Embed Size (px)
Citation preview

1
FENOMENI DITRASPORTO
I trasporti attraversole membrane biologiche

2
La struttura a doppio strato l ipidico delle membrane biologiche condiziona fortementeil movimento di gas, ioni e molecole attraverso le membrane. L’interno idrofobico deldoppio strato l ipidico rende le membrane impermeabili alla maggior parte degli ioni edelle molecole cariche. In generale, mentre i gas come O2, CO2, NO e le molecolelipofi l iche permeano facilmente attraverso le membrane, gli ioni e le molecoleidrosolubil i permeano con molta difficoltà o non permeano affatto attraverso la faselipidica della membrana.Tuttavia, i movimenti di sostanze cariche sono necessari per i l corretto funzionamentocellulare e pertanto tutte le cellule hanno sviluppato e perfezionato adeguati sistemimolecolari per facil itarne il movimento. Anche grosse proteine e particelle possonopermeare la membrana cellulare attraverso specific i sistemi di trasporto.

3
Sistemi di trasporto di membrana
Trasporto passivo Trasporto attivo
Diffusione semplice
Diffusionefacilitata
Trasporto Attivo
primario
Trasporto Attivo
secondario
Indipendenteda un vettore Dipendente da un vettore
Endocitosi EsocitosiTranscitosi
Canale Carrier
I sistemi di trasporto attraverso la membrana possono essere schematizzati come segue:È possibile differenziarli in base al fatto che avvengano secondo gradiente diconcentrazione chimica o elettrochimica di una sostanza in cui non vi è dispendio dienergia metabolica della cellula, ed in questo caso si parla di trasporto passivo, cheviceversa avvengano contro gradiente elettro-chimico e di conseguenza necessitino dienergia metabolica, in questo caso si parla di trasporto attivo.Nell ’ambito del trasporto passivo, ovvero secondo gradiente di concentrazione sipossono a loro volta distinguere processi di trasporto che avvengono per diffusionesemplice, c ioè senza l’ausil io di alcuna proteina trasportatrice, da processi di diffusionefacil itata, ancora una volta secondo gradiente elettrochimico, che però richiedonol’intervento di vettore, ovvero una specifica proteina di trasporto o di un canale.Nel trasporto attivo che come detto avviene contro gradiente di concentrazione, sipossono distinguere 2 modalità: nella prima, denominata trasporto attivo primario, laproteina di membrana uti l izza direttamente l ’energia metabolica fornita dall ’idrolisi delATP per creare ad esempio gradienti ionic i di Na e K tra l ’ambiente intra edextracellulare. Nel secondo caso, ovvero nel trasporto attivo secondario, i l trasporto diuna sostanza (ione o molecola) contro i l proprio gradiente di concentrazione èaccoppiato al movimento generalmente di uno ione secondo il proprio gradienteelettrochimico, dipendente a sua volta dall ’attività di un trasporto attivo primario.Al trasporto attivo appartengono i fenomeni di endo ed esocitosi (internalizzazione oespulsione di proteine ed altre macromolecole o fluidi mediante invaginazione oestroflessione della membrana plasmatica) nonché i processi di transcitosi(attraversamento di un epitelio).

4
La diffusione è il processo mediante il quale si mescolano atomio molecole dotati di moto casuale termico (moto browniano)
Tutte le molecole o particelle in soluzione possiedono una energia termica: esse simuovono casualmente, urtano tra loro e cambiano continuamente direzione. Se inun determinato volume le molecole sono più concentrate queste diffonderannoverso zone a più bassa concentrazione fino a che non si distribuirannouniformemente in ogni parte dello spazio disponibile. Questo processo è noto con iltermine di diffusione. La diffusione è un processo passivo estremamente lento alivello macroscopico (tissutale) mentre è rapido a l ivello microscopico (cellulare).

5
È un processo passivo che nonrichiede fonti di energia.
Le molecole diffondono da una zona amaggiore concentrazione verso zone apiù bassa concentrazione.
La diffusione delle molecole procedefino a quando la concentrazione èuguale in entrambi i lati dellamembrana.
La diffusione è rapida a breve distanza emolto lenta su scala macroscopica.
La diffusione è direttamente proporzionalealla temperatura. Ad alte temperature ilmovimento molecolare è più rapido e ladiffusione più veloce.
La diffusione è inversamenteproporzionale alla dimensionemolecolare.
La diffusione ha alcune proprietà:• È un processo passivo che non richiede fonti di energia. Usa solo l ’energia termica
(cinetica) del movimento molecolare.1. le molecole diffondono da una zona a maggiore concentrazione verso zone a più
bassa concentrazione. Più grande è la differenza di concentrazione e maggiore èla velocità delle molecole che diffondono per unità di tempo e per area dimembrana.
2. la diffusione delle molecole procede fino a quando la concentrazione è uguale inentrambi i lati della membrana. Si raggiunge quindi un equil ibrio dinamico e i lmovimento delle molecole continua senza spostamenti netti.
3. La diffusione è rapida a breve distanza e molto lenta su scala macroscopica.4. la diffusione è direttamente proporzionale alla temperatura. Ad alte temperature i l
movimento molecolare è più rapido e la diffusione più veloce.5. la diffusione è inversamente proporzionale alla dimensione molecolare.6. La diffusione può verificarsi in un sistema aperto o attraverso una barriera che
separa due sistemi.
La diffusione attraverso le membrane è un po’ più complicata rispetto alla diffusione inun sistema aperto. Chiaramente solo le sostanze che possono attraversare i l centrolipidido di una membrana possono muoversi per diffusione.

6
la velocità di diffusione dipendedalla capacità della molecolache diffonde di sciogliersi nellostrato lipidico della membrana
il tasso di diffusione attraversouna membrana è direttamenteproporzionale all’area dellasuperficie della membrana.
il tasso di diffusione attraversouna membrana è inversamenteproporzionale allo spessoredella membrana.
Se una membrana cellulare è permeabile a una molecola, tale molecola può entrareo uscire dalla cellula per diffusione. È necessario però che tali molecole siano ingrado di passare attraverso il centro l ipidico di una membrana cellulare.
La diffusione attraverso attraverso il doppio strato l ipidico della membrana va sotto i lnome di diffusione semplice e possiede, oltre a tutte le proprietà elencate primaanche quelle seguenti:
1. la velocità di diffusione dipende dalla capacità della molecola che diffonde disciogliersi nello strato l ipidico della membrana. E c ioè di quanto la membrana siapermeabile alle molecole che diffondono. Di regola solo i l ipidi, gli steroidi e lepiccole molecole l ipofi le possono attraversare le membrane per diffusionesemplice. Ci sono però delle eccezioni che poi vedremo.
2. i l tasso di diffusione attraverso una membrana è direttamente proporzionaleall ’area della superfic ie della membrana. Quanto più grande è l ’area dellasuperfic ie della membrana tanto più molecole possono diffondere attraverso di essanell ’unità di tempo. È un concetto che può sembrare ovvio ma che ha profondeimplicazioni fisiologiche. Per es. nella malattia polmonare denominata enfisema iltessuto polmonare collassa e viene distrutto questo determina una drasticariduzione dell’area della superfic ie di scambio per la diffusione dell ’ossigeno e diconseguenza meno ossigeno arriva ai tessuti.
3. i l tasso di diffusione attraverso una membrana è inversamente proporzionale allospessore della membrana. Quanto più la membrana è spessa tanto più lenta e lavelocità alla quale la diffusione ha luogo.

7
LEGGE DI FICK
J = velocità di diffusione (flusso)
C1 e C2 = concentrazione della molecolanel compartimento 1 e 2
P = permeabilità di una molecolaattraverso la membrana
Le regole della diffusione semplice attraverso le membrane possono essere espressematematicamente da una equazione nota come Legge di Fick sulla diffusione chepermette di calcolare la velocità di diffusione J (flusso) quando sono note leconcentrazioni iniziali nei due compartimenti (C1 e C2), e la permeabilità di unamolecola attraverso la membrana.

8
LEGGE DI FICK
D = coefficiente di diffusione della molecola
A = area della sezione di scambio
K = coefficiente di partizione della sostanzatra fase acquosa e la fase lipidica dellamembrana
x = distanza dove le due concentrazionisono calcolate
La permeabilità della molecola influenza chiaramente il flusso di un solutoattraverso la membrana. Maggiore è il valore di P, maggiore è il flusso disoluto tra i due compartimenti
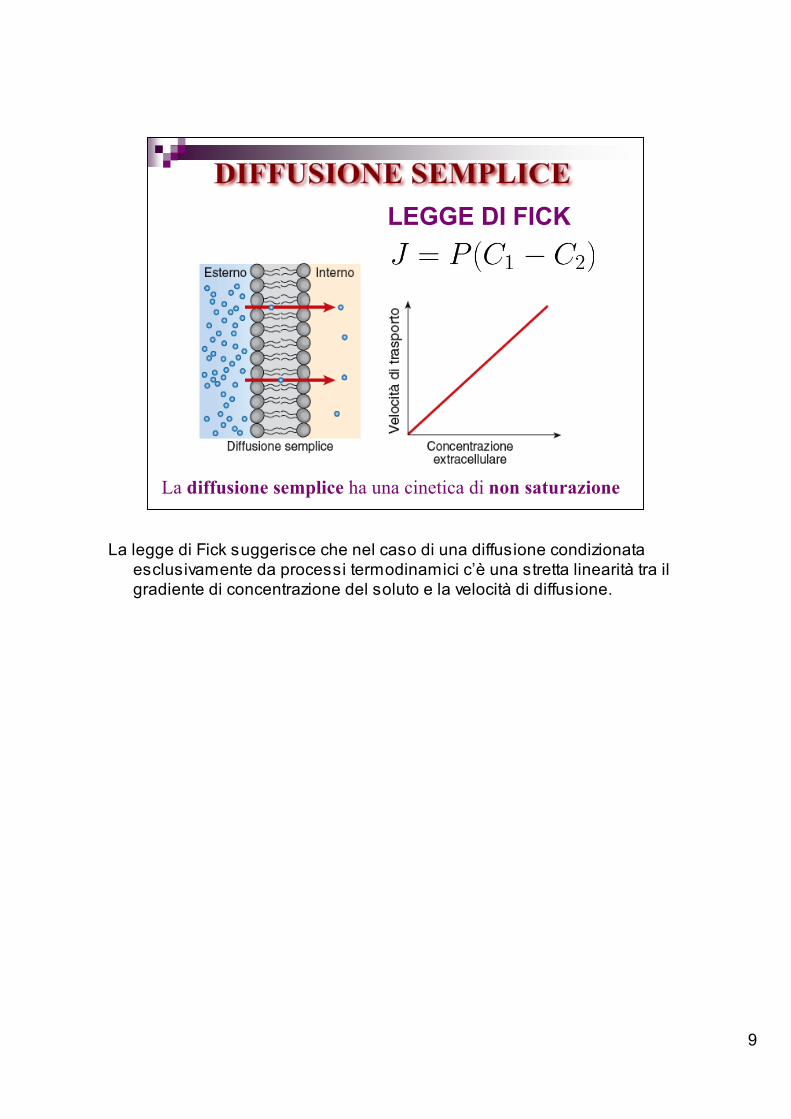
9
LEGGE DI FICK
La diffusione semplice ha una cinetica di non saturazione
La legge di Fick suggerisce che nel caso di una diffusione condizionataesclusivamente da processi termodinamici c’è una stretta linearità tra ilgradiente di concentrazione del soluto e la velocità di diffusione.

10
Per poter abbandonare la faseacquosa ed entrare nella faselipidica, un soluto deve primadi tutto rompere i legamiidrogeno con l’acqua e poi deveessere solubile nella faselipidica (la concentrazione inmembrana aumentaall’aumentare del coefficiente Kdi partizione olio/acqua)
Per poter abbandonare la fase acquosa ed entrare nella fase lipidica, un soluto deve prima ditutto rompere i legami idrogeno con l’acqua (per la rottura di un legame idrogeno sirichiedono 5 kcal/mol). Inoltre la molecola di soluto deve essere solubile nella fase lipidicaed il suo grado di solubilità sarà determinante per l’attraversamento della membrana. (laconcentrazione in membrana aumenta all’aumentare del coefficiente di partizioneolio/acqua, K)

11
Quindi tutte le sostanze che formano pochi legami idrogeno con l’acqua possono entrare piùfacilmente nel doppio strato lipidico mentre gli ioni inorganici non avranno quasi mai lapossibilità di farlo.In genere molecole piccole e non polari come Ossigeno (32 dalton) e anidride carbonica (44dalton) e molecole fortemente idrofobiche (o lipofiliche) diffondono rapidamente attraverso idoppi strati lipidici.Piccole molecole polari come acqua (18 dalton), etanolo (46 dalton) e urea (60 dalton)attraversano rapidamente;
il glicerolo (92 dalton) diffonde meno rapidamente mentre il glucosio, che è una grossamolecola polare, praticamente non diffonde.I doppi strati lipidici sono praticamente impermeabili a molecole cariche e a ioni per quantopiccole esse siano; la carica e l’alto grado di idratazione di queste molecole ne impediscel’ingresso nella fase idrocarburica del doppio strato.
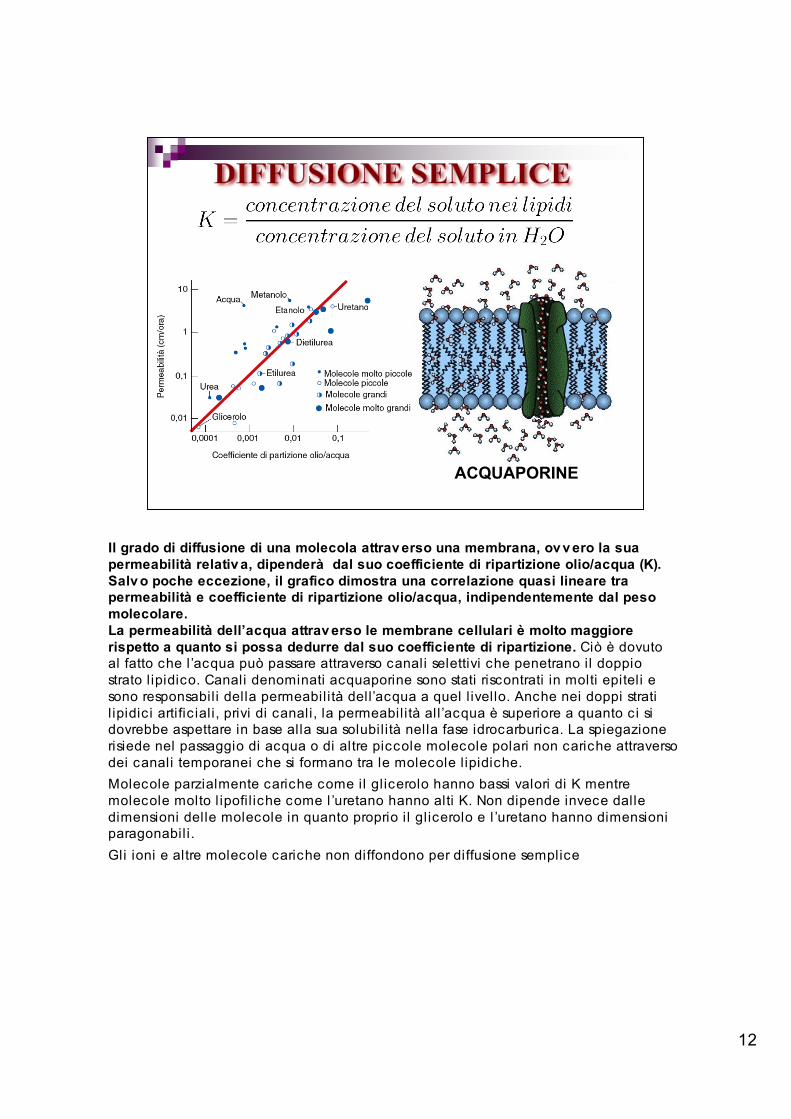
12
ACQUAPORINE
Il grado di diffusione di una molecola attrav erso una membrana, ov v ero la suapermeabilità relativ a, dipenderà dal suo coefficiente di ripartizione olio/acqua (K).Salv o poche eccezione, il grafico dimostra una correlazione quasi lineare trapermeabilità e coefficiente di ripartizione olio/acqua, indipendentemente dal pesomolecolare.La permeabilità dell’acqua attrav erso le membrane cellulari è molto maggiorerispetto a quanto si possa dedurre dal suo coefficiente di ripartizione. Ciò è dovutoal fatto che l ’acqua può passare attraverso canali selettivi che penetrano il doppiostrato l ipidico. Canali denominati acquaporine sono stati riscontrati in molti epiteli esono responsabil i della permeabilità dell’acqua a quel l ivello. Anche nei doppi stratil ipidic i artific iali, privi di canali, la permeabilità all ’acqua è superiore a quanto c i sidovrebbe aspettare in base alla sua solubil ità nella fase idrocarburica. La spiegazionerisiede nel passaggio di acqua o di altre piccole molecole polari non cariche attraversodei canali temporanei che si formano tra le molecole l ipidiche.Molecole parzialmente cariche come il glicerolo hanno bassi valori di K mentremolecole molto l ipofi l iche come l ’uretano hanno alti K. Non dipende invece dalledimensioni delle molecole in quanto proprio i l glicerolo e l ’uretano hanno dimensioniparagonabili.Gli ioni e altre molecole cariche non diffondono per diffusione semplice

13
Non è richiesta energia metabolica. I soluti si muovono lungo un gradiente dipotenziale elettrochimico combinati ad una proteina carrier.
Proteine trasportatrici chetraslocano una sostanza daun ambiente all’altro
Proteine canale cheformano dei pori attraversola membrana consentendoil passaggio di ioni delladimensione e caricaappropriata.
Ioni e piccole molecole cariche e molecole polari come zuccheri, amminoacidi eccnon possono attraversare i l doppio strato l ipidico a causa della loro forte polarità. Il lorotrasporto è mediato da proteine di membrana che possono essere di due tipi: leproteine trasportatric i che traslocano una sostanza da un ambiente all ’altro e leproteine canale che formano dei pori attraverso la membrana consentendo il passaggiodi ioni della dimensione e carica appropriata.Nel caso della diffusione facil itata la relazione è l ineare a basse concentrazioni per poisaturare in maniera diversa ad alte concentrazioni.

14
Ioni e piccole molecole carichediffondono attraverso canali ionici,proteine integrali di membrana(composte da più subunità)
DIFFUSIONE FACILITATA
Il movimento degli ioni avvienesecondo gradienteelettrochimico ed è strettamentecondizionato dalla selettività delcanale. In assenza di ungradiente elettrochimico uncanale ha uguali probabilità ditrasportare uno ione in entrambele direzioni.
In particolare gli ioni e le piccole molecole attravesano la membrana plasmaticaattraverso canali ionic i. Le proteine canale sono costituite da una o più subunitàproteiche i cui componenti attraversano ripetutamente i l doppio strato l ipidico e sidistribuiscono in modo da formare un poro acquoso all ’interno della membrana.Quando il canale è aperto mette in comunicazione diretta i l compartimento intra- edextracellulare e permette i l passaggio rapido e senza ostacoli di decine di milioni diioni al secondo. Il movimento degli ioni avviene secondo gradiente elettrochimico edè strettamente condizionato dalla selettività del canale. In assenza di un gradienteelettrochimico un canale ha uguali probabil ità di trasportare uno ione in entrambe ledirezioni.

15
DIFFUSIONE FACILITATA
VESTIBOLO
FILTRO DISELETTIVITÀ
In generale un canale è strutturalmente costituito da un vestibolo piuttostolargo dove lo ione idratato può accedere facilmente per diffusione e da unfiltro di selettività, più stretto, che forma il poro vero e proprio e seleziona gliioni in base alla carica degli aminoacidi che ne rivestono la parete e aldiametro dell’apertura. Gli ioni devono velocemente deidratarsi per poterpermeare e, nel caso di più siti di legame, gli ioni deidratati si spostandosaltando da un sito all’altro in “fila indiana”.

16
DIFFUSIONE FACILITATA
SELETTIVITÀ
La selettività di un canale èstrettamente determinata dal tipo di
cariche nette (positive o negative) cheformano le pareti del foro di
permeazione.
1.Canali con alta selettività: Canali del Na+, Canalidel K+, Canali del Ca2+, Canali del Cl-
2.Canali con selettività intermedia: Canali del Na+,K+, Canali del Na+, K+, Ca2+
Dal punto di vista funzionale i canali si distinguono in base alla loroselettività e in base alla modalità di attivazione.In base alla selettività si distinguono canali per un solo ione (Na+, K+,Ca2+, Cl-) e canali con selettività intermedia permeabili a Na+ e K+ oppurea Na+, K+ e Ca2+. La selettività di un canale è strettamente determinata daltipo di cariche nette (positive o negative) che formano le pareti del foro dipermeazione. Per quanto riguarda i canali del Na+, K+ e Ca2+, i gruppicarichi delle pareti sono costituiti da gruppi carbossilici (COO-) delle catenelaterali dell’acido glutammico e aspartico a carica netta negativa, mentreper i canali permeabili a Cl- le pareti sono costituite da gruppi amminici(NH3+) degli aminoacidi arginina e lisina a carica netta positiva.

17
DIFFUSIONE FACILITATA
MODALITÀ DI ATTIVAZIONE1. Canali voltaggio dipendenti: la cui probabilità di
apertura dipende strettamente dal potenziale elettricodella membrana.
2. Canali attivati dai ligandi: la cui probabilità di aperturae chiusura è determinata dall’azione di particolarimolecole messaggere.
3. Canali attivati da secondi messaggeri la cui probabilitàdi apertura è condizionata dalla presenza nel liquidointracellulare di ioni e molecole comunemente indicaticome secondi messaggeri intracellulari come Ca2+, ATP,cAMP, cGMP, IP3).
In base alla loro modalità di attivazione i canali si suddividono in tre categorie. In tuttie tre i casi i l canale è strutturato in modo tale da avere due conformazioni: unaconformazione a canale chiuso (non permeabile) e una a canale aperto(permeabile). Si parla anche di canali a porta (gate) che differiscono in base almeccanismo che determina l ’apertura o la chiusura del gate:
1. Canali voltaggio dipendenti: la cui probabil ità di apertura dipende strettamentedal potenziale elettrico della membrana. Il cambiamento di conformazione delcanale da chiuso ad aperto è dovuto allo spostamento di cariche elettriche.
2. canali attivati dai l igandi: la cui probabil ità di apertura e chiusura è determinatadall ’azione di particolari molecole messaggere che agiscono extracellularmente(neurotrasmettitori) e inducono cambi conformazionali capaci di aprire i l poro dipermeazione.
3. canali attivati da secondi messaggeri la cui probabilità di apertura è condizionatadalla presenza nel l iquido intracellulare di ioni e molecole comunenmente indicaticome secondi messaggeri intracellulari come Ca2+, ATP, cAMP, cGMP, IP3).

18
Alcune molecole polari come zuccheri,aminoacidi, nucleotidi e alcunimetaboliti diffondono attraverso lamembrana utilizzando specificheproteine trasportatrici (carrier)attraverso una diffusione facilitata
DIFFUSIONE FACILITATA TRASPORTO ATTIVO SECONDARIO
Alcune molecole polari come zuccheri, aminoacidi, nucleotidi e alcuni metabolitidiffondono attraverso la membrana uti l izzando specifiche proteine trasportatric i (carrier)attraverso una diffusione facil itata. Si sviluppa secondo gradiente in entrambe ledirezioni.Le proteine carrier si legano a substrati specific i e l i trasportano attraverso la membranacambiando conformazione.La molecola da trasportare si lega al carrier da un lato della membrana . Questolegame cambia la conformazione del carrier così che il cancello aperto si chiude.Dopo una breve fase di transizione in cui entrambi i cancell i sono chiusi, i l latoopposto del carrier si apre dall ’altro versante della membrana. Il carrier allora ri lascia lamolecola e i l suo ri lascio determina una variazione conformazionale che fa ritornare i lcarrier nella sua conformazione iniziale.La diffusione facil itata mediante carrier ha carattere di uniporto in quanto vienetrasportato solo un tipo di molecola e si distingue nettamente dal trasporto accoppiatoindicato come cotrasporto che uti l izza anche esso proteine trasportatric i ma si tratta diun trasporto attivo secondario.

19
PROPRIETÀ DEL TRASPORTO FACILITATO• Il trasporto facilitato utilizza il gradiente di potenziale
elettrochimico della specie permeante fino alraggiungimento dell’equilibrio (trasporto passivoequilibrante)
• La velocità di trasporto si riduce in presenza dimolecole che possono legarsi allo stesso sito ditrasporto (inibizione competitiva)
• Il flusso unidirezionale onetto cresce secondo unacurva iperbolica (cinetica disaturazione) in funzionedella concentrazione delsubstrato che può andaread occupare tutti i siti dilegame del carrier.
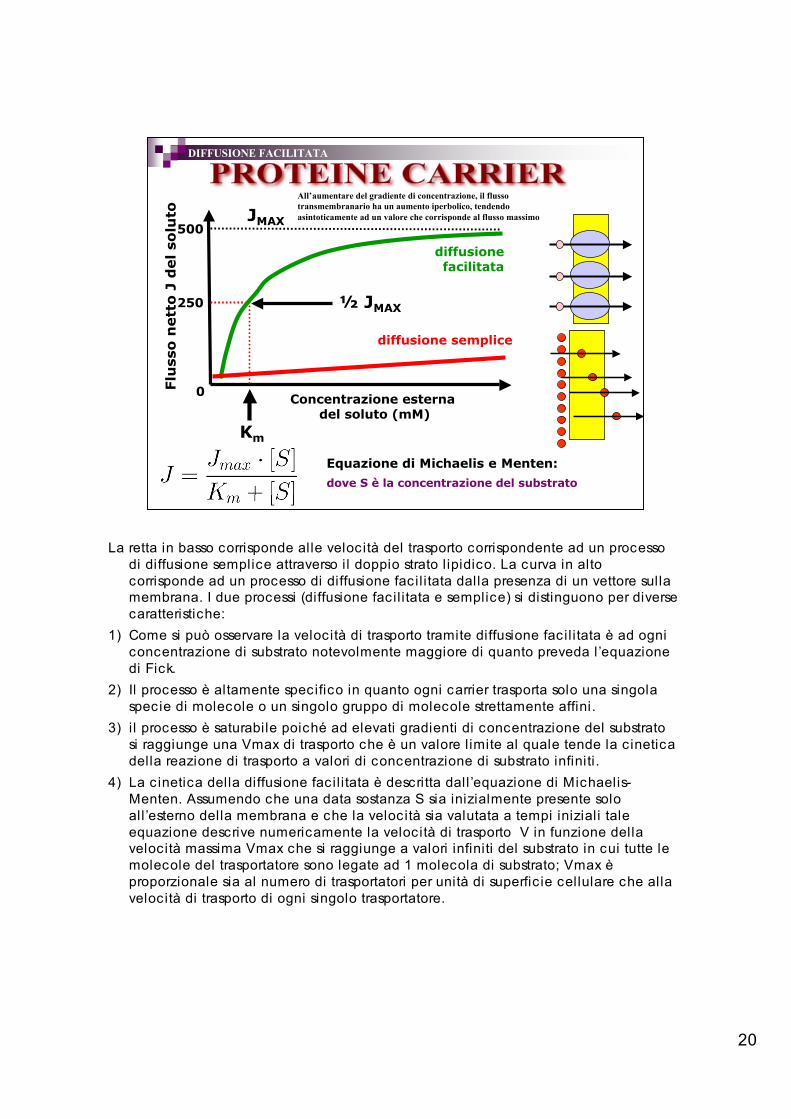
20
JMAX
0
250
500
Flu
sso
nett
o J
del
solu
to
Concentrazione esterna del soluto (mM)
Km
½ JMAX
diffusionefacilitata
diffusione semplice
Equazione di Michaelis e Menten:
dove S è la concentrazione del substrato
DIFFUSIONE FACILITATA
All’aumentare del gradiente di concentrazione, il flussotransmembranario ha un aumento iperbolico, tendendoasintoticamente ad un valore che corrisponde al flusso massimo
La retta in basso corrisponde alle velocità del trasporto corrispondente ad un processodi diffusione semplice attraverso il doppio strato l ipidico. La curva in altocorrisponde ad un processo di diffusione facil itata dalla presenza di un vettore sullamembrana. I due processi (diffusione facil itata e semplice) si distinguono per diversecaratteristiche:
1) Come si può osservare la velocità di trasporto tramite diffusione facil itata è ad ogniconcentrazione di substrato notevolmente maggiore di quanto preveda l ’equazionedi Fick.
2) Il processo è altamente specifico in quanto ogni carrier trasporta solo una singolaspecie di molecole o un singolo gruppo di molecole strettamente affini.
3) i l processo è saturabile poiché ad elevati gradienti di concentrazione del substratosi raggiunge una Vmax di trasporto che è un valore l imite al quale tende la c ineticadella reazione di trasporto a valori di concentrazione di substrato infiniti.
4) La c inetica della diffusione facil itata è descritta dall ’equazione di Michaelis-Menten. Assumendo che una data sostanza S sia inizialmente presente soloall ’esterno della membrana e che la velocità sia valutata a tempi iniziali taleequazione descrive numericamente la velocità di trasporto V in funzione dellavelocità massima Vmax che si raggiunge a valori infiniti del substrato in cui tutte lemolecole del trasportatore sono legate ad 1 molecola di substrato; Vmax èproporzionale sia al numero di trasportatori per unità di superfic ie cellulare che allavelocità di trasporto di ogni singolo trasportatore.

21
DIFFUSIONE FACILITATA
Km (costante di Michaelis-Menten ) corrispondealla concentrazione della sostanza alla quale siraggiunge la velocità semi-massimale
Km corrisponde alla concentrazione della sostanza alla quale si raggiunge la velocitàsemi-massimale; S è la concentrazione della sostanza in esame. Km, c ioè lacostante di Michaelis-Menten è un indice dell’affinità della sostanza in esame perla proteina carrier, ed in particolare è inversamente correlata all’affinità di legamesubstrato-trasportatore; di conseguenza quanto più basso è i l valore di Km tantomaggiore è l ’affinità del substrato per i l trasportatore.
Un’altra caratteristica dei processi di diffusione facil itata che li differenzia dalladiffusione semplice è che essi possono essere inibiti. Nel caso più semplice c ioèl’inibizione competitiva si verifica una riduzione del trasporto di una molecola A,es. D-glucosio in presenza di una molecola B, ed. D- galattosio che le somiglia dalpunto di vista strutturale e compete con il D-glucosio per i l legame al trasportatore.Cineticamente questo tipo di inibizione è facilmente distinguibile da inibizioni ditipo non-competititvo o acompetitivo in quanto si verifica un variazione dellacostante di Michalis-Menten verso valori maggiori, e di conseguenza in direzione diuna riduzione dell ’affinità enzima-substrato.

22
Il trasportatore (uniporto) del glucosio GLUT
Struttura secondaria GLUT
Sito di glicosilazione
Possibile localizzazione del sito dilegame esterno del glucosio
Possibile localizzazione del sito dilegame interno del glucosio
DIFFUSIONE FACILITATA
I carrier GLUT sono proteinedi 45.000 daltons con 12domini transmembranadisposti a cerchio all’internodella membrana.Sono specifici per la forma D-del glucosio e riconosconoanche D-mannosio e D-galattosio.
I trasportatori GLUT sono l’esempio più studiato di diffusione facilitata acarattere di uniporto.

23
DIFFUSIONE FACILITATA
Secondo il modello accettato il trasportatore possiede due staticonformazionali E1 e E2, il primo con i siti leganti il substrato rivolti
verso l’ambiente esterno, mentre lo stato E2 è caratterizzatodall’esposizione dei siti verso l’ambiente intracellulare.
E1E2
Nel GLUT1 il D-glucosio si lega al suo sito, che nellaconformazione E1 è rivolto verso l’ambiente extracellulare.
Questo legame comporta una modificazione nella conformazioneE2 portando il sito di legame verso il lato citoplasmatico.
Nell’ambiente intracellulare le concentrazioni di glucosiosono molto inferiori a quelle vigenti all’esterno e quindi ilsubstrato si stacca dal suo sito di legame. Le concentrazioniintracellulari di D-glucosio si mantengono a bassi livelli inquanto una esochinasi lo trasforma in glucosio-6-fosfato, chenon è riconosciuto dal trasportatore. Di conseguenza ilgradiente di concentrazione favorevole all’ingresso di D-glucosio rimane inalterato. Una volta che lo zucchero si èstaccato il sistema di trasporto scarico può riconvertirsinella conformazione E2 e riprendere così il ciclo di trasporto.

24
Trasportatore del fruttosio
Trasportatore intracellulare(Membrana Ret.endoplasmico)
6-14 mM-GLUT5
GLUT7
Insulino-dipendente(tessuto adiposo e muscolo)
- 2 mMGLUT4
Neuronale-10 mMGLUT3
EpitelialeEpatociti e membrana
basolaterale epiteli
66 mM42 mMGLUT2
Costitutivo(eritrociti; > parte dei tessuti)
5 M20 mMGLUT1
Localizzazione e funzione
Km
(D-Fruct)Km
(D-glucose)Sistema ditrasporto
Il trasportatore (uniporto) del glucosio GLUT
DIFFUSIONE FACILITATA
Al momento si conoscono 6 isoforme di trasportatori GLUT con diversa affinità per i lglucosio. Con l’eccezione di GLUT 5 che trasporta principalmente fruttosio, tutte lealtre isoforme trasportano glucosio e sono distribuite eterogeneamente nei vari tessuti.La prima isoforma denominata GLUT1 è presente in molti tessuti, compresa lamembrana eritrocitaria, ed è denominato costitutivo; l’isoforma 2 è maggiormenteespressa negli epiteli intestinale e renale dove facil ità i l flusso trans-epiteliale delglucosio; l’isoforma 3 è caratteristica del tessuto nervoso; mentre la 4 maggiormenteespressa negli epatociti è estremamente importante nelle patologia del diabetemellito, in quanto è l ’isoforma soggetta al controllo dell ’insulina; infine i l GLUT5espresso a l ivello intestinale, ha una discreta affinità per i l fruttosio favorendo iltrasporto trans-membrana di questo zucchero.

25
È richiesta energia metabolica (ATP)poiché i soluti si muovono CONTROgradiente di potenziale elettrochimico
Tipi di trasporto attivo
Trasportoattivo
primario
Trasportoattivo
secondario
A seconda di come viene uti l izzato l’ATP, i l trasporto attivo può esseresuddiviso in trasporto attivo primario che uti l izza direttamente l’energia chederiva dal legame fosfato dell’ATP e il trasporto attivo secondario che uti l izzainvece l ’energia di un gradiente di concentrazione che è statoprecedentemente creato da un trasporto attivo primario. È ovvio quindi che tuttii trasporti attivi secondari dipendono strettamente dai trasporti attivi primari cheutil izzano ATP per mantenere i gradienti di concentrazioni necessari per i ltrasporto attivo secondario.I meccanismi delle proteine responsabil i del trasporto attivo assomigliano peralcuni aspetti a quell i della diffusione facil itata. I substrati, ioni o molecole, silegano alla proteina carrier, che cambia conformazione ri lasciando il substratonel lato opposto. Esistono però due sostanziali differenze per quanto riguarda leproteine del trasporto attivo primario: 1) i l cambio di conformazione avvienegrazie all ’apporto di energia dell ’ATP, mentre nel caso della diffusionefacil itata non è richiesta alcuna energia metabolica e 2) l ’affinità dellaproteina carrier per i l l igando si modifica nel passaggio da un lato all’altro dellamembrana. Solo in questo modo possono essere legati e ceduti ioni controgradiente

26
.
4 classi di proteine di trasporto ATP-dipendenti.
Le pompe dellaclasse P sono
costituite da 2 peptidi,α e β, e passano in uno
stato fosforilato(subunità α) durante il
ciclo di trasporto.
Le pompe delle classi F e Vnon danno origine ad intermedi
fosforilati.Hanno strutture e proteine simili,
ma le loro subunità non hannoalcuna relazione con quelle delle
pompe della classe P
Le proteine ABC hanno 4domini: 2 transmembrana
(T) e 2 citosolici legantiATP (A) che accoppiano
l'idrolisi di ATP al trasportodel soluto. In certe ABC, i
domini sono presenti comesubunità separate mentrein altre risultano fusi in un
singolo polipeptide.
Il trasporto attivo primario uti l izza proteine di membrana che cambianoconformazione uti l izzando l’energia chimica derivata dall ’idrolisi dell ’ATP inADP e fosfato. In questo senso tali proteine sono dei veri e propri enzimi evengono comunamente definite ATPasi.Possiamo distinguere quattro c lassi di proteine di trasporto ATP-dipendenti. Le pompe della c lasse p (P-type ATPasi) sono costituite da due diversi peptidi,alfa e beta, e passano in uno stato fosforilato durante i l c ic lo di trasporto. Lasequenza attorno al residuo che viene fosforilato, situato nelle subunità alfa èomologa nelle diverse pompe.Le pompe delle c lassi F e V non danno origine ad intermedi fosforilati . Le lorostrutture sono simili e contengono proteine simili, ma le loro subunità nonhanno alcuna relazione con quelle delle pompe della c lasse P.Tutti i membri dell 'ampia superfamiglia di proteine ABC (ATP-binding cassettetransporter) contengono quattro domini essenziali: due domini transmembrana(T) e due domini c itosolic i leganti ATP (A) che accoppiano l 'idrolisi di ATP altrasporto di un dato soluto. In alcune proteine ABC, questi domini sono presenticome subunità separate (come nel caso il lustrato) mentre, in altre, risultano fusiin un singolo polipeptide.

27
ATP
ADP
ATPasi per il trasporto ionico sono le fonti principali e anche i maggiori utilizzatori di ATP
Ca2+
ER
Ca2+- ATPasi(reticolo endo-sarco-plasmatico)SERCA
Ca2+Ca2+-ATPasi(plasmalemma)PMCA
K+
H+
H+/K+ATPasi: Ghiandole gastriche; Dotti collettori; Secrezione HCl (stomaco); Riassorbimento bicarbonato reneK+/H+ATPasi : Colon distale; sensibile a ouabaina e omeprazolo; Assorbimento di K+ ed escrezione di H+
K+
Na+ Na+K+ ATPasiInibitore: ouabainaubiquitaria;funzione house-keeping
H+
AA+
V-type ATPasiLisosomi;endosomi;granuli dideposito;acidificazioneintravescicolare
ATP
H+
ADP+P F-type ATPasiMembranamitocondriale internaProduzione di ATP
Trasportatori ABC(MDR, CFTR)App. gastro-entericoe renale; celluletumorali; epiteliopolmonareEscrezione difarmaci e sostanzeidrofobiche (MDR),di ioni cloro (CFTR)

28
Na+/K+ATPasiLa pompa
Na+/K+ATPasi consistedi 4 subunità principali
(2 α e 2 β)
subunità α = 1000 aa; 110 KDa, 10 segmenti transmembrana.contiene i siti di legame per Na+ e ATP e un sito di fosforilazione neldominio citoplasmatico; nel dominio extracellulare ha i siti di legameper il K+ e la ouabainasubunità β = 300 aa; ~45 KDa, 1 segmento transmembrana, senzaattività enzimatica e di trasporto. La sua associazione con la α-subunitàè necessaria per l’attività della pompa in quanto stabilizza la subunità αall’interno della membrana
TRASPORTO ATTIVO PRIMARIO P-Type ATPasi
La pompa Na+/K+ATPasi è stata laprima pompa ad esserestudiata.

29
Na+/K+ATPasiK+
Na+
La Na+/K+-ATPasi mantiene i gradienti di Na+ e K+ tral’ambiente intra e quello extra-cellulare
[K+] = 145 mM[Na+] = 9,2 mM
[K+] = 2,5 mM
[Na+] = 140 mM
TRASPORTO ATTIVO PRIMARIO P-Type ATPasi
Ha il compito principale di mantenere stabili i gradienti di concentrazione disodio e potassio ai lati della membrana.
Oltre a possedere tutti i requisiti dei sistemi ditrasporto facilitato precedentemente analizzati(velocità, specificità, saturabilità, etc..), in analogia atutti i sistemi di trasporto attivo primario ècaratterizzata funzionalmente dal lavorare controgradiente di concentrazione sia dello ione sodio chedel potassio in quanto trasporta il Na dall’ambienteinterno dove è meno concentrato a quello esternodove è più concentrato; viceversa il potassio vienetraslocato dall’ambiente extracellulare a minorconcentrazione a quello intracellulare aconcentrazione maggiore.

30
Na+/K+ATPasi
TRASPORTO ATTIVO PRIMARIO P-Type ATPasi
La pompa dal versante intracellulare ha un’alta affinità per il Na e unabassa affinità per il K. Ciò facilita l’attacco degli ioni Na ai propri siti dilegame a discapito della loro bassa concentrazione all’interno della cellula(circa 9 mM). Si legano quindi 3 ioni Na. A questo punto avviene lafosforilazione dei siti di legame intracellulari dell’ATP (step 2) che induce ilcambio di conformazione necessario per permettere il cambio di direzionedella pompa e la liberazione del Na nel liquido extracellulare (step 3 e 4).Nello stato fosforilato, la pompa ha un’alta affinità per il K e una bassaaffinità per il Na sul lato extracellulare. Quindi il Na viene rilasciato,nonostante la sua concentrazione all’esterno della cellula sia elevata (145mM) mentre si legano due ioni K pur essendo bassa la loro concentrazioneche è intorno a 4 mM (step 5). L’attacco degli ioni K sul lato extracellulareattiva la liberazione del gruppo fosfato (step 6) che fa variare laconformazione della pompa (step 7) con il rilascio degli ioni K nel latointracellulare contro gradiente di concentrazione (140 mM).

31
TRASPORTO ATTIVO PRIMARIO P-Type ATPasi
In questo modo il ripetersi del ciclo permette di mantenere bassi i livelli diNa e alti i livelli di K all’interno della cellula e una situazione oppostaall’esterno.L

32
La Na+/K+-ATPasi mantiene i gradienti di Na+ e K+ tra l’ambiente intra equello extra-cellulare
Caratteristiche funzionali:• contro gradiente diconcentrazione• elevata selettività• ATP-dipendente• inibito da veleni metabolici e da agenti specifici (ouabaina)• elettrogenico• parametri cinetici
(KmNa = 0.2 mM; KmK = 0.05 mM)
Ouabaina
Na+/K+ATPasiTRASPORTO ATTIVO PRIMARIO P-Type ATPasi
E’ un processo ATP-dipendente, di conseguenza è soggetto all’inibizione da parte di v eleni delmetabolismo che limitano la disponibilità di ATP (disaccoppianti respirazione mitocondriale, etc..) oda agenti specif ici, come il glicoside cardiaco ouabaina. Il sito di legame per l’ouabaina(strofantina G), un glicoside vegetale si trova nelle immediate vicinanze deisiti per il potassio. Il legame della ouabaina porta all’arresto della pompaquando questa passa dalla configurazione E2 a quella E1. Ciò comporta sial’inibizione del trasporto ionico che dell’idrolisi dell’ATP. L’inibizione èreversibile, l’affinità dell’inibitore per la pompa è variabile sia tra speciediverse che, nell’ambito della stessa specie, a livello di tessuti diversi.Inf ine la > parte dei trasporti attiv i primari traslocano cariche elettriche attrav erso la membranadeterminando una separazione di cariche tra i due lati della membrana; in questo modo la Na-K-ATP-asi incide direttamente sul potenziale elettrico esistente f isiologicamente a cav allo dellemembrane plasmatiche oltre a contribuirv i indirettamente creando la distribuzione asimmetrica delsodio e del potassio tra l’ambiente intra ed extra-cellulare. La costante di legame per il Na è di 0,2mM; la Km per il K è di 0,05 mM.

33
Ruolo della Na+/K+ATPasi
I gradienti del Na+ sono importanti per itrasporti attivi secondari di soluti organici
(glucosio, aminoacidi),Ca2+, Cl-, H+
Crea gradienti per Na+ e K+necessari per:
•Potenziali d’azione•Potenziali graduati
TRASPORTO ATTIVO PRIMARIO P-Type ATPasi

34
PMCA (Plasma MembraneCalcium ATPase): utilizzataper espellere all’esterno dellacellula il Ca2+ attraverso lamembrana cellulare
Ca2+ ATPasi: PMCA e SERCATrasporti attivi primari
Idrolisi di ATP accoppiata al movimento del Ca2+
SERCA (Smooth EndoplasmicReticulum Calcium ATPase):Importante per sequestrare ilCa2+ nel reticolosarcoplasmatico/endoplasmatico
TRASPORTO ATTIVO PRIMARIO P-Type ATPasi
La pompa Ca2+ ATPasi è una proteina integrale di membrana (PM 100 kDa)con 10 domini transmembrana. Ha le caratteristiche di un uniporto etrasporta fuori dalla cellula 2 ioni Ca per ogni molecola di ATP utilizzata inogni ciclo. È elettrogenica ed ha l’importante ruolo di mantenere bassa laconcentrazione del Ca intracellulare che è una condizione v itale per lasoprav v iv enza e la funzionalità cellulare. La pompa del Ca è presente nonsolo a liv ello del plasmalemma ma è attiv a anche sulla membrana delreticolo endoplasmatico e sarcoplasmatico e nei mitocondri e aiuta amantenere bassi i liv elli di Ca citoplasmatico trasportabto lo ione all’internodi questi organuli contro un forte gradiente di concentrazione.Le due isoforme hanno div erse caratteristiche molecolari. La PMCA ha unamaggiore affinità per il Ca ed è praticamente sempre in funzione permantenere a liv ello basale la concentrazione di ca intracellulare. La sercaentra in funzione solo quando la concentrazione di Ca aumenta di un ordinedi grandezza. Non si conoscono ancora inibitori specifici per la PMCAmentre la SERCA è inibita dalla tapsigargina.

35
Ca2+ ATPasi: PMCATRASPORTO ATTIVO PRIMARIO P-Type ATPasi
Ca2+
SERCA
PMCA
reticoloendoplasmico
Pompa del calcio della membranaplasmatica (PMCA) Ne esistono diverseisoforme e sono attivate dallacalmodulina; ha il compito di pompareuno ione calcio dal citosol all’esterno.
PMCA 1: ubiquitaria, ha la funzionedi mantenere costante laconcentrazione intracellulare dicalcio. Presenta un sito per lafosforilazione da parte di protein-chinasi A e C (PKA e PKC) (che servead aumentare la velocità della pompa).PMCA 2: neuroni.PMCA 3: muscolo scheletrico ecervello.PMCA 4: in fase di studio
10-7 M
10-3 M
10-3 M
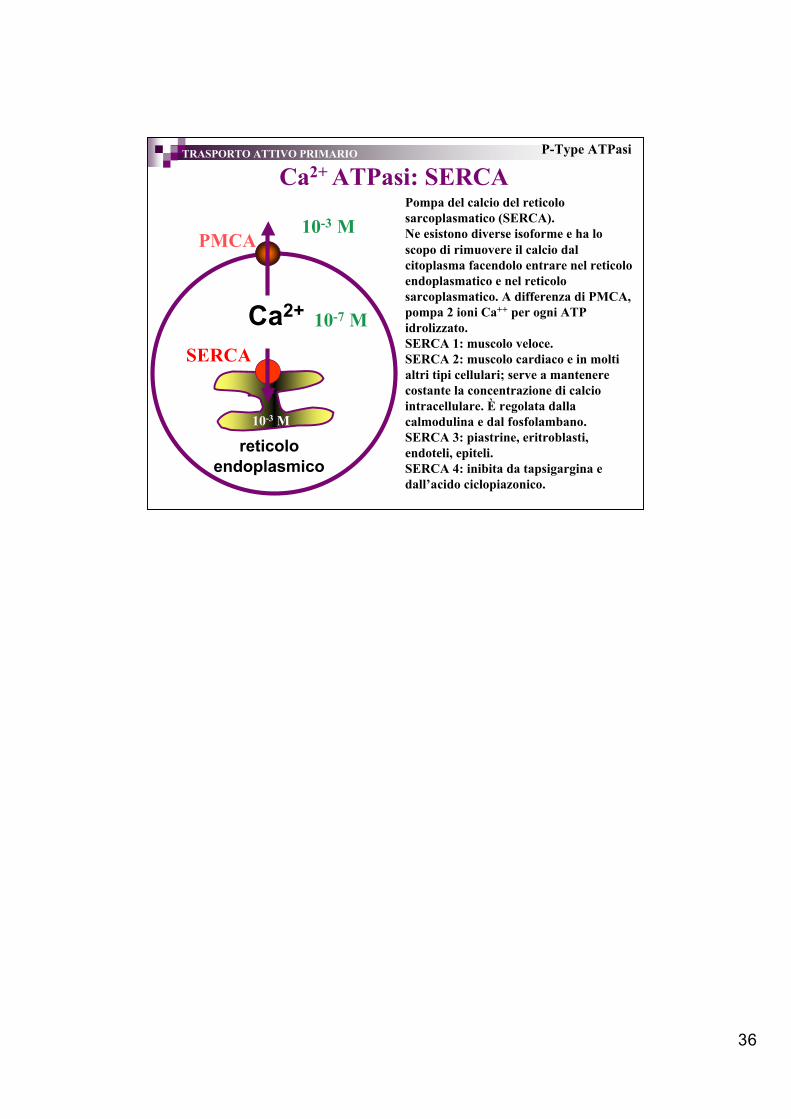
36
Ca2+ ATPasi: SERCATRASPORTO ATTIVO PRIMARIO P-Type ATPasi
Ca2+
PMCA
reticoloendoplasmico
Pompa del calcio del reticolosarcoplasmatico (SERCA).Ne esistono diverse isoforme e ha loscopo di rimuovere il calcio dalcitoplasma facendolo entrare nel reticoloendoplasmatico e nel reticolosarcoplasmatico. A differenza di PMCA,pompa 2 ioni Ca++ per ogni ATPidrolizzato.SERCA 1: muscolo veloce.SERCA 2: muscolo cardiaco e in moltialtri tipi cellulari; serve a mantenerecostante la concentrazione di calciointracellulare. È regolata dallacalmodulina e dal fosfolambano.SERCA 3: piastrine, eritroblasti,endoteli, epiteli.SERCA 4: inibita da tapsigargina edall’acido ciclopiazonico.
10-7 M
10-3 M
10-3 M
SERCA

37
H+/K+ ATPasiTRASPORTO ATTIVO PRIMARIO P-Type ATPasi
Strutturalmente molto simile allaNa+/K+ ATPasi che trasporta 4 H+
fuori dalla cellula e 4 ioni K+
all’interno della cellula.
Si trova maggiormente sulle celluleparietali gastriche ma è ancheriscontrabile nel dotto collettore esulla mucosa del colon responsabiledella secrezione acida nellostomaco.
La sua importanza è anche legata allarecente scoperta che derivatidell’omeprazolo si legano alla subunità αdella pompa e ne bloccano l’attività. Questifarmaci sono attualmente ampiamenteutilizzati come antiulcerosi e per la cura didiverse disfunzioni gastriche.
La pompa H+/K+ ATPasi o pompa protonica è un trasportatore strutturalmente moltosimile alla Na+/K+ ATPasi che trasporta 4 H+ fuori dalla cellula e 4 ioni K+ all’internodella cellula con stechiometria di 1:1 (quindi non è elettrogenica). In questo modopermette l’acidificazione del liquido extracellulare pur essendo bassi i liv elli di H+citoplasmatici (pH 7,4, [H+] = 4*10-8 M).La pompa è localizzata sulla parete delle cellule ossintiche (o parietali) delleghiandole gastriche e sv olge un ruolo centrale nella produzione dell’HCl del succogastrico. Permette la produzione di acido a concentrazioni massime di 100 mM(pH=1) ed è quindi in grado di mantenere elev atissimi gradienti di H+ tra l’interno el’esterno della cellula. La sua importanza è anche legata alla recente scoperta chederiv ati dell’omeprazolo si legano alla subunità α della pompa e ne bloccanol’attiv ità. Questi farmaci sono attualmente ampiamente utilizzati come antiulcerosi eper la cura di div erse disfunzioni gastriche.Il lansoprazolo, come tutti gli inibitori di pompa, inibisce questo processo conconseguente immediato blocco della produzione di acido cloridrico,indipendentemente dallo stimolo che l'ha prodotto. Nel lume gastrico il pH v ienemantenuto forzatamente al di di sopra di 3-4.La H+-K+-ATPasi ha una struttura molto simile alla pompa sodio-potassio: anche quisi riconoscono due subunità α e due β con la α che è responsabile della funzionalitàdella pompa protonica contenendo i siti di legame degli ioni, di fosforilazione e diattiv ità ATPasica.

38
CFTRTRASPORTO ATTIVO PRIMARIO TRASPORTATORI ABC
Si tratta di un canale del Clregolato, che risultaalterato nella fibrosi cistica.La funzione di questocanale è quella ditrasportare il cloroattraverso le membranecellulari a livello dellamembrana apicale dellecellule epiteliali delle vieaeree, del pancreas,dell’intestino, delleghiandole sudoripare, delleghiandole salivari e dei vasideferenti.

39
Spessissimo utilizzano ilgradiente elettrochimico delNa+ come fonte energetica.
Cotrasporti econtrotrasporti
Il Substrato si muovecontro il suo gradiente
Il Na+ si muove secondogradiente
Sono definiti attivi perché avvengonocontro gradiente di concentrazione
Sono definiti secondari perché non utilizzanodirettamente energia metabolica
Il trasporto attivo secondario sfrutta l’energia c inetica di uno ione che si muovepassivamente secondo il suo gradiente di concentrazione per trasportare una molecolao un altro ione contro gradiente.Le molecole o gli ioni cotrasportati possono muoversi nella stessa direzione (simporto)oppure in direzione opposta (antiporto).I più comuni sistemi di trasporto attivo secondario sono mediati da proteinetransmembrana che sfruttano il gradiente di concentrazione del Na+ creato dalla Na+-K+ ATPasi. In tal caso, l ’alta concentrazione extracellulare di Na+ favorisce il legamedello ione sul lato esterno del trasportatore, che a sua volta è facil itato a legare unamolecola o un altro ione e a trasportarli dal lato opposto a quello in cui si trovano,senza ulteriore spesa di energia.
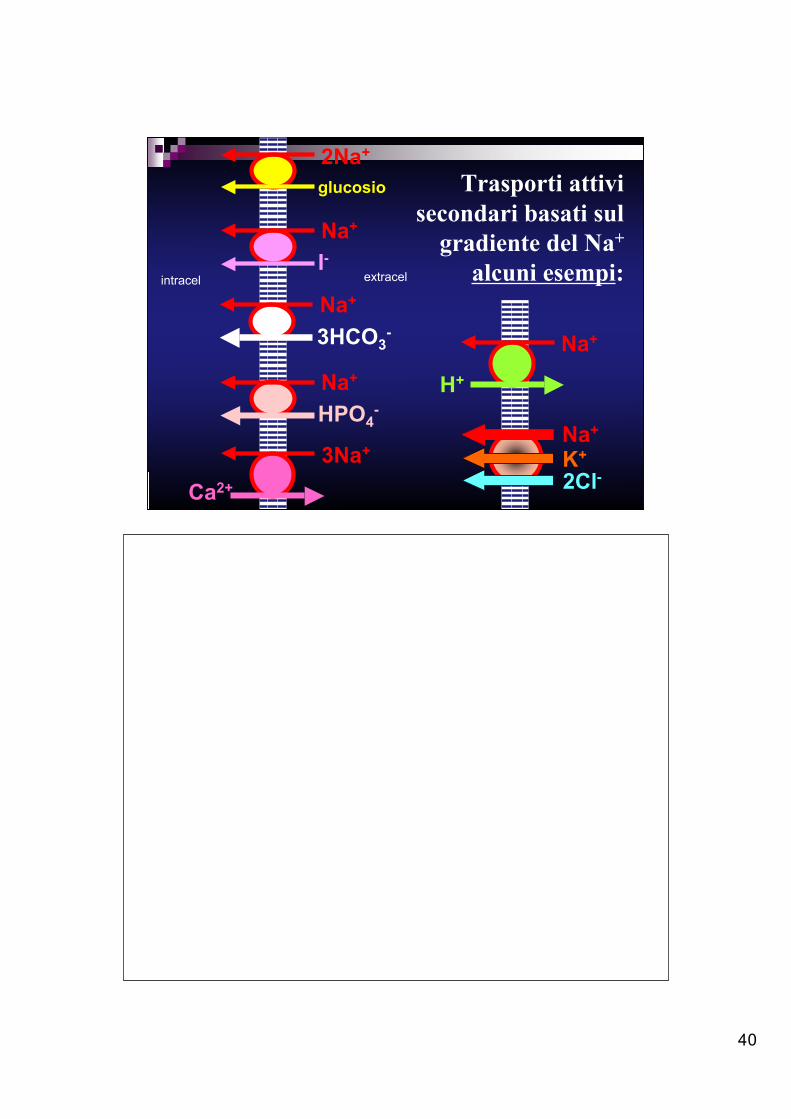
40
Trasporti attivisecondari basati sul
gradiente del Na+
alcuni esempi:
2Na+
glucosio
Na+
I-
Na+
3HCO3- Na+
H+Na+
HPO4-
3Na+
Ca2+
Na+
K+
2Cl-
intracel extracel

41
Antiporto Na+/Ca2+TRASPORTO ATTIVO SECONDARIO
L’antiporto Na+/Ca2+ ha unruolo chiave nel controllo delCa2+ intracellulare in tutte lecellule, dove è costantementeattivato.
È di particolare importanza nellecellule cardiache quando laconcentrazione di Ca2+, cheaumenta durante la contrazionesistolica, deve esserevelocemente riportata al suolivello normale per permettere ilrilasciamento ventricolare
L’antiporto Na+/Ca2+ ha un ruolo chiave nel controllo del Caintracellulare in tutte le cellule, dove è costantemente attivato.Trasporta 3 ioni Na dentro la cellula e 1 ione Ca fuori ed è quindielettrogenico. È di particolare importanza nelle cellule cardiachequando la concentrazione di Ca, che aumenta durante la contrazionesistolica, deve essere velocemente riportata al suo livello normaleper permettere il rilasciamento ventricolare

42
Antiporto Na+/Ca2+ e insufficienza cardiacaTRASPORTO ATTIVO SECONDARIO
Un paziente con insuffic ienza cardiaca possiede un cuore ipodinamicocaratterizzato da una bassa gittata cardiaca (volume di sangue espulso daiventricoli al minuto) insuffic iente a sostenere le richieste metaboliche del suocorpo. Il metodo più efficace per migliorare l ’attività cardiaca consistenell ’aumentare la contratti l i tà miocardica che è marcatamente Ca2+dipendente. Un aumento del Ca c itoplasmatico produce un’aumentatacontratti l i tà cardiaca che a sua volta produce un aumento del volume sistolico(volume di sangue espulso durante un singolo battito cardiaco) e della gittatacardiaca.I glicosidi cardiaci come la digitossina, la digossina e l ’ouabaina, sono usati damoltissimi anni per la cura dell ’insuffic ienza cardiaca. Questi farmaci si leganoselettivamente e inibiscono la Na+/K+-ATPasi della membrana delle celluledel miocardio. In tal modo il Na+ si accumula in cellula riducendonotevolmente l’attività dello scambiatore Na+/Ca2. Come risultato finale i ll ivello c itoplasmatico di Ca aumenta e aumenta di conseguenza la contratti l i tàmiocardica, i l volume sistolico e la gittata cardiaca.

43
L’antiporto Na+/H+ (NHE1 - NHE6)TRASPORTO ATTIVO SECONDARIO
Svolge un ruolo determinantenel controllo del pHintracellulare di tutte le cellule
L’antiporto Na+/H+ èparticolarmente importante alivello renale. La sualocalizzazione sull’orletto aspazzola delle cellule epitelialedel tubulo contorto prossimalepermette, infatti, di arricchire illiquido del lume tubulare di ioniH+ e di acidificare le urine.
L’antiporto Na+/H+ è svolge un ruolo determinante nel controllo delpH intracellulare di tutte le cellule in quanto trasporta ioni H+ fuoridalla cellula e ioni Na dentro in rapporto 1:1. L’antiporto Na+/H+ èparticolarmente importante a livello renale. La sua localizzazionesull’orletto a spazzola delle cellule epiteliale del tubulo contortoprossimale permette, infatti, di arricchire il liquido del lumetubulare di ioni H+ e di acidificare le urine.
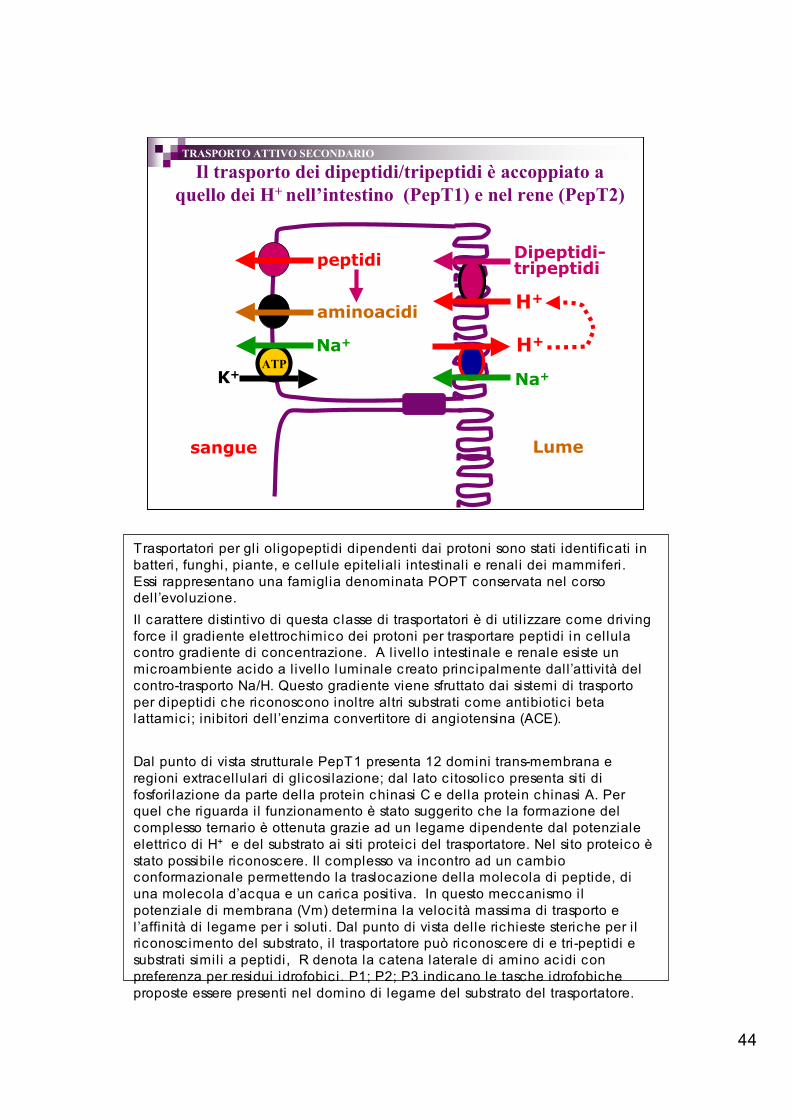
44
Il trasporto dei dipeptidi/tripeptidi è accoppiato aquello dei H+ nell’intestino (PepT1) e nel rene (PepT2)
sangue Lume
H+
Na+ATP
Na+
K+
peptidi
H+
Dipeptidi-tripeptidi
aminoacidi
TRASPORTO ATTIVO SECONDARIO
Trasportatori per gli oligopeptidi dipendenti dai protoni sono stati identificati inbatteri, funghi, piante, e cellule epiteliali intestinali e renali dei mammiferi.Essi rappresentano una famiglia denominata POPT conservata nel corsodell ’evoluzione.Il carattere distintivo di questa c lasse di trasportatori è di uti l izzare come drivingforce il gradiente elettrochimico dei protoni per trasportare peptidi in cellulacontro gradiente di concentrazione. A l ivello intestinale e renale esiste unmicroambiente acido a l ivello luminale c reato principalmente dall’attività delcontro-trasporto Na/H. Questo gradiente viene sfruttato dai sistemi di trasportoper dipeptidi che riconoscono inoltre altri substrati come antibiotic i betalattamici; inibitori dell ’enzima convertitore di angiotensina (ACE).
Dal punto di vista strutturale PepT1 presenta 12 domini trans-membrana eregioni extracellulari di glicosilazione; dal lato c itosolico presenta siti difosforilazione da parte della protein chinasi C e della protein chinasi A. Perquel che riguarda il funzionamento è stato suggerito che la formazione delcomplesso ternario è ottenuta grazie ad un legame dipendente dal potenzialeelettrico di H+ e del substrato ai siti proteic i del trasportatore. Nel sito proteico èstato possibile riconoscere. Il complesso va incontro ad un cambioconformazionale permettendo la traslocazione della molecola di peptide, diuna molecola d’acqua e un carica positiva. In questo meccanismo ilpotenziale di membrana (Vm) determina la velocità massima di trasporto el’affinità di legame per i soluti. Dal punto di vista delle richieste steriche per i lriconoscimento del substrato, i l trasportatore può riconoscere di e tri-peptidi esubstrati simili a peptidi, R denota la catena laterale di amino acidi conpreferenza per residui idrofobic i. P1; P2; P3 indicano le tasche idrofobicheproposte essere presenti nel domino di legame del substrato del trasportatore.
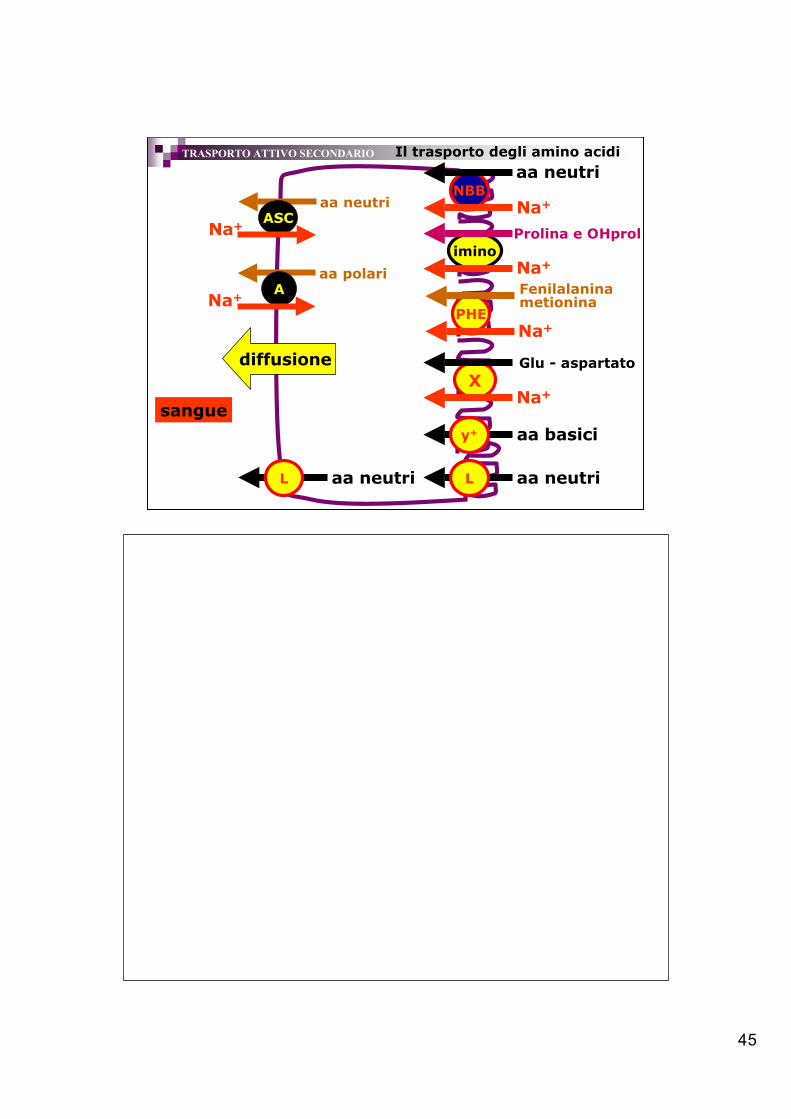
45
iminoNa+
Prolina e OHprol
PHENa+
Fenilalanina metionina
XNa+
Glu - aspartato
NBBNa+
aa neutri
aa basiciy+
L aa neutri
diffusione
L aa neutri
aa neutriASC
Na+
aa polariA
Na+
Il trasporto degli amino acidi
sangue
TRASPORTO ATTIVO SECONDARIO

46
Il cotrasporto Na+-D-glucosiodetermina l’assorbimentointestinale ed ilriassorbimento renale delglucosio
il cotrasporto Na+-D-glucosioTRASPORTO ATTIVO SECONDARIO
Il cotrasporto Na-D-glucosio è un esempio classico di trasporto attiv o secondario. La sua f unzioneprincipale è quella di mediare sia l’assorbimento di glucosio a liv ello intestinale che ilriassorbimento dello zucchero a liv ello renale.

47
il cotrasporto Na+-D-glucosioTRASPORTO ATTIVO SECONDARIO
Il meccanismo di traslocazione è simile a quelli riportati sino ad ora. Anche in questo caso si ha un oscillazione tra duestati conformazionali E1 e E2 con i siti rivolti alternativamente verso l’ambiente extra ed intra-cellulare. In questo casotuttavia l’elemento aggiuntivo è rappresentato dal legame dello ione sodio al trasportatore che induce un cambioconformazionale nella tasca di legame per lo zucchero, aumentandone l’affinità nei confronti del substrato. Una volta che ilD-glucosio si è legato, si ha una traslocazione dei due substrati in direzione dell’ambiente intracellulare, dove il sodio sistacca dal suo sito di legame poiché le sue concentrazioni intracellulari sono basse, il distacco del sodio determina unariduzione dell’affinità per il D-glucosio che si stacca dalla sua tasca di legame, pur essendo le sue concentrazionicitosoliche abbastanza elevate.

48
1:1 3 mMTubulo contortoprossimale (S1)
SGLT-2
2:10.35 mMIntestino tenueTubulo contorto
prossimale (S2 e S3)
SGLT-1
stechiometriaKm D-glucoselocalizzazione
PKC
Struttura secondaria del SGLT-1
PKC PKA PKC
Il cotrasporto Na+-D-glucosioTRASPORTO ATTIVO SECONDARIO
Al momento sono conosciute due isof orme SGLT-1 e SGLT-2 del trasportatore Na-dipendente di D-glucosio.Il primo è presente sia sulle membrane apicali dell’intestino tenue che nella seconda parte deltubulo contorto prossimale, è caratterizzato da una Km di circa 0.1 mM per il D-glucosio, unastechiometria di cotrasporto Na-D-glucosio di 2 ioni sodio per molecola di D-glucosio trasportata; èun trasporto elettrogenico.SGLT-2 è stato riscontrato nelle prime porzioni del tubulo contorto prossimale e si distinguecineticamente da SGLT-1 in quanto ha un’af f inità molto inf eriore ma una v elocità di trasporto moltopiù elev ata rispetto a SGLT-1.Questa distribuzione regionale delle due isof orme di SGLT nel tubulo contorto prossimale assicura ilriassorbimento totale del D-glucosio. Inf atti nelle regioni adiacenti alla capsula di bowman dov epredomina SGLT-2 questo trasporto capta ef f icientemente il D-glucosio ultraf iltrato ad altaconcentrazione, grazie alla sua Km che è v icina alle concentrazioni di glucosio e alla sua altav elocità di trasporto. Allorquando le concentrazioni di D-glucosio nell’ultraf iltrato si riducono, manmano che questo scorre nel tubulo prossimale, questo sistema non sarebbe più ef f iciente acontinuare il processo di riassorbimento a causa della sua bassa af f inità per il D-glucosio e diconseguenza interv iene l’isof orma SGLT-1 localizzata più a v alle, che opera ef f icientemente abasse concentrazioni di D-glucosio a causa della sua elev ata af f inità per lo zucchero.SGLT1 è costituito da 664 residui aacidici. La struttura secondaria del trasportatore si articola in 14alf a-eliche trans-membrana (al contrario dei trasporti f acilitati del glucosio che possiedono solo 12eliche) con dei domini citosolici. La regione del trasportatore che lega e trasloca sodio, identif icatitramite studi con trasportatori chimerici, comprendono la regione N-terminale sino a loopextracellulare che connette i domini trans-membrana 4 e 5; le regioni di legame e di traslocazioneincludono domini TM da 10 a 13. SGLT1, al contrario di altre proteine carrier manca di un grandedominio citosolico, il chè suggerisce l’esistenza di addizionali sub-unità regolatorie. Recentemente èstato proposto da Rehiardt ed altri che una proteina denominata RS1, ancorata alle membranaplasmatiche apicali aumenti la v elocità massima di uptake di D-glucosio e v ada a modularel’attiv ità di altri sistemi di trasporto.Un altro tipo di regolazione sembra essere ef f ettuata dalla protein chinasi C e protein kinasi A datoche domini citosolici del trasportatore contengono regioni suscettibili a f osf orilazione da parte diqueste protein chinasi. Di conseguenza è un sistema di trasporto sottoposto sia a regolazionetramite AMPc, diacil-glicerolo e calcio. Lav ori recenti suggeriscono che PKA e PKC aumentinol’attiv ità del cotrasporto mediante una maggiore espressione di questo a liv ello delle membraneplasmatiche apicali. Il meccanismo di questa regolazione è mediato da endocitosi o esocitosi div escicole situate sotto la membrana pronte ad essere incorporate nel momento della stimolazioneda parte ormonale.SGLT-1 appartiene ad una f amiglia cui appartengono più di 30 classi di trasportatori. Questicondiv idono caratteristiche strutturali e f unzionali omologhe. Posseggono 13 o più eliche trans-membrana, utilizzano il gradiente elettrochimico sia del sodio che dei protoni per trasportate unav arietà di substrati che includono zuccheri, aacidi, anioni organici, urea, v itamine, etc.. Dov rebberocondiv idere un comune meccanismo di trasporto sebbene il substrato cotrasportato possa v ariareconsiderev olmente.
49
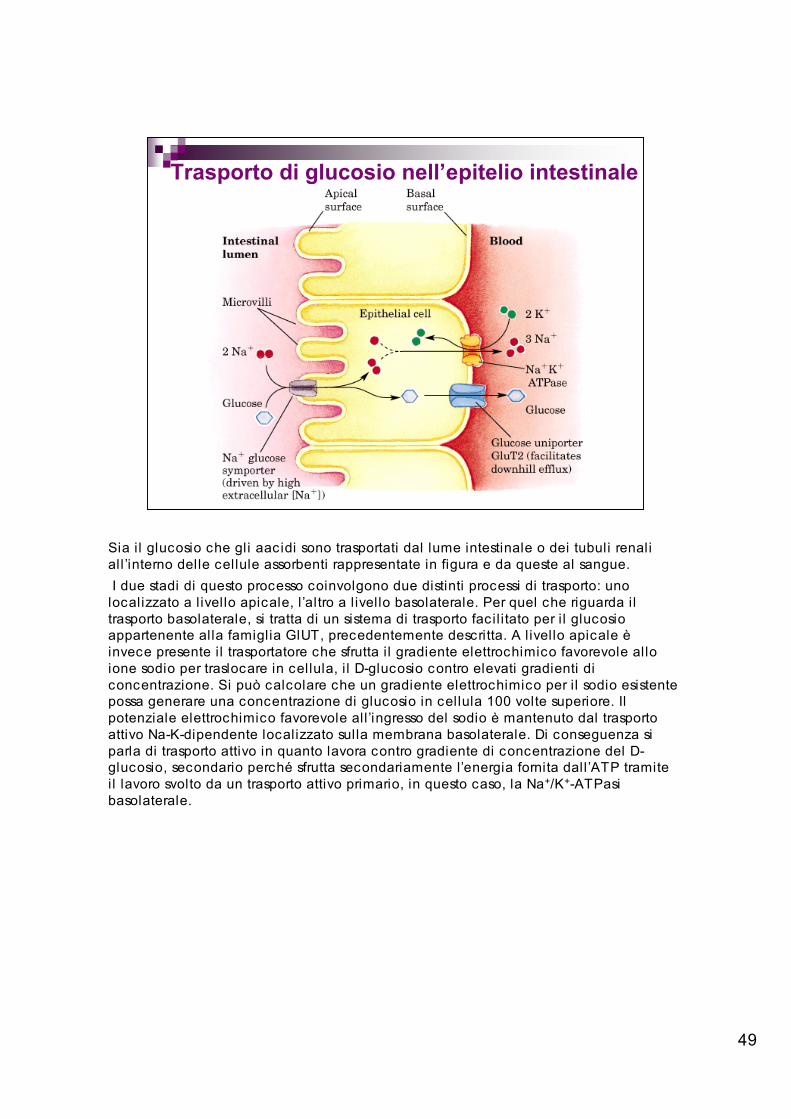
Trasporto di glucosio nell’epitelio intestinale
Sia i l glucosio che gli aacidi sono trasportati dal lume intestinale o dei tubuli renaliall ’interno delle cellule assorbenti rappresentate in figura e da queste al sangue. I due stadi di questo processo coinvolgono due distinti processi di trasporto: unolocalizzato a l ivello apicale, l’altro a l ivello basolaterale. Per quel che riguarda iltrasporto basolaterale, si tratta di un sistema di trasporto facil itato per i l glucosioappartenente alla famiglia GlUT, precedentemente descritta. A livello apicale èinvece presente i l trasportatore che sfrutta i l gradiente elettrochimico favorevole alloione sodio per traslocare in cellula, i l D-glucosio contro elevati gradienti diconcentrazione. Si può calcolare che un gradiente elettrochimico per i l sodio esistentepossa generare una concentrazione di glucosio in cellula 100 volte superiore. Ilpotenziale elettrochimico favorevole all ’ingresso del sodio è mantenuto dal trasportoattivo Na-K-dipendente localizzato sulla membrana basolaterale. Di conseguenza siparla di trasporto attivo in quanto lavora contro gradiente di concentrazione del D-glucosio, secondario perché sfrutta secondariamente l’energia fornita dall ’ATP tramiteil lavoro svolto da un trasporto attivo primario, in questo caso, la Na+/K+-ATPasibasolaterale.

50
Scambiatore Cl-/HCO3-
DIFFUSIONE FACILITATA
Cl-
HCO3-
L’antiporto Cl-/HCO3- (proteina della
banda 3) è importante nel controllodell’equilibrio acido-base a livellocellulare.Trasporta ioni HCO3
- fuori dallacellula e ioni Cl- dentro in unrapporto 1:1.
Oltre al trasporto attivo secondario Na-dipendente esistono anche svariatitrasportatori che non sono Na dipendenti. Tra questi, i l più importante per i l suoruolo nel controllo dell ’equil ibrio acido-base a l ivello cellulare è l ’antiporto Cl-/HCO3
- che trasporta ioni bicarbonato fuori dalla cellula e ioni Cl- dentro in unrapporto 1:1.La fuoriuscita di bicarbonato dalla cellula av v iene secondo gradiente ed èaccoppiata all’ingresso di cloro, che può av v enire anche controgradiente.

51
Scambiatore Cl-/HCO3-
DIFFUSIONE FACILITATA
Lo scambio anionico Cl-/HCO3- o proteina della banda 3 è
presente in notevole quantità nella membrana eritrocitaria.
A livello tissutale, si ha unaelevata tensione parziale di
CO2 e questo determinal’ingresso del gas
nell’eritrocita. La CO2 vieneidratata ad opera dellaanidrasi carbonica con
formazione di HCO3- e H+. Il
bicarbonato viene liberatoverso l’esterno del globulo
rosso, ad opera della banda 3.
Lo scambiatore sv olge un ruolo fondamentale a liv ello degli eritrociti dov econtribuisce all’arricchimento di bicarbonato nel plasma e più in generale altrasporto della CO2 e alla regolazione della respirazione.

52
DIFFUSIONE FACILITATA
A livello dei polmoni c’èuna bassa tensioneparziale di CO2 e si
verifica l’effetto opposto,cioè il flusso di
bicarbonato è direttoall’interno mentre il Cl-
esce dall’eritrocita.
Scambiatore Cl-/HCO3-
Lo scambio anionico Cl-/HCO3- o proteina della banda 3 è
presente in notevole quantità nella membrana eritrocitaria.

53
DIFFUSIONE FACILITATA
Scambiatore Cl-/HCO3-
Il trasportatore Cl/HCO3 è particolarmente importante anche in altri distrettidove svolge importanti funzioni come:•Il riassorbimento di bicarbonato a livello della membrana basolaterale deldigiuno e del tubulo contorto prossimale e distale del rene•L’ingresso di Cl esterno nelle cellule ossintiche dell’epitelio gastrico, cherappresenta la riserva di ioni Cl- utilizzati per la produzione di HCl del succogastrico.•La produzione di bicarbonato del succo pancreatico che serve permantenere il pH del succo pancreatico a valori basici (pH 8-9) che aiuta acontrobilanciare il basso pH del chimo nel lume duodenale e a ottenere pHottimali per il funzionamento degli enzimi pancreatici.
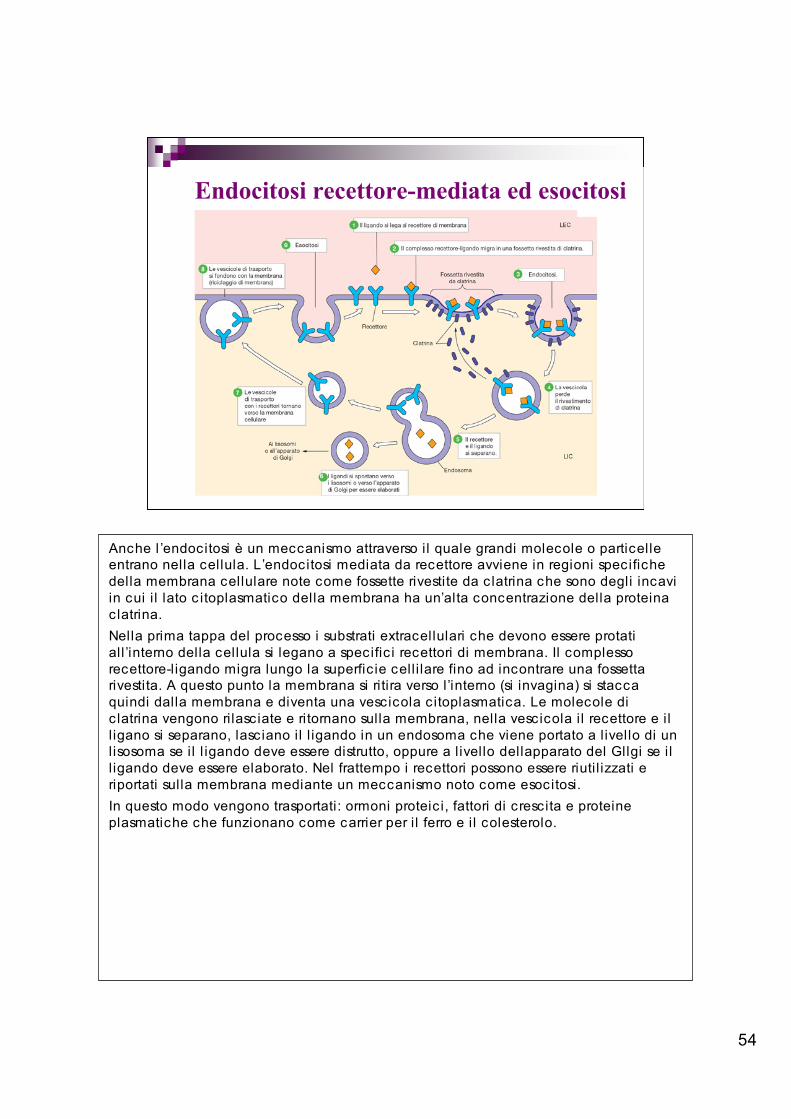
54
Endocitosi recettore-mediata ed esocitosi
Anche l ’endocitosi è un meccanismo attraverso il quale grandi molecole o particelleentrano nella cellula. L’endocitosi mediata da recettore avviene in regioni specifichedella membrana cellulare note come fossette rivestite da c latrina che sono degli incaviin cui i l lato c itoplasmatico della membrana ha un’alta concentrazione della proteinaclatrina.Nella prima tappa del processo i substrati extracellulari che devono essere protatiall ’interno della cellula si legano a specific i recettori di membrana. Il complessorecettore-ligando migra lungo la superfic ie cell i lare fino ad incontrare una fossettarivestita. A questo punto la membrana si ritira verso l’interno (si invagina) si staccaquindi dalla membrana e diventa una vescicola c itoplasmatica. Le molecole dic latrina vengono ri lasciate e ritornano sulla membrana, nella vescicola i l recettore e i ll igano si separano, lasciano il l igando in un endosoma che viene portato a l ivello di unlisosoma se il l igando deve essere distrutto, oppure a l ivello dellapparato del Gllgi se i ll igando deve essere elaborato. Nel frattempo i recettori possono essere riuti l izzati eriportati sulla membrana mediante un meccanismo noto come esocitosi.In questo modo vengono trasportati: ormoni proteic i, fattori di crescita e proteineplasmatiche che funzionano come carrier per i l ferro e i l colesterolo.





























