Embed Size (px)
Citation preview

1
FILOGENIA MOLECULAR Y GENÉTICA DE POBLACIONES DE Alouatta palliata E
INFERENCIAS FILOGENÉTICAS AL INTERIOR DEL GENERO ALOUATTA CON
SECUENCIAS DE ADN MITOCONDRIAL
ÁNGELA MARÍA CERÓN PATÍO
TRABAJO DE GRADO
Presentado como requisito parcial para optar al título
MAGÍSTER EN CIENCIAS BIOLÓGICAS
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
MAESTRÍA EN CIENCIAS BIOLÓGICAS
Bogotá, D. C.
2015

2
FILOGENIA MOLECULAR Y GENÉTICA DE POBLACIONES DE Alouatta palliata E
INFERENCIAS FILOGENETICAS AL INTERIOR DEL GENERO ALOUATTA CON
SECUENCIAS DE ADN MITOCONDRIAL
ÁNGELA MARÍA CERÓN PATÍO
APROBÓ
______________________________ ______________________________
Concepción Judith Puerta Bula PhD Alba Alicia Trespalacios Rangel PhD
Decana Directora Posgrado
Facultad de Ciencias Facultad de ciencias

3

4
NOTA DE ADVERTENCIA
“La Universidad no se hace responsable de los conceptos emitidos por sus alumnos en sus trabajos de
tesis. Solo velará por qué no se publique nada contrario al dogma y a la moral católica y porque la
tesis no contenga ataques personales contra persona alguna, antes bien se vea en ellas el anhelo de
buscar la verdad y la justicia”.
Artículo 23 de la Resolución N° 13 de julio de 1946.

5
AGRADECIMIENTOS
Al Doctor MANUEL RUIZ-GARCÍA que con mucha paciencia me ha permitido hacer parte de
este proyecto y me ha enseñado sobre la genética de poblaciones.
A MYREYA OMAIRA PINEDO CASTRO por haber sido, ser, y espero siga siendo, mi amiga y
maestra en este camino interminable del conocimiento.
A mi familia, en especial a mi papá y mi mamá, ya que sin su apoyo nada de esto hubiera sido
posible.
A mis grandes amigos, gracias por su apoyo.
A la vida, por permitirme crecer como persona integral.

6
TABLA DE CONTENIDO
1. RESUMEN……………………………………………………………………………….................7
2. INTRODUCCIÓN…………………………………………………………………………………..8
3. JUSTIFICACIÓN………………………………………………………….……………………….9
4. MARCO TEÓRICO………………………………………………………………….................…10
4.1 El género Alouatta..........................................................................................................................10
4.1.1Alouatta palliata…………………………………………………………………………...........13
4.1.2Alouatta seniculus……………………………………………………………………………….14
4.1.3Alouatta caraya………………………………………………………………………...……….15
4.1.4Alouatta pigra…………………………….…………………………………….……………….16
4.1.5 Alouatta sara……………………………………………………………...…………...…….…18
5. OBJETIVOS……………………………………………………………………………….…..….19
5.1 Objetivo general…………………………..……………………………………………….......…19
5.2 Objetivos específicos……….…………………………………………………….................……19
6. MATERIALES Y MÉTODOS…………………………………….…………………………...….19
6.1 Obtención de muestras, extracción de ADN, PCR y secuenciación……………..………….........19
6.2 Alineamiento de secuencias de ADN mitocondrial………………………………………...…….22
6.3Análisis filogenéticos……………………………………………..………………………………22
6.4Análisis de diversidad, heterogeneidad y flujo génico...………………………………………….22
6.5Detección de cambios demográficos…………..……………………………………………….…23
7. RESULTADOS…………………………………………………………………………..………..24
7.1 Filogenia………………………………………………………………………………………….24
7.2 Diversidad genética, heterogeneidad genética y flujo génico…..…………………...…………...34
8. DISCUSIÓN Y CONCLUSIONES………………………………………………………………..40
8.1 Filogenia……………………………………………………………………………………….38
8.2 Diversidad genética, heterogeneidad genética y flujo génico…..…………………...………...40
8.3 Cambios demográficos y tiempos de divergencia……………………………………………..41
9. REFERENCIA BIBLIOGRÁFICA...........………………………………………………………...43

7
1. RESUMEN
Para llevar a cabo este trabajo, fueron secuenciadas para el gen mitocondrial COII148 muestras
del género Alouatta (monos aulladores), A. palliata (103), A. coibensis trabeata (3), A. pigra (3),
A. caraya (1), A. sara (6), A. seniculus (11), siendo A. palliata el objetivo principal de este
estudio. Adicionalmente se utilizaron muestras de Lagothrix lagotricha cana (11) como
outgroups. A partir de las secuencias obtenidas se realizó un análisis de las relaciones
filogenéticas. Se analizó si los datos moleculares obtenidos permiten diferenciar las especies
propuestas, se evaluó el estado genético de las poblaciones (diversidad genética, heterogeneidad
genética y flujo génico), y además, se indagó sobre su historia evolutiva (se calcularon tiempos
de divergencia y cambios demográficos de la especie y se relacionaron con posibles causas
climatológicas y geológicas).
Los principales resultados obtenidos fueron los siguientes:
1- A. palliata muestra bajos niveles de diversidad genética mitocondrial respecto a otras
especies de Alouatta.
2- Las muestras que representaron a Alouatta coibensis trabeata (considerada previamente
como subespecie de Allouatta palliata y hoy en día considerada una especie diferente),
ubicadas en la República de Panamá, son indiferenciables molecularmente respecto a la
especie Alouatta palliata.

8
3- Las subespecies Allouatta palliata ssp. palliata (Costa Rica) y Alouatta palliata ssp.
aequatorialis (Colombia y Ecuador) no son diferentes molecularmente, sus diferencias
radican en lo morfológico ya que se deben adaptar a diferentes ambientes. Por lo tanto,
los estudios moleculares no ratificaron esas dos subespecies morfológicas.
2. INTRODUCCIÓN
Durante los últimos 30 años se ha incrementado significativamente el desarrollo de las
investigaciones primatológicas y, paralelamente se han multiplicado los trabajos de campo.
Numerosas características de estos animales son atractivas para nosotros los humanos, que nos
impulsan a querer saber más acerca de ellos. En el nivel más accesible para la mayoría, su
apariencia física llama nuestra atención y nos divierten sus comportamientos complejos y de
varias facetas. En realidad nos recuerdan a nosotros mismos, en un contexto serio o cómico, y en
muchos casos de una manera que para algunos es profundamente inquietante, sobre todo porque
los primates son nuestros parientes más cercanos en el reino animal. (Defler, 2004).
El Orden Primates lo constituyen 237 especies, pertenece a uno de los 18 órdenes dentro de la
Clase Mammalia (Cheney et al., 1986), En este estudio, se analizaran algunas especies de un
género en particular (Allouata), las especies que se trabajaran son A. palliata, A. seniculus, A.
sara, A. pigra y A. caraya. Se obtuvieron secuencias de los genes mitocondriales para 148
muestras. Se hace importante determinar e inferir las características moleculares de las especies,
mediante la construcción de árboles filogenéticos de máxima verosimilitud, Neighbour-Joining,
y bayesianos, además se exploraron las relaciones de parentesco donde se puede vislumbrar y
entender en mayor medida su comportamiento morfológico y geográfico.

9
3. JUSTIFICACIÓN
Los primates del género Alouatta comprenden un gran grupo con varias especies establecidas
a lo largo de Centroamérica y Suramérica. Se han realizado varios estudios para indagar la
evolución de su estructura filogenética y su comportamiento de expansión poblacional. Por
ello se hace muy interesante poder ahondar aún más dentro de su filogenia a través de sus
genes mitocondriales para reconocer los posibles hechos que dieron lugar a dicha evolución.
Los estudios moleculares son herramientas útiles para generar información acerca de la
historia evolutiva de un grupo o especie en particular y ratificar el estado genético de sus
poblaciones con miras a la conservación; permitiendo además, detectar especies crípticas, los
cuales no son reconocidas de manera morfológica.

10
4. MARCO TEORICO
4.1 Género Alouatta
Las especies pertenecientes al género Alouatta tienen nombres comunes como “Aullador” y
“mono cotudo”, el primero de ellos hace alusión a su fuerte vocalización, y el segundo a su
abultada garganta que resulta del exagerado crecimiento del hioides que le sirve de cámara de
resonancia. Los miembros de este género son especies neotropicales de gran talla, los adultos
pueden pesar entre 6-10 Kg, poseen una fuerte cola prensil capaz de soportar el peso de su
cuerpo. (Defler, 2004).
En el estudio de Cortés-Ortiz et al., (2003), se habla de que su distribución es bastante amplia, se
encuentran desde el sur de México hasta el norte de Argentina (fig. 1), y desde la costa pacífica
hasta la selva atlántica del Brasil, se puede encontrar desde el nivel del mar hasta los 3.200
msnm, aunque diferentes especies varían en las preferencias de hábitat (Crockett,
1998).Tradicionalmente los monos aulladores fueron clasificados en 6 especies: A. palliata, A.
pigra, A. seniculus, A. belzebul, A. fusca y A. caraya (Hill, 1962; Hershkovitz, 1972). Varias
subespecies fueron elevadas al rango de especie según dos revisiones sistemáticas posteriores
(Rylands et al., 2000; Groves, 2001; 2005; Gregorin, 2006; Rylands y Mittermeier, 2009).
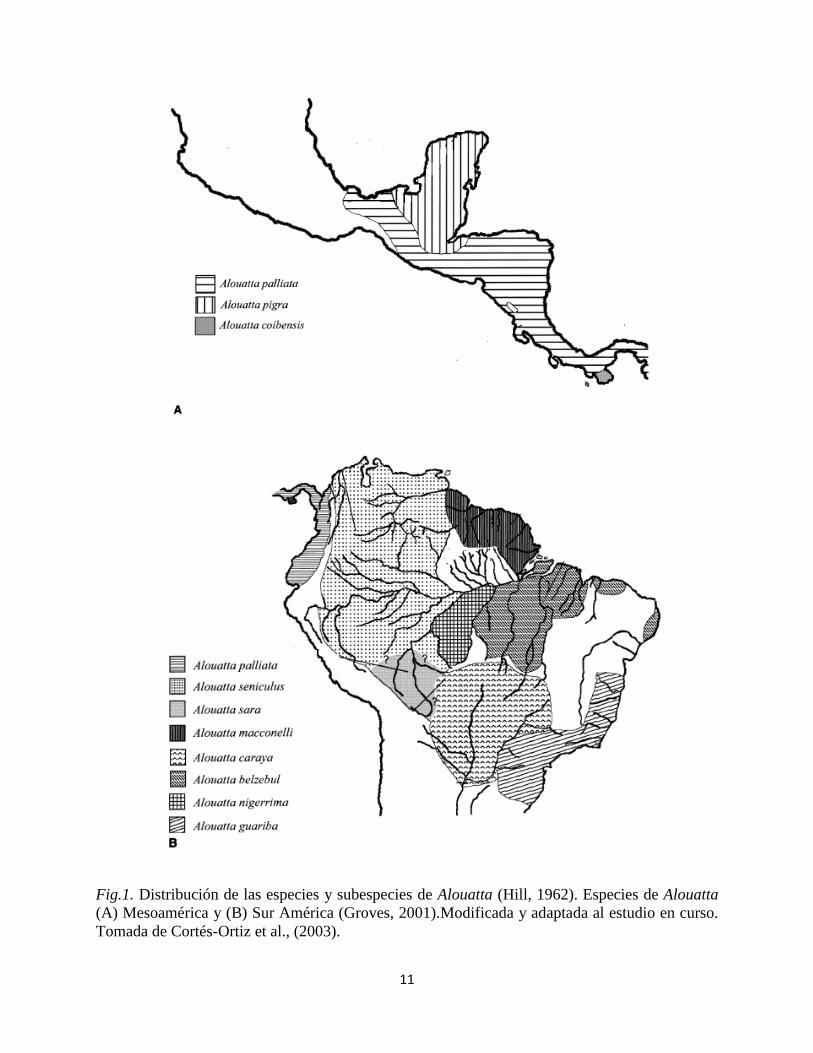
11
Fig.1. Distribución de las especies y subespecies de Alouatta (Hill, 1962). Especies de Alouatta
(A) Mesoamérica y (B) Sur América (Groves, 2001).Modificada y adaptada al estudio en curso.
Tomada de Cortés-Ortiz et al., (2003).

12
Hill 1962 (como se citó en Cortés-Ortíz et all 2003) propone que el género Alouatta está
compuesto de cinco o seis especies; pero los más recientes estudios taxonómicos reconocen entre
9 y 10 especies y hasta 19 subespecies (Groves 2001; Rylands et al., 1995, 2000).
Según los autores se distribuyen de la siguiente manera.
En la (Fig.1A) se muestran las tres especies de Mesoamérica, (1) A. palliata con tres
subespecies, las cuales se encuentran distribuidas desde México hasta Ecuador; (2) A. pigra
restringida a la vertiente del Caribe, desde México hasta el norte de Honduras; (3) A. coibensis
con dos subespecies, una se encuentra en el pacífico de la isla de Coiba y la segunda se
encuentra en la Península de Azuero (Panamá).
En la (Fig. 1B) se encuentran las siete especies restantes en Sudamérica, (1) A. seniculus la cual
tiene de tres a seis subespecies y su distribución limita con el occidente de la cordillera de los
andes en Colombia, y abarca gran parte del norte de Sur América por encima del rio Amazonas.
(2) A. maconnelli (no se reconoce en la taxonomía propuesta por Rylands et al., 2000) se
encuentra en Surinam, Guyana, Guyana Francesa, y Brasil, al norte del Amazonas. (3) A. sara, al
parecer se restringe a la Amazonía Boliviana y sur de Perú; (4) A. belzebul con tres subespecies,
su distribución se extiende desde el sur del tallo principal del Rio Amazonas, y al este del Rio
Tapajos en la costa norte del bosque Atlántico de Brasil; (5) A. nigerrima, delimitada al norte del
Rio Amazonas, al este y oeste de los ríos Trombetas y Madeiras, respectivamente, y al sur en su
límite parapátrico con Alouatta caraya; (6) A. guariba, con dos subespecies distribuidas
alopátricamente en la mitad entre el norte y el sur del bosque Atlántico de Brasil; y (7) A. caraya
distribuido al sur de Brasil, Bolivia, Paraguay y el norte de Argentina.
Entre los primates del nuevo mundo, el género Alouatta se ha estudiado ampliamente en cuanto a
la ecología (Rodríguez-Luna et al., 1993; Estrada et al., 1999; Rodríguez-Luna, 2000; Zunino et

13
al., 2001). Se adaptan muy fácilmente ya que aprovechan los hábitats de acuerdo a lo que
encuentren en ellos, pueden estar en ambientes de selva, xerófilos y con distintos sistemas
sociales (Crockett & Eisenberg, 1987). Una característica relevante es que ambos sexos pueden
dispersarse del grupo natal (Crockett, 1984; Rumiz, 1990).
4.1.1 Alouatta palliata
Comúnmente llamado “aullador negro”, “mono negro” o “mono cotudo”, es un primate de gran
tamaño, robusto; la longitud cabeza-cuerpo varía entre 48.1 y 67.5 cm, con un valor medio para
los machos de 56.1 cm y para las hembras de 52 cm; la longitud cabeza cola varía entre 54.5 cm
y 65.5 cm con un valor medio para los machos de 58.3 cm y para las hembras de 60.9 cm. En su
mayoría son negros, pero tienen en su espalda, costados y hombros un pelaje de color marrón
claro o acanelado. Su hueso hioides está altamente desarrollado y actúa como caja de resonancia
que les permite emitir sus famosos llamados o aullidos. En términos generales consumen hojas y
frutos en cantidades aproximadamente iguales, aunque también consumen flores. Se encuentran
en grupos sociales de entre 6 a 23 individuos, hay usualmente 2 o 3 adultos machos (Defler,
2003).Se distribuye desde México hasta la costa oeste de Colombia y Ecuador (Crockett &
Eisenberg, 1987). Usualmente se encuentran en el estrato medio a alto de árboles altos, en
bosques primarios, siempre verdes o ripários, con varios grados de intervención (Neves &
Rylands, 1991). Se encuentra en el apéndice I de CITES, y es considerado por la UICN como
“LC” (preocupación menor) a nivel global, aunque las poblaciones colombianas han sido
colocadas en la categoría VU (vulnerable) a nivel nacional siguiendo los mismos criterios
(Defler, 2003).

14
Figura 2. Distribución geográfica de A. palliata. <www.iucnredlist.org>.
4.1.2 Alouatta seniculus
Comúnmente llamado “mono colorado”, “mono cotudo”, “roncador”, “araguata”, es uno de los
primates más grandes de Colombia, alcanzando la longitud cabeza-cuerpo de 43.9 cm a 69.0 am
y 54 cm a 79 cm para la cola. Los machos tienen un peso promedio de 7.5 Kg y las hembras de
6.3 Kg. Su pelaje suele ser caoba rojizo, otros pueden exhibir una tonalidad dorada rojiza en la
espalda, el lomo y la porción distal de la cola. Su cabeza es grande, la cara desnuda y de color
negro, al igual que todos los de su género, este primate sobresale por su sonora vocalización,
IUCN (International Union for Conservation of Nature) 2008. Alouatta palliata. The IUCN
Red List of Threatened Species. Version 2015-3

15
siendo el macho uno de los más potentes en la naturaleza. Poseen una fuerte cola prensil, la cual
exhiben mientras forrajean (Defler 2003).
Esta especie se distribuye desde el norte de los Andes colombianos, partiendo de la cordillera
Occidental hacia el oriente y sur, y abarca Venezuela, la isla de Trinidad, el oriente de las
Guayanas, norte de la Amazonia brasileña, Ecuador, Perú y el oeste del Purus en el Brasil y
Bolivia (Emmons y Feer 1997). El 20% del área de la distribución de esta especie está en
Colombia (Defler 2003). El tamaño de los grupos puede variar desde 2 hasta más de 16
individuos, con un tamaño promedio de 6-9 individuos (Crockett & Eisenberg, 1987).
4.1.3 Alouatta caraya
Comúnmente llamado “aullador Negro”, “manechi”, “manechi Negro” (www.iucnredlist.org)es
un primate que vive en las copas de los árboles, en grupos organizados de 3 a 20 individuos con
un macho dominante. Es un primate grande y robusto cuya longitud de tronco-cabeza de 65 cm y
la longitud de la cola es de 70 cm aproximadamente. Su peso en un adulto macho es de 9 Kg y
una adulta hembra es de 5 Kg. Viven en selva de tierra firme y de inundación, monte de zona
tropical y subtropical. Su distribución está en áreas de Brasil, Paraguay y Bolivia. (Bruno 2011).
Por sus características alimentarias son animales folívoros-frugívoros generalistas y de hábitos
diurnos, poseen un largo proceso de digestión (Chivers and Hladik, 1980; Zunino, 1986).
16
Figura 3. Distribución geográfica de A. caraya. <www.iucnredlist.org>.
4.1.4 Alouatta pigra (Lawrence, 1933)
Comúnmente llamado “Araguato De Guatemala”, “Mono Aullador Negro”, “Saraguato”,
“Saraguato Negro”” (www.iucnredlist.org),este primate se encuentra en la península de Yucatán
en México y Belice, y se extiende por el centro y norte de Guatemala. Este primate se encuentra
IUCN (International Union for Conservation of Nature) 2008. Alouatta caraya.
The IUCN Red List of Threatened Species. Version 2015-3
17
principalmente en selva tropical, zonas rivereñas inundadas, en bosques primarios y secundarios.
Comen hojas y frutos, y sus dientes están especialmente adaptados para ello; tienen cola prensil,
su rasgo más característico es que tienen una fuerte cámara de resonancia la cual hacen sonar
muy temprano en la mañana y se puede oír a casi aproximadamente 2.1 Km de distancia.
(Drubbel y Gautier 1993).
Figura 4. Distribución geográfica de A. pigra. <www.iucnredlist.org>.
IUCN (International Union for Conservation of Nature) 2008. Alouatta pigra. The
IUCN Red List of Threatened Species. Version 2015-3

18
4.1.5 Alouatta sara
Comúnmente llamado “Aullador Rojo”, “Aullador Rojo”, “Manechi”, “Manechi Colorado”, es
reconocida como una especie completa. ” (www.iucnredlist.org),se encuentra en Bolivia, se
extiende desde el sur del departamento de Pando a lo largo de la Cordillera de los Andes, y al
este en el centro de Bolivia que incluye la cuenca del rio Mamoré y Beni. Esta especie se
encuentra en bosques tropicales teniendo las mismas características de todos los de su género.
Figura 5. Distribución geográfica de A. sara. <www.iucnredlist.org>.
IUCN (International Union for Conservation of Nature) 2008. Alouatta sara. The
IUCN Red List of Threatened Species. Version 2015-3

19
5. OBJETIVOS
5.1 Objetivo general
Determinarla estructura genética de A. palliata y sus relaciones filogenéticas con otras
especies de Alouatta mediante el marcador mitocondrial COII.
5.2 Objetivos específicos
Determinar los niveles de diversidad genética y heterogeneidad mitocondrial en Alouatta
palliata.
Determinar posibles cambios demográficos durante la historia evolutiva de A. palliata.
Determinar si existen diferencias moleculares apreciables entre las subespecies
morfológicas putativas de A. palliata.
Analizar las relaciones filogenéticas entre A. palliata y otras especies de monos
aulladores.
Estimar relaciones entre los tiempos de divergencia entre los taxones estudiados con
respecto a cambios climáticos y geológicos durante el Mioceno, Plioceno y Pleistoceno.
6. MATERIALES Y METODOS
6.1 Obtención de muestras, extracción de ADN, PCR y secuenciación.
Para la investigación se utilizaron 148 muestras del género Alouatta, con orígenes geográficos
conocidos. Se utilizaron muestras de Lagothrix como outgroups. (Tabla 1).
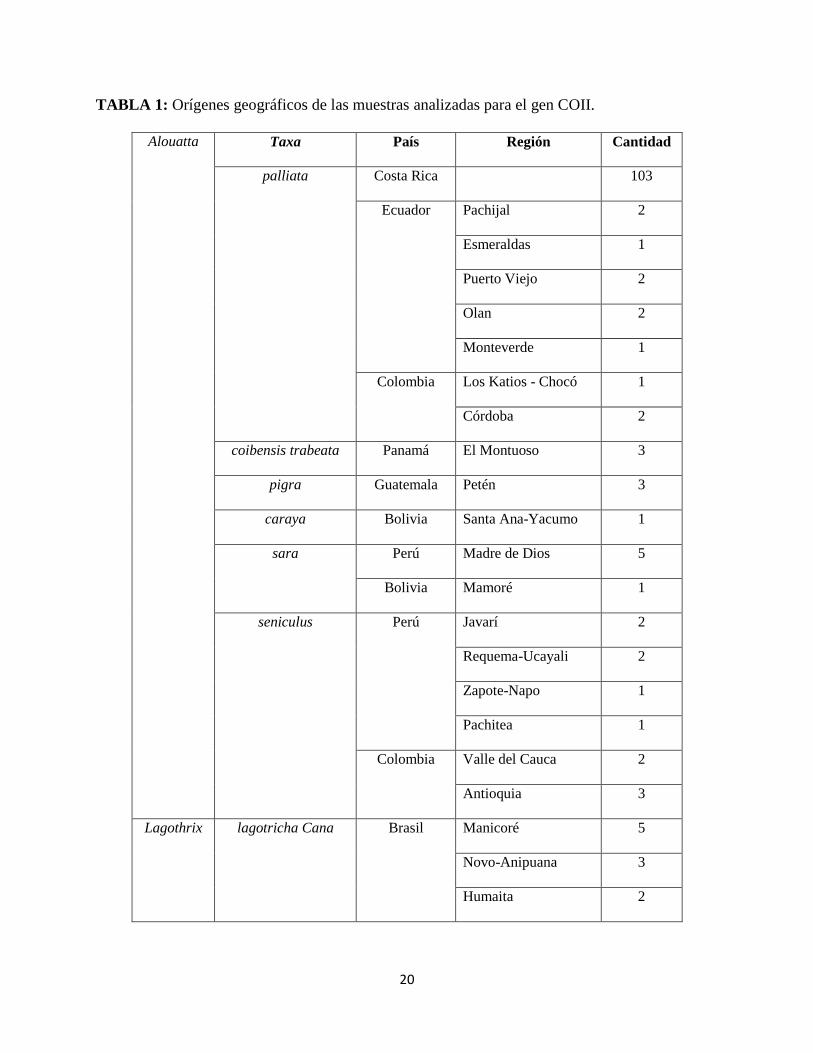
20
TABLA 1: Orígenes geográficos de las muestras analizadas para el gen COII.
Alouatta Taxa País Región Cantidad
palliata Costa Rica 103
Ecuador Pachijal 2
Esmeraldas 1
Puerto Viejo 2
Olan 2
Monteverde 1
Colombia Los Katios - Chocó 1
Córdoba 2
coibensis trabeata Panamá El Montuoso 3
pigra Guatemala Petén 3
caraya Bolivia Santa Ana-Yacumo 1
sara Perú Madre de Dios 5
Bolivia Mamoré 1
seniculus Perú Javarí 2
Requema-Ucayali 2
Zapote-Napo 1
Pachitea 1
Colombia Valle del Cauca 2
Antioquia 3
Lagothrix lagotricha Cana Brasil Manicoré 5
Novo-Anipuana 3
Humaita 2

21
Cabe resaltar que los especímenes utilizados en este estudio fueron cazados por comunidades
indígenas para su alimentación, eran mascotas en comunidades indígenas o de colonos o fueron
capturados en el medio natural para llevar a cabo estudios veterinarios paralelos (caso de las
muestras de Costa Rica)
El ADN fue extraído de piel, músculo, dientes, pelo con bulbo y mancha de sangre. El ADN de
músculo y piel fue obtenido con fenol – cloroformo (Sambrook et al., 1989), mientras que el de
sangre, dientes y pelo fue obtenido con resina Cheelex al 10% (Walsh et al., 1991).
Una vez que se obtuvo el ADN, se realizó PCR (Reacción en Cadena de la Polimerasa) para
amplificar la región del gen mtCOII. Para dicha amplificación se obtuvo un volumen final de
50µl, con 5µl de MgCl2, 6µl de Buffer 10X, 2µl de dNTP’s (4pmol de cada primer), 2µl de
ADN (50-100ng por µl), 2µl de cada uno de los cebadores H7766 (5’-
AACCATTTCATAACTTTGTCAA-3’) y L6955 (5’-CTCTTAATCTTTAACTTAAAAG-3’)
(Ashley & Vaughn 1995).
Para llevar a cabo la PCR se empleó un termociclador Bio-Rad y las condiciones fueron una
denaturación inicial a 95°C por 2min, y de 35 ciclos que constaron de tres pasos: una
denaturación a 95°C por 45s, un segundo paso de anillamiento a 50°C por 30s y un tercer paso
de extensión a 72°C por 35s. Se llevó a cabo una extensión final a 72°C por 5min. Para
determinar la obtención del amplificado esperado, se llevó a cabo una electroforesis en gel de
agarosa al 2% teñido con bromuro de etidio, visualizándose el posible amplificado en un
transiluminador de rayos UV. Posterior al chequeo, las muestras donde se evidenciaran las
bandas claras, fueron enviadas para secuenciación automática de la cadena “L” a MACROGEN
U.S.A.

22
6.2 Alineamiento de secuencias de ADN mitocondrial
Los alineamientos de las secuencias se realizaron manualmente y con la ayuda del programa
ALIGN (Fluxus Technology Ltd.)
.
6.3 Análisis filogenéticos
Los análisis filogenéticos fueron realizados por el programa Modeltest (Posada y Crandall ,
1998) y el programa Mega 06:05 (Tamura et al. , 2013), estos se aplicaron para determinar el
mejor modelo de mutación evolutiva para las secuencias analizadas. Mediante el programa
PAUP*4.0b8 (Swofford, 2002) y el programa MEGA 6.05, se construyeron árboles basados en
los métodos Neighbour-Joining y Maximum-Likelihood. En el árbol Neighbor -joining fue
utilizada la distancia genética 2P Kimura (Kimura 1980). En el árbol de máxima verosimilitud se
utilizó el modelo HKY + G (Hasegawa-Kishino-Yano, 1985) y una distribución discreta gamma
(+ G) con cinco tipos de categorías.El análisis Bayesiano se realizó utilizando el modelo HYK +
G con la tasa de distribución gamma, variando entre los sitios, porque se quería determinar el
mejor modelo utilizado por el programa FindModel. Los análisis Bayesianos se completaron con
el programa BEAST v. 1.8.1 (Drummond et al., 2012).
6.4 Análisis de diversidad, heterogeneidad y flujo génico
Se estimó la heterogeneidad genética y las estimaciones de flujos génico teóricas entre las
diversas especies de Alouatta con las que contó este trabajo, calculando el valor de los
estadísticos HST, KST*, KST, Z, Z* (Hudson et al 1992). Con la ayuda del software DNAsp

23
5.10.01 (Librado y Rozas et al 2010), se determinaron el número de sitios polimórficos (S), el
número de mutaciones (Ἠ) y el número de haplotipos (h) de las secuencias. También se calculó
la diversidad Haplotípica (Hd), la diversidad nucleotídica (π), el número promedio de diferencias
nucleotídicas (k), y el coeficiente de coancestralidad (θ) por secuencia.
6.5 Detección de cambios demográficos
Para determinar los posibles cambios que han tenido las diferentes especies de Alouatta
analizadas se realizaron los siguientes procedimientos:
1. La distribución mismatch (diferencias en los pares de secuencias),se obtuvo siguiendo el
método de Rogers y Harpending (1992) y Rogers et al., (1996).
2. Tests Fu and Li D* and F* (Fu and Li, 1993), el estadístico Fu FS (Fu, 1997), el test D Tajima
(Tajima, 1989) y el estadístico R2 de Ramos-Onsins and Rozas (2002) para determinar posibles
cambios en los tamaños poblacionales de las especies de Alouatta analizadas (Simonsen etal.,
1995; Ramos-Onsins and Rozas, 2002).

24
7. RESULTADOS
7.1 FILOGENIA
Tanto en el árbol de Kimura-2-parameter (Figura 6)y el de máxima verosimilitud (Figura 7),
muestran explicitamente dos grupos de monos aulladoressoportados por diferentes valores de
boostrap para cada árbol. El primero (Figura 6) consta de los aulladores (A. caraya, A. seniculus
y A. sara), teniendo orígenes geográficos provenientes de Colombia, Perú y Bolivia. El segundo
consta de los aulladores (A. pigra y A. palliata), teniendo orígenes geográficos provenientes de
Costa rica, Colombia, Guatemala, Ecuador y Panamá. En el clado uno, las especies son
totalmente diferenciables entre sí, ya que los valores de bootstrap son (99, 99 y 87) para cada una
respectivamente. En cuanto al clado dos, las especies tienen valores de bootstrap de (99 y 99).
Esto nos demuestra que molecularmente todas se pueden distinguir. Se corrobora que las cuatro
muestras obtenidas de Perú (Cuenca del rio Madre de Dios) pertenecen a de A. sara. Las seis
muestras restantes provenientes de Perú (Rio Javarí, Rio Napo, Rio Ucayali y Rio Pachitea), se
encuentran estrechamente relacionadas con los diferentes A. seniculus de Colombia.
En cuanto a los A. palliata que se encuentran en el clado dos, no se distinguen subespecies están,
ya que los individuos se entremezclan unos con otros, no habiendo un punto claro de
diferenciación.
El árbol de máxima verosimilitud, cambia solamente los valores de bootstrap. Clado uno (99 y
97) y clado dos (99 y 97) respectivamente, ya que tiene el mismo comportamiento con las
diferentes especies de aulladores dentro de él.

25
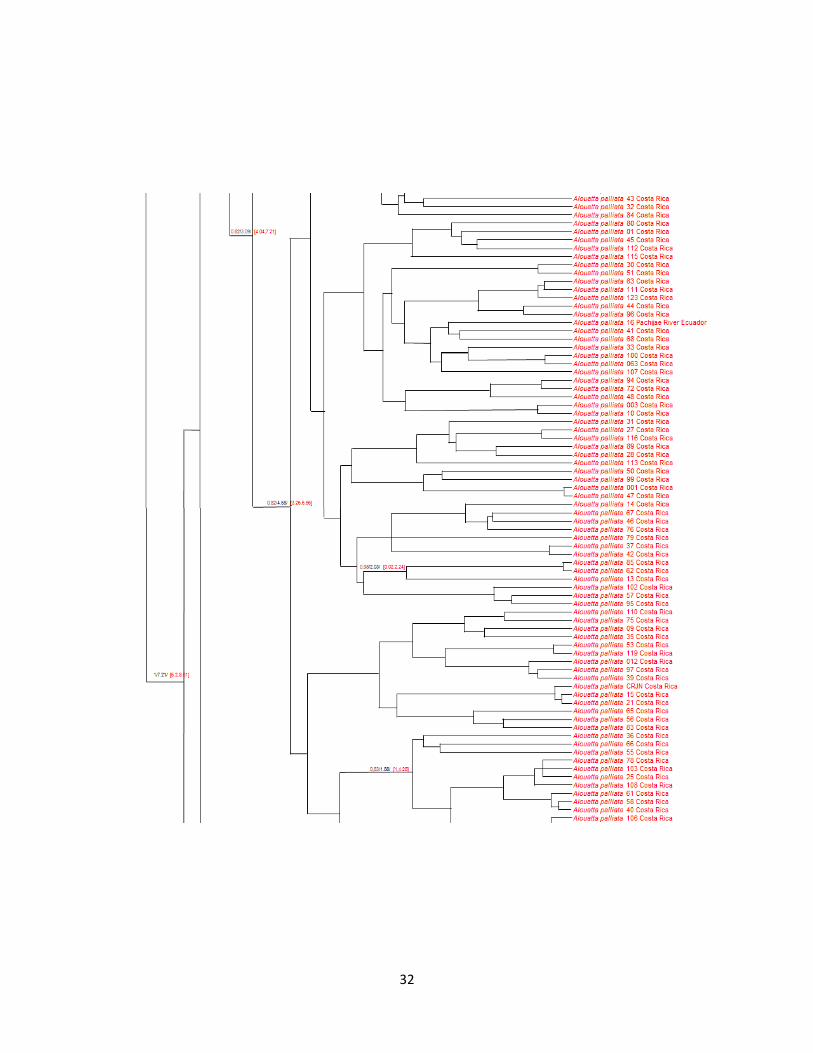
En cuanto al árbol Bayesiano (Figura 8), se evidencia que el género Alouatta se dividió con
respecto a los demás Atélidos hace aproximadamente 15.93 MA(95 % HPD: 14.93-16.85 MA; P
= 1). Específicamente, la división de sus antepasados Lagothrix y Ateles ocurrió alrededor de
11.18 MA (95 % HPD: 10.3-12.23 MA; P = 1). La diversificación dentro del género Alouatta
comenzó sobre los 7.21 MA (95 % HPD: 6.20-8.01 MA; P = 1), siendo A. seniculus el primero
en divergir. Dentro de A. seniculus, la diversificación mitocondrial comenzó alrededor de 3,94
MA (95 % HPD: MA 1,93-5,86; p = 1). La siguiente especie en divergir es A. sara hace
aproximadamente 6.61 MA (P = 0.28). La diversificación mitocondrial de A. sara comenzó
alrededor de 2.27 MA (95 % HPD: 1.2-4.72 MA; P = 1). La división entre las especies de
monos aulladores A. pigra y A. palliata se dio alrededor de 6.06 MA (P = 0.47), dentro de A.
pigra se dio hace aproximadamente 2.12 MA (95 % HPD: 1.7-3.89 MA; P = 1) y dentro de A.
palliata hace aproximadamente 4.58 MA (95 % HPD: 4.04-7.21 MA; P = 0.82).
Es importante resaltar en este caso (Figura 8), la especie A. caraya, esta más relacionada con la
especie A. pigra, aspecto que difiere notablemente de los árboles anteriormente descritos (Figura
6 y 7).

26
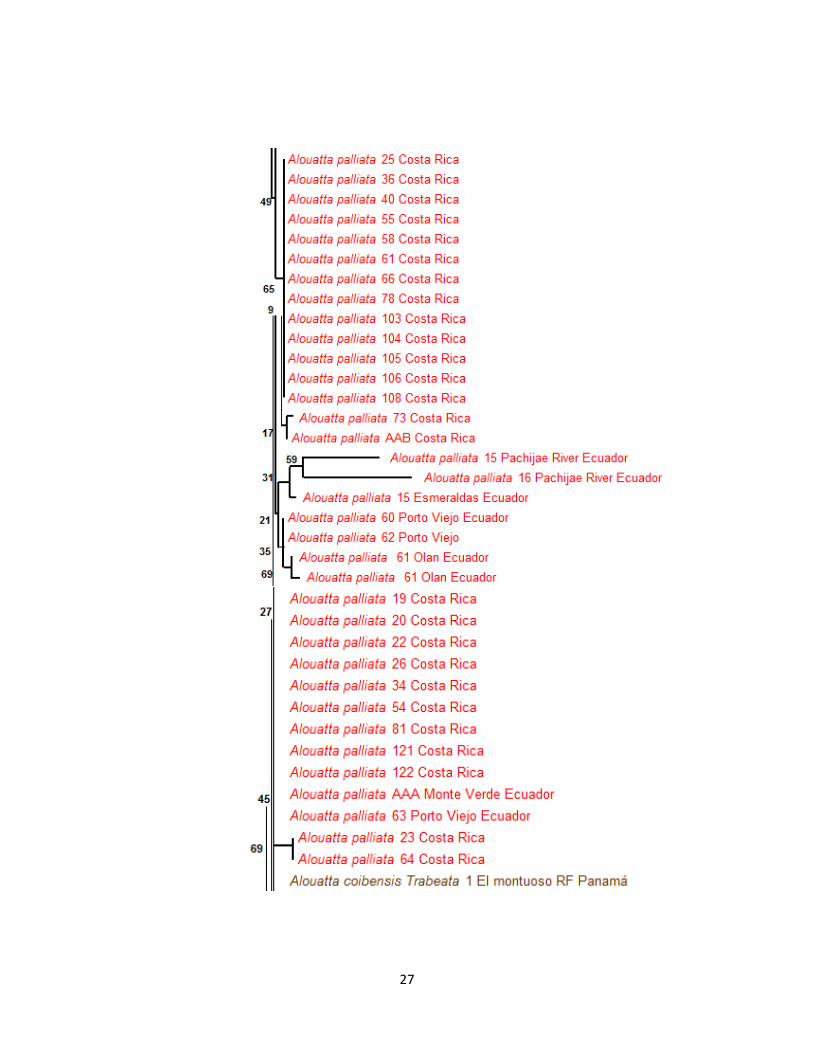
27
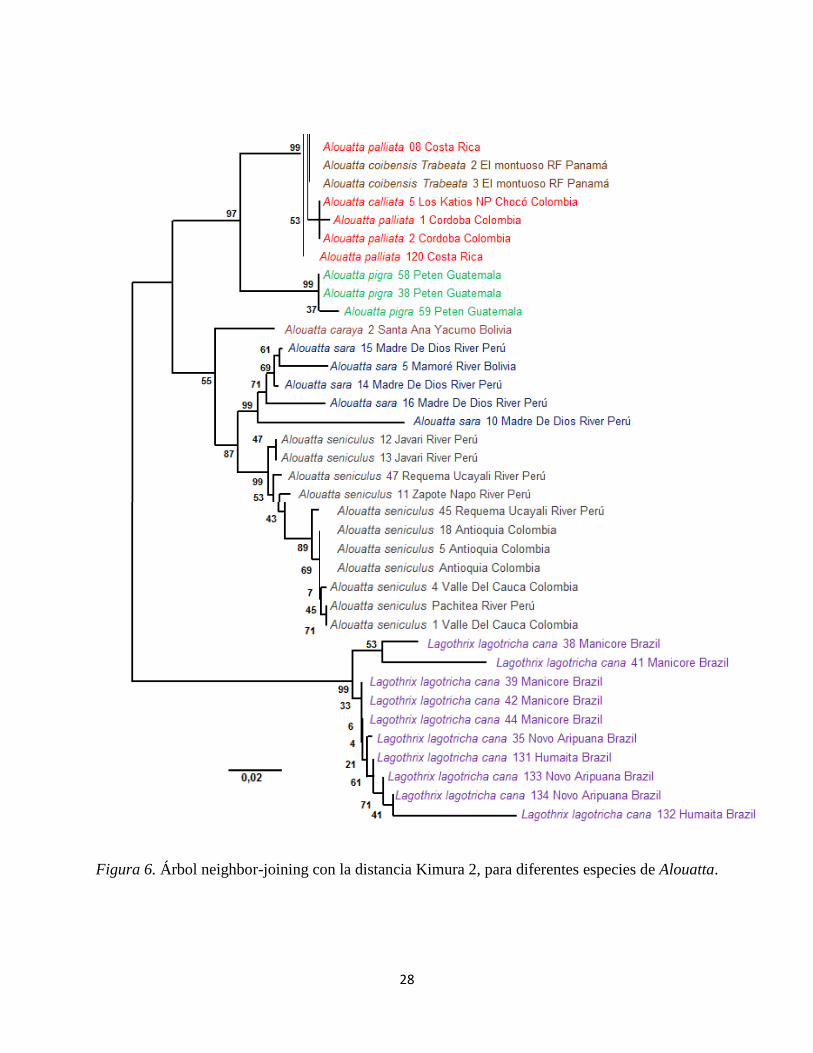
28
Figura 6. Árbol neighbor-joining con la distancia Kimura 2, para diferentes especies de Alouatta.

29
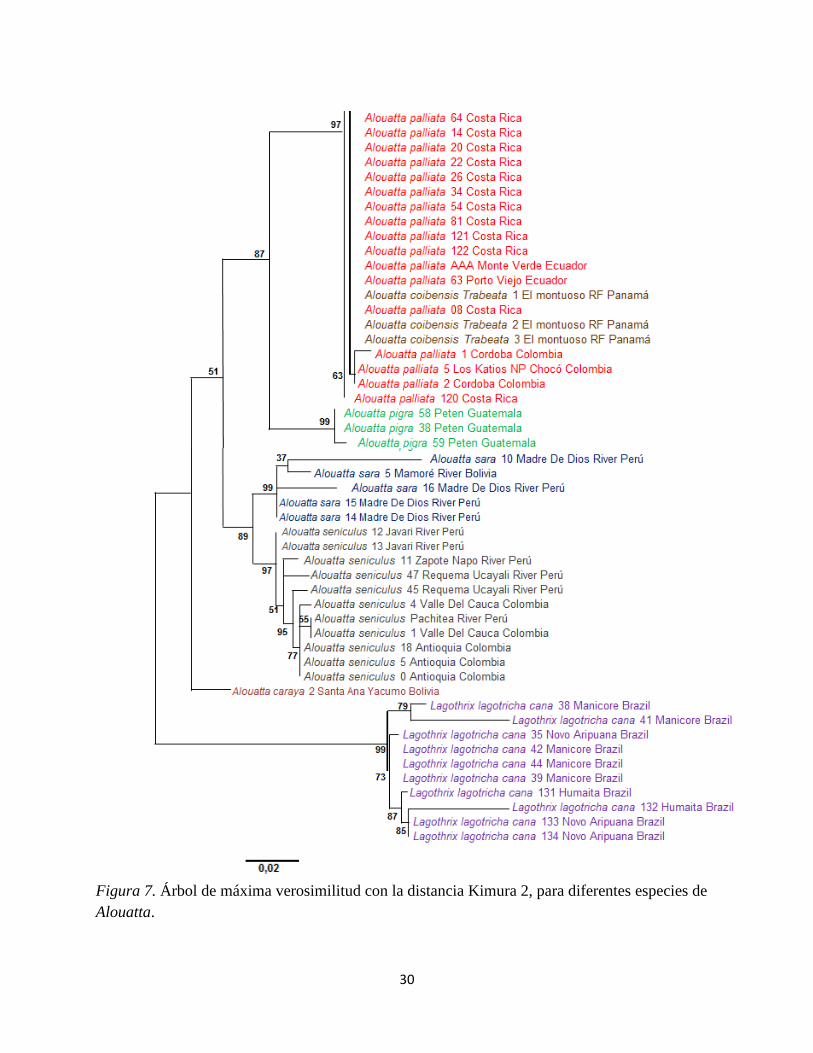
30
Figura 7. Árbol de máxima verosimilitud con la distancia Kimura 2, para diferentes especies de
Alouatta.
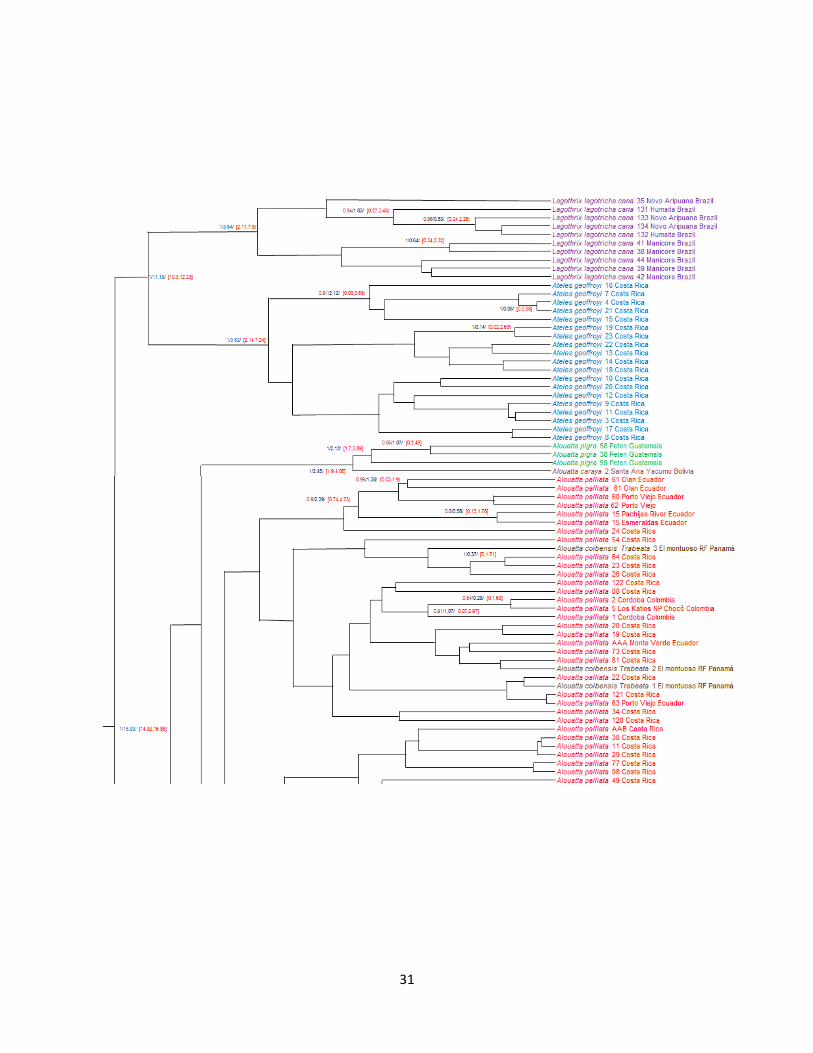
31
32

33
Figura 8. Árbol Bayesiano para diferentes especies de Alouatta.
Las distancias genéticas Kimura 2P (Tabla 2 ) mostraron valores que van desde alrededor de 6 %
a 9 % entre los diferentes tipos de especies deAlouatta estudiadas. Estos son los valores típicos
de especies diferentes (Ascunce et al. , 2003).
Tabla 2. Kimura (1980) 2P distancias genéticas entre diferentes especies de Alouatta. Abajo, los
valores de las distancias genéticas en porcentajes (%); arriba, errores estándar en porcentajes
(%). 1= A. palliata; 2 = A. pigra; 3 = A. seniculus; 4 = A. sara; 5 = A. caraya; 6 = Lagothrix
lagotricha cana
Especies de
Alouatta
1 2 3 4 5 6
1 - 0.8 1.1 1.0 1.2 1.7
2 5.3 - 1.3 1.0 1.2 1.9
3 8.4 8.0 - 0.8 0.9 1.6
4 9.3 8.4 5.5 - 1.7 1.6
5 8.0 7.2 5.5 7.6 - 1.5
6 15.1 16.0 14.3 16.4 13.6 -

34
7.2 DIVERSIDAD GENÉTICA, HETEROGENEIDAD GENÉTICA Y FLUJO GÉNICO
La heterogeneidad genética entre las muestras de especies de monos aulladores analizadas fue
significativamente alta (Tabla 3). Del mismo modo, las estimaciones del flujo génico son muy
bajas (Nm = 0.09-0.13), por tal razón se avala la existencia de que éstas son especies diferentes.
Tabla 3. Etadisticos de heterogeneidad y flujo génico (Nm) para diferentes especies de Alouata.
Probabilidad significativa (P < 0.05).
Estimados de diferenciación
genética P Flujo génico
= 411.000 df = 105 0.0000*
HST = 0.0825 0.0000* γST = 0.7895 Nm = 0.13
KST = 0.7758 0.0000* NST = 0.8429 Nm = 0.09
KST* = 0.4524 0.0000* FST = 0.8387 Nm = 0.10
ZS = 3,674.6439 0.0000*
ZS* = 8.0124 0.0000*
Snn = 1.0000 0.0000*
Los valores más altos de heterogeneidad genética entre especies fueron para A. seniculus-A.
palliata (FST = 0.938) entre A. seniculus-A. pigra (FST = 0.936) (Tabla 4). Entre A. seniculus-A.
sara, el valor también es significativo (FST = 0.641), lo que indica que son dos especies
diferentes, como se comprueba en los diferentes árboles, aun cuando fenotípicamente son
idénticas (Figura 6 y 7).
35
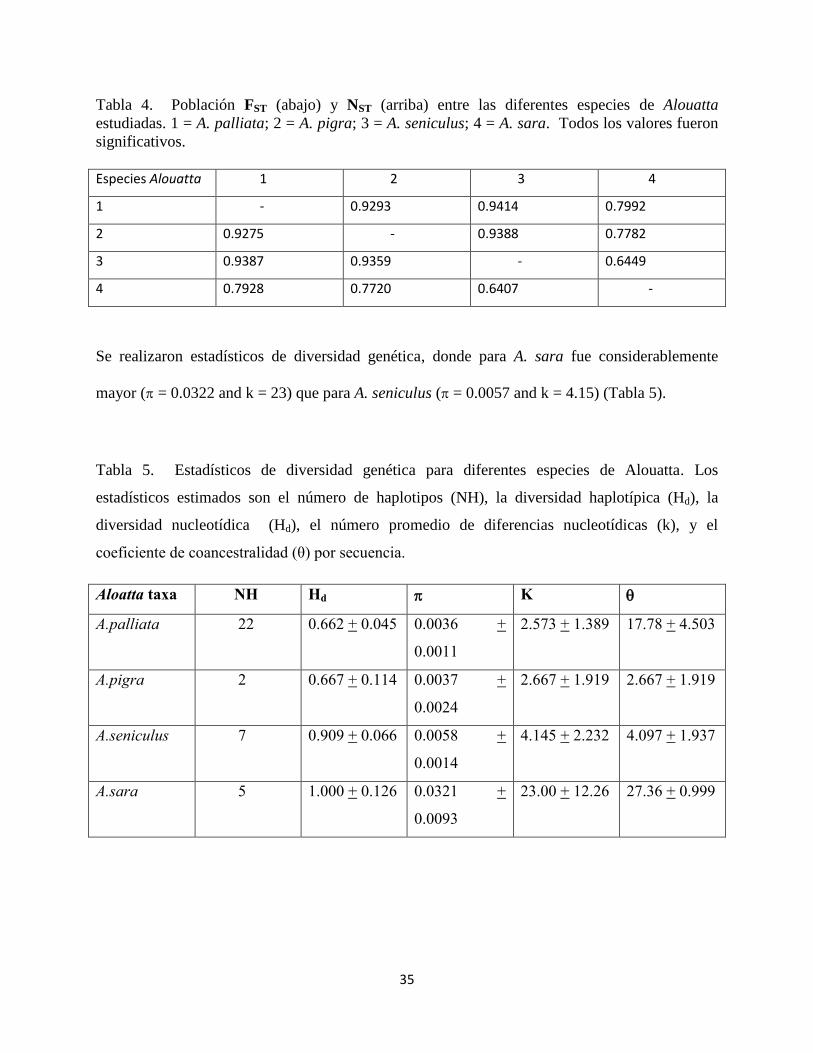
Tabla 4. Población FST (abajo) y NST (arriba) entre las diferentes especies de Alouatta
estudiadas. 1 = A. palliata; 2 = A. pigra; 3 = A. seniculus; 4 = A. sara. Todos los valores fueron
significativos.
Especies Alouatta 1 2 3 4
1 - 0.9293 0.9414 0.7992
2 0.9275 - 0.9388 0.7782
3 0.9387 0.9359 - 0.6449
4 0.7928 0.7720 0.6407 -
Se realizaron estadísticos de diversidad genética, donde para A. sara fue considerablemente
mayor ( = 0.0322 and k = 23) que para A. seniculus ( = 0.0057 and k = 4.15) (Tabla 5).
Tabla 5. Estadísticos de diversidad genética para diferentes especies de Alouatta. Los
estadísticos estimados son el número de haplotipos (NH), la diversidad haplotípica (Hd), la
diversidad nucleotídica (Hd), el número promedio de diferencias nucleotídicas (k), y el
coeficiente de coancestralidad (θ) por secuencia.
Aloatta taxa NH Hd K
A.palliata 22 0.662 + 0.045 0.0036 +
0.0011
2.573 + 1.389 17.78 + 4.503
A.pigra 2 0.667 + 0.114 0.0037 +
0.0024
2.667 + 1.919 2.667 + 1.919
A.seniculus 7 0.909 + 0.066 0.0058 +
0.0014
4.145 + 2.232 4.097 + 1.937
A.sara 5 1.000 + 0.126 0.0321 +
0.0093
23.00 + 12.26 27.36 + 0.999

36
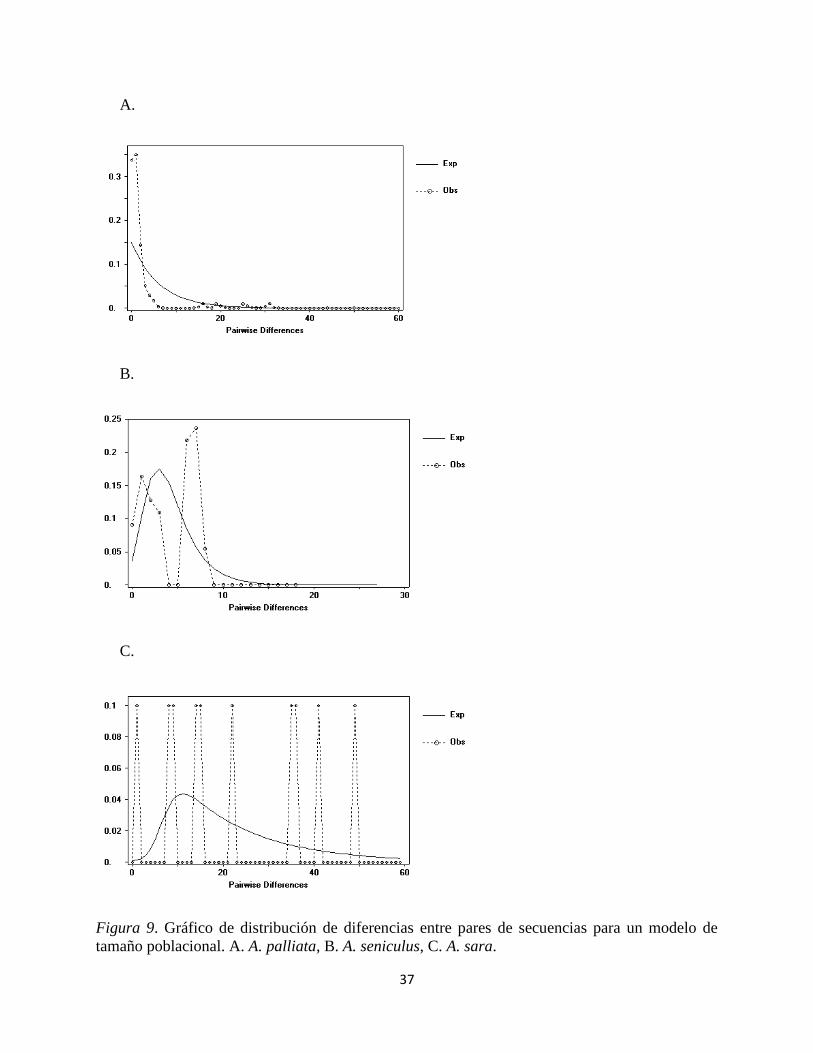
Se analizaron los posibles cambios demográficos históricos de A. seniculus, A. sara y A. palliata
(Tabla 6 y figura 9). Para A. seniculus ningún estadístico mostró claramente que hay un cambio
demográfico en la historia evolutiva de esta especie ya que ningún resultado es significativo.
Para A. sara tres de las pruebas empleadas mostraron evidencia significativa de expansión de la
población (prueba de Tajima D, Fu y Li D y Fu y Li * F *).En el caso de A. palliata todos los test
dieron significativos, por tal razón es una especie que muestra claros síntomas de expansión
poblacional.
Tabla 6. Estadísticos demográficos aplicados a diferentes especies de Alouatta (A. palliata, A.
seniculus, A. sara). * P < 0.05; ** P < 0.01, expansiones poblacionales significativas.
Tajima
D
Fu & Li
D*
Fu & Li
F* Fu’s Fs
raggedness
rg R2
A. palliata
P[D < -
2.789] =
0.001**
P[D* < -
8.811] =
0.001**
P[F* < -
7.438] =
0.001**
P[Fs < -
8.928 ] =
0.004**
P[rg <
0.0527] =
0.296
P[R2 <
0.038] =
0.039*
A. seniculus
P[D <
0.051] =
0.553
P[D* < -
0.071] =
0.426
P[F* < -
0.122] =
0.444
P[Fs < -
0.815] =
0.327
P[rg <
0.103] =
0.460
P[R2 <
0.146] =
0.332
A. sara
P[D < -
1.205] =
0.026*
P[D* < -
1.432] =
0.038*
P[F* < -
1.306] =
0.036*
P[Fs <
0.683] =
0.386
P[rg <
0.140] =
0.229
P[R2 <
0.218] =
0.521
37
A.
B.
C.
Figura 9. Gráfico de distribución de diferencias entre pares de secuencias para un modelo de
tamaño poblacional. A. A. palliata, B. A. seniculus, C. A. sara.

38
Para el conjunto de las 148 muestras, se encontraron 44 haplotipos. La red de haplotipos permitió
separar a las especies, como lo hicieron los árboles filogenéticos (Figura 10)
Figura 10. Red de haplotipos para diferentes especies de Alouatta. Rosa = A. Caraya, azul = A.
seniculus, verde = A. sara, vinotinto = A. palliata, amarillo = A. pigra, negro = Lagothrix.
8. DISCUSIÓN Y CONCLUSIONES
8.1 FILOGENIA
Los árboles filogenéticos resolvieron de manera clara las relaciones entre las especies de
Alouatta con las cuales se trabajaron. Se señalan algunos detalles:

39
En dos de los tres árboles realizados con las distintas especies de Alouatta (Figura 6 y 7) y en la
red de haplotipos (figura 10), la especie A. caraya, tiene un comportamiento definido ya que es
la más basal y diverge del resto de especies con las cuales se trabajaron. Menos lógico es el
resultado obtenido con el árbol Bayesiano, ya que A. caraya queda mezclado entre los A. palliata
y los A. pigra (figura 8), por tal razón se deben realizar estudios más exhaustivos con A. caraya
para determinar qué relaciones presenta con las otras especies de aulladores.
Este estudio nos demuestra que la heterogeneidad genética entre las distintas especies de
Alouatta es significativa. Es claro que las especies A. sara y A. seniculus están estrechamente
relacionadas, ya que desde el punto de vista morfológico son idénticas, pero son totalmente
discernibles entre sí molecularmente. Esto coincide con Cortés- Ortiz et al., (2003), quienes
usando diferentes secuencias de otros genes mitocondriales también diferenciaron esas dos
especies de aulladores. Los genes mitocondriales han resultado ser más efectivos y útiles que los
genes nucleares para poder discriminar las diferencias genéticas entre las distintas especies del
género Alouatta. Un ejemplo de trabajo con genes nucleares lo hicieron en el estudio de Cortés-
Ortiz et al., (2003) tratando de resolver las relaciones filogenéticas entre las diferentes especies
Alouatta. Sin embargo, los genes nucleares no recuperaron adecuadamente las relaciones
filogenéticas entre especies de aulladores, mientras que los genes mitocondriales sí lo hicieron.
Estas fuertes diferencias entre taxones de Alouatta se hacen evidentes mediante estudios
cromosómicos realizados por Minezawa et al., (1985) donde se muestra 2N = 50 para hembras
(con X1X1X2X2) y 2N = 49 para machos (X1X2Y), con un rango de 2-6 microcromosomas
(seis microcromosomas fueron más frecuentes) para A. sara. Luego, Stanyon et al., (1995)
determina 2N = 50 tanto para machos como para hembras (con X1X1X2X2 y X1X2Y1Y2) y 28

40
cromosomas acrocéntricos. Notoriamente, A. sara es cromosómicamente diferenciable de A.
seniculus en Colombia (2N = 44 También con X1X1X2X2 y X1X2Y1Y2, 26 cromosomas
acrocéntricos, 3-4 microcromosomas; Torres y Leibocini 2001). Por lo tanto el marcador
mitocondrial COII (al igual que los cariotipos) puede perfectamente discriminar ejemplares de A.
sara y A. seniculus que son morfológicamente casi idénticos.
Las subespecies A. palliata ssp. palliata y A. palliata ssp. aequatorialis no se diferencian
molecularmente ya que los individuos de ambas subespecies morfológicas quedan
entremezclados independientemente del lugar de su procedencia geográfica (Costa Rica,
Colombia y Ecuador) (figura 6, 7 y 8). Igualmente, las muestras trabajadas de Panamá (A.
trabeata coibensis) tampoco se diferencian molecularmente de A. palliata, ya que en ninguno de
los árboles forma un grupo o clado aislado (figura 6, 7 y 8). Probablemente, los rasgos
morfológicos utilizados para diferenciar esta supuesta especie (dermatoglifos; Frohelich et al.,
1991) y algunas mínimas diferencias en características del pelaje no son buenos caracteres
filogenéticos para establecer relaciones entre esas poblaciones de aulladores en Panamá. Similar
resultado al nuestro fue encontrado por Cortés-Ortiz et al., (2003) con diferentes genes.
8.2 DIVERSIDAD GENÉTICA, HETEROGENEIDAD GENÉTICA Y CAMBIOS
DEMOGRÁFICOS
La diversidad genética encontrada en A. palliata y A. pigra es claramente inferior a la encontrada
en las dos especies sudamericanas (A. seniculus y A. sara). Esto concuerda con la hipótesis que
las formas sudamericanas son más ancestrales y que las dos especies mesoamericanas son formas

41
derivadas más recientes. La heterogeneidad genética al interior de A. palliata es muy pequeña, lo
cual concuerda con los árboles filogenéticos en no detectar supuestas subespecies al interior de la
misma. Los estadísticos demográficos aplicados (Tajima D, Fu & Li D*, Fu & Li F*, Fu’s Fs y
R2) fueron muy importantes para comprobar la expansión poblacional de A. palliata, mientras
que en A. sara y A. pigra (por el tamaño muestral) no se tiene claro si han habido cambios
demográficos o no (Tabla 6 y figura 10). Es de suma importancia tener un número significativo
de muestras de cada especie ya que si hay pocas, esto hace que no se detecten fácilmente los
tipos de expansiones que han tenido a lo largo del tiempo. Lo que resulta evidente es que en A.
palliata (donde el tamaño muestral es elevado) se detecta una muy fuerte evidencia de expansión
poblacional reciente que, a su vez, concuerda con el hecho que es una especie más reciente que
las sudamericanas y con fuerte homogeneidad molecular a su interior.
8.3 TIEMPOS DE DIVERGENCIA
El tiempo de diversificación del genero Alouatta en este trabajo fue de 7.21 millones de años
(MA), esto coincide aproximadamente con Cortés-Ortiz et al., (2003), ya que ellos estimaron un
tiempo de 6.8 MA. Esta información encaja con la formación del norte de los Andes (Lundberg
et al., 1998) y la formación de los ríos amazónicos principales según la hipótesis paleogeográfica
(Hoorn et al., 1995; 2010). La estimación temporal de la diversificación de A. palliata y A. pigra,
con respecto a las formas sudamericanas, fue estimada en unos 3.12 MA, mientras que Cortés-
Ortiz et al., (2003) la estimaron en unos 3 MA, siendo esta similitud bastante importante para la
veracidad de los resultados de este trabajo. Esa datación coincide con la finalización del Istmo
de Panamá (Estrada and Coates-Estrada, 1984); esto incluye y aumenta la importancia de este

42
Istmo en la especiación de los monos centroamericanos. Por tal razón, la evolución molecular de
los monos aulladores suramericanos precede a la evolución molecular de los monos
centroamericanos. Igualmente, la red de haplotipos mostró que A. palliata y A. pigra pueden
proceder de dos migraciones desde Sudamérica diferentes, fenómeno que no es observable en los
árboles de naturaleza jerárquica. Esto es debido a que los árboles jerárquicos únicamente recogen
eventos cladogenéticos, mientras que las redes de haplotipos permiten simultáneamente la
coexistencia de haplotipos ancestrales y descendientes y, por lo tanto, de fenómenos de
evolución reticular u anagénesis (Posada & Crandall, 2001).
En conclusión:
1. Las especies mostraron niveles altos de diversidad genética y bajos niveles de flujo
génico como un todo, lo cual es evidencia de que se analizaron especies bien
diferenciadas. Sin embargo, las dos especies mesoamericanas mostraron niveles de
diversidad genética claramente inferiores a las especies sudamericanas, por lo que
parecen las primeras haber descendido de ancestros de las segundas.
2. El gene mitocondrial COII claramente pudo diferenciar dos especies sinmórficas (A.
seniculus y A. sara) al igual que lo hacen los cariotipos, pero no la morfología. Por
primera vez, hemos encontrado evidencia molecular que los aulladores rojos del sur de la
Amazonia peruana representan al taxón A. sara, con lo cual el mismo no sería endémico
exclusivamente de Bolivia.
3. Las diferencias genéticas entre individuos y poblaciones de A. palliata en Suramérica
(Colombia y Ecuador) y Centro América (Costa Rica) son muy pequeñas lo cual aboga
por la inexistencia de subespecies con claro sentido biológico. Por lo tanto únicamente se

43
reconocería a A. palliata sin subespecies. Igualmente, A. coibensis trabetta no pareciera
ser una entidad real al nivel específico a partir de los resultados moleculares presentados.
Sin embargo, estudios con otros marcadores moleculares, cariotípicos o de aislamiento
reproductivo serían necesarios para determinar la realidad de esa pretendida especie.
4. Claramente, A. palliata muestra haber tenido una clara expansión poblacional con todos
los análisis llevados a cabo. No queda tan claro cuál ha sido el comportamiento
demográfico de los otros taxones analizados debido a que sus tamaños muestrales fueron
menos representativos.
5. Estimamos que el proceso de divergencia al interior del género Alouatta comenzó hace
unos 7 MA, es decir durante la última fase del Mioceno. Sin embargo, la aparición de los
linajes mitocondriales típicos de las dos especies centro-americanas, A. palliata y A.
pigra, se dio en torno a 3.1 MA, lo cual coincide con el cierre definitivo de Istmo de
Panamá durante el Plioceno. Divisiones de linajes haplotípicos al interior de esas dos
especies se dieron básicamente durante el Pleistoceno.
9. REFERENCIA BIBLIOGRAFICA
Abascal, F. 2007. Alineamiento de secuencias. Motivos, perfiles y dominios. Madrid – España.
Centro Nacional de Biotecnología. 71 diapositivas.
Ascunce, M.S., Hasson, E., Mudry, M.D. (2003). COII: a useful tool for inferring phylogenetic
relationships among New World monkeys (Primates, Platyrrhini). Zool. Scripta 32: 397-406.

44
Ashley, M.V. & T.A. Vaughn. 1995. Owl monkeys (Aotus) are highly divergent in mitocondrial
cytochrome c oxidase (COII) sequences. Int. J. Primatol. 5: 793-807.
Cortés-Ortiz, L., Bermingham, E., Rico, C., Rodriguez-Luna, E., Sampaio, I., Ruiz-García, M.
(2003). Molecular systematics and biogeography of the Neotropical monkey genus Alouatta.
Mol. Phylogenet. Evol. 26:64–81.
Cheney, D. L. & Wrangham, R.W. 1987. Predation. En Primates Societies, Smuts, B. B.
;Cheney, D.L.; Seyfarth, R. M.; Wrangham, R. W. y Struhsaker, T. T. (Eds.) Primatesocieties.
The University of Chicago Press. 227-239 pp.
Chivers, D. J. & Hladik, L. M. 1980. Morphology of the gastrointestinal tract in primates:
comparations whith mammals in relation to diet. Journal of Morphology. 166: 337 – 386.
Crockett, C.M., 1998. Conservation biology of the genus Alouatta. Int.J. Primatol. 19, 549–578.
Crockett, C. M. 1984. Family feuds. Natural History 93(8):54-63.
Crockett, C. & Eisenberg, J. 1987. Howlers: variations in group size and demography. En
Primate societies. Chicago IL, USA: University of Chicago Press.
Defler, T. R. 2003. Primates de Colombia. Conservación Internacional. Bogotá DC., Colombia.
543p.

45
Defler, T. R. 2004. Historia natural de los primates colombianos. Bogotá DC., Colombia. 308p.
Drummond, A.J., Suchard, M.A., Xie, D., Rambaut, A. (2012). Bayesian phylogenetics with
BEAUti and the BEAST 1.7. Mol. Biol. Evol. 29: 1969–1973.
Emmons, L. H. and F. Feer. 1997. Neotropical rainforest mammals: A field guide. Second
edition. University of Chicago Press, Chicago, USA.
Estrada, A.; Juan-Solano, S.; Ortiz-Martinez, T. & Coates-Estrada, R. 1999. Feeding and general
activity patterns of a howler monkey (Alouatta palliata) troop living in a forest fragment at Los
Tuxtlas, Mexico. American Journal of Primatology 48(3):167-183.
Estrada, A., Coates-Estrada, R. (1984). Some observations on the present distribution and
conservation of Alouatta and Ateles in southern Mexico. Amer. J. Primatol. 7: 133-137.
Fu, Y-X. (1997). Statistical tests of neutrality against population growth, hitchhiking and
background selection. Genetics 147: 915-925.
Fu, Y., Li, W. (1993). Statistical Tests of Neutrality of Mutations. Genetics 133: 693-709.
Frohelich, J.W., Frohelich, P.H. (1987). The status of Panama’s endemic howling monkeys.
Primate Conservation 8: 58–62.

46
Gregorin, R. (2006). Taxonomia e variação geográfica das espécies do gênero Alouatta Lacépède
(Primates, Atelidae) no Brasil. Revista Brasileira de Zoologia 23: 64-144.
Groves, C.P., 2001. Primate Taxonomy. Smithsonian Institution Press, Washington, DC.
Hasegawa, M., Kishino, H., Yano, T. (1985). Dating of human-ape splitting by a molecular
clock of mitochondrial DNA. Journal of Molecular Evolution 22: 160-174.
Hershkovitz, P. (1972). The recent mammals of the Neotropical region: a zoogeographical and
ecological review. In A. Keast, F. Erk, and B. Glass (Eds.). Evolution, mammals, and southern
continents (pp. 311-431). Albany, New York: State University of New York Press.
Hill, W.C.O., 1962. Primates Comparative Anatomy and Taxonomy. V. Cebidae. Part. B.
Edinburgh University Press, Edinburgh.
Hoorn, C., Wesselingh, F.P., Hovikoski, J., Guerrero, J., 2010b. The development of the
Amazonian mega-wetland (Miocene; Brazil, Colombia, Peru, Bolivia). In: Hoorn, C.,
Wesseling, F.P. (Eds.), Amazonia, Landscape and Species Evolution. Wiley-Blackwell,
Oxford, pp. 123–142.
Hoorn, C., 1994b. Fluvial palaeoenvironments in the intracratonic Amazonas Basin (Early
Miocene–early Middle Miocene, Colombia). Palaeogeogr. Palaeoclimatol. Palaeoecol.
109, 1–54.

47
Hudson, R. 2000. A new statistic for detecting genetic differentiation. Genetics 155: 2011- 2014.
Nei, M. and Chesser, R. K. 1983. Estimation of fixation indices and gene diversities. Ann. Hum.
Genetics 47: 253-259
Hudson, R.R., Boss, D.D., Kaplan, N.L. (1992). A statistical test for detecting population
subdivision. Molecular Biology and Evolution9: 138-151.
Hudson, R., Slatkin, M., Maddison, W. P. 1992. Estimation of levels of gene flow from DNA
sequence data. Genetics 132: 583- 589.
Kimura, M. (1980). A simple method for estimating evolutionary rates of base substitutions
through comparative studies of nucleotide sequences. J. Mol. Evol. 16: 111-120.
Librado, P., Rozas, J. (2009). DnaSP v5: A software for comprehensive analysis of DNA
polymorphism data. Bioinformatics25: 1451-1452 | doi: 10.1093/bioinformatics/btp187.
Lundberg, J.G., Marshall, L., Guerrero, J., Horton, B., Malabarba, C., Wesselingh, F. (1998).
The stage for neotropical fish diversification: history of a tropical South American river. In:
Phylogeny and classification of neotropical fishes. Malabarba, L.R., Reis, R., Vari, R., Lucena,
Z., Lucena, C.A., (Eds). Porto Alegre, Brazil: Pontificia Universidade Católica Do Rio Grande
Do Sul. Pp. 13–48.

48
Lynch, M. and Crease T. 1990. The analysis of population survey data on DNA sequence
variation. Mol Biol. Evol. 7: 377-394
Minezawa, M., Harada, M., Jordan, O.C., Borda, C.J.V. (1985). Cytogenetics of Bolivian
endemic red howler monkeys (Alouattaseniculus sara): accessory chromosomes and Y-autosome
translocation related numerical variations. Kyoto Univ. Overs. Res. Rep. N. W. Monkeys 5: 7–16.
Nei, M. 1987. Molecular Evolutionary Genetics. Columbia University Press. Ney York.USA
Neves, A., and Rylands, A. 1991. Diet of a group of howling monkeys, Alouatta seniculus, in an
isolated forest patch in central Amazonia. A Primatol. Brasil 3: 263-274.
Posada D, Crandall KA. (1998). MODELTEST: testing the model of DNA substitution.
Bioinformatics 14: 817-818.
Posada, D., Crandall, K.A. (2001). Intraspecific gene genealogies: trees grafting into networks.
Trends Ecol. Evol. 16:37–45.
Ramos-Onsins, S.E., Rozas, J. (2002). Statistical Properties of New Neutrality Tests Against
Population Growth. Mol. Biol. Evol. 19: 2092–2100.
Raymond, M.R., Rousset, F. 1995. Genepop version 3.2. Population genetics software for exact
test and ecumenicism. Journal of heredity 86: 248-249

49
Rodríguez-Luna, E.; García-Orduña, F. & Canales- Espinosa, D. 1993. Translocación del mono
aullador Alouatta palliata: una alternativa conservacionista. En Estrada, A.;
Rodríguez- Luna, E; Cortés-Ortís L., Martinez-Contreras, J. (Eds.) Estudios primatológicos en
México. Vol. II. Biblioteca de la Universidad Veracruzana de Xalapa, Veracruz, México. 113-
148 pp.
Rodriguez-Luna, E. 2000. An assessment of thediversity of new world primates. Neotropical
Primates 8(2):61-93.
Rogers, A.R., Harpending, H.C. (1992). Population growth makes waves in the distribution of
pairwise genetic differences. Mol. Biol. Evol. 9: 552-569.
Rogers, A.R., Fraley, A.E., Bamshad, M.J., Watkins, W.S., Jorde, L.B. (1996). Mitochondrial
mismatch analysis is insensitive to the mutational process. Mol. Biol. Evol. 13: 895-902.
Ruiz-García, M, Leguizamón, M, Vásquez, C, Rodríguez, K, Castillo, M. 2010. Métodos
genéticos para la reintroducción de monos de los géneros Saguinus, Aotus y Cebus (Primates:
Cebidae) decomisados en Bogotá, Colombia. 1052-1053.
Rylands, A.B., Schneider, H., Langguth, A., Mittermeier, R.A., Groves, C.P., Rodríguez-Luna,
E. (2000). An assessment of the diversity of New World primates. Neotrop. Prim. 8: 61–93.

50
Rylands, A. B., Mittermeier, R. A. (2009). The diversity of the New World primates
(Platyrrhini): An annotated taxonomy. In South American Primates. Comparative perspectives
in the study of behavior, ecology and conservation. Garber, P. A., Estrada, A., Bicca-Marques,
J.C., Heymann, E. W., Strier, K. B. (Eds.). Springer Science+Business Media, New York, USA.
Pp. 23-54.
Rylands, A.B., Mittermeier, R.A., Rodríguez-Luna, E. (1995). A species list for the New World
primates (Platyrrhini): distribution by country, endemism, and conservation status according to
the Mace-Land system. Neotrop. Prim. 3: 113–160.
Rumiz, D.I. 1990. Alouatta caraya: population density and demography in Northern Argentina.
American Journal of Primatology 21:279–294.
Sambrook, J., Fritsch, E.F., Maniatis, T. 1989. Molecular Cloning. A laboratory manual. 2a.
Edition. Cold Spring Harbor Press.
Simonsen, K., Churchill, G., Aquadro, C. (1995). Properties of Statistical Tests of Neutrality for
DNA Polymorphism Data. Genetics 141: 413-429.
Swofford, D.L. (2002). PAUP*. Phylogenetic analysis using parsimony and other methods.
http://paup.csit.fsu.edu. pp. 1-142.

51
Tajima, F. 1993. Statistical analysis of DNA polymorphism. Japanese Journal of Genetics (68):
567-595
Tajima, F. 1983. Evolutionary relationships of DNA sequences in finite populations. Genetics
105: 437-460.
Tajima, F. (1989). Statistical method for testing the neutral mutation hypothesis by DNA
polymorphism. Genetics 123: 585-595.
Tamura K., Stecher G., Peterson D., Filipski A., and Kumar S. (2013). MEGA6: Molecular
Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution30: 2725-2729.
Torres, O.M., Leibovici, M. (2001). Caracterización del cariotipo del mono aullador colorado
Alouatta seniculus que habita en Colombia. Caldasia 23: 537-548.
Walsh, P., Metzger, D. A. and Higuchi, R. 1991. Chelex 100 as a medium for simple extraction
of DNA for PCR-based typing from forensic material. Bio Techniques. 10: 506- 513.
Weir, B. and Hill, W. 2002. Estimating F-statistics. Annu. Rev. Genet. 2002. 36:721–50
Wright, S. 1951. The genetical structure of populations. Ann. Eugenetics. 15: 323-354

52
Zunino, G.E.; Gonzalez, V.; Kowalewski, M.M.; Bravo, S.P. 2001. Alouatta caraya. Relations
among habitat, density and social organization. Primate Report 61: 37-46.
Zunino, G. 1986. Algunos aspectos de la ecologia y etologia del mono aullador negro
(Alouattacaraya) en habitat fragmentados. Tesis doctoral, Buenos Aires, 152 pp.





























