Embed Size (px)
Citation preview

Evaluation ofMYC and Chromosome 8 Copy Numberin Breast Carcinoma by Interphase Cytogenetics
Daniel W. Visscher,* Tracie Wallis, Solomon Awussah, Anwar Mohamed, and John D. Crissman
Department of Pathology, Harper Hospital and Karmanos Cancer Institute, Detroit, Michigan and Wayne State University School ofMedicine, Detroit, Michigan
We used fluorescence in situ hybridization (FISH) to determine MYC and chromosome 8 copy number on whole nuclearimprint preparations of 24 breast carcinomas, seven benign breast samples, and two phyllodes tumors. None of the benigntissues and neither of the phyllodes tumors demonstrated an increased copy number for MYC or chromosome 8, which wasdefined as greater than two signals in .10% of nuclei. In contrast, 22 of 24 carcinomas demonstrated an increased MYC copynumber. The modal numbers ofMYC copies/nucleus were 0–2 in seven cases (29%), 3–5 in seven cases (29%), 6–9 in five cases(21%), and .9 in five cases (21%). An increased chromosome 8 copy number was observed in 21 of 22 carcinomas with MYCgain, and the modal number of signals/nucleus was either identical to (n5 14; 64%) or less than (n5 8; 36%) the number ofMYCcopies. The number ofMYC copies correlated with cellular DNA content, as determined by using flow cytometry. In peridiploidtumors (DNA index 0.9–1.2; n 5 7), the MYC copy numbers/nucleus were 0–2 in five cases and 3–5 in two cases. In contrast,the modal MYC copy numbers/nucleus among the 11 hyperdiploid tumors (DNA index 1.3–1.9) were 0–2 in one case, 3–5 infour cases, 6–9 in five cases, and .9 in one case. All three tetraploid/hypertetraploid carcinomas exhibited .9 MYCcopies/nucleus. We conclude that an increased MYC copy number, as detected by using interphase cytogenetics, is extremelyfrequent in human breast carcinomas. However, in most cases, MYC gene duplication is probably secondary to polysomy ofchromosome 8 and/or genomic endoreduplication (i.e., DNA aneuploidy). Genes Chromosom Cancer 18:1–7, 1997.
r 1997 Wiley-Liss, Inc.
INTRODUCTION
TheMYC gene, which is located on chromosomeband 8q24, encodes for a nuclear transcriptionfactor that regulates cell proliferation and differen-tiation. Amplification, or an increased gene copynumber for MYC, has been reported in 5–50% ofhuman breast carcinomas (Escot et al., 1986; Bonillaet al., 1988; Meyers et al., 1990). Observed associa-tions betweenMYC amplification and adverse prog-nostic indicators suggest that abnormal expressionof MYC may facilitate progression of breast neopla-sia (Tsuda et al., 1991; Berns et al., 1992; Kreipe etal., 1993; Pertschuk et al., 1993).Most published studies that have evaluatedMYC
amplification have employed Southern hybridiza-tion or differential polymerase chain reaction (Bo-nilla et al., 1988; Kreipe et al., 1993; Fontana et al.,1994). The sensitivity of these techniques is lim-ited by the mixing of DNA from neoplastic cellswith DNA of host-derived stromal or inflammatorycells, which infiltrate breast carcinomas sparsely toextensively, depending on the individual case. Byanalogy, it is also difficult to account for cellularheterogeneity of the gene copy number whenmolecular techniques are used (Pavelic et al., 1992).Finally, most gene amplification studies have notassessed the degree to which aneuploidy, either
chromosomal or genomic, contributes to an increasein gene copy number (i.e., as opposed to intra- orextrachromosomal duplication of DNA segmentscontainingMYC).When DNA sequence probes that are specific for
sequences of MYC and the chromosome 8 centro-meric region are used, it is possible to enumerateand compare gene and chromosome copy numberwithin interphase nuclei (Hopman et al., 1994).This technique—interphase cytogenetics—also pro-vides a means for performing a relatively selectiveanalysis of neoplastic populations. Thus, gene copynumber may be evaluated directly in the context ofaneuploidy. In this study, we employed interphasecytogenetics to studyMYC and chromosome 8 copynumber within intact nuclei of benign and malig-nant breast tissues. Our objective was to obtain amore refined determination of the degree andintratumoral distribution of MYC amplification inbreast carcinomas as well as the extent to whichgene copy number reflects chromosomal or ge-nomic endoreduplication.
*Correspondence to: Daniel W. Visscher, MD, Department ofPathology, Harper Hospital, 3990 John Road, Detroit, MI 48201.Received 29 January 1996; Accepted 20 May 1996.
GENES, CHROMOSOMES & CANCER 18:1–7 (1997)
3333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333 RESEARCH ARTICLES 33333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333333
r 1997 Wiley-Liss, Inc.

MATERIALS AND METHODS
Samples were collected prospectively betweenJuly and November, 1995, from the Department ofSurgical Pathology, Harper Hospital, Detroit, Michi-gan. Fresh, unfixed breast biopsy specimens wereserially sectioned, and whole-cell imprint prepara-tions were made by gentle pressing of silanizedslides (n 5 5) to areas of grossly visible neoplastictissue. The slides were fixed immediately by sub-mersion in methanol/acetic acid. One of the slideswas stained with hematoxylin and eosin to assessfor adequate cellularity and representation of tumorcell populations (at least 80% of total cells). Wemade a formalin-fixed, paraffin- embedded sectionfrom the tissue that was utilized for imprints inorder to confirm the histologic type and differentia-tion of the neoplasm. Twenty-six malignant breastlesions were accrued, including nineteen invasiveductal carcinomas, four invasive lobular carcinomas,one ductal carcinoma in situ, and two phyllodestumors (one high grade and one low grade).Flow-cytometric DNA analysis is performed rou-
tinely on breast tumors submitted to our depart-ment. Our methods have been published else-where (Visscher et al., 1990). DNA indices on allbut two of the invasive epithelial neoplasms wereavailable. Synthesis-phase fractions (SPFs) wereavailable in 19 of the 24 breast carcinomas; in fivecases, calculations were precluded by peak overlap.Seven benign breast cases were collected andstudied as controls, including one case with nonpro-liferative fibrocystic change, three cases with prolif-erative fibrocystic change, one benign granular celltumor, and two cases with lobular benign carcinomain situ (LCIS). Neoplastic cells in the LCIS caseswere extremely focal within the biopsy sample.Therefore, interphase cytogenetic analyses wereactually performed on accompanying benign, fibro-cystic tissue from these cases.Genomic sequence MYC and chromosome 8
a-satellite probes and most reagents necessary forhybridization were purchased fromOncor (Gaithers-burg, MD). TheMYC probe was digoxigenin conju-gated, and the chromosome 8 centromeric probewas biotin labeled. Hybridizations were performedon separate touch preparations essentially accord-ing to the instructions that accompany the probes,with minor modifications designed to optimize thesignal strength of each probe. Briefly, after slidedehydration with graded ethanol washes, denatur-ation was performed by incubation in 70% for-mamide 2 3 standard saline citrate solution (SSC)at 70°C. Hybridization with labeled probes was
carried out in a humidified chamber (4–16 hourincubations at 43°C). After washes in SSC and then1 3 phosphate-buffer detergent solutions, pH 8.0(Oncor), rhodamine antidigoxigenin/FITC-avidinwas applied to slides (45 µl; 15 minutes at 37°C).The nuclei were counterstained with DAPI andthen examined directly by using a fluorescencemicroscope. Only signals in nonoverlapped, appar-ently intact nuclei (n 5 200) were counted. Nucleiapparently corresponding to spindle cells or inflam-matory cell populations were avoided.Individual nuclei were classified as having 0–2
signals, 3–5 signals, 6–9 signals, or .9 signals, andthe percentages of nuclei in each of these categorieswere calculated. Based on evaluation of benigncontrol samples, a case was considered abnormal(i.e., amplified or aneusomic) if signal gain (forMYC, for chromosome 8, or for both) was observedin .10% of the enumerated nuclei. AmplificationofMYC was arbitrarily designated as 88low grade’’ ifsignal gain (i.e., .2 signals) was observed in morethan 10% of nuclei but did not exceed 5 signals/nucleus in more than 10% of cells. ‘‘High-grade’’amplification meant that more than 10% of nucleishowed more than 5 signals/nucleus.
RESULTS
Hybridizations were successful on all seven be-nign control cases and on all 26 malignant neo-plasms. The hybridization efficiency was .90% ineach control sample (i.e., there was less than 10%signal loss). Neither amplification of MYC norchromosome 8 aneusomy was observed in any ofthe control cases. However, 22 of 26 (86%) malig-nant tumors exhibited increased MYC copy num-ber. This was low grade (i.e., 3–5 copies/nucleus) in11 cases (50%) and high grade (i.e., .5 copies/nucleus) in the remaining 11 cases (50%). Althoughchromosome 8 gain was frequent (see below), lossof chromosome 8 signal was not observed.
Benign Control Cases
None of the seven benign control cases exhibitedmore than 5% of nuclei with increased signal countsfor chromosome 8 (see Table 1). Only two speci-mens (cases 5 and 7) were observed to have anyaneusomy (5 and 2%, respectively). Five of theseven controls, however, had signal gains with theMYC probe, ranging from 1 to 8%. The specimenwith 8% abnormal nuclei was from a biopsy speci-men that contained focal lobular carcinoma in situ(case 6). None of the nuclei with MYC or chro-
2 VISSCHER ET AL.

mosome 8 gain contained more than 3–5 signals(Fig. 1).
TumorsWith Absent or Low-GradeMYCAmplification
Four of the malignant tumors (cases 1–4) showed10% or fewer nuclei with MYC signal gain (seeTable 2). Two of these four (cases 2 and 3) werestromal neoplasms (phyllodes tumors), and none ofthe four exhibited a significant signal gain for thechromosome 8 probe. Five additional tumors in thisgroup demonstrated MYC signal gain in 11–49% ofthe counted nuclei. In each of these cases, all of theabnormal nuclei contained 3–5 signals. Two of thefive (cases 8 and 9) also displayed a signal gain forchromosome 8; the degree was very similar to thatof theMYC probe.The remaining six tumors with low-grade MYC
amplification (Table 2, cases 10–15) exhibited amajority (i.e., at least 50%) of nuclei with 3–5signals. All six tumors demonstrated chromosome 8aneusomy, which, once again, was highly analogousto MYC with respect to the number of signals andthe proportion of nuclei involved. This group oftumors differed from the others with low-gradeMYC amplification, because it contained occasionalnuclei (1–10%) with .5 signals for MYC and/orchromosome 8.The neoplasms with low-grade MYC amplifica-
tion, in general, were not highly aneuploid. Flowcytometric DNA indices were ,1.3 in 6 of 11
tumors that were studied, and only 3 of 11 had aDNA index that exceeded 1.3. They also expresseda morphologically differentiated phenotype; only 3of 15 cases (9, 13, and 15) had grade III (i.e., poorlydifferentiated) histology.
TumorsWith High-GradeMYC Amplification
The 11 tumors that were classified as havinghigh-gradeMYC amplification were relatively homo-geneous (see Table 3). Only one tumor (case 1)exhibited a majority of nuclei with only 3–5 MYCsignals/nucleus; however, that tumor demonstratedthat 24% of nuclei had 6–9 signals. The modalnumbers ofMYC signals/nucleus among the remain-ing ten tumors were 6–9 in five cases and .9 in fivecases. Only case 1 failed to demonstrate an increasein chromosome 8 copy number. In the remainingtumors, the modal signal count for chromosome 8was similar to that for MYC in four tumors, was onecategory less increased in three tumors, and wastwo categories less increased in the remaining threetumors.High-grade MYC amplification was also well
correlated with DNA aneuploidy. Only case 1 wasDNA diploid (DNA index 5 1.0), and, interest-ingly, it exhibited normal chromosome 8 signalcounts despite the fact that 24% of nuclei showed6–9 signals for MYC. None of the remaining ninecases with DNA index determinations was peridip-loid, and three were tetraploid (n 5 1) or hypertet-raploid (n 5 2). Finally, as implied by the frequencyand degree of DNA aneuploidy in this group ofcases, tumors with high-grade MYC amplificationwere typically poorly differentiated, with grade IIIhistologic features (8 of 11).
DISCUSSION
Our results show that interphase cytogeneticmethods reveal a high frequency of increased MYCgene copy number in epithelial neoplasms of thebreast. In fact, only two of the 24 breast carcinomasfailed to demonstrate a MYC signal gain relative tothe benign control specimens. A variety of factors,however, may cause or may contribute to theobservedMYC signal gains. First, three or fourMYCgene copies may theoretically be present in nucleithat are in the process of synthesizing DNA inpreparation for cell division. Given the significantSPF of human breast carcinomas, it may be esti-mated conservatively that (low-grade) signal gainsin up to 25% of nuclei may actually be a reflectionof proliferative activity. This factor probably ac-counts for the signal gains observed in cases 5 and 6in Table 2. Observed MYC copy number, in fact,
TABLE 1. Chromosome 8 andMYC Signal Counts in BenignControl Cases
Percent of nuclei with
Case no. Diagnosis Probe 0–2 Signals 3–5 Signals
1 NPFCDa MYC 99 1chr 8b 100 0
2 PFCDc MYC 100 0chr 8 100 0
3 PFCD MYC 100 0chr 8 100 0
4 PFCD MYC 95 5chr 8 100 0
5 GCTd MYC 95 5chr 8 95 5
6 PFCDe MYC 92 8chr 8 100 0
7 PFCDe MYC 96 4chr 8 98 2
aNonproliferative fibrocystic change.bChromosome 8.cProliferative fibrocystic change.dGranular cell tumor.eObtained from a biopsy with focal lobular carcinoma in situ.
3MYC FISH IN BREAST CANCER

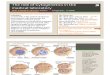
Figure 1. Photomicrographs of MYC and chromosome 8 hybridiza-tions in breast carcinomas (Elite Ektachrome, 400 ASA, Kodak). Top: Adiploid-range breast carcinoma showing normal hybridization patterns(two copies/nucleus) for both chromosome 8 (left) and MYC (right).Middle: A breast carcinoma with ‘‘low-grade’’MYC amplification. Thereare two to three copies of both chromosome 8 (left) and MYC (right)
per nucleus. Bottom: A breast carcinoma with ‘‘high-grade’’ amplifica-tion. Both probes (chromosome 8, left;MYC, right) show more than fivesignals/nucleus, although MYC copies are more numerous. The elon-gated nucleus with two signals for chromosome 8 (midfield on the left)corresponds to host-derived fibroblasts or endothelium.
4 VISSCHER ET AL.

correlated with flow cytometric SPF. The meanSPF for tumors without amplification (n 5 3) was2.3% vs. 5.2% for tumors with low-grade amplifica-tion (n 5 6) and 19.2% for tumors with high- gradeamplification (n 5 10; P 5 0.002; two-sample ttest). Increased cell proliferation would not causean increased chromosome 8 copy number, becausethe number of centromeric signals remains constantduring cell cycle stages (Hopman et al., 1994).Another possible mechanism for increased MYC
copy number is numerical alteration of chromo-some 8. Our results suggest that this is a frequentoccurrence in breast carcinomas, as implied by thenear-uniform presence of an increased chromosome8 copy number in tumors with an increased MYCcopy number. It is noteworthy that both modal andtotal gains of chromosome 8 copy number were
nearly always less than the corresponding gains forMYC. We assume that this reflects the effects ofcellular DNA replication on MYC (but not onchromosome 8) copy number, as previously noted.Without doing karyotype studies, we are unable toascertain the extent to which observed aneusomiesfor chromosome 8 derive from selective chromo-somal duplication, from genomic endoreduplica-tion, or from a combination of these factors. Theapparent relationship between flow cytometricDNAindex and chromosome 8 copy number wouldimply that genomic endoreduplication (i.e.,‘‘tetra-ploidization’’) is the more frequent mechanism. Forexample, only 1 of 7 cases with peridiploid DNAcontent (DNA index 0.9–1.2) showed signal gainwith the chromosome 8 probe. On the other hand,both tumors with hypertetraploid DNA content
TABLE 2. Chromosome 8 andMYC Signal Counts in Malignant Cases With Absent, Minimal, or Low-Grade Amplification
Percent of nuclei with
Case no. Diagnosis DNA index Probe 0–2 Signals 3–5 Signals .5 Signals
1 Inv. duct.a 1.2 MYC 100 0 0chr 8b 100 0 0
2 Phyllodes, low grade NA MYC 92 8 0chr 8 100 0 0
3 Phyllodes, high grade 1.1 MYC 90 10 0chr 8 98 2 0
4 Inv. lob.c 1.0 MYC 100 0 0chr 8 100 0 0
5 Inv. lob. 0.9 MYC 85 15 0chr 8 99 1 0
6 DCIS, low graded NA MYC 87 13 0chr 8 99 1 0
7 Inv. duct. 1.0 MYC 63 37 0chr 8 100 0 0
8 Inv. lob. NA MYC 67 33 0chr 8 53 47 0
9 Inv. duct. 1.9 MYC 51 47 1chr 8 59 41 0
10 Inv. duct. 0.9 MYC 16 84 0chr 8 70 30 0
11 Inv. duct. NA MYC 16 83 1chr 8 15 84 1
12 Inv. duct. 1.3 MYC 9 89 2chr 8 14 86 0
13 Inv. duct. 1.7 MYC 10 83 7chr 8 3 97 0
14 Inv. duct. 1.3 MYC 19 71 10chr 8 19 74 7
15 Inv. duct. 1.7 MYC 0 90 10chr 8 4 94 2
aInvasive ductal carcinoma.bChromosome 8.cInvasive lobular carcinoma.dDuctal carcinoma in situ.boldface numbers 5 modal signal count/nucleus
5MYC FISH IN BREAST CANCER

contained .5 chromosome 8 copies per nucleus inthe majority of cells.The flow cytometric and interphase cytogenetic
data, taken together, suggest that selective intrachro-masomal or extrachromosomal gene sequence repli-cation is an uncommon mechanism ofMYC ‘‘ampli-fication.’’ In our study, in fact, there were only 3 of26 tumors (Table 3, cases 21, 23, and 24) in whichthe modalMYC copy number per nucleus exceededthe modal chromosome 8 copy number by morethan one category. These data correlate with therelatively infrequent occurrence of double minutesor homogeneously stained regions in karyotypes ofbreast carcinoma cells (Geleick et al., 1990). It ispossible that cases with MYC signal gain relative tochromosome 8 have extra copies of the 8q arm.Excluding this possibility could be accomplishedwith interphase cytogenetics, but it would requiresimultaneous dual (two-color) hybridization.Compared with molecular studies, our interphase
cytogenetic analysis reveals a more frequent inci-dence of abnormal MYC copy number in breastcarcinoma. This probably reflects the inherentsensitivity advantage of interphase cytogenetic tech-niques, in which the analysis may be performed
selectively on neoplastic populations. This largelydiminishes the dilutional effect of benign stromal orinflammatory cell populations that constitute, onaverage, 50% of the cells in breast carcinomas.Increased sensitivity is also likely to account for therelatively frequent occurrence of high-grade MYCcopy gain in which .5 copies/nucleus are observed.Molecular studies typically show that such cases arerelatively uncommon.Others have reported that MYC amplification in
breast carcinoma is correlated with an aggressivephenotype or clinical behavior (Tsuda et al., 1991;Berns et al., 1992; Kreipe et al., 1993). Althoughthey are limited by small numbers of cases, theobserved correlations with DNA aneuploidy andpoor differentiation in our study would appear toconfirm this association. It remains uncertain, how-ever, whether such associations are actually medi-ated by increased MYC expression. To date, thereare no published data that correlate elevated MYCmessage or gene product levels with adverse clinico-pathologic features in breast carcinomas. Our datasuggest that, in many cases, the MYC copy numbermay represent a surrogate marker for genetic insta-bility that has led to cellular DNA aneuploidy (i.e.,
TABLE 3. Chromosome 8 andMYC Signal Counts In Malignant Cases With High-Grade Amplification
Percent of nuclei with
Case no. Diagnosis DNA index Probe 0–2 Signals 3–5 Signals 6–9 Signals .9 Signals
16 Inv. duct.a 1.0 MYC 9 67 24 0chr 8b 100 0 0 0
17 Inv. duct. 1.3 MYC 0 44 48 8chr 8 25 72 3 0
18 Inv. duct. 1.5 MYC 0 0 87 13chr 8 6 22 71 1
19 Inv. duct. 1.5 MYC 0 4 84 12chr 8 2 2 72 24
20 Inv. duct. 1.6 MYC 3 7 90 0chr 8 7 85 8 0
21 Inv. duct. 1.7 MYC 0 0 7 93chr 8 5 89 6 0
22 Inv. duct. 1.8 MYC 0 8 78 14chr 8 10 79 10 1
23 Inv. lobc. NA MYC 0 1 6 93chr 8 1 84 15 0
24 Inv. duct. 2.0 MYC 5 9 40 46chr 8 31 66 3 0
25 Inv. duct. 2.4 MYC 0 0 0 100chr 8 0 0 37 63
26 Inv. duct. 3.2 MYC 5 2 7 86chr 8 0 0 18 82
aInvasive ductal carcinoma.bChromosome 8.cInvasive lobular carcinoma.boldface numbers 5 modal signal count/nucleus
6 VISSCHER ET AL.

as opposed to a direct cause of constitutive MYCexpression). Evaluation ofMYC copy number in thecontext of expression level would clearly be re-quired in order to clarify this issue as well as todefine the prognostic relevance of expression inhuman breast neoplasia.
REFERENCES
Berns EM, Klijn JG, van Putten WL, van Staveren IL, Portengen H,Foekens JA (1992) c-myc Amplification is a better prognostic factorthan HER2/neu amplification in primary breast cancer. Cancer Res1:1107–1113.
Bonilla M, Ramirez M, Lopez-Cueto J, Gariglio P (1988) In vivoamplification and rearrangement of c-myc oncogene in humanbreast tumors. J Natl Cancer Inst 80:665–671.
Escot C, Theillet C, Lidereau R, Spyratos F, Champeme MH, GestJ, Callahan R (1986) Genetic alteration of the c-myc protooncogene(MYC) in human primary breast carcinomas. Proc Natl Acad SciUSA 83:4834–4838.
Fontana X, Ferrari P, Milano G, Abbes M, Monticelli J, Namer M,Bussiere (1994) Analysis of c-myc amplification by the differentialpolymerase chain reaction (d-PCR), study in breast cancer. OncolReports 1:311–366.
Geleick D, Muller H, Matter A, Torhorst J, Regenass U (1990)Cytogenetics of breast cancer. Cancer Genet Cytogenet 46:217–229.
Hopman AH, Voorter CE, Ramaeker FC (1994) Detection ofgenomic changes in cancer by in situ hybridization. Biol Reports19:31–44.
Kreipe H, Feist H, Fischer L, Felner J, Heidorn K, Mettler L,Parwaresch R (1993) Amplification of c-myc but not of c-erbB-2 isassociated with high proliferative capacity in breast cancer. CancerRes 53:1956–1961.
Meyers SL, O’Brien MT, Smith T, Dudley JP (1990) Analysis of theint-1, int-2, c-myc, and neu oncogenes in human breast carcinomas.Cancer Res 50:5911–5918.
Pavelic ZP, Pavelic K, Carter C, Pavelic L (1992) Heterogeneity ofc-myc expression in histologically similar infiltrating ductal carcino-mas of the breast. J Cancer Res Clin Oncol 118:16–22.
Pertschuk LP, Feldman JG, Kim DS, Nayeri K, Eisenberg KB,Carter AC, Thelmo WT, Rhong ZT, Benn P, Grossman A (1993)Steroid hormone receptor immunohistochemistry and amplifica-tion of c-myc protooncogene. Cancer 71:162–171.
Tsuda H, Hirohashi S, Hirota T, Shimosato Y (1991) Alterations incopy number of c-erbB-2 and c-myc, proto-oncogenes in advancedstage of human breast cancer. Acta Pathol Jpn 41:19–23.
Visscher DW, Zarbo RJ, Jacobsen G, Kambouris A, Talpos G, Sakr W,Crissman JD (1990) Multiparametric deoxyribonucleic acid andcell cycle analysis of breast carcinomas by flow cytometry. LabInvest 62:370–378.
7MYC FISH IN BREAST CANCER