Embed Size (px)
Citation preview

British Journal of Oral and Maxillofacial Surgery (2004) 42, 315—322
Comparison of the healing process in plated andnon-plated fractures of the mandible in rats
L. Rasubalaa,*, H. Yoshikawaa, A.A.S. Islama, K. Nagatab, T. Iijimab,M. Ohishia
a Department of Oral and Maxillofacial Oncology, Division of Maxillofacial Diagnostic and SurgicalSciences, Faculty of Dental Science, Kyushu University, Fukuoka 812-8582, Japanb Department of Oral Anatomy and Cell Biology, Faculty of Dental Science, Kyushu University,Fukuoka 812-8582, Japan
Accepted 22 February 2004
Available online 6 May 2004
KEYWORDSHealing process;Plated fracture;Mandible
Summary We compared the healing process of plated and non-plated fractures. Themandibles of 72 male Wistar rats were fractured and more either plated or not plated(n = 36 in each group). The healing process of the two conditions was studied histo-logically and immunohistochemically using a specific antibody to bone morphogeneticprotein-2 (BMP-2). The results showed that the healing process in the plated groupwas delayed by one week compared with the non-plated group. Trauma to the sur-rounding soft tissues affected the healing process. BMP-2 was expressed at all stagesin both groups. We conclude that the healing process is disturbed by the fixing of aplate; that periosteum is one of the main sources of osteogenic cells; and that BMP-2is an important regulator of morphogenesis.© 2004 The British Association of Oral and Maxillofacial Surgeons. Published by ElsevierLtd. All rights reserved.
Introduction
The aim of treatment of fractures is to restorethe architecture and function of the bone. Thehealing process is affected by many variables in-cluding the extent of damage to soft and hardtissue, the vascular supply, and the formation ofa haematoma.1—3 Numerous studies of healing offractures have been described histologically andultra-structurally. However, there is less informa-tion on the healing process of fractures fixed withplates, particularly when the soft tissue has been
*Corresponding author. Tel.: +81-92-642-6447;fax: +81-92-642-6386.
E-mail address: [email protected](L. Rasubala).
stripped across a wide area. For oral and maxillofa-cial surgeons, knowledge of the mechanism of bonehealing is essential to minimise deleterious effectsof operations and expedite the healing process.Bone morphogenetic proteins (BMPs) were dis-
covered and named after they were recognised tobe crucial mediators of the unique ability of dem-ineralised, lyophilised bone to induce formationof bone when placed within the muscle tissue ofrabbits.4,5 The effects of BMPs have been char-acterised in vitro; they act as mitogenic, chemo-tactic, and morphogenetic factors.6—8 Numerousstudies have shown that BMP-2 and BMP-4 are ableto restore bony defects in animal experiments.9—13
In a previous study, we examined immuno-histochemically the expressions of BMP-2 andplatelet-derived growth factor-B during the heal-
0266-4356/$ — see front matter © 2004 The British Association of Oral and Maxillofacial Surgeons. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.bjoms.2004.02.020
316 L. Rasubala et al.
ing of non-plated fractures. We found that BMP-2was involved in all stages of healing and seemedto play an important part in the formation of in-tramembranous ossification and in the differen-tiation of osteoblasts and chondrocytes.14 In thepresent study we compared the healing processof plated and non-plated fractures, and studied itafter surgical trauma. We assumed that the differ-ences in the histological appearance of cases wouldbe reflected in the expression of cytokines dur-ing the various stages of healing. As our previousstudy as well as other studies indicated that BMP-2has osteoinductive potential during the healing offractures,9—14 we used this protein in the presentstudy as a marker of the healing process.
Material and methods
Animal
Seventy-two male Wistar rats aged 12 weeks (SLCCo., Ltd., Fukuoka, Japan) and weighing 320—350 gwere used in this study. The rats were maintainedin temperature-controlled room and given unre-stricted access to food and water. The experimen-tal procedures followed the guiding principles forthe care and use of animals described in the KyushuUniversity Journal of Animal Care.
Experimental design
The animals were divided into two groups: one inwhich the fracture site was immobilised by inter-nal fixation of a single plate with two 1mm micro-screws (Mathys osteosynthesis, Tokyo, Japan) and acontrol group of non-plated fractures; each groupcomprised 36 rats. The animals were killed on days3 (n = 16), 7 (n = 16), 14 (n = 16), 21 (n = 12) and28 (n = 12) after the fracture. Radiographs weretaken to confirm the state of each fracture site.
Operation
The animals were anaesthetised intraperitoneallywith pentobarbitone 40mg/kg (Nembutal, AbbotLaboratories, Chicago, IL, USA), and the right sideof the mandible was prepared for aseptic opera-tion by an extra-oral approach. The skin overlyingthe right mandible was shaved and a 1 cm incisionmade. We split the masseter muscle in the directionof its fibres to expose the mandible, and a 0.3mmdiameter drill was used to make a smooth grooveabout 3—5mm behind the incisal root while it wasirrigated continuously with saline solution. A verti-
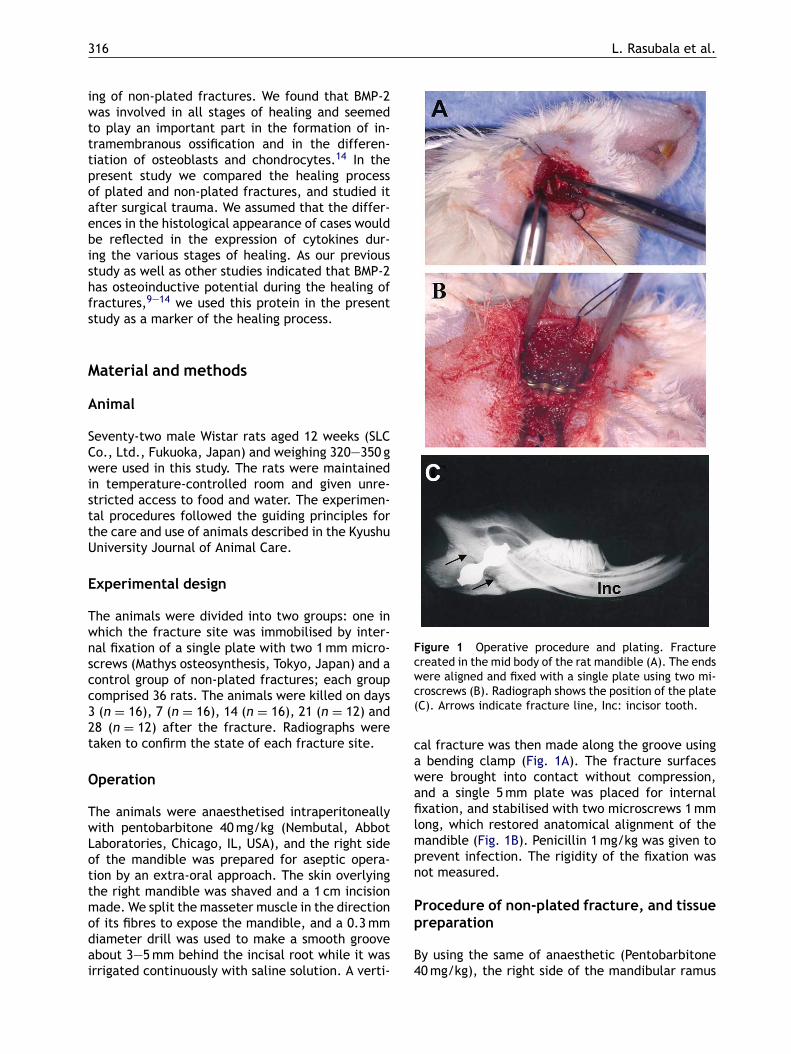
Figure 1 Operative procedure and plating. Fracturecreated in the mid body of the rat mandible (A). The endswere aligned and fixed with a single plate using two mi-croscrews (B). Radiograph shows the position of the plate(C). Arrows indicate fracture line, Inc: incisor tooth.
cal fracture was then made along the groove usinga bending clamp (Fig. 1A). The fracture surfaceswere brought into contact without compression,and a single 5mm plate was placed for internalfixation, and stabilised with two microscrews 1mmlong, which restored anatomical alignment of themandible (Fig. 1B). Penicillin 1mg/kg was given toprevent infection. The rigidity of the fixation wasnot measured.
Procedure of non-plated fracture, and tissuepreparation
By using the same of anaesthetic (Pentobarbitone40mg/kg), the right side of the mandibular ramus

Comparison of the healing process 317
was fractured with a bending clamp without an in-cision.On days 3, 7, 14, 21, and 28 after the opera-
tion, both groups of animals were killed by deepether anaesthesia. The fractured mandibles weredissected and fixed in 4% paraformaldehyde (SigmaChemical, St. Louis, MO, USA) overnight at 4 ◦C,decalcified in 20% ethylene diamine tetra-aceticacid tetrasodium salt, tetrahydrate (Dojin, Ku-mamoto, Japan) in 0.1M Tris—hydrochloric acid(Sigma Chemical), adjusted to a pH of 7.4. After7—10 days the specimens were placed in 20% su-crose solution overnight. Specimens were trimmedand embedded in tissue-freezing medium (SakuraFinetechnical, Tokyo, Japan) and kept at −80 ◦Cuntil they were cryosectioned. Specimens weresectioned sagittally (5�m thick) using a Leicacryostat (Nussloch, Germany). A total of 150 sec-tions was prepared from each specimen. Sectionswere placed on glass slides that were coated with3-aminopropyltriethoxysilane (Sigma Chemical),and stained with haematoxylin and eosin. The ex-pression of BMP-2 was detected by an immunohis-tochemical technique.
Antibody
Affinity-purified goat polyclonal anti-BMP-2 anti-body was bought from Santa Cruz Biotech (SantaCruz, CA, USA). Specificity was checked by peptideblocking experiments and immunoblotting with pu-rified recombinant BMPs. Only BMP-2 and BMP-4reacted with this antibody.
Immunohistochemistry
Sections from all 72 animals were rehydratedwith phosphate buffered saline, and endogenousperoxidase activity was blocked with 0.3% hydro-gen peroxide in absolute methanol for 20min atroom temperature. Non-specific protein bindingwas blocked by incubation with 5% normal don-key serum (Chemicon International Inc., Temecula,CA, USA) for 30min at room temperature. Thepolyclonal anti-BMP-2 goat antibody (1:100 dilu-tion), was incubated with the slides in a humidi-fied chamber at 4 ◦C. After they had been washedwith phosphate buffered saline, the slides wereincubated with a biotinylated secondary antibody(Jackson ImmunoResearch, PA, USA) for 1 h andan avidin—biotin peroxidase complex (DAKO, Ky-oto, Japan) for 30min. Finally the sections weredeveloped with 3,3′-diaminobenzidine (DAB sub-strate kit, Vector Lab, CA, USA), counterstainedwith haematoxylin and examined under a lightmicroscope. Normal goat IgG (Jackson ImmunoRe-
search) was used instead of the primary antibodyas a negative control.
Results
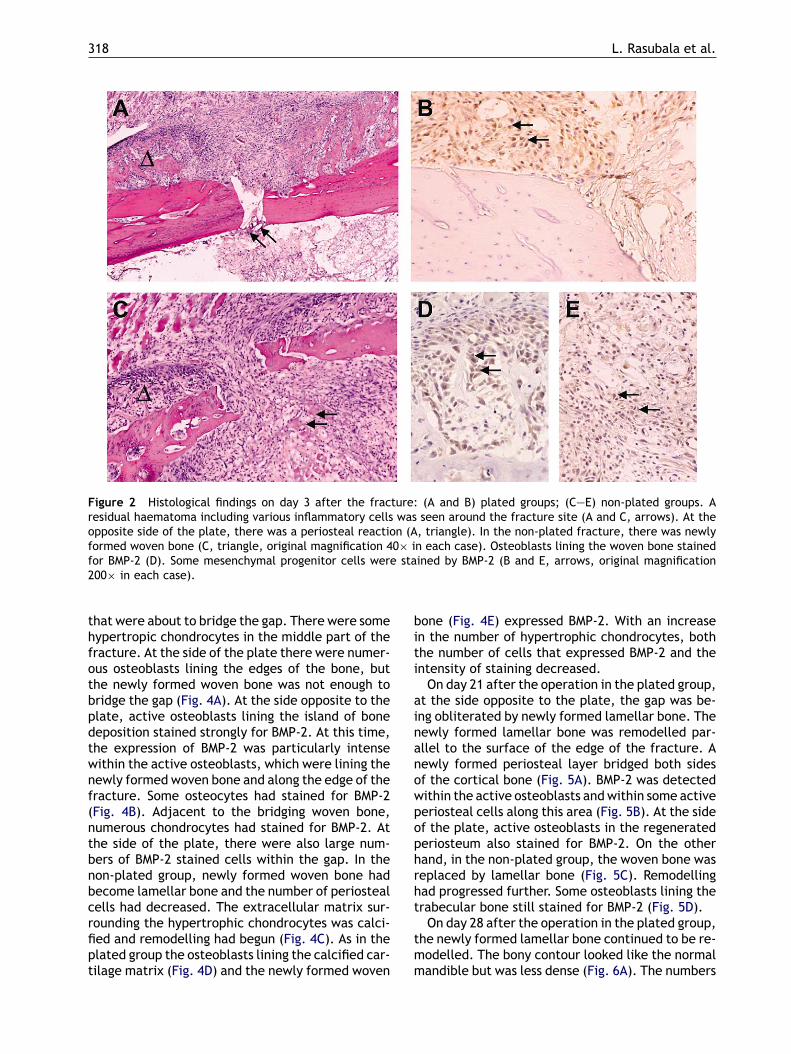
On day 3 after the operation in the plated group,there was a gap between the cortical bone and thesoft tissue. At the plated side, there was a smallgap between the fractured ends, and a slight resid-ual haematoma. There was no periosteal reactionat the plated side. On the side of the bone inter-nal to the plate, a number of mesenchymal cellshad gathered around the fracture ends, and therewas periosteal reaction with the proliferation of pe-riosteal cells (Fig. 2A). In contrast, the fractures inthe non-plated group were characterised by a thick-ened periosteum with proliferation of periostealcells proximal and distal to the fracture site. Therewas a considerable haematoma and various inflam-matory cells and numerous primitive mesenchymalcells (Fig. 2C). BMP-2 was identified in the primitivemesenchymal cells around the fracture site (Fig. 2Band E) and in the thickened periosteum (Fig. 2D).On day 7 after the operation in the plated
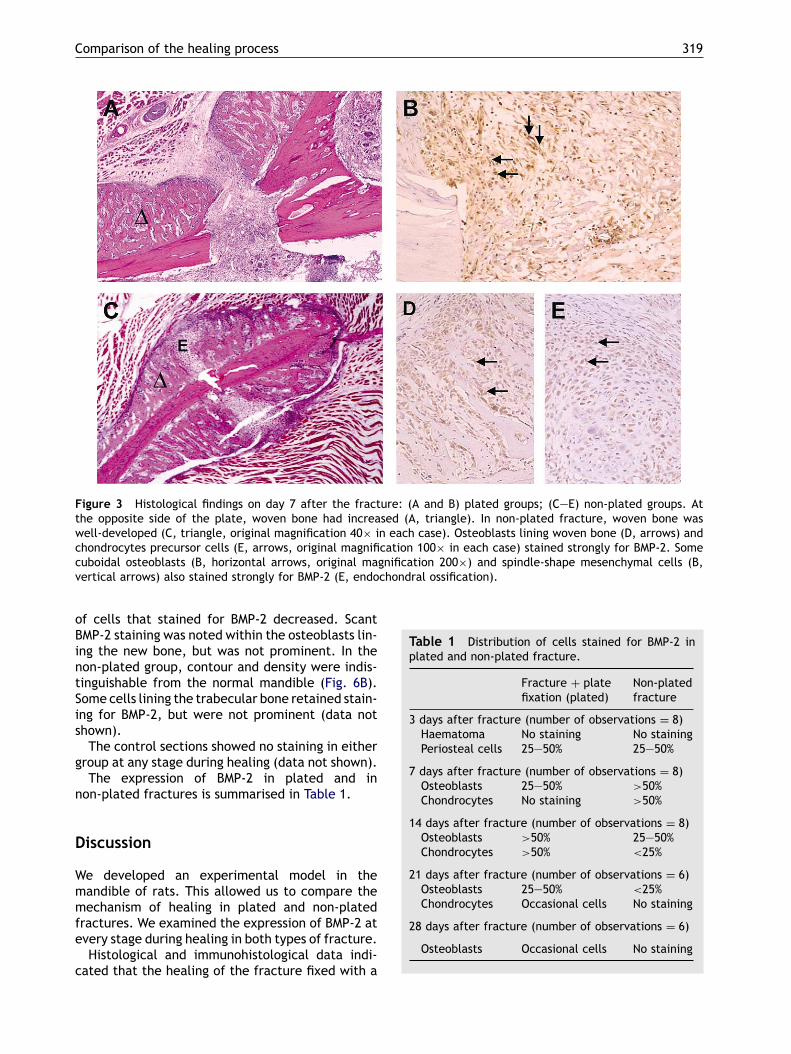
group the haematoma had disappeared, but somenecrotic connective tissue still remained. On theplated side there was still no periosteal reaction.However, a periosteal reaction was seen near thearea where the soft tissue had not been stripped.At the side opposite to the plate, numerous mes-enchymal cells had migrated from the thickenedperiosteum toward the fractured gap and prolifer-ated. Advanced intramembranous ossification wasseen sub-periosteally near the bony ends withoutintermediate cartilage. There were a large numberof osteoblasts lining this immature bone (Fig. 3A).BMP-2 was strongly expressed in cuboidal-shape os-teoblasts lining the newly formed woven bone andalso in the population of spindle-shaped mesenchy-mal cells, which proliferated from the periosteumalong the fractured edge toward the gap (Fig. 3B).In the non-plated group, there were a large numberof osteoblasts that lined the newly formed bone,corresponding to the intramembranous ossificationarea. Endochondral ossification developed fromthe initial haematoma through formation of carti-lage and bridged the fracture gap (Fig. 3C). BMP-2was strongly detected in the cuboidal-shaped os-teoblasts (Fig. 3D), as seen on side opposite to theplate in the plated group. In the area of endochon-dral ossification, precursor cells of chondrocytesalso stained prominently for BMP-2 (Fig. 3E).On day 14 after the operation in the plated group,
at the side opposite to the plate, newly formed wo-ven bone was accompanied by abundant osteoblasts
318 L. Rasubala et al.
Figure 2 Histological findings on day 3 after the fracture: (A and B) plated groups; (C—E) non-plated groups. Aresidual haematoma including various inflammatory cells was seen around the fracture site (A and C, arrows). At theopposite side of the plate, there was a periosteal reaction (A, triangle). In the non-plated fracture, there was newlyformed woven bone (C, triangle, original magnification 40× in each case). Osteoblasts lining the woven bone stainedfor BMP-2 (D). Some mesenchymal progenitor cells were stained by BMP-2 (B and E, arrows, original magnification200× in each case).
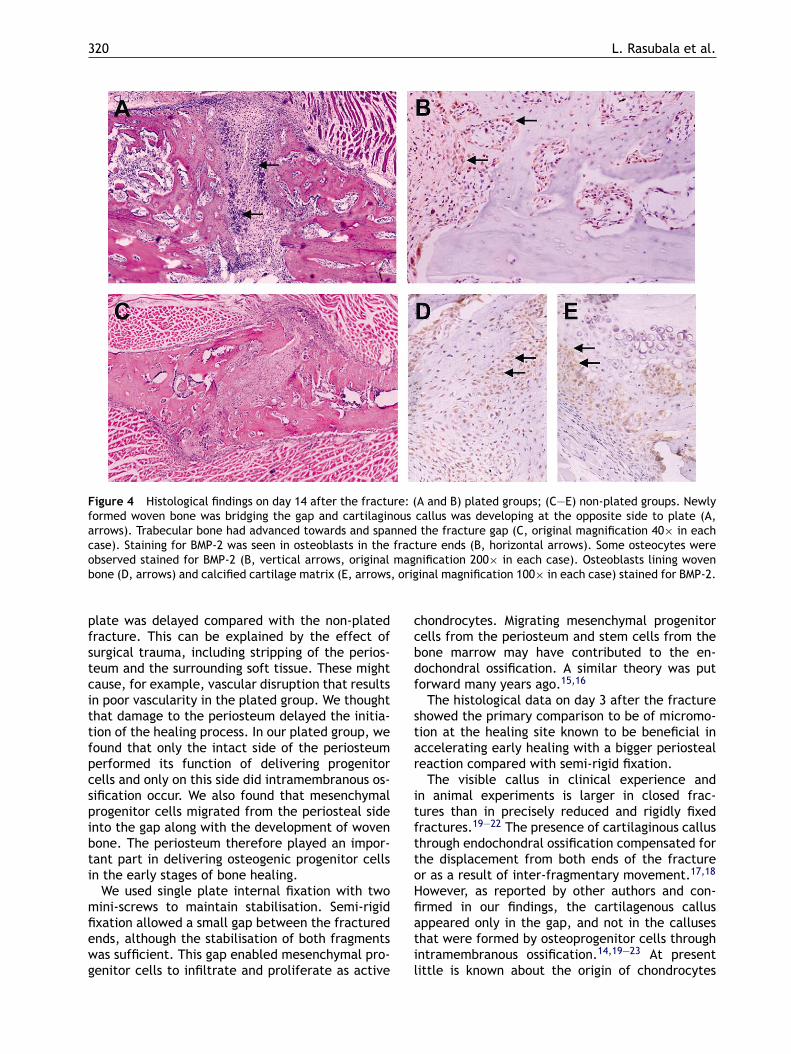
that were about to bridge the gap. There were somehypertropic chondrocytes in the middle part of thefracture. At the side of the plate there were numer-ous osteoblasts lining the edges of the bone, butthe newly formed woven bone was not enough tobridge the gap (Fig. 4A). At the side opposite to theplate, active osteoblasts lining the island of bonedeposition stained strongly for BMP-2. At this time,the expression of BMP-2 was particularly intensewithin the active osteoblasts, which were lining thenewly formed woven bone and along the edge of thefracture. Some osteocytes had stained for BMP-2(Fig. 4B). Adjacent to the bridging woven bone,numerous chondrocytes had stained for BMP-2. Atthe side of the plate, there were also large num-bers of BMP-2 stained cells within the gap. In thenon-plated group, newly formed woven bone hadbecome lamellar bone and the number of periostealcells had decreased. The extracellular matrix sur-rounding the hypertrophic chondrocytes was calci-fied and remodelling had begun (Fig. 4C). As in theplated group the osteoblasts lining the calcified car-tilage matrix (Fig. 4D) and the newly formed woven
bone (Fig. 4E) expressed BMP-2. With an increasein the number of hypertrophic chondrocytes, boththe number of cells that expressed BMP-2 and theintensity of staining decreased.On day 21 after the operation in the plated group,
at the side opposite to the plate, the gap was be-ing obliterated by newly formed lamellar bone. Thenewly formed lamellar bone was remodelled par-allel to the surface of the edge of the fracture. Anewly formed periosteal layer bridged both sidesof the cortical bone (Fig. 5A). BMP-2 was detectedwithin the active osteoblasts andwithin some activeperiosteal cells along this area (Fig. 5B). At the sideof the plate, active osteoblasts in the regeneratedperiosteum also stained for BMP-2. On the otherhand, in the non-plated group, the woven bone wasreplaced by lamellar bone (Fig. 5C). Remodellinghad progressed further. Some osteoblasts lining thetrabecular bone still stained for BMP-2 (Fig. 5D).On day 28 after the operation in the plated group,
the newly formed lamellar bone continued to be re-modelled. The bony contour looked like the normalmandible but was less dense (Fig. 6A). The numbers
Comparison of the healing process 319
Figure 3 Histological findings on day 7 after the fracture: (A and B) plated groups; (C—E) non-plated groups. Atthe opposite side of the plate, woven bone had increased (A, triangle). In non-plated fracture, woven bone waswell-developed (C, triangle, original magnification 40× in each case). Osteoblasts lining woven bone (D, arrows) andchondrocytes precursor cells (E, arrows, original magnification 100× in each case) stained strongly for BMP-2. Somecuboidal osteoblasts (B, horizontal arrows, original magnification 200×) and spindle-shape mesenchymal cells (B,vertical arrows) also stained strongly for BMP-2 (E, endochondral ossification).
of cells that stained for BMP-2 decreased. ScantBMP-2 staining was noted within the osteoblasts lin-ing the new bone, but was not prominent. In thenon-plated group, contour and density were indis-tinguishable from the normal mandible (Fig. 6B).Some cells lining the trabecular bone retained stain-ing for BMP-2, but were not prominent (data notshown).The control sections showed no staining in either
group at any stage during healing (data not shown).The expression of BMP-2 in plated and in
non-plated fractures is summarised in Table 1.
Discussion
We developed an experimental model in themandible of rats. This allowed us to compare themechanism of healing in plated and non-platedfractures. We examined the expression of BMP-2 atevery stage during healing in both types of fracture.Histological and immunohistological data indi-
cated that the healing of the fracture fixed with a
Table 1 Distribution of cells stained for BMP-2 inplated and non-plated fracture.
Fracture + platefixation (plated)
Non-platedfracture
3 days after fracture (number of observations = 8)Haematoma No staining No stainingPeriosteal cells 25—50% 25—50%
7 days after fracture (number of observations = 8)Osteoblasts 25—50% >50%Chondrocytes No staining >50%
14 days after fracture (number of observations = 8)Osteoblasts >50% 25—50%Chondrocytes >50% <25%
21 days after fracture (number of observations = 6)Osteoblasts 25—50% <25%Chondrocytes Occasional cells No staining
28 days after fracture (number of observations = 6)
Osteoblasts Occasional cells No staining
320 L. Rasubala et al.
Figure 4 Histological findings on day 14 after the fracture: (A and B) plated groups; (C—E) non-plated groups. Newlyformed woven bone was bridging the gap and cartilaginous callus was developing at the opposite side to plate (A,arrows). Trabecular bone had advanced towards and spanned the fracture gap (C, original magnification 40× in eachcase). Staining for BMP-2 was seen in osteoblasts in the fracture ends (B, horizontal arrows). Some osteocytes wereobserved stained for BMP-2 (B, vertical arrows, original magnification 200× in each case). Osteoblasts lining wovenbone (D, arrows) and calcified cartilage matrix (E, arrows, original magnification 100× in each case) stained for BMP-2.
plate was delayed compared with the non-platedfracture. This can be explained by the effect ofsurgical trauma, including stripping of the perios-teum and the surrounding soft tissue. These mightcause, for example, vascular disruption that resultsin poor vascularity in the plated group. We thoughtthat damage to the periosteum delayed the initia-tion of the healing process. In our plated group, wefound that only the intact side of the periosteumperformed its function of delivering progenitorcells and only on this side did intramembranous os-sification occur. We also found that mesenchymalprogenitor cells migrated from the periosteal sideinto the gap along with the development of wovenbone. The periosteum therefore played an impor-tant part in delivering osteogenic progenitor cellsin the early stages of bone healing.We used single plate internal fixation with two
mini-screws to maintain stabilisation. Semi-rigidfixation allowed a small gap between the fracturedends, although the stabilisation of both fragmentswas sufficient. This gap enabled mesenchymal pro-genitor cells to infiltrate and proliferate as active
chondrocytes. Migrating mesenchymal progenitorcells from the periosteum and stem cells from thebone marrow may have contributed to the en-dochondral ossification. A similar theory was putforward many years ago.15,16
The histological data on day 3 after the fractureshowed the primary comparison to be of micromo-tion at the healing site known to be beneficial inaccelerating early healing with a bigger periostealreaction compared with semi-rigid fixation.The visible callus in clinical experience and
in animal experiments is larger in closed frac-tures than in precisely reduced and rigidly fixedfractures.19—22 The presence of cartilaginous callusthrough endochondral ossification compensated forthe displacement from both ends of the fractureor as a result of inter-fragmentary movement.17,18
However, as reported by other authors and con-firmed in our findings, the cartilagenous callusappeared only in the gap, and not in the callusesthat were formed by osteoprogenitor cells throughintramembranous ossification.14,19—23 At presentlittle is known about the origin of chondrocytes
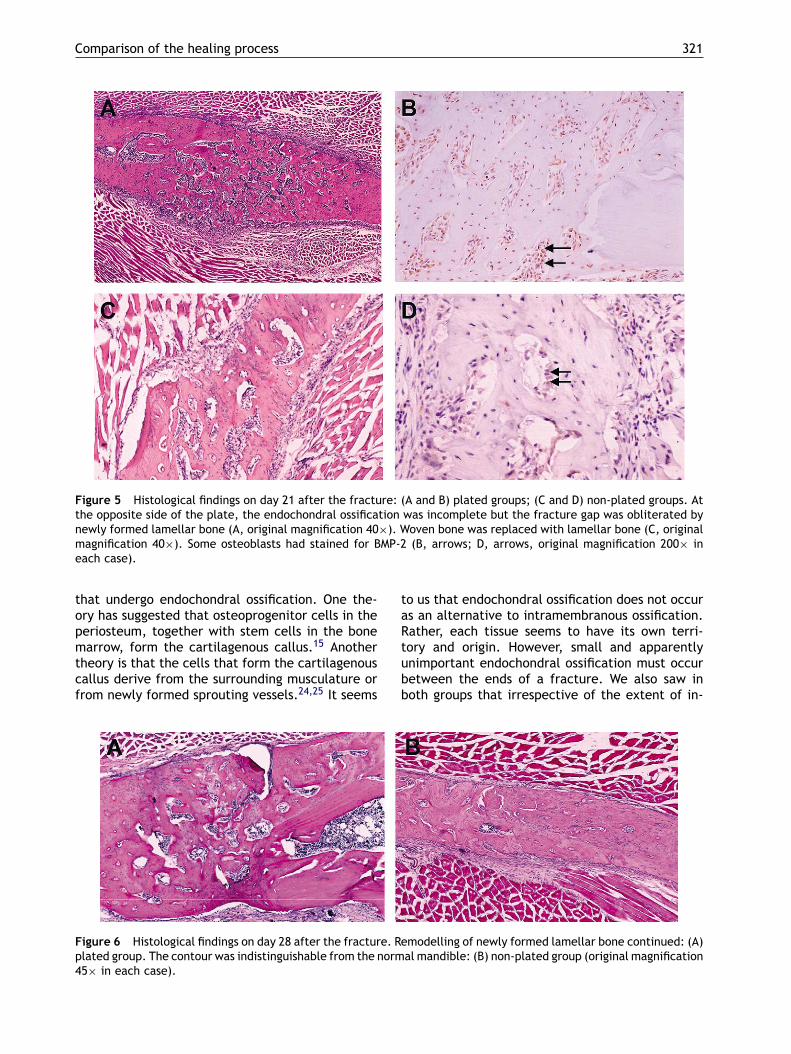
Comparison of the healing process 321
Figure 5 Histological findings on day 21 after the fracture: (A and B) plated groups; (C and D) non-plated groups. Atthe opposite side of the plate, the endochondral ossification was incomplete but the fracture gap was obliterated bynewly formed lamellar bone (A, original magnification 40×). Woven bone was replaced with lamellar bone (C, originalmagnification 40×). Some osteoblasts had stained for BMP-2 (B, arrows; D, arrows, original magnification 200× ineach case).
that undergo endochondral ossification. One the-ory has suggested that osteoprogenitor cells in theperiosteum, together with stem cells in the bonemarrow, form the cartilagenous callus.15 Anothertheory is that the cells that form the cartilagenouscallus derive from the surrounding musculature orfrom newly formed sprouting vessels.24,25 It seems
Figure 6 Histological findings on day 28 after the fracture. Remodelling of newly formed lamellar bone continued: (A)plated group. The contour was indistinguishable from the normal mandible: (B) non-plated group (original magnification45× in each case).
to us that endochondral ossification does not occuras an alternative to intramembranous ossification.Rather, each tissue seems to have its own terri-tory and origin. However, small and apparentlyunimportant endochondral ossification must occurbetween the ends of a fracture. We also saw inboth groups that irrespective of the extent of in-

322 L. Rasubala et al.
tramembranous ossification at the point of bridgingthe gap endochondral ossification took place at theborder of the bony fragments.The healing process in the plated groups was
delayed by a week. BMP-2 was expressed at allstages in both groups with different levels of in-tensity. BMP-2 seems to be a useful indicator offracture healing, correlating with the differen-tiation of mesenchymal cells into osteoblasts orchondrocytes, and acting in autocrine or paracrinemanner.In conclusion, the natural healing process was dis-
turbed in the plated group. The intact periosteumwas the main source of osteogenic progenitor cells.BMP-2 was one of important regulatory factors ofthe healing process in both groups.
References
1. Hulth A. Current concepts of fracture healing. Clin Orthop1989;249:265—84.
2. Brand RA, Rubin CT. Fracture healing. In: Albright JA, BrandRA, editors. The scientific basis of orthopaedics. 2nd ed.Norwalk: Appleton & Lange; 1987. p. 325–46.
3. Grundnes O, Reikeras O. The importance of thehematoma for fracture healing in rats. Acta Orthop Scand1993;63:340—2.
4. Van de Putte KA, Urist MR. Osteogenesis in the interiorof intramuscular implants of decalcified bone matrix. ClinOrthop 1965;43:257—70.
5. Urist MR. Bone: formation by autoinduction. Science1965;150:893—9.
6. Sakou T. Bone morphogenetic proteins: from basic studiesto clinical approaches. Bone 1998;22:591—603.
7. Yang S, Wei D, Wang D, et al. In vitro and in vivo syn-ergistic interactions between the Runx2/Cbfa1 transcrip-tion factor and bone morphogenetic protein-2 in stimulat-ing osteoblast differentiation. Bone Miner 2003;18:705—15.
8. Suzuki T, Bessho K, Segami N, Iizuka T, Nojima T. Immuno-histochemical localization of bone morphogenetic protein-2in the oral and maxillofacial area of the human embryo. BrJ Oral Maxillofac Surg 2001;39:289—93.
9. Gerhart TN, Kirker-Head CA, Kriz MJ, Holtrop ME, HennigGE, Hipp J, et al. Healing segmental femoral defects insheep using recombinant human bone morphogenetic pro-tein. Clin Orthop 1993;293:317—26.
10. Seto I, Asahina I, Oda M, Enomoto S. Reconstruction ofprimate mandible by the combination graft of recombinanthuman bone morphogenetic protein-2 (rhBMP-2) and bonemarrow. J Oral Maxillofac Surg 2001;59:53—61.
11. Boyne PJ, Nath A, Nakamura A. Human recombinant BMP-2in osseus reconstruction of stimulated cleft palate defects.Br J Oral Maxillofac Surg 1998;36:84—90.
12. Hecht BP, Fischgrund JS, Herkowitz HN. The use of recom-binant human bone morphogenetic protein 2 (rhBMP-2) topromote spinal fusion in a nonhuman primate anterior in-terbody fusion model. Spine 1999;24:629—36.
13. Spector JA, Luchs JS, Mehrara BJ, Greenwald JA, SmithLP, Longaker MT. Expression of bone morphogenetic pro-teins during membranous bone healing. Plast Reconstr Surg2000;107:124—34.
14. Rasubala L, Yoshikawa H, Nagata K, Iijima T, Ohishi M.Platelet-derived growth factor and bone morphogeneticprotein in the healing of mandibular fractures in rats. Br JOral Maxillofac Surg 2003;41:173—8.
15. Kernek CB, Wray JB. Cellular proliferation in the formationof fracture callus in the rat tibia. Clin Orthop 1973;91:197—209.
16. Owen M. The origin of bone cells in the postnatal organism.Arthritis Rheum 1980;23:1073—80.
17. Park SH, O’Connor K, McKellop H, Sarmiento A. The in-fluence of active shear or compressive motion on fracturehealing. J Bone Joint Surg Am 1998;80:868—78.
18. Sarmiento A, Mullis DL, Latta LL, Tarr RR, Alvarez R. A quan-titative comparative analysis of fracture healing under theinfluence of compression plating vs closed weight-bearingtreatment. Clin Orthop 1980;149:232—9.
19. Molster A, Gjerdt NR, Raugstad TS, et al. Effect of insta-bility on experimental fracture healing. Acta Orthop Scand1982;53:521—6.
20. Park SH, O’Conner K, Sung R, McKellop H. Comparison ofhealing process in open osteotomy model and closed frac-ture model. J Orthop Trauma 1999;13:114—20.
21. Si X, Jin Y, Tipoe GL, White FH. Expression of BMP-2 andTGF-�1 mRNA during healing of the rabbit mandible. Eur JOral Sci 1997;105:325—30.
22. Henricson A, Hulth A, Johnell O. The cartilaginous fracturecallus in rats. Acta Orthop Scand 1987;57:244—8.
23. Onishi T, Ishidou Y, Nagamine T, Yone K, Imamura T, KatoM, et al. Distinct and overlapping pattern of localization ofbone morphogenetic protein (BMP) family members and aBMP type II receptor during fracture healing in rats. Bone1988;22:605—12.
24. McLean FC, Urist MR. Bone: fundamentals of physiology ofskeletal tissue. 3rd ed. Chicago: Chicago University Press;1968. p. 5–12.
25. Nathanson MA. Bone matrix-directed chondrogenesis ofmuscle in vitro. Clin Orthop 1985;200:142—58.





























