Embed Size (px)
Citation preview

TIIE JOURNAL OF l3IoLooICAL CriEalrsTRV Vol. ZAG, No. 19,Issws of October 10, pp. 6029443G. 1971
Printed in U.S.A.
Electron Paramagnetic Resonance and Proton Relaxation Rate
Studies of Spin-labeled Creatine Kinase and Its Complexes*
(Received for publication, March 26, 1971)
JUNE s. Tar~ort,$ iZ~.m MCLAUGHLIN, AND mLDRED COHN$
From the Departwent oj Biophysics and Physical Biochelnist~y, Johnson Reseawh Foundation, University oj Pennsylvania Xchool sf Jteclicine, Philadelphia, Pennsylvania 19104
SUMMARY
A spin-labeled derivative of rabbit muscle creatine kinase has been prepared by reaction of the essential sulfhydryl group at the active sites with a stoichiometric amount of the nitroxide radical N-( I-oxyl-2,2,5,5-tetramethyl-d-pyrroli- dinyl) iodoacetamide. The spin-labeled enzyme retains the ability to bind nucleotide substrates although it lacks the phosphotransferase and ATPase activities of the native en- zyme. Two magnetic resonance properties of the specifically spin-labeled enzyme and its substrate complexes were used to probe the local environment of the covalently bound radical: (a) the electron paramagnetic resonance (EPR) spectrum and (b) the enhancement of the proton relaxation rate of water protons. The protein-bound radical shows a broad, asymmetric EPR spectrum resembling a solid state powder spectrum; the outer peaks are separated by 62 gauss compared with 31 gauss for the free radical and the amplitude of the central peak is 14 times greater in the unbound than the bound spin label. In addition, the spin-labeled protein enhances the water proton relaxation rate by approximately 1 order of magnitude. Both phenomena indicate a high degree of immobilization of the bound spin label. Metal nucleotide binding increases immobilization of the spin label as evi- denced by the EPR spectrum. The spectral changes repre- sent the first direct evidence for a proposed conformational change in the region of the active site on addition of divalent ions to the enzyme-nucleotide binary complex. The proton relaxation rate of water is even more sensitive to local struc- tural changes and reveals differences between the enzyme and its binary nucleotide complexes as well as between binary and ternary enzyme-metal nucleotide complexes. Creatine alone or in combination with metal nucleotides fails to affect either the EPR spectrum or the water proton relaxation rate of the spin-labeled enzyme.
The nucleotide complexes of the alkali earth cations,
* This investigation was supported in part by Grant GM-12446 from the National Institutes of Health. United States Public Health Service, and by Grant GB-18487 from the National Science Foundation. A preliminary report of some of this work has been published (I).
Y$ Recipient of Predoctoral Training Grant 5 TOl-GM-OOG94 from t.he United States Public Health Service. Present address, De- partment, of Biochemistry, Tale University, New Haven, Con- necticut OG510.
$ Career Investigator of the American Heart Association.
Mg(II), Ca(II), Sr(II), and Ba(II), produce approximately the same changes in the EPR of the spin-labeled enzyme, amounting to a diminution of 20% in central peak amplitude upon addition of metal-ADP and a somewhat smaller effect with the ATP complexes. Quantitative differences in the properties of the different metal complexes are most easily observed by measurement of the proton relaxation rate of water under optimal temperature and frequency conditions. The ternary complexes of the irreversibly inhibitory Zn(I1) nucleotides with spin-labeled enzyme have qualitatively dif- ferent EPR spectra than do the corresponding complexes of alkali earth ions. Apparent dissociation constants have been calculated from both EPR and proton relaxation rate titra- tions for the dissociation of the alkali earth nucleotides from the ternary complexes. All are of the same order of magni- tude as for the native enzyme, about 0.1 mM for ADP ternary complexes.
The t.ernary metal nucleotide-enzyme complexes of creatine kinase have been studied extensively by a number of workers (l-7) to determine the structural features of the complexes which affect the catalytic activity. Magnetic resonance studies of manganese nucleotide-enzyme complexes have provided strong evidence that the metal ion does not funct.ion as a bridge between enzyme and substrate, but is bound to the nucleotide only (2, 3). There is evidence that specificity of nucleotide substrates mani- fested by variation in maximum reaction velocity is correlated with conformational changes induced in the active site by the substrates. The pa,rallel variation in water prot.on relaxation rate, Vmax, and the sulfhydryl reactivity towards iodoacetate (4) for a series of met,al nucleotides, ADP > deoxyADP > IDP suggests that binding of metal nucleotide induces a conforma- tional change, the nature of which is sensitive to the structure of the nucleotide. It has been suggested that the different effects of nucleotide inhibitors and metal nucleotide substrates on the iodoacet,ate reaction are due to a combination of conformational and charge contributions in the respective enzyme complexes (5).
In the present studies, a spin label covalently bound to the sulfhydryl group at the active sites of creatine kinase has been used to investigate conformational changes in the neighborhood of the spin label which occur upon nucleot,ide and met,al nucleo-
6029
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6030 EPR ancl PRR of Spin-labeled Cyeatine Kinase Vol. 246, Xo. 19
tide binding. This approach is more direct than t,hose used prc-
viously since the observed changes in the electron paramagnetic resonance spectrum can be interpreted unequivocally in terms of local conformational changes (8, 9) in the region of the CJS-
teinc residue of the active site. It also became possible to inves- tigatc the question whether the relative effectiveness of various alkali earth cations in bringing about conformational change is related to their efficacy as activators of t,he enzymatic reaction. The spin-labeled enzyme makes it possible to extend the range of complexes studied by EPR’ and PRR to those containing no met.al ions, and to complexes with a series of divalent ions of rarying charge densities and coordination properties.
MATERIALS AND ~IIcTEIOD~
Xnterials-The sodium salt of Al)P \vas obtained from P-L Biochemicals and the sodium salt of ATP from P-L Biochemicals and from Sigma. Met,al salts were spectrographically analyzed reagents purchased from Johnson, Matthcy, and Company, Ltd., London. Creatine was obtained from Fisher Scientific and phosphocreatine from Boehringer-Mannheim. The spin label, N-( I-oxyl-2,2,5,5-tetramethyl- 3 - pyrrolidinyl) iodoacetamide, was synthesized by the method of Rozantzev and Krinitzkaya (10) or purchased from Synvar Associates, Palo Alto, California. N-I~th~lmorpholine was obtained from K and I< Laboratories and redistilled under reduced pressure. Tris (Trizma base) was purchased from Sigma. All other reagents were reagent grade and used without further purification. Quartz double distilled water was used throughout.
&zyme Pyeparatirm-Creatine kinasc was prepared from rab- bit skeletal muscle by Method B of Kuby, Noda, and Lardy (II). The lengthy crystallization procedure was often replaced by a chromatographic procedure which yielded enzyme of maxi- mal activity and purity. Fraction IV obtained by their proce- dure was chromatographed on a column of W’hatman microgran- ular DEAE-cellulose, equilibrated with 0.01 AI Tris-HCl, 0.01 M NaCl, pH 8.5, at 4”, and elutcd with a linear gradient of 0.01 M to 0.10 M NaCl in 0.01 M Tris-HCl, pH 8.5. Enzyme so pre- pared has a specific activity of 240 pcq of base consumed per min per mg of enzyme when tested at 30” in the pH-stat assay of Mahowald, Noltman, and Kuby (12). The enzyme displays a single band upon cellulose acetate strip electrophoresis at pH 4 and 8 and in persulfate-polymerized gels at pH 8.1 (13).
Magnetic Resonance Measurements-EPR spectra were re- corded on a Varian Associates E-3 spectrometer operating at 9.1 GHz. EPR measurements were also performed at 35 GHz on a Varian Associates 4503 spectrometer. Unless otherwise stated, the spectra were taken at 9.1 GHz, 32 milliwatt power, and 1.25 gauss modulation amplitude. The positions of the peaks in the magnetic field were reported from the dial readings of the spectrometer. A solution of 1 rnnf MnClz in 1 mu HCl Q-as used as a standard in order to compare amplitudes of de- rivative spectra taken at different times. Pulsed NMR meas- urements of t.he longitudinal relaxation time, T1, of water pro- tons using the Carr-Purcell pulsing sequence (14) were made at 8, 15, 24.3, 40, and 60 MHz at various temperatures on an in- strument constructed in this laboratory, equipped with a Varian model V-4340 variable temperature accessory. No attempt was made to remove 02 from the solutions. Titrations were per- formed by addition of aliquots of a solution cont.aining enzyme,
1 The abbreviations used are: EPR, electron paramagnetic resonance; PRR, proton relaxation rate.
metal, and concentrated nucleot.idc to a solution containing en- zyme and metal only; for titrations, EPR measurements and PRR measurements (at 24.3 MHz) were made in rapid succes- sion on the same solution. In the studies on frequency and temperature dependence of the proton relaxation rates of the various complexes, only single metal nucleotidc concentrations in the presence of 0.21 mar enzyme (active sites) were measured over the entire range of frequency and temperature. In the ATP experiments, saturating concentrations were used for mag- nesium, calcium, and strontium (25 m&f) ; it was not est,ablished whether 25 mM spermidine ATP was saturating. Zinc was used at 10 mM which was shown to be saturating. In the ADP CX-
periments, the met.al nucleotide concentrations were as follows: magnesium, 25 mr; calcium, 8 mnl; zinc, 10 mar; spermidine, 25 mar; magnesium, calcium, and zinc concentrations were Fat- urating but spermidine concentrat.ion was not.
RESULTS
Preparation of Spin-labeled Enxywze
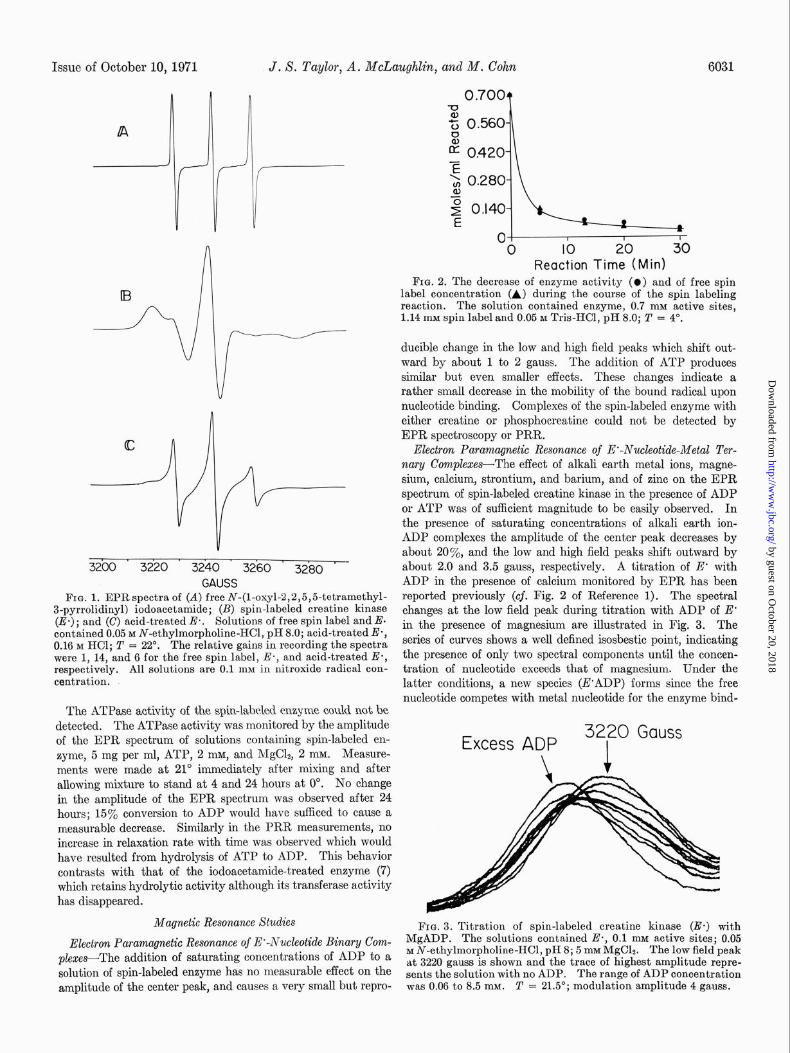
Creatine kinase specifically labeled at the sulfhydryl group of the active site is prepared by reacting enzyme (40 mg per ml, 1.0 mM active sites) with spin-labeled iodoacet,amide, 1.10 ml1 N-(1-oxy-2,2,5,5-tetramethyl-3-pyrrolidinyl) iodoacetamide, in 0.05 M N-ethylmorpholine-HCl, pH 8.0, at 0” for a period of approximately 1 hour. The extent of reaction is monitored bj measuring residual enzyme activity. When the activity has fallen to less than 1% of the initial value, excess free label is removed by gel filtration at 4” on Sephadex G-25 equilibrated with 0.05 M N-ethylmorpholine-II-ICl, ~1-18, followed by overnight dialysis against 200 volumes of the same buffer. Fig. 1 shows EPR derivative spectra of solutions of free spin label (Curve A), of spin-labeled enzyme (hereafter designated E’) (Curve B), and of acid-treated E’ (Curve C). The asymmetrical EPR spectrum of E’ in neutral solution resembles a powder spectrum of the radical, so that the bound radical must be highly immobilized (9). The spectrum (B) has a positive peak at ~3220 gauss, a central peak at ~3242 gauss with a maximum amplitude 14 times less than that of the isotropic spectrum (A) of the spin label, and a negative peak at -3282 gauss. The peaks of the unbound spin label spectrum (A) fall at about 3227, 3242, and 3258 gauss. Upon acidification in 0.16 M HCl, the EPR spec- trum of E’ reverts to the typical three-line spectrum (C) charac- teristic of a rapidly tumbling nitroxide radical with slightly less symmetry than the unbound free radical (cf. Fig. 1). The bound form in the acid-treated protein is distinguishable from t.he free form by the broader line widths, about 2 gauss at 20” for t,he former compared with line widths of <l gauss for the latter.
The spin label reacts with creatine kinase at the same rate as iodoacetamide which has been shown to alkylate the reactive sulfhydryl group at the active site stoichiometrically (15). The concentration of spin label remaining at any time during t,he reaction with native enzyme may be followed by measuring the amplitude of its distinctive peak at 3258 gauss. The stoichiom- etry of the spin-labeling reaction was checked by observing si- multaneously the decrease in enzyme activity and the decrease in free spin label concentration (Fig. 2). Aliquots were diluted loo-fold to stop the reaction and the activity was measured in a coupled enzymatic assay (16). Within the error of the meas- urements (,t3’%), 1 mole of spin label reacts per mole of enzyme subunits.
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of October 10, 1971
A
B
J. 8. Taylor, A. McLaughlin, and ill. Cohn 6031
/
V iL-----
3200 3220 3240 3260 3280 GAUSS
FIG. 1. EPRspectra of (A) free N-(I-oxyl-2,2,5,5-tetramethyl- 3-pyrrolidinyl) iodoacetamide; (B) spin-labeled creatine kinase (II*); and (C) acid-treated.E.. Solutions of free spin label and E. contained 0.05 M N-ethylmorpholine-HCl, pH 8.0; acid-treated E*, 0.16 M HCl; T = 22'. The relative gains in recording the spectra were 1, 14, and 6 for the free spin label, E-, and acid-treated E*, respectively. All solutions are 0.1 mu in nitroxide radical con- centration.
Electron Paramqnetic Resonance of E’-Nuckotide-Metal Ter- nary CompZexes-The effect of alkali earth metal ions, magne- sium, calcium, stront.ium, and barium, and of zinc on the EPR spectrum of spin-labeled creatine kinase in the presence of ADP or ATP was of sufficient magnitude to be easily observed. In the presence of saturating concentrations of alkali earth ion- ADP complexes the amplitude of the center peak decreases by about 20%, and the low and high field peaks shift outward by about 2.0 and 3.5 gauss, respectively. A titration of E* with ADP in the presence of calcium monitored by EPR has been reported previously (cf. Fig. 2 of Reference 1). The spectral changes at the low field peak during titration with ADP of E’ in the presence of magnesium are illustrated in Fig. 3. The series of curves shows a well defined isosbestic point, indicating the presence of only two spectral components until the concen- tration of nucleotide exceeds that of magnesium. Under the latter conditions, a new species (E’ADP) forms since the free nucleotide competes with metal nucleotide for the enzyme bind-
The ATPase activity of the spin-labeled enzyme could not be detected. The ATPase activity was monitored by the amplitude of the EPR spectrum of solutions containing spin-labeled en- zyme, 5 mg per ml, ATP, 2 mM, and MgC12, 2 InM. Measure- ments were made at 21” immediately after mixing and after allowing mixture to stand at 4 and 24 hours at 0”. No change in the amplitude of the EPR spectrum was observed after 24 hours; 15% conversion to ADP would have sufficed to cause a measurable decrease. Similarly in the PRR measurements, no increase in relaxation rate with time was observed which would have resulted from hydrolysis of ATP to ADP. This behavior contrasts with that of the iodoacetamide-treated enzyme (7) which retains hydrolytic activity although its transferase activity has disappeared.
Excess ADP 32?o Gauss
Magnetic Resonance Studies FIG. 3. Titration of spin-labeled creatine kinase (E*) with MgADP. The solutions-contained E*, 0.1 mu active sites; 0.05 M N-ethvlmornholine-HCI. DH 8: 5 mM MrrCL The low field neak Electron Paramagnetic Resonance of E*-Nucleotick Binary Com-
plexes-The addition of saturating concentrations of ADP to a solution of spin-labeled enzyme has no measurable effect on the amplitude of the center peak, and causes a very small but repro-
IS ,
at 3220 gauss;s shown and the trace of highest amplitude repre- sents the solution with no ADP. The range of ADP concentration was 0.06 to 8.5 mM. T = 21.5’; modulation amplitude 4 gauss.
0.700 T¶ 1
$ 0.560
- h g 0.420 =G \, 0.280 -I\
01 I I I 0 IO 20 30
Reaction Time (Mid FIG. 2. The decrease of enzyme activity (0) and of free spin
label concentration (A) during the course of the spin labeling reaction. The solution contained enzyme, 0.7 mM active sites, 1.14 mM spin label and 0.05 M Tris-HCI, pH 8.0; T = 4".
ducible change in the low and high field peaks which shift out- ward by about 1 to 2 gauss. The addition of ATP produces similar but even smaller effects. These changes indicate a rather small decrease in the mobility of the bound radical upon nucleotide binding. Complexes of the spin-labeled enzyme with either creatine or phosphocreatine could not be detected by EPR spectroscopy or PRR.
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from
6032 EPR and PRR of #pin-labeled Creatine Kinase Vol. 246, No. 19
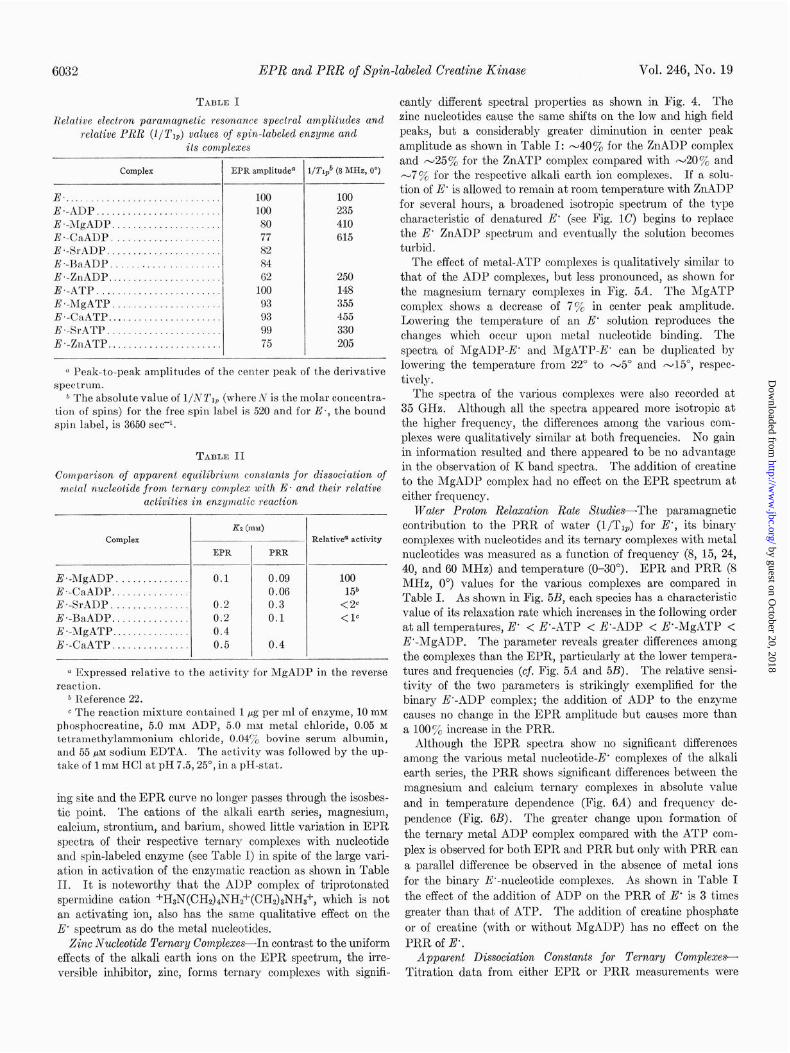
TABLE I
Relative electron paramagnetic resonance spectral amplitudes and relative PRR (l/T,,) values of spin-labeled enzyme and
its complexes
Complex EPR amplitude” l/Txpb (8 MHz, 0’)
E.. 100 100 E .-ADP . . . . 100 235 E .-MgADP. . 80 410 E .-CaADP . 77 615 E .-SrADP 82 E .-BaADP 84 E .-ZnADP. 62 250 E.-ATP. . 100 148 E ,-MgATP 93 355 E .-CaATP. . 93 455 E .-SrATP 99 330 E.-ZnATP.. . . . . 75 205
a Peak-to-peak amplitudes of the center peak of the derivative spectrum.
b The absolute value of l/ATT,, (where N is the molar concentra- tion of spins) for the free spin label is 520 and for E., the bound spin label, is 3650 see-l.
TABLE II
Comparison of apparent equilibrium constants for dissociation of metal nucleotide from ternary complex with E’ and fheir relative
activities in enzymatic reaction
Complex
E .-MgADP . E.-CaADP. E .-SrADP E.-BaADP., E .-MgATP . E .-CaATP . .
Kz (mar)
EPR PRR
0.1 0.09 0.06
0.2 0.3 0.2 0.1 0.4 0.5 0.4
Relatives activity
100 15b
<2= < 1c
a Expressed relative to the activity for MgADP in the reverse reaction.
* Reference 22. c The reaction mixture contained 1 pg per ml of enzyme, 10 mM
phosphocreatine, 5.0 rnb% ADP, 5.0 mM metal chloride, 0.05 M
tetramethylammonium chloride, 0.04% bovine serum albumin, and 55 PM sodium EDTA. The activity was followed by the up- take of 1 mM HCl at pH 7.5,25”, in a pH-stat.
ing site and the EPR curve no longer passes through the isosbes- tic point. The cations of the alkali earth series, magnesium, calcium, strontium, and barium, showed little variation in EPR spectra of their respective ternary complexes with nucleotide and spin-labeled enzyme (see Table I) in spite of the large vari- ation in activation of the enzymatic reaction as shown in Table II. It is noteworthy that the ADP complex of triprotonated spermidine cation +H3N(CH2)4NHiC(CH&NH3+, which is not an activating ion, also has the same qualitative effect on the E’ spectrum as do the metal nucleotides.
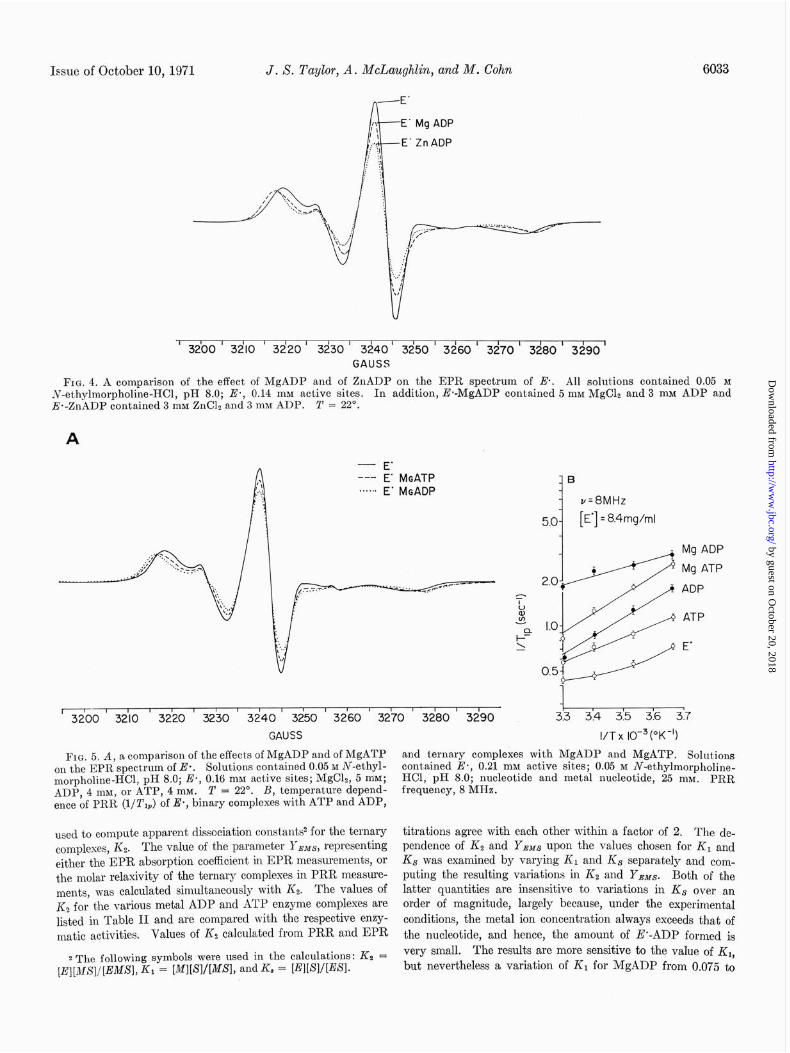
Zinc Nudeotide Ternary Complexes-In contrast to the uniform effects of the alkali earth ions on the EPR spectrum, the irre- versible inhibitor, zinc, forms ternary complexes with signifi-
cantly different spectral properties as shown in Fig. 4. The zinc nucleotides cause the same shifts on the low and high field peaks, but a considerably greater diminution in center peak amplitude as shown in Table I: ~40% for the ZnADP complex and ~25% for the ZnATP complex compared with -20% and -7% for the respective alkali earth ion complexes. If a solu- tion of E’ is allowed to remain at room temperature with ZnADP for several hours, a broadened isotropic spectrum of the type characteristic of denatured E’ (see Fig. 1C) begins to replace the E’ ZnADP spectrum and eventually the solution becomes turbid.
The effect of metal-ATP complexes is qualitatively similar to that of the ADP complexes, but less pronounced, as shown for the magnesium ternary complexes in Fig. 5A. The MgATP complex shows a decrease of 7% in center peak amplitude. Lowering the temperature of an E’ solution reproduces the changes which occur upon metal nucleotide binding. The spectra of MgADP-E’ and MgATP-E’ can be duplicated by lowering the temperature from 22” to ~5~ and -15”, respec- tively.
The spectra of the various complexes were also recorded at 35 GHz. Although all the spectra appeared more isotropic at the higher frequency, the differences among the various com- plexes were qualitatively similar at both frequencies. No gain in information resulted and there appeared to be no advantage in the observation of K band spectra. The addition of creatine to the MgADP complex had no effect on the EPR spectrum at either frequency.
Water Proton Relaxation Rate Studies-The paramagnetic contribution to the PRR of water (l/T& for E’, its binary complexes with nucleotides and its ternary complexes with metal nucleotides was measured as a function of frequency (8, 15, 24, 40, and 60 MHz) and temperature (O-30”). EPR and PRR (8 MHz, 0”) values for the various complexes are compared in Table I. As shown in Fig. 5B, each species has a characteristic value of its relaxation rate which increases in the following order at all temperatures, E” < E’-ATP < E’-ADP < E’-MgATP < E’-MgADP. The parameter reveals greater differences among the complexes than the EPR, particularly at the lower tempera- tures and frequencies (cj. Fig. 5A and 5B). The relative sensi- tivity of the two parameters is strikingly exemplified for the binary E’-ADP complex; the addition of ADP to the enzyme causes no change in the EPR amplitude but causes more than a 100% increase in the PRR.
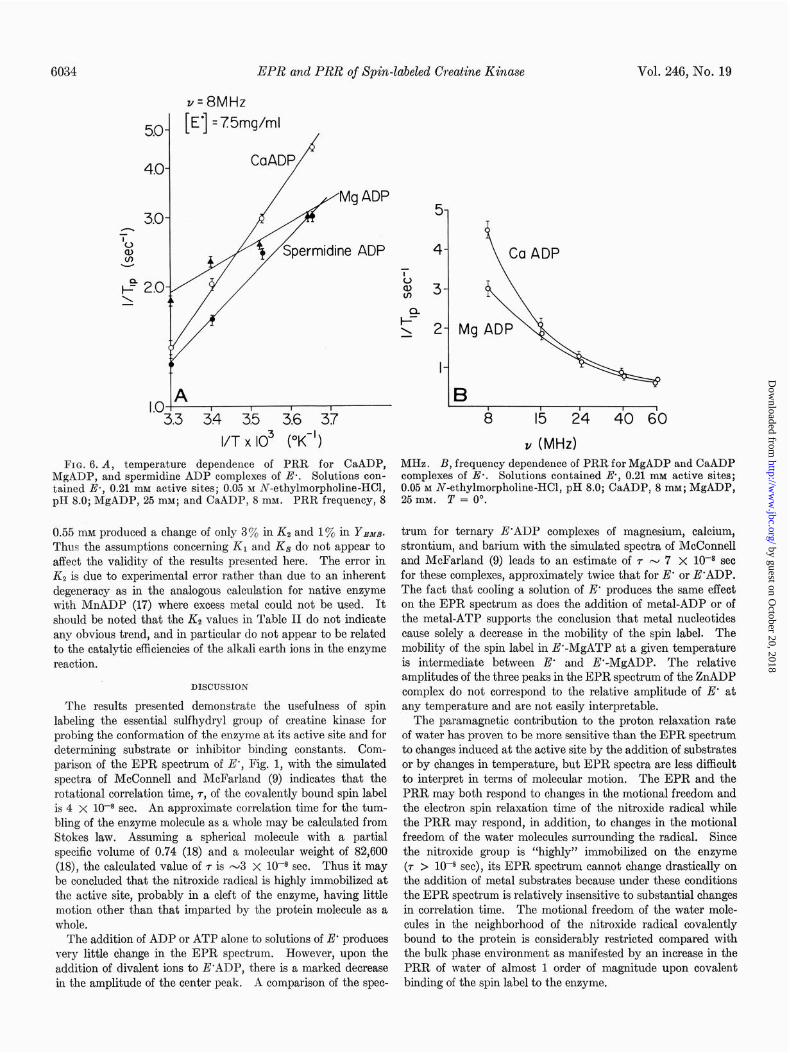
Although the EPR spectra show no significant differences among the various metal nucleotide-E’ complexes of the alkali earth series, the PRR shows significant differences between the magnesium and calcium ternary complexes in absolute value and in temperature dependence (Fig. 6A) and frequency de- pendence (Fig. 6B). The greater change upon formation of the ternary metal ADP complex compared with the ATP com- plex is observed for both EPR and PRR but only with PRR can a parallel difference be observed in the absence of metal ions for the binary E’-nucleotide complexes. As shown in Table I the effect of the addition of ADP on the PRR of E’ is 3 times greater than that of ATP. The addition of creatine phosphate or of creatine (with or without MgADP) has no effect on the PRR of E’.
Apparent Dissociation Constants for Ternary Complexes- Titration data from either EPR or PRR measurements were
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Issue of October 10, 1971 J. S. Taylor, A. McLaughlin, and M. Cohn 6033
Mg ADP
Zn ADP
' 3200 ' 3dlO # I I I I I I I , , I I I
3220 3230 3240 3250 3260' 3270' 3280' 3290' GAUSS
FIG. 4. A comparison of the effect of MgADP and of ZnADP on the EPR spectrum of E. All solut.ions contained 0.05 M iv-ethylmorpholine-HCl, pH 8.0; E., 0.14 mM active sites. In addition, E*-MgADP contained 5 rnM MgClz and 3 mM ADP and E*-ZnADP contained 3 mM ZnClr and 3 mM ADP. T = 22”.
A
- E’ --- E’ MGATP .**..+ E’ MGADP
11 1 t 1 ’ I”’ 1 ’ 1 ” “1 11 3200 3210 3220 3230 3240 3250 3260 3270 3280 3290
GAUSS
FIG. 5. A, a comparison of the effects of MgADP and of MgATP on the EPR spectrum of Es. Solutions contained 0.05 M N-ethyl- morpholine-HCl, pH 8.0; E*, 0.16 mM active sites; MgC12, 5 mM; ADP, 4 mM, or ATP, 4 mM. T = 22’. B, temperature depend- ence of PRR (l/T,*) of E*, binary complexes with ATP and ADP,
used to compute apparent dissociation constants2 for the ternary complexes, K2. The value of the parameter YBMs, representing either the EPR absorption coefficient in EPR measurements, or the molar relaxivity of the ternary complexes in PRR measure- ments, was calculated simultaneously with Kz. The values of Kz for the various metal ADP and ATP enzyme complexes are listed in Table II and are compared with the respective enzy- matic activities. Values of & calculated from PRR and EPR
2 The following symbols were used in the calculations: KS = [E][MS]/[EMSI, K1 = MtSlI[MSl, and K = LWWWI.
B
v=8MHz
5.0 [E’] = 8.4mg/ml
3.3 3.4 3.5 3.6 3.7
I/T x lO-3 (“K-‘1
and ternary complexes with MgADP and MgATP. Solutions contained Es, 0.21 mM active sites; 0.05 M N-ethylmorpholine- HCl, pH 8.0; nucleotide and metal nucleotide, 25 mM. PRR frequency, 8 MHz.
titrations agree with each other within a factor of 2. The de- pendence of Kz and YsMs upon the values chosen for K1 and K8 was examined by varying KI and Ks separately and com- puting the resulting variations in KS and YEMS. Both of the latter quantities are insensitive to variations in KS over an order of magnitude, largely because, under the experimental conditions, the metal ion concentration always exceeds that of the nucleotide, and hence, the amount of E’-ADP formed is very small. The results are more sensitive to the value of K,, but nevertheless a variation of K1 for MgADP from 0.075 to
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from
6034 EPR and PRR of Spin-labeled Creatine Kinase Vol. 246, No. 19
v=8MHz
[ 1 E’ = 75mg/ml
CaADP P
A I I I I
3 34 3.5 3.6 37
I/T x IO3 (OK-‘)
FIG. 6. A, temperature dependence of PRR for CaADP, MHz. B, frequency dependence of PRR for MgADP and CaADP MgADP, and spermidine ADP complexes of Ea. Solutions con- complexes of E*. Solutions contained E, 0.21 rnM active sites; tained E*, 0.21 mu active sites; 0.05 M N-ethylmorpholine-HCl, 0.05 M N-ethylmorpholine-HCl, pH 8.0; CaADP, 8 mM; MgADP, pH 8.0; MgADP, 25 mM; and CaADP, 8 mM. PRR frequency, 8 25 mM. 2’ = 0’.
0.55 mu produced a change of only 3% in Kz and 1% in Y.w.T. Thus the assumptions concerning K1 and Ks do not appear to affect the validity of the results presented here. The error in Kz is due to experimental error rather than due to an inherent degeneracy as in the analogous calculation for native enzyme with MnADP (17) where excess metal could not be used. It should be noted that the Kz values in Table II do not indicate any obvious trend, and in particular do not appear to be related to the catalytic efficiencies of the alkali earth ions in the enzyme reaction.
DISCUSSION
The results presented demonstrate the usefulness of spin labeling the essential sulfhydryl group of creatine kinase for probing the conformation of the enzyme at its active site and for determining substrate or inhibitor binding constants. Com- parison of the EPR spectrum of E’, Fig. 1, with the simulated spectra of McConnell and McFarland (9) indicates that the rotational correlation time, 7, of the covalently bound spin label is 4 X 10-S sec. An approximate correlation time for the tum- bling of the enzyme molecule as a whole may be calculated from Stokes law. Assuming a spherical molecule with a partial speci6c volume of 0.74 (18) and a molecular weight of 82,600 (18), the calculated value of r is ~3 X lo-* sec. Thus it may be concluded that the nitroxide radical is highly immobilized at the active site, probably in a cleft of the enzyme, having little motion other than that imparted by the protein molecule as a whole.
The addition of ADP or ATP alone to solutions of E' produces very little change in the EPR spectrum. However, upon the addition of divalent ions to E’ADP, there is a marked decrease
B I I I I I 8 I5 24 40 60
v (MHz)
trum for ternary E’ADP complexes of magnesium, calcium, strontium, and barium with the simulated spectra of McConnell and McFarland (9) leads to an estimate of r N 7 X lo-* see for these complexes, approximately twice that for E' or E’ADP. The fact that cooling a solution of E* produces the same effect on the EPR spectrum as does the addition of metal-ADP or of the metal-ATP supports the conclusion that metal nucleotides cause solely a decrease in the mobility of the spin label. The mobility of the spin label in E’-MgATP at a given temperature is intermediate between E’ and E’-MgADP. The relative amplitudes of the three peaks in the EPR spectrum of the ZnADP complex do not correspond to the relative amplitude of E* at any temperature and are not easily interpretable.
The paramagnetic contribution to the proton relaxation rate of water has proven to be more sensitive than the EPR spectrum to changes induced at the active site by the addition of substrates or by changes in temperature, but EPR spectra are less difficult to interpret in terms of molecular motion. The EPR and the PRR may both respond to changes in the motional freedom and the electron spin relaxation time of the nitroxide radical while the PRR may respond, in addition, to changes in the motional freedom of the water molecules surrounding the radical. Since the nitroxide group is “highly” immobilized on the enzyme (7 > lo-* set), its EPR spectrum cannot change drastically on the addition of metal substrates because under these conditions the EPR spectrum is relatively insensitive to substantial changes in correlation time. The motional freedom of the water mole- cules in the neighborhood of the nitroxide radical covalently bound to the protein is considerably restricted compared with the bulk phase environment as manifested by an increase in the PRR of water of almost 1 order of magnitude upon covalent
in the amplitude of the center peak. A comparison of the spec- binding of the spin label to the enzyme.
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of October 10, 1971 J. X. Taylor, A. McLaughlin, and M. Cohn 6035
Differences in PRR values among the species are most easily observed at low frequencies since, in general, relaxation effects on the water protons decrease with increasing frequency (19). Similarly, since the correlation times associated with most re- laxing mechanisms have negative temperature coefficients, the PRR effects are largest at low temperatures (Fig. 5B).
In an attempt to interpret the mechanism of the proton re- laxation rates, the temperature and frequency dependences of the PRR were investigated. Theoretical analysis of the data3 indicates that two forms of water contribute to the PRR: “free” water in the active site and “bound” water either in hydration spheres of the bound substrates or on amino acid residues near the active site. There is no evidence for a stable hydration sphere around the nitroxide radical, such as exist,s around an aqueous metal ion. The observed paramagnetic contribution to the relaxation rate is the average of the con- tributions from the two classes of water. The enhancement of the relaxation rate on covalent binding of the nitroxide to the enzyme and again on the addition of substrates is due to de- creases in the diffusional mobility of the water immediately surrounding the nitroxide, as well as to changes in the amount and location of the “bound” water near the radical.
O’Sullivan and Cohn (4) postulated differences in conforma- tion at the active site for the native enzyme and each of its binary and ternary complexes on the basis of differences in re- activity of the essential -SH groups. The EPR spectra pro- vide suggestive evidence for, firstly, a conformational change upon addition of metal ion to the protein-nucleotide complex, and secondly, a difference in protein conformation between ATP and ADP ternary complexes. The PRR measurements are consistent with both of these findings. Furthermore PRR values discriminate among the three conformations: E., E’-ADP, and E’-ATP.
Thus EPR and PRR measurements with enzyme spin labeled at the essential sulfhydryl group are sufficiently sensitive to furnish physical evidence for differing conformational states at the act,ive site of the enzyme alone, its inhibitor (nucleotide) complexes, its substrate (metal-nucleotide) complexes and even to distinguish between nucleotide di- and triphosphates. Tem- perat,ure-jump studies of the kinetics of conformational changes induced by ATP, ADP, metal-ATP, and metal-ADP reported by Hammes and Hurst (20), establish the fact that the con- formational change occurring upon addition of nucleotide or metal nucleotide occurs much faster than the time required for a single turnover of the enzyme, and thus meets an essential criterion for steps involved in the catalytic process.
Since the EPR and PRR of spin-labeled enzyme complexes reflect conformational changes between active and inactive species, e.g. the striking differences between nucleotide and metal nucleotide complexes, it was disappointing that a correlation with metal ion specificity could not be established. The EPR spectra of the alkali earth ion complexes (magnesium, calcium, strontium, and barium) showed no significant differences in spite of the large differences in enzymatic activity. Further- more, when the organic cation spermidine, a competitive in- hibitor of magnesium, replaces an alkali earth ion in the ternary complex, it gives qualitatively similar spectra although it is enzymatically inactive.
The difference between the magnesium and calcium complexes (Fig. 6B) observed in PRR measurements reflects a difference
in the nature of the bound water in each comp1ex.a The calcium complex would have either more bound water or have an equivalent amount bound at a distance closer to the radical. It is not obvious, however, that this structural difference is related to the difference in enzymatic activity for the two metals.
It is not surprising that ZnADP, which is an irreversible in- hibitor of the enzyme and leads to its eventual denaturation, should have an EPR spectrum in its ternary complex different from all other species. In particular, its structure differs in symmetry as well as motional freedom from the spin-labeled enzyme. It has been proposed that zinc irreversibly inhibits the enzyme by binding to the essential -SH groups (al), but the results for enzyme with spin label covalently bound to sulf- hydryl groups do not support this explanat,ion.
It could be argued that the active site conformations for the various alkali earth ions would differ in complexes with the native enzymes but not with the modified spin-labeled enzyme. Such a loss of ability t,o induce conformational change resulting from enzyme modification is not likely in view of the finding that the binding constants (Table II) remain the same (22). Also the environment of the binding site in the modified enzyme is not very different from the native as monitored both by the bind- ing constant of MnADP and the PRR of water due to man- ganese in the respective ternary E.-manganese nucleotide com- plexes as shown in the following paper. The possibility that the changes in conformation do not affect the mobility of the spin label and that another probe would reveal differences among the various metal ternary complexes cannot be ruled out.
We should like to propose that the ternary enzyme-metal nucleotide complexes of different metal ions have essentially the same conformation, but that the quaternary complexes do not. Whereas the presence of MgADP or BaADP affects the -SH reactivity towards iodoacetate to the same extent, MgADP plus creatine affords the enzyme almost comp1et.e protection against inhibition by iodoacetamide (7), yet the presence of BaADP plus creatine does not.4 Unfortunately, other methods will have to be devised to test this proposal since creatine kinase modified at the essential sulfhydryl groups, upon addition of creatine, no longer exhibits any change detectable by EPR or PRR of water.
Acknowledgments-We wish to acknowledge the use of the University of Pennsylvania Medical School Computer Facility under Grant RR-15 of the United States Public Health Service.
1.
2. 3.
4.
5.
6.
7.
8.
REFERENCES
TAYLOR, J. S., LEIGH, J. S., JR., AND COHN, M., Proc. Nat. Acad. Sci. U. S. A., 64, 219 (1969).
COHN, M., Biochemistry, 2, 623 (1963). O’SULLIVAN, W. J., AND COHN, M., J. Biol. Chem., 241, 3104
(1966). O’SULLIVAN, W. J., AND COHN, M., J. Biol. Chem., 241, 3116
(1966). O’SULLIVAN, W. J., DIEFENBACH, I-I., AND COHN, M., Bio-
chemistry, 6, 2666 (1966). LUI, N. S. T., AND CUNNINGHAM, L., Biochemistry, 6, 144
(1966). O’SULLIVAN, W. J., AND COHN, M., J. Biol. Chem., 243, 2737
(1968). SMITH, I., in J. R. BOLTON, D. BORG, AND H. SWARTZ (Edi-
tars), Biological applications of electron spin resonance spectroscopy, John Wiley-Interscience, in press.
4 W. J. O’Sullivan, Dersonal communication. ̂3 A. McLaughlin, unpublished observations.
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6036 EPR and PRR of Spin-labeled Creatine Kinase Vol. 246, No. 19
9. MCCONNELL, H. M., AND MCFARLAND, B. G., Quart. Rev. Biophys., 3, 91 (1970).
10. ROZANTZEV. E. G., AND Kn~w~zrcaya, L. A., Tetrahedron, 21, 491 (1965j. ’
Il. KUBY, S. A., NODA, L., AND LARDY, H. A., J. Biol. Chem., 209, 191 (1954).
12. MA’IAHO~AL~, T. A., NOLTMANX, E. A., AND KuB~, S. A., J. Biol. Chem., 237, 1535 (1962).
13. TAYLOIL, J. S., Ph.D. thesis, University of Pennsylvania, 1969, p. 35.
14. CARR, H. Y., AND PURCELL, E. M., Phys. Rev., 94, 630 (1954). 15. WATTS, D. C., AND RABIN, B. R., Biochem. J., 85, 507 (1962). 16. ROTTHAUWE, H. W., A~\TD CERQUEIRO-RODI~IGUEZ, M., Gin.
Chem. Ada, 10, 134 (1964).
17. REED, G. H., COHN, ill., AND O’SULLIVAN, W. J., J. Biol.
Chem., 245, 6547 (1970). 18. YUE, R. H., PALMIERI, It. II., OLSOX, 0. E., AND KUBP, S. A.,
Biochemistry, 6, 3204 (1967).
19. PEACOCKE, A. R., RICHARDS, R. E., AND SHEARD, B., ~lol.
Phys., 16, 177 (1969).
20. HAMA~ES, G. G., AND HURST, J. IL, Biochemistry, 8, 1083
(1969).
21. O’SULLIVAN, W. J., AND MORRISON, J. I?., Biochim. Biophys.
Acta, 7’7, 142 (1963). 22. MORRISON, J. F., AND UHR, M. L., Biochim. Biophys. Acta, 122,
57 (1966).
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from

June S. Taylor, Alan McLaughlin and Mildred CohnSpin-labeled Creatine Kinase and Its Complexes
Electron Paramagnetic Resonance and Proton Relaxation Rate Studies of
1971, 246:6029-6036.J. Biol. Chem.
http://www.jbc.org/content/246/19/6029Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/246/19/6029.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on October 20, 2018
http://ww
w.jbc.org/
Dow
nloaded from









![Proton NMR Spin – Lattice Relaxation Time in …H NMR relaxation times T 1 value [14-16], therefore, to study the effect of temperature on the chemical shift and relaxation time,](https://img.dokumen.tips/doc/110x75/5f085b3a7e708231d4219ae9/proton-nmr-spin-a-lattice-relaxation-time-in-h-nmr-relaxation-times-t-1-value.jpg)



















