. 2.2.9.5) ergab keine Hinweise auf unspezifische Nebenprodukte oder Primer-
Dimere. Die Schmelzkurven der cDNA-Proben (s. Abbildung 3.13) zeigten außer einem
Hauptmaximum keine weiteren Spitzen. Das Maximum der Schmelzkurven lag einheitlich bei
einer Schmelztemperatur von 78° C und entsprach damit dem theoretisch errechneten Wert
für das Zielfragment. Die Schmelzkurven der Negativkontrollen wiesen keinen Schmelzpunkt
auf, enthielten also keine doppelsträngige DNA.
T
C
n
Schmelzkurven der Negativkontrollen
AgroßeZelliseinheiauf, en
D
Reinh
Beurte
die S
Seque
Schmelztemperatur in °
dF/d
koll eines quantitativen
die Agarose-Gelelektrophorese und die Analyse m
3.13: Dissoziationsproto IL-10 Fragment mit SybrGreen. Es wurden sechs olierungen und zwei Negativkontrollen mitgefühtlichen Schmelzpunkt bei 78° C. Die Negativkonthalten also keine doppelsträngige DNA.
eit des amplifizierten PCR-Produktes überprüft
ilung der Spezifität wurden die PCR-Produkte s
equenzidentität mit dem gewünschten Zielfragm
nzanalyse ergab in allen drei Fällen 100 %ige
Schmelzkurven der cDNA-Probe
real-time PCR-Laufs für das 86 bp
it SybrGreen wurden die Größe und
Ansätze mit cDNA aus verschiedenen rt. Alle cDNA-Proben zeigen einen trollen weisen keinen Schmelzpunkt
und bestätigt. Zur abschließenden
equenziert, um auch Aufschluss über
ent zu erhalten. Das Ergebnis der
Übereinstimung mit den 86 bp der
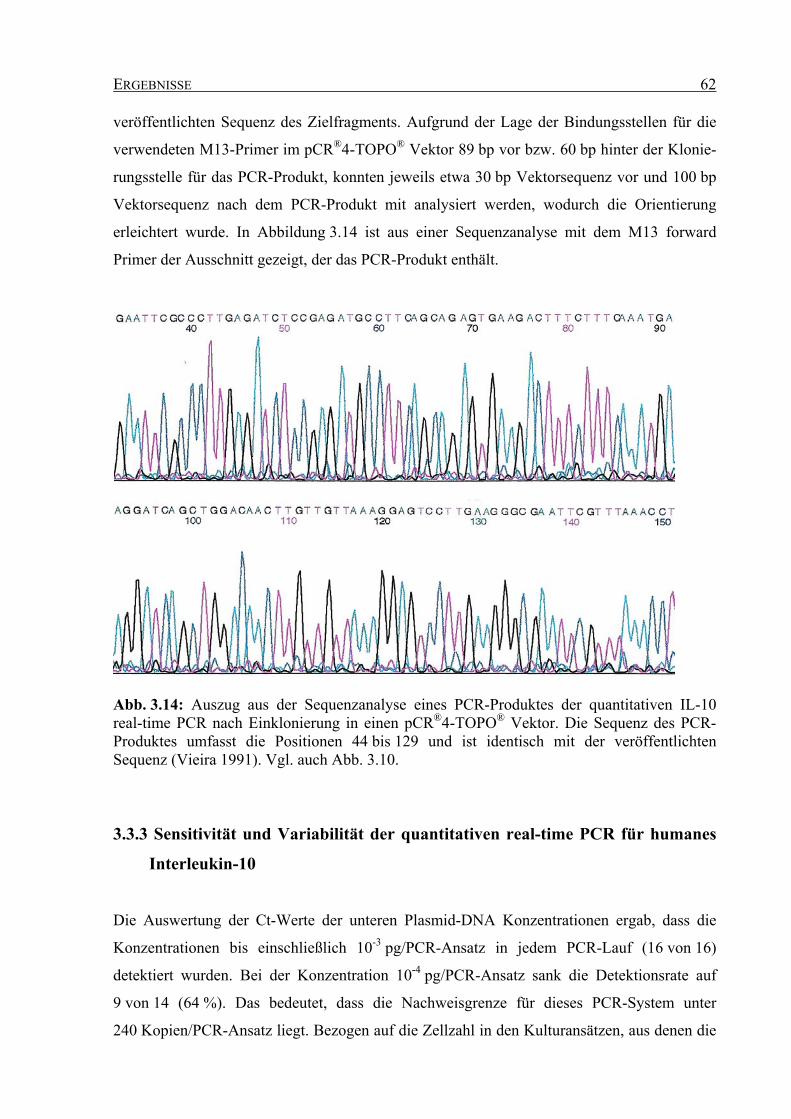
ERGEBNISSE 62
veröffentlichten Sequenz des Zielfragments. Aufgrund der Lage der Bindungsstellen für die
verwendeten M13-Primer im pCR®4-TOPO® Vektor 89 bp vor bzw. 60 bp hinter der Klonie-
rungsstelle für das PCR-Produkt, konnten jeweils etwa 30 bp Vektorsequenz vor und 100 bp
Vektorsequenz nach dem PCR-Produkt mit analysiert werden, wodurch die Orientierung
erleichtert wurde. In Abbildung 3.14 ist aus einer Sequenzanalyse mit dem M13 forward
Primer der Ausschnitt gezeigt, der das PCR-Produkt enthält.
bb. 3.14: Auszug aus der Sequenzanalyse eines PCR-Produktes der quantitativen IL-10
® ®
.3.3 Sensitivität und Variabilität der quantitativen real-time PCR für humanes
ie Auswertung der Ct-Werte der unteren Plasmid-DNA Konzentrationen ergab, dass die
Areal-time PCR nach Einklonierung in einen pCR 4-TOPO Vektor. Die Sequenz des PCR-Produktes umfasst die Positionen 44 bis 129 und ist identisch mit der veröffentlichten Sequenz (Vieira 1991). Vgl. auch Abb. 3.10.
3
Interleukin-10
D
Konzentrationen bis einschließlich 10-3 pg/PCR-Ansatz in jedem PCR-Lauf (16 von 16)
detektiert wurden. Bei der Konzentration 10-4 pg/PCR-Ansatz sank die Detektionsrate auf
9 von 14 (64 %). Das bedeutet, dass die Nachweisgrenze für dieses PCR-System unter
240 Kopien/PCR-Ansatz liegt. Bezogen auf die Zellzahl in den Kulturansätzen, aus denen die
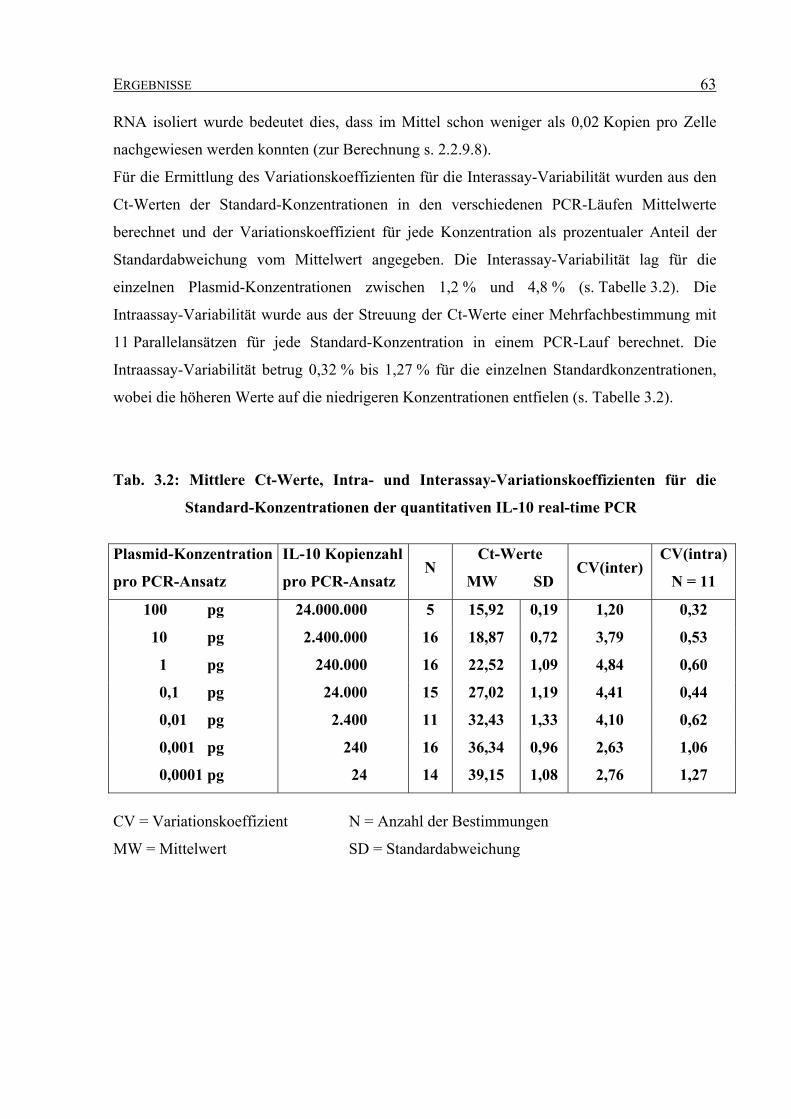
ERGEBNISSE 63
RNA isoliert wurde bedeutet dies, dass im Mittel schon weniger als 0,02 Kopien pro Zelle
nachgewiesen werden konnten (zur Berechnung s. 2.2.9.8).
Für die Ermittlung des Variationskoeffizienten für die Interassay-Variabilität wurden aus den
e
ab. 3.2: Mittlere Ct-Werte, Intra- und Interassay-Variationskoeffizienten für die
lasmid-Konzentration IL-10 Kopienzahl
N Ct-Werte
MW SD CV(inter)
CV(intra)
Ct-Werten der Standard-Konzentrationen in den verschiedenen PCR-Läufen Mittelwerte
berechnet und der Variationskoeffizient für jede Konzentration als prozentualer Anteil der
Standardabweichung vom Mittelwert angegeben. Die Interassay-Variabilität lag für die
einzelnen Plasmid-Konzentrationen zwischen 1,2 % und 4,8 % (s. Tabelle 3.2). Die
Intraassay-Variabilität wurde aus der Streuung der Ct-W rte einer Mehrfachbestimmung mit
11 Parallelansätzen für jede Standard-Konzentration in einem PCR-Lauf berechnet. Die
Intraassay-Variabilität betrug 0,32 % bis 1,27 % für die einzelnen Standardkonzentrationen,
wobei die höheren Werte auf die niedrigeren Konzentrationen entfielen (s. Tabelle 3.2).
T
Standard-Konzentrationen der quantitativen IL-10 real-time PCR
P
pro PCR-Ansatz pro PCR-Ansatz N = 11
100 pg 5 1,20 24.000.000 15,92 0,19 0,32
10 pg 2.400.000 16 18,87 0,72 3,79 0,53
1 pg 240.000 16 22,52 1,09 4,84 0,60
0,1 pg 24.000 15 27,02 1,19 4,41 0,44
0,01 pg 2.400 11 32,43 1,33 4,10 0,62
0,001 pg 240 16 36,34 0,96 2,63 1,06
0,0001 pg 24 14 39,15 1,08 2,76 1,27
V = Variationskoeffizient N = Anzahl der Bestimmungen C
MW = Mittelwert SD = Standardabweichung

ERGEBNISSE 64
3.4 Untersuchungen zur funktionellen Bedeutung der Interaktion zwischen
RSV und T-Zellen in vitro
3.4.1 Vitalität der virusstimulierten T-Zell-Kulturen
Im Rahmen der funktionellen Charakterisierung RSV-stimulierter T-Zellen wurde zunächst
der Einfluss der RSV-Exposition auf die Vitalität der Zellkulturen mittels Trypanblau-
Färbung (s. 2.2.1.4) überprüft. Nach 2h bis 48h Inkubation betrug die Vitalität der RSV-
stimulierten Kulturen 87,0 ± 7,6 % (N = 14) und unterschied sich damit nicht von der Vitalität
in den Kontrollkulturen, die bei 87,2 ± 5,8 % lag. Auch die Abnahme der Vitalität mit
zunehmender Kulturdauer verlief mit und ohne RSV etwa gleich. Nach 2h lag die Vitalität bei
92,7 % (RSV) bzw. 91,0 % (Kontrollen), nach 24h bei 85,0 % bzw. 85,7 % und nach 48h bei
83,5 % bzw. 85,8 %. Damit war sichergestellt, dass die folgenden Zytokinmessungen nicht
durch unterschiedliche Vitalität der Zellen beeinflusst wurden.
3.4.2 IL-10 mRNA-Expression in RSV-stimulierten T-Zellkulturen
Zunächst sollte geprüft werden, ob RSV-Stimulierung die IL-10 mRNA Expression von
T-Zellen beeinflusst. Dazu wurde die Expression von IL-10 mRNA in CD3+14- Kulturen im
Zeitverlauf mit der oben beschriebenen quantitativen real-time PCR untersucht (Abb. 3.15).
Frisch isolierte CD3+14- Zellen exprimierten sehr geringe, aber mit der quantitativen PCR
messbare Mengen an IL-10 mRNA. Die entsprechenden Ct-Werte lagen bei 37,31 ± 1,22
(N = 12), das entspricht einen Kopienzahl von etwa 5 pro 1000 Zellen. In der Zellkultur stieg
die IL-10 mRNA-Expression sowohl in unstimulierten als auch in RSV-stimulierten Kulturen
zunächst an (Abb. 3.15). Während diese Zunahme in den unstimulierten Kulturen bereits nach
1 h wieder rückläufig war, hielt sie in RSV-stimulierten Kulturen insgesamt länger an,
erreichte spenderabhängig nach 18 bis 24h ihr Maximum und sank dann ebenfalls ab.
In Abbildung 3.15 A ist exemplarisch der Verlauf in der frühen Phase für die unstimulierte
sowie für RSV-stimulierte Kulturen (m.o.i. 1 und 3) eines Spenders gezeigt. In Abbil-
dung 3.15 B sind die Ergebnisse der Kinetiken von unstimulierten und RSV(m.o.i. 3)-
stimulierten Kulturen zusammengefasst (N = 4). Den Auswertungen liegen die Ergebnisse der

ERGEBNISSE 65
relativen Quantifizierung zugrunde, bei der für jede Probe ein ∆Ct-Wert aus den PCR-
Ergebnissen für IL-10 und das Referenz-Gen β-Glucuronidase ermittelt wurde, so dass
Ergebnisse aus verschiedenen Experimenten direkt miteinander verglichen werden konnten
(s. 2.2.9.4). Für die Darstellung wurde die Nulllinie auf ein ∆Ct von –10 gelegt.
0
2
4
6
8
0h 0.5h 1h 2h 18h 24h0
2
4
6
8
0h 2h 24h 48h
RSV m.o.i. 3RSV m.o.i. 1unstimuliert
Kulturdauer [h]
B A ∆ Ct
Abb. 3.15: Kinetik der IL-10 mRNA-Expression in CD3+14- T-Zellkulturen A) Verlauf in den ersten 24h der Kultur für unstimulierte und RSV-stimulierte Ansätze eines
Spenders B) Verlauf über 48h für unstimulierte und RSV(m.o.i. 3)-stimulierte Ansätze von 4 Spendern
(Mittelwerte und Standardabweichung).
Aufgrund der ermittelten Zeitkinetik für die IL-10 mRNA Expression in T-Zellen wurde für
die folgenden Experimente zur Stimulation von CD3+14- Zellen durch RSV eine Kulturdauer
von 24h gewählt. Für die Auswertung wurden aus der Differenz der ∆Ct-Werte der RSV-
stimulierten Proben und dem ∆Ct-Wert der entsprechenden unstimulierten 24h-Probe ∆∆Ct-
Werte berechnet, die den ‚Netto’-Effekt der Virusstimulation darstellen. Aus den ∆∆Ct-
Werten wurde für jede Probe der Faktor der Zunahme der IL-10 Expression durch die RSV-
Stimulation ermittelt (s. 2.2.9.4). Die Ergebnisse sind in Abbildung 3.16 gezeigt. Von
insgesamt 12 Spendern reagierten die CD3+14- Zellen eines Spenders nicht mit einer
Zunahme der IL-10 Expression auf die RSV-Stimulierung in der Kultur. In den übrigen
Kulturen nahm die IL-10 mRNA bei RSV-Stimulierung um den Faktor 1,1 bis 12 bei einer
m.o.i von 1 und um den Faktor 1,2 bis 14 bei einer m.o.i. von 3 zu. Dabei war in der Hälfte
der Kulturen, die parallel mit m.o.i. 1 und 3 stimuliert wurden (N = 8) ein weiterer Anstieg
der IL-10 mRNA Expression bei der höheren m.o.i. zu beobachten. Zusammengefasst zeigte
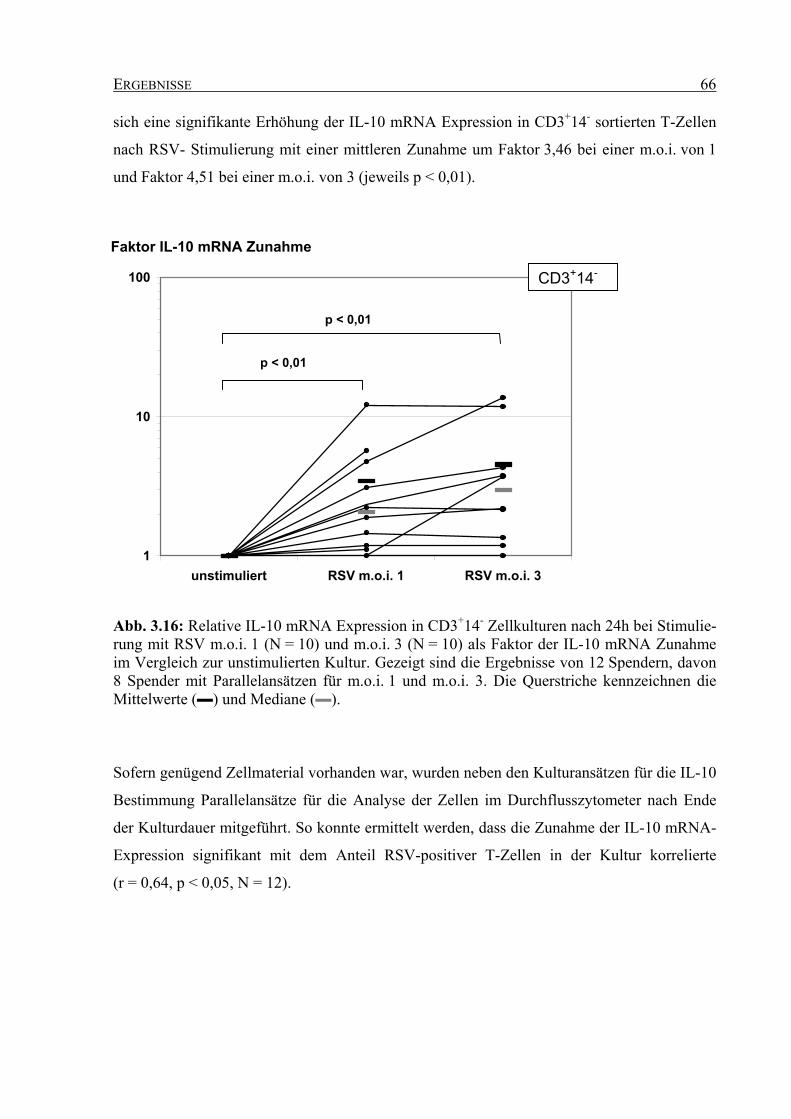
ERGEBNISSE 66
sich eine signifikante Erhöhung der IL-10 mRNA Expression in CD3+14- sortierten T-Zellen
nach RSV- Stimulierung mit einer mittleren Zunahme um Faktor 3,46 bei einer m.o.i. von 1
und Faktor 4,51 bei einer m.o.i. von 3 (jeweils p < 0,01).
e
Faktor IL-10 mRNA Zunahm1
10
100
unstimuliert RSV m.o.i. 1 RSV m.o.i.
p < 0,01
p < 0,01
Abb. 3.16: Relative IL-10 mRNA Expression in CD3+14- Zellkurung mit RSV m.o.i. 1 (N = 10) und m.o.i. 3 (N = 10) als Faktorim Vergleich zur unstimulierten Kultur. Gezeigt sind die Ergebn8 Spender mit Parallelansätzen für m.o.i. 1 und m.o.i. 3. Die QMittelwerte (▬) und Mediane (▬).
Sofern genügend Zellmaterial vorhanden war, wurden neben den
Bestimmung Parallelansätze für die Analyse der Zellen im Dur
der Kulturdauer mitgeführt. So konnte ermittelt werden, dass die
Expression signifikant mit dem Anteil RSV-positiver T-Zelle
(r = 0,64, p < 0,05, N = 12).
CD3+14-
3
lturen nach 24h bei Stimulie- der IL-10 mRNA Zunahme isse von 12 Spendern, davon uerstriche kennzeichnen die
Kulturansätzen für die IL-10
chflusszytometer nach Ende
Zunahme der IL-10 mRNA-
n in der Kultur korrelierte

ERGEBNISSE 67
3.4.3 IL-10 mRNA-Expression in T-Zellkulturen mit anderen Stimuli
Um beurteilen zu können, ob die beobachteten Zunahme der IL-10 Expression in CD3+14-
Zellen eine spezifische Antwort auf RSV darstellte, wurden parallel zur RSV-Stimulation
auch Ansätze mit Newcastle Disease Virus (NDV), einem dem RSV verwandten, aber nicht
humanpathogenen Virus aus der Gruppe der Paramyxoviren, stimuliert. Zusätzlich wurde der
Einfluss von antiCD28-Antikörpern als nicht-viraler Stimulus allein und in Kombination mit
den viralen Stimuli untersucht.
Nach Stimulierung mit NDV in einer Konzentration von 3,2 hämagglutinierenden Einheiten
pro ml Zellkultur (s. 2.2.3) zeigte sich kein einheitlicher Trend der IL-10 mRNA Expression
im Vergleich zu den Kontrollen. In zwei von fünf Fällen lag die IL-10 mRNA Expression
jeweils unter dem Wert der unstimulierten Proben. Im Mittel war die Expression in den NDV-
stimulierten Ansätzen mit einem Faktor von 1,3 [0,2 – 4,1] gegenüber den unstimulierten
Kontrollen nur leicht erhöht, gegenüber den RSV-stimulierten Proben aber um den Faktor 2,0
[0,6 - 28,8; m.o.i. 1] bzw. 2,9 [0,5 – 83,3; m.o.i. 3] erniedrigt.
Zugabe von antiCD28-Antikörper zu den Kulturen (Endkonzentration 1 µg/ml) sollte eine
mäßige Aktivierung der T-Zellen durch suboptimale polyklonale Stimulation bewirken.
Dadurch sollte die IL-10 Antwort aktivierter T-Zellen auf die RSV-Stimulierung untersucht
werden, ohne dass durch den nicht-viralen Stimulus allein bereits eine maximale Stimulation
der Zellen erfolgte.
Es zeigte sich, dass Zusatz von antiCD28 in den Kulturen ohne Virusstimulation und in den
Ansätzen mit NDV zu einer deutlich gesteigerten IL-10 Expression führte, während die
Expressionsrate in den RSV-stimulierten Kulturen nur leicht zunahm. Diese geringe Zunahme
korrelierte jedoch wiederum mit einem erhöhten Anteil an RSV-positiven T-Zellen in den
RSV/antiCD28-stimulierten Kulturen (r = 0,71, p < 0,05, N = 10). Die Ergebnisse sind in
Abbildung 3.17 zusammengestellt.
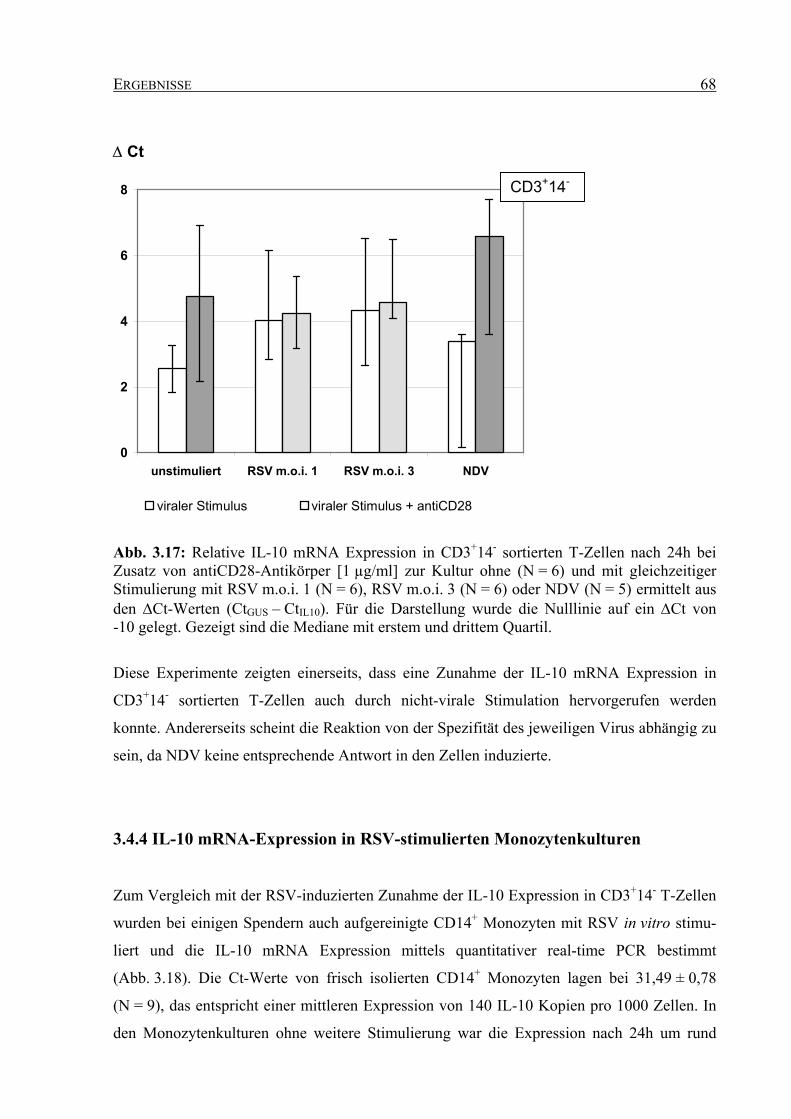
ERGEBNISSE 68
0
2
4
6
8
unstimuliert RSV m.o.i. 1 RSV m.o.i. 3 NDV
viraler Stimulus
∆ Ct
viraler Stimulus + antiCD28
Abb. 3.17: Relative IL-10 mRNA Expression in CD3+14- Zusatz von antiCD28-Antikörper [1 µg/ml] zur Kultur ohnStimulierung mit RSV m.o.i. 1 (N = 6), RSV m.o.i. 3 (N = 6den ∆Ct-Werten (CtGUS – CtIL10). Für die Darstellung wurd-10 gelegt. Gezeigt sind die Mediane mit erstem und drittem
Diese Experimente zeigten einerseits, dass eine Zunahme
CD3+14- sortierten T-Zellen auch durch nicht-virale Sti
konnte. Andererseits scheint die Reaktion von der Spezifität
sein, da NDV keine entsprechende Antwort in den Zellen ind
3.4.4 IL-10 mRNA-Expression in RSV-stimulierten M
Zum Vergleich mit der RSV-induzierten Zunahme der IL-10
wurden bei einigen Spendern auch aufgereinigte CD14+ Mo
liert und die IL-10 mRNA Expression mittels quantita
(Abb. 3.18). Die Ct-Werte von frisch isolierten CD14+ M
(N = 9), das entspricht einer mittleren Expression von 140 I
den Monozytenkulturen ohne weitere Stimulierung war die
CD3+14-
sortierten T-Zellen nach 24h bei e (N = 6) und mit gleichzeitiger ) oder NDV (N = 5) ermittelt aus e die Nulllinie auf ein ∆Ct von Quartil.
der IL-10 mRNA Expression in
mulation hervorgerufen werden
des jeweiligen Virus abhängig zu
uzierte.
onozytenkulturen
Expression in CD3+14- T-Zellen
nozyten mit RSV in vitro stimu-
tiver real-time PCR bestimmt
onozyten lagen bei 31,49 ± 0,78
L-10 Kopien pro 1000 Zellen. In
Expression nach 24h um rund
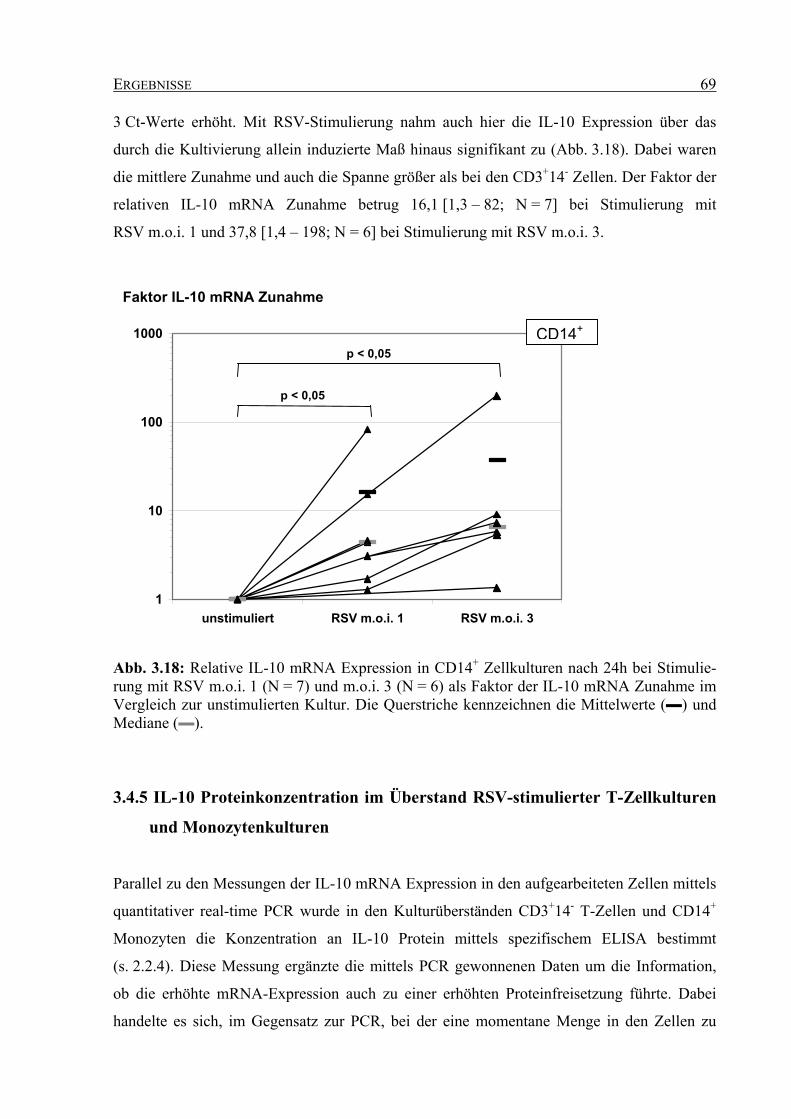
ERGEBNISSE 69
3 Ct-Werte erhöht. Mit RSV-Stimulierung nahm auch hier die IL-10 Expression über das
durch die Kultivierung allein induzierte Maß hinaus signifikant zu (Abb. 3.18). Dabei waren
die mittlere Zunahme und auch die Spanne größer als bei den CD3+14- Zellen. Der Faktor der
relativen IL-10 mRNA Zunahme betrug 16,1 [1,3 – 82; N = 7] bei Stimulierung mit
RSV m.o.i. 1 und 37,8 [1,4 – 198; N = 6] bei Stimulierung mit RSV m.o.i. 3.
e
Faktor IL-10 mRNA Zunahm1
10
100
1000
unstimuliert RSV m.o.i. 1 RSV m.o.i. 3
p < 0,05
p < 0,05
Abb. 3.18: Relative IL-10 mRNA Expression in CD14+ Zellkulrung mit RSV m.o.i. 1 (N = 7) und m.o.i. 3 (N = 6) als Faktor deVergleich zur unstimulierten Kultur. Die Querstriche kennzeichnMediane (▬).
3.4.5 IL-10 Proteinkonzentration im Überstand RSV-sti
und Monozytenkulturen
Parallel zu den Messungen der IL-10 mRNA Expression in den a
quantitativer real-time PCR wurde in den Kulturüberständen C
Monozyten die Konzentration an IL-10 Protein mittels spe
(s. 2.2.4). Diese Messung ergänzte die mittels PCR gewonnenen
ob die erhöhte mRNA-Expression auch zu einer erhöhten Prot
handelte es sich, im Gegensatz zur PCR, bei der eine momenta
CD14+
turen nach 24h bei Stimulie-r IL-10 mRNA Zunahme im en die Mittelwerte (▬) und
mulierter T-Zellkulturen
ufgearbeiteten Zellen mittels
D3+14- T-Zellen und CD14+
zifischem ELISA bestimmt
Daten um die Information,
einfreisetzung führte. Dabei
ne Menge in den Zellen zu

ERGEBNISSE 70
einem bestimmten Zeitpunkt (hier 24h) ermittelt wurde, beim ELISA um eine kumulative
Messung, der die gesamte Zellaktivität im Verlauf der 24h Kulturdauer zugrunde lag.
Es zeigte sich, dass die IL-10 Konzentrationen in CD3+14- T-Zellkulturen bei 10 von 12
Spendern nach 24h und 48h sowohl unstimuliert als auch mit RSV-Stimulierung unterhalb der
quantitativen Messgrenze des ELISA von 7,8 pg/ml lagen. Lediglich bei zwei Spendern
ließen sich mit 9,4 pg/ml und 10,0 pg/ml Werte für die RSV(m.o.i. 3)-stimulierten Proben
ermitteln, während die korrespondierenden RSV m.o.i. 1- und unstimulierten Ansätze unter
7,8 pg/ml lagen. In den Kulturen mit Zusatz von antiCD28-Antikörpern waren bei den 24h-
Kulturen 2 von 8 und bei den 48h-Kulturen 2 von 4 im ELISA quantifizierbar mit einer
mittleren IL-10 Proteinkonzentration von 25,2 [0 - 229,6] pg/ml. Dabei lagen die Konzen-
trationen nach 48h jeweils höher als die vergleichbaren 24h-Werte.
In den Überständen der CD14+ Zellkulturen waren unstimuliert nach 24h 2 von 8 Kulturen
quantifizierbar mit einer mittleren IL-10 Proteinkonzentrationen von 12,9 [0 – 80,2] pg/ml. In
den korrespondierenden RSV-stimulierten Ansätzen waren die IL-10 Konzentrationen
signifikant (p < 0,05) erhöht mit 6 von 7 Proben (m.o.i. 1) und 5 von 5 (m.o.i. 3) > 7,8 pg/ml
und mittleren Konzentrationen von 65,2 [0 – 173,0] bzw. 205,6 [25,9 – 579,2] pg/ml. Die
vorhandenen 48h-Überstände wiesen wieder jeweils höhere Konzentrationen auf als
vergleichbare 24h-Kulturen.
In Verbindung mit den zugehörigen PCR-Daten der CD3+14- und der CD14+ Zellen (s. Abb.
3.16 und 3.18) ergab sich eine positive Korrelation zwischen der IL-10 Expressionsrate und
der IL-10 Proteinkonzentration in den Kulturüberständen. Bei Korrelation der beiden Werte
aus den 24h-Kulturen (Abb. 3.19) ergab sich für beide Populationen zusammengefasst ein
Korrelationskoeffizient von r = 0,80 (p < 0,001; N = 96). Korrelierte man die ELISA-Daten
von 48h-Kulturen mit den PCR-Werten der zugehörigen 24h-Kulturen so stieg der
Korrelationskoeffizient auf r = 0,91 (p < 0,001; N = 37). Darin kommt zum Ausdruck, dass
eine hohe Expressionsrate zum Zeitpunkt 24h erst zeitversetzt nach Ablauf der Protein-
biosynthese und Ausschleusung der Proteinmoleküle zu einer erhöhten Zytokinfreisetzung
führt, die im Kulturüberstand mittels ELISA messbar wird.
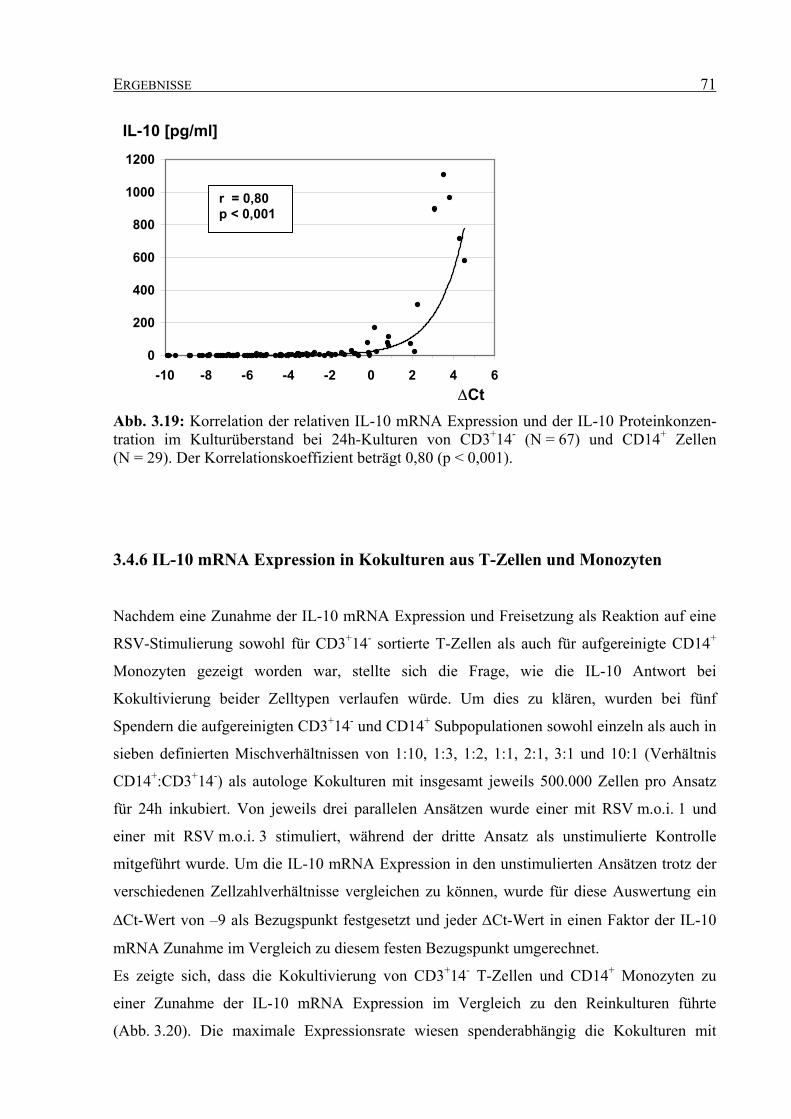
ERGEBNISSE 71
0
200
400
600
800
1000
1200
-10 -8 -6 -4 -2 0 2 4 6
IL-10 [pg/ml]
r = 0,80 p < 0,001
∆CtAbb. 3.19: Korrelation der relativen IL-10 mRNA Expression und der IL-10 Proteinkonzen-tration im Kulturüberstand bei 24h-Kulturen von CD3+14- (N = 67) und CD14+ Zellen (N = 29). Der Korrelationskoeffizient beträgt 0,80 (p < 0,001).
3.4.6 IL-10 mRNA Expression in Kokulturen aus T-Zellen und Monozyten
Nachdem eine Zunahme der IL-10 mRNA Expression und Freisetzung als Reaktion auf eine
RSV-Stimulierung sowohl für CD3+14- sortierte T-Zellen als auch für aufgereinigte CD14+
Monozyten gezeigt worden war, stellte sich die Frage, wie die IL-10 Antwort bei
Kokultivierung beider Zelltypen verlaufen würde. Um dies zu klären, wurden bei fünf
Spendern die aufgereinigten CD3+14- und CD14+ Subpopulationen sowohl einzeln als auch in
sieben definierten Mischverhältnissen von 1:10, 1:3, 1:2, 1:1, 2:1, 3:1 und 10:1 (Verhältnis
CD14+:CD3+14-) als autologe Kokulturen mit insgesamt jeweils 500.000 Zellen pro Ansatz
für 24h inkubiert. Von jeweils drei parallelen Ansätzen wurde einer mit RSV m.o.i. 1 und
einer mit RSV m.o.i. 3 stimuliert, während der dritte Ansatz als unstimulierte Kontrolle
mitgeführt wurde. Um die IL-10 mRNA Expression in den unstimulierten Ansätzen trotz der
verschiedenen Zellzahlverhältnisse vergleichen zu können, wurde für diese Auswertung ein
∆Ct-Wert von –9 als Bezugspunkt festgesetzt und jeder ∆Ct-Wert in einen Faktor der IL-10
mRNA Zunahme im Vergleich zu diesem festen Bezugspunkt umgerechnet.
Es zeigte sich, dass die Kokultivierung von CD3+14- T-Zellen und CD14+ Monozyten zu
einer Zunahme der IL-10 mRNA Expression im Vergleich zu den Reinkulturen führte
(Abb. 3.20). Die maximale Expressionsrate wiesen spenderabhängig die Kokulturen mit
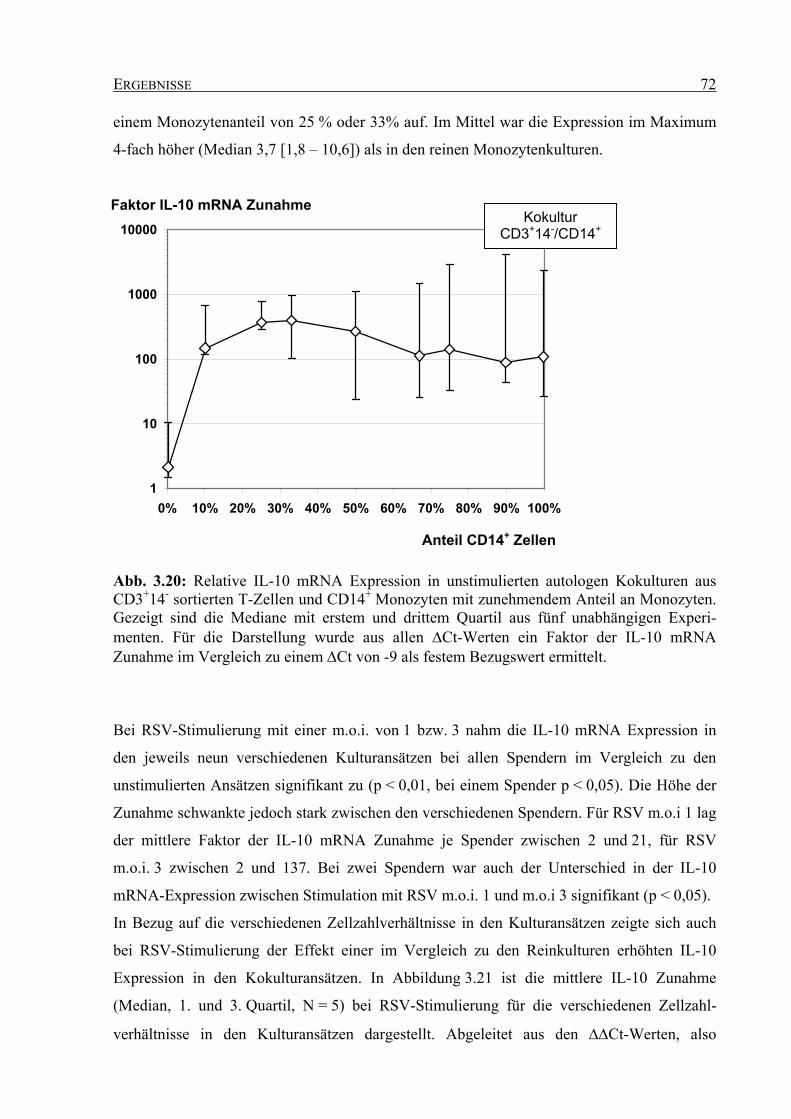
ERGEBNISSE 72
einem Monozytenanteil von 25 % oder 33% auf. Im Mittel war die Expression im Maximum
4-fach höher (Median 3,7 [1,8 – 10,6]) als in den reinen Monozytenkulturen.
1
10
100
1000
10000
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
Faktor IL-10 mRNA Zunahme Kokultur
CD3+14-/CD14+
Anteil CD14+ Zellen Abb. 3.20: Relative IL-10 mRNA Expression in unstimulierten autologen Kokulturen aus CD3+14- sortierten T-Zellen und CD14+ Monozyten mit zunehmendem Anteil an Monozyten. Gezeigt sind die Mediane mit erstem und drittem Quartil aus fünf unabhängigen Experi-menten. Für die Darstellung wurde aus allen ∆Ct-Werten ein Faktor der IL-10 mRNA Zunahme im Vergleich zu einem ∆Ct von -9 als festem Bezugswert ermittelt.
Bei RSV-Stimulierung mit einer m.o.i. von 1 bzw. 3 nahm die IL-10 mRNA Expression in
den jeweils neun verschiedenen Kulturansätzen bei allen Spendern im Vergleich zu den
unstimulierten Ansätzen signifikant zu (p < 0,01, bei einem Spender p < 0,05). Die Höhe der
Zunahme schwankte jedoch stark zwischen den verschiedenen Spendern. Für RSV m.o.i 1 lag
der mittlere Faktor der IL-10 mRNA Zunahme je Spender zwischen 2 und 21, für RSV
m.o.i. 3 zwischen 2 und 137. Bei zwei Spendern war auch der Unterschied in der IL-10
mRNA-Expression zwischen Stimulation mit RSV m.o.i. 1 und m.o.i 3 signifikant (p < 0,05).
In Bezug auf die verschiedenen Zellzahlverhältnisse in den Kulturansätzen zeigte sich auch
bei RSV-Stimulierung der Effekt einer im Vergleich zu den Reinkulturen erhöhten IL-10
Expression in den Kokulturansätzen. In Abbildung 3.21 ist die mittlere IL-10 Zunahme
(Median, 1. und 3. Quartil, N = 5) bei RSV-Stimulierung für die verschiedenen Zellzahl-
verhältnisse in den Kulturansätzen dargestellt. Abgeleitet aus den ∆∆Ct-Werten, also
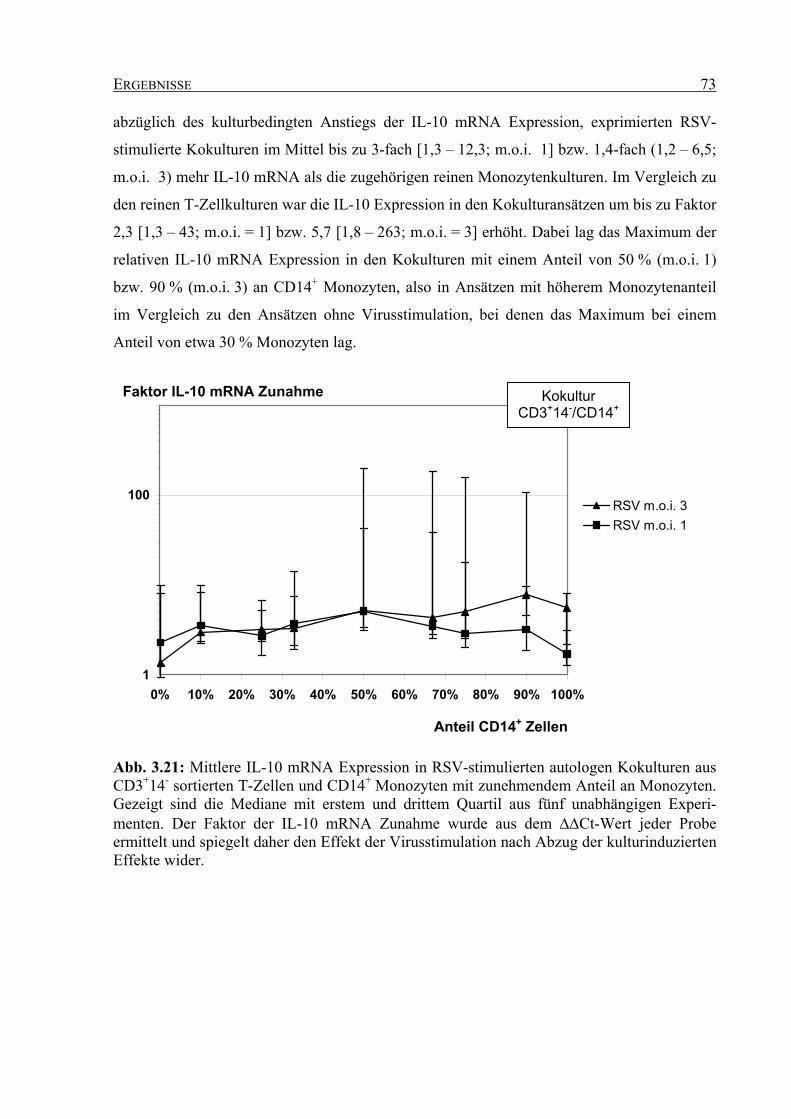
ERGEBNISSE 73
abzüglich des kulturbedingten Anstiegs der IL-10 mRNA Expression, exprimierten RSV-
stimulierte Kokulturen im Mittel bis zu 3-fach [1,3 – 12,3; m.o.i. 1] bzw. 1,4-fach (1,2 – 6,5;
m.o.i. 3) mehr IL-10 mRNA als die zugehörigen reinen Monozytenkulturen. Im Vergleich zu
den reinen T-Zellkulturen war die IL-10 Expression in den Kokulturansätzen um bis zu Faktor
2,3 [1,3 – 43; m.o.i. = 1] bzw. 5,7 [1,8 – 263; m.o.i. = 3] erhöht. Dabei lag das Maximum der
relativen IL-10 mRNA Expression in den Kokulturen mit einem Anteil von 50 % (m.o.i. 1)
bzw. 90 % (m.o.i. 3) an CD14+ Monozyten, also in Ansätzen mit höherem Monozytenanteil
im Vergleich zu den Ansätzen ohne Virusstimulation, bei denen das Maximum bei einem
Anteil von etwa 30 % Monozyten lag.
1
100
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
RSV m.o.i. 3RSV m.o.i. 1
e Kokultur CD3+14-/CD14+
Abb. 3.21: Mittlere IL-10 mRNA Expression inCD3+14- sortierten T-Zellen und CD14+ MonozyGezeigt sind die Mediane mit erstem und dritmenten. Der Faktor der IL-10 mRNA Zunahmermittelt und spiegelt daher den Effekt der VirusEffekte wider.
Anteil CD14+ Zellen
Faktor IL-10 mRNA Zunahm
RSV-stimulierten autologen Kokulturen aus ten mit zunehmendem Anteil an Monozyten. tem Quartil aus fünf unabhängigen Experi-e wurde aus dem ∆∆Ct-Wert jeder Probe
stimulation nach Abzug der kulturinduzierten

ERGEBNISSE 74
3.4.7 IL-10 Proteinkonzentration im Überstand RSV-stimulierter Kokulturen aus
T-Zellen und Monozyten
Auch bei den Kokulturen aus CD3+14- T-Zellen und CD14+ Monozyten wurde parallel zu den
Messungen der IL-10 mRNA Expression in den aufgearbeiteten Zellen mittels quantitativer
real-time PCR die IL-10 Konzentrationen in den Kulturüberständen mittels spezifischem
ELISA bestimmt. Wie in Abbildung 3.22 zu sehen ist, zeigte sich auch in den Kokulturen eine
positive Korrelation zwischen der IL-10 mRNA Expression und der IL-10 Protein-
konzentration im Überstand bei den 24h-Ansätzen (r = 0,90, p < 0,001, N = 129).
0
500
1000
1500
2000
2500
-10 -8 -6 -4 -2 0 2 4 6
IL-10 [pg/ml] Kokultur
CD3+14-/CD14+
r = 0,90 p < 0,001
∆Ct
Abb. 3.22: Korrelation zwischen relativer IL-10 mRNA Expression und IL-10 Proteinkon-zentration im Überstand von Kokulturen aus CD3+14- mit CD14+ Zellen. Der Auswertung liegen die jeweils 9 unstimulierten und RSV-stimulierten 24h-Ansätze mit verschiedenen Zellzahlverhältnissen zugrunde (N = 129). Der Korrelationskoeffizient beträgt 0,90 (p < 0,001).
Wie bei der mRNA Expression zeigte sich auch bei der IL-10 Proteinkonzentration im
Überstand eine Zunahme in den Mischkulturen gegenüber den Reinkulturen (Abb. 3.23). Für
die Auswertung wurden die Werte der RSV-stimulierten Kulturen mit den Werten der
unstimulierten Kultur jeweils saldiert, um den RSV-induzierten Effekt bereinigt um den
kulturbedingten Anstieg zu erhalten.
Bei Stimulierung mit RSV m.o.i. 1 war die IL-10 Konzentration im Maximum, das bei einem
Monozytenanteil von 33 % lag, 4,4 [1,9 – 44,6] mal höher als in den reinen Monozyten-
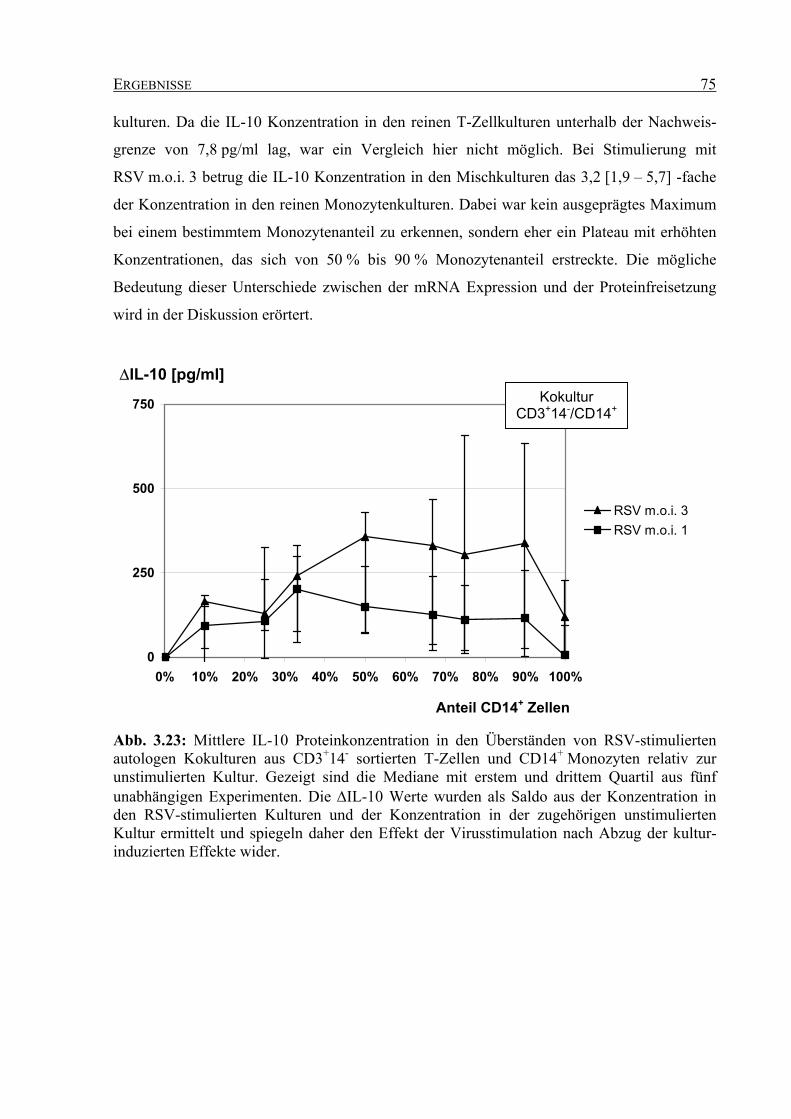
ERGEBNISSE 75
kulturen. Da die IL-10 Konzentration in den reinen T-Zellkulturen unterhalb der Nachweis-
grenze von 7,8 pg/ml lag, war ein Vergleich hier nicht möglich. Bei Stimulierung mit
RSV m.o.i. 3 betrug die IL-10 Konzentration in den Mischkulturen das 3,2 [1,9 – 5,7] -fache
der Konzentration in den reinen Monozytenkulturen. Dabei war kein ausgeprägtes Maximum
bei einem bestimmtem Monozytenanteil zu erkennen, sondern eher ein Plateau mit erhöhten
Konzentrationen, das sich von 50 % bis 90 % Monozytenanteil erstreckte. Die mögliche
Bedeutung dieser Unterschiede zwischen der mRNA Expression und der Proteinfreisetzung
wird in der Diskussion erörtert.
0
250
500
750
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
RSV m.o.i. 3RSV m.o.i. 1
∆IL-10 [pg/ml] Kokultur
CD3+14-/CD14+
Abb. 3.23: Mittlere IL-10 Proteinkonzentrationautologen Kokulturen aus CD3+14- sortierten Tunstimulierten Kultur. Gezeigt sind die Medianunabhängigen Experimenten. Die ∆IL-10 Werteden RSV-stimulierten Kulturen und der KonzeKultur ermittelt und spiegeln daher den Effekt dinduzierten Effekte wider.
Anteil CD14+ Zellen
in den Überständen von RSV-stimulierten -Zellen und CD14+ Monozyten relativ zur e mit erstem und drittem Quartil aus fünf wurden als Saldo aus der Konzentration in ntration in der zugehörigen unstimulierten er Virusstimulation nach Abzug der kultur-

ERGEBNISSE 76
3.4.8 IL-10 Proteinkonzentration im Überstand RSV-stimulierter Kokulturen aus
Alveolarmakrophagen und T-Zellen
Im Zusammenhang mit einer möglichen Bedeutung von RSV-Infektionen für Exazerbationen
der COPD wurden auch Kokulturen aus Alveolarmakrophagen und T-Zellen untersucht.
Alveolarmakrophagen aus der Spülflüssigkeit bronchoalveolärer Lavagen (s. 2.2.1.3) wurden
zusammen mit autologen CD3+-sortierten T-Zellen aus dem peripheren Blut in vitro mit
RSV m.o.i. 1 und PHA [Endkonzentration 2,4 µg/ml] oder nur mit PHA stimuliert und die
IL-10 Proteinkonzentration mittels ELISA in den Kulturüberständen nach 48h gemessen. Bei
der Variation der Zellzahlverhältnisse wurden aufgrund der begrenzten Zahl an Alveolar-
makrophagen je 500.000 T-Zellen mit 50.000, 150.000 oder 500.000 Alveolarmakrophagen in
einem Volumen von 500 µl inkubiert. Als Vergleichskulturen wurden parallel je 500.000
T-Zellen bzw. Alveolarmakrophagen allein in 500 µl kultiviert. Die Ergebnisse sind in Abbil-
dung 3.24 zusammengefasst. Auch in diesen Kokulturen zeigte sich eine starke Zunahme der
IL-10 Proteinkonzentration in den Überständen der Mischkulturen im Vergleich zu den reinen
CD3+ T-Zellkulturen und den reinen Alveolarmakrophagenkulturen. Mit zunehmendem
Anteil an Alveolarmakrophagen (AM) steigt die IL-10 Konzentration in den RSV-stimulier-
ten Mischkulturen von 3,5 [1,9 - 4,9] ng/ml bei 50.000 AM auf 5,1 [2,8 – 6,9] ng/ml bei
150.000 AM und 6,4 [3,6 - 10,5] ng/ml bei 500.000 AM, während 500.000 AM in Reinkultur
lediglich 0,1 [0 - 0,1] ng/ml IL-10 im Überstand bei RSV-Stimulierung aufwiesen. Daraus
lässt sich ableiten, dass das Zusammentreffen beider Zelltypen zu einem synergistischen
Effekt bei der IL-10 Freisetzung führte, da die Zunahme in den verschiedenen Mischkulturen
den reinen Anteil der Alveolarmakrophagen bei weitem übertraf.
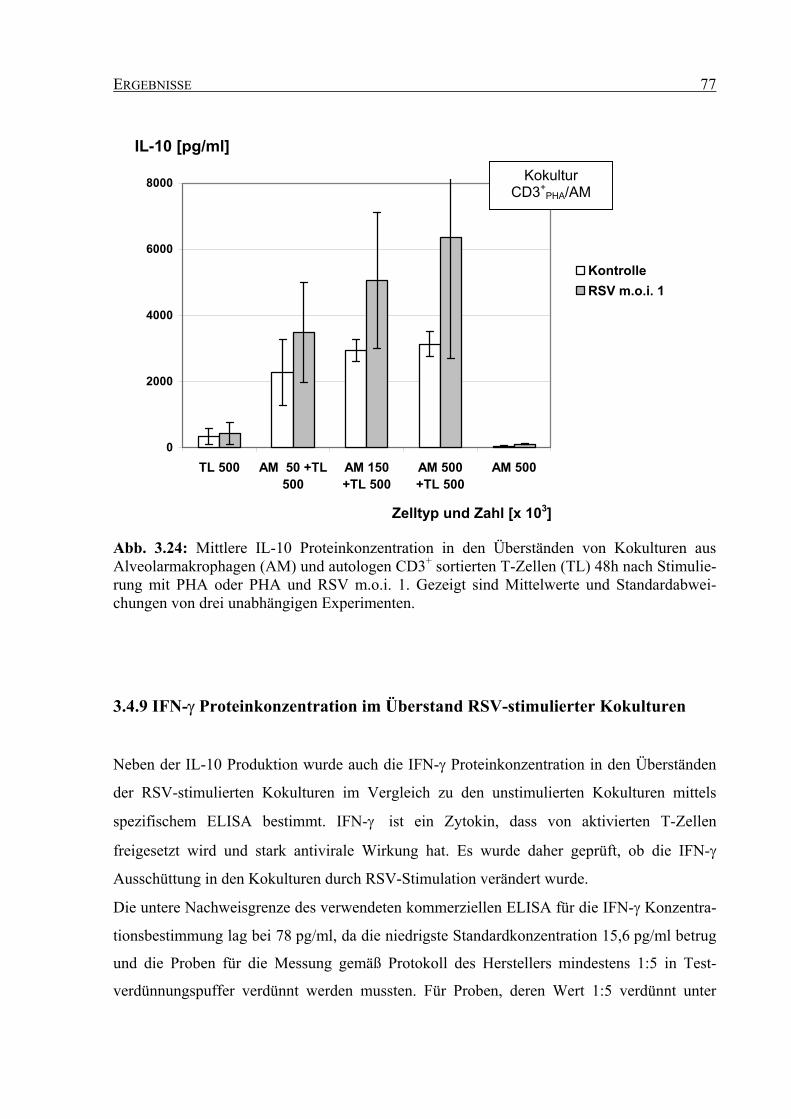
ERGEBNISSE 77
0
2000
4000
6000
8000
TL 500 AM 50 +TL500
AM 150+TL 500
AM 500+TL 500
AM 500
Kontrolle
IL-10 [pg/ml] Kokultur
CD3+PHA/AM
RSV m.o.i. 1
Zelltyp und Zahl [x 103]
Abb. 3.24: Mittlere IL-10 Proteinkonzentration in den Überständen von Kokulturen aus Alveolarmakrophagen (AM) und autologen CD3+ sortierten T-Zellen (TL) 48h nach Stimulie-rung mit PHA oder PHA und RSV m.o.i. 1. Gezeigt sind Mittelwerte und Standardabwei-chungen von drei unabhängigen Experimenten.
3.4.9 IFN-γ Proteinkonzentration im Überstand RSV-stimulierter Kokulturen
Neben der IL-10 Produktion wurde auch die IFN-γ Proteinkonzentration in den Überständen
der RSV-stimulierten Kokulturen im Vergleich zu den unstimulierten Kokulturen mittels
spezifischem ELISA bestimmt. IFN-γ ist ein Zytokin, dass von aktivierten T-Zellen
freigesetzt wird und stark antivirale Wirkung hat. Es wurde daher geprüft, ob die IFN-γ
Ausschüttung in den Kokulturen durch RSV-Stimulation verändert wurde.
Die untere Nachweisgrenze des verwendeten kommerziellen ELISA für die IFN-γ Konzentra-
tionsbestimmung lag bei 78 pg/ml, da die niedrigste Standardkonzentration 15,6 pg/ml betrug
und die Proben für die Messung gemäß Protokoll des Herstellers mindestens 1:5 in Test-
verdünnungspuffer verdünnt werden mussten. Für Proben, deren Wert 1:5 verdünnt unter

ERGEBNISSE 78
15,6 pg/ml lag, wurde bei der weiteren Auswertung die Nachweisgrenze von 78 pg/ml
angesetzt.
Von den Kokulturen aus CD3+14- T-Zellen und CD14+ Monozyten (s. 3.4.7) wurden bei
den Proben mit den Zellzahlverhältnissen 10:1, 2:1 und 1:3 (Verhältnis CD3+14- : CD14+)
sowie bei den reinen T-Zell- und Monozytenkulturen die IFN-γ Konzentrationen im Über-
stand der unstimulierten und der mit RSV m.o.i. 3 stimulierten Proben bei jeweils vier
Spendern bestimmt. Es zeigte sich, dass nur in den Proben mit einem Zellzahlverhältnis von
10:1 oder 2:1 die IFN-γ Konzentrationen oberhalb der Nachweisgrenze lagen. Dabei war der
Effekt der RSV-Stimulation weniger einheitlich als beim IL-10. Bei 5 von 8 Proben nahm die
IFN-γ Konzentration unter RSV-Stimulation zu (MdRPMI 87 [78 – 263] pg/ml; MdRSV 892
[279 - 3022] pg/ml), bei den übrigen 3 Proben ab (MdRPMI 340 [118 – 416] pg/ml; MdRSV 122
[78 - 310] pg/ml).
Bei den Kokulturen aus Alveolarmakrophagen und PHA-stimulierten CD3+ T-Zellen
erfolgte die Messung der IFN-γ Konzentration in Überständen von unstimulierten und RSV
m.o.i. 1-stimulierten Ansätzen mit einem Zellzahlverhältnis von 1:1 sowie in den zugehörigen
reinen T-Zell- und Makrophagenkulturen bei sieben Spendern. In den reinen Makrophagen-
kulturen konnte kein IFN-γ in den Überständen nachgewiesen werden. In den reinen
T-Zellkulturen lag die IFN-γ Konzentration aufgrund der in diesem Kokulturmodell vorge-
nommenen PHA-Stimulierung weit über der Nachweisgrenze. Sie betrug in den Kontroll-
Kulturen ohne RSV 6,3 ± 2,6 ng/ml und sank nach RSV-Stimulierung im Mittel auf
4,8 ± 3,3 ng/ml bei einer Probe mit gegenläufiger Tendenz. In den Kokulturansätzen sank sie
im Mittel von 32,1 ± 22,6 ng/ml in den Kulturen ohne Virus auf 23,2 ± 20,6 ng/ml in den
RSV-stimulierten Ansätzen, wobei der Trend bei zwei Spendern gegenläufig war.
Zusammengefasst zeigte sich in den beiden Kokulturmodellen in Bezug auf die IFN-γ
Konzentration mit und ohne RSV-Stimulierung eine tendenzielle IFN-γ Abnahme durch die
Virus-Stimulation. Dabei reagierten aber 4 von insgesamt 11 Spendern mit einer gesteigerten
IFN-γ Freisetzung auf die RSV-Stimulation in vitro.

ERGEBNISSE 79
3.4.10 Verhältnis von IL-10 und IFN-γ Produktion in RSV-stimulierten
Kokulturen
Zur Klärung, inwieweit die RSV-Infektion zu einer Veränderung des Zytokinverhältnisses
zwischen IL-10 und IFN-γ führte, wurde aus den jeweiligen Einzelwerten für IL-10 und IFN-γ
ein Quotient IL-10/IFN-γ berechnet und mit und ohne RSV-Stimulierung jeweils verglichen.
Eine Erhöhung des Quotienten ergibt sich zum einen, wenn die IL-10 Konzentration durch die
RSV-Stimulierung steigt und die IFN-γ Konzentration sinkt, wie es in mehreren Ansätzen der
Fall war. Der Quotient steigt aber auch dann, wenn beide Konzentrationen durch die RSV-
Stimulation steigen oder sinken, dabei aber IL-10 stärker ansteigt als IFN-γ oder IFN-γ stärker
abnimmt als IL-10.
Interessanterweise stieg der Quotient IL-10/IFN-γ trotz des nicht einheitlichen Trends beim
IFN-γ und den interindividuellen Schwankungen in der absoluten Höhe der Zytokinproduk-
tion in allen PHA-stimulierten CD3+ T-Zellkulturen (N = 5, p = 0,068) und in allen Kokultu-
ren aus Alveolarmakrophagen und PHA-stimulierten CD3+ T-Zellen (N = 7, p = 0,018) durch
die RSV-Stimulation an. Bei den Kokulturansätzen aus CD3+14- T-Zellen und CD14+ Mono-
zyten, bei denen mindestens einer der beiden Zytokinwerte oberhalb der Nachweisgrenze lag,
stieg der Quotient in 12 von 15 Ansätzen an (p < 0,01).
Das bedeutet, dass bei mehreren Ansätzen, in denen die IL-10 und die IFN-γ Konzentration
durch die RSV-Stimulation gestiegen waren, die Zunahme beim IL-10 verhältnismäßig größer
war als die Zunahme beim IFN-γ.
Die Veränderungen der Quotienten durch die RSV-Stimulierung sind in Abbildung 3.25
zusammengefasst.

ERGEBNISSE 80
Quotient IL-10/IFN-γ
0
0,1
0,2
0,3
0,4
0,5
0,6
ohne RSV mit RSV
0
0,1
0,2
0,3
0,4
0,5
0,6
ohne RSV mit RSV
A B C
CD3+PHA
0,1
1
10
100
ohne RSV mit RS
C CD
Abb. 3.25: Quotient IL-10/IFN-γA) PHA-stimulierten CD3+ T-ZelB) Kokulturen aus Alveolarmak
verhältnis 1:1, RSV-StimulatiC) Kokulturen aus CD3+14- T-Ze
Abweichend von A) und B) sGezeigt sind die Einzelwerte sow
In den Kokulturen aus Alveolarm
bei allen Spendern auf und w
Kokultur 3+14-/CD14+
V
in RSV-stimulierten Kulturen und Kontrollen bei lkulturen, RSV-Stimulation mit m.o.i. 1 rophagen und PHA-stimulierten CD3+ T-Zellen, on mit m.o.i. 1 llen und CD14+ Monozyten, RSV-Stimulation mit
ind die Werte auf einer logarithmischen Skala dargeie die Mittelwerte (▬) und Mediane (▬).
akrophagen und T-Zellen trat diese Verschiebung e
ar mit einer mittleren Zunahme um den Faktor
Kokultur D3+
PHA/AM
Zellzahl-
m.o.i. 3. stellt.
inheitlich
3,3 ± 1,9

ERGEBNISSE 81
deutlicher ausgeprägt als in den reinen T-Zellkulturen (Faktor Zunahme 2,0 ± 0,6) oder den
Kokulturen mit CD14+ Monozyten (Faktor Zunahme 2,6 ± 1,4).
Neben einer signifikanten Zunahme der IL-10 mRNA-Expression in aufgereinigten T-Zellen
und einer synergisitischen Steigerung der IL-10 Produktion in Kokulturen aus T-Zellen und
Monozyten/Makrophagen bewirkte RSV in vitro auch eine signifikante Erhöhung des
Quotienten aus IL-10 und IFN-γ insbesondere in den Kokulturen aus Alveolarmakrophagen
und T-Zellen.

82
4. Diskussion
4.1 Gewinnung hochreiner T-Zellen aus peripherem Blut
In der vorliegenden Arbeit wurden aufgereinigte T-Zellen nach einer Stimulierung mit RSV
in vitro mittels Durchflusszytometrie und CLS-Mikroskopie phänotypisch charakterisiert.
Darüber hinaus sollte eine mögliche funktionelle Bedeutung der in vitro beobachteten
Interaktion zwischen RSV und T-Zellen untersucht werden. Für die Analysen der RSV-
stimulierten T-Zellen im Durchflusszytometer und Immunfluoreszenzmikroskop, die einen
gleichzeitigen Nachweis des Zelltyps ermöglichten, wurden T-Zellen in einer einstufigen
magnetischen Zellsortierung aus PBMC durch Bindung von antiCD3-Antikörpern CD3-
positiv sortiert. Der Nachweis einer funktionellen Bedeutung dieser Virus-Zell-Interaktion
erforderte eine besonders hohe Reinheit der T-Zellpopulation, um bei den angestrebten
Zytokinmessungen andere Zellen als Produzenten sicher ausschließen zu können.
Insbesondere beim Interleukin-10, das aufgrund seiner immunsuppressiven Wirkung in
Zusammenhang mit der möglichen Bedeutung einer RSV-Infektion bei der COPD von
Interesse war, sind Monozyten ebenfalls als Produzenten beschrieben (de Waal Malefyt
1991). Daher wurde das Sortierschema für die funktionellen Analysen auf eine zweistufige
magnetische Zellsortierung mit vorgeschalteter CD14-Depletion erweitert. Der Anteil
verunreinigender CD14+-Monozyten konnte durch diesen zusätzlichen Sortierschritt von
durchschnittlich 3 % in den CD3+ Fraktionen auf 0,2 % in den CD3+14- sortierten Fraktionen
gesenkt werden. In Verbindung mit einer ermittelten Zellausbeute von über 20 % der
eingesetzten PBMC erwies sich diese zweistufige magnetische Zellsortierung als eine
effektive Methode zur Gewinnung sehr reiner T-Zellen aus PBMC. Darüber hinaus
ermöglichte sie gleichzeitig die Gewinnung einer Fraktion CD14+ Monozyten mit ebenfalls
hoher Reinheit, die für Vergleichsexperimente und Kokulturansätze aus T-Zellen und
Monozyten verwendet werden konnte.
4.2 Phänotypische Charakterisierung RSV-stimulierter T-Zellen
Die Ergebnisse der phänotypischen Charakterisierung aufgereinigter T-Zellen nach einer
Stimulierung mit RSV in vitro mittels Durchflusszytometrie und CLS-Mikroskopie zeigten

DISKUSSION 83
eine Bindung von RSV-Partikeln an die Oberfläche CD4+ und CD8+ T-Zellen und belegen
das intrazelluläre Vorkommen von RSV in CD3+ sortierten T-Zellen.
In bisherigen Arbeiten zu dieser Frage (Thurau 1998, Diaz 1999) wurden RSV-Antigene
intrazellulär mittels indirekter Immunfluoreszenz im Durchflusszytometer nach Permeabilisie-
rung der Zellen nachgewiesen. Diese Behandlung ermöglicht ein Eindringen der Fluoreszenz-
gekoppelten Antikörper in das Zytoplasma der Zelle. Die Antikörper können aber auch an die
Zelloberfläche gebundenes Antigen erkennen. Im Gegensatz zu Antigenen, die nur intrazel-
lulär exprimiert werden wie z.B. Zytoskelettbestandteile, kann bei einem Virusnachweis mit
dieser Methode nicht ausgeschlossen werden, dass es sich um außen an der Zelle anhaftende
Viruspartikel handelt, da sich die Signale nicht unterscheiden. In der Arbeit von Diaz wurde
zudem bei der Analyse der Infektionsrate in CD4+ oder CD8+ isolierten Kulturen keine
Markierung mit T-zellspezifischen Antikörpern durchgeführt. Daher könnten die 6 % bzw.
5 % RSV-positiven Zellen bei einer Reinheit der Kulturen von 95 % auch beinahe vollständig
anderen Subpopulationen angehören, zumal für B-Zellen und Monozyten deutlich höhere
Infektionsraten in jener bzw. anderen Arbeiten (Midulla 1989) nachgewiesen worden waren.
In der vorliegenden Arbeit wurde bei den Analysen im Durchflusszytometer ausschließlich
mit extrazellulärer Markierung gearbeitet bei gleichzeitiger Markierung mit T-Zell-
spezifischen Antikörpern (antiCD3, antiCD4 oder antiCD8).
Die ersten Beobachtungen zur Bindung von RSV an T-Zellen, die den Anlass zu der hier
vorliegenden Arbeit gaben, stammten aus eigenen Untersuchungen an PHA-stimulierten
T-Zellen. Bei hoher Infektionsdosis (m.o.i. = 6) und gleichzeitiger PHA-Stimulierung belief
sich der Anteil RSV-positiver T-Zellen auf bis zu 23 %. Daher stellte sich die Frage, ob es
sich möglicherweise um eine indirekte Bindung über das Pflanzenlektin handeln könnte. In
den hier beschriebenen Experimenten ließ sich jedoch zeigen, dass Inkubation von T-Zellen
mit RSV auch ohne gleichzeitige PHA-Stimulierung zu einem signifikanten Anstieg des
Anteils RSV-positiver Zellen im Vergleich zu den Kontrollen ohne Virus führte. Der Anteil
RSV-positiver T-Zellen stieg mit zunehmender Virusdosis sowohl in den Kulturen ohne
weiteren Stimulus als auch in den Ansätzen mit antiCD28- oder PHA-Zusatz an. Bei PHA-
Stimulierung nahm der Anteil RSV-positiver Zellen im Bereich niedriger m.o.i. zunächst
stark, bei höherer m.o.i. nur noch wenig zu. Der sich daraus ergebende Kurvenverlauf in
Abbildung 3.4 entspricht einer Sättigungskurve. Bei gleicher m.o.i. war der Anteil RSV-
positiver T-Zellen in den PHA-stimulierten Ansätzen signifikant höher als in den Kulturen

DISKUSSION 84
mit antiCD28-Zusatz und in diesen höher als ohne weiteren Stimulus. Zusammen legen diese
Beobachtungen nahe, dass es sich bei der Bindung von RSV an T-Zellen um eine spezifische
Bindung an eine Oberflächenstruktur handelt, die je nach Aktivierungszustand der T-Zellen
unterschiedlich stark exprimiert wird.
Die zusätzlich zu den Analysen im Durchflusszytometer hier durchgeführten CLSM-
Untersuchungen ermöglichten eine schichtweise Durchleuchtung der Proben. Durch
Vergleich der Bilder aus verschiedenen Ebenen konnte die Lage der angefärbten Strukturen
abgeleitet werden. Durch Etablierung einer Immunfluoreszenzmarkierung, die neben dem
RSV-Nachweis in derselben Probe eine Markierung des T-zellspezifschen CD3-Oberflächen-
antigens in der CLS-Mikroskopie zuließ, war die Identifizierung des Zelltyps auch hier
jederzeit gewährleistet. In CD3+ sortierten T-Zellen konnte 20h nach Stimulation mit PHA
und RSV m.o.i. 1 auf diese Weise RSV intrazellulär nachgewiesen werden.
Darüber hinaus zeigten diese Aufnahmen, dass die Bindung der RSV-Partikel an die
Oberfläche der T-Zellen räumlich getrennt vom TZR-Komplex erfolgte, da die RSV-Signale
nicht mit den CD3-Signalen kolokalisiert waren. Der Anteil von bis zu 7 % RSV-positiven
T-Zellen ohne weitere Stimulierung legt nahe, dass RSV auch an T-Zellen bindet, die nicht
RSV-spezifisch sind. Im Mausmodell ist gezeigt worden, dass der Anteil antigen-spezifischer
T-Zellen während einer Virusinfektion unter 1 % lag (Tripp 1995), also weit niedriger ist als
der Anteil RSV-positiver Zellen in den hier untersuchten T-Zellkulturen.
Im Rahmen der phänotypischen Analyse der T-Zellen wurde auch die Expression von CD69
durch Markierung mit spezifischen monoklonalen Antikörpern im Durchflusszytometer
ermittelt. CD69, früher auch als EA1, MLR3 oder Leu-23 bezeichnet, ist als ein früher
Aktivierungsmarker auf T-Zellen, B-Zellen und NK-Zellen beschrieben (Hara 1986; Risso
1989; Lanier 1988). Frisch isolierte CD3+14- T-Zellen exprimierten erwartungsgemäß kein
CD69. Stimulierung mit RSV m.o.i. 1 oder m.o.i. 3 führte wie die unstimulierte Kultur in
Medium nach 24h zu einer geringen Expression auf rund 2 % der Zellen. Zugabe von
antiCD28-Antikörpern führte zu einer gesteigerten Expression mit durchschnittlich 4 % und
maximal bis zu 8 % CD69-positiven Zellen. Diese Rate wurde bei Kombination von
antiCD28- und RSV-Stimulierung nicht weiter gesteigert. Diese Ergebnisse zeigen, dass die
positive Isolierung der T-Zellen allein nicht zu einer Aktivierung führte, während Zugabe von
antiCD28-Antikörpern eine Aktivierung der T-Zellen bewirkte.

DISKUSSION 85
Ein Rezeptor für RSV auf den Zellen, für die eine RSV-Infektion bereits nachgewiesen
wurde, konnte bisher nicht eindeutig identifiziert werden (Teng 2002). ICAM-1 wurde als
einer von vielen möglichen Rezeptoren für RSV auf den T-Zellen untersucht. ICAM-1 ist
zusammen mit seinem Liganden LFA-1 an der Bindung von T-Zellen an antigenpräsentie-
rende Zellen und der Wanderung von T-Zellen beteiligt (Dustin 1986). Darüber hinaus ist
ICAM-1 als Rezeptor für humanes Rhinovirus (HRV) beschrieben (Greve 1989, Staunton
1989). Auf humanen epithelialen Zellen (Zelllinie A549) war die Expression von ICAM-1
nach RSV-Infektion in vitro signifikant erhöht (Arnold 1996; Wang 2000). Zur Überprüfung
einer möglichen Bindung von RSV an ICAM-1 wurden die T-Zellen vor und während der
Stimulierung mit RSV mit einem antiICAM-1 Antikörper inkubiert. Der Anteil RSV-positiver
T-Zellen wurde davon nicht beeinflusst. Es konnte jedoch nicht geklärt werden, ob es sich bei
dem verwendeten antiICAM-1 Antikörper um einen blockierenden Antikörper handelte. Eine
Bindung von RSV an ICAM-1 kann daher aufgrund dieser Experimente nicht sicher
ausgeschlossen werden. Zusammen mit der nur minimalen Expressionsrate von < 1 % auch
nach Stimulierung mit RSV und PHA scheint es allerdings eher unwahrscheinlich, dass RSV
an ICAM-1 auf T-Zellen bindet.
Nachdem durch die CLSM-Analysen das intrazelluläre Vorkommen von RSV in T-Zellen
nachgewiesen worden war, stellte sich die Frage, ob RSV sich in T-Zellen auch repliziert.
Diese Frage wurde mittels einer quantitativen real-time PCR für das RSV-Fusionsprotein
untersucht, die kürzlich in unserer Arbeitsgruppe etabliert worden ist (Borg 2002). Mit diesem
PCR-System wurde ein 76 bp großes Fragment aus dem RSV-Genom im Zeitverlauf von 2h
bis 48h nach RSV-Stimulation in CD3+14- sortierten T-Zellen quantifiziert. Bei der Nuklein-
säure-Isolierung aus den Zellkulturen war ein Ansatz für die Gewinnung von GesamtRNA
gewählt worden, der auch virale RNA umfasste. Die anschließende Reverse Transkription
wurde mit random Hexamer-Primern durchgeführt. Auf diesem Wege hergestellte cDNA
konnte sowohl für die RSV-PCR als auch für die IL-10 PCR als Ausgangsmaterial eingesetzt
werden.
Die Messung der Menge an viraler RNA in den T-Zellkulturen im Zeitverlauf ergab ∆Ct-
Werte von +0,1 bis +1,0 für die 24h- und 48h-Ansätze im Vergleich zu den 2h-Ansätzen.
Dazu ist anzumerken, dass zwei Werte nicht sicher als verschieden angesehen werden
können, sofern die Messunterschiede kleiner als die Intraassayvarianz sind, die für dieses
PCR-System mit einem ∆Ct von 0,56 für den oberen Messbereich angegeben ist (Borg 2002).
Bei den T-Zellkulturen zeigte sich also keine wesentliche Veränderung der Menge an RSV-

DISKUSSION 86
RNA im Verlauf von 48h. Durch Einbeziehen von 48h-Werten bei den T-Zellkulturen war
gewährleistet, dass die Replikation auch dann detektiert werden konnte, wenn sie in T-Zellen
eventuell langsamer als in gut infizierbaren Zelltypen ablaufen sollte. Der Vergleich mit der
Entwicklung der RSV-RNA nach Infektion einer permissiven humanen Epithelzelllinie
(Hep-2 Zellen) ließ darauf schließen, dass sich das Virus in den T-Zellen nicht produktiv
replizierte.
Bei der Interpretation dieser Ergebnisse ist zu berücksichtigen, dass durch die Verwendung
von random hexamer Primern bei der cDNA-Herstellung mit diesem PCR-System nicht
zwischen zugefügtem und neu synthetisiertem Virusmaterial unterschieden werden konnte.
Die Veränderung der in den Kulturen gemessenen Menge an viraler RNA im Zeitverlauf
entspricht dem Saldo von Abbau und eventueller Neusynthese. Eine geringe Replikation von
RSV in den T-Zellen kann daher nicht ausgeschlossen werden. Der Vergleich mit den Daten
aus der Infektion von HEp-2 Zellen zeigt allerdings deutlich, welche Zunahme an RSV-RNA
in permissiven Zellen mit einer produktiven Infektion einhergeht.
Das Vorhandensein von viraler Nukleinsäure in Zellen ohne oder mit nur geringer Replikation
ist kennzeichnend für eine latente Infektion in Verbindung mit viraler Persistenz (de la Torre
1991). Für RSV wird die Möglichkeit einer Persistenz in humanen Zellen diskutiert.
O’Donnell (1998) wies mittels PCR bei Kindern mit akuter RSV-Bronchiolitis RSV-RNA
auch in Blutzellen nach. RSV verbreitet sich also auch über die Atemwege hinaus im
menschlichen Körper. Die molekularen Mechanismen, die zur Persistenz führen, sind bei
einem RNA-Virus schwerer vorstellbar, als bei DNA-Viren, bei denen sich Teile des viralen
Genoms in das zelluläre Genom integrieren können. Für Adenoviren wurde dies bereits nach-
gewiesen und eine Verbindung zu der Pathogenese des steroid-resistenten kindlichen Asthmas
aufgezeigt (Macek 1994). Für das Masernvirus, das wie RSV zur Gruppe der Paramyxoviren
gehört, ist eine persistierende Infektion von T- und B-Zellen bekannt. Sie führt zu einer gene-
ralisierten Immunsuppression über im Detail noch unbekannte Mechanismen (Oldstone
1989).
Das in dieser Arbeit beschriebene intrazelluläre Vorkommen von RSV in T-Zellen ohne
produktive Replikation in vitro weist darauf hin, dass das RS-Virus T-Zellen als Träger einer
latenten oder persistierenden Infektion nutzen kann. Dies wäre in weiterführenden
Experimenten, z.B. durch den Versuch eines Nachweises von RSV-RNA ex vivo in T-Zellen
aus dem peripheren Blut gesunder Spender, systematisch zu untersuchen. Aufgrund der
wahrscheinlich niedrigen Kopienzahl müsste dabei mit größeren Mengen an T-Zellen

DISKUSSION 87
gearbeitet werden, als sie hier in den unstimulierten Ansätzen zur Verfügung standen. Außer
dem Nachweis des intrazellulären Vorkommens von RSV in T-Zellen 20h nach RSV-
Exposition in vitro stehen weitere Beweise für eine Infektion der T-Zellen durch RSV noch
aus. Bei den Experimenten zur Klärung der funktionellen Bedeutung der Interaktion von RSV
und T-Zellen wurde daher der nicht die Bezeichnung RSV-infizierte, sondern der Begriff
RSV-stimulierte T-Zellen verwendet.
4.3 Nachweis einer funktionellen Bedeutung der Interaktion von RSV und
T-Zellen in vitro
Außer über direkten Zell-Zell-Kontakt üben die Zellen des Immunsystems ihre Funktion über
die Freisetzung von Zytokinen und die Schaffung eines sie umgebenden Zytokinmilieus aus.
Der Eingriff in die Zytokinproduktion einer Zelle ist daher eine Möglichkeit für ein Virus, die
Immunantwort des Wirtsorganismus zu seinen Gunsten zu beeinflussen. Gleichzeitig
vermitteln Zytokine die Aktivierung der Abwehrmechanismen gegen Pathogene. Zur
Überprüfung der funktionellen Bedeutung der beobachteten Interaktion von RSV und
T-Zellen wurde daher die Zytokinproduktion der Zellen mit und ohne Virusstimulation
verglichen.
Neben sogenannten proinflammatorischen Zytokinen wie IL-6, IL-8, TNF-α und antiviralen
Stoffen wie Interferon-α (IFN-α) stellt IL-10 ein Schlüsselmolekül dar, da durch seine
Induktion die Viren der Erkennung und Eliminierung durch das Immunsystem des Wirts zu
entgehen versuchen. Diese Schlüsselrolle beruht auf der breiten immunsuppressiven Wirkung
von IL-10 (s. Einleitung) und wird durch die Codierung viraler IL-10 Homologe in den
Genomen verschiedener Viren eindrucksvoll belegt. IL-10 Homologe mit immunsuppressiven
Funktionen wurden unter anderem für das Epstein-Barr-Virus (EBV) (Hsu 1990, de Waal
Malefyt 1991a) und das Cytomegalievirus (CMV) nachgewiesen (Kotenko 2000, Spencer
2002). Für Rhinovirus wurde gezeigt, dass es in vitro IL-10 in Monozyten induziert und zu
einer verminderten Expression von MHC-II Molekülen auf diesen Zellen führt (Stöckl 1999).
Eine Methode, die einen Zytokinnachweis mit der Möglichkeit der Zelltyp-Charakterisierung
verbindet, ist der intrazelluläre Zytokinnachweis im Durchflusszytometer (Jung 1993). Diese
Methode erfordert aber meist eine starke Stimulation der Zellen mit künstlichen Stimuli wie

DISKUSSION 88
Phorbol-12-Myristat-13-Azetat (PMA) in Verbindung mit Ionomycin, einem Kalzium-
ionophor (Thurau 1998). Da nachgewiesen wurde, dass die Art der Stimulation erheblichen
Einfluss auf die Zytokinproduktion einer Zelle hat (Yssel 1992), ist der Nachweis einer
funktionellen Bedeutung der RSV-Bindung in der vorliegenden Arbeit ohne zusätzliche
Stimulanzien erfolgt. Dies erforderte eine sehr sensitive Messmethode, wie sie die
quantitative PCR darstellt (Bustin 2000). Daher wurde für die Messung von IL-10 in CD3+14-
aufgereinigten RSV-stimulierten T-Zellen eine quantitative real-time PCR für humanes
IL-10 etabliert.
Möglichen Fallstricken der Methode (Übersicht in Freeman 1999) wurde durch entsprechende
Verfahrensweisen begegnet. So wurden die kritischen Schritte der RNA-Isolation und
RT-Reaktion (s. 2.2.7 und 2.2.8) für alle Proben mit den gleichen kommerziellen Kits und
von einer Person durchgeführt. Geringe Schwankungen in der Zahl der Zellen pro Kultur-
ansatz sowohl innerhalb eines Experimentes als auch von einer Zellisolierung zur nächsten
wurden durch die Quantifizierung relativ zu einem Haushaltsgen ausgeglichen (s. 2.2.9.4).
Dabei war zu berücksichtigen, dass die Expression des Haushaltsgens unter den gewählten
experimentellen Bedingungen einer Virus-Infektion in vitro konstant sein muss. Statt der
häufig verwendeten Haushaltsgene β-Aktin oder GAPDH, die prozessierte Pseudogene
besitzen, die das PCR-Ergebnis erheblich beeinflussen können (Jung 2000) wurde daher in
diesem Fall ß-Glucuronidase (GUS) als Haushaltsgen eingesetzt. Dieses Gen hatte sich in
einer Analyse in unserer Arbeitsgruppe bei virusinfizierten PBMC als konstantestes von 11
untersuchten Haushaltsgenen erwiesen (Löseke 2002).
Eine Verfälschung der Ergebnisse durch die Amplifikation von genomischer DNA wurde
dadurch verhindert, dass das PCR-Produkt auf genomischer Ebene in zwei benachbarten
Exons liegt, die durch 865 bp intronische Sequenz voneinander getrennt sind.
Neben diesen grundlegenden methodischen Überlegungen ist nach der Einrichtung eines
PCR-Systems für ein bestimmtes Gen eine sorgfältige Validierung für dies bestimmte Labor
wesentlich (Freeman 1999). Daher wurden Sensitivität, Spezifität und Reproduzierbarkeit als
wesentliche Qualitätsmerkmale einer Methode zur mRNA-Quantifizierung (Bustin 2000) für
die in dieser Arbeit beschriebene quantitative real-time PCR für humanes IL-10 eingehend
untersucht. Mit Variationskoeffizienten von 1,3 % und 4,8 % für die Intra- bzw. Interassay-
varianz und einer Nachweisgrenze von unter 50 mRNA-Molekülen erwies sich das System als
sehr sensitiv, zuverlässig reproduzierbar und mit anderen real-time PCR-Systemen vergleich-
bar (Bustin 2000). Die Spezifität wurde mittels Schmelzkurven-Analyse, Gelelektrophorese

DISKUSSION 89
und Sequenzierung der PCR-Produkte überprüft. Dabei fanden sich keine Hinweise auf
unspezifische Amplifikation.
Nach abgeschlossener Validierung konnte die IL-10 mRNA-Expression in CD3+14-
T-Zellkulturen mit diesem PCR-System analysiert werden.
Es konnte bereits nachgewiesen werden, dass IL-10 von verschiedenen T-Zellpopulationen
produziert wird. Bei Mäusen wurden IL-10 exprimierende T-Zellen stets als TH2-Zellen
gemäß ihres sonstigen Zytokinprofils identifiziert. Im Gegensatz dazu fand sich eine IL-10
Produktion bei humanen TH1-Zellen, TH2-Zellen und auch CD8+ zytotoxische T-Zellen
(Yssel 1992, del Prete 1993). Kürzlich wurde eine CD4+ Subpopulation beschrieben, die
zusammen mit TGF-β große Mengen an IL-10 und mäßige Mengen an IFN-γ und IL-5 frei-
setzt. Aufgrund ihrer veränderten Funktion nannte man diese T-Zellen Tr1-Zellen (Groux
1997, Jonuleit 2000, Levings 2000). In Verbindung mit einer RSV-Infektion in vitro war eine
IL-10 Produktion bisher nur für Alveolarmakrophagen (Panuska 1995) und unseparierte
PBMC (König 1996, Diaz 1999) gezeigt worden.
In frisch isolierten CD3+14- T-Zellen konnte eine geringe IL-10 mRNA Expression von etwa
5 Kopien pro 1000 Zellen nachgewiesen werden. Dies könnte Ausdruck einer konstitutiven
Expression oder einer in vivo stattgefundenen Aktivierung der Zellen sein. Die relativ
geringen Schwankungen der Expressionsrate zwischen den verschiedenen Spendern (∆Ct
37,3 ± 1,2) deuten eher auf eine konstitutive Expression hin. Yssel (1992) untersuchte die
IL-10 mRNA Expression in humanen CD4+ T-Zellen mittels Northern-Blot-Analyse und fand
keine konstitutive Expression, sondern eine Induktion von IL-10 8h nach Stimulierung mit
antiCD3-Antikörpern und Tetradecanoyl-Phorbolazetat (TPA). Diese Unterschiede können
zum einen daraus resultieren, dass die Sensitivität einer PCR wesentlich höher ist als die des
Northern-Blot-Verfahrens. Zum anderen handelte es sich aber auch um unterschiedliche
T-Zellpräparationen, da in der zitierten Arbeit die Northern-Blot-Analysen mit expandierten
T-Zellklonen durchgeführt wurden, und nur bei ELISA-Messungen vergleichende Experi-
mente mit aufgereinigten T-Zellpopulationen aus dem peripheren Blut erfolgten.
Außer dieser Abweichung in Bezug auf eine schwache IL-10 mRNA Expression in den
unkultivierten Zellen entsprach die in der vorliegenden Arbeit gezeigte Kinetik der IL-10
mRNA Expression dem in der Literatur beschriebenen Verlauf (Barth 2000, Yssel 1992). In
den unstimulierten Kulturen fiel die IL-10 mRNA Expression nach einem kurzen Anstieg

DISKUSSION 90
wieder ab. Dieser Verlauf steht in Einklang mit der Beobachtung von Barth (2000), wonach
bei PBMC die Kultur in Medium zu einer kurzfristig erhöhten Expression bei verschiedenen
anderen Zytokinen (IL-10 wurde nicht untersucht) führte. Dieser Effekt wurde auf eine
vorübergehende Serum-induzierte Stimulation der Transkription zurückgeführt und war nach
24h vollständig abgeklungen. In den RSV-stimulierten Kulturen stieg die IL-10 Expression
über die anfängliche Zunahme hinaus weiter an und erreichte spenderabhängig nach 18h bis
24h ihr Maximum. Auch in der oben bereits erwähnten Untersuchung von Yssel lag das
Maximum der IL-10 Expression bei 24h. Für die weiteren Experimente zur IL-10 Expression
wurde daher eine Kulturdauer von 24h gewählt.
Die Auswertung ergab eine signifikante Zunahme der IL-10 mRNA-Expression in
CD3+14- T-Zellkulturen 24h nach Stimulierung mit RSV m.o.i. 1 oder m.o.i. 3. Dabei
reagierten die Zellen der verschiedenen Spender nahezu einheitlich in der Tendenz, aber
unterschiedlich stark auf die Stimulation mit RSV. Nur die Zellen eines Spenders (von 12
Spendern) reagierten nicht mit einer Zunahme der IL-10 Expression. Die Expression stieg
durchschnittlich auf das 3,5 fache bei m.o.i. 1 und das 4,5 fache bei m.o.i. 3. Der Anstieg der
IL-10 mRNA Expression war signifikant mit dem Anteil RSV-positiver Zellen korreliert, der
durch extrazelluläre Markierung durchflusszytometrisch bestimmt worden war. Zusammen
mit der strengen Kontrolle identischer Inkubationsbedingungen für die RSV-Ansätze und
Kontrollen unterstreicht dies, dass es wirklich die RSV-Partikel und nicht andere Faktoren
waren, die zur Zunahme der IL-10 Expression führten.
Das intrazelluläre Vorkommen von RSV in den T-Zellen war aus methodischen Gründen
nicht exakt quantifizierbar. Daher ist aus den Ergebnissen nicht ableitbar, ob allein die
Bindung der RSV-Partikel an die Zelloberfläche oder erst das Eindringen des Virus in die
Zelle die Signale für die erhöhte mRNA Expression auslösten. Zur Beantwortung dieser Frage
wurden Experimente mit UV-inaktiviertem RSV-Material durchgeführt (Daten im
Ergebnisteil nicht gezeigt). Diese erwiesen sich als nicht auswertbar, da das UV-inaktivierte
Virus starke zytopathische Effekte in den Zellkulturen verursachte. In HEp-2 Zellen führte es
zu einem höheren Anteil an abgestorbenen Zellen und mikroskopisch veränderter
Morphologie in den noch lebenden Zellen. Auch in PBMC-Kulturen bewirkte die Inkubation
mit inaktiviertem RSV-Material eine deutlich verminderte Zellvitalität. 48h nach Stimulation
mit inaktiviertem RSV war die Vitalität auf 25 % gesunken im Vergleich zu über 80 %
Zellvitalität in den RSV-stimulierten Ansätzen und den Kontroll-Kulturen ohne Virus
(J. Kronsbein, persönliche Mitteilung).

DISKUSSION 91
Zur Klärung der Frage, wie und in welchem Maße RSV im vorliegenden Zellkulturmodell
IL-10 in den T-Zellen aktivierte, sollte der Effekt einer RSV-Stimulierung auch in
Verbindung mit einem mitogenen Stimulus untersucht werden. Damit eine eventuelle
Steigerung bei Zusammentreffen beider Stimuli gut detektiert werden konnte, musste dabei
eine sehr starke Aktivierung der IL-10 Expression durch den nicht-viralen Stimulus allein,
wie sie z.B. bei PHA beobachtet werden konnte (vgl. Abb. 3.24), vermieden werden. Durch
die Zugabe von antiCD28-Antikörpern sollte eine Aktivierung der T-Zellen durch
suboptimale polyklonale Stimulierung erzielt werden, die nicht allein schon zu einer starker
IL-10 Induktion führte und ohne dass von anderen Zelltypen produziertes IL-10 die
Ergebnisse beeinträchtigen konnte. Dass antiCD28-Antikörper stimulatorische Funktion bei
den CD3+14- sortierten T-Zellen in vorliegenden Zellkultursystem ausübten, war durch die
Induktion der CD69-Expression (s. 3.2.1.3) bereits gezeigt worden. Darüber hinaus ist
beschrieben, dass CD28 in Verbindung mit einer Stimulierung der T-Zellen über den
TZR-Komplex über eine nur mäßige stimulatorische Kapazität in Bezug auf die IL-10
Sekretion von T-Zellen verfügt (Hutloff 1999).
Die Ergebnisse der quantitativen IL-10 PCR bestätigten den stimulatorischen Effekt einer
CD28-Bindung in den CD3+14- T-Zellkulturen, da die IL-10 Expression in diesen Kulturen
nach 24h im Mittel um den Faktor 4 erhöht war (vgl. Abb. 3.17). In den RSV-stimulierten
Kulturen stieg die IL-10 Expression bei gleichzeitiger antiCD28-Stimulierung jedoch nur
geringfügig an und war deutlich geringer als das theoretische additive Ergebnis der beiden
Effekte. Dies könnte in Zusammenhang mit der von Akdis (2000) beschriebenen
Beobachtung stehen, dass IL-10 die Phosphorylierung von CD28-Tyrosin und damit den
initialen Schritt des CD28-Signalweges inhibiert. Allerdings waren die spenderabhängigen
Schwankungen der IL-10 Antwort in den antiCD28-stimulierten Kulturen ohne viralen
Stimulus sehr groß. Eine sichere Beurteilung dieser Zusammenhänge bedürfte daher weiterer
Experimente.
Parallel zu diesen Experimenten wurde auch der Effekt einer Stimulierung mit Newcastle-
Disease-Virus (NDV), einem RSV verwandten, aber nicht human pathogenen Virus aus der
Familie der Paramyxoviridae, ebenfalls mit und ohne Zusatz von antiCD28-Antikörpern
untersucht. Die Ergebnisse der Stimulierung mit NDV zeigten, dass die Zellen der
verschiedenen Spender auf NDV weniger einheitlich als auf RSV reagierten. Bei zwei von
5 Spendern konnte keine Erhöhung der IL-10 Expression beobachtet werden, obwohl beide

DISKUSSION 92
auf RSV-Stimulierung mit einer erhöhten IL-10 Expression antworteten. Bei dem Vergleich
zwischen NDV und RSV ist zu berücksichtigen, dass die virale Dosis von RSV und NDV
aufgrund der verschiedenen Verfahren bei der Bestimmung des Virustiters nicht direkt
miteinander verglichen werden können. Allerdings reagierten die Spender, bei denen NDV zu
einer Erhöhung der IL-10 Expression führte, in ähnlicher Höhe wie bei der RSV-Stimulation.
Im Gegensatz zu den Beobachtungen beim IL-10 führte NDV außerdem in PBMC-Kulturen
- in der gleichen Konzentration wie in dieser Arbeit eingesetzt - zu einer wesentlich höheren
Interferon-α Produktion als RSV (Löseke 2002). Dies legt nahe, dass die beobachteten
Unterschiede nicht auf die unterschiedliche Virusdosis, sondern auf verschiedene
Wirkmechanismen zurückzuführen sind. Diese Mechanismen werden in unserer Arbeits-
gruppe zur Zeit weiter untersucht.
Parallel zu den T-Zellkulturen wurde die IL-10 mRNA Expression in CD14+ Monozyten-
kulturen mit und ohne RSV-Stimulierung bestimmt. Die Infizierbarkeit von Monozyten aus
peripherem Blut durch RSV in vitro ist ausreichend belegt, ebenso wie für
Alveolarmakrophagen (Midulla 1989, Panuska 1990), für die darüber hinaus eine erhöhte
IL-10 Produktion nach RSV-Stimulierung gezeigt wurde (Panuska 1995). Für beide Zelltypen
ist auch eine Replikation von RSV in vitro in diesen Zellen beschrieben worden (Krilov 1987,
Panuska 1990).
In den hier untersuchten CD14+ Monozytenkulturen war die IL-10 mRNA Expression 24h
nach Stimulation mit RSV ebenfalls signifikant erhöht. Die durchschnittliche Zunahme lag
mit Faktor 16 bei m.o.i. 1 und Faktor 38 bei m.o.i. 3 deutlich über dem Anstieg der IL-10
Expression in den CD3+14- T-Zellkulturen. Diese bei gleicher Stimulation vermehrte IL-10
Expression der CD14+ Monozyten im Vergleich zu den T-Zellen steht in Einklang mit ihrer
Beschreibung als Hauptproduzenten von IL-10 im humanen System (Borish 1998).
Parallel zur Messung der IL-10 mRNA Expression in den Zellen wurde in den Kulturüber-
ständen die Konzentration an IL-10 Protein mittels spezifischem ELISA bestimmt. Auch
hier ergab sich eine Zunahme nach RSV-Stimulation von CD3+14- oder CD14+ Zellen in
vitro. In Verbindung mit den zugehörigen PCR-Daten zeigte sich eine signifikante
Korrelation zwischen der IL-10 Expressionsrate und der IL-10 Proteinkonzentration im
Überstand. Die beste Korrelation ergab sich zwischen den 24h PCR-Daten und den ELISA-
Daten der zugehörigen 48h-Kulturen. Darin kommt zum Ausdruck, dass eine hohe mRNA-
Expression zum Zeitpunkt 24h erst zeitversetzt nach Ablauf der Proteinbiosynthese und

DISKUSSION 93
Ausschleusung der Proteinmoleküle zu einer erhöhten Zytokinfreisetzung führt, die im
Kulturüberstand mittels ELISA messbar wird. Dass trotzdem auch die 24h-Werte gut
miteinander korrelierten deutet an, dass einer höheren mRNA-Expression zum Zeitpunkt 24h
auch höhere Expression zu früheren Zeitpunkten voran ging, woraus kumulativ bereits eine
erhöhte Proteinkonzentration im Überstand nach 24h resultierte. Insgesamt belegen diese
Ergebnisse, dass nach RSV-Stimulierung der T-Zellen und Monozyten nicht nur mehr IL-10
mRNA in den Zellen vorhanden war, sondern diese mRNA auch translatiert und das
synthetisierte IL-10 Proteinmolekül freigesetzt wurde, so dass die erhöhte mRNA Expression
auch potentiell biologisch wirksam werden konnte.
4.4 Die Rolle RSV-stimulierter T-Zellen in verschiedenen Kokultur-
modellen
Mit den Ergebnissen aus der quantitativen real-time PCR für IL-10 und den zugehörigen
ELISA-Daten konnte nachgewiesen werden, dass die phänotypisch charakterisierte
Interaktion von RSV mit reinen T-Zellen auch zu einer veränderten Funktion der T-Zellen in
Form einer erhöhten IL-10 Produktion führte. Die absolute Höhe dieser Antwort war unter
Berücksichtigung der Ergebnisse der aufgereinigten CD14+ Zellen aber vergleichsweise
gering. Eine trotzdem möglicherweise entscheidende Bedeutung dieser zusätzlichen IL-10
Produktion durch T-Zellen nach RSV-Kontakt zeichnete sich in den Ergebnissen der
Kokultur-Experimente ab. Zumal die Kokulturen die in vivo Bedingungen wesentlich besser
reflektieren als Untersuchungen an einzelnen aufgereinigten Zellpopulationen.
RSV-Stimulierung von Kokulturen aus CD3+14- T-Zellen und CD14+ Monozyten mit
unterschiedlichen Anteilen der beiden Zelltypen führte zu einer Verschiebung des Maximums
der relativen IL-10 mRNA Expression. In unstimulierten Kokulturen war die IL-10
Expression bei einem Anteil von ca. 30 % Monozyten in der Kultur am höchsten. In den
RSV-stimulierten Kokulturen lag die höchste Expressionsrate in Kulturen mit einem
Monozytenanteil von 50 % bei m.o.i. 1 und 90 % bei m.o.i. 3. In dieser Verschiebung der
Maxima zu höheren Monozytenanteilen in den RSV-stimulierten Kulturen spiegelt sich die
bereits gezeigte stärkere Antwort der Monozyten auf RSV wider. Bei einem rein additiven
Ergebnis der IL-10 Produktion von T–Zellen und Monozyten müsste das Maximum in den

DISKUSSION 94
Kulturen mit dem höchsten Monozytenanteil, also in der jeweiligen CD14+-Reinkultur liegen.
Jedoch fiel die IL-10 Expression bei beiden RSV-Stimulationen in den Kulturen mit 100 %
Monozytenanteil wieder ab, woraus sich ein synergistisches Zusammenwirken der beiden
Zelltypen in diesem Kokultursystem ableiten lässt.
Die ELISA-Ergebisse der Kokulturen zeigten ebenfalls diesen Synergismus mit erhöhter
IL-10 Freisetzung bei Anwesenheit beider Zelltypen in den RSV-stimulierten Kulturen im
Vergleich zu den reinen T-Zell- und Monozytenkulturen. Die beschriebenen Unterschiede
zwischen den Kurvenverläufen der PCR- und ELISA-Daten beruhen wahrscheinlich auf den
verschiedenen Informationsgehalten der beiden Messgrößen. Während die PCR-Daten eine
momentane Menge zu einem bestimmten Zeitpunkt angeben, handelt es sich bei den ELISA-
Daten um kumulative Messergebnisse, die sich aus der Zellaktivität der gesamten Kulturdauer
ergeben. Aufgrund der großen Streuung in der absoluten Höhe der Antwort auf die RSV-
Stimulierung bei den einzelnen Spendern ist es aber auch möglich, dass sich diese
Unterschiede zwischen mRNA-Expression und Proteinfreisetzung zufällig ergaben und sich
bei anderer Spenderzusammensetzung nicht gezeigt hätten.
Festzuhalten bleibt, dass unabhängig von den Schwankungen zwischen den einzelnen Spen-
dern und den Unterschieden zwischen mRNA-Expression und Proteinkonzentration im Über-
stand, die IL-10 Antwort auf RSV stets in den Kulturen am höchsten war, in denen ein hoher
Anteil Monozyten in Anwesenheit weniger T-Zellen kultiviert wurde.
Dieser Trend zeigte sich ebenso in Kokulturen aus PHA-stimulierten CD3+ T-Zellen und
Alveolarmakrophagen, die aus der Spülflüssigkeit bronchoalveolärer Lavagen gewonnen
wurden. Wenngleich die Ansätze in diesem Modell in Bezug auf die Zellzahlen nicht ganz
vergleichbar mit denen aus dem CD3+14-/14+-Modell waren, so lag auch hier die IL-10
Konzentration in den Kulturüberständen bei Anwesenheit beider Zelltypen um ein Vielfaches
höher als in den reinen T-Zell- oder Makrophagen-Kulturen. Auch in diesem Kokulturmodell
zeigte sich also ein synergistisches Zusammenspiel von RSV-stimulierten T-Zellen und
Alveolarmakrophagen bei der virusinduzierten IL-10 Antwort.
Im Gegensatz zur Zunahme des immunsuppressiven IL-10 nach RSV-Stimulierung ergab sich
bei den Messungen der IFN-γ Konzentration in beiden Modellen eine tendenzielle Abnahme
des IFN-γ durch die RSV-Stimulation. Dabei war der Trend weniger einheitlich als beim

DISKUSSION 95
IL-10, da sich bei etwa einem Drittel der Spender auch eine gesteigerte IFN-γ Konzentration
fand.
Die Abnahme der IFN-γ Konzentration in den RSV-stimulierten Kulturen könnte IL-10
vermittelt sein (Joss 2000). Dabei ist offen, ob die in diesen Ansätzen erhöhte IL-10
Konzentration die IFN-γ Konzentration direkt beeinflusste oder ob es sich um einen
indirekten Einfluss über andere IFN-γ regulierende Faktoren handelte. Für die
T-Zellproliferation konnte ein direkter hemmender Einfluss von IL-10 durch funktionelle
Blockade des IL-10 Rezeptors gezeigt werden (Joss 2000). Eine Inhibierung der Produktion
verschiedener Zytokine (darunter IFN-γ) durch IL-10 wurde in derselben Arbeit durch
Vergleich der IFN-γ Konzentrationen in PBMC-Kulturen mit und ohne Zusatz von exogenem
IL-10 nachgewiesen. Ein direkter Einfluss von IL-10 auf die IFN-γ Produktion von T-Zellen
ist daher nicht eindeutig belegt. Die bereits weiter oben erwähnte Blockierung des CD28-
Signalweges durch IL-10 (Akdis 2001) legt eine direkte Einwirkung jedoch nahe. Durch
D’Andrea (1993) ist aber auch beschrieben, dass IL-10 die IFN-γ Produktion indirekt über
eine Suppression von IL-12 inhibieren kann. IL-12 wird unter anderem von Monozyten und
Makrophagen ausgeschüttet und induziert oder verstärkt die IFN-γ Produktion von T-Zellen
(Kobayashi 1989; Manetti 1993). Daher ist sowohl in den in der vorliegenden Arbeit
untersuchten Kokultursystemen als auch in PBMC-Kulturen ein indirekter Einfluss von IL-10
auf die IFN-γ Produktion ebenfalls möglich.
Entgegen dem uneinheitlichen Trend bei der RSV-induzierten IFN-γ Antwort stieg der
Quotient aus IL-10 und IFN-γ nach RSV-Stimulierung in den untersuchten Proben signifi-
kant an. Außer in den Ansätzen, in denen IL-10 nach RSV-Stimulierung zunahm und IFN-γ
abnahm, stieg der Quotient auch in mehreren Proben mit gleichzeitiger Zu- oder Abnahme
von IL-10 und IFN-γ dadurch an, dass der IL-10 Anstieg höher war als die Zunahme beim
IFN-γ oder IFN−γ stärker gesunken war als IL-10. In den Kokulturen aus PHA-stimulierten
CD3+ T-Zellen und Alveolarmakrophagen mit einem Zellzahlverhältnis von 1:1 war der
Anstieg des IL-10/IFN-γ Quotienten mit einer mittleren Zunahme um den Faktor 3,3 und
maximaler Zunahme um den Faktor 7,5 besonders deutlich ausgeprägt. Sofern das Verhältnis
zwischen zwei in ihrer Wirkung gegenläufigen Zytokinen wie IL-10 und IFN-γ eher entschei-
dend ist für die Immunantwort als die absolute Höhe der einzelnen Zytokine, führt RSV in
vitro zu einer signifikanten Verschiebung der Immunantwort in Kokulturen aus T-Zellen und
Monozyten/Makrophagen. Dass das Verhältnis zweier Zytokine und nicht die absoluten

DISKUSSION 96
Konzentrationen entscheidend sein können, wird durch folgende Arbeiten beispielhaft belegt.
So entscheidet z.B. das Verhältnis von IL-12 und Prostaglandin E2 (PGE2) über die IFN-γ
Produktion aktivierter humaner CD4+ T-Zellen (Hilkens 1996). Meynaard (1996) konnte
zeigen, dass die Proliferation von T-Zellen maßgeblich durch das Verhältnis von IL-12 und
IL-10 bestimmt wird.
4.5 Möglicher Einfluss RSV-stimulierter T-Zellen auf die virusinduzierte
Immunantwort bei COPD
Die Ergebnisse aus den Kokulturmodellen erbringen Hinweise, wie T-Zellen an der
Pathogenese der COPD beteiligt sein könnten. Wie in der Einleitung dargelegt, finden sich in
der Bronchialschleimhaut von COPD-Patienten im Vergleich zur gesunden Schleimhaut
vermehrt aktivierte Alveolarmakrophagen, T-Zellen und Granulozyten. Die in dieser Arbeit
vorgelegten Daten zeigen, dass insbesondere aktivierte T-Zellen mit einer verstärkten IL-10
Antwort auf RSV-Stimulierung in vitro reagierten. Darüber hinaus führte RSV-Stimulierung
in den Kokulturmodellen mit Monozyten/Makrophagen und autologen T-Zellen bei
Anwesenheit beider Zelltypen zu einer synergistisch verstärkten IL-10 Produktion und zu
einem erhöhten IL-10/IFN-γ Quotienten. Die Zellzahlverhältnisse in den Kokulturansätzen
mit der höchsten IL-10 Antwort nach RSV-Stimulierung waren in Bezug auf das Verhältnis
von Monozyten und T-Zellen den Verhältnissen bei der COPD ähnlich. Die Ergebnisse aus
dem Kokulturmodell mit Alveolarmakrophagen und T-Zellen erscheinen in diesem
Zusammenhang besonders interessant, da sie eher ein lokales Equivalent der Lunge
widerspiegeln als PBMC-Kulturen (Varga 2002). Interessanterweise zeigte gerade der
Quotient aus IL-10 und IFN-γ Unterschiede in der Antwort auf eine RSV-Stimulierung in
vitro zwischen dem Kokulturmodell mit Monozyten aus dem peripheren Blut und dem Modell
mit Alveolarmakrophagen aus der Lunge. Die Verschiebung des IL-10/IFN-γ Quotienten
zugunsten des IL-10 war nur in den Kokulturansätzen mit Alveolarmakrophagen und T-Zellen
einheitlich bei allen Spendern zu finden und darüber hinaus auch deutlicher ausgeprägt als in
den Kokulturen mit peripheren Monozyten. Es liegt daher die Vermutung nahe, dass sich bei
einer RSV-Infektion der unteren Atemwege eines COPD-Patienten eine ähnliche
Verschiebung des IL-10/IFN-γ Verhältnisses ergeben und zu einer Schwächung der
antiviralen Immunantwort führen könnte.

DISKUSSION 97
Auch Daten aus verschiedenen Mausmodellen unterstützen die hier vermutete Verbindung
zwischen RSV-Infektion und obstruktiven Lungenerkrankungen wie COPD und Asthma. So
konnte gezeigt werden, dass das Ausmaß einer Lungeneosinophilie nach RSV-Infektion mit
verminderter IFN-γ und erhöhter IL-10 Produktion von T-Zellen aus bronchoalveolärer
Lavage korrelierte (Hussell 1998).
Die Daten von Takanashi (1999), der im induzierten Sputum von Patienten mit Asthma
(N = 16) und COPD (N = 7) erniedrigte IL-10 Konzentrationen nachwies, stehen hierzu
scheinbar im Widerspruch. Zum einen sind jedoch in der vorliegenden Arbeit nicht direkt
Zellen von COPD-Patienten untersucht worden und zum anderen ist in der zitierten Arbeit
nicht beschrieben, dass bei den Proben ein Virusnachweis durchgeführt wurde. Daher ist es
möglich, dass diese COPD-Patienten zu der Gruppe ohne zugrundeliegende Virusinfektionen
zählten, bei denen andere Mechanismen zur COPD führten. In Bezug auf Asthma steht die
Arbeit von Takanashi im Widerspruch zu einer Studie von Robinson (1996), die bei Asthma-
Patienten eine erhöhte IL-10 mRNA Expression in CD3+ T-Zellen und CD68+
Alveolarmakrophagen aus der Spülflüssigkeit bronchoalveolärer Lavagen ermittelte.
In einer klinischen COPD-Studie (Rohde 2002) hatte RSV einen Anteil von 22 % an den bei
einer Exazerbation nachgewiesenen Viren, wurde im Gegensatz zu Influenza A- und B-,
Parainfluenza- und Picornaviren aber bei keinem der Kontrollpatienten in der stabiler Phase
der COPD gefunden. Die Verwendung sensitiverer Messmethoden in der Fortsetzung dieser
Studie ergab jedoch einen gleich häufigen RSV-Nachweis bei Exazerbation und in der
stabilen Phase mit je 28 % (Borg 2002). Ein ähnliches Ergebnis fand sich in einer anderen
klinischen COPD-Studie (Seemungal 2001), die RSV bei 14 % der Exazerbationen und 23 %
der Proben aus der stabilen Phase der COPD nachwiesen. Während die Ergebnisse von Rohde
(2002) zunächst einen direkten Zusammenhang von RSV-Infektion und Exazerbation der
COPD nahe legten, lassen die Ergebnisse von Seemungal (2001) und Borg (2002) mit Nach-
weis von RSV in der stabilen Phase der COPD auf einen eher indirekten Zusammenhang
schließen. Die hier gezeigten in vitro Daten deuten an, dass eine persistierende RSV-Infektion
in der Lunge von COPD-Patienten aufgrund des T-Zellanteils zu einem erhöhten IL-10
Spiegel und einem in Richtung IL-10 verschobenen IL-10/IFN-γ Verhältnis führen könnte. Es
ist zu erwarten, dass die daraus hervorgerufene Schwächung der antiviralen Immunantwort
den Weg für Infektionen mit anderen Atemwegsviren bahnen könnte. In jüngst

DISKUSSION 98
veröffentlichten Untersuchungen zeigte sich eine Assoziation zwischen Exazerbationen der
COPD und der Isolierung neuer Stämme bakterieller Atemwegspathogene wie Streptococcus
pneumoniae und Moraxella catarrhalis (Sethi 2002). Auch hier wäre es möglich, dass das
durch eine RSV-Infektion induzierte, IL-10 dominierte Zytokinmilieu den neuen Keimen die
in diesem Zusammenhang vermutete Umgehung der Immunabwehr (Anthonisen 2002)
erleichtert.
Der Aspekt der Bahnung der einen Virusinfektion durch die andere könnte insbesondere bei
Rhinoviren, die bei Exazerbationen der COPD besonders häufig nachgewiesen wurden
(Seemungal 2001), eine wichtige Rolle spielen. Außer der Veränderung des Zytokinmilieus
durch die RSV-Infektion ist bei Rhinoviren, deren zellulärer Rezeptor ICAM-1 ist (s. oben),
noch eine weitere Verbindung zu einer RSV-Infektion denkbar. So könnte die verstärkte
Expression von ICAM-1 in der Bronchialschleimhaut bei COPD (Di Stefano 1994) durch eine
RSV-Infektion bedingt sein, da eine Hochregulierung von ICAM-1 auf Epithelzellen nach
RSV-Infektion in vitro nachgewiesen wurde (Arnold 1996; Wang 2000).
__________
Diese Arbeit wurde gefördert durch das BMBF (01GC9802/8) und ein Stipendium der Heinrich-und-Alma-Vogelsang-Stiftung der Medizinischen Fakultät der Ruhr-Universität Bochum.

99
Literatur
Akdis CA, Joss A, Akdis M, Faith A, Blaser K. 2000. A molecular basis for T cell
suppression by IL-10: CD28-associated IL-10 receptor inhibits CD28 tyrosine
phosphorylation and phosphatidylinositol 3-kinase binding. FASEB J 14:1666-1668.
Akdis CA, Blaser K. 2001. Mechanism of interleukin-10-mediated immune suppression.
Immunology 103:131-136.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic local alignment search
tool. J Mol Biol 215:403-410.
Alwan WH, Record FM, Openshaw PJM. 1993. Phenotypic and functional characterisation of
T cell lines specific for individual respiratory syncytial virus proteins. J Immunol
150:5211-5218.
Anthonisen NR. 2002. Bacteria and exacerbations of chronic obstructive pulmonary disease.
N Engl J Med 347:526-527.
Arnold R, Humbert B, Werchau H, Gallati H, König W. 1994. Interleukin-8, interleukin-6,
and soluble tumour necrosis factor receptor type I release from a human pulmonary
epithelial cell line (A549) exposed to respiratory syncytial virus. Immunology 82:126-
133.
Arnold R, König B, Gallati H, Werchau H, König W. 1995. Cytokine (IL-8, IL-6, TNF-alpha)
and soluble TNF receptor-I release from human peripheral blood mononuclear cells
after respiratory syncytial virus infection. Immunology 85:364-372.
Arnold R, König W. 1996. ICAM-1 expression and low-molecular-weight G-protein
activation of human bronchial epithelial cells (A549) infected with RSV. J Leukoc
Biol 60:766-771.
Barnes PJ. 2000. Chronic obstructive pulmonary disease. N Engl J Med 343:269-280.

LITERATUR 100
Barnes PJ. 2000a. Mechanism in COPD - differences from asthma. Chest 117:10S-14S.
Barth S, Kleinhappl B, Gutschi A, Jelovcan S, Marth E. 2000. In vitro cytokine mRNA
expression in normal human peripheral blood mononuclear cells. Inflamm Res 49:266-
274.
Becker S, Quay J, Soukup J. 1991. Cytokine (Tumor necrosis factor, IL-6, and IL-8)
production by respiratory syncytial virus-infected human alveolar macrophages.
J Immunol 147:4307-4312.
Becker S, Soukup J, Yankaskas JR. 1992. Respiratory syncytial virus infection of human
primary nasal and bronchial epithelial cell cultures and bronchoalveolar macrophages.
Am J Respir Cell Mol Biol 6:369-374.
Becker S, Reed W, Henderson FW, Noah TL. 1997. RSV infection of human airway
epithelial cells causes production of beta-chemokine RANTES. Am J Physiol
272:L512-L520.
Bernard P, Gabant P, Bahassi EM, Couturier M. 1994. Positive-selection vectors using the F
plasmid ccdB killer gene. Gene 148:71-74.
Bont L, Heijnen CJ, Kavelaars A, van Aalderen WM, Brus F, Draaisma JTM, Geelen SM,
Kimpen JL. 2000. Monocyte IL-10 production during respiratory syncytial virus
bronchiolitis is associated with recurrent wheezing in a one-year follow-up study.
Am J Respir Crit Care Med 161:1518-1523.
Borg I, Rohde G, Löseke S, Bittscheidt J, Schultze-Werninghaus G, Stephan V, Bufe A. 2002.
Evaluation of a quantitative real-time PCR assay for the detection of respiratory
syncytial virus in pulmonary diseases. Eur Respir J, eingereicht.
Borish L. 1998. IL-10: Evolving concepts. J Allergy Clin Immunol 101:293-297.
Böyum A. 1968. Isolation of mononuclear cells and granulocytes from human blood.
Isolation of mononuclear cells by one centrifugation, and of granulocytes by
combining centrifugation and sedimentation at 1 g. Scan J Clin Lab Invest 97:77-89.

LITERATUR 101
Bustin SA. 2000. Absolute quantification of mRNA using real-time reverse transcription
polymerase chain reaction assays. J Mol Endocrinol 25:169-193.
Cherrie AH, Anderson K, Wertz GW, Openshaw PJM. 1992. Human cytotoxic T cells
stimulated by antigen on dendritic cells recognise the N, SH, F 22K and 1b proteins of
respiratory syncytial virus. J Virol 66:2102-2110.
Collins PL, Chanock RM, Murphy BR. 2001. Respiratory syncytial virus. In: Knipe D,
(editor). Fundamental Virology. 4th edition, Raven press, New York, p. 1443-1485.
Corinti S, Albanesi C, la Sala A, Pastore S, Girolomoni G. 2001. Regulatory activity of
autocrine IL-10 on dendritic cell functions. J Immunol 166:4312-4318.
Croxton TL, Weinmann GG, Senior RM, Hoidal JR. 2002. Future research directions in
chronic obstructive pulmonary disease. Am J Respir Crit Care Med 165:838-844.
D'Andrea A, Aste-Amezaga M, Valiante NM, Ma X, Kubin M, Trinchieri G. 1993.
Interleukin 10 (IL-10) inhibits human lymphocyte interferon gamma-production by
suppressing natural killer cell stimulatory factor/IL-12 synthesis in accessory cells.
J Exp Med 178:1041-1048.
Dakhama A, Vitalis TZ, Hegele RG. 1997. Persistence of respiratory syncytial virus (RSV)
infection and development of RS-specific IgG1 response in a guinea-pig model of
acute bronchiolitis. Eur Respir J 10:20-26.
de la Torre JC, Borrow P, Oldstone MB. 1991. Viral persistence and disease: cytopathology
in the absence of cytolysis. Med Bull 47:838-851.
de Waal Malefyt R, Abrams J, Bennett B, Figdor CG, de Vries JE. 1991. Interleukin 10
(IL-10) inhibits cytokine synthesis by human monocytes: an autoregulatory role of
IL-10 produced by monocytes. J Exp Med 174:1209-1220.
de Waal Malefyt R, Haanen J, Spits H, Roncarolo M, te Velde A, Figdor C, Johnson K,
Kastelein R, Yssel H, de Vries JE. 1991a. Interleukin 10 (IL-10) and viral IL-10
strongly reduce antigen-specific human T cell proliferation by diminishing the

LITERATUR 102
antigen-presenting capacity of monocytes via downregulation of class II major
histocompatibility complex expression. J Exp Med 174:915-924.
del Prete G, De Carli M, Almerigogna F, Giudizi MG, Biagiotti R, Romagnani S. 1993.
Human IL-10 is produced by both type 1 helper (Th1) and type 2 helper (Th2) T cell
clones and inhibits their antigen-specific proliferation and cytokine production.
J Immunol 150:353-360.
Di Stefano A, Maestrelli P, Roggeri A, Turato G, Calabro S, Potena A, Mapp CE, Ciaccia A,
Covacev L, Fabbri LM, et al. 1994. Upregulation of adhesion molecules in the
bronchial mucosa of subjects with chronic obstructive bronchitis. Am J Respir Crit
Care Med 149:803-810.
Diaz PV, Calhoun WJ, Hinton KL, Avendano LF, Gaggero A, Simon V, Arredondo SM,
Pinto R, Diaz A. 1999. Differential Effects of respiratory syncytial virus and
adenovirus on mononuclear cell cytokine responses. Am J Respir Crit Care Med
160:1157-1164.
Ding L, Linsley PS, Huang L, Germain RN, Shevach EM. 1993. IL-10 inhibits macrophage
costimulatory activity by selectively inhibiting the up-regulation of B7 expression.
J Immunol 151 :1224-1234.
Dustin ML, Rothlein R, Bhan AK, Dinarello CA, Springer TA. 1986. Induction by IL1 and
interferon-gamma: tissue distribution, biochemistry, and function of a natural
adherence molecule (ICAM-1). J Immunol 137:245-254.
Eriksson S. 1996. A 30-year perspective on alpha 1-antitrypsin deficiency. Chest 110:237S-
242S.
Finlay GA, O'Driscoll LR, Russell KJ, D'Arcy EM, Masterson JB, Fitzgerald MX,
O'Connor CM. 1997. Matrix metalloproteinase expression and production by alveolar
macrophages in emphysema. Am J Respir Crit Care Med 156:240-247.
Freeman WM, Walker SJ, Vrana KE. 1999. Quantitative RT-PCR: Pitfalls and Potential.
BioTechniques 26:112-125.

LITERATUR 103
Greve JM, Davis G, Meyer AM, Forte CP, Yost SC, Marlor CW, Kamarck ME,
McClelland A. 1989. The major human rhinovirus receptor is ICAM-1. Cell 56:839-
847.
Groux H, O'Garra A, Bigler M, Rouleau M, Antonenko S, de Vries JE, Roncarolo M. 1997.
A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis.
Nature 389:737-742.
Hall CB. 2001. Respiratory syncytial virus and parainfluenza virus. N Engl J Med 344:1917-
1928.
Hara T, Jung LK, Björndahl JM, Fu SM. 1986. Human T cell activation. III. Rapid induction
of a phosphorylated 28 kD/32 kD disulfide-linked early activation antigen (EA 1) by
12-o-tetradecanoyl phorbol-13-acetate, mitogens, and antigens. J Exp Med 164:1988-
2005.
Higham MA, Pride NB, Alikhan A, Morrell NW. 2000. Tumor necrosis factor-α gene
promotor polymorphism in chronic obstructive pulmonary disease. Eur Respir J
15:281-284.
Hilkens CMU, Snijders A, Vermeulen H, van der Meide PH, Wierenga EA, Kapsenberg ML.
1996. Accessory cell-derived IL-12 and Prostaglandin E2 determine the IFN-γ level of
activated human CD4+ T cells. J Immunol 156:1722-1727.
Hogg JC. 1999. Childhood viral infection and the pathogenesis of asthma and chronic
obstructive lung disease. Am J Respir Crit Care Med 160:S26-S28.
Hogg JC. 2001. Viral infection and exacerbations of chronic obstructive pulmonary disease.
Am J Respir Crit Care Med 164:1555-1556.
Hsu DH, de Waal Malefyt R, Fiorentino DF, Dang MN, Vieira P, de Vries JE, Spits H,
Mosmann TR, Moore KW. 1990. Expression of interleukin-10 activity by Epstein-
Barr virus protein BCRF1. Science 250:830-832.

LITERATUR 104
Huang SL, Su CH, Chang SC. 1997. Tumor necrosis factor-alpha gene polymorphism in
chronic bronchitis. Am J Respir Crit Care Med 156:1436-1439.
Hussell T, Baldwin CJ, O'Garra A, Openshaw PJ. 1997. CD8+ T cells control Th2-driven
pathology during pulmonary respiratory syncytial virus infection. Eur J Immunol
27:3341-3349.
Hussell T, Georgiou A, Sparer TE, Matthews S, Pala P, Openshaw PJM. 1998. Host genetic
determinants of vaccine-induced eosinophilia during respiratory syncytial virus
infection. J Immunol 161:6215-6222.
Hutloff A, Dittrich AM, Beier KC, Eljaschewitsch B, Kraft R, Anagnostopoulos I, Kroczek
RA. 1999. ICOS is an inducible T-cell co-stimulator structurally and functionally
related to CD28. Nature 397:263-266.
Jackson M, Scott R. 1996. Different patterns of cytokine induction in cultures of respiratory
syncytial (RS) virus-specific human TH cell lines following stimulation with RS virus
and RS virus proteins. J Med Virol 49:161-169.
Janeway CA, Travers P. 1997. Immunologie, 2. Auflage. Spektrum Akademischer Verlag,
Heidelberg. Kapitel 1, 4 und 7.
Jonuleit H, Schmitt E, Schuler G, Knop J, Enk AH. 2000. Induction of interleukin 10-
producing, nonproliferating CD4+ T cells with regulatory properties by repetitive
stimulation with allogeneic immature human dendritic cells. J Exp Med 192:1213-
1222.
Joss A, Akdis M, Faith A, Blaser K, Akdis CA. 2000. IL-10 directly acts on T cells by
specifically altering the CD28 co-stimulation pathway. Eur J Immunol 30:1683-1690.
Jung R, Soondrum K, Neumaier M. 2000. Quantitative PCR. Clin Chem Lab Med 38:833-
836.
Jung T, Schauer U, Heusser C, Neumann C, Rieger CHL. 1993. Detection of intracellular
cytokines by flow cytometry. J Immunol Methods 159:197-207.

LITERATUR 105
Keatings VM, Collins PD, Scott DM, Barnes PJ. 1996. Differences in interleukin-8 and tumor
necrosis factor-α in induced sputum from patients with chronic obstructive pulmonary
disease or asthma. Am J Respir Crit Care Med 153:530-534.
Kemeny DM, Vyas B, Vukmanovic-Stejic M, Thomas MJ, Noble A, Loh L, O'Connor BJ.
1999. CD8+ T cell subsets and chronic obstructive pulmonary disease. Am J Respir
Crit Care Med 160:S33-S37.
Kobayashi M, Fitz L, Ryan M, Hewick RM, Clark SC, Chan S, Loudon R, Sherman F,
Perussia B, Trinchieri G. 1989. Identification and purification of natural killer cell
stimulatory factor (NKSF), a cytokine with multiple biologic effects on human
lymphocytes. J Exp Med 170:827-845.
König B, Streckert HJ, Krusat T, König W. 1996. Respiratory syncytial virus G-protein
modulates cytokine release from human peripheral blood mononuclear cells. J Leukoc
Biol 59:403-406.
Kotenko SV, Saccani S, Izotova LS, Mirochnitchenko OV, Pestka S. 2000. Human
cytomegalovirus harbors its own unique IL-10 homolog (cmvIL-10). Proc Natl Acad
Sci U S A 97:1695-1700.
Krilov LR, Hendry RM, Godfrey E, McIntosh K. 1987. Respiratory virus infection of
peripheral blood monocytes: correlation with ageing of cells and interferon production
in vitro. J Gen Virol 68:1749-1753.
Lanier LL, Buck DW, Rhodes L, Ding A, Evans E, Barney C, Phillips JH. 1988. Interleukin-2
activation of natural killer cells rapidly induces the expression and phosphorylation of
the Leu-23 activation antigen. J Exp Med 167:1572-1585.
Levings MK, Roncarolo M. 2000. T-regulatory 1 cells: a novel subset of CD4+ T cells with
immunoregulatory properties. J Allergy Clin Immunol 106:S109-S112.
Liu AN, Mohammed AZ, Rice WR, Fiedeldey DT, Lieberman JS, Whitsett JA, Braciale TJ,
Enelow RI. 1999. Perforin-independent CD8+ T-cell-mediated cytotoxity of alveolar

LITERATUR 106
epithelial cells is preferentially mediated by tumor necrosis factor-α: relative
insensitivity to Fas ligand. Am J Respir Cell Mol Biol 20:849-858.
Longo MC, Berninger MS, Hartley JL. 1990. Use of uracil DNA glycosylase to control carry-
over contamination in polymerase chain reactions. Gene 93:125-128.
Lopez AD, Murray CC. 1998. The global burden of disease, 1990-2020. Nat Med 4:1241-
1243.
Löseke S, Grage-Griebenow E, Wagner A, Gehlhar K, Bufe A. 2002. Differential expression
of IFN-α subtypes in human PBMC; evaluation of novel real-time PCR assays.
J Immunol Methods, eingereicht.
Macek V, Sorli J, Kopriva S, Marin J. 1994. Persistent adenoviral infection and chronic
airway obstruction in children. Am J Respir Crit Care Med 150:7-10.
Mahadeva R, Lomas DA. 1998. Genetics and respiratory disease. 2. Alpha 1-antitrypsin
deficiency, cirrhosis and emphysema. Thorax 53:501-505.
Manetti R, Parronchi P, Giudizi MG, Piccinni MP, Maggi E, Trinchieri G, Romagnani S.
1993. Natural killer cell stimulatory factor (interleukin 12 [IL-12]) induces T helper
type 1 (Th1)-specific immune responses and inhibits the development of IL-4-
producing Th cells. J Exp Med 177:1199-204.
Matsuse T, Hayashi S, Kuwano K, Keunecke H, Jefferies WA, Hogg JC. 1992. Latent
adenoviral infection in the pathogenesis of chronic airway obstruction. Am Rev Respir
Dis 146:177-184.
Mäkelä MJ, Kanehiro A, Dakhama A, Borish L, Joetham A, Tripp R, Anderson L, Gelfand
EW. 2002. The failure of interleukin-10-deficient mice to develop airway
hyperresponsiveness is overcome by respiratory syncytial virus infection in allergen-
sensitized/challenged mice. Am J Respir Crit Care Med 165:824-831.

LITERATUR 107
Meynaard L, Hovenkamp E, Otto SA, Miedema F. 1996. IL-12-induced IL-10 production by
human T cells as a negative feedback for IL-12-induced immune responses. J Immunol
156:2776-2782.
Midulla F, Huang YT, Gilbert IA, Cirino NM, McFadden ER, jr., Panuska JR. 1989.
Respiratory syncytial virus infection of human cord and adult blood monocytes and
alveolar macrophages. Am Rev Respir Dis 140:771-777.
Midulla F, Villani A, Panuska JR, Dab I, Kolls JK, Merolla R, Ronchetti R. 1993. Respiratory
syncytial virus lung infection in infants: immunregulatory role of infected alveolar
macrophages. J Infect Dis 168:1515-1519.
Miltenyi S, Müller W, Weichel W, Radbruch A. 1990. High gradient magnetic cell separation
with MACS. Cytometry 11:231-238.
Modrow S, Falke D. 1998. Molekulare Virologie, 1. korrigierter Nachdruck. Spektrum
Akademischer Verlag, Heidelberg. Kapitel 14.2:203-226.
Mosmann TR, Coffman RL. 1989. TH1 and TH2 cells: different patterns of lymphokine
secretion lead to different functional properties. Annu Rev Immunol 7:145-173.
Mullis KB, Faloona FA. 1987. Specific synthesis of DNA in vitro via a polymerase-catalyzed
chain reaction. Methods Enzymol 155:335-350.
Noah TL, Becker S. 1993. Respiratory syncytial virus-induced cytokine production by a
human bronchial epithelial cell line. Am J Physiol 265:L472-L478.
O'Donnell DR, McGarvey MJ, Tully JM, Balfour-Lynn IM, Openshaw PJ. 1998. Respiratory
syncytial virus RNA in cells from the peripheral blood during acute infection.
J Pediatr 133:272-274.
O'Shaughnessy TC, Ansari TW, Barnes NC, Jeffery PK. 1997. Inflammation in bronchial
biopsies of subjects with chronic bronchitis: inverse relationship of CD8+
T lymphocytes with FEV1. Am J Respir Crit Care Med 155:852-857.

LITERATUR 108
Oldstone MB. 1989. Viral Persistence. Cell 56:517-520.
Olszewska-Pazdrak B, Casola A, Saito T, Alam R, Crowe SE, Mei F, Ogra PL, Garofalo P.
1998. Cell-specific expression of RANTES, MCP-1, and MIP-1α by lower airway
epithelial cells and eosinophils infected with respiratory syncytial virus. J Virol
72:4756-4764.
Openshaw PJM. 1995. Immunopathological mechanism in respiratory syncytial virus disease.
Springer Semin Immunopathol 17:187-201.
Panuska JR, Cirino NM, Midulla F, Despot JE, McFadden ER, Jr., Huang YT. 1990.
Productive Infection of isolated human alveolar macrophages by respiratory syncytial
virus. J Clin Invest 86:113-119.
Panuska JR, Merolla R, Rebert NA, Hoffmann SP, Tsivitse P, Cirino NM, Silverman RH,
Rankin JA. 1995. Respiratory syncytial virus induces interleukin-10 by human
alveolar macrophages. Suppression of early cytokine production and implications for
incomplete immunity. J Clin Invest 96:2445-2453.
Parkin J, Cohen B. 2001. An overview of the immune system. Lancet 357:1777-1789.
Pauwels RA, Buist AS, Calverley PMA, Jenkins CR, Hurd SS. 2001. Global strategy for the
diagnosis, management, and prevention of chronic obstructive pulmonary disease.
Am J Respir Crit Care Med 163:1256-1276.
Risso A, Cosulich ME, Rubartelli A, Mazza MR, Bargellesi A. 1989. MLR3 molecule is an
activation antigen shared by human B, T lymphocytes and T cell precursors. Eur J
Immunol 19:323-328.
Robinson DS, Tsicopoulos A, Meng Q, Durham S, Kay AB, Hamid Q. 1996. Increased
interleukin-10 messenger RNA expression in atopic allergy and asthma. Am J Respir
Cell Mol Biol 14:113-117.
Rohde G, Wiethege A, Borg I, Kauth M, Bauer TT, Gillissen A, Bufe A, Schultze-
Werninghaus G. 2002. Respiratory viruses in exacerbations of chronic obstructive

LITERATUR 109
pulmonary disease requiring hospitalization - a case-control study. Thorax, zur
Veröffentlichung angenommen.
Saetta M, Di Stefano A, Maestrelli P, Ferraresso A, Drigo R, Potena A, Ciaccia A,
Fabbri LM. 1993. Activated T-lymphocytes and macrophages in bronchial mucosa of
subjects with chronic bronchitis. Am Rev Respir Dis 147:301-306.
Saetta M, Di Stefano A, Turato G, Facchini FM, Corbino L, Mapp CE, Maestrelli P,
Ciaccia A, Fabbri LM. 1998. CD8+ T-lymphocytes in peripheral airways of smokers
with chronic obstructive pulmonary disease. Am J Respir Crit Care Med 157:822-826.
Salgame P, Abrams JS, Clayberger C, Goldstein H, Convit J, Modlin RL, Bloom BR. 1991.
Differing lymphokine profiles of functional subsets of human CD4 and CD8 T cell
clones. Science 254:279-282.
Sambrook J, Russell DW. 2001. Molecular cloning: a laboratory manual. 3rd edition. Cold
Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA.
Sanger F, Nicklen S, Coulson AR. 1977. DNA sequencing with chain-terminating inhibitors.
Proc Natl Acad Sci U S A 74:5463-5467.
Schwarze J, Cieslewicz G, Joetham A, Ikemura T, Hamelmann E, Gelfand EW. 1999. CD8
T cells are essential in the development of respiratory syncytial virus-induced lung
eosinophilia and airway hyperresponsiveness. J Immunol 162:4207-4211.
Schwarze J, Cieslewicz G, Hamelmann E, Joetham A, Shultz LD, Lamers MC, Gelfand EW.
1999a. IL-5 and eosinophils are essential for the development of airway
hyperresponsiveness following acute respiratory syncytial virus infection. J Immunol
162:2997-3004.
Schwarze J, Mäkelä MJ, Cieslewicz G, Dakhama A, Lahn M, Ikemura T, Joetham A,
Gelfand EW. 1999b. Transfer of the enhancing effect of respiratory syncytial virus
infection on subsequent allergic airway sensitization by T lymphocytes. J Immunol
163:5729-5734.

LITERATUR 110
Seemungal T, Harper-Owen R, Bhowmik A, Moric I, Sanderson G, Message S,
MacCallum P, Meade TW, Jeffries DJ, Johnston SL, Wedzicha JA. 2001. Respiratory
viruses, symptoms, and inflammatory markers in acute exacerbation and stable chronic
obstructive pulmonary disease. Am J Respir Crit Care Med 164:1618-1623.
Sethi S. 2000. Bacterial infection and the pathogenesis of COPD. Chest 117:286S-291S.
Sethi S. 2000a. Infectious etiology of acute exacerbations of chronic bronchitis. Chest
117:380S-385S.
Sethi S, Evans N, Grant BJB, Murphy TF. 2002. New strains of bacteria and exacerbations of
chronic obstructive pulmonary disease. N Engl J Med 347:465-471.
Shapiro SD. 1999. The macrophage in chronic obstructive pulmonary disease. Am J Respir
Crit Care Med 160:S29-S32.
Shuman S. 1994. Novel approach to molecular cloning and polynucleotide synthesis using
vaccinia DNA topoisomerase. J Biol Chem 269:32678-32684.
Sigurs N. 2001. Epidemiologic and clinical evidence of a respiratory syncytial virus-reactive
airway disease link. Am J Respir Crit Care Med 163:S2-S6.
Simoes EAF. 2001. Treatment and prevention of respiratory syncytial virus lower respiratory
tract infection. Am J Respir Crit Care Med 163:S14-S17.
Spencer JV, Lockridge KM, Barry PA, Lin G, Tsang M, Penfold MET, Schall TJ. 2002.
Potent immunosuppressive activities of cytomegalovirus-encoded interleukin-10.
J Virol 76:1285-1292.
Staunton DE, Merluzzi VJ, Rothlein R, Barton R, Marlin SD, Springer TA. 1989. A cell
adhesion molecule, ICAM-1, is the major surface receptor for rhinoviruses. Cell
56:849-853.

LITERATUR 111
Stein RT, Sherrill D, Morgan WJ, Holberg CJ, Halonen M, Taussig LM, Wright AL,
Martinez FD. 1999. Respiratory syncytial virus in early life and risk of wheeze and
allergy by age 13 years. Lancet 354:541-545.
Stockley RA. 1999. Neutrophils and protease/antiprotease imbalance. Am J Respir Crit Care
Med 160:S49-S52.
Stöckl J, Vetr H, Majdic O, Zlabinger G, Kuechler E, Knapp W. 1999. Human major group
rhinoviruses downmodulate the accessory function of monocytes by inducing IL-10.
J Clin Invest 104:957-965.
Takanashi S, Hasegawa Y, Kanehira Y, Yamamoto K, Fujimoto K, Satoh K, Okamura K.
1999. Interleukin-10 level in sputum is reduced in bronchial asthma, COPD and in
smokers. Eur Respir J 14:309-314.
Teng MN, Collins PL. 2002. The central conserved cystine noose of the attachment G Protein
of human respiratory syncytial virus is not required for efficient viral infection in vitro
or in vivo. J Virol 76:6164-6171.
Thurau AM, Streckert HJ, Rieger CHL, Schauer U. 1998. Increased number of T cells
committed to IL-5 production after respiratory syncytial virus (RSV) infection of
human mononuclear cells in vitro. Clin Exp Immunol 114:450-455.
Tripp RA, Hou S, McMickle A, Houston J, Doherty PC. 1995. Recruitment and proliferation
of CD8+ T cells in respiratory virus infections. J Immunol 154:6013-6021.
Varga SM, Braciale TJ. 2002. RSV-induced immunopathology: dynamic interplay between
the virus and the host immune response. Virology 295:203-207.
Vestbo J, Lange P. 2002. Can GOLD stage 0 provide information of prognostic value in
chronic obstructive pulmonary disease. Am J Respir Crit Care Med 166:329-332.
Vieira P, de Waal Malefyt R, Dang MN, Johnson KE, Kastelein R, Fiorentino DF, de Vries
JE, Roncarolo M, Mosmann TR, Moore KW. 1991. Isolation and expression of human

LITERATUR 112
cytokine synthesis inhibitory factor cDNA clones: homology to Epstein-Barr virus
open reading frame BCRFI. Proc Natl Acad Sci U S A 88:1172-1176.
Vukmanovic-Stejic M, Vyas B, Gorak-Stolinska P, Noble A, Kemeny DM. 2000. Human Tc1
and Tc2/Tc0 CD8 T-cell clones display distinct cell surface and functional
phenotypes. Blood 95:231-240.
Wang S, Hallsworth PG, Dowling KD, Alpers JH, Bowden JJ, Forsyth KD. 2000. Adhesion
molecule expression on epithelial cells infected with respiratory syncytial virus.
Eur Respir J 15:358-366.
Weigl JAI, Puppe W, Schmitt HJ. 2001. Incidence of respiratory syncytial virus-positive
hospitalizations in Germany. Eur J Clin Microbiol Infect Dis 20:452-459.
Weiner HL. 2001. The mucosal milieu creates tolerogenic dendritic cells and Tr1 and Th3
regulatory cells. Nat Immunol 2:671-672.
Wilson R, Grossman R. 2000. Introduction: the role of bacterial infection in chronic
bronchitis. Semin Respir Infect 15:1-6.
World Health Organization. 2000. World Health Report. Geneva. Available from URL:
http://www.who.int/whr/2000/en/statistics.htm, table 4.
Yssel H, de Waal Malefyt R, Roncarolo M, Abrams J, Lahesmaa R, Spits H, de Vries JE.
1992. IL-10 is produced by subsets of human CD4+ T cell clones and peripheral blood
T cells. J Immunol 149:2378-2384.
Zakrzewski JT, Barnes NC, Costello JF, Piper PJ. 1987. Lipid mediators in cystic fibrosis and
chronic obstructive pulmonary disease. Am Rev Respir Dis 136:779-782.
Zhang L, Peeples ME, Boucher RC, Collins PL, Pickles RJ. 2002. Respiratory syncytial virus
infection of human airway epithelial cells is polarized, specific to ciliated cells, and
without obvious cytopathology. J Virol 76:5654-5666.

Danksagung
Ich danke Herrn Prof. Dr. med. A. Bufe und Herrn Prof. Dr. med. G. Schultze-Werninghaus
für die Überlassung des Themas und die gute Betreuung im Verlauf dieser Arbeit.
Bei Herrn Prof. Dr. Lübbert bedanke ich mich für die Übernahme des Korreferats.
Frau Dr. rer. nat. E. Grage-Griebenow, Frau Dr. rer. nat. K. Gehlhar, Frau I. Borg, Herrn
S. Löseke und den übrigen Mitgliedern der Arbeitsgruppe Experimentelle Pneumologie danke
ich für Ihre vielfältige Unterstützung, stetige Diskussionsbereitschaft und die gute Zusam-
menarbeit.
Herrn O. Anhenn vom Institut für Pathologie der Ruhr-Universität Bochum danke ich für die
wertvolle Unterstützung bei den Immunfluoreszenz-Aufnahmen.
Allen Mitgliedern der Abteilung für Pneumologie, Allergologie und Schlafmedizin, insbeson-
dere Frau B. Schärling und Frau S. Werner danke ich für viele praktische Tipps und das
freundliche Arbeitsklima.
Insbesondere möchte ich mich bei Frau Dr. rer. nat. E. Grage-Griebenow und Frau
Dr. rer. nat. K. Gehlhar für das Lesen des Manuskripts und die wertvollen Anregungen
bedanken.

Liste der Veröffentlichungen und Vorträge
Rohde G, Wiethege A, Borg I, Kauth M, Bauer TT, Gillissen A, Bufe A, Schultze-
Werninghaus G. 2002. Respiratory viruses in exacerbations of chronic obstructive pulmonary
disease requiring hospitalization – a case control study. Thorax, zur Veröffentlichung
angenommen.
Kauth M, Werner S, Wiethege A, Rohde G, Schultze-Werninghaus G, Bufe A. 2002. Einfluss
einer Infektion mit respiratory syncytial virus (RSV) auf die Interleukin-10-Expression
verschiedener PBMC-Populationen in vitro. Pneumologie 56:S64.
Kauth M, Rohde G, Werner S, Wiethege A, Bufe A, Schultze-Werninghaus G. 2000. RSV-
induced effects on oxidative burst and cytokine production of alveolar macrophages and co-
cultures T-lymphocytes in vitro. Pneumologie 54:551.
Kauth M, Bartling A, Rohde G, Kronsbein J, Gillissen A, Schultze-Werninghaus G. 1999.
Effects of RSV-exposition on human T-lymphocytes in vitro. Postervortrag, Symposium
„RSV after 43 years“ 8.-11.11.1999 in Stuart, Florida, USA.
Kauth M, Bartling A, Rohde G, Kronsbein J, Gillissen A, Schultze-Werninghaus G. 1999.
Effekte der RSV-Exposition auf menschliche T-Lymphozyten in vitro. Pneumologie 53:570.
Jäckel S, Epplen JT, Kauth M, Miterski B, Tschentscher F, Epplen C. 1998. Polymerase chain
reaction - single strand conformation polymorphism or how to detect reliably and efficiently
each sequence variation in many samples and many genes. Electrophoresis 19:3055-3061.
Kauth M, Engel S, Lastimoza LL, Curio E. 1998. Observations on the breeding biology of the
Writhed-billed Hornbill (Aceros waldeni) in the Philippines, Journal für Ornithologie
139:475-483.
D'Souza M, Kauth M, Frank G, Epplen JT, Pöhlau D, Sindern E, Haupts M, Weber F, Epplen
C. 1997. Polymorphisms of HLA-DRB1 and TCRBV genes confer susceptibility for multiple
sclerosis. Medizinische Genetik 9:474.

Lebenslauf
persönliche Daten
Name: Marion Kauth
Geburtsdatum: 25.07.1966
Geburtsort: Wattenscheid
Familienstand: ledig
Schul- und Berufsausbildung
08/72 - 06/76 Grundschule in Wattenscheid
08/76 - 06/85 Hellweg-Gymnasium in Bochum, Abschluß: Abitur
09/85 - 10/88 Ausbildung zur Industriekauffrau und zur Betriebswirtin (VWA) bei der Steag AG, Essen / Verwaltungs- und Wirtschaftsakademie, Bochum
Studium
10/92 - 03/98 Biologiestudium an der Ruhr-Universität Bochum
03/96 - 05/96 Auslandsaufenthalt anlässlich der Mitarbeit in einem Natur- und Arten-schutzprojekt auf den Philippinen (Leitung Prof. Dr. E. Curio)
12/96 - 01/97 Diplomhauptprüfung in den Fächern Mikrobiologie, Zoologie und Molekulare Humangenetik
02/97 - 03/98 Diplomarbeit bei Prof. Dr. J. T. Epplen, Abteilung für Molekulare Humangenetik, Medizinische Fakultät; Thema: Polymorphismen der immunmodulierenden Gene CD40 und CD40L und ihre Assoziation zur Multiplen Sklerose
Berufstätigkeit und Promotion
10/88 - 09/92 Kaufmännische Sachbearbeiterin bei der Steag AG, Essen
07/98 – 07/01 Wissenschaftliche Angestellte in der Abteilung für Pneumologie, Allergologie und Schlafmedizin, Universitätsklinik Bergmannsheil, Bochum (Prof. Dr. G. Schultze-Werninghaus), im Verlauf dieser Tätigkeit Beginn der Arbeiten für die Promotion
08/01 – 07/02 Stipendiatin der Heinrich-und-Alma-Vogelsang-Stiftung, Fortsetzung der Promotionsarbeiten, Betreuung durch Prof. Dr. A. Bufe
08/02 – lfd. Wissenschaftliche Angestellte in der Abteilung für Experimentelle Pneumologie, Medizinische Fakultät, Ruhr-Universität Bochum (Prof. Dr. A. Bufe), Abgabe der Dissertation zum 15.10.2002





























