Embed Size (px)
Citation preview

1
AMG 595, an anti-EGFRvIII Antibody Drug Conjugate, Induces Potent Anti-Tumor Activity Against
EGFRvIII Expressing Glioblastoma
Kevin J. Hamblett1 , Carl J. Kozlosky1, Sophia Siu1, Wesley S. Chang2, Hua Liu1, Ian N Foltz3, Esther S.
Trueblood1, David Meininger1, Taruna Arora4*, Brian Twomey4, Steven L. Vonderfecht4, Qing Chen4, John
S. Hill4, William C. Fanslow1
1 Amgen Inc., Seattle, WA
2 Amgen Inc., South San Francisco, CA
3 Amgen Inc., Burnaby, BC
4 Amgen Inc., Thousand Oaks, CA
Running title: AMG 595 an anti-EGFRvIII ADC for glioblastoma
Keywords: EGFRvIII, Glioblastoma Multiforme, antibody drug conjugate
All authors were employed by Amgen, Inc. and in some cases own Amgen stock.
Corresponding author:
Kevin J. Hamblett
Current Address: Kevin Hamblett, Zymeworks Biopharmaceuticals, Seattle, WA
Phone: 206-237-1031
e-mail: [email protected]
*Current Address: Taruna Arora, NGM Biopharmaceuticals, South San Francisco, CA
Word count: 4,904
Six figures and tables
Amgen, Inc. funded the research.
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

2
Abstract
Epidermal growth factor receptor variant III (EGFRvIII) is a cancer-specific deletion mutant observed in
approximately 25-50% of Glioblastoma multiforme (GBM) patients. An antibody drug conjugate, AMG
595, composed of the maytansinoid DM1 attached to a highly selective anti-EGFRvIII antibody via a non-
cleavable linker was developed to treat EGFRvIII positive GBM patients. AMG 595 binds to the cell
surface and internalizes into the endo-lysosomal pathway of EGFRvIII expressing cells. Incubation of
AMG 595 with U251 cells expressing EGFRvIII led to potent growth inhibition. AMG 595 treatment
induced significant tumor mitotic arrest, as measured by phospho-histone H3, in GBM subcutaneous
xenografts expressing EGFRvIII. A single intravenous injection of AMG 595 at 17 mg/kg (250 μg DM1/kg)
generated complete tumor regression in the U251vIII subcutaneous xenograft model. AMG 595
mediated tumor regression in the D317 subcutaneous xenograft model that endogenously expresses
EGFRvIII. Finally, AMG 595 treatment inhibited the growth of D317 xenografts orthotopically implanted
into the brain as determined by magnetic resonance imaging. These results demonstrate that AMG 595
is a promising candidate to evaluate in EGFRvIII expressing GBM patients.
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

3
Introduction:
An estimated 23,380 new cases of brain cancer will be diagnosed in the US in 2014 (1). Roughly half of
all brain cancers are grade IV, also known as Glioblastoma multiforme (GBM), which has an extremely
poor prognosis and a median survival of 14.6 months after diagnosis (2, 3). Yet, in the past twenty years
only two agents have been approved for the treatment of GBM; temozolomide for first-line patients as a
combination with adjuvant radiotherapy followed by temozolomide monotherapy (3) and bevacizumab
for patients with recurrent GBM (4). Sequencing of GBM by The Cancer Genome Atlas (TCGA) effort and
others is generating a clearer picture of the molecular basis of GBM (5). The TCGA determined that
amplification of EGFR is almost exclusively observed in the classical subtype of GBM. Epidermal growth
factor receptor (EGFR) expression is associated with a malignant phenotype in multiple cancers including
colon cancer (6), head and neck squamous cell carcinoma (7), and GBM (8). Amplification of EGFR is
observed in approximately 40-70% of GBM patients (9, 10). EGFR gene alterations are observed in
tumors with amplified EGFR (11). The most common genetic alteration found in GBM is EGFR variant III
(EGFRvIII) wherein exons 2-7 of EGFR have been deleted (12). Juxtaposition of EGFR exons 1 and 8
forms a novel glycine residue between these two exons. Deletion of EGFR’s exons 2-7 yields a protein
that is incapable of binding to EGFR ligands (13) but remains constitutively activate leading to a
malignant phenotype (14). EGFRvIII expression in GBM samples is reported between 25-50%,
depending on the method used for detection (15-17).
EGFRvIII is expressed in tumors, and its lack of normal tissue expression makes it an ideal candidate for
pursuing a targeted therapeutic (13). Multiple modalities are being investigated to target EGFRvIII
expressing tumors including kinase inhibitor combinations (18), peptide vaccination (19), and antibody
targeting (20). Antibodies can be modified to improve potency through enhanced Fc receptor binding
and effector function (21), engagement of T cells (22), or attachment of a cytotoxic drug (23). Recently
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

4
the antibody drug conjugates (ADCs) Adcetris (24) and Kadcyla (25) were approved for the treatment of
CD30 expressing Hodgkin lymphoma and anaplastic large cell lymphoma and Her2 expressing breast
cancer, respectively.
We have developed a fully human antibody that binds solely to EGFRvIII and does not bind to wild-type
EGFR. This fully human anti-EGFRvIII specific IgG1 antibody was conjugated to the anti-tubulin agent,
DM1, via the non-cleavable MCC linker to generate AMG 595. In this report we describe AMG 595, and
show its in vitro functional characterization and demonstrate its ability to mediate tumor regression in
EGFRvIII expressing subcutaneous and orthotopic xenografts.
Materials and Methods:
Reagents
U251, U251vIII, and D317 GBM cells were obtained through an agreement with Dr. Darell Bigner (Duke
University) and held in a repository at Amgen. EGFRvIII is a genomic deletion mutant that has only been
observed in human tumors, there are no reports that EGFRvIII is expressed in normal tissues (13).
EGFRvIII expression is lost from patient derived Glioblastoma cells cultured in vitro (26, 27). EGFRvIII
expression by flow cytometry or IHC was used to authenticate cell lines, cells were not passaged in vitro
for more than three months. U251vIII and U251 cells were cultured in 10% Dulbecco’s Modified Eagle
Medium with 10% fetal bovine serum. To generate fully human anti-EGFRvIII antibodies, Xenomouse®
animals (28) were immunized with either the EGFRvIII junctional peptide LEEKKGNYVVTDHC conjugated
to keyhole limpet hemocyanin (KLH) or cells expressing EGFRvIII. Following immunization, B cells were
either collected for generation of hybridomas or isolated using the Selected Lymphocyte Antibody
Method (29). Specific anti-EGFRvIII binding antibodies were selected for additional analysis. Antibodies
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

5
were conjugated to the maytansinoid DM1 (ImmunoGen, Inc.) using a non-cleavable linker (30). Briefly,
the anti-EGFRvIII antibody was modified with the hetero-bifunctional linker succinimidyl-4-(N-
maleimidomethyl)cyclohexane-1-carboxylate (SMCC). Following removal of the excess SMCC,
maleimide labeled antibody was incubated with DM1 to generate anti-EGFRvIII Ab-MCC-DM1 (AMG
595). Antibody and DM1 concentrations were calculated by solving a pair of Beer's Law equations at
252 and 280 nm (31). The human anti-EGFRvIII antibody constant regions were modified to murine
constant regions to generate a chimeric anti-EGFRvIII human variable/murine IgG1 constant domain
antibody for use in immunohistochemistry.
BIAcore Affinity Measurement
Binding of AMG 595 and anti-EGFRvIII antibody to human EGFRvIII was measured by solution
equilibrium binding assay using Surface Plasmon Resonance (BIAcore). EGFRvIII was immobilized on the
second flow cell of a CM5 chip using amine coupling (reagents provided by GE Healthcare, Piscataway,
NJ) with an approximate density of 7000 RU. The first flow cell was used as a background control. Three
hundred pM, 1.0 nM, or 3.0 nM of AMG 595 or anti-EGFRvIII antibody were mixed with a serial dilution
of EGFRvIII (ranging from 6.6 pM to 390 nM) in PBS plus 0.1 mg/mL BSA, 0.005% P20 and incubated at
room temperature for 4 hours. Binding of the free antibodies in the mixed solutions was measured by
injecting over the EGFRvIII coated chip surface. Non-linear regression curve fit was used to calculate
antibody affinity and 95% confidence intervals (CI).
Immunohistochemistry of GBM patient samples
Formalin fixed paraffin embedded (FFPE) GBM sections were heat treated in Biocare’s DIVA solution
using Biocare’s decloaking chamber (Biocare, Concord, CA) for SP1 125 C for 30 seconds and SP2 90 C
for 10 seconds. Frozen GBM sections were fixed with acetone at 4 C for 5 minutes. Tissue sections
were incubated for 60 minutes with 0.69 μg/ml of the chimeric anti-EGFRvIII human variable/murine
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

6
constant antibody, followed by EnVision+ mouse HRP polymer (DAKO, Carpinteria, CA) and DAB
chromogen to visualize EGFRVIII expression. Mouse IgG1 (Invitrogen, Carlsbad, CA) was used as an
isotype negative reagent control on all tissue sections.
Quantitative PCR of GBM patient samples
cDNA was synthesized from total RNA using the High-Capacity cDNA Archive Kit (Life Technologies). Per
sample, 1 g of RNA was reverse transcribed in a 100 l reaction volume. Quantitative PCR (qPCR) was
performed with a 7900HT SDS instrument using the following reagents: EGFRvIII forward primer:
5’GAGTCGGGCTCTGGAGGAA3’, reverse primer: 5’GGCCCTTCGCACTTCTTACAC 3’, probe: 5’[FAM]-
TCACCACATAATTACCTTTCT-[quencher]3’. The EGFRvIII primers were synthesized at Amgen and the
probe was synthesized at Applied Biosystems (Life Technologies, Grand Island, NY). EGFRvIII positive
and negative control DNAs were assayed with each sample batch. Each qPCR reaction nominally
contained 100 ng of cDNA. Cycle Threshold (CT) values were extracted with the “CT Analysis” settings
configured for “Automatic Baseline” and “Manual CT”, using the default threshold setting of 0.2.
Internalization of AMG 595
To assess cell surface binding, internalization, and trafficking of AMG 595, U251vIII cells were plated
onto collagen-coated glass chamber slides (Thermo Scientific, Waltham, MA) and grown to ~60%
confluence. For live cell surface labeling of EGFRvIII, cells were transferred to ice and incubated with
AMG 595 at 1 μg/mL in complete media containing lysosomal protease inhibitors leupeptin (10 μg/mL)
and pepstatin (5 μM) for 30 minutes at 4°C. Cells were washed twice in cold media containing lysosomal
protease inhibitors and either fixed immediately in 3% formaldehyde in PBS for 20 minutes on ice (t=0)
or incubated at 37°C for 30 minutes, 4 hours, or 24 hours, and then fixed. Cells were permeabilized and
blocked in TBST + 5% donkey serum + 0.1% TX-100, then AMG 595 was detected with a donkey anti-
human IgG–AlexaFluor 488 (Jackson ImmunoResearch, West Grove, PA). Early endosome marker EEA1
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

7
was detected using a rabbit antibody (Cell Signaling Technology, Danvers, MA) followed by anti-rabbit
IgG-Alexa 647 conjugated antibody (Invitrogen, Carlsbad, CA). Lysosomal marker LAMP1 was detected
with a mouse antibody (BD Biosciences, San Jose, CA), followed by anti-mouse Alexa 568 conjugated
antibody (Invitrogen, Carlsbad, CA). Cells were coverslipped with ProLong Gold + DAPI antifade reagent
(Invitrogen) and images were acquired using a 100X (N.A. 1.4) oil immersion objective lens on a Zeiss
LSM 510 confocal microscope system (Carl Zeiss, Jena GmBH).
Co-localization Image Analysis
Images were cropped in Adobe Photoshop (Adobe, San Jose, CA) to select regions of interest (ROI) for
analysis. 8-bit gray scale images of EEA1 and LAMP1 staining were pseudo-colored red and merged with
images of human IgG staining in green to generate composite images for quantitative co-localization
analysis using Columbus software (Perkin-Elmer, Waltham , MA). Co-localization analysis was performed
on pairs of channels within the composite image (hIgG/EEA1, hIgG/LAMP1) to generate Pearson’s R
values at each time point.
AMG 595 Mediated Cell Growth Inhibition
U251 and U251vIII cells were cultured in a 96-well tissue culture plate and incubated at 37°C, 5% CO2 for
approximately 4-6 hours. After the incubation, serial dilutions of AMG 595, control conjugate, DM1 or
Lysine-MCC-DM1 were added to cell cultures and continuously incubated at 37 C, 5% CO2 for 4 days
prior to measurement of cellular ATP levels using the CellTiter-Glo reagent (Promega, Madison, WI). The
ATP levels were directly correlated with viable cell number. Luminescence was measured using a Wallac
EnVision 2103 multilabel reader (Perkin Elmer, Waltham, MA) with a reading time of 0.1 second per
well. The IC50 value was determined from the dose response curve by using nonlinear regression
analysis (sigmoidal curve fit) of log transformed concentration data using Prism 6.03 (GraphPad
Software, La Jolla, CA).
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

8
Animal Care and Use
Female CB17 SCID or CD-1 nu/nu mice between 4 and 6 weeks of age were cared for in accordance to
the Guide for the Care and Use of Laboratory Animals, 8th Edition. Mice were group-housed at an
AAALAC international-accredited facility in sterile ventilated micro-isolator (or static) housing on corn
cob bedding. All research protocols were approved by the Institutional Animal Care and Use
Committee. Animals had ad libitum access to pelleted feed and water via automatic watering system or
water bottle. Animals were maintained on a 12:12 hr light: dark cycle in rooms and had access to
enrichment opportunities. Tumor volume and animal weights were measured two or three times a
week. Tumor length and width were measured with electronic digital calipers. Tumor volume (mm3)
was calculated as (W2 X L)/2 where width (W) is defined as the smaller of the two measurements and
length (L) is defined as the larger of the two measurements. Subcutaneous efficacy experiments were
performed in a masked fashion with one individual measuring tumor volume and animal weights while a
separate individual prepared test articles and injected the animals.
Serial in vivo propagation of D317
Fragments of the D317 tumor (derived from a patient with a GBM and shown to express EGFRvIII) were
serially propagated in immunocompromised mice to maintain EGFRvIII expression (26). Animals
bearing D317 human GBM xenografts were euthanized and tumors removed under sterile conditions.
Tumors were aseptically sectioned into similar sized fragments that fit into 13 gauge implant trocars.
Tumors were implanted via trocar into the flanks of naïve CB-17/SCID mice.
Pharmacodynamics of AMG 595
Mice bearing subcutaneous D317 tumors were randomized into treatment groups based on tumor
volume. Animal cohorts received vehicle, control conjugate, or AMG 595 by intravenous injection
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

9
fourteen days after implantation. Forty hours post treatment, animals were euthanized to collect
tumors that were processed for immunohistochemistry assessment of phospho-histone H3 levels as a
measure of mitotic arrest. Results are reported as the number of positive phospho-histone H3 cells in
an area of 1 mm2 (phH3(+)/mm2). The average number of phospho-histone H3 positive cells per mm2 in
the treatment groups was compared to the number observed in the control treatment groups.
Efficacy of AMG 595 in subcutaneous U251vIII xenografts
Ten million U251vIII cells mixed with growth factor reduced matrigel (BD Biosciences, San Jose, CA) were
implanted subcutaneously into CD-1 nu/nu mice. Once tumor volumes were approximately 300 mm3
animals were randomized into treatment groups of ten animals each. Animals were administered a
single intravenous dose of either a non-targeted control conjugate Ab-MCC-DM1 at 14.4 mg/kg (250 μg
DM1/kg) or AMG 595 at 1.7, 5.6 or 17 mg/kg based on antibody dose (25, 82, and 250 μg DM1/kg,
respectively). Earlier experiments demonstrated no difference in the vehicle and control conjugate
groups (Supplementary Figure S1).
D317 subcutaneous xenograft AMG 595 dose response and re-treatment
CB-17/SCID mice bearing subcutaneous D317 tumors were measured and once tumors reached
approximately 240 mm3 animals were randomized into treatment groups of ten animals each for
treatment initiation. Treatment was administered intravenously via the tail vein once weekly for four
injections. Animal cohorts received vehicle, a non-targeted control conjugate (Ab-MCC-DM1) at 26.8
mg/kg (375 μg DM1/kg), 7.3, 14.6, or 22 mg/kg of AMG 595 (125, 250, and 375 μg DM1/kg,
respectively). Five animals that achieved complete regression following initial treatment that recurred
were re-treated with two doses of AMG 595, administered one week apart on days 94 and 101.
Ex vivo EGFRvIII expression analysis by flow cytometry
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

10
Three animals bearing D317 subcutaneous xenografts treated with the initial four doses of AMG 595
were euthanized and tumors that re-grew were excised. Tumors were minced with scissors and passed
through a cell screen prior to incubation with red blood cell lysis buffer (Sigma, St. Louis, MO) for 10
minutes, followed by centrifugation. Tumor cells were counted and then dual stained with either a
human IgG1 control antibody or human IgG1 anti-EGFRvIII antibody and a mouse IgG1 kappa antibody
(BD Biosciences, San Jose, CA) or mouse anti-human HLA-ABC antibody (BD Biosciences, San Jose, CA) at
4 C for approximately 30-40 minutes. Cells were washed and incubated for approximately 30-40
minutes at 4 C with an anti-human IgG-Alexa 647 conjugated antibody and an anti-mouse IgG-Alexa 488
conjugated antibody (Grand Island, NY). Cells were analyzed on a FACSCalibur (BD Biosciences, San Jose,
CA).
Efficacy of AMG 595 in orthotopic xenograft
CB-17/SCID mice were exposed to whole body irradiation with a γ-source (1.44 Gy). Three days later
animals were stereotactically implanted into the right frontal lobe with 1x105 D317 cells. Animal cohorts
of eight animals each received vehicle or 22 mg/kg (375 μg DM1/kg) AMG 595 by intravenous injection
three days after D317 orthotopic implantation. Treatment was administered twice weekly for two
weeks. Twelve and twenty days following implant animals tumors T2-weighted images were collected
using magnetic resonance imaging (MRI) with a 4.7 Tesla magnet.
Statistics
Group comparisons for pharmacodynamic data were performed using the Mann-Whitney test using
GraphPad Prism version 6.03.
Group tumor volumes are shown as means plus or minus standard error of the mean and plotted as a
function of measurement time post implantation. Statistical significance of observed differences
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

11
between growth curves was evaluated by repeated measures analysis of covariance of the log
transformed tumor volume data with Dunnett adjusted multiple comparisons post test. Efficacy
statistical calculations were made through the use of JMP software v7.0 interfaced with SAS v9.1 (SAS
Institute, Inc., Cary, NC).
Results:
Generation of AMG 595
To generate a specific anti-EGFRvIII fully human antibody Xenomouse® animals were immunized with
the EGFRvIII peptide-KLH or cells expressing EGFRvIII. Fully human antibodies were assessed for binding
to EGFRvIII and counter-screened for binding to wild-type EGFR. The lead antibody was selected based
on its binding affinity for EGFRvIII (740 pM, 95% CI [550, 1000]), and lack of binding to wild-type EGFR.
This antibody was conjugated to DM1 with the non-cleavable linker MCC (Figure 1A) to generate AMG
595. Binding affinity of AMG 595 to EGFRvIII was 610 pM, 95% CI [440, 840], similar to that observed for
the unconjugated antibody. A chimeric form of the anti-EGFRvIII antibody was generated to stain
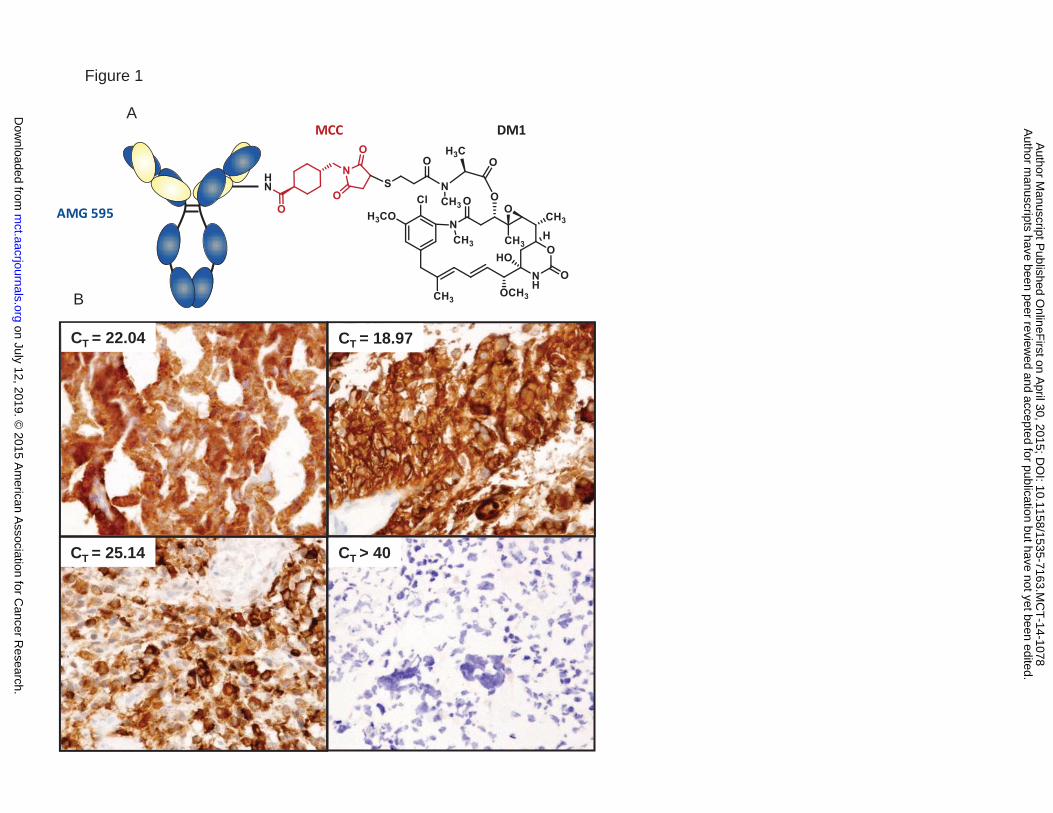
human tissue sections for EGFRvIII expression. Figure 1B shows representative tissue section staining
for EGFRvIII in GBM samples, five out of sixteen patient samples were positive for EGFRvIII expression by
IHC and qPCR. One sample was positive by IHC only, another was positive by qPCR only, whereas in the
remaining 14 sample EGFRvIII expression status was matched for qPCR and IHC (87.5% concordance)
demonstrating the EGFRvIII specificity of the anti-EGFRvIII antibody used in AMG 595.
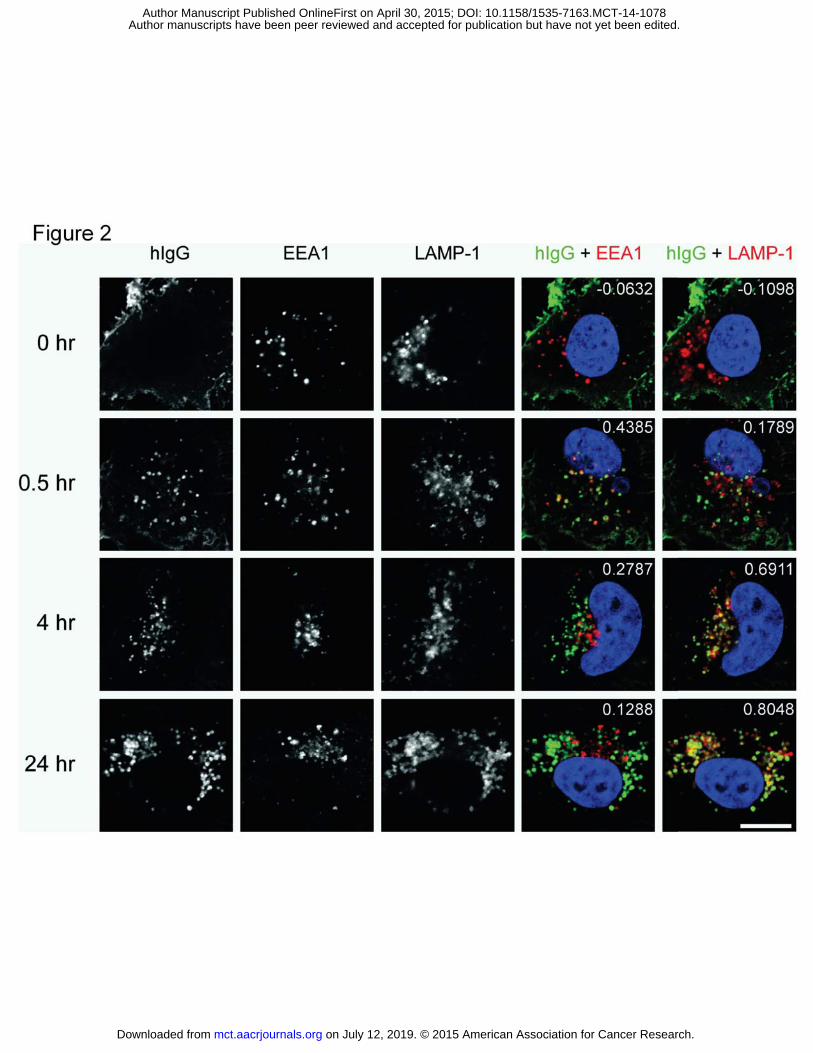
Internalization of AMG 595
The ability of AMG 595 to internalize into EGFRvIII expressing cells, and its sub-cellular localization was
explored in U251vIII cells, which had a CT value of 23.10, similar to that identified for the EGFRvIII
positive glioblastoma patient samples described above. AMG 595 bound to the cell surface of U251vIII
cells (Figure 2, top row) at 4 C, t=0 timepoint. A peripheral membrane pattern of staining was observed,
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

12
with no-localization with endosome or lysosomal markers. Upon elevating the temperature to 37 C for
0.5 hours, the staining shifted to a punctate pattern, with AMG 595 localized within EEA1+ early
endosomes (Figure 2). After 4 hours, most of the internalized AMG 595 had trafficked to LAMP1+
lysosomes as evidenced by the co-localization coefficient 0.69 and visually in Figure 2 (far right column).
At 24 hours, in the presence of LPI, internalized AMG 595 was still present in the lysosomes.
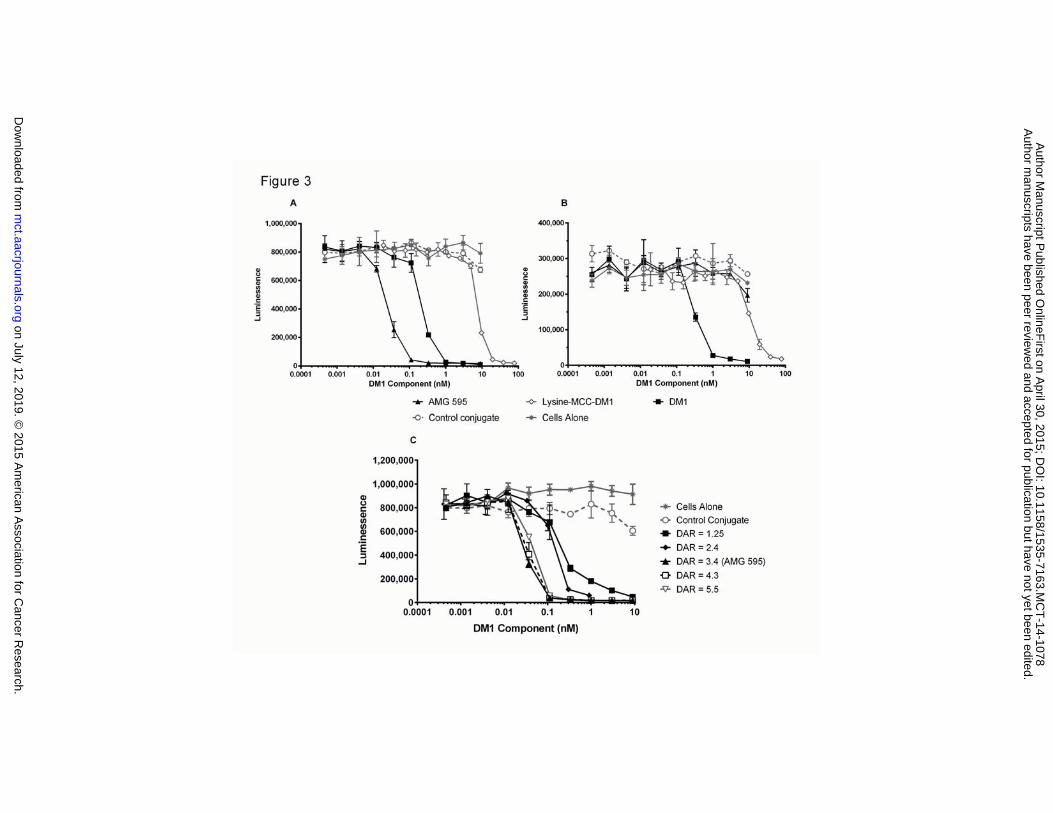
AMG 595 Potently Inhibits Cell Growth
Cell growth was assessed by measuring ATP levels which correlated with cell number. In U251vIII cells,
AMG 595 was more potent at killing the cells than DM1 alone with an IC 50 of 25 ± 3 pM , based on DM1
concentration, (Figure 3A). However in non-EGFRvIII expressing U251 cells AMG 595 did little to affect
growth and was similar to the effect observed with control conjugate (Figure 3B), this was also
demonstrated in A431 cells that overexpress wild-type EGFR (Supplementary Figure S2). In U251vIII and
U251 cells DM1 showed similar potency at ~ 0.25 nM. The IC50 of Lysine-MCC-DM1, the catabolite
formed in lysosomes of cells treated with AMG 595 or other non-cleavable linker DM1 ADC, was
approximately 300 fold less potent than AMG 595 in EGFRvIII expressing cells.
To define the optimal drug antibody ratio (DAR), anti-EGFRvIII antibody was conjugated with different
levels of DM1 ranging from 1.2 to 5.5 DM1 molecules per antibody and ability to kill EGFRvIII expressing
cells measured (Figure 3C). The two lowest DARs, 1.2 and 2.4, showed decreased cell growth inhibition
IC50 as compared to the higher DAR conjugates, which ranged from 3.4 to 5.5. With no improvement in
the IC50 at higher DAR values, the optimal DAR for AMG 595 was determined to be approximately 3.5.
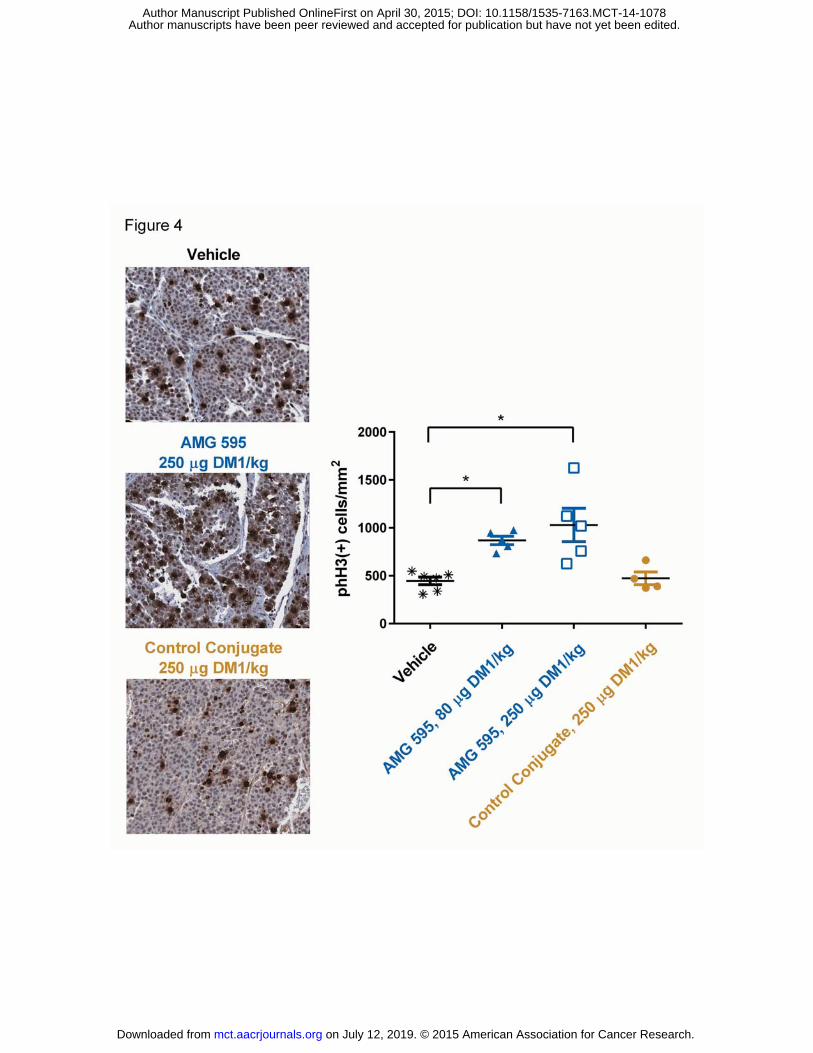
Pharmacodynamics of AMG 595
To assess the induction of mitotic arrest following treatment with AMG 595 in vivo, subcutaneous D317
tumors from AMG 595 or control treated animals were removed and assessed for levels of phospho-
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

13
histone H3 by immunohistochemistry. Tumors in animals treated with 5.3 mg/kg of AMG 595 (80 μg
DM1/kg) generated an average of 868 phospho-histone H3 positive cells/mm2, approximately two-fold
increase compared to vehicle (p = 0.0043) and control conjugate (p = 0.0159) as shown in Figure 4. At
the 16.7 mg/kg dose (250 μg DM1/kg), AMG 595 treatment also demonstrated an increased number of
phospho-histone H3 expressing cells compared to vehicle (p = 0.0043) and control conjugate (p =
0.0317).
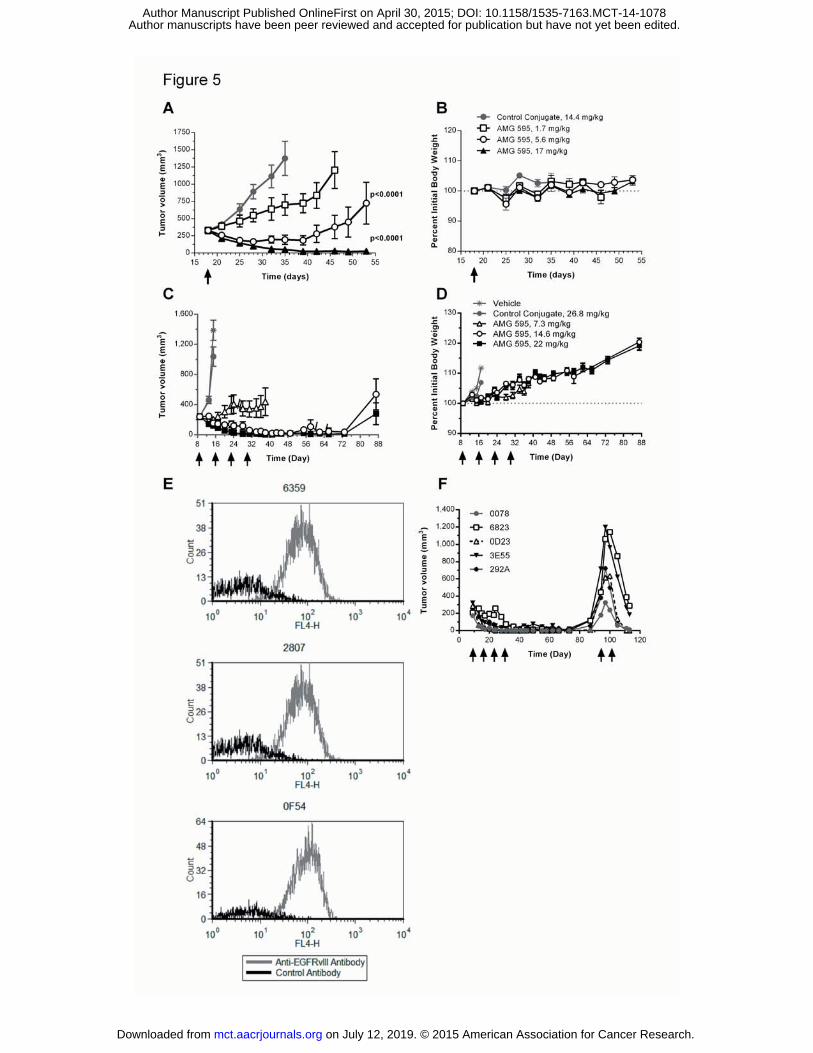
Efficacy of AMG 595 in subcutaneous U251vIII xenografts
Animals bearing U251vIII human GBM subcutaneous xenografts were randomized for treatment with
either AMG 595 or control conjugate. Tumors in animals treated with control conjugate grew from an
average of 328 ± 28 mm3 on day 18 to an average of 1374 ± 248 mm3 on day 35 post implantation
(Figure 5A). Although there was a trend for a delay in tumor growth rate mediated by the AMG 595 at
1.7 mg/kg (25 μg DM1/kg) dose compared to the control conjugate, it was not statistically significant (p
= 0.1164). AMG 595 significantly delayed tumor growth at the 5.6 mg/kg (82 μg DM1/kg) (p < 0.0001)
and the 17 mg/kg (250 μg DM1/kg) (p < 0.0001) dose levels compared to control conjugate. From
day 35 to 53 all animals treated with AMG 595 at 17 mg/kg possessed tumors less than 75 mm3 in size.
There was no significant percent difference in body weight in animals treated with AMG 595 at any dose
compared to control conjugate (Figure 5B). Serum concentrations of AMG 595 were dose proportional
from one to twenty-one days after administration (Supplementary Figure S3).
D317 subcutaneous xenograft AMG 595 dose response and re-treatment
To assess the ability of AMG 595 to impact tumors that endogenously express EGFRvIII, mice bearing
D317 xenografts were treated with either a dose response of AMG 595, control conjugate, or vehicle.
AMG 595 at the 7.3 mg/kg (125 μg DM1/kg) dose level inhibited tumor growth compared to both the
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

14
vehicle and control conjugate treated cohorts (p < 0.0001) (Figure 5C). Tumor regression was observed
in all animals treated with 14.6 and 22 mg/kg of AMG 595 (250 and 375 μg DM1/kg, respectively) (p <
0.0001). At the 22 mg/kg dose level seven of ten animals achieved complete regression. No body
weight loss was observed in animals throughout the course of the study (Figure 5D). After an initial
response to treatment with AMG 595 D317 tumors re-grew in some animals. To assess if tumors that
re-grew expressed EGFRvIII, tumors were removed from three animals, tumor disaggregates were
produced and analyzed by flow cytometry. Human tumor cells collected from the recurrent xenografts
expressed EGFRvIII (Figure 5E). To explore if recurrent tumors remained responsive to AMG 595 five
tumors that re-grew, ranging in size from 178 to 715 mm3, were re-treated with two doses of AMG 595
at 22 mg/kg (375 μg DM1/kg) starting 94 days following implant. The five individual tumor volumes are
shown in Figure 5F from initial treatment to re-treatment. Immediately following the first re-treatment
dose tumors continued to grow, however, following the second re-treatment dose the tumors were all
significantly smaller than their largest volume, three of which were unmeasurable 113 days following
implant.
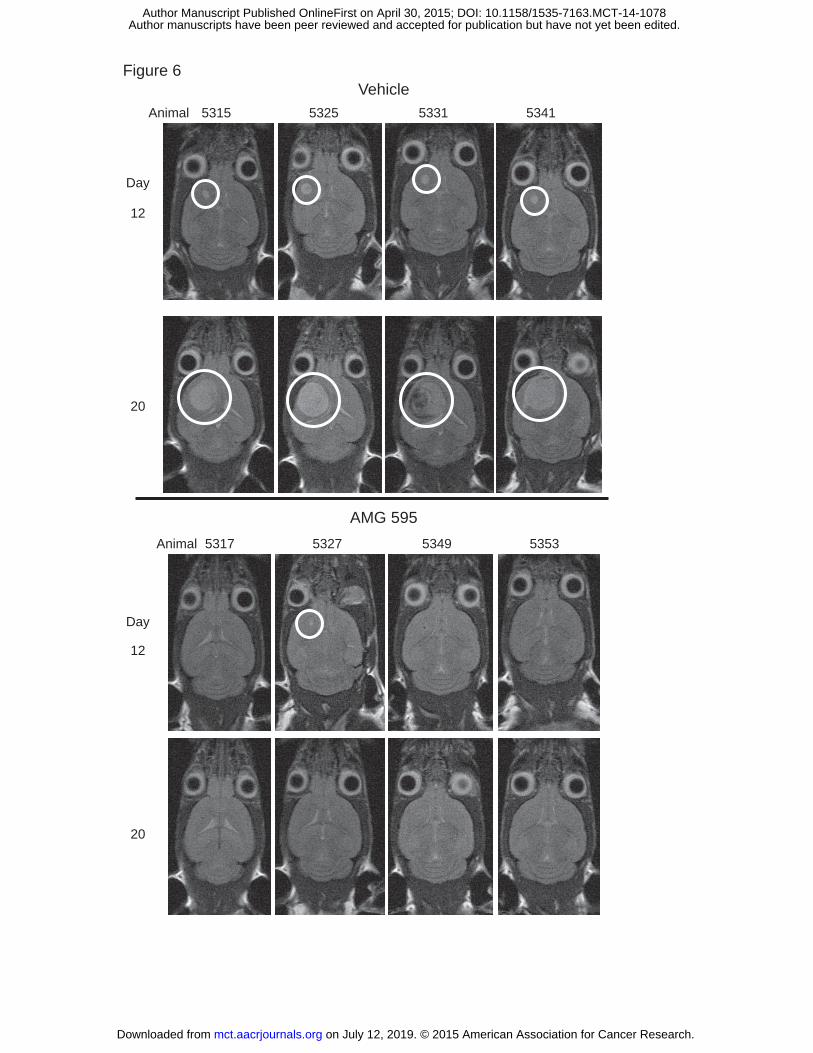
Efficacy of AMG 595 in orthotopic D317 GBM Model
An orthotopically implanted intracranial xenograft model of EGFRvIII expressing GBM was developed to
assess the ability of AMG 595 to mediate anti-tumor activity in a tumor growing in the brain. Three days
following stereotactic implant of D317 cells treatment was initiated with vehicle or AMG 595. All eight
vehicle treated animals had measurable tumors by MRI twelve days after treatment initiation (Figure 6).
Seven of eight animals treated with AMG 595 had no evidence of tumor in the brain on day 12. By day
20 tumor volumes increased in all vehicle treated animals, with two animals succumbing to disease prior
to the day 20 MRI image. In contrast, no tumors were observed by MRI in any AMG 595 treated animals
on day 20.
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

15
Discussion:
Exploiting molecular targets such as EGFR is one strategy to develop therapeutics with the potential to
improve survival in patients with GBM. Endogenous expression of EGFR on the skin and other tissues
represents a potential source of on-target toxicity or exaggerated phenomenon for an ADC mechanism
of action directed against wild-type EGFR. The EGFR deletion mutant EGFRvIII provides a bonafide
tumor specific antigen which could minimize any potential on-target toxicity effects that might be
observed with a wild-type EGFR targeted conjugate. Although EGFRvIII is only expressed in a subset of
GBM patients, it can be used as a biomarker to prospectively select patients for treatment moving
towards personalized therapy (32, 33). The antibody component of AMG 595 was shown to stain GBM
samples with a high level of correlation with EGFRvIII mRNA indicating a high degree of specificity for
the anti-EGFRvIII Ab used in AMG 595. The sample size of GBM samples examined here was too small to
provide a reliable estimate of EGFRvIII prevalence and was beyond the scope of the current study.
Robust internalization is required for non-cleavable linker ADC technology to be effective (34). The AMG
595/EGFRvIII complex rapidly internalized into the cellular endo-lysosomal pathway first localizing to
endosomes followed by time dependent localization in the lysosomes, similar to what has been reported
for other ADCs (31, 35, 36). Once in the lysosome, AMG 595 is catabolized into Lysine-MCC-DM1 (data
not shown), similar to other Ab-MCC-DM1 conjugates (37). The potency of the free warhead Lysine-
MCC-DM1 is much less than AMG 595 on EGFRvIII expressing cells, demonstrating that the catabolized
warhead will not lead to significant bystander killing of normal cells which was demonstrated to occur
when employing a cleavable linker by Kovtun et al. (38). A key factor in selecting a non-cleavable linker
for AMG 595 was to minimize potential bystander toxicity of normal cells in the brain.
Two glioblastoma models were evaluated in these experiments, U251vIII, a transfected cell line with
484,000 EGFRvIII receptors/cell and D317, a patient derived model with 199,000 EGFRvIII receptors/cell
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

16
(Supplementary Table S1). Endogenous EGFRvIII expression is difficult to maintain in standard cell
culture conditions (26, 27), which limited the evaluation of D317 to in vivo experiments. D317 human
GBM xenografts had a high basal level of phospho-histone H3 which was expected with a fast growing
tumor such as D317. U251vIII human GBM xenografts grew at a slower rate than D317 human GBM
xenografts and had significantly lower basal phospho-histone H3 levels (data not shown). The increased
phospho-histone H3 levels following AMG 595 treatment of D317 xenografts demonstrated the
induction of mitotic arrest, as expected, upon intracellular delivery of a maytansinoid, similar to the anti-
tubulin agent vinblastine (39). No dose response was observed with AMG 595 treatment, however, the
dynamic range of the assay was merely two-fold, to improve the sensitivity of the assay further
optimization of the assay itself and the collection time are likely required. Recent clinical studies testing
aurora kinase inhibitors included exploratory pharmacodynamic endpoints to assess phospho-histone h3
levels in normal skin (40, 41). Preclinical efficacy has been observed with several conjugates, whereas
translation into the clinic based on a maximally effective minimal dose has been difficult to determine
due largely to the lack of pharmacodynamic markers that can be employed in preclinical and clinical
studies. Incorporation of pharmacodynamic endpoints, such as phospho-histone H3 in the case of
tubulin inhibitor-based ADCs, into clinical trials may lead to improved preclinical to clinical translation.
Comparison of ADC preclinical efficacy is challenging with the different antibody targets, linkers,
warheads, tumor type, tumor burden pharmacokinetics and model relevance. The presence or absence
of bystander activity with cleavable or non-cleavable linkers and the warhead employed are arguably
the most essential factors in comparing ADCs. Only a handful of ADCs employing the MCC non-cleavable
linker attached to DM1 have entered clinical trials (42), of which only two have reported results, IMGN
529, reported preliminary data in a phase I study (43) and Kadcyla (25). Preclinical efficacy data was
reported in multiple models for trastuzumab-MCC-DM1 which required 15 mg/kg to generate
regressions as a single or multiple dose dependent on the model (44). AMG 595 in vivo dose levels to
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

17
generate regressions in two subcutaneous models is within 1.5-two-fold compared to trastuzumab-
MCC-DM1.
EGFRvIII expression in animals bearing D317 tumors that initially responded to treatment and later
recurred were shown to express EGFRvIII. Furthermore re-treatment of recurring tumors led to
significant reduction of tumor volume, demonstrating that tumors expressed EGFRvIII and remained
responsive to AMG 595 after initial treatment. In a Phase II clinical trial exploring the efficacy of an
EGFRvIII vaccine in GBM patients that expressed EGFRvIII, upon recurrence 9/11 patients were found to
lack EGFRvIII expression (45). Sampson et al. measured EGFRvIII expression by IHC, EGFRvIII has a
precise epitope and the lack of EGFRvIII staining by IHC could reflect either the presence of patient-
generated anti-EGFRvIII antibodies blocking the specific EGFRvIII epitope or GBM cells that lack EGFRvIII
expression as a result of either elimination of all EGFRvIII cells or treatment induced down-regulation of
EGFRvIII. Orthogonal methods to verify EGFRvIII protein expression in patients will be necessary to
understand the mechanism of this change in EGFRvIII expression following vaccine treatment. Detailed
analysis of clinical samples pre and post AMG 595 treatment will reveal if patients that respond to AMG
595 continue to express EGFRvIII upon recurrence, similar to what was observed preclinically.
The goal of the subcutaneous U251vIII and D317 models was to demonstrate that targeted delivery of
an anti-EGFRvIII ADC could eliminate EGFRvIII expressing tumor cells specifically. Despite the ability of
AMG 595 to generate regression in subcutaneous tumor models, one of the potential limitations of
treating brain tumors is the blood-brain-barrier significantly impairing the ability of large and small
molecules to access the tumor. In contrast to other diseases in the brain where the blood-brain-barrier
remains intact, initial de-bulking surgery, the subsequent radiation, and the tumor itself are thought to
compromise the blood-brain barrier (46). Reports of radiolabeled antibodies accumulating in the brain
such as the 806 antibody (20) suggest that in some cases a large molecule can reach tumors within the
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

18
brain. Orthotopic models of GBM in mice can’t completely mimic the human disease, however, they can
provide a closer approximation compared to subcutaneous xenograft models. Despite the approximate
molecular weight of AMG 595 at 150 kD, which is expected to be excluded from entry into the brain by
the blood-brain barrier, it was encouraging that AMG 595 mediated inhibition of tumor growth in the
D317 orthotopic GBM model. One of the limitations of this orthotopic D317 experiment was initiation
of AMG 595 treatment was three days following implant, which may only allow partial reformation of
the blood-brain-barrier for the first dose of AMG 595. The efficacy of AMG 595 in established orthotopic
tumors will be explored in a future publication. Ultimately, the only way to address if AMG 595 can
access GBM tumors in the brain is by testing the hypothesis in a carefully designed clinical trial.
The tumor-specific nature of EGFRvIII, internalization, and minimal potency as an unmodified anti-
EGFRvIII antibody make EGFRvIII an attractive target to explore with a selective ADC. In this report we
described the generation of an anti-EGFRvIII specific fully human ADC, AMG 595. AMG 595
demonstrated potent efficacy in vitro and in preclinical subcutaneous models/orthotopic model of GBM.
The ability to target EGFRvIII expressing GBM cells is an attractive option for patients and warrants
clinical exploration. Currently, AMG 595 is being tested in a phase I clinical trial enrolling patients shown
to have a GBM that expresses EGFRvIII, with early signs of efficacy (47).
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

19
Acknowledgements:
The authors thank Dr. Darell Bigner for providing the cell lines and patient tumor samples, Amgen
Burnaby for the generation of the anti-EGFRvIII antibody, ImmunoGen Inc. for supplying DM1, Chris Hale
for assistance in co-localization analysis, and Oncodesign for performing the orthotopic D317 GBM
experiment.
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

20
References
1. Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA: a cancer journal for clinicians. 2014;64:9-29. 2. Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, Janzer RC, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. The Lancet Oncology. 2009;10:459-66. 3. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. The New England journal of medicine. 2005;352:987-96. 4. Moustakas A, Kreisl TN. New treatment options in the management of glioblastoma multiforme: a focus on bevacizumab. OncoTargets and therapy. 2010;3:27-38. 5. Verhaak RG, Hoadley KA, Purdom E, Wang V, Qi Y, Wilkerson MD, et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer cell. 2010;17:98-110. 6. O'Dwyer P J, Benson AB, 3rd. Epidermal growth factor receptor-targeted therapy in colorectal cancer. Seminars in oncology. 2002;29:10-7. 7. Kalyankrishna S, Grandis JR. Epidermal growth factor receptor biology in head and neck cancer. Journal of clinical oncology : official journal of the American Society of Clinical Oncology. 2006;24:2666-72. 8. Bigner SH, Burger PC, Wong AJ, Werner MH, Hamilton SR, Muhlbaier LH, et al. Gene amplification in malignant human gliomas: clinical and histopathologic aspects. Journal of neuropathology and experimental neurology. 1988;47:191-205. 9. Jeuken J, Sijben A, Alenda C, Rijntjes J, Dekkers M, Boots-Sprenger S, et al. Robust detection of EGFR copy number changes and EGFR variant III: technical aspects and relevance for glioma diagnostics. Brain pathology. 2009;19:661-71. 10. Weller M, Kaulich K, Hentschel B, Felsberg J, Gramatzki D, Pietsch T, et al. Assessment and prognostic significance of the epidermal growth factor receptor vIII mutation in glioblastoma patients treated with concurrent and adjuvant temozolomide radiochemotherapy. International journal of cancer Journal international du cancer. 2014;134:2437-47. 11. Wong AJ, Ruppert JM, Bigner SH, Grzeschik CH, Humphrey PA, Bigner DS, et al. Structural alterations of the epidermal growth factor receptor gene in human gliomas. Proceedings of the National Academy of Sciences of the United States of America. 1992;89:2965-9. 12. Nicholas MK, Lukas RV, Jafri NF, Faoro L, Salgia R. Epidermal growth factor receptor - mediated signal transduction in the development and therapy of gliomas. Clinical cancer research : an official journal of the American Association for Cancer Research. 2006;12:7261-70. 13. Pedersen MW, Meltorn M, Damstrup L, Poulsen HS. The type III epidermal growth factor receptor mutation. Biological significance and potential target for anti-cancer therapy. Annals of oncology : official journal of the European Society for Medical Oncology / ESMO. 2001;12:745-60. 14. Wikstrand CJ, Reist CJ, Archer GE, Zalutsky MR, Bigner DD. The class III variant of the epidermal growth factor receptor (EGFRvIII): characterization and utilization as an immunotherapeutic target. Journal of neurovirology. 1998;4:148-58. 15. Biernat W, Huang H, Yokoo H, Kleihues P, Ohgaki H. Predominant expression of mutant EGFR (EGFRvIII) is rare in primary glioblastomas. Brain pathology. 2004;14:131-6.
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

21
16. Wikstrand CJ, Hale LP, Batra SK, Hill ML, Humphrey PA, Kurpad SN, et al. Monoclonal antibodies against EGFRvIII are tumor specific and react with breast and lung carcinomas and malignant gliomas. Cancer research. 1995;55:3140-8. 17. Yoshimoto K, Dang J, Zhu S, Nathanson D, Huang T, Dumont R, et al. Development of a real-time RT-PCR assay for detecting EGFRvIII in glioblastoma samples. Clinical cancer research : an official journal of the American Association for Cancer Research. 2008;14:488-93. 18. Mellinghoff IK, Wang MY, Vivanco I, Haas-Kogan DA, Zhu S, Dia EQ, et al. Molecular determinants of the response of glioblastomas to EGFR kinase inhibitors. The New England journal of medicine. 2005;353:2012-24. 19. Del Vecchio CA, Li G, Wong AJ. Targeting EGF receptor variant III: tumor-specific peptide vaccination for malignant gliomas. Expert review of vaccines. 2012;11:133-44. 20. Scott AM, Lee FT, Tebbutt N, Herbertson R, Gill SS, Liu Z, et al. A phase I clinical trial with monoclonal antibody ch806 targeting transitional state and mutant epidermal growth factor receptors. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:4071-6. 21. Niwa R, Sakurada M, Kobayashi Y, Uehara A, Matsushima K, Ueda R, et al. Enhanced natural killer cell binding and activation by low-fucose IgG1 antibody results in potent antibody-dependent cellular cytotoxicity induction at lower antigen density. Clinical cancer research : an official journal of the American Association for Cancer Research. 2005;11:2327-36. 22. Molhoj M, Crommer S, Brischwein K, Rau D, Sriskandarajah M, Hoffmann P, et al. CD19-/CD3-bispecific antibody of the BiTE class is far superior to tandem diabody with respect to redirected tumor cell lysis. Molecular immunology. 2007;44:1935-43. 23. Alley SC, Okeley NM, Senter PD. Antibody-drug conjugates: targeted drug delivery for cancer. Current opinion in chemical biology. 2010;14:529-37. 24. Senter PD, Sievers EL. The discovery and development of brentuximab vedotin for use in relapsed Hodgkin lymphoma and systemic anaplastic large cell lymphoma. Nature biotechnology. 2012;30:631-7. 25. Verma S, Miles D, Gianni L, Krop IE, Welslau M, Baselga J, et al. Trastuzumab emtansine for HER2-positive advanced breast cancer. The New England journal of medicine. 2012;367:1783-91. 26. Bigner SH, Humphrey PA, Wong AJ, Vogelstein B, Mark J, Friedman HS, et al. Characterization of the epidermal growth factor receptor in human glioma cell lines and xenografts. Cancer research. 1990;50:8017-22. 27. Pandita A, Aldape KD, Zadeh G, Guha A, James CD. Contrasting in vivo and in vitro fates of glioblastoma cell subpopulations with amplified EGFR. Genes, chromosomes & cancer. 2004;39:29-36. 28. Mendez MJ, Green LL, Corvalan JR, Jia XC, Maynard-Currie CE, Yang XD, et al. Functional transplant of megabase human immunoglobulin loci recapitulates human antibody response in mice. Nature genetics. 1997;15:146-56. 29. Babcook JS, Leslie KB, Olsen OA, Salmon RA, Schrader JW. A novel strategy for generating monoclonal antibodies from single, isolated lymphocytes producing antibodies of defined specificities. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:7843-8. 30. Chari RV, Martell BA, Gross JL, Cook SB, Shah SA, Blattler WA, et al. Immunoconjugates containing novel maytansinoids: promising anticancer drugs. Cancer research. 1992;52:127-31. 31. Polson AG, Yu SF, Elkins K, Zheng B, Clark S, Ingle GS, et al. Antibody-drug conjugates targeted to CD79 for the treatment of non-Hodgkin lymphoma. Blood. 2007;110:616-23. 32. Cloughesy TF, Cavenee WK, Mischel PS. Glioblastoma: from molecular pathology to targeted treatment. Annual review of pathology. 2014;9:1-25. 33. Olar A, Aldape KD. Using the molecular classification of glioblastoma to inform personalized treatment. The Journal of pathology. 2014;232:165-77.
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

22
34. Polson AG, Calemine-Fenaux J, Chan P, Chang W, Christensen E, Clark S, et al. Antibody-drug conjugates for the treatment of non-Hodgkin's lymphoma: target and linker-drug selection. Cancer research. 2009;69:2358-64. 35. Law CL, Gordon KA, Toki BE, Yamane AK, Hering MA, Cerveny CG, et al. Lymphocyte activation antigen CD70 expressed by renal cell carcinoma is a potential therapeutic target for anti-CD70 antibody-drug conjugates. Cancer research. 2006;66:2328-37. 36. Sutherland MS, Sanderson RJ, Gordon KA, Andreyka J, Cerveny CG, Yu C, et al. Lysosomal trafficking and cysteine protease metabolism confer target-specific cytotoxicity by peptide-linked anti-CD30-auristatin conjugates. The Journal of biological chemistry. 2006;281:10540-7. 37. Erickson HK, Park PU, Widdison WC, Kovtun YV, Garrett LM, Hoffman K, et al. Antibody-maytansinoid conjugates are activated in targeted cancer cells by lysosomal degradation and linker-dependent intracellular processing. Cancer research. 2006;66:4426-33. 38. Kovtun YV, Audette CA, Ye Y, Xie H, Ruberti MF, Phinney SJ, et al. Antibody-drug conjugates designed to eradicate tumors with homogeneous and heterogeneous expression of the target antigen. Cancer research. 2006;66:3214-21. 39. Juan G, Traganos F, James WM, Ray JM, Roberge M, Sauve DM, et al. Histone H3 phosphorylation and expression of cyclins A and B1 measured in individual cells during their progression through G2 and mitosis. Cytometry. 1998;32:71-7. 40. Dittrich C, Fridrik MA, Koenigsberg R, Lee C, Goeldner RG, Hilbert J, et al. A phase 1 dose escalation study of BI 831266, an inhibitor of Aurora kinase B, in patients with advanced solid tumors. Investigational new drugs. 2014. 41. Moreno L, Marshall LV, Pearson AD, Morland B, Elliott M, Campbell-Hewson Q, et al. A Phase I Trial of AT9283 (a Selective Inhibitor of Aurora Kinases) in Children and Adolescents with Solid Tumors: A Cancer Research UK Study. Clinical cancer research : an official journal of the American Association for Cancer Research. 2015;21:267-73. 42. Mullard A. Maturing antibody-drug conjugate pipeline hits 30. Nature reviews Drug discovery. 2013;12:329-32. 43. Stathis A, Freedman, A.S., Flinn, I.W., Maddocks, K.J., Weitman, S., Berdeja, J.G., Mejia, A.V., Zucca, E., Green, R., Romanelli, A., Zildjian, S.H., Ruiz-Soto, R., and Palomba, L. A Phase I Study of IMGN529, an Antibody-Drug Conjugate (ADC) Targeting CD37, in Adult Patients with Relapsed or Refractory B-Cell Non-Hodgkin’s Lymphoma (NHL). Blood. 2014;124:1760. 44. Lewis Phillips GD, Li G, Dugger DL, Crocker LM, Parsons KL, Mai E, et al. Targeting HER2-positive breast cancer with trastuzumab-DM1, an antibody-cytotoxic drug conjugate. Cancer research. 2008;68:9280-90. 45. Sampson JH, Heimberger AB, Archer GE, Aldape KD, Friedman AH, Friedman HS, et al. Immunologic escape after prolonged progression-free survival with epidermal growth factor receptor variant III peptide vaccination in patients with newly diagnosed glioblastoma. Journal of clinical oncology : official journal of the American Society of Clinical Oncology. 2010;28:4722-9. 46. Grossman SA, Batara JF. Current management of glioblastoma multiforme. Seminars in oncology. 2004;31:635-44. 47. Santostefano M, Engwall, M, Everds N, Guzman, R, Chow, V, Upreti, V, Hamblett, KJ, Vargas, HM, and Hill, JS. AMG 595: A novel ADC with therapeutic potential in glioblastoma. 35th Annual Meeting of the American College of Toxicology Program; 2014; Orlando, FL.
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

23
Figure Legends
Figure 1. A) Schematic representation of AMG 595 consisting of the anti-EGFRvIII antibody conjugated to
the maytansine derivative DM1 by the non-cleavable linker MCC. B) GBM patient samples analyzed for
EGFRvIII expression by qPCR (CT values) and immunohistochemistry.
Figure 2. Cell surface binding, internalization and time-dependent trafficking of AMG 595 to endosomes
and lysosomes in U251vIII cells. Cells were live-labeled with AMG 595 at 4 C to detect surface EGFRvIII, a
single representative cell is shown in each image (top row). Cells were incubated at 37 C for 30
minutes, 4 hours, and 24 hours and then fixed and permeabilized. Immunofluorescent labeling with
antibodies against human IgG (AlexaFluor 488), early endosome marker EEA1 (AlexaFluor 647) and
lysosomal marker LAMP-1 (AlexaFluor 555) was used to detect the internalized ADC as it moved through
the endo-lysosomal pathway. Co-localization images and coefficients are included in the two right
columns for hIgG + EEA1 and hIgG + LAMP-1.
Figure 3. A. AMG 595 Demonstrates Potent Cell Growth Inhibition of EGFRvIII Expressing Cells. A.
U251vIII and B. U251 cells were plated and exposed to media, AMG 595, control conjugate, DM1, or Lys-
MCC-DM1 for 96 hours. Viable cell number remaining in the cultures was assessed using Cell TiterGlo
and measurement of luminescence. C. DM1 was conjugated to anti-EGFRvIII antibody yielding
conjugates with drug antibody ratios from 1.25 to 5.5 and exposed to U251vIII cells for 96 hours. Cell
viability was assessed using Cell TiterGlo as in A and B. The data are expressed as the mean ± standard
error of the mean (SEM) for duplicate measurements (n = 2).
Figure 4 AMG 595 Induces Tumor Mitotic Arrest In Vivo As Measured By Increased Phospho-Histone H3.
Mice bearing D317 human GBM subcutaneous xenografts were treated with either vehicle, AMG 595 at
two dose levels 5.3 mg/kg and 16.7 mg/kg (80 and 250 μg DM1/kg, respectively), or a non-target control
conjugate at 16.7 mg/kg (250 μg DM1/kg). Forty hours after treatment tumors were collected and
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

24
sectioned for IHC analysis of phospho-histone H3. Representative 20X images are shown on the left
panel. Phospho-histone H3 quantification for each group is shown on the right panel. * p < 0.05 vs
vehicle as determined by Mann-Whitney test.
Figure 5 Regression of U251vIII and D317 Human GBM Subcutaneous Xenografts Following AMG 595
Treatment.
A & B) U251vIII human GBM cells were subcutaneously implanted into CD-1 nu/nu mice. Mice were
randomized into different groups (n=10) and treated with one of three dose levels of AMG 595: 1.7, 5.8,
and 17 mg/kg (25, 82, and 250 μg DM1/kg, respectively), or a non-targeted control conjugate at 14.4
mg/kg (250 μg DM1/kg). (A) Tumor volume (mm3) and Percent initial body weight (B) are shown. C-F)
D317 human GBM cells were subcutaneously implanted into CB-17/SCID mice. Mice were randomized
into cohorts of 10 mice each and treated with vehicle, 7.3, 14.6, and 22 mg/kg of AMG 595 (125, 250, or
375 μg DM1/kg, respectively) or 26.8 mg/kg of control conjugate (375 μg DM1/kg) control conjugate
once weekly for four weeks. C) Tumor volume, hash marks depict one animal euthanized due to large
tumor volume. D) Percent initial body weight. E) Three animals in which tumors re-grew had tumors
removed and assessed for EGFRvIII by flow cytometry (four digit code represents individual animal
numbers). F) Individual tumor volumes for five animals that had tumors re-grow following initial
treatment were re-treated with AMG 595, numbers denote the last four digits of identification chips.
(four digit code represents individual animal numbers). For A and C (tumor volume) and B and D
(percent initial body weight) data shown are mean ± SEM.
Figure 6:
Efficacy of AMG 595 in an EGFRvIII orthotopic model: CB17 SCID mice were stereotactically implanted
with D317 cells in the right frontal lobe. Three days after cell implant animals were treated with Vehicle
or AMG 595 (n=8/group). MRI was performed on study groups twelve and twenty days following
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

25
implantation. Four representative animals from each group are shown; tumors are indicated with the
white circles. Prior to day 20 (study end) two animals in the vehicle treated group succumbed to
disease.
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078
DM1MCC
AMG 595
CT = 22.04 CT = 18.97
CT = 25.14 CT > 40
A
B
Figure 1
on July 12, 2019. © 2015 A
merican A
ssociation for Cancer R
esearch. m
ct.aacrjournals.org D
ownloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on A
pril 30, 2015; DO
I: 10.1158/1535-7163.MC
T-14-1078
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078
on July 12, 2019. © 2015 A
merican A
ssociation for Cancer R
esearch. m
ct.aacrjournals.org D
ownloaded from
Author m
anuscripts have been peer reviewed and accepted for publication but have not yet been edited.
Author M
anuscript Published O
nlineFirst on A
pril 30, 2015; DO
I: 10.1158/1535-7163.MC
T-14-1078
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078
Day
12
20
Vehicle 5315 5325 5331 5341 Animal
Figure 6
AMG 595 5317 5327 5349 5353
Day
12
20
Animal
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078

Published OnlineFirst April 30, 2015.Mol Cancer Ther Kevin J Hamblett, Carl J. Kozlosky, Sophia Siu, et al. GlioblastomaPotent Anti-Tumor Activity Against EGFRvIII Expressing AMG 595, an anti-EGFRvIII Antibody Drug Conjugate, Induces
Updated version
10.1158/1535-7163.MCT-14-1078doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2015/05/02/1535-7163.MCT-14-1078.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/early/2015/04/30/1535-7163.MCT-14-1078To request permission to re-use all or part of this article, use this link
on July 12, 2019. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on April 30, 2015; DOI: 10.1158/1535-7163.MCT-14-1078





























