Embed Size (px)
Citation preview

Flower colour manipulation: a floral facelift Jos Mol
F l o w e r s a r e n o t o n l y a p l a n t ' s c r o w n i n g g l o r y b u t t h e y c o n t a i n its r e p r o d u c t i v e o r g a n s . C o l o u r e d p e t a l s
a n d s t a m e n s a r e v e r y a t t r a c t i v e t o p o l l i n a t o r s a n d t h e r e f o r e t r e m e n d o u s l y i m p o r t a n t f o r r e p r o d u c t i v e
success . I n s t r u c t i o n s f o r t he p r o d u c t i o n o f f l o w e r p i g m e n t s a r e e n c o d e d o n t he D N A . I t is n o w p o s s i b l e
to p r o v i d e a p l a n t w i th n e w g e n e s o r to swi tch o f f ex i s t ing o n e s , a n d g e n e t i c i s t s a r e l o o k i n g at t h e
m a n i p u l a t i o n o f f l o w e r c o l o u r s w i th f r e sh e n t h u s i a s m .
Plants grow by cell division at the grow- ing points (meristems). Whether the newly formed cells will develop into a flower depends on external signals from their surroundings, such as light, temperature, or day length, and on internal factors, such as age and hor- mone balances. Flower development starts with a reorganisation of the meris- tem into concentric groups of cells which from the outside inwards will become sepals, petals, stamens, and pistil.
It is now possible to locate the genes responsible for stimulating plant cells to produce these orderly structures. A re- cent breakthrough came with the de- velopment of the in vitro flower bud system, in which isolated meristems are induced in the test tube to form shoots, roots, or buds. This system is used both to help identify factors which promote flowering and to find and isolate genes which are switched on in the earliest stages of flower bud development (figure 1).
Each of the five types of plant hor- mone (or rather: growth regulator), namely auxin, cytokinin, gibberellin, abscisin, and ethene) play some role or other in flower formation. At least we know that the various cytokinins cause
Jos Mol, Ph.D.
Studied chemistry, specializing in molecular biology. In 1980 was appointed at the Free University, Amsterdam, where he is now Pro- fessor of molecular genetics. His main interest is in the molecular details of f lower develop- ment and reproduction.
EURO-ARTICLE. (see page ii) This article is published in association with Natuur & Techniek, The Netherlands. Translated by Anne Veerman.
Endesvour, New Series, Volume 15, No. 2, 1991. 0160-9327/91 $3.00 + 0.00. Pergamon Press plc. Printed in Great Britain.
42
different reactions in the in vitro sys- tem. Application of zeatin will, for ex- ample, cause shoot formation while kinetin stimulates bud development. A very recent discovery is the fact that oligosaccharides (small sugar polymers from the wall of the cell) will also induce flower formation in this system. This demonstrates that flower develop- ment is controlled by a whole range of interacting factors.
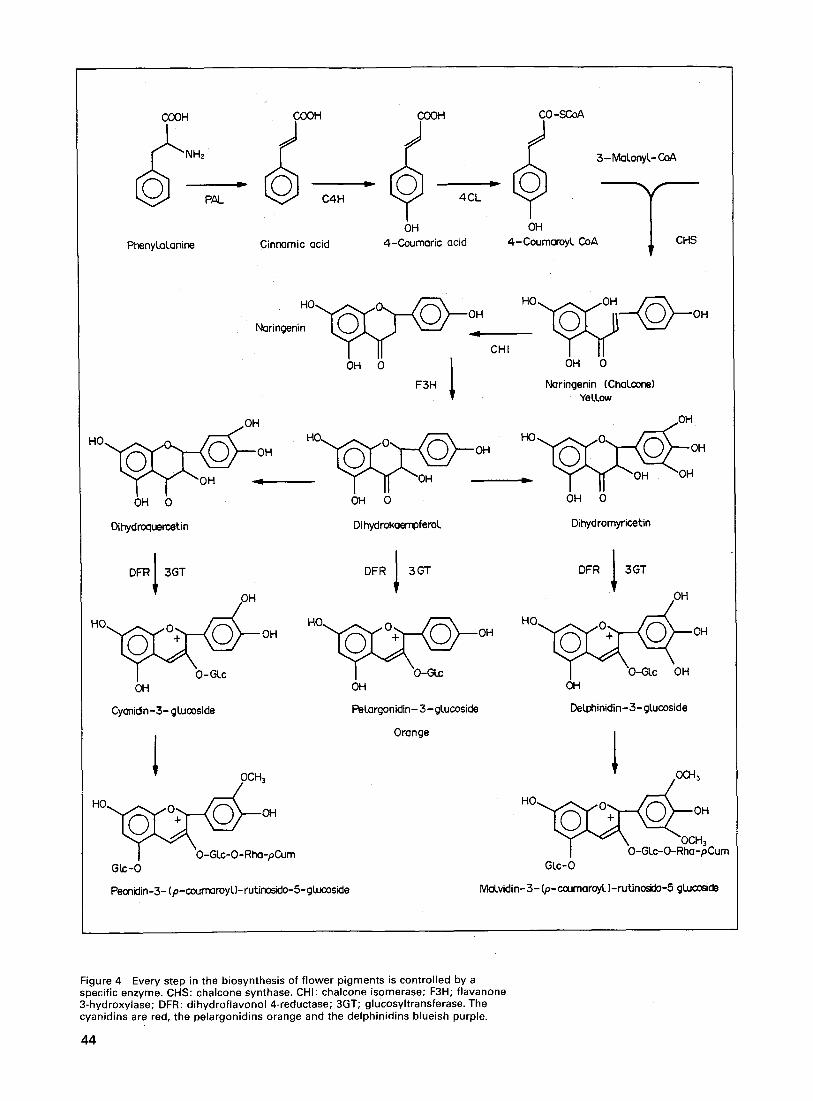
Some plants have separate male or female flowers, but most flowers con- tain both types of reproductive organ in addition to sepals and petals. These surround and protect the reproductive organs and attract the insects essential for pollination. Before this can take place, the male cells (pollen) must de- velop in the anthers of the stamens and the female cells in the ovary. Contact between pollen and stigma is one of the first steps in reproduction. To enable successful fusion of male and female gametes, a pollen grain grows a pollen tube through the style down which the male cell can pass and reach its female counterpart (figure 2(a) and (b)).
After fertilisation the flower still plays a key role in the life of the plant. Petals and sepals wilt and die, but in the protected environment of the ovary the fertilised egg cells develop into seeds. After ripening the seeds can germinate and the cycle starts all over again.
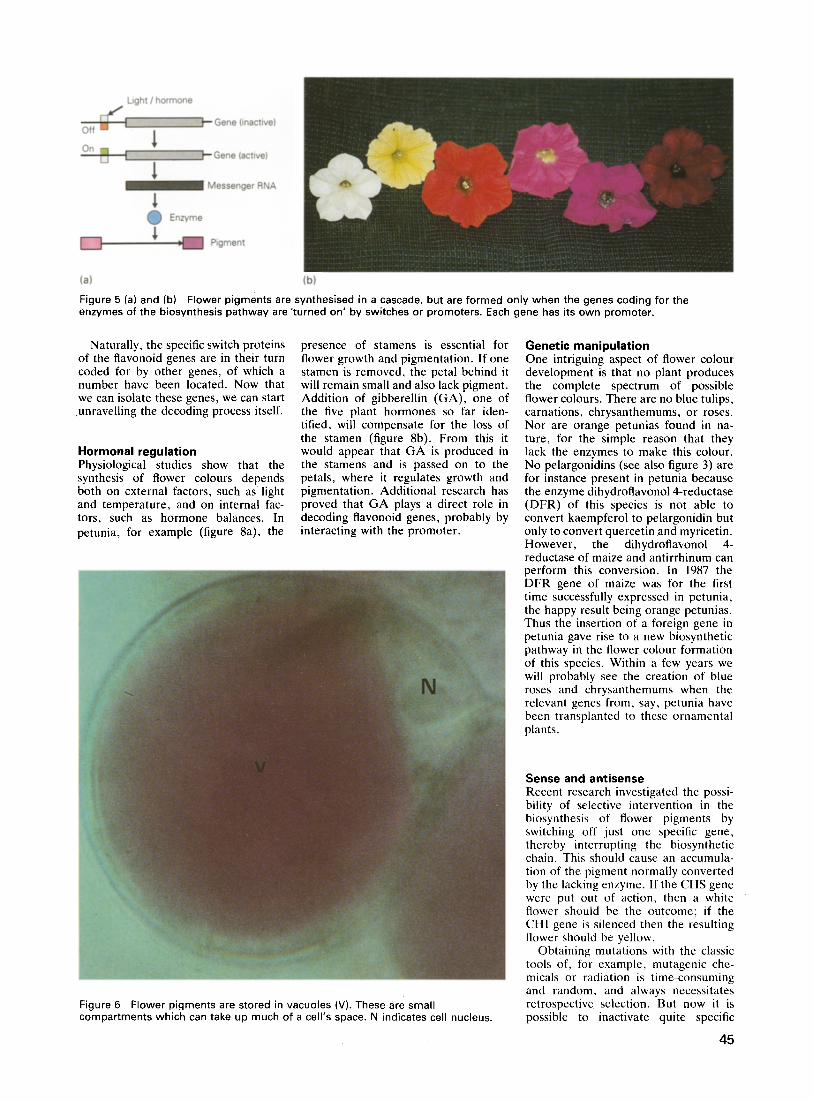
If an organism is to develop success- fully, the genetic instructions residing on the DNA must be translated into a structural plan at the right moment, in the right place, and the right way. Until recently, cell biologists could deduce this process only from various observ- able phenomena and were not able to describe it in fine detail beyond the level of the cell. The advent of recombinant DNA technology in the early 1970s opened the way for studying develop- mental processes at the DNA level. Today, many plants can also receive new genetic information through micro- surgery, using the soil microorganism Agrobacterium tumefaciens (figure 3), or through mechanical means such as
microinjection and microprojectile bombardment.
Normal flower colours and their mutants can generally be recognized at a glance, and this makes them attractive subjects for scientific research, a fact which caught the attention of Gregor Mendel, the founder of genetics, in the last century. In the first half of this century the chemical structures of flower pigments were clarified, and by now most key enzymes involved in their synthesis are also known.
The development of recombinant DNA technology has made it possible to isolate some of the most important genes controlling flower colour synth- esis and to study their structure and expression in depth. Since 1982 plants can even be provided with new genetic information. This technology has made possible novel approaches to flower (colour) breeding at the molecular level, and some spectacular successes have already been obtained.
Flower colour synthesis Flavonoids comprise the main category of flower pigments. Their structure is simple and based on the amino acid phenylalanine. Through a number of enzymatic steps phenylalanine is con- verted into coumaric acid which in its turn forms the precursor for the synth- esis of the first flavonoid, the yellow naringenin (chalcone). Many subtle modifications of this basic structure en- able flowers to produce a whole array of pigments varying in colour from light orange to deep violet. Each of these conversions depends on a specific en- zyme and therefore a vast number of genes is involved (figure 4).
Classic genetic experiments - mainly with petunia, maize, and antirrhinum - have shown that at least 35 genes play a role in producing flower colours. Some of them code for the biosynthesis en- zymes, but others control specific reg- ulatory or transport functions. This is hardly surprising, as during the de- velopment of a flower all the flower colour genes must be brought into play
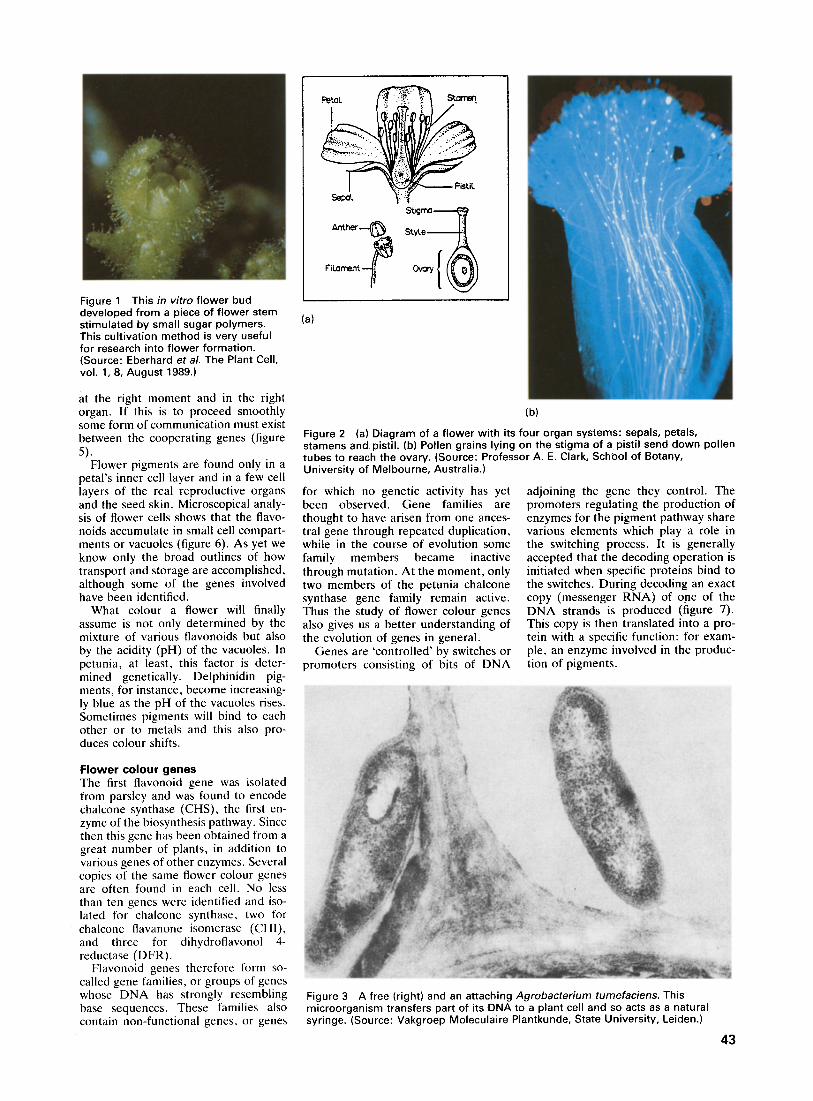
Figure 1 This in vitro flower bud developed from a piece of flower stem stimulated by small sugar polymers. This cultivation method is very useful for research into flower formation. (Source: Eberhard et al. The Plant Cell, vol. 1, 8, August 1989.)
(a)
Se L
Anther
F i ~ m e n t - . ~
$~Jgrna {
S~.yLe
Ovary
at the right moment and in the right organ. If this is to proceed smoothly some form of communication must exist between the cooperating genes (figure 5).
Flower pigments are found only in a petal 's inner cell layer and in a few cell layers of the real reproductive organs and the seed skin. Microscopical analy- sis of flower cells shows that the flavo- noids accumulate in small cell compart- ments or vacuoles (figure 6). As yet we know only the broad outlines of how transport and storage are accomplished, although some of the genes involved have been identified.
What colour a flower will finally assume is not only determined by the mixture of various flavonoids but also by the acidity (pH) of the vacuoles. In petunia, at least, this factor is deter- mined genetically. Delphinidin pig- ments, for instance, become increasing- ly blue as the pH of the vacuoles rises. Sometimes pigments will bind to each other or to metals and this also pro- duces colour shifts.
Flower colour g e n e s The first flavonoid gene was isolated from parsley and was found to encode chalcone synthase (CHS), the first en- zyme of the biosynthesis pathway. Since then this gene has been obtained from a great number of plants, in addition to various genes of other enzymes. Several copies of the same flower colour genes are often found in each cell. No less than ten genes were identified and iso- lated for chalcone synthase, two for chalcone flavanone isomerase (CHI), and three for dihydroflavonol 4- reductase (DFR).
Flavonoid genes therefore form so- called gene families, or groups of genes whose DNA has strongly resembling base sequences. These families also contain non-functional genes, or genes
(b)
Figure 2 (a) Diagram of a flower with its four organ systems: sepals, petals, stamens and pistil. (b) Pollen grains lying on the stigma of a pistil send down pollen tubes to reach the ovary. (Source: Professor A. E. Clark, SchOol of Botany, University of Melbourne, Australia.)
for which no genetic activity has yet been observed. Gene families are thought to have arisen from one ances- tral gene through repeated duplication, while in the course of evolution some family members became inactive through mutation. At the moment, only two member s of the petunia chalcone synthase gene family remain active. Thus the study of flower colour genes also gives us a better understanding of the evolution of genes in general.
Genes are 'controlled' by switches or promoters consisting of bits of DNA
adjoining the gene they control. The promoters regulating the production of enzymes for the pigment pathway share various elements which play a role in the switching process. It is generally accepted that the decoding operation is initiated when specific proteins bind to the switches. During decoding an exact copy (messenger RNA) of one of the DNA strands is produced (figure 7). This copy is then translated into a pro- tein with a specific function: for exam- ple, an enzyme involved in the produc- tion of pigments.
Figure 3 A free (right) and an attaching Agrobacterium tumefaciens. This microorganism transfers part of its DNA to a plant cell and so acts as a natural syringe. (Source: Vakgroep Moleculaire Plantkunde, State University, Leiden.)
43
COOH COOH
PAL C4H
PhenyLaLanine Cinnamic acid
COOH CO -SCoA
~ 3- Malonyl- CoA
; " T 4CL
OH OH 4-Coumaric ac id 4-CoumaroyL CoA CHS
.o~..A~oh_ ~ Naringenin ~
OH 0 1 F3H
OH
CHI OH 0
Naringenin (ChaLcone) YelLow
OH
OH
H O ' ~ ~ O ~ ~ OH
T 1" -OH -_ OH 0
Dihydrocluercetin
H O ~ ' ~ O ~ ~ - ~ OH
T 11 -OH OH 0
DihydrokoempferoL
OH
H O ' ~ O ~ OH
. " ~ "1~ "OH "OH OH 0
Dihydromyricet.in
DFR 1 3GT
OH
H O ~ _ _ ~ OH
I O-GLc OH
Cyanidin-3- gLucoside
OCH 3 HO oGlco O m Gk;-O
Peonidin-3-. (p-coumaroyU- r ut.inosido- 5- g Lucoside
/ DFR ~ 3GT
OH
OH
PeLargonidin- 3-gLucoside
Orange
DFR I 3GT
• OH
I O-GLc OH OH
DeLphinidin-3- glucoside
OH
00.i 3
OH
" r v \ -OC_,H3 I O-GLc-O-Rha-pCum
Glc-O
MaLvidin- 3 - (p- coumaroyL)-rutinosido-5 gLucoside
Figure 4 Every step in the biosynthesis of flower pigments is controlled by a specific enzyme. CHS: chalcone synthase. CHI: chalcone isomerase; F3H; flavanone 3-hydroxylase; DFR: dihydroflavonol 4-reductase; 3GT; glucosyltransferase. The cyanidins are red, the pelargonidins orange and the delphinidins blueish purple.
44
Light / hormone
O f f - - ~ l l l l i l t E j ~ Gene (inactive)
1 ~ G e n e (active)
1 Messenger RNA
O Enzyme
. ~ Pigment
(a) (b)
Figure 5 (a) and (b) Flower pigments are synthesised in a cascade, but are formed only when the genes coding for the enzymes of the biosynthesis pathway are 'turned on' by switches or promoters. Each gene has its own promoter.
Naturally, the specific switch proteins of the flavonoid genes are in their turn coded for by other genes, of which a number have been located. Now that we can isolate these genes, we can start
unravelling the decoding process itself.
Hormonal regulation Physiological studies show that the synthesis of flower colours depends both on external factors, such as light and temperature, and on internal fac- tors, such as hormone balances. In petunia, for example (figure 8a), the
presence of stamens is essential for flower growth and pigmentation. If one stamen is removed, the petal behind it will remain small and also lack pigment. Addition of gibberellin (GA), one of the five plant hormones so far iden- tified, will compensate for the loss of the stamen (figure 8b). From this it would appear that G A is produced in the stamens and is passed on to the petals, where it regulates growth and pigmentation. Additional research has proved that G A plays a direct role in decoding flavonoid genes, probably by interacting with the promoter.
Genetic manipulation One intriguing aspect of flower colour development is that no plant produces the complete spectrum of possible flower colours. There are no blue tulips, carnations, chrysanthemums, or roses. Nor are orange petunias found in na- ture, for the simple reason that they lack the enzymes to make this colour. No pelargonidins (see also figure 3) are for instance present in petunia because the enzyme dihydroflavonol 4-reductase (DFR) of this species is not able to convert kaempferol to pelargonidin but only to convert quercetin and myricetin. However, the dihydroflavonol 4- reductase of maize and antirrhinum can perform this conversion. In 1987 the DFR gene of maize was for the first time successfully expressed in petunia, the happy result being orange petunias. Thus the insertion of a foreign gene in petunia gave rise to a new biosynthetic pathway in the flower colour formation of this species. Within a few years we will probably see the creation of blue roses and chrysanthemums when the relevant genes from, say, petunia have been transplanted to these ornamental plants.
Figure 6 Flower pigments are stored in vacuoles (V). These are small compartments which can take up much of a cell's space. N indicates cell nucleus.
Sense and antisense Recent research investigated the possi- bility of selective intervention in the biosynthesis of flower pigments by switching off just one specific gene, thereby interrupting the biosynthetic chain. This should cause an accumula- tion of the pigment normally converted by the lacking enzyme. If the CHS gene were put out of action, then a white flower should be the outcome; if the CHI gene is silenced then the resulting flower should be yellow.
Obtaining mutations with the classic tools of, for example, mutagenic che- micals or radiation is time-consuming and random, and always necessitates retrospective selection. But now it is possible to inactivate quite specific
45

Promoter
c ~ ( ; -
Anti-sense __T-- ~ DNA strond DNA strand ~_
Direction of ~-- ~;;:"
I V i a ~ U ~ n O r ; bi~°gicaL
Direct.ion of tronscripUon
DoubLe-stranded l~lA
4 ~ :.~! :i!~!:..:ii
x~ ";~ ,:::::
Messenger (sense) RNA
!.E
Antisense RNA $ G 0 G C 4 A
Protein
DoubLe-stronded DNA
~ ~ G
~ : ~ " ~",'~A
No protein
No protein
Figure 7 The principle of sense and antisense. If a DNA strand produces messenger RNA and the wrong, complementary strand of a second, identical gene is copied at the same time, double-stranded RNA may be formed. This is genetically inactive as it cannot be translated into a protein such as a pigmentation enzyme.
genes by inactivating their own particu- lar messenger RNA as the latter can then no longer be translated into pro- teins: This RNA elimination is achieved with the so-called antisense technique.
Only one of the two complementary (fitting together) DNA strands is trans- cribed into messenger RNA. This RNA is translated directly into a protein by ribosomes and is therefore called 'sense' RNA. As this sense RNA is the comple- ment of one of the two DNA strands, the coding DNA strand is called the 'antisense' strand. The non-coding or sense strand of DNA can in principle produce antisense RNA, but this cannot code for a protein. If, however, a situa- tion is created in which both sense and antisense RNA are present, then they can merge to form a double-stranded RNA chain. Ribosomes cannot trans- late such structures into a protein and the final effect thus resembles a muta- tion in which the gene has lost its func- tion.
Generally speaking, this antisense effect can be brought to bear on any gene through genetic manipulation. For this purpose a gene in a bacterium is turned back to front with respect to its promoter by means of recombinant DNA techniques. This antisense gene is then introduced into a plant. In addition
46
to the normal gene the plant will now also transcribe the wrong strand. The resulting antisense RNA can then in- activate the normal messenger RNA.
I f we block the CHS gene in this way the effect will be directly visible. Plants expressing an antisense CHS gene dis- play flower colour intensities ranging from normal to fully white (figure 9). This is probably caused by the fact that the antisense gene, introduced with the aid of Agrobacterium tumefaciens, is incorporated in a more or less random fashion in one of the plant's chromo- somes. Depending on its position, the expression of the transferred gene will be strong or weak and will inhibit pig- mentation to a greater or lesser degree.
Rings and stars After antisense treatment colour pat- terns such as rings and stars appear in the flower. These features are extreme- ly sensitive to light, temperature fluctuations, and hormone treatments, for example with GA. Patterns are also known from 'natural' varieties of petu- nia, violet, and impatiens but they have never been seen in cut flowers such as rose, carnation, and gerbera. Thanks to antisense techniques it is feasible to grow two-coloured cultivars of these plants too.
If more of the same flower colour genes (sense) that the plant already possesses are added, this would be ex- pected to lead to increased enzyme pro- duction manifested in higher pigment concentrations. In practice, however, the effect is quite opposite and the same, sometimes complete, reduction of coloration is observed as when anti- sense genes are added. Occasionally pattern formation also results (although the sense and antisense patterns may differ).
The results of sense and antisense experiments clearly show that the level of CHS gene expression is regulated very precisely. Both increase and reduc- tion cause fairly severe disturbance. In all probability the sense technique will also help create new (two-) coloured cut flower varieties.
Longevity It has for some time been known that aging flowers of cultivated species such as petunia and carnation produce ethene (ethylene, C2H4) and that ethene speeds up the aging process. Not only the production of ethene but also sensitivity to this substance seems to increase with age. This means that aging of flowers can be retarded by inhibitors of ethene biosynthesis or by substances which block the effects of ethene but also sensitivity to this substance seems to increase with age. This means that aging of flowers can be retarded by inhibitors of ethene biosynthesis or by substances which block the effects of ethene. Not all flower species produce ethene during senescence, exceptions being, among others, chrysanthemum and gerbera, which in any case proved insensitive to ethene. Therefore a diffe- rent aging mechanism is at work in their flowers.
Pollination frequently increases the rate of aging. In some orchid species the flowers survive for only a few hours after pollination. As pollination entails touching of stamens and pistil, some signal from these organs must reach the petals to trigger wilting. In petunia, ethene production is stepped up after pollination, which explains the rapid aging of the flower. Recent studies show that growing pollen tubes stimulate the production of ACC synthase in the style (ACC is a biochemical precursor of ethene). This ACC is then transported to the petals and there converted to ethene.
The antisense technique enables us to prolong the life of ornamental plants. This is achieved by blocking ethene biosynthetic genes on the one hand and by preventing the synthesis of ethene receptors (b!nding sites of ethene) on the other.
The advent of gene isolation, man- ipulation, and transformation heralds a new era of molecular flower breeding

Figure 8 (a) A natural variety of petunia: red star. (b) A petunia from which the stamens have been removed in the bud stage suffers from growth and pigmentation defects (middle). Spraying with the plant hormone GA leads to recovery (right).
(a) (b)
for colour, Manipulation of the genes involved in flower pigmentation and longevity will result in more w~rieties with new colours and longer life.
Some production characteristics such as synchronisation and timing of flower- ing are regulated by hormones. Genetic manipulation of these developmental processes will have to wait until we have a more fundamental knowledge of the hormonal regulation involved.
Not all cultivated plants are good
subjects for gene transfer with the aid of A. tumeJ~wiens. The development of mechanical methods for these recalcit- rant species (bombardment with DNA- coated gold pellets or micro-injection) is a good alternative (figure 10). Resear- chers must have 'green fingers' to raise adult plants from plant cells provided with a new gene. The right type of tissue in the right developmental stage must be placed in a perfectly balanced growth medium to obtain shoot formation.
Even today, little success has been re- ported with some species, including carnations.
The possible dangers of introducing transgenic plants into nature are still under debate, as is the discrepancy which exists between patent law and the plant breeder's rights used in this field up to now. Despite these complicating factors the first transgenic plants will probably appear on the market within the next five years.
47

Figure 9 The var ious patterns in these f lowers were obta ined by int roducing ant isense (a) and sense (b, c) CHS genes.
Bibliography [1] Botterman, J. 'Gcnmanipulatic van plan-
ten' , Natuur & Techniek, 57, No. 5, 378-387, 1989.
[2] Broertjes, C. 'Veredeling van siergewas- sen Bloemkleurcn uit het laborator- ium', Natuur & Techniek, 49, No. 6, 42(~439, 1981.
[3] Drews, G. and Goldberg, R. 'Genetic control of flower development ' . Trends in Genetics, 5,256-261, 1989.
[4] Lindhout, P. and Kramcr, Th. 'Tomaten - Van massaselectie tot manipulatie' , Natuur & Techniek, 58, No. 5,352-364, 1990.
[5] Mol. J., c.s. "Saying it with genes: molecular flower breeding' , Trends in Biotechnology, 7, 148-153, 1989.
[6] Voesenek, L. A. C. J., Harren, F. J. M. and Woltering, E. J. 'H2C/CH2 - Een gas als hormoon' , Natuur & Techniek, 58, No. 3, 2114-215, 1990.
[7] Weintraub, H. M. 'Antisense RNA and DNA' , Scient. Am., 262, No. 1, 34-40, 199(I.
Figure 10 The DNA-gun. In this technique a special gun is used to shoot smal l gold or p la t inum part icles covered wi th DNA wi th great force into l iv ing cells. The effects can be dramat ic, as seen in this maize kernel (b) which was peppered wi th a lacking-colour gene. Colour fo rmat ion is resumed in cells that were hit (a).
48





























