Embed Size (px)
Citation preview

Eolss Publishers Co. Ltd., UK Copyright © 2017 Eolss Publishers/ UNESCO Information on this title: www.eolss.net/eBooks ISBN- 978-1-78021-040-7 e-Book (Adobe Reader) ISBN- 978-1-78021-540-2 Print (Color Edition) The choice and the presentation of the facts contained in this publication and the opinions expressed therein are not necessarily those of UNESCO and do not commit the Organization. The designations employed and the presentation of material throughout this publication do not imply the expression of any opinion whatsoever on the part of UNESCO concerning the legal status of any country, territory, city, or area, or of its authorities, or the delimitation of its frontiers or boundaries. The information, ideas, and opinions presented in this publication are those of the Authors and do not represent those of UNESCO and Eolss Publishers. Whilst the information in this publication is believed to be true and accurate at the time of publication, neither UNESCO nor Eolss Publishers can accept any legal responsibility or liability to any person or entity with respect to any loss or damage arising from the information contained in this publication. All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage or retrieval system, without prior permission in writing from Eolss Publishers or UNESCO. The above notice should not infringe on a 'fair use' of any copyrighted material as provided for in section 107 of the US Copyright Law, for the sake of making such material available in our efforts to advance understanding of environmental, political, human rights, economic, democracy, scientific, and social justice issues, etc. If you wish to use copyrighted material from this e-book for purposes of your own that go beyond 'fair use', you must obtain permission from the EOLSS Publishers. Every effort has been made to trace and credit all the copyright holders, but if any have been inadvertently overlooked, UNESCO and Eolss Publishers will be pleased to make the necessary arrangements at the first opportunity. British Library Cataloguing-in-Publication Data A catalogue record of this publication is available from the British Library. Library of Congress Cataloging-in-Publication Data A catalog record of this publication is available from the library of Congress Singapore

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
DISEASES CAUSED BY VIRAL PATHOGENS Toshihiro Nakai Graduate School of Biosphere Science, Hiroshima University, 1-4-4 Kagamiyama, Higashi-Hiroshima 739-8528, Japan Motohiko Sano, Department of Marine Biosciences, Tokyo University of Marine Science and Technology, Konan 4-5-7, Minato-ku, Tokyo 108-8477, Japan. Mamoru Yoshimizu, Hisae Kasai Laboratory of Biotechnology and Microbiology, Faculty of Fisheries Sciences, Hokkaido University, Hakodate, Hokkaido 041-8611, Japan Toshiaki Itami Department of Aquaculture, Faculty of Agriculture, University of Miyazaki, 1-1 Gakuen Kibanadai-nishi, Miyazaki 889-2192, Japan. Raja Sudhakaran School of Biosciences and Technology, VIT University, Vellore 600 014, Tamilnadu, India Keywords: Viral disease; Freshwater fish, SVC, CCVD, KHVD, HVHN, EHN, Red sea bream iridoviral disease (RSIVD), Viral nervous necrosis (VNN), Megalocytivirus, Betanodavirus, Salmonid disease, IPN, IHN, VHS, OMVD, ISA, shrimp, virus Contents 1. Freshwater Fish 2. Marine Fish 3. Salmon and Trout Viral Diseases 4. Shrimp Diseases Glossary Bibliography Biographical Sketches Summary Viral diseases cause particularly severe damages in aquaculture in the world, mainly due to lack of effective chemotherapeutics and limited numbers of commercially available vaccines. Some of the diseases have spread among wild populations of fish as well as cultured ones, resulting in considerable loss in fisheries and the natural resources. In this section, we selected important viral diseases of fish and shellfish (shrimp), and describe their disease agents, geographical distributions, host ranges, and diagnostic and control methods. These include five diseases (SVC, CCVD, KHVD, HVHN, EHN) in freshwater fish except for warm-water fish, two (RSIVD, VNN) in marine fish except for salmonids, five (IPN, IHN, VHS, OMVD, ISA) in salmon and trout, and six (WSD, YHD, IHHN, TS, WTD, IMN) in penaeid shrimp. Most of the diseases are currently listed by the World Organization for Animal Health (OIE) and thus are of great importance for the
278
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
international trade of aquatic animals. 1. FRESHWATER FISH Motohiko Sano 1.1. Synopsis/Abstract This section describes the viral diseases affecting warm-water fin-fish in fresh water. Although many viral diseases and the causative viruses have been reported to date, this section focuses on the five most important viral diseases. 1.2. Introduction Fresh water fish form the largest segment of the world aquaculture production and a major part of the catch-fish industry in inland waters. The harvest of farmed warm-water fish is increasing, and consequently, fry production of fish species for aquaculture has also rapidly increased and disease problems have become the largest obstacle in aquaculture operations. Some diseases of cultured fish (e.g. koi herpesvirus disease) have spread to wild populations of fish which pose serious threats to the ecosystem. . Wolf (1988) described 59 fish viruses, and, subsequently, the number of fish viruses reported continues to increase so far. Viruses of warm-fresh water fish of major importance are shown in Table 1.1 (Sano et al, 2011). This section focuses on diseases caused by 5 of the viruses in the table: spring viremia of carp, channel catfish virus disease, koi herpesvirus disease, herpesviral hematopoietic necrosis and epizootic hematopoietic necrosis. These diseases are transboundary aquatic animal diseases, of international concern due to their significance in the international trade of aquatic animals. Most of the diseases in this section are currently listed by the World Organisation for Animal Health (OIE) (Aquatic Animal Health Code 2012).
Virus Taxonomy (Family)
Main Host
DNA Virus Carp herpesvirus (CHV) (=CyHV-1) Alloherpesviridae Carp (Cyprinus carpio) Goldfish hematopietic necrosis virus (GFHNV) (=CyHV-2)
Alloherpesviridae Goldfish (Carassius auratus)
Koi herpesvirus (KHV) (=CyHV-3) Alloherpesviridae Carp Channel catfish virus (CCV) (=IcHV-1)
Alloherpesviridae Channel catfish (Ictalurus punctatus)
Ictalurus melas herpesvirus (IcmHV) (=IcHV-2)
Alloherpesviridae Black bullhead (Ameiurus melas)
Herpesvirus anguillae (HVA) (AngHV-1)
Alloherpesviridae Japanese eel (Anguilla japonica), European
White sturgeon herpesvirus 1 (WSHV-1) (=AciHV-1)
Alloherpesviridae White sturgeon (Acipenser transmontanus)
White sturgeon herpesvirus 2 (WSHV-2) (=AciHV-2)
Alloherpesviridae White sturgeon
279
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Epizootic hematopoietic necrosis virus (EHNV)
Iridoviridae Redfin perch (Perca fluviatilis)
European catfish virus (ECV) Iridoviridae Black bullhead European sheatfish virus (ESV) Iridoviridae Sheatfish (Silurus glanis) Largemouth bass virus (LMBV) Iridoviridae Largemouth bass
(Micropterus salmoides) White sturgeon iridovirus (WSIV) Iridoviridae White sturgeon Infectious spleen and kidney necrosis virus (ISKNV)
Iridoviridae Various freshwater species
Carp edema virus (CEV) Pox-like virus Carp RNA Virus Spring viremia of carp virus (SVCV) Rhabdoviridae Carp Viral hemorrhagic septicaemia virus (VHSV) (genogroup IVb)
Rhabdoviridae Various freshwater species
Pike fry rhabdovirus (PFRV) Rhabdoviridae Northern pike (Esox lucius)Perch rhabdovirus (PRV) Rhabdoviridae Redfin perch Snakehead rhabdovirus (SHRV) Rhabdoviridae Snakehead fish
(Ophicephalus striatu) Aquabirnaviruses (e.g. Eel virus European)
Birnaviridae Various freshwater species
Golden shiner virus (GSV) Reoviridae Golden shiner (Notemigonus crysoleucas)
Grass carp reovirus (GCRV) Reoviridae Grass carp (Ctenopharyngodon idella)
Table 1.1. Major viruses of warm water freshwater species.
1.3. Spring Viremia of Carp 1.3.1. Introduction Spring viremia of carp (SVC) is an acute, systemic, contagious disease caused by a rhabdovirus. SVC in carp typically occurs at 11-17°C, predominantly in spring. Mortality can reach to 30-70 %. This disease is currently listed by the OIE. Selected references or reviews: (Wolf, 1988; Sano et al, 2011; Aquatic Animal Health Code 2012; Fijan et al, 1971; Ahne et al, 2002; Walker and Winton, 2010; OIE, 2013; Stone et al, 2003). 1.3.2. Disease Agent Virus: spring viremia of carp virus (SVCV) (available collection: ATCC VR-1390) Virus taxonomy: genus Vesiculovirus, family Rhabdoviridae, order Mononegavirales Morphology: typical bullet shaped virion with 60-90 nm wide and 80-180 nm long Virion proteins: 5 structural proteins: L (238 kDa), G (57 kDa), N (47 kDa), P (35 kDa) and M (25 kDa) Genome: single-stranded negative-sense RNA of ca. 11,000 nucleotide bases, encoding the structural proteins in the order 3'- N-P-M-G-L-5'; complete genome sequence: accession No. U18101, AJ318079, DQ097384, DQ491000, EU177782
280

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Serotype: single (SVCV and pike fry rhabdovirus (PFRV) can be two serotypes of a single virus species) Genotype: single genotype (Genogroup I) with 4 subgenogroups (Ia to Id) consisted with the geographical origin 1.3.3. Geographical Distribution European countries, USA, Canada and China 1.3.4. Host Range Carp (Cyprinus carpio) and other cyprinid species (eg. goldfish (Carassius auratus), grass carp (Ctenopharyngodon idella), crucian carp (Carassius carassius)). 1.3.5. Diagnostic Methods Clinical signs: External signs are non-specific, but likely include skin darkening, abdominal distension, exophthalmos, petechial hemorrhage in the skin and gills, and pale gills. Gross pathology: Internal signs are dominated by edema in all organs, hemorrhage, peritonitis and catarrhal enteritis. Excess ascites may be bloody. Petechia is evident in the muscles and internal organs including swim-bladder. Histopathology: Changes including hemorrhage, hyperemia, multiple focal necrosis, perivascular inflammation, and edema and necrosis of blood vessels can be observed in all major organs, especially liver, kidney and spleen.
Figure 1.1. CPE on EPC cells following infection with SVCV. Diagnosis: Specific diagnosis is generally based on the isolation of SVCV in cell culture (EPC or FHM) at 20°C by inoculation with homogenates of kidney, spleen, liver and
281

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
encephalon, followed by the identification. The typical CPE is of the rounded cells [Figure 1.1]. Isolated virus is identified using serological techniques (virus neutralization, IFAT, ELISA) and nucleotide based methods (RT-PCR, real-time RT-PCR or LAMP). 1.3.6. Control SVCV is generally transmitted horizontally. Outbreaks can be prevented or stopped by raising water temperatures above 20°C. No chemotherapeutic treatments and commercial vaccines are currently available. General biosecurity measures and regular hygiene practices on farm level are applicable. Avoidance of crowding during winter and early spring is essential to reduce spread of the virus. A fish selection program resulted in high resistance of the Krasnodar strain of carp. 1.4. Channel Catfish Virus Disease 1.4.1. Introduction Channel catfish virus disease (CCVD) is an acute, systemic, contiguous and highly species-specific disease of young channel catfish in the USA caused by an alloherpesvirus. CCVD in channel catfish typically occurs at high water temperature ranging approximately 20-30°C. Mortality can occasionally approach 100%. Selected references or reviews: (Wolf, 1988; Sano et al, 2011; Wolf and Darlington, 1971; Plumb, 1989 ; Davison, 1992 ; Camus, 2004 ; Hanson et al, 2011 ; Waltzek et al, 2009) 1.4.2. Disease Agent Virus: Channel catfish virus (CCV) (available collection: ATCC VR-665) Virus taxonomy: genus Ictalurivirus, family Alloherpesviridae, order Herpesvirales Morphology: 175-200 nm virion consisted of 100 nm icosahedral nucleocapsid with an envelop Virion proteins: 32 polypeptides detected Genome: double-stranded DNA of 134 kbp with 90 genes predicted; complete genome sequence: accession No. NC_001493 Serotype: single Genotype: not available 1.4.3. Geographical Distribution USA, Mexico 1.4.4. Host Range Channel catfish (Ictalurus punctatus) and blue catfish (Ictalurus furcatus). White catfish (Ictalurus cams) is susceptible to experimental infection. 1.4.5. Diagnostic Methods Clinical signs: Signs can vary and depend on the degree of kidney damage due to CCV
282

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
multiplication, and usually include distension of abdomen, exophthalmia, swollen and protruding vent, hemorrhage at the base of ventral and caudal fins, in gills and skin. Gross pathology: The peritoneal cavity is hyperemic and contains a clear, yellowish or slightly reddish fluid. Liver and kidney may be pale, with or without hemorrhage or petechiae. The spleen is congested and dark. A yellowish mucoid material is present in the intestine. Histopathology: Severe changes, consisting of edema, hemorrhage and necrosis can be observed in the internal organs, especially in kidney. The hematopoietic tissue shows an increase in lymphoid cells, edema, necrosis and accumulation of macrophages. Necrosis and occasional hemorrhage develop in nephrons. Diagnosis: Specific diagnosis is generally based on the isolation of CCV in cell culture (CCO and BB) at 25°C by inoculation with homogenates of kidney and spleen, followed by the identification. The typical CPE is of cell enlargement and syncytium formation. Isolated virus is identified using serological techniques (virus neutralization, IFAT, ELISA) and also nucleotide based methods (PCR, real-time RT-PCR). PCR is at present, the most useful method. 1.4.6. Control CCV is generally transmitted horizontally. The reduction of water temperature to 19°C or lower may decrease the mortality. No chemotherapeutic treatments and commercial vaccines are currently available. General biosecurity measures and regular hygiene practices on farm level are applicable. Lower stocking densities for production of fingerlings and appropriate daily feeding rates are important, especially during high water temperatures. Fingerlings should be harvested and handled only below 20°C. Breeding for resistance and hybridization of channel catfish strains is a promising approach. 1.5. Koi Herpesvirus Disease 1.5.1. Introduction Koi herpesvirus disease (KHVD) is an acute, systemic, highly contagious disease caused by an alloherpesvirus. KHVD typically occurs at 17-28°C. Mortality can approach 100%. This disease is currently listed by the OIE. Selected references or reviews: (Wolf, 1988; Sano et al, 2011; Aquatic Animal Health Code 2012; OIE, 2013; Hanson et al, 2011; Waltzek et al, 2009; Hedrick et al, 2000; Haenen et al, 2004; Haenen and Hedrick, 2006; Michel et al, 2010 ; Kurita et al, 2009)[1-3, 7, 13-19]. 1.5.2. Disease Agent Virus: koi herpesvirus (KHV) (=cyprinid herpesvirus 3(CyHV-3)) (available collection: ATCC VR-1592) Virus taxonomy: genus Cyprinivirus, family Alloherpesviridae, order Herpesvirales Morphology: 170-200 nm virion consisted of 110-120 nm icosahedral nucleocapsid with an envelop
283

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Virion proteins: 40 polypeptides detected Genome: double-stranded DNA of 295 kbp with 163 genes predicted; complete genome sequence: accession No. NC_009127 (DQ177346 (strain I); DQ657948 (strain U); AP008984 (strain J=TUMST1) Serotype: single Genotype: two distinct (European and Asian) lineages 1.5.3. Geographical Distribution European countries, Asian countries, North America, Israel, South Africa 1.5.4. Host Range Carp (Cyprinus carpio) (including ornamental varieties such as koi) 1.5.5. Diagnostic Methods Clinical signs: The most consistent sign is an irregular discoloration of the gills consistent with necrosis (Figure 1.2). Other signs include anorexia, exophthalmia, fin erosion, hemorrhage on the skin and base of the fins, pale irregular patches on the skin associated with excess mucus secretion.
Figure 1.2. A common carp with KHVD showing necrosis of the gill filaments and enophthalmia.
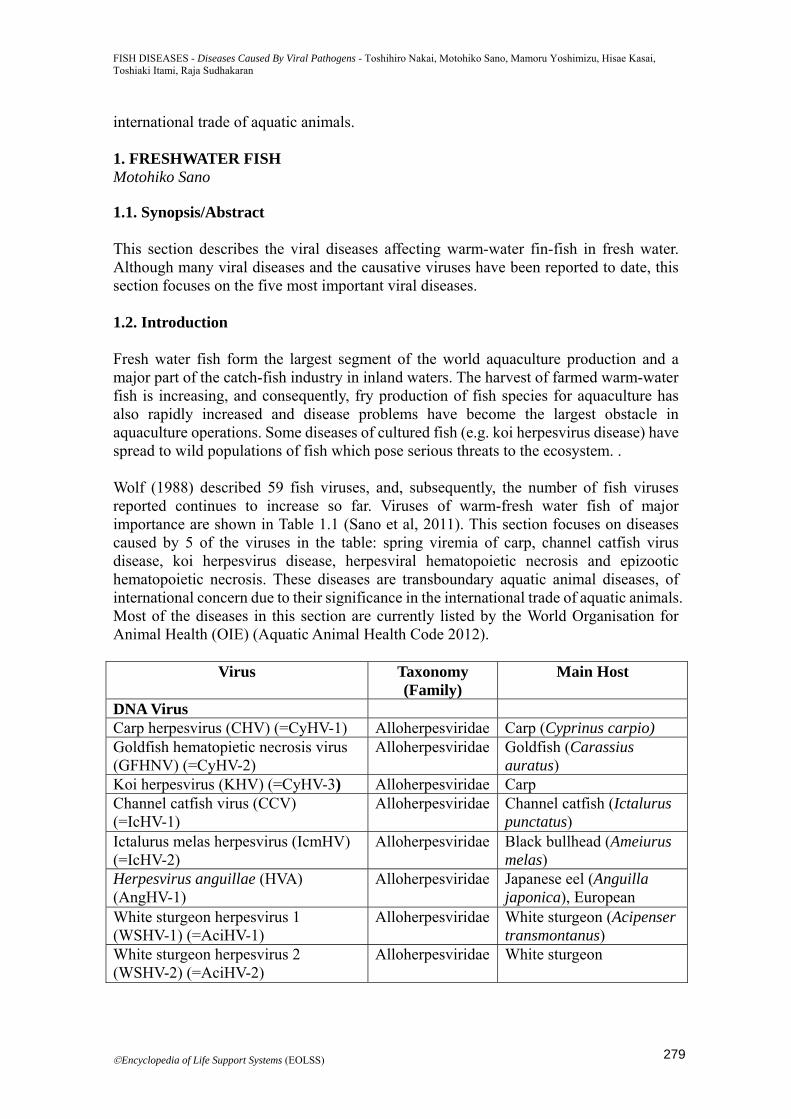
Gross pathology: Internal gross signs are inconsistent, but enlarged kidney, swollen spleen and heart are occasionally observed. Histopathology: Changes are not consistent, but necrosis of gill tissues and hyperplasia and hypertrophy of the branchial epitherial cells and fusion of adjacent secondary lamellae are commonly found (Figure 1.3). Inflammation, necrosis, and nuclear swelling, margination of chromatin and plae diffuse eosinophilic inclusions may be observed in the organs including gill, kidney, gastrointestinal system and skin.
284
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
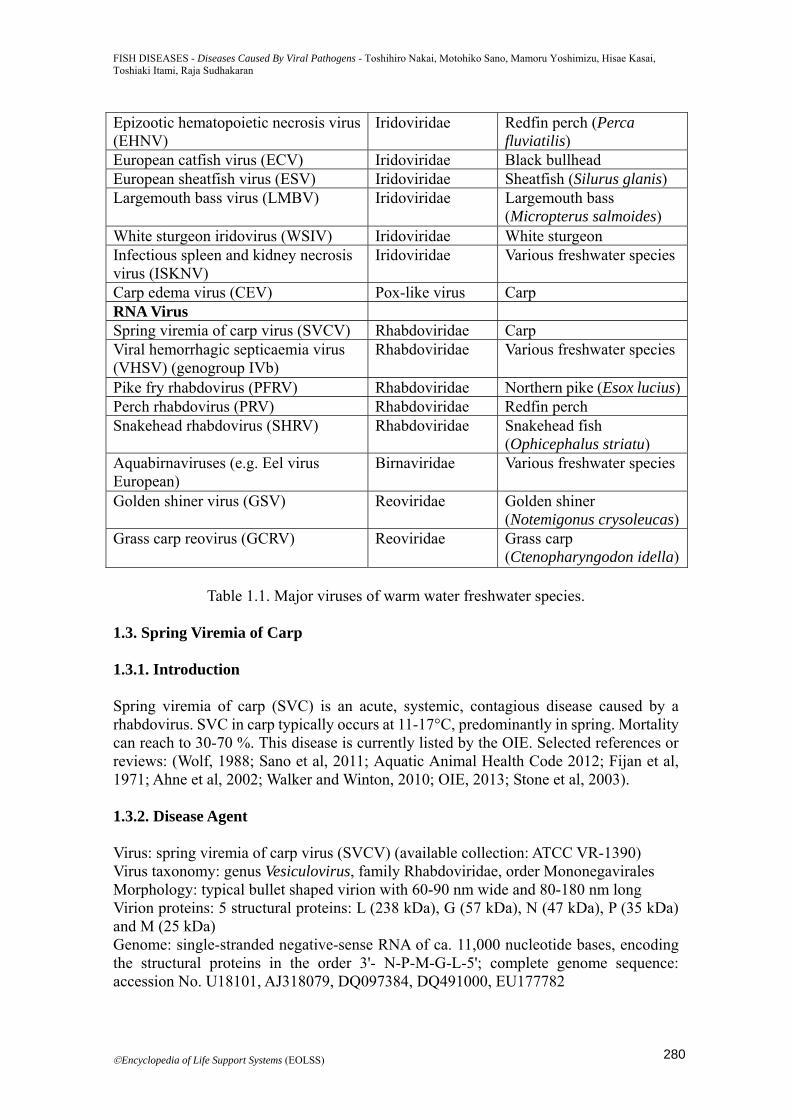
Diagnosis: Specific diagnosis is based on direct method such as virus isolation, viral antigen detection using IFAT or ELISA, and viral DNA amplification assay using PCR, real-time PCR or LAMP. PCR is currently considered most reliable method. Virus isolation can be done in KF-1 or CCB at 20°C, but is probably difficult to achieve reliably. The typical CPE is of syncytium formation and intense cytoplasmic vacuolation (Figure 1.4). Antibody-capture ELISA is helpful for screening the fish experienced with the disease.
Figure 1.3. Tissue section of the gills of common carp infected with KHV showing fusion of secondary lammelae. Courtesy of Dr. S. Miwa.
Figure 1.4. CPE on CCB cells following infection with KHV. 1.5.6. Control KHVD is generally transmitted horizontally. Outbreaks can be prevented or stopped by raising water temperatures above 28°C. No chemotherapeutic treatments are available. A vaccine using attenuated virus is commercially licensed in Israel. Survivor fish in KHVD, which is persistently or latently infected with the virus, is considered a potential risk as an
285

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
infection source through worldwide trade, and, therefore, koi farm should aim to produce KHV-free fish. Egg or fish should be introduced in aquaculture facility from KHV-free farm. Breeding for resistance of carp strains is a promising approach. 1.6. Herpesviral Hematopoietic Necrosis 1.6.1. Introduction Herpesviral hematopoietic necrosis (HVHN) is an acute, systemic, highly contagious disease caused by an alloherpesvirus. HVHN in goldfish typically occurs at 15-25°C. Mortality can reach 100%. Selected references: (Waltzek et al, 2009; Jung and Miyazaki 1995; Waltzek et al, 2009; Goodwin et al, 2006; Li et al, 2003; Goodwin et al, 2009; Jeffery et al, 2007; Wang et al, 2012). 1.6.2. Disease Agent Virus: goldfish hematopoietic necrosis virus (GFHNV) (=cyprinid herpesvirus 2(CyHV-2)) Virus taxonomy: genus Cyprinivirus, family Alloherpesviridae, order Herpesvirales Morphology: 170-220 nm virion consisted of 115-117 nm icosahedral nucleocapsid with an envelop Virion proteins: not available Genome: double-stranded DNA Serotype: not available Genotype: not available 1.6.3. Geographical Distribution Japan, Taiwan, China, USA, UK, Australia, Czech (possibly distributed by hobbyist aquariums worldwide) 1.6.4. Host Range Goldfish (Carassius auratus) and Prussian carp (Carassius gibelio) 1.6.5. Diagnostic Methods Clinical signs: Diseased fish show a pale skin and necrotic pale gills sometimes with white patches, and occasionally abdominal distension and exophthalmia. Gross pathology: Pale gills and liver, ascites, splenomegaly with white nodular lesions, swollen pale kidney, and an intestine devoid of food are often observed. Histopathology: The most severe lesions are found in the kidney and spleen. Necrosis in the hematopoietic cells and renal tubular epithelia and glomeruli in the kidney, and extensive necrosis of pulp and sheathed arteries are observed. Diagnosis: Specific diagnosis is based on direct detection by PCR or real-time PCR.
286

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
GFHNV is difficult to isolate in cell culture and is typically lost after several passages in EPC, FHM and KF-1. Successful propagation using GFF (GF-1) derived from goldfish fin has been reported. The typical CPE is of intense cytoplasmic vacuolation similar with those of KHV. 1.6.5. Control GFHNV is generally transmitted horizontally. Outbreaks in goldfish can be stopped by raising water temperatures at 33°C for over 4 days, and the treated fish may become carrier of the virus. No chemotherapeutic treatments and commercial vaccines are currently available. General biosecurity measures and regular hygiene practices on farm level are applicable. Susceptibility to the virus is different among goldfish varieties, and breeding for resistance of goldfish strains is a promising approach. 1.7. Epizootic Hematopoietic Necrosis 1.7.1. Introduction Epizootic hematopoietic necrosis (EHN) is an acute, systemic, contagious disease in mostly redfin perch in Australia caused by an iridovirus. EHN in redfin perch occurs at 12-21°C. Mortality, especially in juvenile redfin perch, can approach 100%. This disease is currently listed by the OIE. Selected references or reviews: (Sano et al, 2011; Aquatic Animal Health Code 2012; OIE, 2013 ; Langdon et al, 1986 ; Chinchar, 2002 ; Chinchar et al, 2009; Whittington et al, 2010; Jancovich et al, 2010). 1.7.2. Disease Agent Virus: Epizootic hematopoietic necrosis virus (EHNV) Virus taxonomy: genus ranavirus, family Iridoviridae Morphology: approximately 175 nm icosahedral virion with an inner lipid layer Virion proteins: 30 polypeptides detected (Frog virus 3 (FV-3), a type strain of Ranavirus)) Genome: double-stranded DNA of 127 kbp with 100 genes predicted; complete genome sequence: accession No. FJ433873 Serotype: not available (EHNV and ictalurid isolates, European catfish virus (ECV) and European sheatfish virus (ESV), show serological and genetic relatedness to FV-3) Genotype: not available 1.7.3. Geographical Distribution Australia 1.7.4. Host Range Redfin perch (Perca fluviatilis) and rainbow trout (Oncorhynchus mykiss). A number of other species such as Macquarie perch (Macquaria australasica), silver perch (Bidyamus bidyanus), mosquito fish (Gambusia affinis), mountain galaxias (Galaxia olidus) are susceptible to experimental infection.
287

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
1.7.5. Diagnostic Methods Clinical signs: Signs are non-specific, but likely include skin darkening, erythema around the brain and nostrils, and hemorrhage around bases of fins. Gross pathology: Internal gross signs are inconsistent, but some fish show enlargement of kidney, liver or spleen. There may be focal whitish lesions in the liver corresponding to areas of necrosis. Histopathology: Changes consist of acute focal, multifocal or locally extensive coagulative or liquefactive necrosis of liver, hematopoietic kidney and spleen, and also necrosis in heart, pancreas, gastrointestinal tract, gill and pseudobranch. Diagnosis: Specific diagnosis is generally based on the isolation of EHNV in cell culture (BF-2 or FHM) at 22°C by inoculation with homogenates of liver, kidney and spleen, followed by the identification. The typical CPE is of the rounded cells [Figure 1.5]. Isolated virus is identified using serological techniques (ELISA) and nucleotide based methods (PCR-restriction endonuclease analysis). 1.7.6. Control EHNV is generally transmitted horizontally. Outbreaks may be prevented by shifting water temperatures below 12°C. No chemotherapeutic treatments and commercial vaccines are currently available. Since EHN is endemic in the limited water basin of Australia, general biosecurity measures are applicable for prevention of spreading this disease.
Figure 1.5. CPE on BF-2 cells following infection with EHNV.
288

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
2. Marine Fish
Toshihiro Nakai
2.1. Synopsis
With the rapid development of mass seed production techniques in fish, aquaculture
targeting a variety of marine fish species has been intensively conducted in various
areas of the world since 1980s. During fish rearing in hatcheries and farms, however,
infectious diseases with high mortalities have frequently occurred, resulting in severe
economical losses. It is no doubt that disease is one of the most important limiting
factors for further development of marine aquaculture (Liao, 2009). This chapter
describes viral diseases of marine fish excepting for salmonids, with particular reference
to iridoviral disease caused by megalocytiviruses (Iridoviridae) and viral nervous
necrosis caused by betanodaviruses (Nodaviridae), both of which are of great
importance in world marine aquaculture due to their high geographical distribution, host
range and virulence.
2.2. Introduction
Major viruses of non-salmonid marine fish causing diseases are listed in Table 2.1.
Flounder herpesvirus (FHV), which is classified in Alloherpesviridae, causes often mass
mortality in hatchery-reared larvae of Japanese flounder (Paralichthys olivaceus) in
Japan (Iida et al, 1989, 2008). The disease is characterized by epidermal hyperplasia on
the fins and skin with vacuolar degeneration of the Malpighian cells. Cell culture
isolation and propagation of FHV have not succeeded. Another herpesvirus belonging to
Alloherpesviridae, Pilchard herpesvirus (PHV), was found in the affected Australasian
pilchard (Sardinops sagax neopilchardus) during a large-scale epizootic in Australian
and New Zealand coastlines (Whittington et al, 1997; Hyatt et al, 1997). Significant
lesions are confined to the gills where the virus is consistently present. Aquatic
birnaviruses, which belong to the genus Birnavirus (Birnaviridae), are differentiated
from infectious pancreatic necrosis virus (IPNV) causing disease in salmonid fish
(Smail and Snow 2011). Specified clinical cases in non-salmonid marine fish species by
aquatic birnaviruses are limited to yellowtail (Seriola quinqueradiata) (Sorimachi and
Hara 1985) and some flatfish species such as turbot (Psetta maxima), Atlantic halibut
(Hippoglossus hippoglossus) and Senegalese sole (Solea senegalensis) (Castric et al,
1987; Mortensen et al, 1990 ; Rodriguez Saint-Jean et al, 1997).
Virus Taxonomy
Family / Genus
Main host fish Reference
Flounder
herpesvirus (FHV)
Alloherpesviridae Japanese flounder
(Paralichthys olivaceus)
Iida et al (1989, 2008)
Pilchard herpesvirus
(PHV)
Alloherpesviridae Pacific sardine (Sardinops
sagax)
Whittington et al
(1997), Hyatt et al
(1997)
Aquabirnaviruses
Yellowtail ascites
virus (YTAV)
and unspecified
viruses
Birnaviridae /
Aquabirnavirus
Yellowtail (Seriola
quinqueradiata)
Turbot (Psetta maxima)
Atlantic halibut
(Hippoglossus
Sorimachi and Hara T.
(1985), Castric et al
(1987), Mortensen et
al (1990), Rodriguez
Saint-Jean et al (1997)
289
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
hippoglossus)
Senegalese sole (Solea
senegalensis)
Lymphocystis
disease virus
(LCDV)
Iridoviridae /
Lymphocystisvirus
Various marine and
freshwater species
Wolf et al. (1966)
Red sea bream
iridovirus (RSIV)
Infectious spleen
and kidney necrosis
virus (ISKNV)
Iridoviridae /
Megalocytivirus
Various marine and
brackishwater species
Inouye et al. (1992),
OIE (2012), Jancovich
et al (2011)
Betanodaviruses
(NNV)
Nodaviridae /
Betanodavirus
Various marine species Mori et al (1992),
Munday et al (2002)
Viral haemorrhagic
septicaemia virus
(VHSV)
Rhabdoviridae
/Novirhabdovirus
Flatfish, cod and others Schlotfeldt et al
(1991), Ross et al
(1994), Isshiki et al
(2001) Kim et al
(2009)
Hirame rhabdovirus
(HIRRV)
Rhabdoviridae
/Novirhabdovirus
Japanese flounder Kimura et al (1986),
Nishizawa et al (1991)
Modified from Sano et al. (2011)
Table 2.1. Major viruses of non-salmonid marine fish species
Lymphocystis disease virus (LCDV) belonging to the genus Lymphocystis
(Iridoviridae) causes benign, wart-like lesions comprising of grossly hypertrophoid cells
in the body surface of fish. The disease (lymphocystis) has been recorded in a variety of
cultured/wild marine and freshwater fish species worldwide (Wolf et al, 1966). Hirame
rhabdovirus (HIRRV) infection has caused mortalities in hatchery-reared juvenile
Japanese flounder in Japan (Kimura et al, 1986; Nishizawa et al, 1991). The disease can
be controlled experimentally by rearing fish at the water temperatures higher than 16°C
(Oseko et al, 1988). Compared with these viral diseases, which have caused somewhat
limited damages in some regions, the other three diseases, i.e. red sea bream iridoviral
disease (RSIVD) caused by megalocytiviruses (Iridoviridae), viral nervous necrosis
(VNN) caused by betanodaviruses (Nodaviridae), and viral hemorrhagic septicaemia
(VHS) caused by viral hemorrhagic septicaemia virus (VHSV) belonging to the genus
Novirhabdovirus in the family Rhabdoviridae, have higher geographical distribution,
host range and virulence, thus, they have higher socio-economic importance in world
marine aquaculture. Both RSID and VHS are listed in the aquatic animal health code by
the World Organization for Animal Health (OIE, 2011). VHS has long been known as
the most serious disease in farmed rainbow trout (Oncorhynchus mykiss) in Europe
since 1950s, and the disease to date is recognized as a disease of both farmed salmonid
and non-salmonid fish including a wide range of wild fish, with a global distribution
(Smail and Snow 2011). Although VHS outbreaks have been reported in farmed non-
salmonid marine fish, they are limited to turbot (Scophthalmus maximus) in Europe
(Schlotfeldt et al, 1991; Ross et al, 1994) and Japanese flounder in Japan and Korea
(Isshiki et al, 2001; Kim et al, 2009). In this chapter, RSIVD and VNN regarding short
history of the disease, property of the virus, diagnostic methods and control of the
disease are briefly described.
290

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
2.3. Red Sea Bream Iridoviral Disease (RSIVD) Red sea bream iridoviral disease (RSIVD) was first described in farmed juvenile red sea bream (Pagrus major) in 1990 in Japan (Inouye et al, 1992). Currently, more than 30 cultured marine fish species have been recorded as susceptible hosts in south-western parts of Japan, mostly fish belonging to Perciformes (Matsuoka et al, 1996; Kawakami and Nakajima, 2002). OIE (2012) listed 41 host fish species including 2 hybrid species for RSIVD (OIE, 2012). Similar diseases were reported in East and Southeast Asian countries, which were caused by infectious spleen and kidney necrosis virus (ISKNV) isolated from cultured mandarin fish (Siniperca chuatsi) (He et al, 2000, 2001) or other related viruses (Shi et al, 2004), resulting in severe mortalities from juvenile to marketable fish. Some review papers on RSIVD have been published (Nakajima et al, 2001; Sano et al, 2011). The RSIV and ISKNV are currently classified into the genus Megalocytivirus of the family Iridoviridae, in which the Ranavirus and Lymphocystivirus genera are also included as fish-pathogenic iridoviruses. All of the present megalocytiviruses are regarded as strains of the same or closely-related viral species (Jancovich et al, 2011). Virions of the Megalocytivirus (type species: ISKNV) possess icosahedral symmetry with about 140-200 nm in diameter, and contain a single, linear dsDNA molecule; the genome size of ISKNV is 111,362 bp with 54.8% G+C content. The complete genome sequences of other megalocytiviruses have been reported, the genome sizes being approximately 112-121 kbp (Kurita et al, 2002; He et al, 2001 ; Do et al, 2004 ; Lu et al, 2005; Ao and Chen, 2006). Figure 2.1 shows RSIV virions in the spleen of the infected red sea bream (Inouye et al, 1992). RSIV are unenveloped but sensitive to ether and chloroform, suggesting the presence of a thin lipid layer. Several fish cell lines, i.e. BF-2, CHSE-214, FHM, GF, JSKG, KRE-3, RYG-2 and YTF, are available to propagate RSIV at optimal incubation temperatures of 20-25°C. Among these cell lines, GF (grunt fin) is the most permissive to the virus, but virus productivity remains at low level (105 TCID50/ml) (Nakajima and Sorimachi, 1994). Recently, a stably susceptible cell line (MFF-1) to ISKNV was reported (Dong et al, 2008).
Figure 2.1. Red sea bream iridovirus (RSIV) virions in the spleen of the infected red sea bream (Inoue et al. 1992).
291

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
RSIVD occurs in the summer season at water temperature of 25°C and above in Japan (Matsuoka et al, 1996; Kawakami and Nakajima, 2002; Jun et al, 2009). Juveniles are more susceptible than adults. However, the disease has never been observed in hatcheries (Muroga, 2001). Gloss findings of the affected red sea bream are severe anemia, petechiae of the gills and enlargement of the spleen. The disease is most characterized by the presence of enlarged cells in the spleen, heart, kidney, intestine and gills of the affected fish. The enlarged cells (about 20 μm in diameter) are basophilic and Feulgen-positive, and may originate in infected leucocyte (Inouye et al, 1992). Cell culture isolation of the virus is not reliable due to lower permissibility of the cultured cells. IFAT with a specific monoclonal antibody (named M10) is a simple and reliable diagnostic method in overt infection cases of RSIV (Nakajima and Sorimachi, 1995; Nakajima et al, 1995). OIE manual (2012) recommends spleen as the most appropriate organ in the IFAT. Many PCR and nested PCR methods have been reported to identify and detect megalocytiviruses (Kurita et al, 1998; He et al, 2001; Chao et al, 2002; Jeong et al, 2003). A real-time PCR assay and LAMP method have been developed for RSIV detection (Caipang et al, 2003, 2004). Water borne route is the principal mode of virus transmission. Virus-harboring trash fish may be hazardous to farmed susceptible fish (Kim et al, 2007). RSIV is inactivated by a treatment at 55°C for 30 min, and ISKNV is susceptible to some disinfectants (Nakajima and Sorimachi, 1994; He et al, 2002). The effectiveness of formalin-killed vaccine by intraperitoneal injection was demonstrated in red sea bream under laboratory and field settings (Nakajima et al, 1997, 1999). The inactivated vaccine by injection method has been licensed in Japan since 1999, and to date is available for red sea bream, striped jack (Pseudocaranx dentex), Malabar grouper (E. malabaricus), orange-spotted grouper (E. coioides) and fish species of the genus Seriola. However, the vaccine is not effective to protect fish of the genus Oplegnathus from RSIVD. RSIVD is of great importance for aquacultures particularly in East and Southeast Asian countries, and there still remain many subjects to be studied in the future. 2.4. Viral Nervous Necrosis (VNN) Viral nervous necrosis (VNN), also known as viral encephalopathy and retinopathy (VER), is one of the most important limiting factors in successful seed production in marine aquaculture worldwide, because of its wide range of host species and severe mortalities, often up to 100%. The disease was first described in 1990 in hatchery-reared Japanese parrotfish (Oplegnathus fasciatus) in Japan and Asian sea bass (barramundi, Lates calcarifer) in Australia (Yoshikoshi and Inoue, 1990; Glazebrook et al, 1990). In the following 1991-1992 years, the disease was documented in larvae of turbot (Scophthalmus maximus) in Norway, European sea bass (Dicentrarchus labrax) in France and striped jack (Pseudocarnx dentex) in Japan, and in larvae/juveniles of red-spotted grouper (Epinephelus akaara) in Japan (Bloch et al, 1991; Breuil et al, 1991 ; Mori et al, 1991 ; Mori et al, 1992). Although the disease is most serious during hatchery-rearing period, several fish species such as sevenband grouper, European sea bass, Atlantic halibut (Hippoglossus hippoglossus) and Atlantic cod (Gadus morhua) remain susceptible to the causative virus through older stages (Fukuda et al, 1996; Le Breton et al, 1997 ; Aspehaug et al, 1999 ; Patel et al, 2007). The number of host fish species of VNN is still increasing, to date reached more than 40 species (24 families and 8 orders). Some instances of the disease in
292

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
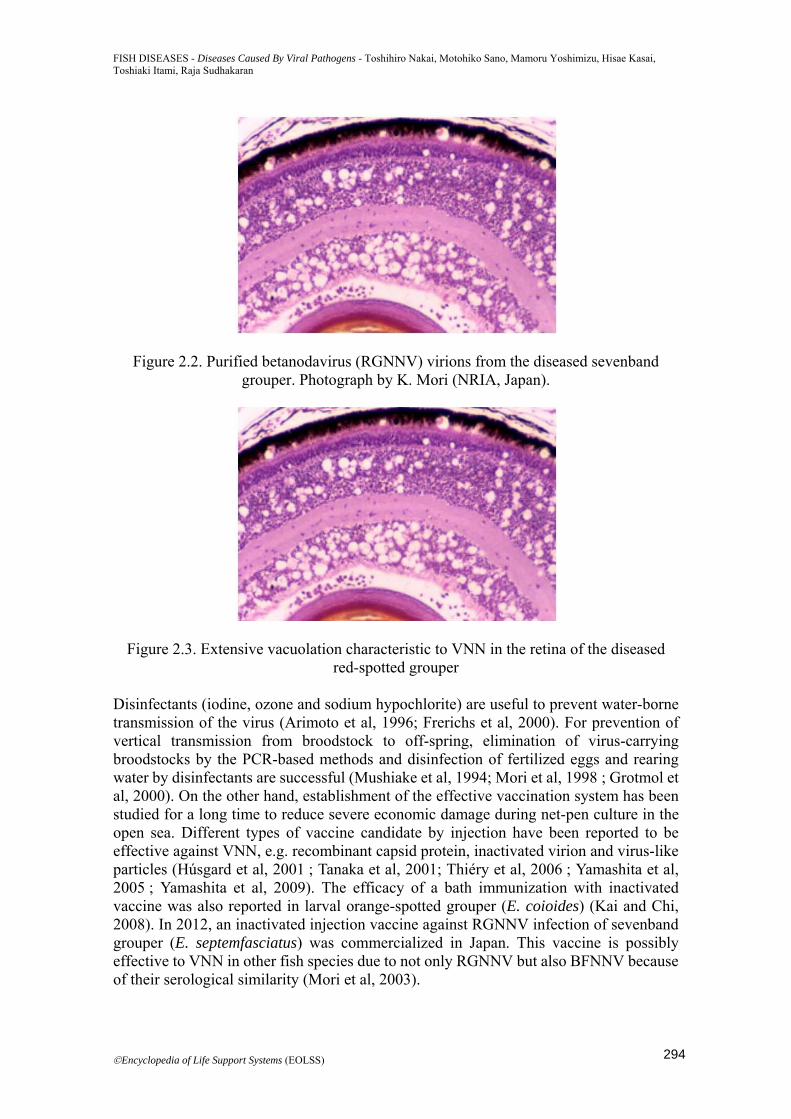
freshwater fish species or freshwater farms have been recorded. No overt VNN has been observed in salmonids, though Atlantic salmon (Salmo salar) is susceptible to experimental infection (Korsnes et al, 2005). Some review papers have been published for VNN and the causative agents (Munday et al, 2002; Sano et al, 2011). The causative agent of VNN was first characterized in diseased larval striped jack (Pseudocaranx dentex), named as striped jack nervous necrosis virus (SJNNV), and identified as a member of the family Nodaviridae (Mori et al, 1992). Agents in diseased larval Asian sea bass and European sea bass were also purified and identified as nodavirus (Comps et al, 1994; Chi et al, 2001). The current virus taxonomy classifies piscine nodaviruses into the genus Betanodavirus, while the genus Alphanodavirus is for insect nodaviruses (Thiéry et al, 2012). Betanodaviruses are non-enveloped and round-shaped with 25-30 nm in diameter. The genome consists of two positive sense ssRNA molecules; RNA1 (RNA-dependent RNA polymerase gene: 3.1 kb) and RNA2 (coat protein gene: 1.4 kb). Both molecules lack poly (A) tails at their 3’-ends. A subgenomic RNA3 (0.4 kb), which is derived from RNA1 in infected cells, encodes a protein with an RNA silencing-suppression activity. Based on the coat protein gene sequences, betanodaviruses are subgrouped into four major genotypes; designated SJNNV (type species of the genus), TPNNV, RGNNV and BFNNV (Nishizawa et al, 1995, 1997). Most of isolates from diseased fish are either RGNNV genotype or BFNNV genotype. Complete nucleotide genome sequences were reported for SJNNV, RGNNV and BFNNV (Iwamoto et al, 2001; Tan et al, 2001; Sommerset and Nerland, 2004;), and the established infectious RNA transcription system demonstrated that viral RNA2 and/or encoded coat protein control host specificity in SJNNV and RGNNV (Iwamoto et al, 2004). These genotypes are related closely to the serotypes, i.e. serotype A for SJNNV, serotype B for TPNNV and serotype C for both BFNNV and RGNNV (Mori et al, 2003). There are also a close relationship between the genotypes and the host fish from which virus was isolated. The RGNNV has been isolated from warm water fish, while the BFNNV has been isolated from cold water fish, suggesting that water temperature is an important factor influencing disease outbreaks (Iwamoto et al, 1999). Figure 2.2 shows the purified RGNNV virions which were isolated from the diseased sevenband grouper. There are no clinical signs on the body surface and gills of affected fish at any stages. The affected juveniles or older fishes are characterized by a range of erratic swimming behaviors: spiral, whirling movement or belly-up floating with inflation of swim bladder, or lying down at rest. Histopathologically, severely extended necrosis and vacuolation are observed in the affected brain, spinal cord and retina (Figure 2.3). PCR-based methods are most rapid and convenient to diagnose clinically affected fish. For a confirmative diagnosis, however, either histopathology or virus isolation in cell culture followed by immunological procedures (FAT, IHC, ELISA) or sequencing analysis has to be performed. SSN-1 and E-11 cell lines are now officially provided to isolate and propagate any of betanodaviruses (Frerichs et al, 1996; Iwamoto et al, 2000). PCR-based or cell culture isolation method demonstrated that betanodaviruses are frequently detected in asymptomatic cultured and wild marine fish including trash fish (Castric et al, 2001; Barker et al, 2002; Gomez et al, 2004). These subclinically or persistently infected fish might be important as a potential infection source for farmed fish susceptible to the virus.
293
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Figure 2.2. Purified betanodavirus (RGNNV) virions from the diseased sevenband grouper. Photograph by K. Mori (NRIA, Japan).
Figure 2.3. Extensive vacuolation characteristic to VNN in the retina of the diseased red-spotted grouper
Disinfectants (iodine, ozone and sodium hypochlorite) are useful to prevent water-borne transmission of the virus (Arimoto et al, 1996; Frerichs et al, 2000). For prevention of vertical transmission from broodstock to off-spring, elimination of virus-carrying broodstocks by the PCR-based methods and disinfection of fertilized eggs and rearing water by disinfectants are successful (Mushiake et al, 1994; Mori et al, 1998 ; Grotmol et al, 2000). On the other hand, establishment of the effective vaccination system has been studied for a long time to reduce severe economic damage during net-pen culture in the open sea. Different types of vaccine candidate by injection have been reported to be effective against VNN, e.g. recombinant capsid protein, inactivated virion and virus-like particles (Húsgard et al, 2001 ; Tanaka et al, 2001; Thiéry et al, 2006 ; Yamashita et al, 2005 ; Yamashita et al, 2009). The efficacy of a bath immunization with inactivated vaccine was also reported in larval orange-spotted grouper (E. coioides) (Kai and Chi, 2008). In 2012, an inactivated injection vaccine against RGNNV infection of sevenband grouper (E. septemfasciatus) was commercialized in Japan. This vaccine is possibly effective to VNN in other fish species due to not only RGNNV but also BFNNV because of their serological similarity (Mori et al, 2003).
294

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
3. Salmon and Trout Viral Diseases Hisae Kasai and Mamoru Yoshimizu 3.1. Synopsis Transmissible diseases of socio-economic importance must be controlled within national boundaries. Salmon and trout viral diseases such as infectious hematopoietic necrosis (IHN), infectious salmon anemia (ISA) and viral hemorrhagic septicemia (VHS) are listed as diseases notifiable to the World Organization for Animal Health (OIE). Formerly Oncorhynchus masou virus disease was also listed. For prevention and treatment of diseases, see “Prevention and Treatment of Diseases Caused by Fish Pathogens”. 3.2. Introduction Salmonid fish including chum salmon (Oncorhynchus keta), pink salmon (O. gorbuscha), sockeye (O. nerka), Chinook (O. tshawytscha), coho (O. kisutch) and masu salmon (O. masou) are important species that are hatchery reared and released fish in to the North Pacific Ocean. Atlantic salmon (Salmo salar), and rainbow trout (O. mysiss) are important for aquaculture. Viral diseases such as infectious pancreatic necrosis (IPN), infectious hematopoietic necrosis (IHN), viral hemorrhagic septicemia (VHS), Oncorhynchus masou virus disease (OMVD), infectious salmon anemia (ISA), erythrocytic inclusion body syndrome (EIBS), pancreas disease (PD), sleeping disease (SD), viral wiring disease (VWD), and viral erythrocytic necrosis (VEN) limit successful propagation and aquaculture of these species. Here, we discuss the characteristics, genome size, serological classification, molecular classification and pathogenesis of representative viruses, outlined in the OIE manual of diagnostic methods for aquatic animals (OIE Manual of diagnostic tests for aquatic animals), AFS-FHS blue book (2010), and fish diseases and disorders (Patrick and Bruno, 2011). 3.3. Representative Viral Diseases and their Characteristics 3.3.1. Infectious Pancreatic Necrosis Infectious pancreatic necrosis (IPN) is an acute contagious systemic birnavirus disease of fry and fingerling trout. The disease most characteristically occurs in rainbow trout, brook trout (Salvelinus fontinalis), brown trout (Salmo trutta) and Atlantic salmon. However IPNV and IPN-like birnaviruses have been isolated from a variety of non-salmonid fishes and invertebrates from freshwater, estuarine, and marine environments. IPNV (and IPN-like birnaviruses) is among geographically dispersed groups of viruses. 3.3.1.1 Disease Agent IPNV is the type species of the genus Aquabirnavirus within the viral family designated Birnaviridae by the International Committee on Taxonomy of Viruses (ICTV) (King et al, 2012). The IPNV virion is non-enveloped, single shelled and measures approximately 60 nm in diameter. The viral genome consistes of a bi-segmented, double-stranded RNA. Aquatic birnavirus including IPNV is classified into two serogroups (A and B).
295

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Serogroup A isolates are divided into nine serotypes, termed A1-A9. Serogroup B contains only one serotype, but this is based on a limited number of isolates. Aquatic birnavirus including IPNV was classified into seven genogroup by sequence analysis of the VP2/NS junction region. 3.3.1.2 Diagnostic Methods In salmonid fry, symptoms of infection include anorecia and a violent whirling pattern. Non-specific external symptoms include darkening of the skin, abdominal swelling, exophthalmia, pale gills and petechial hemorrhages. IPN exhibits marked pancreatic necrosis and severe lesions in the intestinal mucosa. Pancreatic lesions can vary from small fogi to extensive acinar cell necrosis, with nuclear pyknosis, karyorrhexis and basophilic cytoplasmic inclusions. Virological examination coupled with serological or molecular identification is required for confirmation of clinical or subclinical infection with IPNV. Cell cultures of blue gill fry (BF-2), Chinook salmon embryo (CHSE-214), or rainbow trout gonad (RTG-2) are commonly used for evaluation of clinical materials for detection of IPNV in salmonid fishes. Confirmatory identification of IPNV can be accomplished using immunologic-based or molecular-based assay formats. 3.3.1.3 Control Most IPN epizootics have been linked to the importation of infected eggs or fry. Selection of pathogen free eggs and broodstock is important. IPNV is tolerant to low pH but not alkaline conditions (pH 12). It takes hours to inactivate IPNV using heat, unless the temperature is well above 70°C. Although iodophors are relatively effective against IPNV in vitro, it is insufficient for use as a disinfection procedure for IPNV contaminated eggs. Chemotherapy is not available. Promising vaccines have been tested in field trials, and mixed vaccine against five pathogens including recombinant IPNV protein is commercially available in Norway. 3.3.2. Infectious Hematopoietic Necrosis Infectious hematopoietic necrosis (IHN) is an acute, systemic, and usually virulent rhabdoviral disease that occurs in the wild, but is more typically seen in epizootic proportion among young trout and certain Pacific salmon under husbandry in coastal North America from California to Alaska, Europe, and the Far East. The disease most affects rainbow/ steelhead trout, cutthroat trout (Salmo clarki), brown trout, Atlantic salmon, and Pacific salmon including Chinook, sockeye/ kokanee, chum, masu/ yamame and amago salmon (O. rhodurus). Under experimental conditions, infections have been reported in non-salmonids like pike fry, sea bream and turbot. 3.3.2.1 Disease Agent: IHNV is the type species of the genus Novirhabdovirus within the viral family designated Rhabdoviridae by the ICTV (King et al, 2012). IHNV virion is bullet-shaped and measures 170×70 nm. Its viral genome consists of complementary, single-stranded RNA. IHNV isolates are not classified serologically but can be grouped into three genetic types by sequence analysis of viral glycoprotein gene.
296

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
3.3.2.2 Diagnostic Methods Moribund fish are lethargic, swim high in the water column, and are anorexic. They exhibit exophthalmia, darkening of body color, abdominal distension, pale gills, and hemorrhages at the bases of fins. Fecal casts trailing from the vent have been reported but are not always observed. Necrosis of splenic hematopoietic tissue and of the endocrine and exocrine tissue of the pancreas is diffuse, but the liver can have areas of focal necrosis. Virological examination coupled with serological or molecular identification is required for confirmation of clinical or subclinical infection with IHNV. Cell cultures of BF-2, CHSE-214, RTG-2 or Epithelioma papulosum cyprini (EPC) are commonly used for evaluation of clinical materials for detection of IHNV in salmonid fishes. Confirmatory identification is same as IPNV. 3.3.2.3 Control Most IHN epizootics have been linked to the importation of infected eggs or fry. Eggs should be disinfected with and iodophor solution, and virus-free water should be used to incubate eggs and raise fry (see Prevention and Treatment of Diseases Caused by Fish Pathogens). IHNV is readily inactivated by most common disinfectants. In addition, this virus can be inactivated by drying, or by heating to 60°C for 15minutes. Chemotherapy is not available. Promising vaccines have been tested in field trials, and DNA vaccines are commercially available in Canada. 3.3.3. Viral Hemorrhagic Septicemia Viral hemorrhagic septicemia (VHS) is an important source of mortality for cultured and wild fish in freshwater and marine environments in several regions of the northern hemisphere. Over 60 species of freshwater and marine fish are currently known to be natural hosts of VHSV but rainbow trout are especially susceptible. 3.3.3.1 Disease Agent VHSV is classified to the genus Novirhabdovirus within the viral family designated Rhabdoviridae by the ICTV (King et al, 2012). VHSV virion is bullet-shaped and measures 180-200×75 nm. The viral genome is complementary, single-stranded RNA. VHSV was classified into three serogroups, however there was considerable overlap of strains within and between these serotypes. VHSV can be grouped into four genetic types by sequence analysis. These genotypes show a geographic basis for their distribution rather than host specificity. 3.3.3.2 Diagnostic Methods External clinical signs of disease can include exophthalmia, abdominal distention, darkened coloration, anemia, lethargy, hyperactivity, and hemorrhages in the eyes, skin, gills, and at the base of fins. Internally, visceral mesenteries can show diffuse hemorrhage, the kidneys and liver can be hyperemic, swollen, and discolored, and hemorrhages can occur in skeletal muscle. In the liver, kidney and spleen, focal to extensive necrotic changes can occur. Virological examination is same as IHNV. Confirmatory identification is same as IPNV.
297

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
3.3.3.3 Control Same as IHNV. Promising vaccines have been tested in field trials, but no vaccines are commercially available. 3.3.4. Oncorhynchus Masou Virus Disease Oncorhynchus masou virus disease (OMVD) is an economically significant disease of farmed salmonid fish in Japan. This infection also occurs in wild fish. The disease most characteristically occurs in kokanee, coho, masu salmon and rainbow trout. Surviving salmon often develop cutaneous tumors, particularly around the mouth. 3.3.4.1 Disease Agent OMV is taxonomically known as Salmonid herpesvirus 2; SalHV-2. This virus is classified to the genus salmonivirus within the viral family designated Alloherpesviridae by the ICTV (King et al, 2012). Diameter of the OMV virion is 200-240 nm and the diameter of the icosahedral nucleocapsid formed with 162 capsomeres is 115 nm. The viral genome is double stranded DNA. OMV isolates are not classified serologically. 3.3.4.2 Diagnostic Methods External clinical signs in infected fish include appetent exophthalmia, and petechiae on the body surface, especially beneath the lower jaw. Agonal or abnormal swimming behavior has not been observed. Internally, the liver shows white spot lesions, and in advanced cases the whole liver becomes pearly white. In some cases the spleen is found to be swollen. Necrosis of epithelial cells and kidney were observed in the young one-month old moribund specimens while partial necrosis of the liver, spleen and pancreas was seen in older moribund specimens. Virological examination coupled with serological or molecular identification is required for confirmation of clinical or subclinical infection with OMV. Cell cultures of CHSE-214 or RTG-2 are commonly used for evaluation of clinical materials for detection of OMV in salmonid fishes. Confirmatory identification is same as IPNV. 3.3.4.3 Control Same as IHNV. Chemotherapy using anti-herpesvirus agents is effective but commercially unavailable for fish. Promising vaccines have been tested in field trials, but no vaccines are commercially available. 3.3.5. Infectious Salmon Anemia Infectious salmon anemia (ISA) is one of the most important viral diseases of farmed Atlantic salmon. Sea-run brown trout, rainbow trout, and Atlantic herring (Clupea harengus) are potential asymptomatic carriers of the virus. Infectious salmon anemia has been found in Norway, the Faroe Islands, and the United Kingdom (Scotland and Shetland Islands). Infectious salmon anemia outbreaks occur periodically in Norway and Chile, as well as in a limited region of North America shared by the U.S. and Canada.
298

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
3.3.5.1 Disease Agent ISAV is classified as the type species of the genus Isavirus within the viral family designated Orthomyxoviridae by the ICTV (King et al, 2012). ISAV is a pleomorphic enveloped virus, 100-130 nm in diameter, with 10-12 nm surface projections. The viral genome consists of eight single-stranded RNA segments of negative polarity. ISAV has been divided into two major clusters or genotypes, called the North American and the European, and the third genotype may exist. 3.3.5.2 Diagnostic Methods ISA should be a consideration in Atlantic salmon with increased mortality and signs of anemia, or lesions consistent with this disease. It should always be investigated if the hematocrit is less than 10%. External clinical signs in infected fish are pale gills and muscle, petechial hemorrhage, dark liver and spleen, ascites are all present. The presence of renal interstitial hemorrhaging, and tubular epithelial degeneration, necrosis, and casting within the posterior kidney have all been noted. Liver sections show multifocal to confluent hemorrhagic hepatic necrosis, focal congestion, and dilatation of hepatic sinusoids. 3.3.5.3Control Same as IHNV. Mixed vaccines are commercially available in Norway. 6) Other diseases Erythrocytic inclusion body syndrome (EIBS), pancreas disease (PD), sleeping disease (SD), viral wiring disease (VWD) and viral erythrocytic necrosis (VEN) are reported. In case of EIBS, erythrocytic inclusions and typical virus particles have been detected in spawned adult salmon, but in fresh water the disease seems to be limited to juvenile and smolting fish. In salt water, EIBS has been diagnosed in fish as large as 500 g and has caused problems in netpen culture. Naturally occurring infections have been demonstrated in coho, Chinook and Atlantic salmon. The agent that causes EIBS has neither been isolated nor fully characterized. PD and SD are infectious diseases of farmed Atlantic salmon and rainbow trout, respectively. Salmonid alphaviruses are recognized as pathogen. The main pathological lesions are similar in PD and SD, with extensive loss of pancreatic acinar cells during the viraemic phase, concurrent cardiomyocytic degeneration and inflammation and subsequent skeletal muscle degeneration and fibrosis. VWD was first recognized in coho salmon in Japan (Oh et al, 1995). VEN is a condition characterized by the presence of viroplasmic inclusion bodies within the cytoplasm of affected erythrocytes. The causative agent is tentatively placed in the Iridoviridae. 4. Shrimp Diseases Toshiaki Itami and Raja Sudhakaran 4.1. Synopsis Penaeid shrimp culture is an important industry in Asia and Americas. The total annual production in 2010 reached over 3.7 million tons in the world (FAO, 2012). However,
299

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
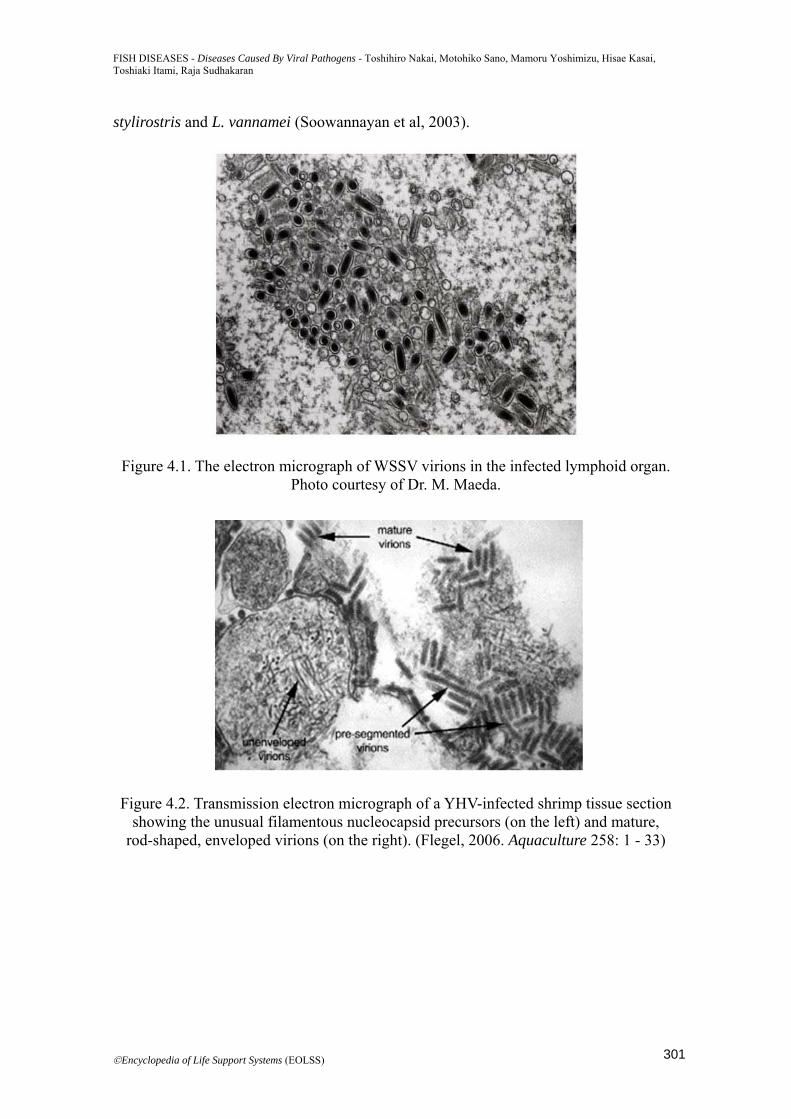
diseases, especially the viral diseases, are being a major negative factor to this industry apart from the bacterial, fungal diseases and parasites. Therefore, six important viral diseases are reviewed in this section. 4.2. Introduction The shrimp culture industries have been spreading worldwide since the 1970s, originally starting from Japan. With their expansion, various viral and bacterial diseases have been occurring and causing serious damage to the shrimp culture industries worldwide. Vibrio penaeicida infection spread from the late 1980s, and then after 1990s, major diseases in shrimp aquaculture changed from bacterial diseases to viral diseases. White spot syndrome (WSS) which is viral disease and has high mortality rate has spread in shrimp culture in Asia and in central and south Americas (Lightner et al, 2012; Lightner2011; Flegel, 2012). The World Animal Health Organization (the OIE: Office International des Epizooties) lists 6 virus diseases out of 8 diseases of crustacean including fungal and bacterial diseases (Table 4.1) (OIE, 2012). In this section, white spot syndrome, yellow head disease, infectious hypodermal and hematpoieitc necrosis, Taura syndrome, white tail disease and infectious myonecrosis are discussed which are damaging shrimp aquaculture industries. 4.3. Disease Agent (Characteristics, Genome Size, Serological Classification, Molecular Classification, Pathogenesis) 4.3.1 White Spot Syndrome The causative agent of this disease is white spot syndrome virus (WSSV). WSSV is a large (80-120 x 250-380 nm), enveloped, double-stranded DNA virus and was assigned to a new genus Whispovirus, and family Nimaviridae (Mayo, 2002,2005) (Figure 4.1). Virions show tail-like appendages. The size of genome of this virus is ~300 kb: 305,107 bp (GenBankaccession No.AF332093), 292,967 bp (AF369029) and 307,287 bp (AF440570). Virulence is quite high and the mortality reaches up to 80-90 % in 3-10 days in juvenile shrimp. WSSV has a wide host range among over 70 species of crustaceans including shrimp, crabs and crayfish (Flegel, 2006). 4.3.2 Yellow Head Disease The causative agent of this disease is yellow head virus (YHV, genotype 1), gill-associated virus (GAV, genotype 2) (Spann et al, 1997) and other related viruses known as genotype 3-6 (OIE, 2012 Mayo, 2002; Walker et al, 2005). YHV, the genus Okavirus in the family of Roniviridae, is enveloped, rod-shaped (40-60×150-200nm), single stranded RNA virus with a genome of ~27 kb (Figure 4.2). YHV (genotype 1) was observed mainly in Southeast Asian countries and Mexico, and GAV and other genotypes were found in Australia, Mozambique and Southeast Asia (Wijegoonawardane et al, 2008). Black tiger shrimp (Penaeus monodon) is the major susceptible host species and shows the high and rapid mortality. Natural infections were reported in other penaeid shrimp, e.g. Marsupenaeus japonicus, Fenneropenaeus merguiensis, Litopenaeus
300
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
stylirostris and L. vannamei (Soowannayan et al, 2003).
Figure 4.1. The electron micrograph of WSSV virions in the infected lymphoid organ. Photo courtesy of Dr. M. Maeda.
Figure 4.2. Transmission electron micrograph of a YHV-infected shrimp tissue section showing the unusual filamentous nucleocapsid precursors (on the left) and mature,
rod-shaped, enveloped virions (on the right). (Flegel, 2006. Aquaculture 258: 1 - 33)
301
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
causative agents susceptible species, stage and target organ Disease name
virus name taxon species stage target organ/tissue white spot syndrome, white spot disease
WSSV, WSDV family Nimaviridae genus Whispovirus
penaeid shrimp PL - adults ectodermal and mesodermal origin
yellow head disease YHV (gill-associated virus: GAV)
family Roniviridae genus Okavirus
penaeid shrimp PL15-adults ectodermal and mesodermal origin
infectious hypodermal and hematopoietic necrosis
IHHNV genus Brevidensovirus family Parvoviridae
penaeid shrimp egg - adults ectodermal and mesodermal origin
Taura syndrome TSV family Dicistroviridae genus Aparavirus
penaeid shrimp PL - adults cuticular epithelium (the general exoskeleton, foregut, hindgut, gills and appendages), connective tissues, haematopoietic tissues, lymphoid organ, antennal gland
white tail disease MrNV XSV
MrNV: family NodaviridaeXSV: extra small virus
Macrobrachium rosenbergii
larvae - PL - juveniles
gill tissue, head muscle, heart, abdominal muscle, ovaries, pleopods tail muscle
infectious myonecrosis IMNV
family Totiviridae(closely related to Giardia lamblia virus)
penaeid shrimp juveniles - subadults
striated muscles, connective tissues, haemocytes, lymphoid organ
Table 4.1. OIE listed virus diseases in shrimp as of 2012 (OIE, 2012) 302

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
4.3.3 Infectious Hypodermal and Hematpoieitc Necrosis The causative agent of this disease is infectious hypodermal and hematpoieitc necrosisvirus (IHHNV). This virus was assigned to the genus Brevidensovirus, family Parvoviridae. IHHNV is a 20-22 nm, non-enveloped icosahedron, containing single-stranded DNA with estimated size of 3.9 kb (Lightner2011; OIE, 2012; Lightner et al, 1983; Bonami and Lightner, 1991; Bonami et al, 1990 ; Rai et al, 2011). IHHNV has at least three genotypes: IHHNV-I from Americas/Philippines, IHHNV-II from Southeast Asia and IHHNV-III from East Africa and Australia (Lightner et al, 2012). The most penaeid shrimp show the susceptibility to IHHNV-I and IHHNV-II, and principal hosts are L. stylirostris, L. vannamei and P. monodon. IHHNV causes the acute epizootics and mass mortality in L.stylirostris (>90%), while this virus causes runt-deformity syndrome (RDS) and reduced, irregular growth, rather than heavy mortalities, in L. vannamei (Kalagayan et al, 1991). P. monodon usually show the subclinical infection, however IHHNV-infected shrimp have been reported to show RDS and low growth rate, causing economic damage (Chayaburakul et al, 2005; Primavera et al, 2000). 4.3.4 Taura Syndrome The causative virus Taura syndrome virus (TSV) is 32 nm diameter and nonenveroped icosahedron (Figure 4.3). The genome consists of a linear single-stranded RNA of 10.2 kb. TSV has at least four genotypes: the Americas group, Southeast Asian group, the Belize group and the Venezuelan group (OIE, 2012; Wertheim et al, 2009). TSV has been assigned to the newly created genus Aparavirus in new family Dicistroviridae in the order of Picornavirales in the 9th report of the ICTV (Lightner et al, 2012; Chen et al, 2012). Major hosts of this virus are L. vannameiandL.stylirostrisand other penaeid species can be infected with TSV by experimental challenge.
Figure 4.3. TSV by TEM. The electron micrograph above shows TSV in the cytoplasm by normal TEM using thin tissue sections from TSV infected P. vannamei. The upper inset
micrograph shows purified TSV virions by negative staining. (Brock et al., 1997). (Flegel, 2006. Aquaculture 258: 1 - 33)
303

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
4.3.5 White Tail Disease The causative agents of this disease are two viral pathogens: Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV) (Qian et al, 2003). MrNV is necessary for the outbreak of WTD in M. rosenbergii but the role of XSV is not clear, known as satellite virus. MrNV is a non-envelped, icosahydron (26-27 nm diameter), consisting of 2.9 kb and 1.26 kb single-stranded RNA (Figure 4.4) . XSV is a non-envelped, icosahydron (15nm diameter), consisting of 0.9 kb single-stranded RNA (Figure 4.4) (Bonami et al, 2005; Wang et al, 2008). The principal host species is M.rosenbergii, giant freshwater prawn, and are a few other host reported (Sudhakaran et al, 2006a; Sudhakaran et al, 2006b; Bonami and Widada, 2011). Severe mortality is observed in larvae and post larvae stages of M.rosenbergii and adults become resistant carriers (Sahul Hameed et al, 2004; Sudhakaran et al, 2006).
Figure 4.4. Purified XSV (a) and MrNV (b) particles after the CsCl gradient. TEM. PTA 2%. Bars =100 nm. Inset in b: higher magnification of MrNV; bar= 50 nm. (Bonami and
Widada, 2011. Journal of Invertebrate Pathology 106:131 - 142)
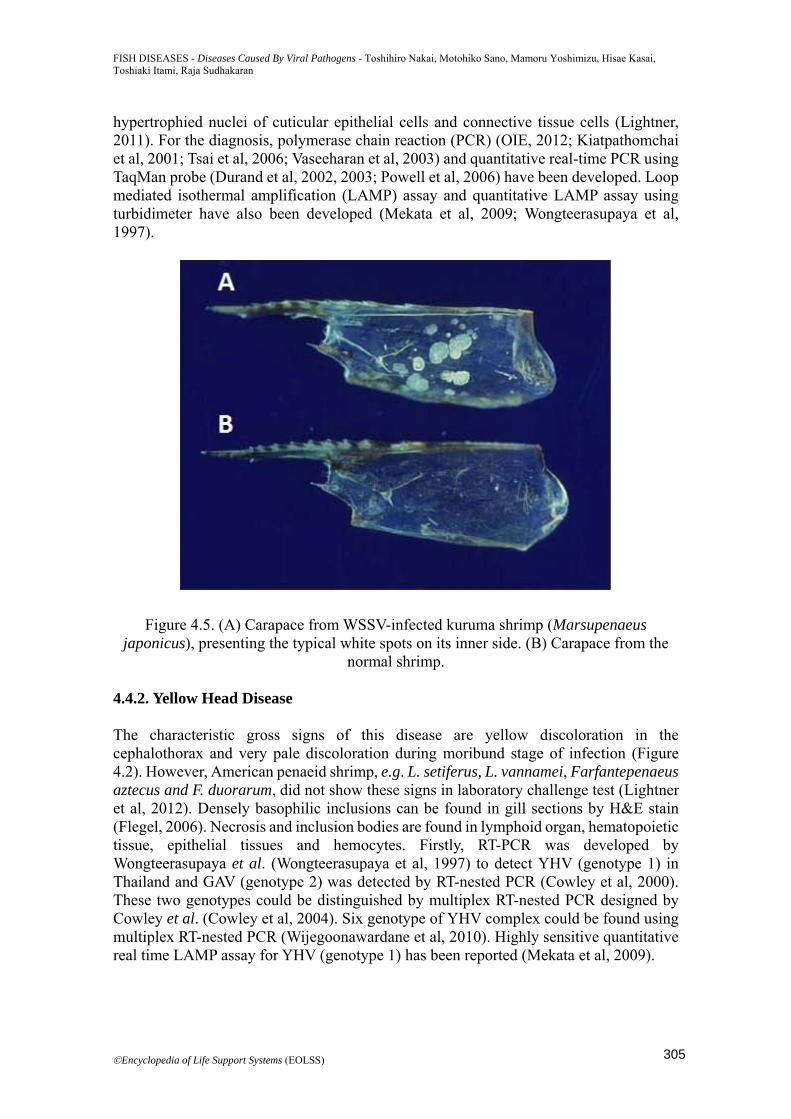
4.3.6 Infectious Myonecrosis Infectious myonecrosis virus (IMNV) is icosahedron (40 nm in diameter) having double stranded RNA of 7.56 kb as genetic material. IMNV is closely related to genus Giardiavirus, family Totiviridae (Poulos et al, 2006). The principal host species is L. vannamei, L.stylirostris and P. Monodon is susceptible by experimental infection (Tang et al, 2005). Juveniles and sub-adults of L. vannamei are most severely infected by IMNV (Lightner, 2011). 4.4. Diagnostic Methods 4.4.1. White Spot Syndrome The clinical sign of this disease is observation of typical white spots (0.5-2.0 mm in diameter) on the carapace (Figure 4.5). Moribund shrimp show a pinkish or reddish coloration of the exoskeleton. Typical histopathology of this disease is presence of eosinophilic to pale basophilic, Feulgen-positive intranuclear inclusion bodies in
304
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
hypertrophied nuclei of cuticular epithelial cells and connective tissue cells (Lightner, 2011). For the diagnosis, polymerase chain reaction (PCR) (OIE, 2012; Kiatpathomchai et al, 2001; Tsai et al, 2006; Vaseeharan et al, 2003) and quantitative real-time PCR using TaqMan probe (Durand et al, 2002, 2003; Powell et al, 2006) have been developed. Loop mediated isothermal amplification (LAMP) assay and quantitative LAMP assay using turbidimeter have also been developed (Mekata et al, 2009; Wongteerasupaya et al, 1997).
Figure 4.5. (A) Carapace from WSSV-infected kuruma shrimp (Marsupenaeus japonicus), presenting the typical white spots on its inner side. (B) Carapace from the
normal shrimp. 4.4.2. Yellow Head Disease The characteristic gross signs of this disease are yellow discoloration in the cephalothorax and very pale discoloration during moribund stage of infection (Figure 4.2). However, American penaeid shrimp, e.g. L. setiferus, L. vannamei, Farfantepenaeus aztecus and F. duorarum, did not show these signs in laboratory challenge test (Lightner et al, 2012). Densely basophilic inclusions can be found in gill sections by H&E stain (Flegel, 2006). Necrosis and inclusion bodies are found in lymphoid organ, hematopoietic tissue, epithelial tissues and hemocytes. Firstly, RT-PCR was developed by Wongteerasupaya et al. (Wongteerasupaya et al, 1997) to detect YHV (genotype 1) in Thailand and GAV (genotype 2) was detected by RT-nested PCR (Cowley et al, 2000). These two genotypes could be distinguished by multiplex RT-nested PCR designed by Cowley et al. (Cowley et al, 2004). Six genotype of YHV complex could be found using multiplex RT-nested PCR (Wijegoonawardane et al, 2010). Highly sensitive quantitative real time LAMP assay for YHV (genotype 1) has been reported (Mekata et al, 2009).
305

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Figure 4.6. Gross signs of yellow head infection are seen here in the 3 shrimp on the right. They are generally bleached in color with a yellowish discoloration of the cephalothorax
region when compared to shrimp of normal appearance on the left. (Flegel, 2006. Aquaculture 258:1 - 33)
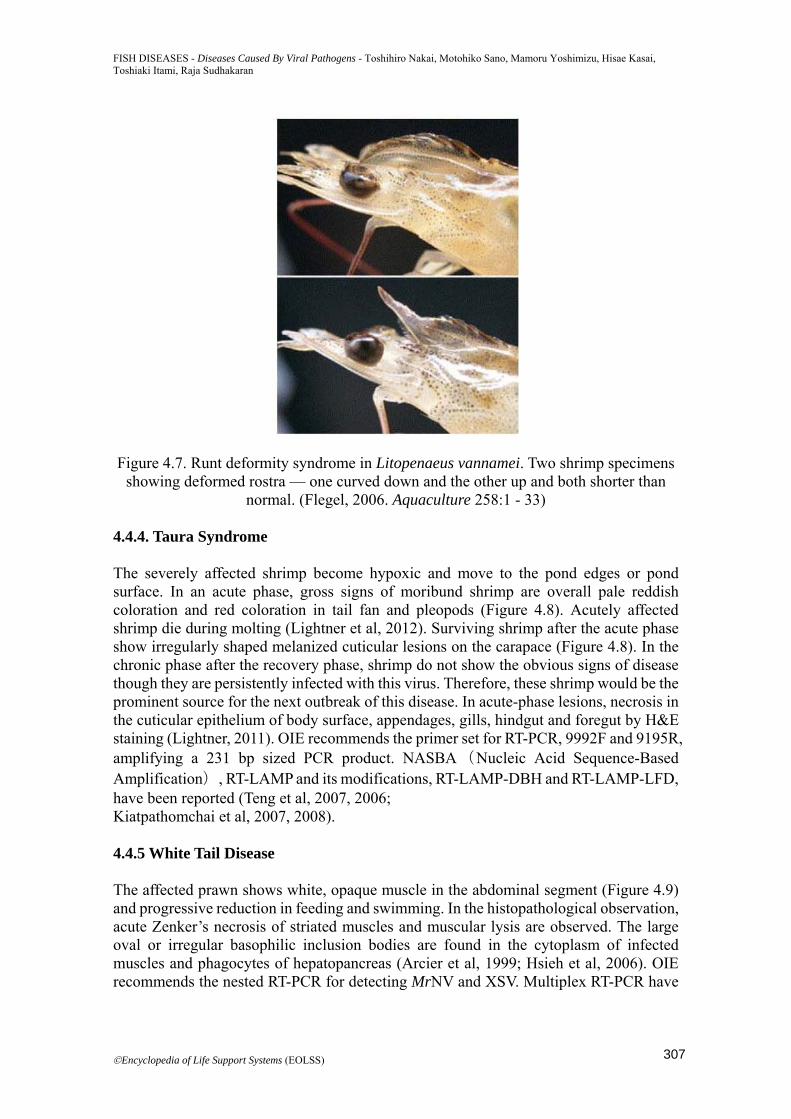
4.4.3. Infectious Hypodermal and Hematpoieitc Necrosis The gross signs of this disease in L.stylirostris are lethargic surface swimming, inverted sinking to the bottom and lack of mobility (Vega-Heredia et al, 2012). In L. vannamei, runt-deformity syndrome (RDS) and reduced, irregular growth can be found (Figure 4.7) (Kalagayan et al, 1991). However, high mortality and significant reduced or irregular growth have not been observed in P. monodon (Chayaburakul et al, 2005). Cowdry type A intranuclear inclusions are found in nerve cord, gills, cuticular hypodermis, haematopoietic tissues and connective tissues (Lightner, 1996). Aquatic Animal Health Manual (OIE, 2012) recommends the primer sets for PCR detection of IHHNV. Quantitative real-time PCR using TaqMan probe and SYBR have been developed (Tang and Lightner, 2001; Yue et al, 2006 ; Dhar et al, 2001). Qualitative (Sun et al, 2006) and quantitative LAMP assay (Sudhakaran et al, 2008), multiplex LAMP assay (He et al, 2011) and LAMP-LFD (chromatographic lateral flow dipstick) (Arunrut et al, 2011) assays have been reported.
306
FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Figure 4.7. Runt deformity syndrome in Litopenaeus vannamei. Two shrimp specimens showing deformed rostra — one curved down and the other up and both shorter than
normal. (Flegel, 2006. Aquaculture 258:1 - 33) 4.4.4. Taura Syndrome The severely affected shrimp become hypoxic and move to the pond edges or pond surface. In an acute phase, gross signs of moribund shrimp are overall pale reddish coloration and red coloration in tail fan and pleopods (Figure 4.8). Acutely affected shrimp die during molting (Lightner et al, 2012). Surviving shrimp after the acute phase show irregularly shaped melanized cuticular lesions on the carapace (Figure 4.8). In the chronic phase after the recovery phase, shrimp do not show the obvious signs of disease though they are persistently infected with this virus. Therefore, these shrimp would be the prominent source for the next outbreak of this disease. In acute-phase lesions, necrosis in the cuticular epithelium of body surface, appendages, gills, hindgut and foregut by H&E staining (Lightner, 2011). OIE recommends the primer set for RT-PCR, 9992F and 9195R, amplifying a 231 bp sized PCR product. NASBA(Nucleic Acid Sequence-Based Amplification), RT-LAMP and its modifications, RT-LAMP-DBH and RT-LAMP-LFD, have been reported (Teng et al, 2007, 2006; Kiatpathomchai et al, 2007, 2008). 4.4.5 White Tail Disease The affected prawn shows white, opaque muscle in the abdominal segment (Figure 4.9) and progressive reduction in feeding and swimming. In the histopathological observation, acute Zenker’s necrosis of striated muscles and muscular lysis are observed. The large oval or irregular basophilic inclusion bodies are found in the cytoplasm of infected muscles and phagocytes of hepatopancreas (Arcier et al, 1999; Hsieh et al, 2006). OIE recommends the nested RT-PCR for detecting MrNV and XSV. Multiplex RT-PCR have
307

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
been developed by Yoganandhan et al. (Yoganandhan et al, 2005) and Tripathy et al. (Haridas et al, 2010) to detect both viruses at the same time. RT-LAMP with loop-primer (Phillai et al, 2006; Puthawibool et al, 2010; Haridas et al, 2010) and real-time RT-PCR with TaqMan probe (Zhang et al, 2006) have been established.
Figure 4.8. Gross signs of Taura syndrome. On the left is a tail fan of L. vannamei with reddish necrotic areas (arrow). The right photo shows black lesions in the cuticle
characteristic of the recovery stage of TSV infection (Lightner, 1996). (Flegel, 2006. Aquaculture 258:1 - 33)
Figure 4.9. Postlarvae of M. rosenbergii heavily infected with WTDV, showing whitish coloration of the muscle of head part.
4.4.6 Infectious Myonecrosis The gross signs are extensive white necrotic areas in striated muscles in distal abdominal segments and tail fan (Figure 4.10). Lymphoid organs show hypertrophy that is 2-4 times
308

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
their normal size. In histopathological features, IMN shows the characteristic coagulative necrosis of striated muscle fibers, accompanied by infiltration and accumulation of hemocytes (Lightner, 2011). OIE recommends the nested RT-PCR (Poulos et al, 2006). Real-time RT-PCR using TaqMan probe and RT-LAMP have been developed (Andrade et al, 2007; Andrade et al, 2009).
Figure 4.10. Gross signs of infectious myonecrosis in naturally infected farmed L. vannamei, exhibiting various degrees of skeletal muscle necrosis, visible as an opaque,
whitish discoloration of the abdomen. Source: DV Lightner (http://library.enaca.org/Health/FieldGuide/html/cv045im.htm#)
4.5. Control and Recent Topics (Prevention, Chemotherapy, Vaccine) Routine virus monitoring by PCR or other detecting methods would help the shrimp farmers to reduce the mortality and to avoid the disease outbreak by partial harvest. This can be achieved standardizing the stocking density and by feeding the additional vitamins and immunostimulants to enhance the immune response. In the laboratory experiment level, DNA vaccine and recombinant vaccine for WSSV have been demonstrated to be effective to control this disease. WSSV has five major proteins: VP28, VP26, VP24, VP19 and VP15. VP28 and VP19 are the envelope protein and others are nucleocapsid-associated proteins (Van Hulten et al, 2000; Van Hulten et al, 2000). DNA vaccines contracted by the plasmids encoding VP28 and/or VP281 showed the significant level of protection against WSSV artificial challenge test when the DNA vaccine was injected. Protection by DNA vaccination lasts till 7 weeks after the vaccination whereas the recombinant protein vaccination failed to protect after 3 weeks (Rout et al, 2007; Rajesh Kumar et al, 2008; Kono et al, 2009). The oral delivery of DNA construct containing VP28 gene of WSSV encapsulated in chitosan nanoparticles were effective to control this disease (Rajesh Kumar et al, 2009).
309

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Oral administration of recombinant proteins, rVP28 or rVP26, were effective for prophylaxis of this disease. Although the duration of the vaccine is relatively short (within 55-75 days in this experiments), the booster administration of the homologous vaccine could extend its duration (Satoh et al, 2009, 2008; Ning et al, 2011). An application of RNA interference technology against WSSV are promising and effective antiviral strategy. Sequence-specific viral inhibition in shrimp by long synthetic dsRNAs or siRNAs was highly efficient in inhibiting WSSV gene expression (Sarathi et al, 2008; Shekhar and Lu, 2009; Xu et al, 2007; Kim et al, 2007; Sudhakaran et al, 2010) and YHV gene expression (Tirasophon et al, 2007). Since 2009, a newly emerging disease has been occurring in China, Vietnam (estimated loss: 75 million US dollar) and Thailand. This disease is characterized by massive degeneration of the hepatopancreas but the causative agent has not been identified yet. D.V. Lightner (University of Arizona) has named this condition acute hepatopancreatic necrosis syndrome (AHPNS). Close attention should be paid on the spread and impact of this disease on shrimp culture production (Flegel, 2012). Glossary WSSV : White spot syndrome virus
YHV : Yellow head virus
IHHNV : Infectious hypodermal and hematopoietic necrosis virus
TSV : Taura syndrome virus
WTD : White tail disease
MrNV : Macrobrachium rosenbergii nodavirus
XSV : Extra small virus
IMNV : Infectious myonecrosis virus
RT-PCR : Reverse transcription polymerase chain reaction
LAMP : Loop mediated isothermal amplification
RT-LAMP : Reverse transcription loop mediated isothermal amplification
H&E staining
: Hemotoxylin and eosin staining
Bibliography Bibliography (Section 1. Freshwater Fish)
Wolf K. (1988). Fish Viruses and Fish Viral Diseases, 476 pp. Ithaca, NY, USA.
Sano M., Nakai N., Fijan N. (2011). Viral diseases and agents of warm-water fish. In Fish Diseases and Disorders: Volume 3. Woo, P.T.K. and Bruno, D.W. (eds), pp 166-244. Oxfordshire, UK.
Aquatic Animal Health Code 2012, http://www.oie.int/international-standard-setting/aquatic-code/access-online/
310

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Fijan N., Petrinec Z., Sulimanović D., Zwillenberg L.O. (1971). Isolation of the viral causative agent from the acute form of infectious dropsy of carp. Veterinarski Arhiv 41, 125-138.
Ahne W., Bjorklund H.V., Essbauer S., Fijan N., Kurath G., Winton J.R. (2002). Spring viremia of carp (SVC). Diseases of Aquatic Organisms, 52, 261-272.
Walker P.J., Winton J.R. (2010). Emerging viral diseases of fish and shrimp. Veterinary Research 41, 51.
OIE (2013). Manual of Diagnostic Tests for Aquatic Animals 2013. http://www.oie.int/international-standard-setting/aquatic-manual/access-online/
Stone D.M., Ahne W., Denham K.L., Dixon P.F., Liu C.T., Sheppard A.M., Taylor G.R. Way K. (2003). Nucleotide sequence analysis of the glycoprotein gene of putative spring viraemia of carp virus and pike fry rhabdovirus isolates reveals four genogroups. Diseases of Aquatic Organisms 53, 203-210.
Wolf K., Darlington R.W. (1971). Channel catfish virus: a new herpesvirus of ictalurid fish. Journal of Virology 8, 525-533.
Plumb J.A. (1989). Channel catfish herpesvirus. In Viruses of Lower Vertebrates. pp 198-216. Berlin, Germany.
Davison A.J. (1992). Channel catfish virus: a new type of herpesvirus. Virology 186, 9-14.
Camus A.C. (2004). Channel catfish virus disease. Southern Regional Aquaculture Center Publication No. 4702.
Hanson L. Dishon A, Kotler M. (2011). Herpesviruses that infect fish. Viruses 3, 2160-2191.
Waltzek T.B., Kelley G.O., Alfaro M.E., Kurobe T., Davison A.J. Hedrick R.P. (2009). Phylogenetic relationships in the family Alloherpesviridae. Diseases of Aquatic Organisms 84, 179-194.
Hedrick R.P., Gilad O., Yun S., Spangenberg J.V., Marty G.D., Nordhausen R.W., Kebus M.J., Bercovier H., Eldar A. (2000). A herpesvirus associated with mass mortality of juvenile and adult koi, a strain of common carp. Journal of Aquatic Animal Health 12, 44–57.
Haenen O.L.M., Way K., Bergmann S.M., Ariel E. (2004). The emergence of koi herpesvirus and its significance to European aquaculture. Bulletin of the European Association of Fish Pathologists 24, 293–307.
Haenen O.L.M, Hedrick R.P. (2006). Koi herpesvirus workshop. Bulletin of the European Association of Fish Pathologists 26, 26–37.
Michel B., Fournier G,, Lieffrig F,, Costes B,, Vanderplasschen A. (2010). Cyprinid herpesvirus 3. Emerging infectious diseases 16, 1835-1843.
Kurita J., Yuasa K., Ito T., Sano M., Hedrick R.P., Engelsma M.Y., Haenen O.L.M., Sunarto A., Kholidin E.B., Chou H-Y., Tung M.-C., de la Peña L., Lio-Po G., Tu C., Way K., Iida T. (2009). Molecular epidemiology of koi herpesvirus Fish Pathology 44, 59-66.
Jung S.J., Miyazaki T. (1995). Herpesviral haematopoietic necrosis of goldfish, Carassius auratus (L.). Journal of Fish Diseases 18, 211-220.
Waltzek T.B., Kurobe T., Goodwin A.E., Hedrick R.P.. (2009). Development of a polymerase chain reaction assay to detect cyprinid herpesvirus 2 in goldfish. Journal of Aquatic Animal Health 21, 60-67.
Goodwin A.E., Merry G.E., Sadler J.. (2006). Detection of the herpesviral hematopoietic necrosis disease agent (Cyprinid herpesvirus 2) in moribund and healthy goldfish: validation of a quantitative PCR diagnostic method. Diseases of Aquatic Organisms 69, 137-143.
Li X., Fukuda H. (2003). In vitro culture of goldfish cell sensitive to goldfish herpesvirus. Journal of Shanghai Fisheries University 12, 12-18.
Goodwin A.E., Sadler J., Merry G.E., Marecaux E.N.. (2009). Herpesviral haematopoietic necrosis virus (CyHV-2) infection: case studies from commercial goldfish farms. Journal of Fish Diseases 32, 271-278.
Jeffery K.R., Bateman K., Bayley A., Feist S.W., Hulland J., Longshaw C., Stone D., Woolford G., Way K. (2007). Isolation of a cyprinid herpesvirus 2 from goldfish, Carassius auratus (L.), in the UK. Journal of Fish Diseases 30, 649-656.
311

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Wang L., He J., Liang L., Zheng X., Jia P., Shi X., Lan W., Xie J., Liu H., Xu P. (2012). Mass mortality caused by Cyprinid Herpesvirus 2 (CyHV-2) in Prussian carp (Carassius gibelio) in China. Bulletin of the European Association of Fish Pathologists 32, 164-173.
Langdon J.S., Humphrey J.D., Williams L.M., Hyatt A.D., Westbury, H.A. (1986). First virus isolation from Australian fish: iridovirus-like pathogen from redfin perch, Perca fluviatilis. Journal of Fish Diseases 9, 263-268.
Chinchar V.G. (2002). Ranaviruses (family Iridoviridae): emerging cold-blooded killers. Archive Virology 147, 447-470.
Chinchar V.G., Hyatt A., Miyazaki T., Williams, T. (2009). Family Iridoviridae: poor viral relations no longer. Current Topics in Microbiology and Immunology 328, 123-170.
Whittington R.J,, Becker J.A,, Dennis M.M. (2010). Iridovirus infections in finfish - critical review with emphasis on ranaviruses. Journal of Fish Diseases 33, 95-122.
Jancovich J.K., Bremont M., Touchman J.W., Jacobs B.L.. (2010). Evidence for multiple recent host species shifts among the Ranaviruses (family Iridoviridae). Journal of Virology 84, 2636-2647.
Bibliography (Section 2. Marine fish)
Liao, I.C. (2009) The role of aquaculture in upcoming food crisis. Fish Pathol., 44, 1-8.
Iida Y., Masumura K., Nakai T., Sorimachi M., Matsuda H. (1989) A viral disease in larvae and juveniles of the Japanese flounder, Paralichthys olivaceus. Journal of Aquatic Animal Health 1, 7-12.
Iida Y., Hiroi J., Namba K., Nakai T. (2008) Dysfunction in respiration and osmotic regulation of larval Japanese flounder affected by viral epidermal hyperplasia. Fish Pathology 43, 72-78.
Whittington R.J., Jones J.B., Hine P.M., Hyatt A.D. (1997) Herpesvirus that caused epizootic mortality in 1995 and 1998 in pilchard, Sardinops sagax neopilchardus (Steindachner), in Australia is now endemic. Diseases of Aquatic Organisms 28, 1-16.
Hyatt A.D., Hine P.M., Jones J.B., Whittington R.J., Kearns C., Wise T.G., Crane M.S., Williams L.M. (1997) Epizootic mortality in the pilchard Sardinops sagax neopilchardus in Australia and New Zealand in 1995. II. Identification of a herpesvirus within the gill epithelium. Diseases of Aquatic Organisms 28, 17-29.
Smail D.A., Snow M. (2011) Viral haemorrhagic septicaemia. In Fish Diseases and Disorders: Volume 3. Woo P.T.K. and Bruno D.W. (eds), pp. 110-142, CABI, Oxfordshire, UK.
Sorimachi M., Hara T. (1985) Characteristics and pathogenicity of a virus isolated from yellowtail fingerlings showing ascites. Fish Pathology 19, 231-238.
Castric J., Baudin-Laurencin G., Coustans M.F., Auffret M. (1987) Isolation of infectious pancreatic necrosis virus, Ab serotype, from an epizootic in farmed turbot, Scophthalmus maximus. Aquaculture 67, 117-126.
Mortensen S.H., Hjeltnes B., RØdseth O., Krogsrud J., Christie K.E. (1990) Infectious pancreaticnecrosis virus, serotype N1, isolated from Norwegian halibut (Hippoglossus hippoglossus), tubot (Scophthalmus maximus) and scallops (Pecten maximus). Bulletin of the European Association of Fish Pathologists 10, 42-43.
Rodriguez Saint-Jean S., Vilas M.P., Guiterrez M.C., Perez-Prieto S.I. (1997) Isolation and preliminary characterisation of a birnavirus from the sole Solea senegalensis in southwest Spain. Journal of Aquatic Animal Health 9, 295-300.
Wolf K., Gravell M., Malsberger R.G. (1966) Lymphocystis virus: isolation and propagation in centrarchid fish cell lines. Science 151, 1004-1005.
Kimura T., Yoshimizu M., Gorie S. (1986) A new rhabdovirus isolated in Japan from cultured hirame (Japanese flounder) Paralichthys olivaceus and ayu Plecoglossus altivelis. Diseases of Aquatic Organisms 1, 209-217.
312

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Nishizawa T., Yoshimizu M., Winton J.R., Ahne W., Kimura T. (1991) Characterization of structural proteins of hirame rhabdovirus, HRV. Diseases of Aquatic Organisms 10, 167-172.
Oseko N., Yoshimizu M., Kimura T. (1988) Effect of water temperature on artificial infection of Rhabdovirus olivaceus (hirame rhabdovirus: HRV) to hirame (Japanese flounder, Paralichthys olivaceus). Fish Pathology 23, 125-132.
OIE (2011) Aquatic Animal Health Code, 14th ed. World Organisation for Animal Health, Paris, 286 pp.
Schlotfeldt H.-J., Ahne W., Vestergard-JØrgensen R.E., Glende W. (1991) Bulletin of the European Association of Fish Pathologists 11, 105-107.
Ross K., McCarthy U., Huntly P.J., Wood B.P., Stuart D., Rough E.I., Smail D.A., Bruno D.W. (1994) An outbreak of viral hemorrhagic septicemia (VHS) in turbot (Scophthalmus maximus) in Scotland. Bulletin of the European Association of Fish Pathologists 14, 213-214.
Isshiki T., Nishizawa T., Kobayashi T., Nagano T., Miyazaki T. (2001) An outbreak of VHSV (viral hemorrhagic septicemia virus) infection in farmed Japanese flounder Paralichthys olivaceus in Japan. Diseases of Aquatic Organisms 47, 87-99.
Kim W.-S., Kim S.-R., Kim D., Kim J.-O., Park M.-A., Kitamira S., Kim H.-Y., Kim D.-H., Han H.-J., Jung S.-J., Oh M.-J. (2009) An outbreak of VHSV (viral hemorrhagic septicemia virus) infection in farmed olive flounder Paralichthys olivaceus in Korea. Aquaculture, 165-168.
Inouye K., Yamano K., Maeno Y., Nakajima K., Matsuoka M., Wada Y., Sorimachi M. (1992) Iridovirus infection of cultured red sea bream, Pagrus major. Fish Pathology 27, 19–27.
Matsuoka S., Inouye K., Nakajima K. (1996) Cultured fish species affected by red seabream iridoviral disease from 1991 to 1995. Fish Pathology 31, 233-234.
Kawakami H., Nakajima K. (2002) Cultured fish species affected by red sea bream iridoviral disease from 1996 to 2000. Fish Pathology 37, 45–47.
OIE (2012) Chapter 2.3.7. Red sea bream iridoviral disease In: Manual of Diagnostic Tests for Aquatic Animals 7th ed., World Organisation for Animal Health, Paris, pp. 345-356.
He J.G., Wang S.P., Zeng K., Huang Z.J., Chan S.M. (2000) Systemic disease caused by an iridoviruslike agent in cultured mandarin fish, Siniperca chuatsi (Basilewsky), in China. Journal of Fish Diseases 23, 219–222.
He J.G., Deng M., Weng S.P., Li Z., Zhou S.Y., Long Q.X., Wang X.Z., Chan S.M. (2001) Complete genome analysis of the mandarin fish infectious spleen and kidney necrosis iridovirus. Virology 291, 126-139.
Shi C.Y., Wang Y.G., Yang S.L., Huang J., Wang Q.Y. (2004) The first report of an iridovirus-like agent infection in farmed turbot, Scophthalmus maximus, in China. Aquaculture 236, 11-25.
Nakajima K., Kurita J., Ito T., Inouye K., Sorimachi M. (2001) Red sea bream iridoviral disease in Japan. Bulletin of National Research Institute of Aquaculture Suppl 5, 23–5.
Sano M., Nakai T., Fijan N. (2011) Viral diseases and agents of warm water fish. In Fish Diseases and Disorders: Volume 3. pp. 166-244, Woo P.T.K., Bruno D.W. (eds), CABI, Oxfordshire, UK.
Jancovich J.K., Chinchar V.G., Hyatt A., Miyazaki T., Williams T., Zhang Q.Y. (2011) Family Iridoviridae In Virus Taxonomy Nineth Report of the International Committee on Taxonomy of Viruses. pp. 193-210, King A.M.Q., Adams M.J., Carstens E.B., Lefkowitz E.J. (eds), Academic Press, San Diego.
Kurita J., Nakajima K., Hirono I., Aoki T. (2002) Polymerase chain reaction (PCR) amplification of DNA of red sea bream iridovirus (RSIV). Fisheries Sciences 68, Suppl. II, 1113-1115.
Do J.W., Moon C.H., Kim H.J., Ko M.S., Kim S.B., Son J.H., Kim J.S., An E.J., Kim M.K., Lee S.K., Han M.S., Cha S.J., Park M.S., Park M.A., Kim Y.C., Kim J.W., Park J.W. (2004) Complete genomic DNA sequence of rock bream iridovirus. Virology 325, 351-363.
Lu L., Zhou S.Y., Chen C., Weng S.P., Chan S.M., He J.G. (2005) Complete genome sequence analysis of an iridovirus isolated from the orange-spotted grouper, Epinephelus coioides. Virology 339, 81–100.
313

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Ao J., Chen X. (2006) Identification and characterization of a novel gene encoding an RGD-containing protein in large yellow croaker iridovirus. Virology 355, 213-222.
Nakajima K., Sorimachi M. (1994) Biological and physicochemical properties of iridovirus isolated from cultured red sea bream, Pagrus major. Fish Pathology 24, 29-33.
Dong C., Weng S., Shi X., Xu X., Shi N., He J. (2008) Development of a mandarin fish Siniperca chuatsi fry cell line suitable for the study of infectious spleen and kidney necrosis virus (ISKNV). Virus Research 135, 273-281.
Jun L.J., Jeong J.B., Kim J.H., Nam J.H., Shi K.W., Kim J.K., Kang J-C., Jeong H.D. (2009) Influence of temperature shifts on the onset and development of red sea bream iridoviral disease in rock bream Oplegnathus fasciatus. Diseases of Aquatic Organisms 84, 201-208.
Muroga K. (2001) Viral and bacterial diseases of marine fish and shellfish in Japanese hatcheries. Aquaculture, 202, 23-44.
Nakajima K., Sorimachi M. (1995) Production of monoclonal antibodies against red sea bream iridovirus. Fish Pathology 30, 47-52.
Nakajima K., Maeno Y., Fukudome M., Fukuda Y., Tanaka S., Matsuoka S., Sorimachi M. (1995) Immunofluorescence test for the rapid diagnosis of red sea bream iridovirus infection using monoclonal antibody. Fish Pathology 30, 115-119.
Kurita J., Nakajima K., Hirono I., Aoki, T. (1998) Polymerase chain reaction (PCR) amplification of DNA of red sea bream iridovirus (RSIV). Fish Pathology 33, 17-23.
Chao C-B., Yang S-C., Tsai H-Y., Chen C-Y., Lin C-S., Huang H.-T. (2002) A Nested PCR for the detection of grouper iridovirus in Taiwan (TGIV) in cultured hybrid grouper, giant seaperch, and largemouth bass. Journal of Aquatic Animal Health 14, 104–13.
Jeong J.B., Jun L.J., Yoo M.H., Kim M.S., Komisar J.L., Jeong H.D. (2003) Characterization of the DNA nucleotide sequences in the genome of red sea bream iridoviruses isolated in Korea. Aquaculture 220, 119–133.
Caipang C.M., Hirono I., Aoki T. (2003) Development of a real-time PCR assay for the detection and quantification of red seabream iridovirus (RSIV). Fish Pathology 38, 1–7.
Caipang C.M., Haraguchi I., Ohira T., Hirono I., Aoki T. (2004) Rapid detection of a fish iridovirus using loop-mediated isothermal amplification (LAMP). Journal of Virological Methods 121, 155-161.
Kim J.H., Gomez D.K., Choresca Jr. C.H., Park S.C. (2007) Detection of major bacterial and viral pathogens in trash fish used to feed cultured flounder in Korea. Aquaculture, 272, 105-110.
He J.G., Zeng K., Weng S.P., Chan S.M. (2002) Experimental transmission, pathogenicity and physical- chemical properties of infectious spleen and kidney necrosis virus (ISKNV). Aquaculture 204, 11–24.
Nakajima K., Maeno Y., Kurita J., Inui Y. (1997) Vaccination against red sea bream iridoviral disease in red sea bream. Fish Pathology 32, 205-209.
Nakajima K., Maeno Y., Honda A., Yokoyama K., Tooriyama T., Manabe S. (1999) Effectiveness of a vaccine against red sea bream iridoviral disease in a field trial test. Diseases of Aquatic Organisms 36, 73-75.
Yoshikoshi K., Inoue K. (1990) Viral nervous necrosis in hatchery-reared larvae and juveniles of Japanese parrotfish, Oplegnathus fasciatus (Temminck & Schlegel). Journal of Fish Diseases 13, 69–77.
Glazebrook J.S., Heasman M.P., de Beer S.W. (1990) Picorna-like viral particles associated with mass mortalities in larval barramundi, Lates calcarifer Bloch. Journal of Fish Diseases 13, 245–249.
Bloch B., Gravningen K., Larsen J.L. (1991) Encephalomyelitis among turbot associated with a piconavirus-like agent. Diseases of Aquatic Organisms 10, 65–70.
Breuil G., Bonami J.R., Pepin J.F., Pichot Y. (1991) Viral- infection (picorna-like virus) associated with mass mortalities in hatchery-reared sea-bass (Dicentrarchus labrax) larvae and juveniles. Aquaculture 97, 109–116.
314

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Mori K., Nakai T., Nagahara, M. Muroga, K. Mukuchi T., Kanno T. (1991) A viral disease in hatcheryreared larvae and juveniles of redspotted grouper. Fish Pathology 26, 209–210.
Mori K., Nakai T., Muroga K., Arimoto M., Mushiake K., Furusawa I. (1992) Properties of a new virus belonging to nodaviridae found in larval striped jack (Pseudocaranx dentex) with nervous necrosis. Virology 187, 368–371.
Fukuda Y., Nguyen H.D., Furuhashi M., Nakai T. (1996) Mass mortality of cultured sevenband grouper, Epinephelus septemfasciatus, associated with viral nervous necrosis. Fish Pathology 31, 165–170.
Le Breton A., Grisez L., Sweetman J., Olievier F. (1997) Viral nervous necrosis (VNN) associated with mass mortalities in cage-reared sea bass, Dicentrarchus labrax (L.). Journal of Fish Diseases 20, 145–151.
Aspehaug V., Devold M., Nylund A. (1999) The phylogenetic relationship of nervous necrosis virus from halibut (Hippoglossus hippoglossus). Bulletin of the European Association of Fish Pathologists 19, 196–202.
Patel S., Korsnes K., Bergh Ø., Vik-Mo F., Pedersen J., Nerland A.H. (2007) Nodavirus in farmed Atlantic cod Gadus morhua in Norway. Diseases of Aquatic Organisms 77, 169-173.
Korsnes K., Devold M., Nerland A.H., Nylund A. (2005) Viral encephalopathy and retinopathy (VER) in Atlantic salmon Salmo salar after intraperitoneal challenge with a nodavirus from Atlantic halibut Hippoglossus hippoglossus. Diseases of Aquatic Organisms 68, 7-15.
Munday B.L., Kwang J., Moody N. (2002) Betanodavirus infections of teleost fish: a review. Journal of Fish Diseases 25, 127–142.
Comps M., Pepin J.F., Bonami J.R. (1994) Viral infection (picorna-like virus) associated with mass mortalities in hatchery-reared Lares calcarifer and Dicentrarchus labrax. Aquaculture, 123, 1–10.
Chi S.C., Lo C.F., Lin S.C. (2001) Characterization of grouper nervous necrosis virus. Journal of Fish Diseases 24, 3–13.
Thiéry R., Johnson K.L., Nakai T., Schneemann A., Bonami J.R., Lightner D.V. (2012) Family Nodaviridae. In King A.M.Q., Adams M.J., Carstens E.B. and Lefkowitz E.J. (eds) Virus Taxonomy, Ninth Report of the International Committee on Taxonomy of Viruses. pp. 1061-1067, Academic Press, San Diego, California.
Nishizawa T., Mori K., Furuhashi M., Nakai T., Furusawa I., Muroga K. (1995) Comparison of the coat protein genes of five fish nodaviruses, the causative agents of viral nervous necrosis in marine fish. Journal of General Virology 76, 1563–1569.
Nishizawa T., Furuhashi M., Nagai T., Nakai T., Muroga K. (1997) Genomic classification of fish nodaviruses by molecular phylogenetic analysis of the coat protein gene. Applied and Environmental Microbiology 63, 1633–1636.
Iwamoto T., Mise K., Mori K., Arimoto M., Nakai T., Okuno T. (2001) Establishment of an infectious RNA transcription system for Striped jack nervous necrosis virus, the type species of the betanodaviruses. Journal of General Virology 82, 2653–2662.
Tan C., Huang B., Chang S.F., Ngoh G.H., Munday B.L., Chen S.C., Kwang J. (2001) Determination of the complete nucleotide sequences of RNA1 and RNA2 from greasy grouper (Epinephelus tauvina) nervous necrosis virus, Singapore strain. Journal of General Virology 82, 647–653.
Sommerset I., Nerland A.H. (2004) Complete sequence of RNA1 and subgenomic RNA3 of Atlantic halibut nodavirus (AHNV). Diseases of Aquatic Organisms 58, 117–125.
Iwamoto T., Okinaka Y., Mise K., Mori K., Arimoto M., Okuno T., Nakai T. (2004) Identification of host-specificity determinants in betanodaviruses by using reassortants between striped jack nervous necrosis virus and sevenband grouper nervous necrosis virus. Journal of Virology 78, 1256–1262.
Mori K., Mangyoku T., Iwamoto T., Arimoto M., Tanaka S., Nakai T. (2003) Serological relationships among genotypic variants of betanodavirus. Diseases of Aquatic Organisms 57, 19–26.
Iwamoto T., Mori K., Arimoto M., Nakai T. (1999) High permissivity of the fish cell line SSN-1 for piscine nodaviruses. Diseases of Aquatic Organisms 39, 37–47.
Frerichs G.N., Rodger H.D., Peric Z. (1996) Cell culture isolation of piscine neuropathy nodavirus from
315

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
juvenile sea bass, Dicentrarchus labrax. Journal of General Virology 77, 2067–2071.
Iwamoto T., Nakai T., Mori K., Arimoto M., Furusawa I. (2000) Cloning of the fish cell line SSN-1 for piscine nodaviruses. Diseases of Aquatic Organisms 43, 81–89.
Castric J., Thiéry R., Jeffroy J., de Kinkelin P., Raymond J.C. (2001) Sea bream Sparus aurata, an asymptomatic contagious fish host for nodavirus. Diseases of Aquatic Organisms 47, 33–38.
Barker D.E., MacKinnon A.M., Boston L., Burt M.D.B., Cone D.K., Speare D.J., Griffiths S., Cook M., Ritchie R., Olivier G. (2002) First report of piscine nodavirus infecting wild winter flounder Pleuronectes americanus in Passamaquoddy Bay, New Brunswick, Canada. Diseases of Aquatic Organisms 49, 99–105.
Gomez D.K., Sato J., Mushiake K., Isshiki T., Okinaka Y., Nakai T. (2004) PCR-based detection of betanodaviruses from cultured and wild marine fish with no clinical signs. Journal of Fish Diseases 27, 603–608.
Arimoto M., Sato J., Maruyama K., Mimura G., Furusawa I. (1996) Effect of chemical and physical treatments on the inactivation of striped jack necrosis virus (SJNNV). Aquaculture 143, 15–22.
Frerichs G.N., Tweedie A., Starkey W.G., Richards R.H. (2000) Temperature, pH andelectrolyte sensitivity, and heat, UV and disinfectant inactivation of sea bass (Dicentrarchus labrax) neuropathy nodavirus. Aquaculture 185, 13–24.
Mushiake K., Nishizawa T., Nakai T., Furusawa I., Muroga K. (1994) Control of VNN in striped jack: selection of spawners based on the detection of SJNNV gene by polymerase chain reaction (PCR). Fish Pathology 29, 177–182.
Mori K., Mushiake K., Arimoto M. (1998) Control measures for viral nervous necrosis in striped jack. Fish Pathology 33, 443–444.
Grotmol S., Totland G.K. (2000) Surface disinfection of Atlantic halibut Hippoglossus hippoglossus eggs with ozonated sea-water inactivates nodavirus and increases survival of the larvae. Diseases of Aquatic Organisms 39, 89–96.
Húsgard S., Grotmol S., Hjeltnes B., RØdseth O.M., Biering E. (2001) Immune response to a recombinant capsid protein of striped jack nervous necrosis virus (SJNNV) in turbot Scophthalmus maximus and Atlantic halibut Hippoglossus hippoglossus, and evaluation of a vaccine against SJNNV. Diseases of Aquatic Organisms 45, 33–44.
Tanaka S., Mori K., Arimoto M., Iwamoto T., Nakai T. (2001) Protective immunity of sevenband grouper, Epinephelus septemfasciatus Thunberg, against experimental viral nervous necrosis. Journal of Fish Diseases 24, 15–22.
Thiéry R., Cozien J., Cabon J., Lamour F., Baud M., Schneemann, A. (2006) Induction of a protective immune response against viral nervous necrosis in the European sea bass Dicentrarchus labrax by using betanodavirus virus-like particles. Journal of Virology 80, 10201–10207.
Yamashita Y., Fujita Y., Kawakami H., Nakai T. (2005) The efficacy of inactivated virus vaccine against viral nervous necrosis (VNN). Fish Pathology 40, 15–21.
Yamashita H., Mori K., Kuroda A., Nakai, T. (2009) Neutralizing antibody levels for protection against betanodavirus infection in sevenband grouper, Epinephelus septemfasciatus (Thunberg), immunized with an inactivated virus vaccine. Journal of Fish Diseases 32, 767-775.
Kai Y.H., Chi S.C. (2008) Efficacies of inactivated vaccines against betanodavirus in grouper larvae (Epinephelus coioides) by bath immunization. Vaccine 26, 1450-1457.
Bibliography (Section 3. Salmon and Trout Viral Diseases) World Organization for Animal Health, Manual of diagnostic tests for aquatic animals, 383 p, OIE, Paris.
AFS-FHS (American Fisheries Society-Fish Health Section) (2010). FHS blue book: suggested procedures for the detection and identification of certain finfish and shellfish pathogens, 2010 edition. AFS-FHS, Bethesda, Maryland.
Patrick W.T.K. and Bruno D.W. (2011). Fish diseases and disorders, volume 3: Viral, Bacterial and Fungal Infections, 2nd edition, 930 pp. CABI, Oxfordshire, UK.
316

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
King A.M.Q., Adams M.J., Carstens E.B. and Lefkowiz E.J. (ed) (2012). Virus taxonomy: Ninth report of the international committee on taxonomy of viruses, 1338 p. Elsevier, Diego, CA USA.
Oh M.J., Yoshimizu M., Kimura T. and Egusa Y. (1995). A new virus isolated from salmonid fish. Fish Pathology, 30, 23-32.
Bibliography (Section 4. Shrimp Diseases) FAO (2012) http://www.fao.org/fishery/statistics/software/fishstat/en,
Lightner, D.V. , Redman, R.M., Pantoja, C.R., Tang, K.F.J., Noble, B.L., Schofield, P., Mohney, L.L., Nunan, L.M., Navarro, S.A.(2012)Historic emergence, impact and current status of shrimp pathogens in the Americas. Journal of Invertebrate Pathology, 110(2), 174–183.
Lightner, D.V. (2011)Virus diseases of farmed shrimp in the Western Hemisphere (the Americas): A review. Journal of Invertebrate Pathology, 106(1), 110–130.
Flegel, T.W. (2012) Historic emergence, impact and current status of shrimp pathogens in Asia. Journal of Invertebrate Pathology, 110(2),166-173.
OIE. (2012) http://www.oie.int/en/international-standard-setting/aquatic-manual/access-online/.
Mayo, M.A. (2002) A summary of taxonomic changes recently approved by ICTV. Archives of Virology147(8), 1655-1656.
Mayo, M.A. (2005) Changes to virus taxonomy 2004. Archives of Virology 150(1), 189-198.
Flegel, T.W. (2006) Detection of major penaeid shrimp viruses in Asia, a historical perspective with emphasis on Thailand. Aquaculture, 256(1-4), 1-33.
Spann, K.M., Cowley, J.A., Walker, P.J., Lester, R.J.G. (1997) A yellow-head-like virus from Penaeus monodon cultured in Australia. Diseases of Aquatic Organisms 31(3), 169-179.
Walker, P. J., Bonami, J. R., et al., (2005) Family Roniviridae. In "Virus Taxonomy 8th Report" Elsevier, USA, 975-979.
Wijegoonawardane, P.K.M., Cowley, J.A., Phan, T., Hodgson, R.A.J., Nielsen, L., Kiatpathomchai, W., Walker, P.J. (2008) Genetic diversity in the yellow head nidovirus complex. Virology, 380(2), 213–225.
Soowannayan, C., Flegel, T.W., Sithigorngul, P., Slater, J., Hyatt, A., Cramerri, S., Wise, T., Crane, M.St.J., Cowley, J.A., McCulloch, R.J., Walker, P.J., (2003) Detection and differentiation of yellow head complex viruses using monoclonal antibodies. Diseases of Aquatic Organisms, 57(3), 193-200.
Lightner, D.V., Redman, R.M., Bell, T.A. (1983) Infectious hypodermal and hematopoietic necrosis, a newly recognized virus disease of penaeid shrimp. Journal of Invertebrate Pathology, 42(1), 62-70.
Bonami, J. R. and Lightner, D. V. (1991) Unclassified viruses of Crustacea. In "Atlas of Invertebrate Viruses", CRC Press, Boca Raton, FL, 597-622.
Bonami, J.R., Trumper,B., Mari, J., Brehelin, M., Lightner, D.V. (1990) Purification and characterization of the infectious hypodermal and haematopoietic necrosis virus of penaeid shrimps. Journal of General Virology. 71(11), 2657-2664.
Rai, P., Safeena, M.P., Karunasagar, I., Karunasagar, I. (2011) Complete nucleic acid sequence of Penaeus stylirostris densovirus (PstDNV) from India. Virus Research, 158(1-2), 37-45.
Kalagayan, H., Godin, D., Kanna, R., Hagino, G., Sweeney, J., Wyban, J., Brock, J., (1991) IHHN virus as an etiological factor in Runt-deformity syndrome (RDS) of juvenile Penaeus vannamei cultured in Hawaii. J. World Aquaculture Society. 22(4), 235-243.
Chayaburakul, K. Lightner, D.V. Sriurairattana, S. Nelson, K.T. Withyachumnarnkul, B (2005) Different responses to infectious hypodermal and hematopoietic necrosis virus (IHHNV) in Penaeus monodon and P. vannamei. Diseases of Aquatic Organisms, 67(3), 191-200.
Primavera, J.H. and Quinitio, E.T. (2000) Runt-Deformity Syndrome in cultured giant tiger prawn Penaeus monodon. Journal of Crustacean Biology, 20(4), 796–802.
Wertheim, J.O., Tang, K.F.J., Navarro, S.A., Lightner, D.V. (2009) A quick fuse and the emergence of Taura syndrome virus. Virology, 390(2), 324–329.
317

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Chen, Y.P., Nakashima, N., et al., (2012) In: King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J. (Eds.), Virus Taxonomy, 9th Report of the ICTV, pp. 840–845 (1327p).
Qian, D., Shi, Z., Zhang, S., Cao, Z., Liu, W., Li, L., Xie, Y., Cambournac, I., Bonami J.R. (2003). Extra small virus-like particles (XSV) and nodavirus associated with whitish muscle disease in the giant freshwater prawn, Macrobrachium rosenbergii. Journal of Fish Diseases, 26(9), 521–527.
Bonami, J. R., Shi, Z., Qian, D. Widada, J.(2005)White tail disease of the giant freshwater prawn, Macrobrachium rosenbergii: Separation of the associated virions and characterization of MrNV as a new type of nodavirus. Journal of Fish Diseases, 28(1), 23-31.
Wang, C.S., Chang, J.S., Wen, C.M., Shih, H.H., Chen, S.N. (2008) Macrobrachium rosenbergii nodavirus infection in M. rosenbergii (de Man) with white tail disease cultured in Taiwan. Journal of Fish Diseases, 31(6), 415-422.
Sahul Hameed, A.S., Yoganandhan, K., Sri Widada, J.b., Bonami, J.R.(2004) Experimental transmission and tissue tropism of Macrobrachium rosenbergii nodavirus (MrNV) and its associated extra small virus (XSV) Diseases of Aquatic Organisms, 62(3), 191–196.
Sudhakaran R., Ishaq Ahmed V.P., Haribabu, P., Mukherjee, S.C., Sri Widada, J., Bonami, J.R., Sahul Hameed, A.S. (2006) Experimental vertical transmission of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV) from brooders to progeny in Macrobrachium rosenbergii and Artemia. Journal of Fish Diseases, 30(1), 27-35.
Poulos, B.T., Tang, K.F.J., Pantoja, C.R., Bonami, J.R., Lightner, D.V. (2006) Purification and characterization of infectious myonecrosis virus of penaeid shrimp. Journal of General Virology, 87(4), 987-996.
Tang, K.F.J., Pantoja, C.R., Poulos, B.T. Redman, R .M., Lightner D.V.(2005)In situ hybridization demonstrates that Litopenaeusvannamei, L. stylirostris and Penaeus monodon are susceptible to experimental infection with infectious myonecrosis virus (IMNV). Diseases of Aquatic Organisms, 63(2-3), 261-265.
Kiatpathomchai, W., Boonsaeng, V., Tassanakajon, A., Wongteerasupaya, C., Jitrapakdee, S., Panyim, S. (2001) A non-stop, single-tube, semi-nested PCR technique for grading the severity of white spot syndrome virus infections in Penaeus monodon. Diseases of Aquatic Organisms, 47(3), 235-239.
Tsai, J.-M., Wang, H.-C., Leu, J-H., Wang, A.H.-J., Zhuang, Y., Walker, P. J., Kou, G-H., Lo. C-F.(2006)Identification of the nucleocapsid, tegument, and envelope proteins of the shrimp white spot syndrome virus virion. Journal of Virology, 80(6), 3021-3029.
Vaseeharan, B., Jayakumar, R., Ramasamy, P. (2003) PCR-based detection of white spot syndrome virus in cultured and captured crustaceans in India. Letters in Applied Microbiology, 37(6), 443-447.
Durand, S.V. and Lightner, D.V. (2002) Quantitative real time PCR for the measurement of white spot syndrome virus in shrimp. Journal of Fish Diseases, 25(7), 381-389.
Durand, S.V., Redman, R.M., Mohney, L.L., Tang-Nelson, K.F.J., Bonami, J.R., Lightner, D.V (2003) Qualitative and quantitative studies on the relative virus load of tails and heads of shrimp acutely infected with WSSV. Aquaculture, 216(1-4), 9-18.
Powell J.W.B., Burge, E.J., Browdy, C.L., Shepard, E.F. (2006) Efficiency and sensitivity determination of Shrimple, an immunochromatographic assay for white spot syndrome virus (WSSV), using quantitative real-time PCR. Aquaculture, 257 (1–4), 167–172.
Kono, T., Savan, R., Sakai, M., Itami, T. (2004) Detection of white spot syndrome virus in shrimp by loop-mediated isothermal amplification. Journal of Virological Methods,115(1), 59–65.
Mekata, T., Sudhakaran, R., Kono, T., Supamattaya, K., Linh, N.T.H., Sakai, M., Itami, T. (2009) Real-time quantitative loop-mediated isothermal amplification as a simple method for detecting white spot syndrome virus. Letters in Applied Microbiology, 48(1), 25–32.
Wongteerasupaya, C., Thongchuea, W., Boonsaeng, V., Panyim, S., Tassanakajon, A., Withyachumnarnkul, B., Flegel, T. (1997) Detection of yellow head-virus (YHV) for Penaeus monodon by RT-PCR amplification. Diseases of Aquatic Organisms, 31(3), 181–186.
318

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Cowley, J.A., Dimmock, C.M., Spann, K.M., Walker, P.J. (2000) Detection of Australian gill-associated virus (GAV) and lymphoid organ virus (LOV) of Penaeus monodon by RT-nested PCR. Diseases of Aquatic Organisms, 39(3), 159–167.
Cowley, J.A., Cadogan, L.C., Wongteerasupaya, C., Hodgson, R.A.J., Spann, K.M., Boonsaeng, V., Walker,P.J. (2004) Multiplex RT-nested PCR differentiation detection of gill-associated virus (Australia) and yellow head virus (Thailand) of Penaeus monodon. Journal of Virological Methods, 117(1), 49–59.
Wijegoonawardane, P.K.M., Cowley, J.A., Walker, P.J. (2010) A consensus real-time PCR for detection of all genotypic variants of yellow-head virus of penaied shrimp. Journal of Virological Methods, 167(1), 5–9.
Mekata, T., Sudhakaran, R., Kono, T., U-taynapun, K., Supamattaya, K., Suzuki, Y., Sakai, M., Itami, T. (2009) Real-time reverse transcription loop-mediated isothermal amplification for rapid detection of yellow head virus in shrimp. Journal of Virological Methods, 162(1), 81-87.
Vega-Heredia, S., Mendoza-Cano, F., Sánchez-Paz, A. (2012) The infectious hypodermal and haematopoietic necrosis virus: a brief review of what we do and do not know. Transboundary and Emerging Diseases, 59 (2) 95-105.
Chayaburakul, K., Lightner, D.V., Sriurairattana, D., Tang, K.F.J., Nelson, K., Withyachumnarnkul, B. (2005) Different responses to infectious hypodermal and hematopoietic necrosis virus (IHHNV) in Penaeus monodon and P. vannamei. Diseases of Aquatic Organisms, 67(3), 191–200.
Lightner, D. V. (1996) Epizootiology, distribution and the impact on international trade of two penaeid shrimp viruses in the Americas. Revue scientifiqueet technique / Office international des epizooties.15(2), 579-601.
Tang, K. F. J. and Lightner, D. V. (2001) Detection and quantification of infectious hypodermal and hematopoietic necrosis virus in penaeid shrimp by real-time PCR. Diseases of Aquatic Organisms, 44(2), 79-85.
Yue, Z.Q., Liu, H., Wang, W., Lei, Z.W., Liang, C.Z., Jiang, Y.L., (2006) Development of real-time polymerase chain reaction assay with TaqMan probe for the quantitative detection of infectious hypodermal and hematopoietic necrosis virus from shrimp. Journal of AOAC International, 89(1), 240–244.
Dhar, A.K., Roux, M.M., Klimpel, K.R. (2001) Detection and quantification of infectious hypodermal and hematopoietic necrosis virus and white spot virus in shrimp using real-time quantitative PCR and SYBR green chemistry. Journal of Clinical Microbiology, 39(8), 2835 -2845.
Sun, Z.F., Hu, C.Q., Ren, C.H., Shen, Q. (2006) Sensitive and rapid detection of infectious hypodermal and hematopoietic necrosis virus (IHHNV) in shrimps by loop-mediated isothermal amplification. Journal of Virological Methods, 131(1), 41–46.
He, L. and Xu, H.S. (2011).Development of a multiplex loop-mediated isothermal amplification (mLAMP) method for the simultaneous detection of white spot syndrome virus and infectious hypodermal and hematopoietic necrosis virus in penaeid shrimp. Aquaculture, 311(1), 94-99.
Arunrut, N., Prombun, P., Saksmerprome, V., Flegel, T.W., Kiatpathomchai, W. (2011) Rapid and sensitive detection of infectious hypodermal and hematopoietic necrosis virus by loop-mediated isothermal amplification combined with a lateral flow dipstick. Journal of Virological Methods, 171(1), 21–25.
Sudhakaran, R., Mekata, T., Kono, T, Supamattaya, K., Linh, N., Sakai, M., Itami, T. (2008) Rapid detection and quantification of infectious hypodermal and hematopoietic necrosis virus in white leg shrimp Penaeus vannamei using real-time loop-mediated isothermal amplification. Fish Pathology, 43(4), 170–173.
Teng, P.H., Chen, C.L., Sung, P.F., Lee, F.C., Ou, B.R., Lee, P.Y. (2007) Specific detection of reverse transcription-loop-mediated isothermal amplification amplicons for Taura syndrome virus by colorimetric dot-blot hybridization. Journal of Virological Methods, 146(1), 317–326.
Teng, P.H., Chen, C.L., Wu, C.N., Wu, S.Y., Ou, B.R., Lee, P.Y. (2006) Rapid and sensitive detection of Taura syndrome virus using nucleic acid-based amplification. Disease of Aquatic Organisms, 73(1), 13-22.
Kiatpathomchai, W., Jareonram, W., Jitrapakdee, S., Flegel, T.W. (2007) Rapid and sensitive detection of Taura syndrome virus by reverse transcription loop-mediated isothermal amplification. Journal of
319

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Virological Methods, 146(1), 125–128
Kiatpathomchai, W., Jaroenram, W., Arunrut, N., Jitrapakdee, S., Flegel, T.W. (2008) Reverse transcription loop-mediated isothermal amplification combined with the lateral flow dipstick test for specific detection of the Taura syndrome virus. Journal of Virological Methods, 153(2), 214–217.
Arcier, J.M., Herman, F., Lightner, D.V., Redman, R., Mari, J., Bonami, J.R. (1999). A viral disease associated with mortalities in hatchery-reared postlarvae of the giant fresh water prawn Marcobrachium rosenbergii. Diseases of Aquatic Organisms 38(3), 177-181.
Hsieh, C.Y., Wu, Z.B., Tung, M.C., Tu, C., Lo, S.P., Chang, T.C., Chang, C.D., Chen, S.C., Hsieh, Y.C., Tsai, S.S. (2006) In situ hybridization and RT-PCR detection of Marcobrachium rosenbergii nodavirus in giant fresh water prawn, Marcobrachium rosenbergii (de Man), in Taiwan. Journal of Fish Diseases 29(11), 665-671.
Yoganandhan, K., Sri Widada, J., Bonami, J.R., Sahul Hameed, A.S. (2005) Simultaneous detection of Macrobrachium rosenbergii nodavirus and extra small virus by a single tube, one-step multiplex RT–PCR assay. Journal of Fish Diseases, 28(2), 65–69.
Tripathy, S., Sahoo, P.K., Kumari, J., Mishra, B.K., Sarangi, N., Ayyappan, S. (2006) Multiplex RT-PCR detection and sequence comparison of viruses MrNV and XSV associated with white tail disease in Macrobrachium rosenbergii. Aquaculture, 258(1-4), 134–139.
Phillai, D., Bonami, J.R., Sri Widada, J. (2006) Rapid detection of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV), the pathogenic agents of white tail disease of Macrobrachium rosenbergii (De Man), by loop-mediated isothermal amplification. Journal of Fish Diseases, 29(5), 275–283.
Puthawibool, T., Senapin, S., Flegel, T.W., Kiatpathomchai, W. (2010) Rapid and sensitive detection of Macrobrachium rosenbergii nodavirus in giant freshwater prawns by reverse transcription loop-mediated isothermal amplification combined with a lateral flow dipstick. Molecular and Cellular Probes, 24(5), 244–249
Haridas, D.V., Pillai, D., Manojkumar, B., Nair, C.M., Sherief, P.M. (2010) Optimization of reverse transcriptase loop-mediated isothermal amplification assay for rapid detection of Macrobrachium rosenbergii nodavirus and extra small virus in Macrobrachium rosenbergii. Journal of Virological Methods, 167(1), 61-67.
Zhang, H., Wang, J., Yuan, J., Li, L., Zhang, J., Bonami, J-R., Shi, Z. (2006) Quantitative relationship of two viruses (MrNV and XSV) in white-tail disease of Macrobrachium rosenbergii. Diseases of Aquatic Organisms, 71(1), 11-17.
Poulos, B.T. and Lightner, D.V. (2006)Detection of infectious myonecrosis virus (IMNV) of penaeid shrimp by reverse-transcriptase polymerase chain reaction (RT-PCR). Diseases of Aquatic Organisms, 73(1), 69-72.
Andrade, T.P.D., Srisuvan, T., Tang, K.F.J., Lightner, D.V.(2007)Real-time reverse transcription polymerase chain reaction assay using TaqMan probe for detection and quantification of infectious myonecrosis virus (IMNV). Aquaculture, 264(1-4), 9-15.
Andrade, T.P.D. and Lightner, D.V. (2009) Development of a method for the detection of infectious myonecrosis virus by reverse-transcription loop-mediated isothermal amplification and nucleic acid lateral flow hybrid assay. Journal of Fish Diseases, 32(11), 911-924.
Van Hulten, M.C.W., Westenberg, M., Goodall, S.D., Vlak, J.M. (2000) Identification of two major virion protein genes of white spot syndrome virus of shrimp. Virology, 266(2), 227-236.
Van Hulten, M.C.W., Tsai, M.F., Schipper, C.A., Lo, C.F., Kou, G.H., Vlak, J.M. (2000) Analysis of a genomic segment of white spot syndrome virus of shrimp containing ribonucleotide reductase genes and repeat regions. Journal of General Virology, 81(2), 307-316.
Rout, N., Kumar, S., Jaganmohan, S., Murugan, V. (2007) DNA vaccines encoding viral envelope proteins confer protective immunity against WSSV in black tiger shrimp. Vaccine, 25(15), 2778–2786.
Rajesh Kumar, S., Ishaq Ahamed, V.P., Sarathi, M,. Nazeer Basha, A., Sahul Hameed, A.S. (2008)
320

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
Immunological responses of Penaeus monodon to DNA vaccine and its efficacy to protect shrimp against white spot syndrome virus (WSSV). Fish and Shellfish Immunology, 24(4), 467-478.
Kono, T., Sonoda, K., Kitao, Y., Mekata, T., Itami, T., Sakai, M. (2009) The expression analysis of innate immune-related genes in Kuruma shrimp Penaeus japonicus after DNA vaccination against penaeid rod-shaped DNA virus. Fish Pathology, 44 (2), 94–97
Rajesh Kumar, S., Venkatesan, C., Sarathi, M., Sarathbabu, V., Thomas, J., Anver Basha, K., Sahul Hameed, A.S. (2009) Oral delivery of DNA construct using chitosan nanoparticles to protect the shrimp from white spot syndrome virus (WSSV). Fish and Shellfish Immunology, 26(3), 429-437.
Satoh, J., Nishizawa, T., Yoshimizu, M. (2009) Duration and booster effect of phylactic response against white spot syndrome virus infection in kuruma shrimp orally administrated with recombinant viral proteins, rVP26 and rVP28. Fish Pathology, 44(3), 120–127.
Satoh, J., Nishizawa, T., Yoshimizu, M. (2008﴿ Protection against white spot syndrome virus (WSSV) infection in kuruma shrimp orally vaccinated with WSSV rVP26 and rVP28. Diseases of Aquatic Organisms, 82(2), 89-96.
Ning, D., Leng, X., Li, Q., Xu, W. (2011) Surface-displayed VP28 on Bacillus subtilis spores induce protection against white spot syndrome virus in crayfish by oral administration. Journal of Applied Microbiology, 111 (6), 1327–1336.
Sarathi, M., Simon, M.C., Ishaq-Ahmed, V.P., Rajesh-Kumar, S., Sahul-Hameed, A.S. (2008) Silencing VP28 gene of white spot syndrome virus of shrimp by bacterially expressed dsRNA. Marine Biotechnology, 10 (2), 198–206.
Shekhar M.S. and Lu, Y. (2009).Application of nucleic-acid-based therapeutics for viral infections in shrimp aquaculture. Marine Biotechnology, 11(1), 1–9.
Xu, J.Y., Han, F., Zhang, X.B. (2007). Silencing shrimp white spot syndrome virus (WSSV) genes by siRNA. Antiviral Research, 73(2), 126–131.
Kim, C. S., Kosuke, Z., Nam, Y. K., Kim, S. K., Kim, K. H. (2007) Protection of shrimp (Penaeus chinensis) against white spot syndrome virus (WSSV) challenge by double-stranded RNA. Fish and Shellfish Immunology, 23 (1), 242–246.
Sudhakaran, R., Mekata, T., Kono, T., Inada, M., Okugawa, S., Yoshimine, M., Yoshida, T., Sakai, M., Itami, T. (2010) Double-stranded RNA-mediated silencing of the white spot syndrome virus VP28 gene in kuruma shrimp, Marsupenaeus japonicus. Aquaculture Research, 42(8), 1153-1162.
Tirasophon, W., Yodmuang, S., Chinnirunvong, W., Plongthongkum, N., Panyim, S. (2007) Therapeutic inhibition of yellow head virus multiplication in infected shrimps by YHV-protease dsRNA. Antiviral Research, 74 (2), 150–155.
Sudhakaran, R., Syed Musthaq, S., Haribabu, P., Mukherjee, S.C., Gopal, C., Sahul Hameed, A.S. (2006a) Experimental transmission of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV) in three species of marine shrimp (Penaeus indicus, Penaeus japonicus and Penaeus monodon). Aquaculture, 257, 136–141.
Sudhakaran, R., Yoganandhan, K., Ishaq Ahmed, V.P., Sahul Hameed A.S. (2006b) Artemia as a possible vector for Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus transmission (XSV) to Macrobrachium rosenbergii post-larvae. Diseases of Aquatic Organisms, 70(1-2), 161–166.
Bonami, J.R. and Widada J.S. (2011) Viral diseases of the giant fresh water prawn Macrobrachium rosenbergii: a review. Journal of Invertebrate Pathology, 106 (1), 131–142.
Biographical Sketches Motohiko Sano is currently a professor for Department of Marine Biosciences, Tokyo University of Marine Science and Technology. He obtained Ph.D. degrees from Tokyo University of Fisheries in 1990. Dr. Sano has worked with fish disease for many years. Toshihiro Nakai is currently a professor in Graduate School of Biosphere Science, Hiroshima University.
321

FISH DISEASES - Diseases Caused By Viral Pathogens - Toshihiro Nakai, Motohiko Sano, Mamoru Yoshimizu, Hisae Kasai, Toshiaki Itami, Raja Sudhakaran
©Encyclopedia of Life Support Systems (EOLSS)
He obtained M.Agr. from Hiroshima University in 1979 and Ph.D. from The University of Tokyo in 1986. Dr. Nakai has worked in the field of fish pathology for more than 30 years, particularly with bacterial and viral diseases of cultured fish. Hisae Kasai is currently an associate professor for Faculty of Fisheries Sciences, Hokkaido University. She obtained Ph.D. degrees from Hokkaido University in 2005. Dr. Kasai has worked with fish disease, food safety for many years. Mamoru Yoshimizu is a professor for Faculty of Fisheries Sciences, Hokkaido University. He obtained Ph.D. degrees from Hokkaido University. Dr. Yoshimizu has researched with fish disease and prevention of pathogens for many years. Toshiaki Itami is currently a professor for Faculty of Agriculture, University of Miyazaki, Japan. He obtained Ph.D. degrees from Kyoto University. Dr. Itami has worked with shrimp disease and bio-defense system for many years.. Raja Sudhakaran is currently an assistant professor for Aquatic Animal Health Laboratory, School of Biosciences and Technology, VIT University in India. His major research is shrimp disease and bio-defense system.
322





























