Embed Size (px)
Citation preview

FOTOSINTEZA
• Svetloba kot energija: E = h.√ = h.c/λ(400-700 nm); E = energija; √ =frekvenca;
• λ = val. dolžina svetlobe; h = Plankovakonstanta (6,625 .10 34 J/s); c = hitrostsvetlobe
• Enota : 1μmol s -1 m -2 = 1 μE s -1 m -2= 6,023 . 10 17 fotonov s -1 m -2=
• 6,023 .10 17 kvantov s -1 m -2

FOTOSINTEZA - ASIMILACIJA OGLJIKA (energetika celice
• RASTLINE - AVTOTROFNI ORGANIZMI višje rastline -brstnice;
• cianobakterije, fotosintetskoaktivnebakterije,
• evkariontske alge, • lišaji

KEMOSINTEZA• 1. Nitrifikacijske bakterije (Nitrosomonas)• 2 NH3 + 3 02 = 2 HNO2 + 2 H2O + 158 kcal• 2. Nitratne bakterije (Nitrobacter)• 2 HNO2 + 02 = 2 HNO3 + 38 kcal (solitri; • guano; Čile)• 3. Žveplove bakterije:• 2H2S + O2 ⇒ H20 + S2• S2 + 02 + H20 ⇒ 2 H2SO4• H2S + 2 02 ⇒ H2SO4 + 115kcal

ZAČETEK FOTOSINTEZE
• 4. Purpurne žveplove bakterije• CO2 + 2 H2S(donor e-, H+) ⇒(svetloba) CH20 +H2O +2S
• Svetloba je vir energije• Žveplovodik vir elektronov in protonov• Sprošča se elementarno žveplo

Ostale oblike kemosinteze
• 5. Železove bakterije (Crenothrix, Leptothrix)
• FeCO3 (Fe++) (Fe2(CO3)3 ⇒ Fe(OH)3(Fe+++)
• 6. Metanove bakterije (Bacillus methanicus)
• CH4 + 2O2 ⇒ CO2 + 2H2O

FOTOSINTEZA VIŠJIH RASTLIN, EV. ALG, CIANOBAKTERIJ
• 6CO2 + 6 H2O(donor H+, e-) + 675 kcal ⇒(klorofil, h() C6H12O6 + 6O2
• n H2O + n CO2 ⇒(h.√) (CH2O)n + nO2 ; endergon proces!
• (Go = + n. 477KJ ⇒(+ n. 114 KCAL); n=6• 1. 2H2O + 2A ⇒(h.√) 2AH2 + O2• 2. CO2 + 2AH2⇒ CH2O +H2O + 2A• 1+2( CO2 + 2 H2O ⇒(h.√) CH2O + O2 +
H2O

BISTVO FOTOSINTEZE:
• pretvorba svetlobne energije v kemijsko• oksidacija vode (fotoliza); sprošča se kisik• redukcija ogljik.dioksida; nastanek org.
spojin (sladkorji, amino kisline, maščobe)• PQ (fotosintetski količnik) = O2/CO2 = 1
(samo v idealnih razmerah!)

FAZE FOTOSINTEZE
I. SVETLOBNA FAZA: pretvorba svetlobne energije v kemično: ATP; fotoliza vode: NADPH2(2e-, 2H+); nastanek kisika: O2
- Poteka na tilakoidnih membranah kloroplasta- II. TEMOTNA FAZA: redukcija CO2: nastanek
sladkorjev, aminokislin, maščob,..-poteka v stromi kloroplasta
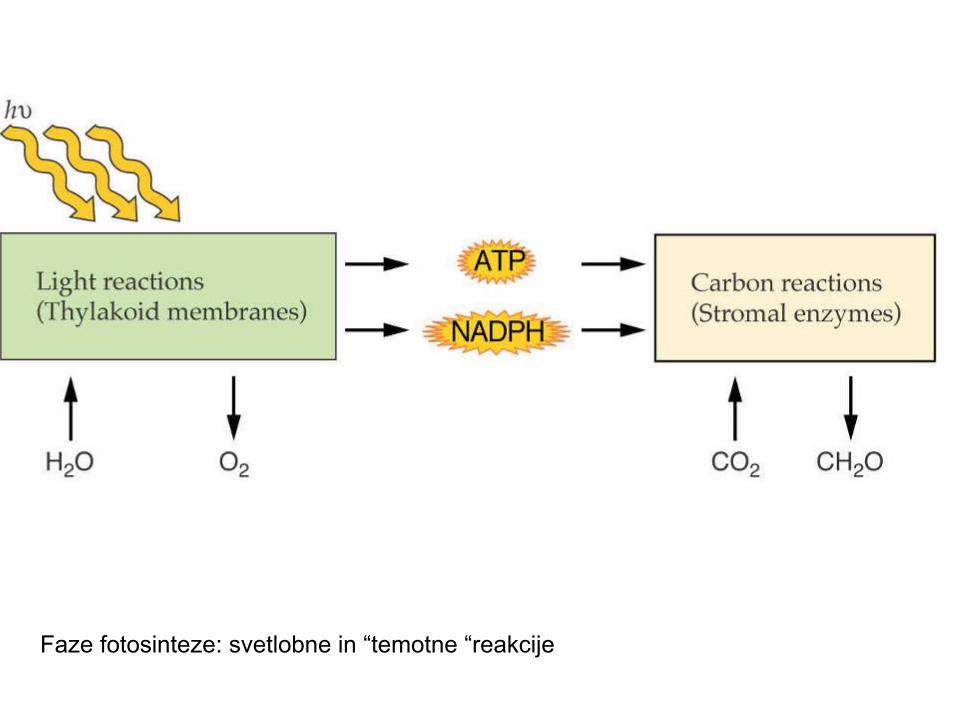
Faze fotosinteze: svetlobne in “temotne “reakcije

I. SVETLOBNA FAZA FOTOSINTEZE
• Pomen svetlobe za rastline:• vir energije• regulator rasti in razvoja (fitokromi,
kriptokromi)• toplotni učinek• svetloba kot stres (preveč, premalo,..)• Učinki so odvisni od spektralnega sestava,
jakosti sevanja in trajanja sevanja

a) Fotofizikalne reakcije:• - absorbcija svetlobne energije:E¸= h.√; fotoni;
kvanti; molekule klorofila; ant. kompleksi, reakcijski centri - prenos energije do reakcijskihcentrov; PSI (700nm), PSII (680nm)
• - ekscitacija in deekscitacija molekul klorofila a; glavni in pomožni pigmenti;
• - Emersonov efekt; PSI, PSII, "red drop"; pomen(; vzburjenje elektrona;
• izbitje elektrona; fluorescenca; toplota; hiperenergetsko stanje kloroplasta

Pomen fotosinteznih barvil:
• A) Glavna: Klorofil a (bakterioklorofil): absorbcija svetlobe in pretvorba energije fotonov v energijo elektronov
• B) Pomožna (antenski pigmenti): pomočpri absorbciji svetlobe (spektralni sestav; okolje!): klorofil b (c, d, e), karotenoidi, fikobilini);zaščita kloroplasta v stresnih razmerah: karotenoidi(karoteni→ksantofili; ksantofilski cikel)

FOTOSINTEZNI UČINEK SVETLOBE
• PhAR(phtosynthetic active radiation): 400 (380) – 700 (710) nm
• Učinek je odvisen od spektralne sestave (=energetske vsebnosti sevanja: E=hν=hc/λ (npr. 1 mol fotonov modre svetlobe -490 nm- ima energijo 240 kJ, 1 mol rdeče svetlobe z val. dol 700 nm pa le 170kJ(1E)

Pomen karotenoidov
• Pomožna absorbcija v modrem delu PhAR• zaščita kloroplasta pred fotokemijsko
oksidacijo fotoinhibicijo• Pomen kemijske zgradbe karotenoidov• Vloga ksantofilskega cikla – odaptacija
kloroplasta na svetlobno okolje

B) Fotokemične reakcije• Z-shema, Hill & Bendal 1960• - izbitje elektronov v reakcijskih centrih (PSI,
PSII); fotoliza vode (PSII)• - vektorski transport elektronov iz vode ( pomoč
PSI, PSII) preko prenašalcev na končneprenašalce (NADP+; O) in poraba v temotni fazi(CO2, NO3
-, SO4--,
• ....; Z - shema);• - vektorski transport protonov H+ iz strome v
notranji lumen tilakoidnih membran;• - fotofosforilizacija (tvorba ATP; ADP + Pi(ADP)

Funkcionalna organizacija tilakoid
• Fotosintezna barvila (klorofili, karotenoidi) so vezana v reakcijske centre (PSI 700, PSII 680) in njim priključene žetvene komplekse (LHC-I, LHC-II)
• V reakcijskih centrih je samo kl a, v “antenah” poleg kl a še kl b, feofitin, karotenoidi in prijemniki elektronov
• Vsi klorofili so vezani na beljakovine, večji del tilakoidnihbeljakovin so del LHC kompleksov
• Za funkcioniranje je pomembna prostorska in topografska organizacija tilakoid (bližina pomeni prenos energije; odklaplanje in priklapljanje LHC na PS
• Pri rdečih algah in cianobakterijah so fikobilini (fikoeritrinin fikocian) organizirani v vodotopne LHC imenovane fikobilisome)

PSI in PSII tvorita funkcionalno enoto
• Samo fotosintetsko aktivne bakterije in cianobakterije imajo 1 PS
• Evkariontske alge in vse višje rastline imajo PS I in PSII, ki delujeta v navezi
• Delež/število PSI/PSII je odvisno od jakosti sevanja
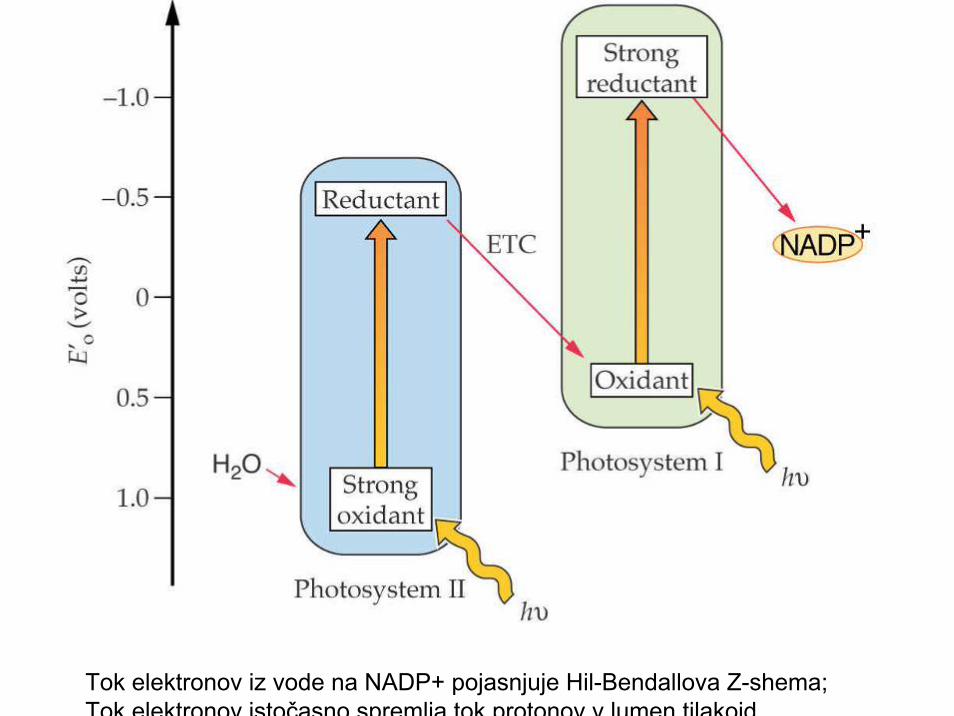
Tok elektronov iz vode na NADP+ pojasnjuje Hil-Bendallova Z-shema;Tok elektronov istočasno spremlja tok protonov v lumen tilakoid

Tvorba ATP- fotofosforilizacija• Teorije nastanka ATP: kemijska hipoteza (Arnon et al. 1954, Slater 1954)• kemoosmotska hipoteza (Mitchell 1966, Nobel 1978):• Fotofosforilizacija poteka na intaktnih (topografsko celih) tilakoidah, ki so
relativno nepropustne za H+ in OH- ione.• Vir e- in H+ je voda. Prenos e- iz vode (fotoliza vode!, PSII) na prejemnike
je vektorski - preko prenašalcev, vezanih za oba PS v tilakoidni membrani.• Prenos e- je istočasno spremljan s prenosom H+ iz strome kloroplasta v
notranji lumen zlepljenih (naloženih) tilakoid. Nastanek koncentracijskegagradienta (H+!) in el. polja preko tilakoidne membrane.
• Kombinacija konc. gradienta in el. polja tvorita visoko energetsko stanje s težnjo po izmetu protonov (H+) iz notranjega lumna tilakoid.
• V tilakoidah obstaja vektorsko vezana ATP sintetaza (F1-Fo kompleks; integralni, transmembranski protein), ki na račun energije elektrokemijskegapotenciala in gradienta protonov sintetizira ATP iz ADP in Pi.

Kaj potrjuje hipotezo?
• - poskusi na izoliranih in intaktnih tilakoid. membranah (meritve fluorescence, sproščanje O2, meritve pH, toplote, količine ATP, NADPH+H,...)
• - analize tilakoidnih membran z el. mikroskopom in biokemične analizesestave
• proteinov v tilaoidnih membranah.

Prikazana pot elektronov med PS II in PS I
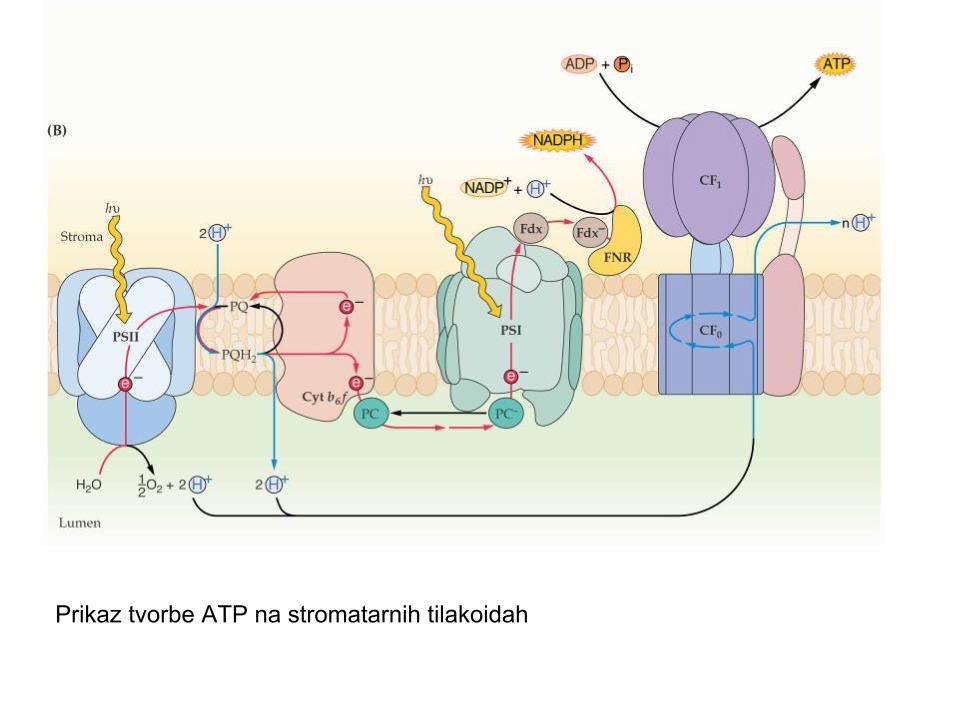
Prikaz tvorbe ATP na stromatarnih tilakoidah
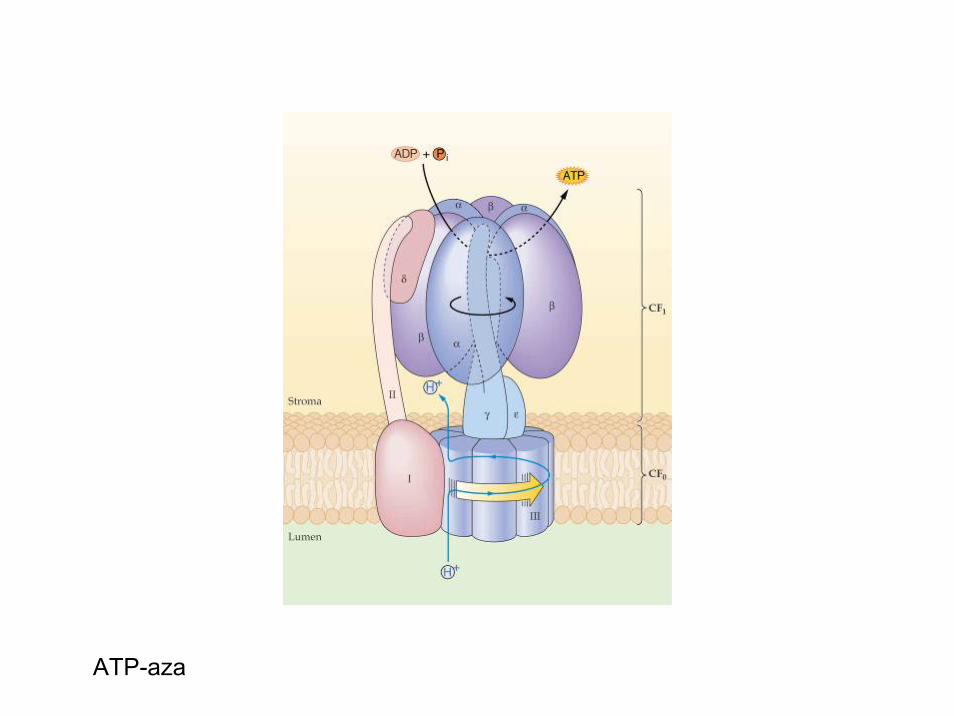
ATP-aza
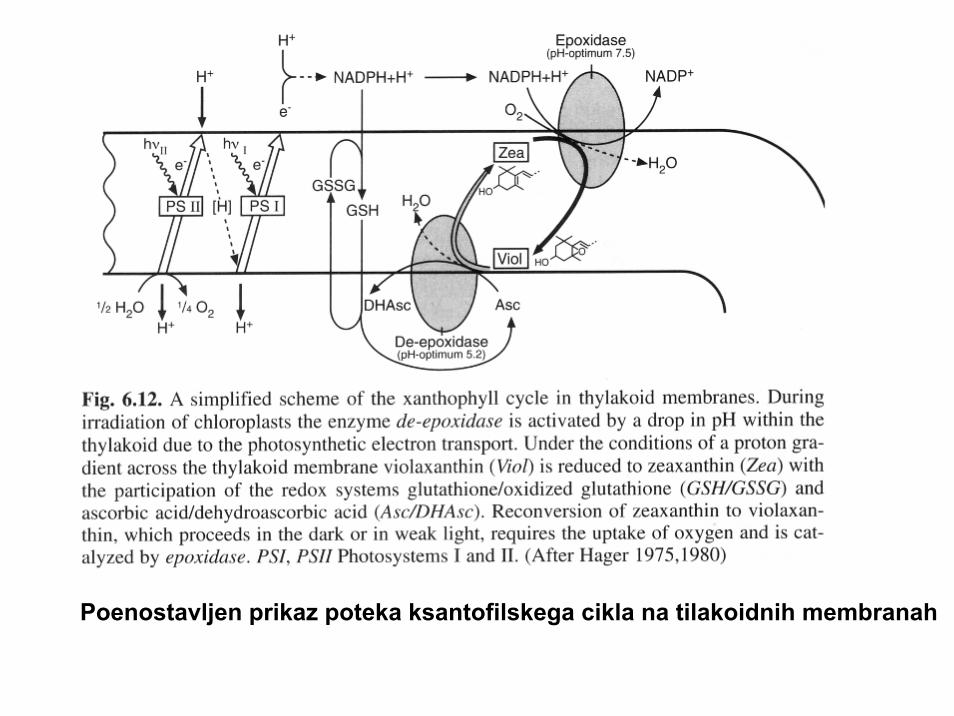
Poenostavljen prikaz poteka ksantofilskega cikla na tilakoidnih membranah

Načini tvorbe ATP
• V normalnih razmerah poteka neciklična fotofosforilizacija, pot elektrona iz vode na NADP+
• Kadar se NADPH+H ne porablja (težave v Kalvinovem ciklu) steče na PS I ciklična fotofosforilizacija, tvori se samo ATP
• Ob popolni redukciji PS I lahko sprejme elektron kisik – Maehlerjeva reakcija-psevdociklična fosforilizacija

Povzetek• - ciklična: PSI; pot e-: PSI⇒PSI (kl a 700nm);• - neciklična: PSII; pot e-: H2O⇒PSII⇒PSI ⇒NADP+:• fotoliza vode: 2H2O⇒(Mn++) ⇒O2+ 4H+ +4e-;• prenos e- na NADP+; PSII (Mn -protein!); Kok et al. 1970;
(H2O + NADP+ + h.√⇒ 1/2O2 + NADPH+H+)• - pseudociklična : PSII,PSI; pot e-: H2O⇒e- iz
H2O⇒PSII⇒PSI⇒O⇒O2• (2H+ + O2+ 2e- ⇒H2O2 (katalaza) ⇒H2O + 1/2O2);
Edwards & Walker 1983; "Mehlerjeva"reakcija)

POVZETEK REAKCIJ IN PRODUKTI SVETLOBNE FAZE:
• 2H2O + 2 NADP+ + 3 ADP + 3Pi (H2PO4) + 8-12 FOTONOV⇒ O2 + 2 NADPH + 2H+ + 3ATP + 3H2O;
• 1 molekula H2O⇒ 1/2O2 + NADPH +H+ + 1,5 ATP;
• za redukcijo 1CO2 potrebujemo: 3 ATP + 2 NADPH +H+
• zadošča že neciklična fosforilizacija, a v kloroplastu ne poteka le redukcija ogljika; stresne razmere!! (poraba ATP in NADPH za reparacijo, delo,..)

TEMOTNA FAZA FOTOSINTEZE = KALVINOV CIKEL= RPP CIKEL• Povzetek reakcij:• 6CO2+ 18 ATP + 12 NADPH + 12H+
⇒C6H1206+ 18ADP + 18 Pi + 12NADP+• Proces ne poteka v temi• Sodelujejo številni encimi-temperaturna
občutljivost• Tvorijo se različni c-skeleti, ki se usmerjajo
glede na potrebe rastline

Regulacija Kalvinovega cikla
• Encimatska – 13 encimov, 3 le v kloroplastu (RUBISCO, sedoheptoloze-1,7-bifosfataza, fosforibulokinaza)
• Stanje reducentov in energije (ATP, NADPH)
• Metabolitna (snovi, ki vzstopajo in izstopajo: CO2, O2, 1. akceptor(R-BP,C5), drugi sladkorji)
• Okolje (svetloba, temperatura, voda)

Faze Kalvinovega cikla
• Karboksilacija• Redukcija• Regeneracija substrata• Avtokataliza
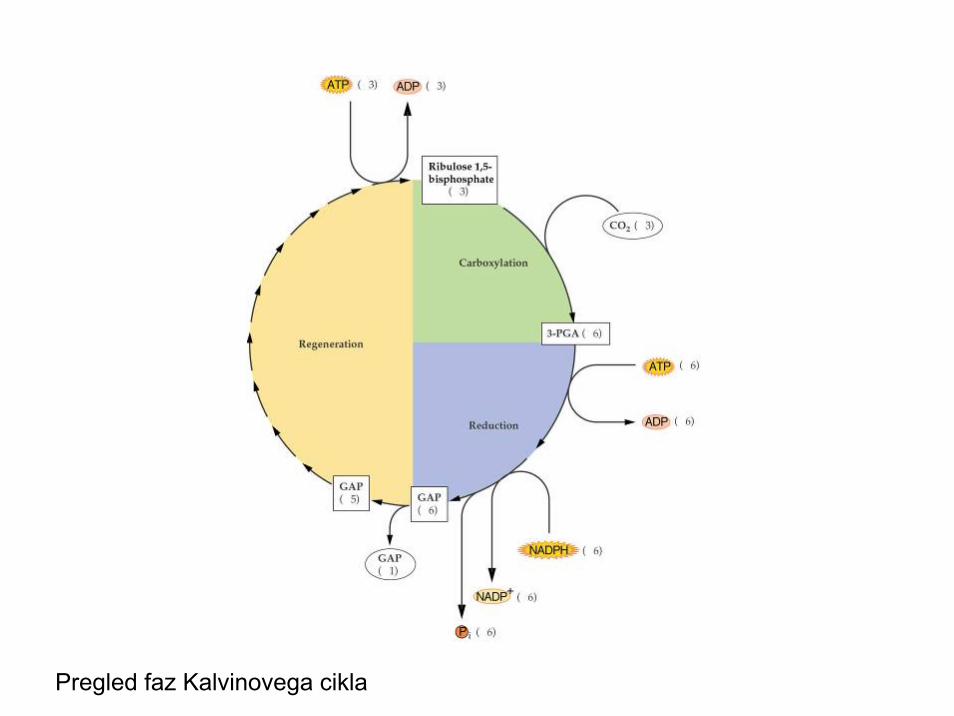
Pregled faz Kalvinovega cikla

Karboksilacija
• Vezava CO2 na prvi akceptor, sladkor ribuloza bifosfat
• Akceptor mopra biti aktiviran • Problem je pomanjkanje CO2, še posebej
ob zaprtih režah; tedaj vezava kisika, RUBISCO postane RUBISO
• Potek pri večini rastlin:• CO2 (C1) RuBP (C5)→(C6) 2PGK (C3)
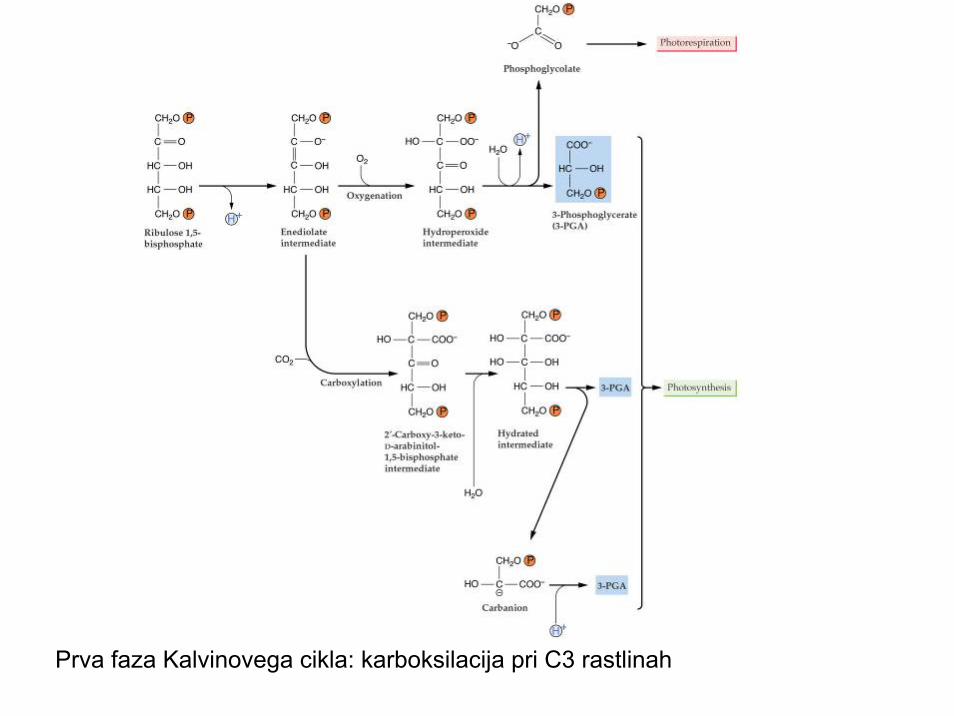
Prva faza Kalvinovega cikla: karboksilacija pri C3 rastlinah

FOTORESPIRACIJA• Ob zaprtih režah zmanjka CO2• Na isti substrat se veže kisik ob katalizaciji
istega encima:• O2 (C0) + RuBP (C5)→(C5) 1PG (C3)+1PG (C2,
fosfoglikolat)• Fosfoglikolat se presnavlja v peroksisomih, ob
sodelovanju klooroplastov in mitohondrijev –fotorespiracija (pomen: stres, povezava metabolizma C in N)
• Rešitve: C4, CAM in C1 tipi fotosinteze

C4 FOTOSINTEZA
• Pretežno tropske trave (Poaceae, Amarantaceae, Aizoaceae, Chenopodiaceae)
• Pretežno zerlišča, bogatih, dobro osvetljenih in na hranilih bogatih tal
• Dva tipa karboksilacije v listu; dimorfizemkloroplastov in asimilacijskega parenhima
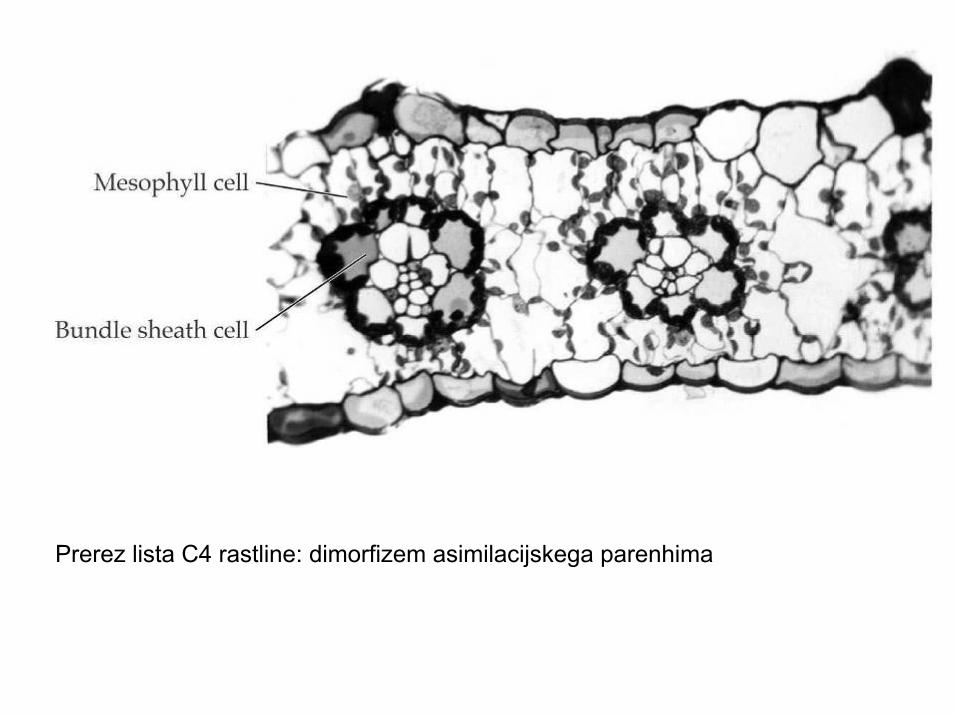
Prerez lista C4 rastline: dimorfizem asimilacijskega parenhima
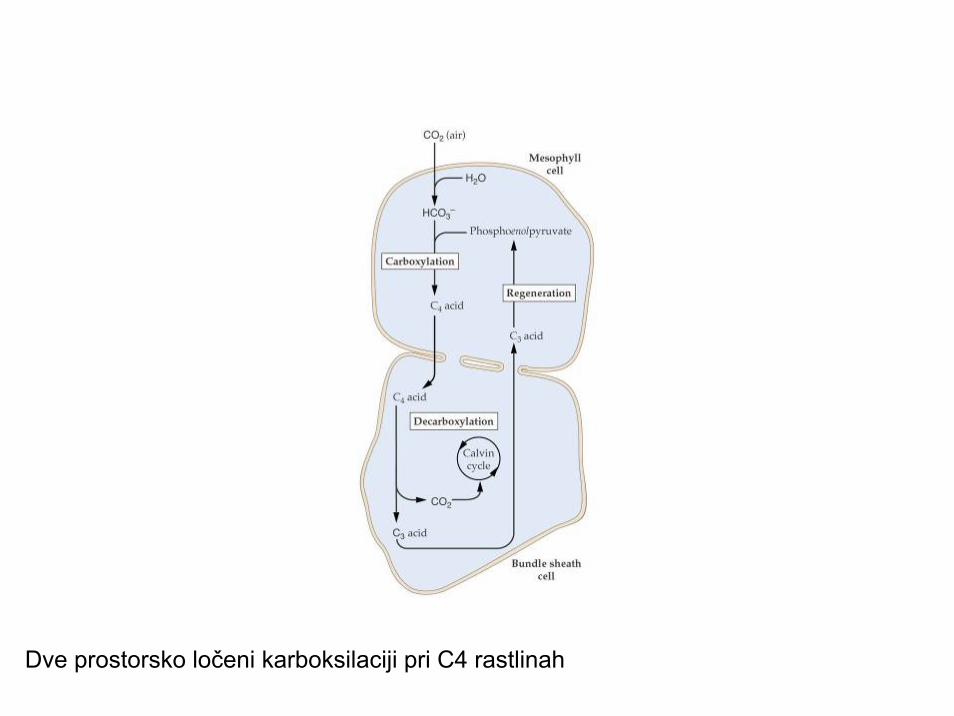
Dve prostorsko ločeni karboksilaciji pri C4 rastlinah

Dimorfizem kloroplastov pri C4 rastlinah: agranularni kloroplasti celicžilnega ovoja nimajo PSII
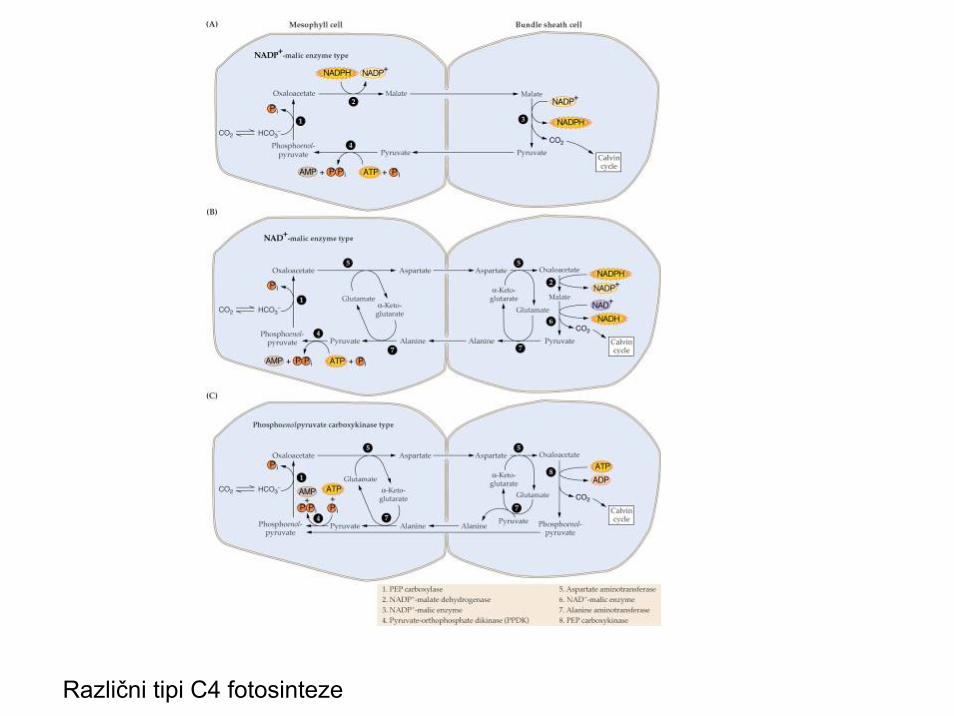
Različni tipi C4 fotosinteze

CAM tip fotosinteze: Crassualacean Acid Metabolism
• Problem odprtosti rež pri pomanjkanju vode; Crassulaceae, Euphorbiaceae, Aizoaceae,Bromeliaceae, Chenopodiaceae,..
• Odprte reže ponoči (manjše sevanje, manjša transpiracija, vezava CO2 iz zraka)
• Podnevi reže zaprte in izraba CO2 iz rezervoarja C4-kislin v vakuoli
• Majhna produkcija a preživetje v zelo sušnih območjih (puščave, polpuščave, epifiti,..
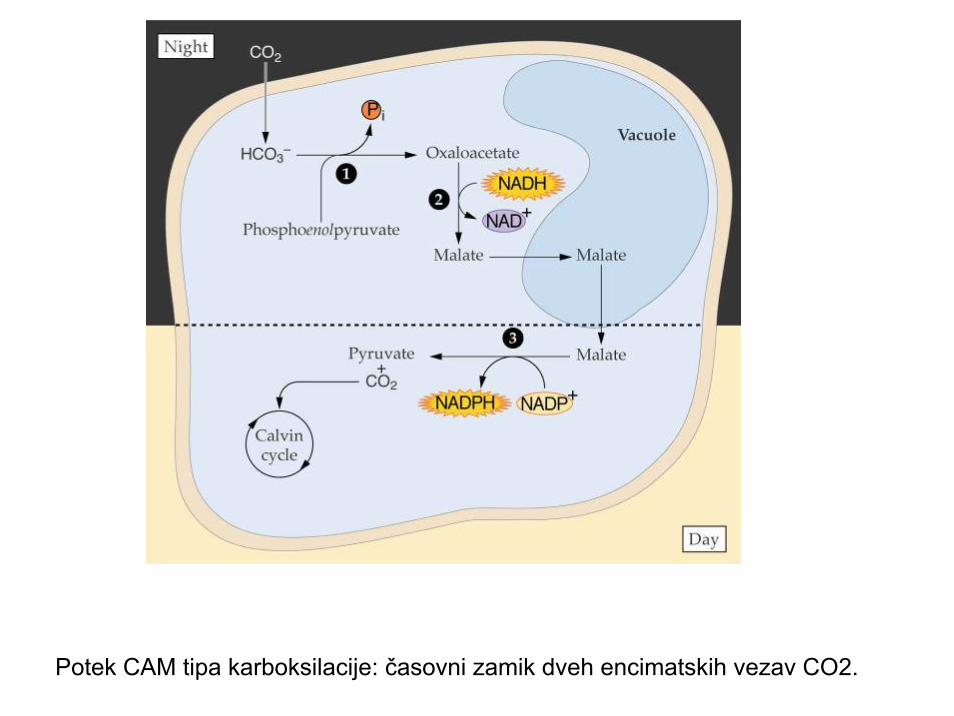
Potek CAM tipa karboksilacije: časovni zamik dveh encimatskih vezav CO2.
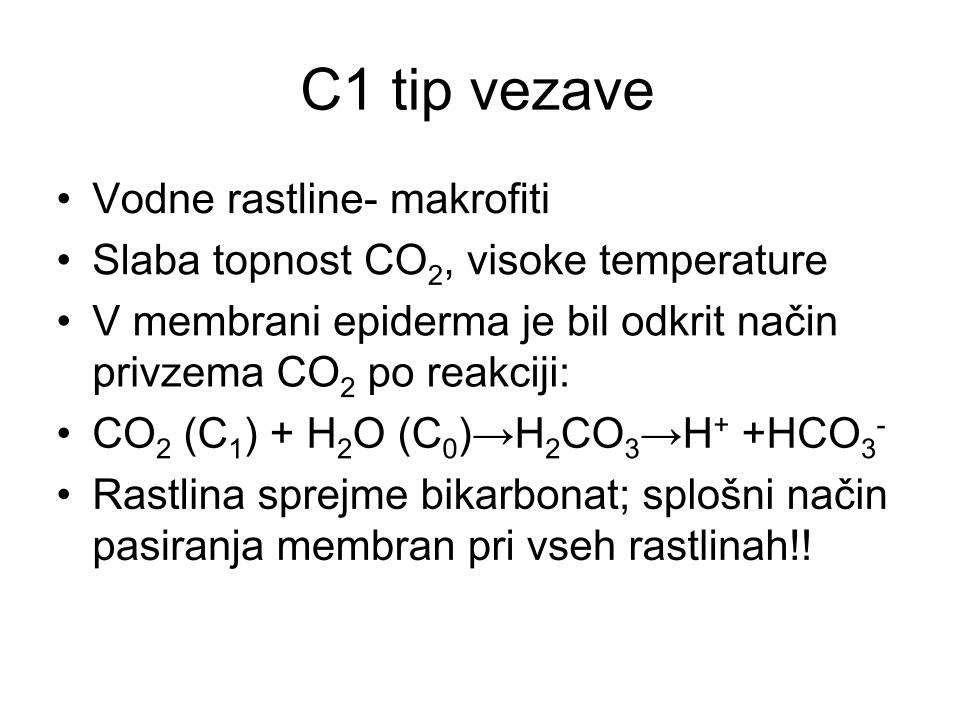
C1 tip vezave
• Vodne rastline- makrofiti• Slaba topnost CO2, visoke temperature• V membrani epiderma je bil odkrit način
privzema CO2 po reakciji:• CO2 (C1) + H2O (C0)→H2CO3→H+ +HCO3
-
• Rastlina sprejme bikarbonat; splošni način pasiranja membran pri vseh rastlinah!!

REDUKCIJA
• Pomeni redukcijo fosfoglicerata v triozo: fofoglicerin aldehid, dihidroksi aceton fosfat
• Porabita se ATP in NADPH2• Reakcija pomeni energetski prag• PGA + NADPH+H + + ATP + h.( (trioze
fosfat; C3 spojina

Regeneracija akceptorja- RuBP
• 6 C5 (ribuloza bifosfat) + 6 C1(CO2) ⇒12 C3⇒6C5, ⇑ 1C6!!
• “feedback” regulacija; ker CO2 pogosto primankuje, je vedno dovolj RuBP, da Kalvinov cikel normalno teče
• Dokazano z izotopi ogljika
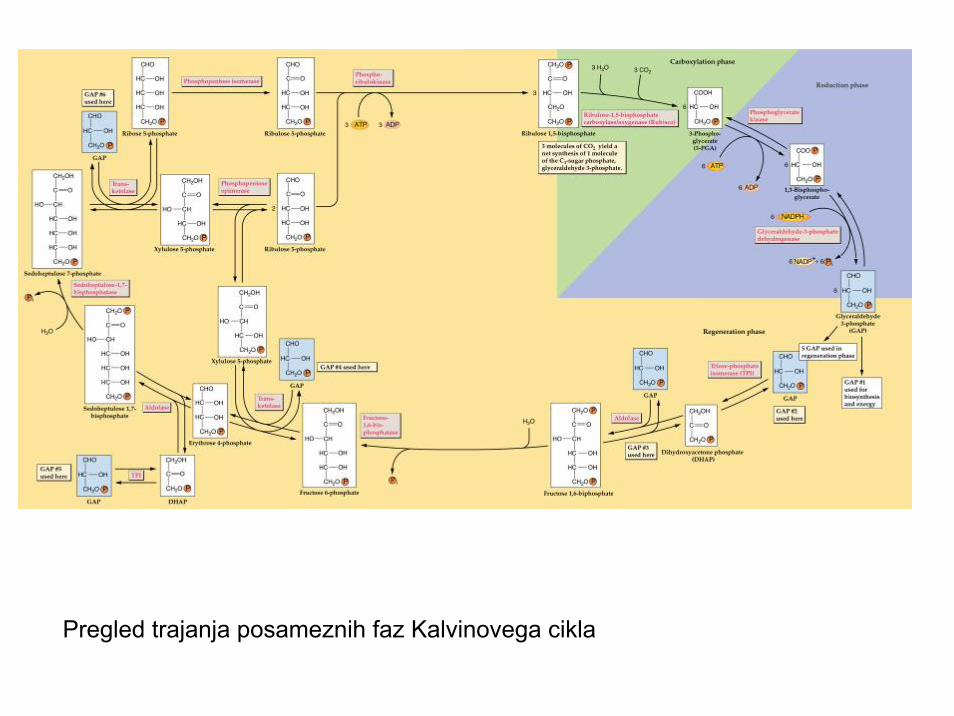
Pregled trajanja posameznih faz Kalvinovega cikla

AVTOKATALIZA
• - uklanjanje C3, C6 spojin iz strome; sinteza škroba (asimilacijski škrob); eksport saharoze, trioz; povezava s sintezo drugih spojin (eritroza ⇒ riboza⇒RNK; deoksiroboza(DNK; ); sek. metabolizem
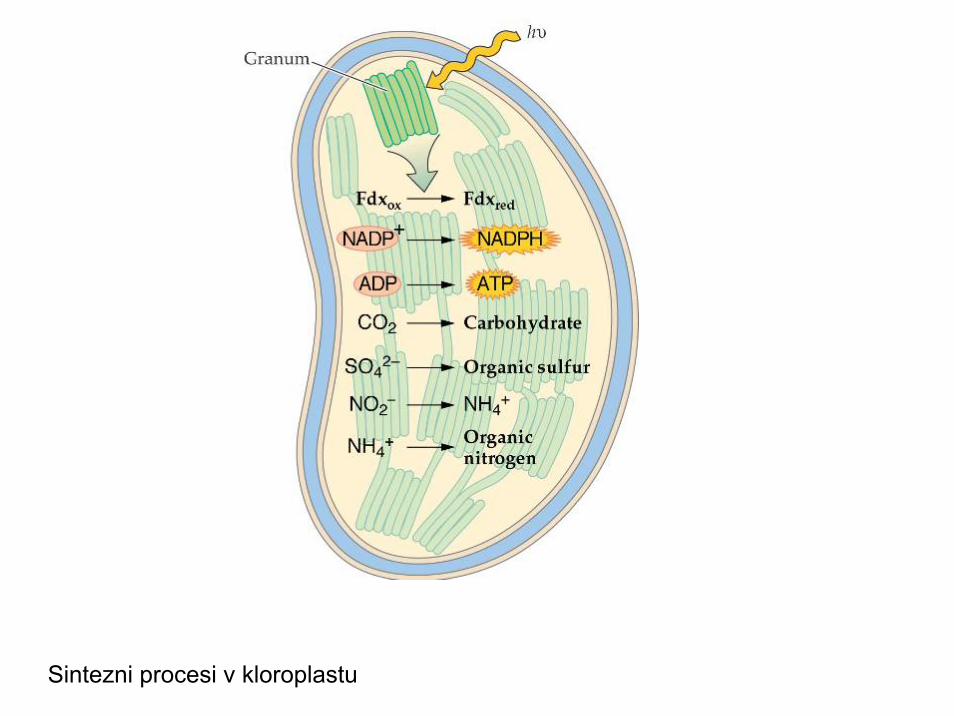
Sintezni procesi v kloroplastu
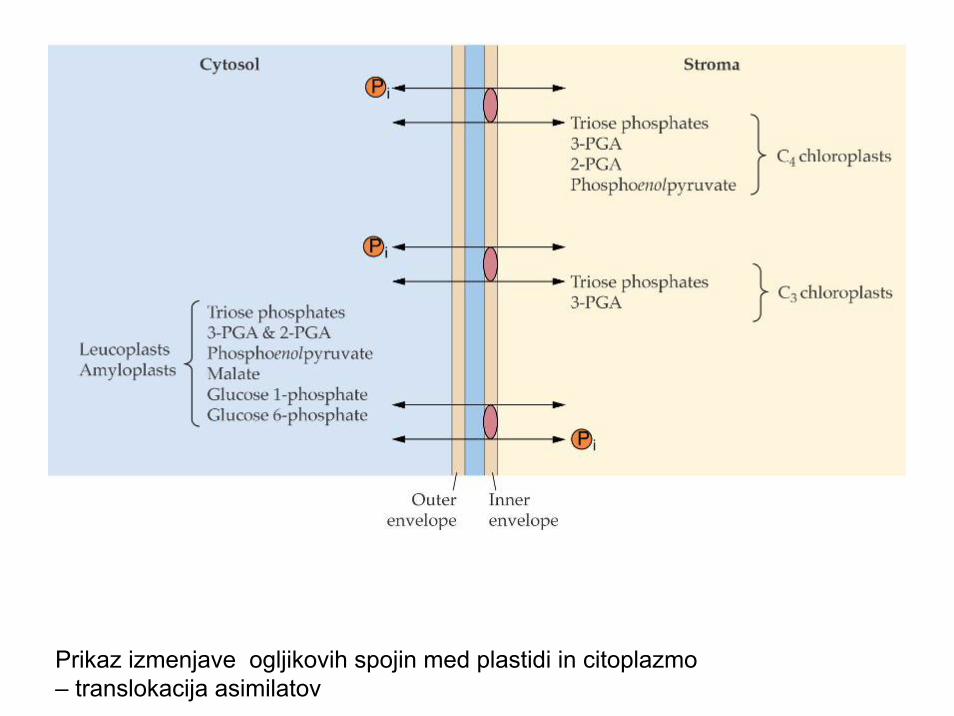
Prikaz izmenjave ogljikovih spojin med plastidi in citoplazmo – translokacija asimilatov
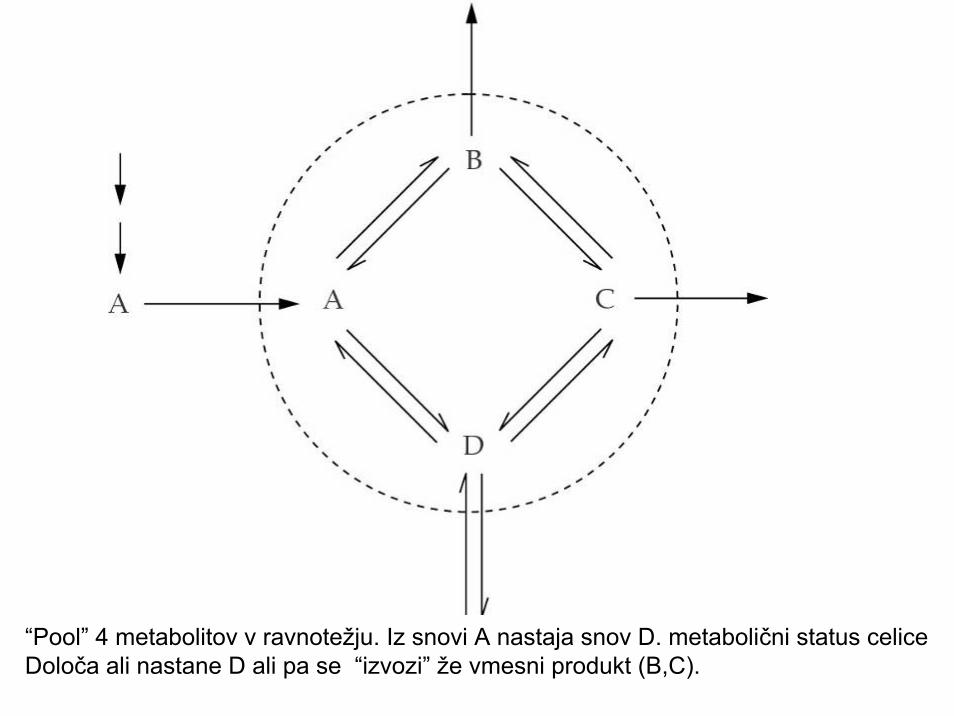
“Pool” 4 metabolitov v ravnotežju. Iz snovi A nastaja snov D. metabolični status celice Določa ali nastane D ali pa se “izvozi” že vmesni produkt (B,C).
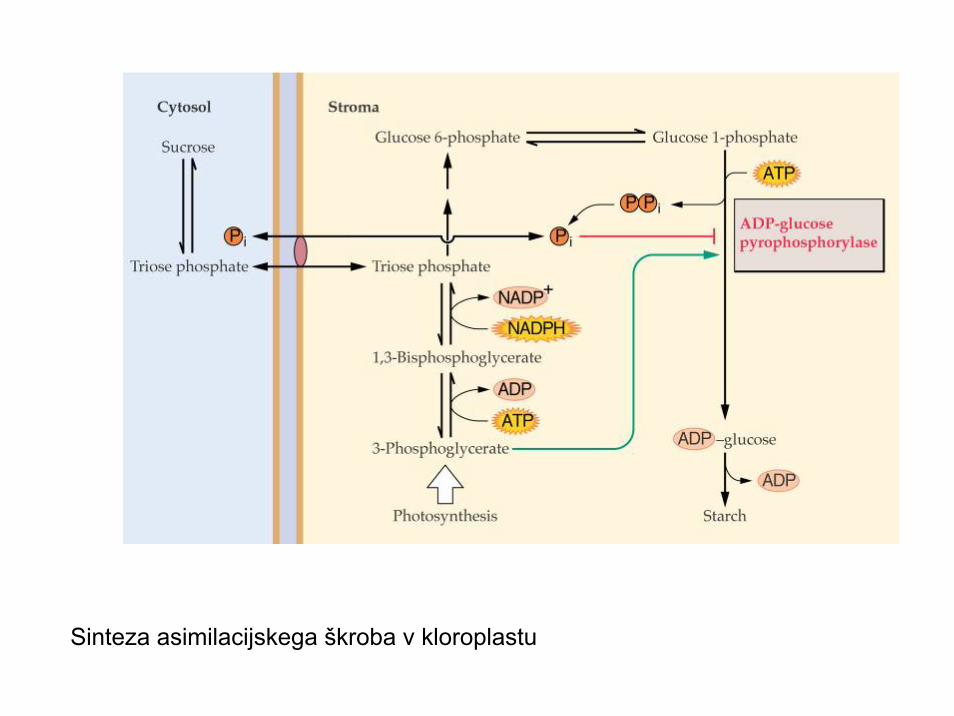
Sinteza asimilacijskega škroba v kloroplastu
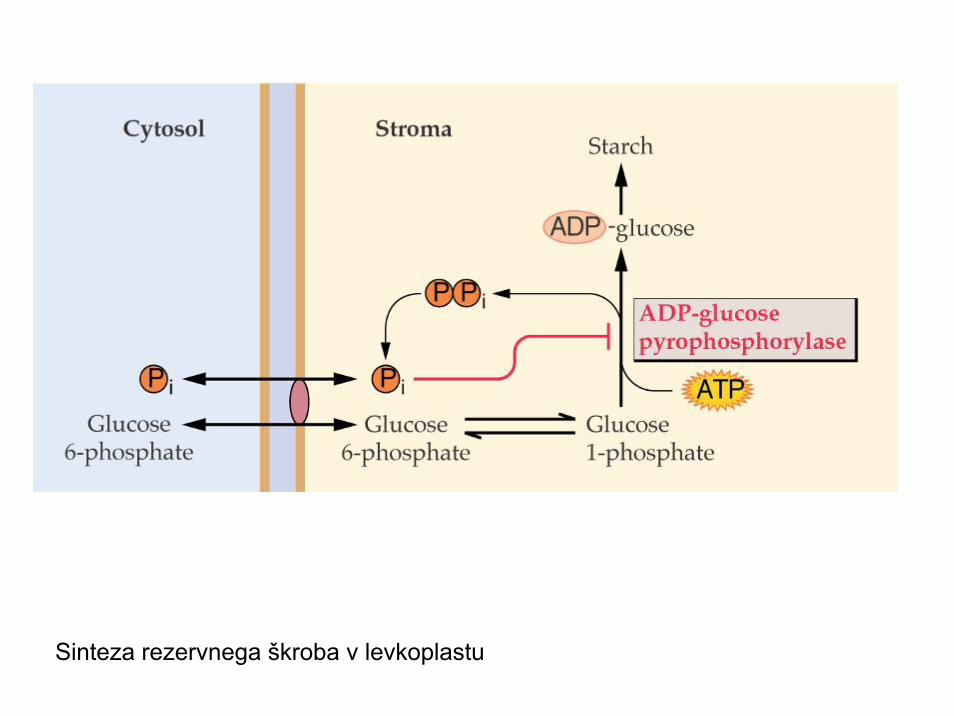
Sinteza rezervnega škroba v levkoplastu

VPLIVI NA FOTOSINTEZO• I. INTERNI: Stanje in vrsta rastline• II. ZUNANJI: Okolje:• 1. [CO2]• Q= D. ΔC.S/d; D= dif. koeficient (cm2s-1); (c= razlika v konc. CO2
med okoljem in celico; S = povprečna debelina lista; d= dolžina potiza CO2
• H2O/CO2; prevodnost , upornost; rs, ra, ri• 2. [O2]• FOTORESPIRACIJA; DIHANJE• 3. SVETLOBA; 4. TEMPERATURA• 5.VODA; 6. MINERALNA PREHRANA• 7. BIOTIČNI DEJAVNIKI OKOLJA: Herbivori; paraziti (glive, živali,
prokarionti)

Interni dejavniki:RASTLINA
• Vrsta, sorta• Starost• Fenofaza• Organ (list, steblo, plod,..)• Tkiva:klorenhimi (asimilacijski
parenhimi),ostala tkiva• Cela rastlina• Trenutne meritve; akumulirana vezava

Načini meritve fotosinteze
• PQ = O2/CO2 = 1 (idealne razmere)• Vodni ekosistemi: meritve sproščanja O2
• Kopenski ekosistemi: meritve vezave CO2
• Meritve fluorescence (oddane svetlobe)• Meritve “pridelka” (žetveni indeks)• Meritve biomase v časovnih presledkih

Pomen svetlobe pri meritvah fotosinteze:
• Multipli vplivi svetlobe, odvisni od jakosti in spektralnega sestava
• Tipala za meritve učinkov svetlobe (kvantna-fotosinteza, radiacijska-energija celotnega spektra, meritve osvetlitve)
• Meritve fluorescence
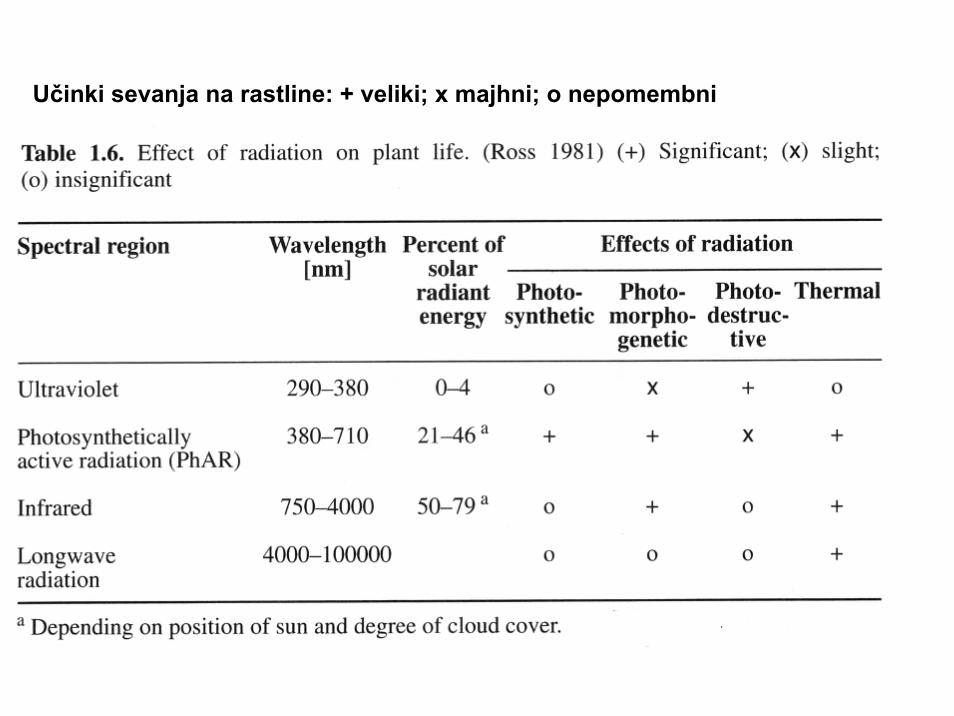
Učinki sevanja na rastline: + veliki; x majhni; o nepomembni
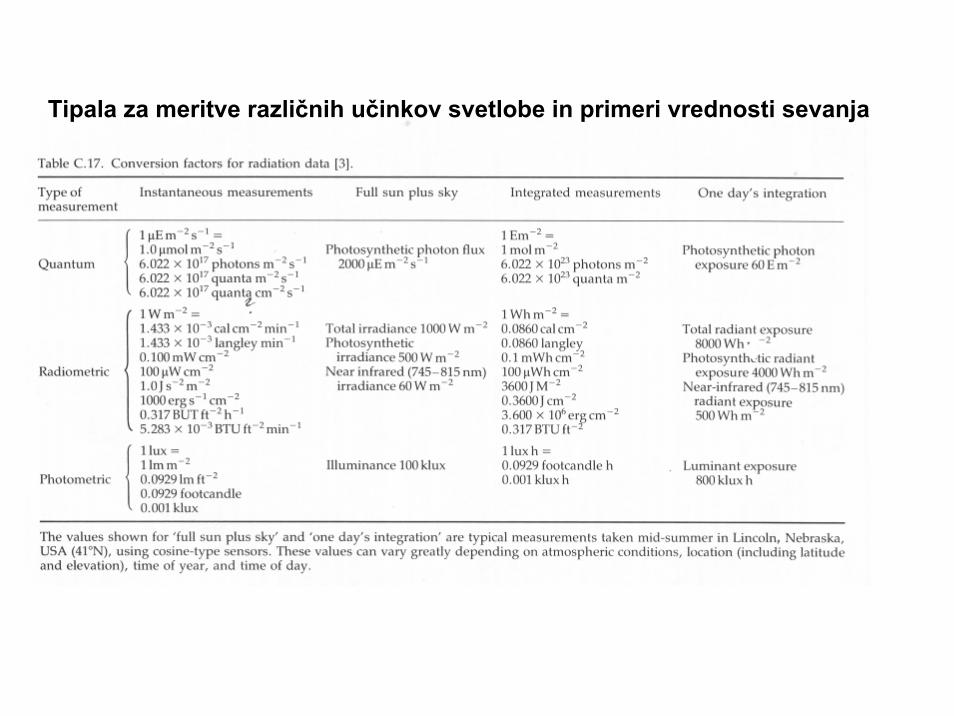
Tipala za meritve različnih učinkov svetlobe in primeri vrednosti sevanja
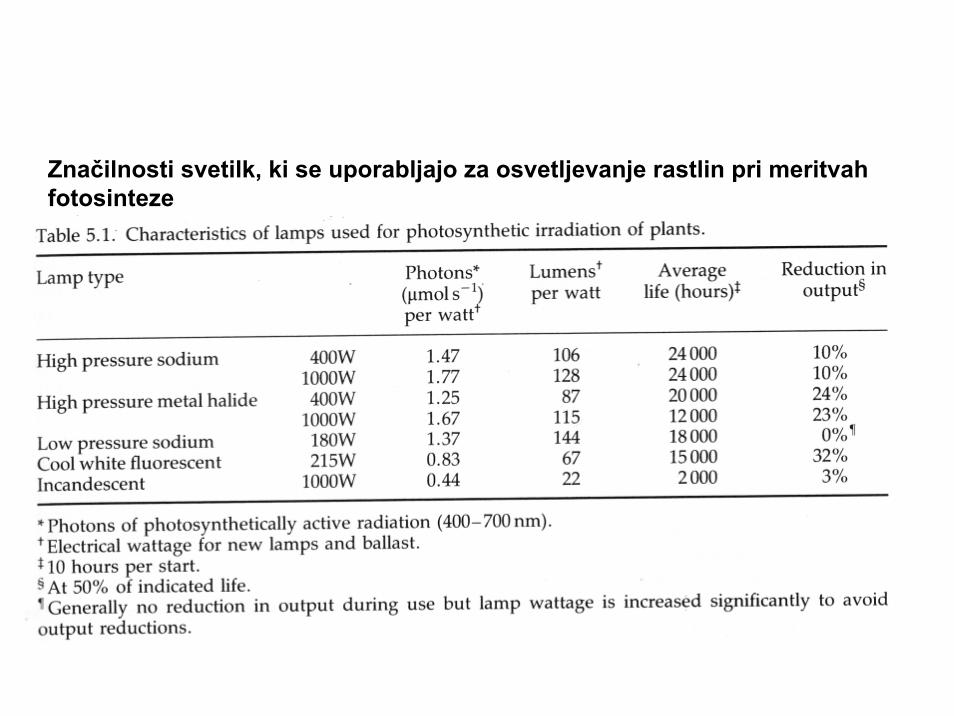
Značilnosti svetilk, ki se uporabljajo za osvetljevanje rastlin pri meritvahfotosinteze
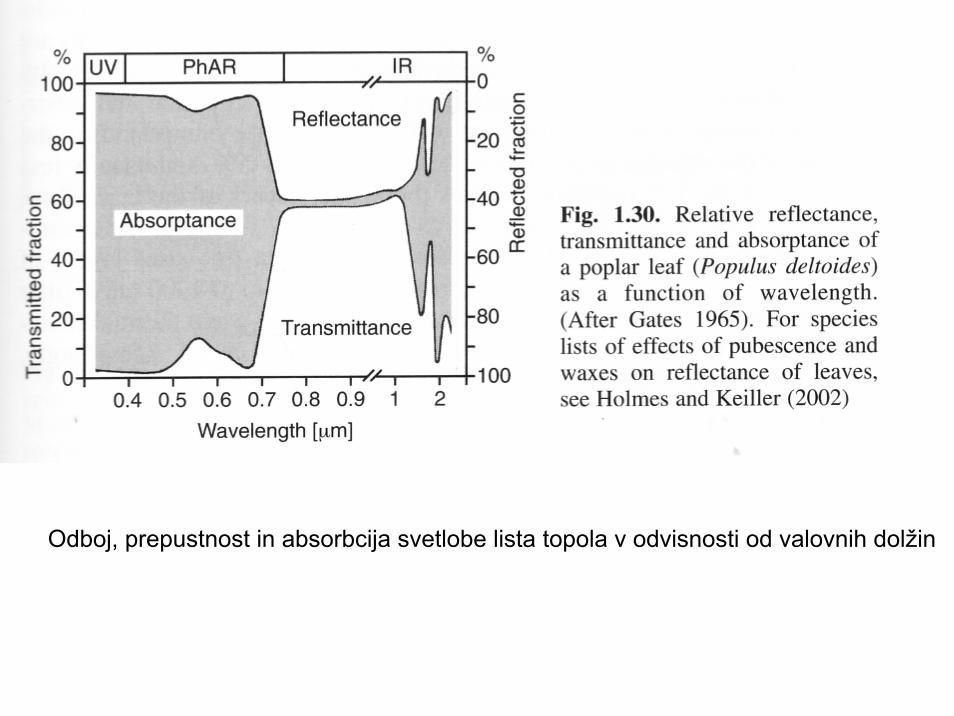
Odboj, prepustnost in absorbcija svetlobe lista topola v odvisnosti od valovnih dolžin
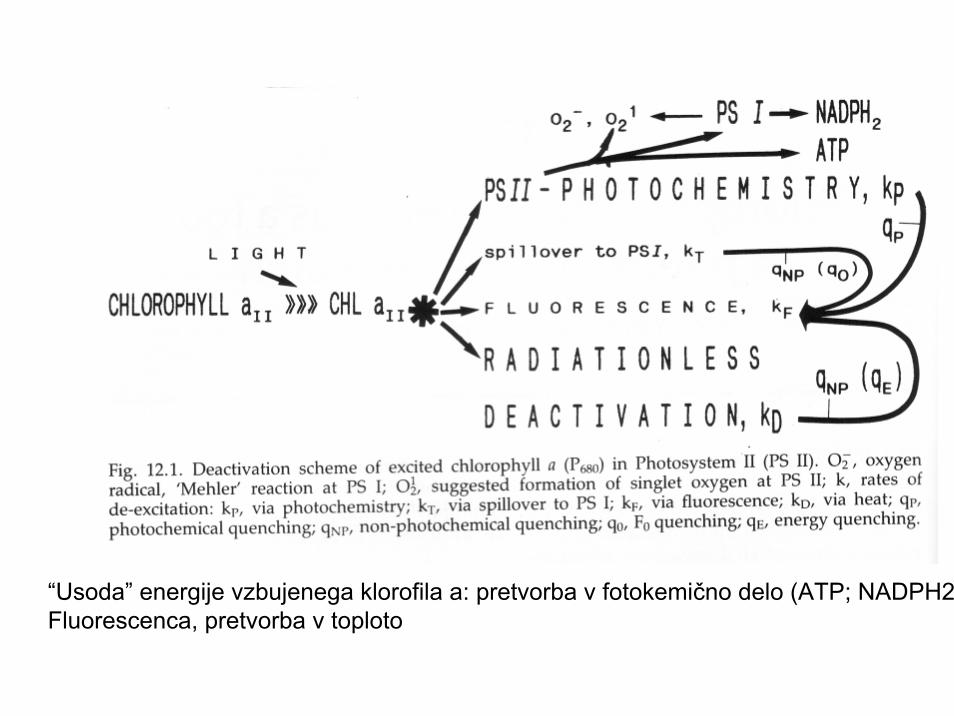
“Usoda” energije vzbujenega klorofila a: pretvorba v fotokemično delo (ATP; NADPH2Fluorescenca, pretvorba v toploto
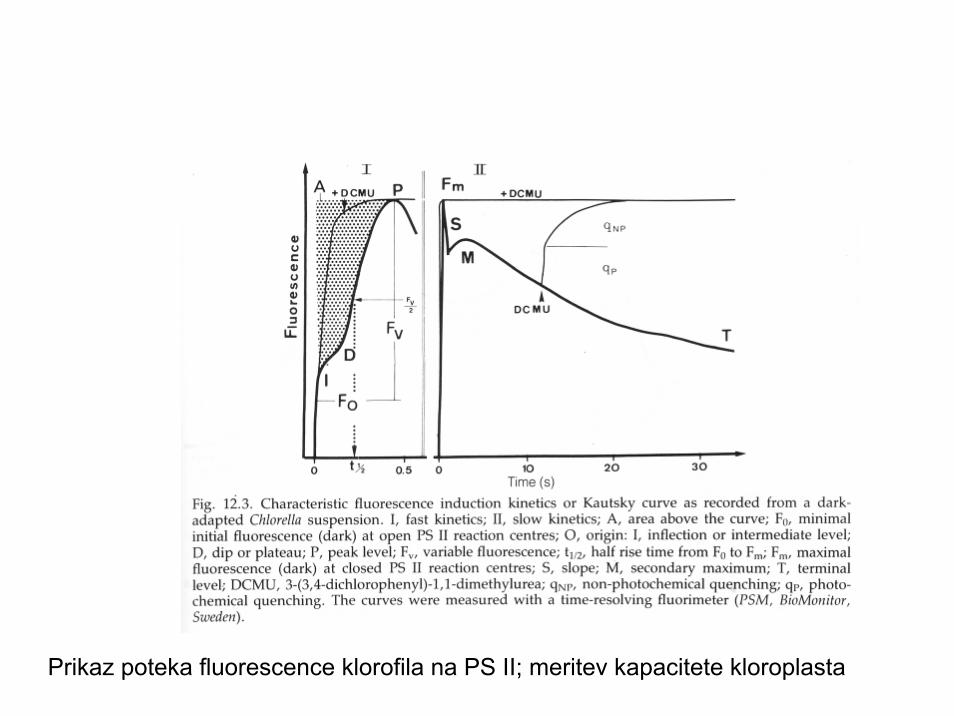
Prikaz poteka fluorescence klorofila na PS II; meritev kapacitete kloroplasta

Meritve izmenjave CO2
• IRGA metode (Infra Red Gas Analysis)• Meritve fotosinteze v kopenskih ekosistemih• Meritve neto fotosinteze: Pn=Pg-R (Pn=neto fotosinteze,
Pg=bruto fotosinteza,R= dihanje)• Kardinalne točke neto fotosinteze: spodnja
kompenzacijska, saturacijska, zgornja kompenzacijska (minimum, optimum, pesimum)
• Spreminjanje vplivnih dejavnikov in meritve fotosinteze; standarne razmere
• Svetlobna, CO2 saturacijska krivulja,..• Odprti in zaprti IRGA sistemi; absolutni in diferencialni
način meritev; meritve spremljajočih parametrov
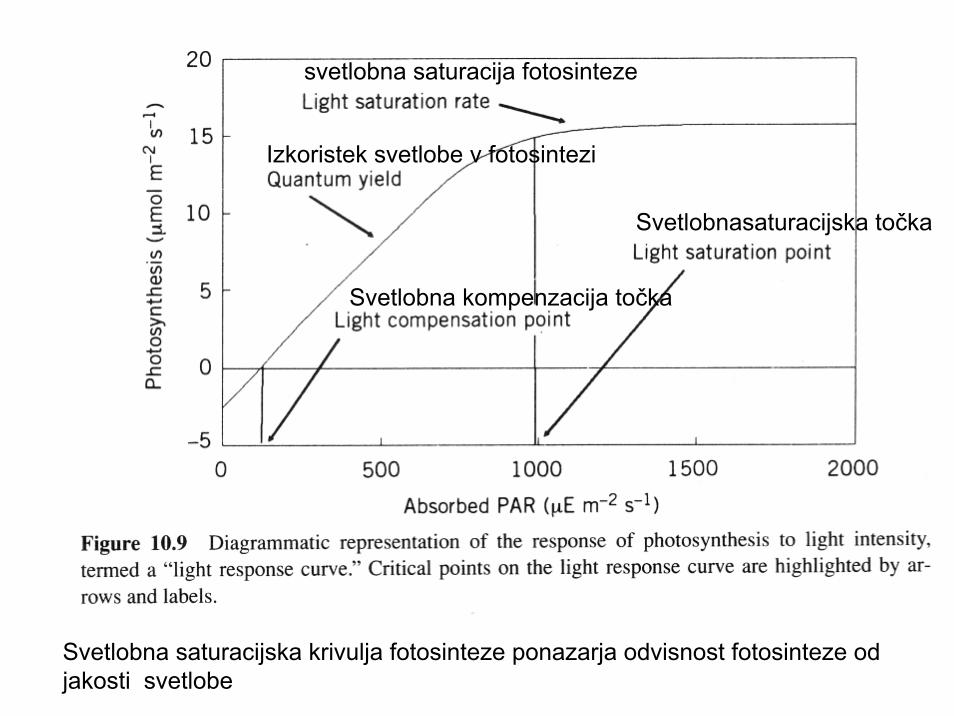
Svetlobna saturacijska krivulja fotosinteze ponazarja odvisnost fotosinteze odjakosti svetlobe
svetlobna saturacija fotosinteze
Svetlobnasaturacijska točka
Svetlobna kompenzacija točka
Izkoristek svetlobe v fotosintezi
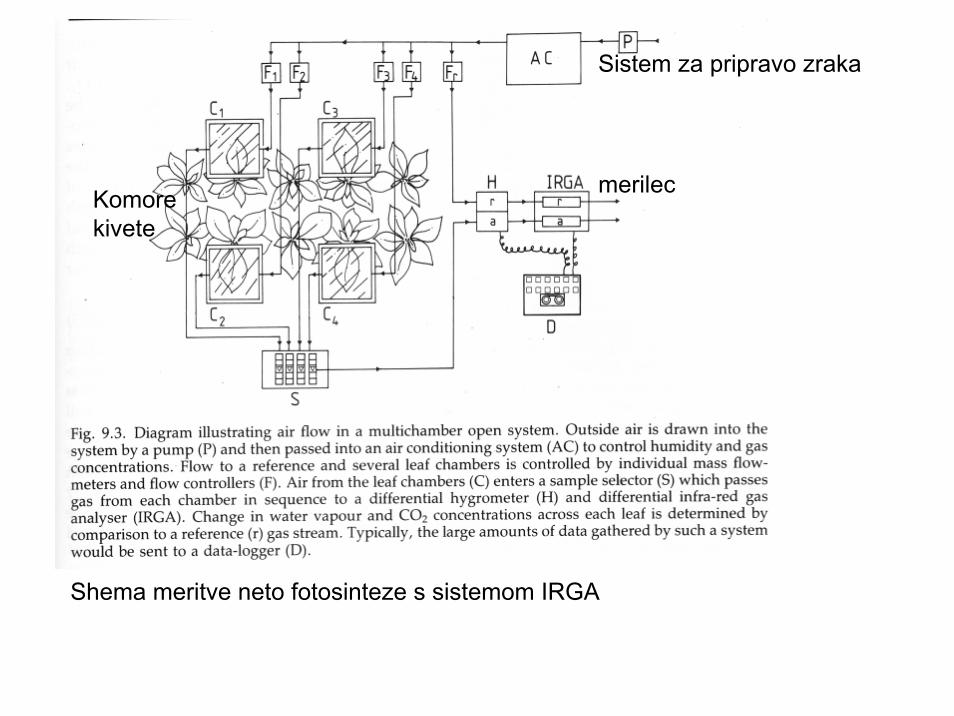
Shema meritve neto fotosinteze s sistemom IRGA
Sistem za pripravo zraka
merilecKomorekivete

Meritve velikosti fotosinteze so lahko na podlagi izmenjave CO2 posameznihrastlin/njihovih organov-največkrat lista (slika levo) ali pa merimo izmenjavo CO2na ravni tipa vegetacije (“eddy covariance metoda, slika desno).
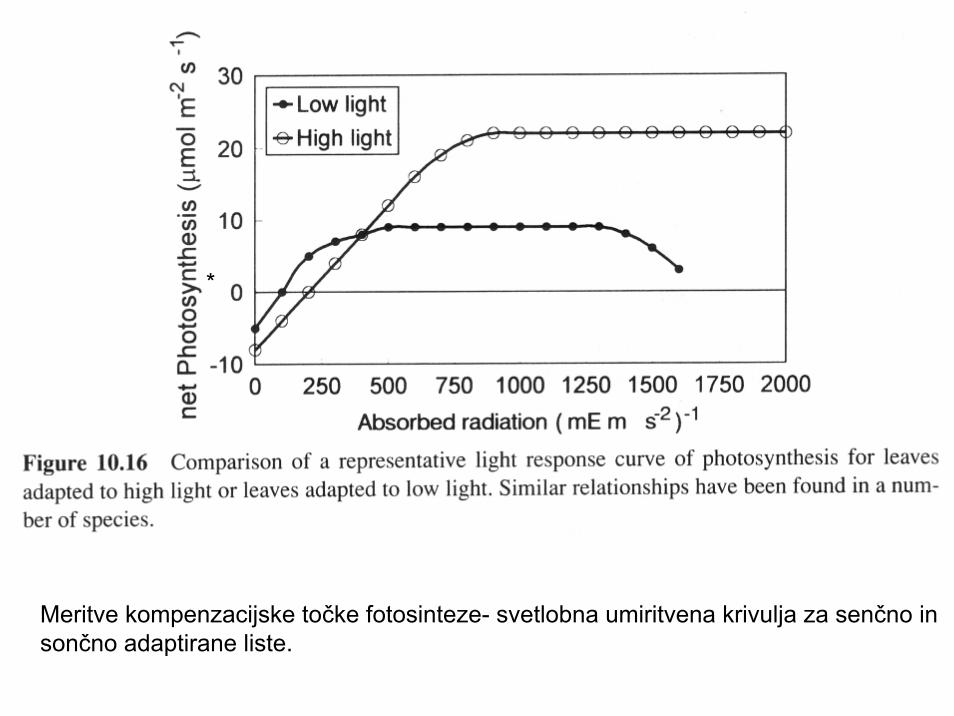
Meritve kompenzacijske točke fotosinteze- svetlobna umiritvena krivulja za senčno insončno adaptirane liste.
*
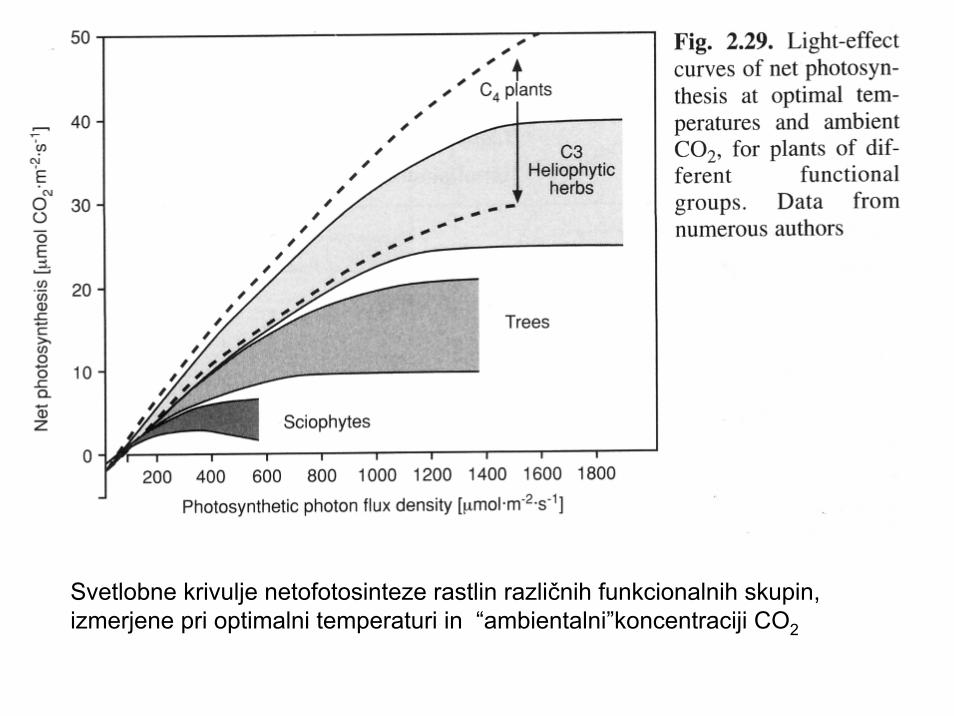
Svetlobne krivulje netofotosinteze rastlin različnih funkcionalnih skupin,izmerjene pri optimalni temperaturi in “ambientalni”koncentraciji CO2
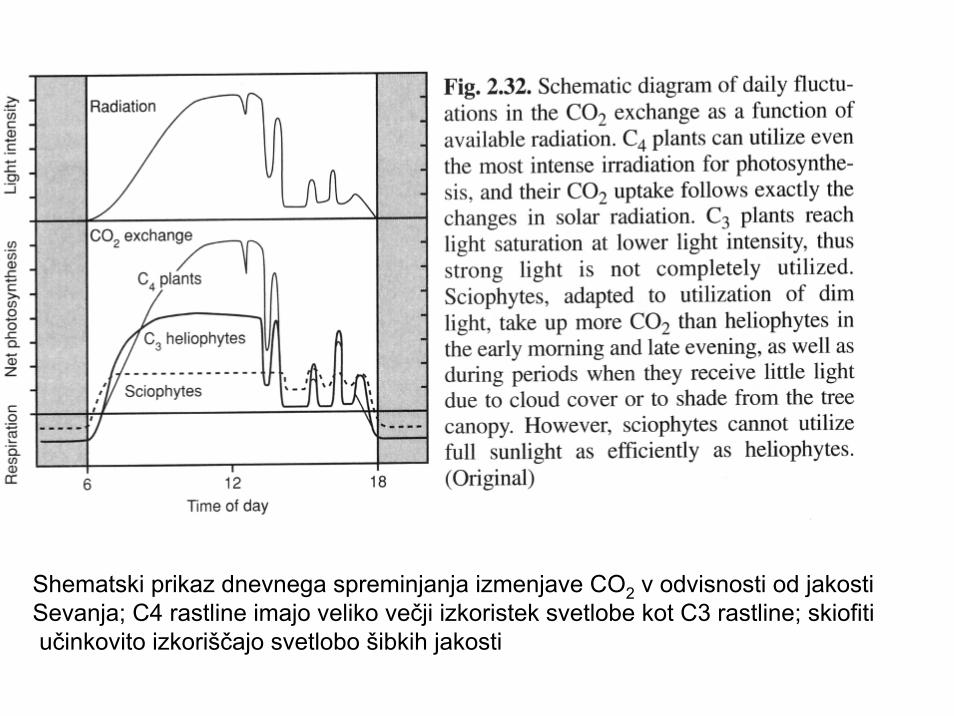
Shematski prikaz dnevnega spreminjanja izmenjave CO2 v odvisnosti od jakosti Sevanja; C4 rastline imajo veliko večji izkoristek svetlobe kot C3 rastline; skiofitiučinkovito izkoriščajo svetlobo šibkih jakosti
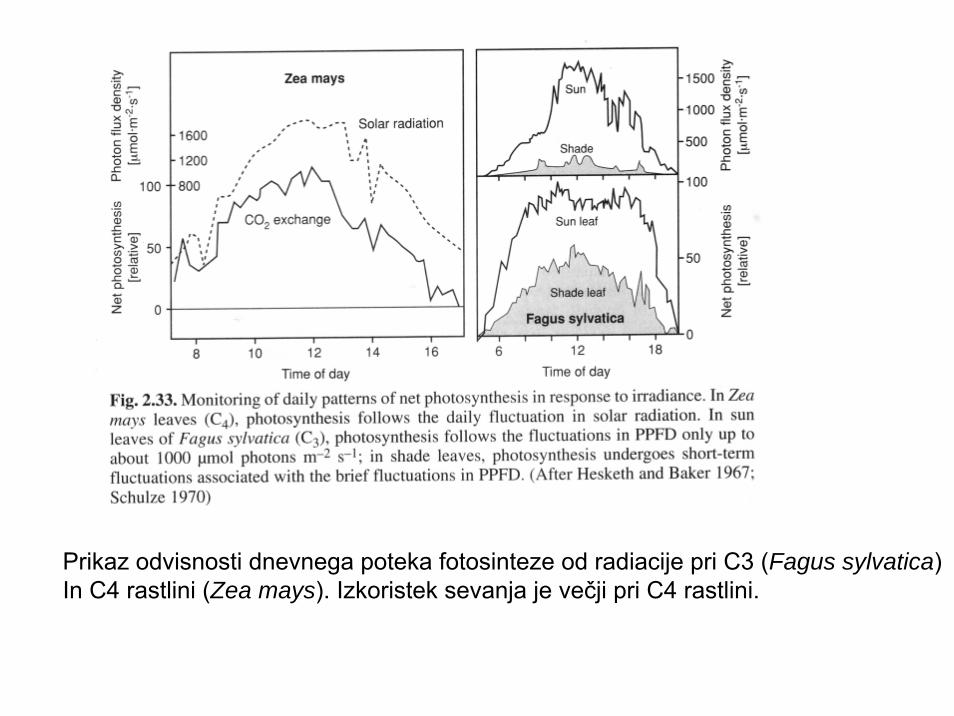
Prikaz odvisnosti dnevnega poteka fotosinteze od radiacije pri C3 (Fagus sylvatica)In C4 rastlini (Zea mays). Izkoristek sevanja je večji pri C4 rastlini.
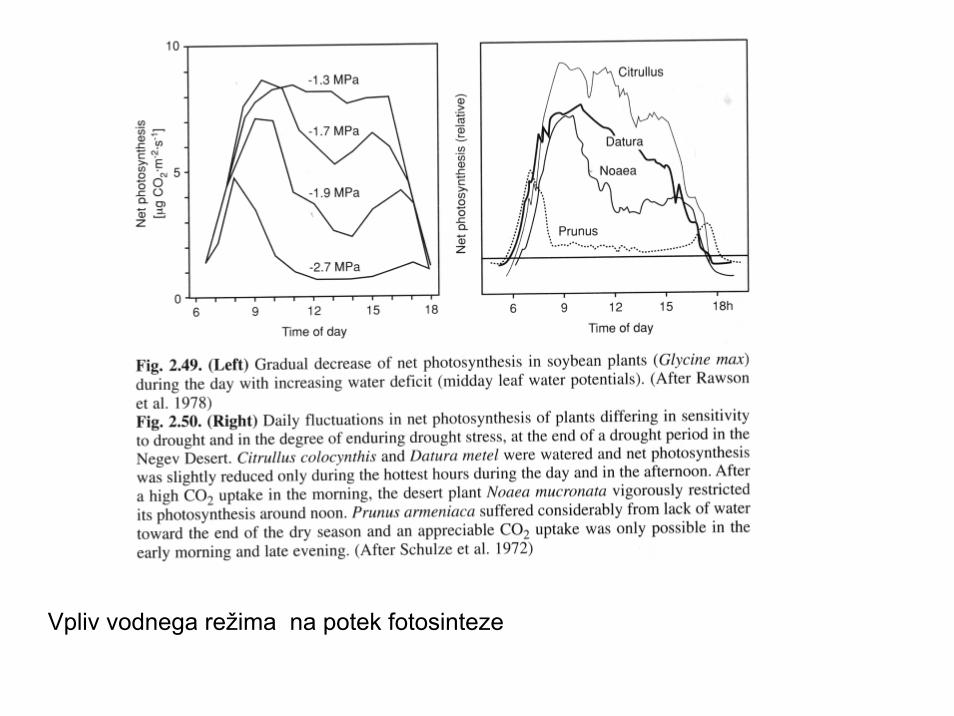
Vpliv vodnega režima na potek fotosinteze
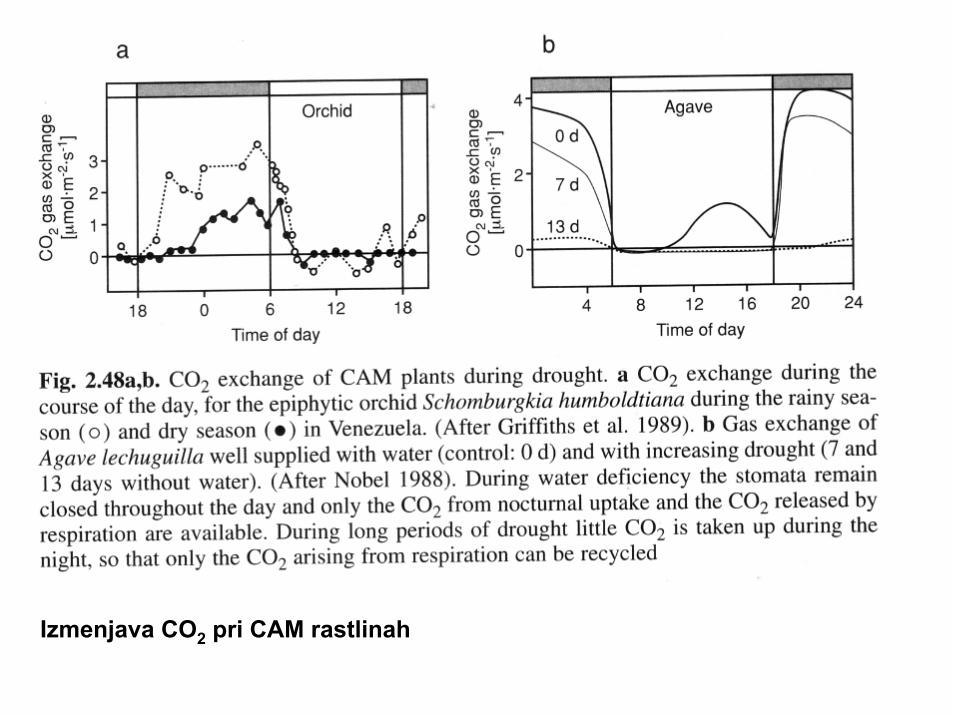
Izmenjava CO2 pri CAM rastlinah
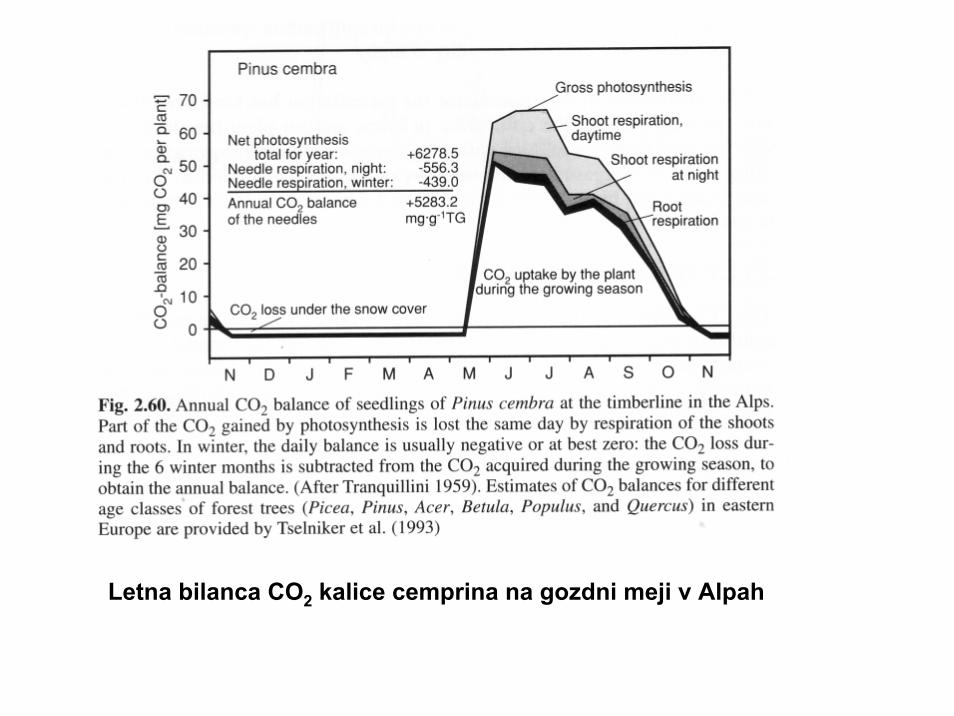
Letna bilanca CO2 kalice cemprina na gozdni meji v Alpah














