Embed Size (px)
Citation preview

Indian Journal of Experimental Biology Vol. 37, January 1999, pp. 61-65
A comparative i n·.;e~· j· :gation of differential scanning calorimetric transitions of Orevolving photosystem II enriched membranes isolated
from beet spinach (Beta vulgaris palanga) leaves and greened cucumber (Cucumis sativus) cotyledons
[ B Jha· , Sujata R Mishra#, Rajiv Bhat· & Prasanna Mohanty··
· Centre For Biotechnology, Jawaharlal Nehru University, New Delhi 110067, India
··School of Life Sciences, Jawaharlal Nehru University, New Delhi 110067, India
Received 6 August 1998; revised 9 October 1998
PS !I is a multicomponent thylakoid membrane protein complex which converts solar radiation into chemical potential. PS II enriched membrane preparations lose their oxygen evolution activity upon thermal denaturation of their associated proteins. PS II enriched membranes were prepared from beet spinach (Beta vulgaris palanga) leaves, and 6 day and 20 day oid greened cotyledons of Cucumis salivus. SDS-PAGE profiles of these rS II preparations are similar and devoid of any PS I contamination. The thermal denaturation of PS II enriched membrane preparations from beet spinach and greened C. sativus cotyledons were monitored by Differential Scanning Calorimetry (DSC). The DSC profiles of PS II preparations from spinach gave rise to the characteristic five endothermic transitions (A" Az, B, C and D) in the 30°-70°C temperature range whereas in the greened cotelydons of C. sativus peak AI was not resolved. However, the other four transitions are similar although we observed a considerable shift in the endothermic transition peaks of PS II preparations from both 6 and 20 day old greened cucumis cotyledons compared to that from spinach leaves. The highest transition peaks (the C, 0 peaks) of PS II enriched membrane preparations were observed at 62.9°, 66.8°C and 67.7°C for .beet spinach, 6 and 20 day old greened cucumis cotyledons, respectively. The difference in the position of highest transition peaks indicates the presence of better heat stability in the PS II protein system in the greened cotyledons of C. sativus.
Photosystem CPS) II IS a multi-component thylakoid membrane protein complex which uti lizes light energy to drive uphill electron transport reactions and oxidizes wate r. Several Or evolving photosystem II (PS II) enriched thylakoid membranes such as BBY, K&M and YUSN have been prepared in various laboratories.I-) These PS II enriched membranes have been well characterized and are used for the study of PS II structure and func tion4
. These PS II preparations lose activity with increasing temperature because of thermal perturbation of PS II associated proteins. Thermal denaturation studies of PS II have provided insight into the structural organization of the multimeric protein complex and the probable roies of the component proleins5
. In recent years,
*·Address for correspondence: School of Life Sciences, lawaharlal Nehru Un iversity, l\'ew Deihi ! 10 067_ India #Present address: Institute of Li fe Sciences, Bhubaneswar 7S I 007. India
Differential Scanning Calorimetry (DSC) has been used to study association of com plex, multicomponent membrane systems6
-9
, so as to obtain information about the stabil ity imd interactions of specific components in the system. DSC has also been used to study the thermal transitions of thylakoid membranes 10. The thermal denaturation profiie of the PS II membrane protein complex from Bela palanga (spinach) has been examined using Differential Scanning Caiorimetr/ ,II. For complex systems a peak in the DSC trace usually corresponds to the cooperative unfolding of a protein domain. Cotyledonary leaves of pumpkins, cucumber etc . have been used as model systems to monitor the etiop last to chlorop last transit ion in light. It is of interest to probe the developmental aspects of th is conversion using PS II enriched O2
evolving membranes from coty ledonary leaves . The thermal denaturation profiles obtained from DSC can give an idea about the natu re of the

62 INDIAN J EXP BlOL, JANUARY 1999
associated proteins during the entire course of development of chloroplast and thus, can be used as potential probe in such studies.
The present work is aimed at preparing O2-evolving PS II enriched membranes (PS II) from greened cotyledons of Cucumis and comparing it with that of beet spinach (Beta vulgaris palanga) PS II enriched membranes with respect to their thermal stabi lity. Furthermore, DSC has been used to obtain the thermal denaturation profiles of the PS II enriched membranes isolated from beet spinach, and 6 day and 20 day old greened cucumber cotyledons. The differential nature of DSC transitions obtained is analysed and discussed .
Materials and Methods lsolation of 0] evolving PS II enriched
membranes-PS II enriched membranes were prepared according to the procedures of Berthold et. al.1 and Van Leeuwen et. al. 12
, albeit with certain modifications. Healthy spinach leaves were grown in an experimental garden and Cucumber (Cucumis sativus, var. Poinsette) was grown as described in ref. 13 . Leaves or greened cotyledons were homogenized in ice-cold 20 mM MES-NaOH buffer (PH 6.0), 15 mM NaCI, and 5 mM CaCho The homogenate was filtered through 4 layers of Mira cloth . The filtrate was centrifuged and carefully washed thrice in grinding buffer [20 mM MES NaOH (PH 6.0), 15 mM NaCI and 5 mM CaCI2] at 5000 g for 5 min . Triton-treated thylakoids were centrifuged at 40,000 g for 30 min ., washed (at least 4 times) and resuspended in 20 mM MES NaOH (PH 6.0), 15 mM NaCI and 0.4 M sucrose and centrifuged again at 5000 g for 5 min. and finally at 40,000 g for 30 min. At the end, the PS II enriched fraction was suspended in 20 mMMES-NaOH buffer (PH 6.5), 20 mM MgCI2, 5 mM CaCI2, 10 mM MgS04, 0.4 M sucrose and stored at -80°C. ChI was estimated according to Porra el. al. 14 with dimethyl formam ide as solvent. O2 evolution activity of PS II enriched membranes was estimated polarographically wi th dichlorobenzoquinone (DCBQ) as the electron acceptor l 5
.
The PS 11 enriched preparation from Cucumis yie lded 260-280 ' Ilmo le Oi mg Chl/hr and, in the case of spinach , the PS II enriched preparations yie lded approximately 400 Ilmo le 0 2/mg Chl/hr.
Protein separation--SDS PAGE ( 12% and 10-15% gradient) were carried out according to the methods ofLaemmeli l6
.
Differential scanning calorimet~ The DSC experiments were conducted in a Microcal MC-2 Differential Scanning Calorimeter at a scan rate of 1°C/min. For all DSC measurements concentrations were 1.12 mg of Chl/mt. The scan was recorded between 10° and 90°C, however al l the transitions were observed in the range of 30°-70°C. Three sets of DSC experiments were carried out for a given condition and the scans were found to be highly reproducible. For each condit ion the PS II membranes were' isolated three times and DSC scans were obtained for each PS II preparations. The data were analysed by using Origin ™ software from Microcal, Inc. For all the calorimetric calculations, concentrations of the samples were converted into ' mi ll imoles' by approximating the molecular weight as 324±64 kDa for the PS II membrane prote in complex.
Results and Discussion In all of our PS II enriched membrane
preparations from spinach leaves and Cucumis cotyledons , the ChI alb ratio was the measure of purity. Samples of PS II enriched membranes with Chi alb ratio 1.7 and below were found to be pure and completely devo id of PS 1. All PS II enriched membrane preparations showed 0 2-evo lution activity indicating thereby the presence of active 02-evolving complexes.
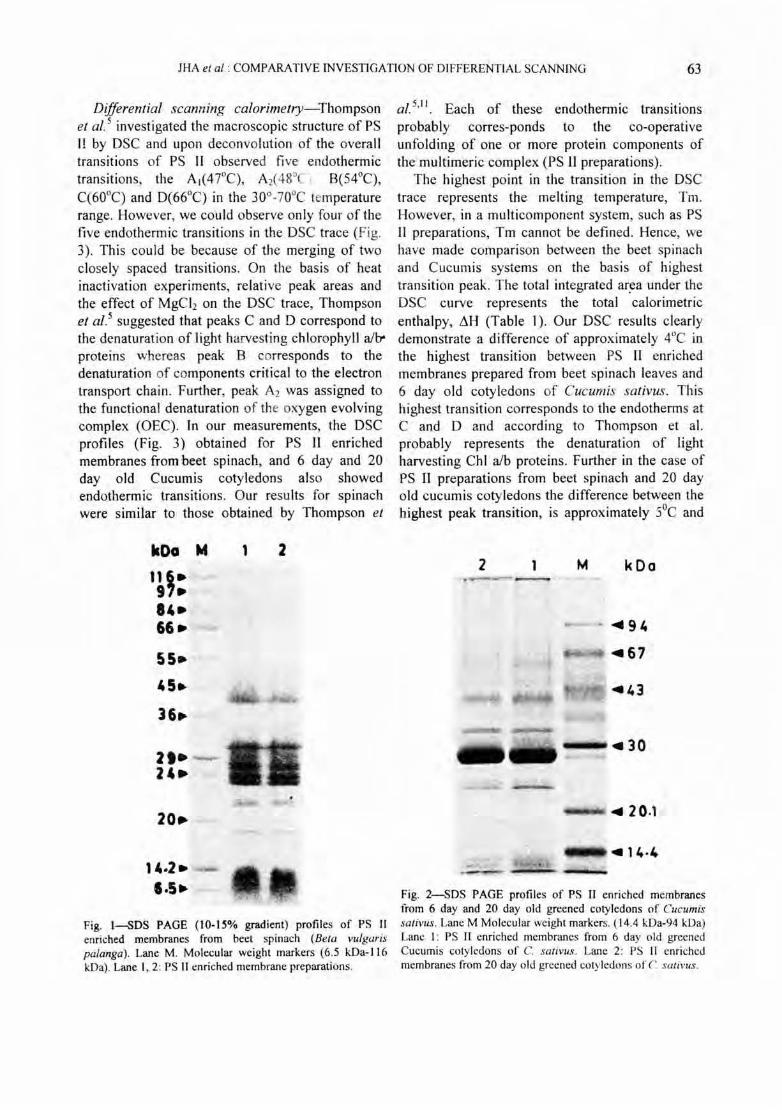
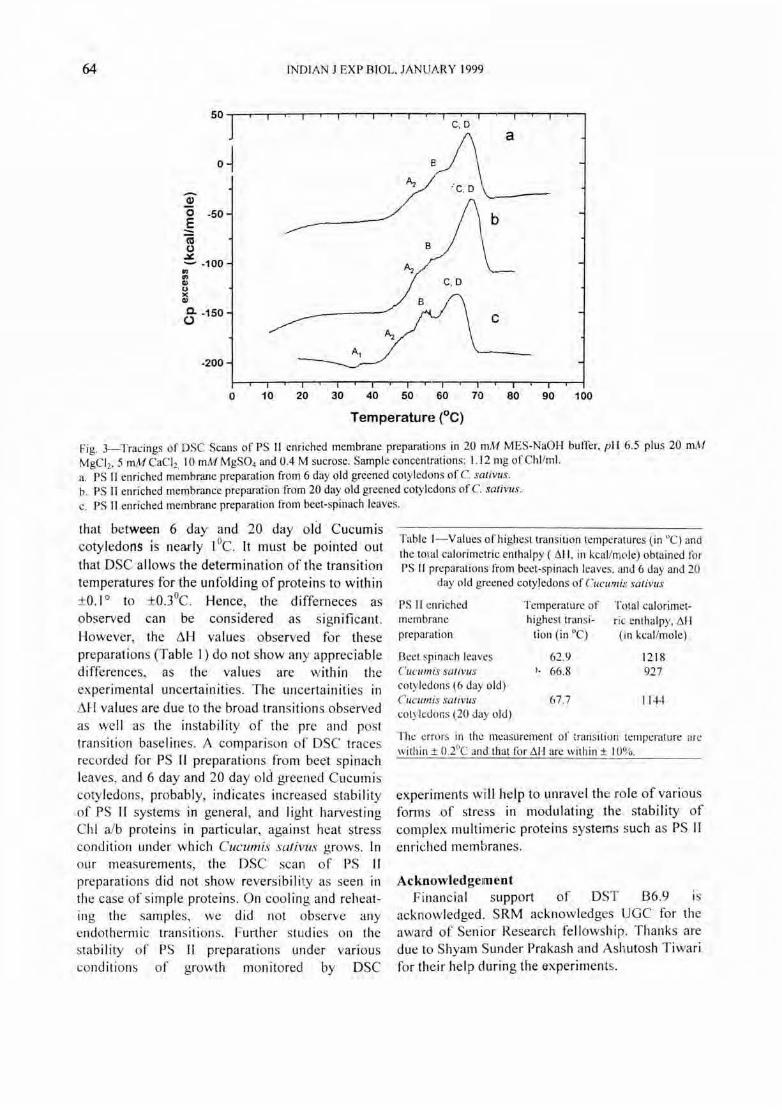
SDS-PAGE profile-The isolated PS II enriched membranes from beet spinach leaves were run on a 10-1 5% linear gradient SDS-PAGE (Fig. I) whereas PS II enriched membranes from greened 6 day and 20 day old Cucumis salivus coty ledons were run on 12% polyacry lamide ge l (F ig.2). SDSPAGE profil es of PS II enriched membranes from spinach leaves and Cucumi s cotyledons were devo id of any PS I contam ination. The SDS PAGE profiles of all the PS II enriched membrane preparations showed the presence of ChI alb protein, LHC at 24-25 kDa l7
, PS II react ion centre proteins at 45 kDa 18, 19 and a prominent band at 32 kDa20
. The two polypeptides at 17 and 22 kDa are probably the extr insic lumenal proteins assoc iated with the oxygen evo lving mechanism21-23 .
JHA el al.: COMPARATIVE INVESTIGATION OF DIFFERENTIAL SCANNING 63
Differential scanning calorimetry-Thompson e/ al. 5 investigated the macroscopic structure of PS I! by DSC and upon deconvolution of the overall transitions of PS II observed fi ve endothermic transitions, the A I (47°C), A2( 48°('! B( 54°C), C(60°C) and D(66°C) in the 30°-70°C temperature range. However, we could observe only four of the five endothermic transitions in the DSC trace (Fig. 3). This could be because of the merging of two closely spaced transitions. On the basis of heat inactivation experiments, relative peak areas and the effect of MgCh on the DSC trace, Thompson et al.5 suggested that peaks C and 0 correspond to the denaturation of light harvesting chlorophyll albproteins whereas peak B corresponds to the denaturation of components critical to the electron transport chain. Further, peak A2 was assigned to the functional denaturation of the oxygen evolving complex (OEC). In our measurements, the DSC profiles (Fig. 3) obtained for PS II enriched membranes from beet spinach, and 6 day and 20 day old Cucumis cotyledons also showed endothermic transitions. Our results for spinach were similar to those obtained by Thompson et
liDo M
116-91-1'-66.
1 2
Fig. l-SDS PAGE (10-15% gradient) profiles of PS II enriched membranes from beet spinach (Bela vulgaris palanga). Lane M. Molecular weight markers (6 .5 kDa-116 kOa). Lane 1,2: PS II enriched membrane preparations.
al.5,11 . Each of these endothermic transitions
probably corres-ponds to the co-operative unfolding of one or more protein components of the multimeric complex (PS II preparations).
The highest point in the transition in the DSC trace represents the melting temperature, Tm. However, in a multicomponent system, such as PS II preparations, Tm cannot be defined. Hence, we have made comparison between the beet spinach and Cucumis systems on the basis of highest transition peak. The total integrated ar,ea under the DSC curve represents the total calorimetric enthalpy, ~H (Table I). Our DSC results clearly demonstrate a difference of approximately 4°C in the highest transition between PS II enriched membranes prepared from beet spinach leaves and 6 · day old cotyledons of Cucumis sativus. This highest transition corresponds to the endotherms at C and 0 and according to Thompson et al. probably represents the denaturation of light harvesting Chi alb proteins. Further in the case of PS II preparations from beet spinach and 20 day old cucumis cotyledons the difference between the highest peak transition, is approximately SoC and
2 1 M kDo
.1' 414.4 trrM' ) i j,
Fig. 2-SDS PAGE profiles of PS II enriched membranes from 6 day and 20 day old greened cotyledons of Cucumis salivus. Lane M Molecular weight markers. (14.4 kOa-94 kOa) Lane 1: PS II enriched membranes from 6 day old greened Cucumis cotyledons of C. salivus. Lane 2: PS 11 enriched membranes from 20 day old greened cotyledons of C. sa[ivus.
64 INDI AN J EX P BIOL, JAN UARY 1999
C, D
-Q)
(5 -50 .§ co o ,:t;
BJ\ a
~)'C~':-~ B \_ - -100 .,
'" Q) (J >< Q)
a. -150 o
-200
o
A,
10 20 30
c
--40 50 60 70 80 90 100
Fig. 3-Tracings of DSC Scans of PS " enriched membrane preparations in 20 mM MES-NaOH buffer, pH 6. 5 plus 20 mM MgCIz, 5 mM CaCI2, 10 mM MgS04 and 0.4 M sucrose. Sample concentrat ions: 1. 12 mg of Chllm l. a. PSI/ enriched membrane preparation from 6 day old greened cotyledons of C. sativ lIs. b. PSI/ enriched membrance preparation from 20 day old greened cotyledons of C. sativlIs .
c. PSI/ enriched membrane preparation from beet-spinach leaves.
that between 6 day and 20 day o ld Cucumis cotyledons is nearly 1°C. It must be pointed out
that DSC allows the determination of the transition temperatures for the unfolding of proteins to within
±O.l o to ±0.3°C. Hence, the differneces as observed can be considered as significant.
However, the L1 H values observed fo r these preparations (Table I) do not show any appreciable differences, as the values are w ithin the experimental uncertainities. The uncertainities in
L1H values are due to the broad trans iti ons o bserved as well as the instability of the pre and post trans iti on base lines. A comparison of DSC traces recorded fo r PS II preparations from beet spinach leaves, and 6 day and 20 day o ld greened Cucumis cotyledons, probably, indicates increased stability of PS II systems in general, and light harvesting Ch i al b proteins in particular, against heat stress condition under which Cucumis salivus grows. In o ur measurements, the DSC scan of PS 11 preparations did not show revers ibili ty as seen in th e case of s imple prote ins. On cooling and reheatin g the samples, we did not observe any endothermi c trans itions. Further studies o n the stability of PS II preparatio ns under various co nditions of growth monitored by DSC
Tab le I-Values of highest transi tion temperatures (in 0c) and
the toral calorimetric enthalpy ( ~H , in kcallmole) obtained for PS " preparati ons from beet-spinach leaves, and 6 day and 20
day o ld greened cotyledons of Cucllmis salivlIs
PS [I enriched Temperature of Total calorimet-membrane highest transi- ric enthalpy, ~H preparation tion (in 0c) (in kcal/mole)
Beet spinach leaves 62.9 12 18 Cllcllmis salivlIs ) . 66.8 927 coty ledons (6 day old) CuclImis salivlIs 67.7 I i 44 coty ledons (20 day old)
The errors in the measurement of transition temperature are with in ± 0.2°C and that for ~H are within + 10%.
experiments will help to unravel the ro le of various forms of stress in modulating the stability of complex multimeric proteins systems such as PS II enriched membranes ,
Acknowledgement Financial support of DST 86.9 IS
acknowledged. SRM acknowledges UGC for the award of Senior Research fel lowshi p. Thanks are due to Shyam Sunder Prakash and Ashutosh Ti wari for the ir help during the axperiments.

JHA et at.: COMPARATIVE INVESTGATON OF DIFFERENTIAL SCANNING 65
Referenc~s I Berthold D A, Babcock G T & Yocum C F, FEBS Lett.
134 (1981) 23 I. 2 Kuwabara T & Murata N, Plant Cell Physiol, 24 (1983)
741. 3 Yamamoto Y, Ueda T, Shinkai H & Nishimura M,
Biochim Biophys Acta. 679 ( 1982) 347. 4 Dunahay TG, Staehelin L A, Seibert M, Ogilvie P D &
Berg S P, Biochim Biophys Acta. 764 (1984) 179. 5 Thompson L K, Sturtevant J M & Brudvig G W
Biochemistry. 25 (1986) 6 161 . 6 Brandts J F, Erickson L, Lysko K, Schwartz T A &
Taverna R D, Biochemistry, 16 (1977) 3450. 7 Brandts J F, Taverna R D, Sadasivan E & Lysko K A,
Biochim Biophys Acta, 512 (1978) 566. 8 Lysko K A, Karlson R, Taverna R, Snow J & Brandts J F,
Biochemistry, 20 ( 1981 ) 5570. •
9 Haltia T, Semo N, Arrondo J L, Goni r M & Freire 1';:, Biochemistry, 33(32) ( ! 994) 9731 .
10 Cramer W A, Whi tmarsh J & Low P S, Biochemistry, 20 (1981) 157.
11 Thompson L K, Blaylock R, Sturtevant J M & Brudvig G W, Biochemistry, 28 (1989) 6686.
12 Van Leeuwen P J, Nieveen M L, Vantede Meent E J, Dekker J P & Van Garkom H J, Photosynth Res, 28 (1991) 149.
13 Prakash J S S, Baig M A & Mohanty P, Photosynthetica. ( 1998) (in press).'
14 Porra R J, Thompson W A & Kriedemann P E, Biochim Biophys Acta, 975 (1989) 384.
15 Mohanty P & Seibert M, Indian J Biochim. Biophys, 34 (1997) 241.
16 Laemmli U K Nature. 227 ( 1970) 680.
17 Thornber J P & Markwell J P, Trends Biochem Sci. 6 (19S1) 122.
18 Satoh K, Biochim Biophys Acta. 546 ( 1979) 84. 19 Green 0 R & Camm E L, J Cell Biochem, 7B (1983) 322.
20 Mullet J E & Arntzen C J, Biochim Biophys Acta, 635 ( 1981) 236.
21 Akerlund H E, Jansson C & Anderson B, Biochim Biophys Acta, 681 (1982) I.
22 Kuwabara T & Murata N, Plant Cell Physiol, 23 ( 1982) 533.
23 Toyoshima Y & Fukutaka E, FEBS Left , 150 (1982) 223.




![Pemphigus Vulgaris [Print] - eMedicine Dermatology Vulgaris .pdf · emedicine.medscape.com eMedicine Specialties > Dermatology > Bullous Diseases Pemphigus Vulgaris Bassam Zeina,](https://img.dokumen.tips/doc/110x75/5c984ab609d3f21c3a8b874e/pemphigus-vulgaris-print-emedicine-vulgaris-pdf-emedicinemedscapecom.jpg)
























