Embed Size (px)
Citation preview



www.isleroyalewolf.org
This document is printed on recycled paper, produced by a chlorine-free process.
Michigan Technological University is an equal opportunity educational institution/equal opportunity employer.
Ecological Studies of Wolves on Isle Royale
Annual Report 2006-2007*by
John A.Vucetichand
Rolf O. PetersonSchool of Forest Resources and Environmental Science
Michigan Technological UniversityHoughton, Michigan USA 49931-1295
31 March 2007*During the past year, major support for these studies was received from the National Park Service (Co-op Agreement No.
CA-6310-9-8001), National Science Foundation (DEB-0424562), Earthwatch, Inc., and the Robert Bateman endowment atthe Michigan Tech Fund.
Additional contributions were received from the following organizations and individuals: Dorthey L. Behrend, Sheri A.Buller, Alison J. Clarke, Donald C. Close, Greg Capito, Steven & Polly Eberhardt, Ronald & Barbara Eckoff, Edith N. Greene,John H. Heidtke, Mary Hindelang & Mark Silver, Horace & Mary Jackson, Dr. H. Robert Krear, Stephen & Deborah Laske,Jodi L. Louth, Dr. Brian E. McLaren, Richard & Bea Ann Murray, Michael & Kari Palmer, Janet L. Parker, Tony & ThelmaPeterle, Rolf & Carolyn Peterson, Joseph & Rose Potvin, Marcel Potvin & Anne Chouinard,Valerie R. Quant, Nathaniel P.Reed, John & Linda Schakenbach, Fred & Joyce Scharringhausen, Beverly Stencel,William & Cynthia Veresh, John Weisel &Johanna Fine, and Dorothy Zeller.
Unless otherwise noted, all photographs are by Rolf O. Peterson or John A.Vucetich.
Important contributions and personal time and financial assistance from the following Earthwatch volunteers are grate-fully acknowledged:
Team I—No team this year.Team IIA—Tim Pacey (leader), Paul Janovicz, Barrett Warming, Erik Freeman, Anna JanoviczTeam IIB—Rolf and Candy Peterson (leaders), Ron Eckoff, Kim Thomas, Mike ThomasTeam III—Jeff Holden (leader), David Norris, Thom Giomi, Clay Ecklund, Chuck MinkTeam IVA—Matt Abbotts (leader), Alex Jones, Steve Gibson, John Agee, Laura Boswell, Kathy WestTeam IVB—Rolf Peterson (leader), Saenger Ellis, Michelle Girts, Katie Crowell, Danica Dahlquist, Sue Dell
Tax-deductible donations to support continuing research on Isle Royale wolves and moose can be sent to Wolf-MooseStudy, Michigan Tech Fund, Michigan Technological University, 1400 Townsend Drive, Houghton, Michigan 49931-1295.THANK YOU to all who help!
Results reported here are preliminary and, in some cases, represent findings of collaborators; please do not cite withoutconsulting the authors.
5/07

2
Ecological Studiesof
Wolves on Isle Royale
“And the end of all our exploring will be to arrive where we started and know the place forthe first time.”
—T.S. Eliot
Personnel and LogisticsIn summer 2006, Rolf Peterson and John Vucetich
directed ground-based fieldwork, aided by MattAbbotts, Sam Gardner, Marcy Erickson, Dan Graham,Melissa Watkins, Erin Largent, Carolyn Peterson, andLeah Vucetich. Fieldwork continued from early Maythrough late October. In 2007, the annual winter studyextended from January 15 to March 5. Because temper-atures were unseasonably warm during late Decemberand early January, ice conditions were very uncertainjust prior to the start of the winter field season. Toaccommodate the uncertain conditions, the U.S. CoastGuard (Lt. Mike Baker, pilot; Lt. Shawn Geragthy, co-pilot; Chief Warrant Officer Jeff Hansen) flew MarkRomanski and Rolf Peterson to the island by helicopteron January 11. They found only 1 in. of ice on Washing-ton Harbor, but the research aircraft (PA-18 Supercub)
was able to land safely on 4 in. of ice by January 15. Theresearch aircraft was used to ferry arriving and depart-ing people and supplies from Feldtmann Lake (13 in.ice) to Washington Harbor until January 23, when icethickness had increased to 12 in. Rolf Peterson, JohnVucetich, and pilot Don E. Glaser participated in theentire study, assisted in the field by Leah Vucetich andthe following personnel from Isle Royale National Park:Mark Romanski, Valena Hofman, Beth Kolb, DieterWeise, Cindy Glase, and Marshall Plumer. U.S. ForestService pilots Wayne Erickson, Dean Lee, and Pat Loweflew several supply flights to Isle Royale from Min-nesota. George Desort participated in the winter study,shooting still photos and video. During the course ofthe year, many park staff and visitors contributed keyobservations and reports of wolves and moose bones.

Figure 1. Wolf and moose fluctuations, Isle Royale National Park, 1959-2007. Moose population estimates during 1959-1993 werebased on population reconstruction from recoveries of dead moose,whereas estimates from 1994-2007 were based on aerial surveys.
3
During 2006-2007, the wolf population dropped from 30to 21, while the moose population declined from 450 toabout 385 (fig. 1). The ratio of moose to wolves remainsvery low, ~20 to 1. Three territorial wolf packs raised atotal of four pups, and 43% of last year’s wolves died.An additional group of two or three wolves formed atleast temporarily in the middle of the island, exhibitingsome territorial behavior and killing moose on theirown.Wolves killed at a relatively low rate (on a per wolf
basis), and consumed a greater proportion of what theykilled than is usual. Moose calves were relatively rarethis winter, and mortality rates among moose were high(~3.6% per month). Spring 2006 was the fourth year ina row that moose suffered from heavy loads of moosetick. The summer was also particularly hot (along withthree of the four previous summers). Hot summers neg-atively impact moose because moose respond to hotweather by foraging less.
The Wolf PopulationDuring the 2007 winter study, the wolf population con-
tained 21 individuals, a 30% decline from last year’s 30wolves (fig. 1). The decline is ultimately attributable tolack of food. This challenge for wolves was compoundedby a lack of snow, which makes moose more difficult tokill. The wolf population was comprised of the samethree packs that have existed since 2000. Maximumpack sizes for all three packs were observed during aneight-day period, and this is our best estimate of popu-lation size. (We assume that all wolves were counted inone of the packs during this period.) Our count was:
East Pack III (EP) . . . . . . . . . . . . . . . . . . . . . .6Chippewa Harbor Pack (CHP) . . . . . . . . . . .7Middle Pack II (MP) . . . . . . . . . . . . . . . . . . . .8 2007 Total . . . . . . . . . . . . . . . . . . . . . . . . . .21
Significant this winter study were the poor conditionsfor aerial observations, which arose from a combinationof factors. First, only one wolf (in East Pack) wore a func-tioning radio collar. Second, because snow depths werevery low, wolves were able to walk anywhere in the for-est, so their travel was not concentrated as usual onlakes and shorelines. Third, when wolves did walk onfrozen lakes, they did not leave tracks because of lackof snow. Fourth, for about one month, we received nosnowfall. Consequently, the snow-covered landscapeaccumulated a dense maze of tracks that made it diffi-cult to locate wolves or determine their travel routes.Finally, many days of high winds reduced the time wecould spend flying.
Moose-Wolf Populations1959–2007
Wol
ves
Moo
se
1955 1965 1975 1985 1995 2005
2500
2000
1500
1000
500
0
50
40
30
20
10
0
Summary
4
We observed Middle Pack only twice(once with five wolves and once witheight wolves). CHP was observed eighttimes (on seven days), and only oncewith seven wolves. Although weobserved the radio-collared East Packmany times, they were usually in the for-est and we were often unable to makecomplete counts of the pack. Twice weobserved East Pack with six wolves, andfive times we observed five wolves inthis pack. We observed loners on just afew occasions, including one wolf with a“rope tail” (fig. 2). Rope-tailed wolveswere very common in the East Pack inthe 1970s, but since then they rarelyappear. Because of these poor circum-stances, we will rely on DNA analysis ofscats to confirm our winter count.
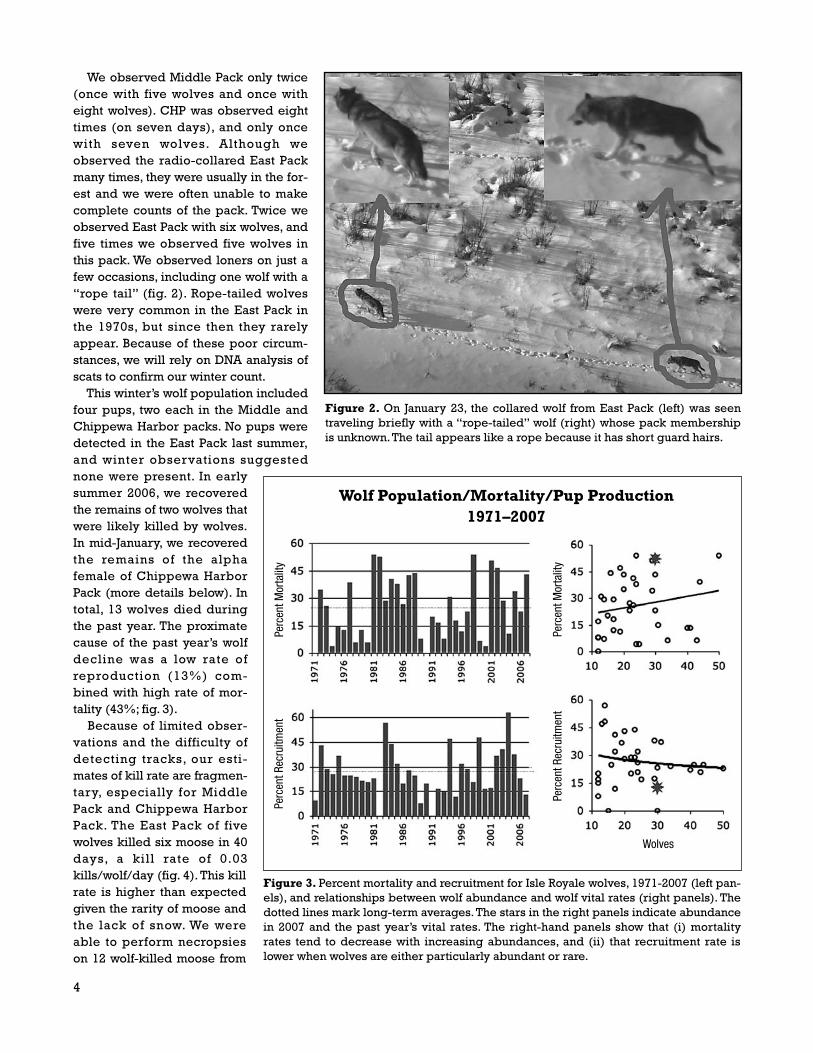
This winter’s wolf population includedfour pups, two each in the Middle andChippewa Harbor packs. No pups weredetected in the East Pack last summer,and winter observations suggestednone were present. In earlysummer 2006, we recoveredthe remains of two wolves thatwere likely killed by wolves.In mid-January, we recoveredthe remains of the alphafemale of Chippewa HarborPack (more details below). Intotal, 13 wolves died duringthe past year. The proximatecause of the past year’s wolfdecline was a low rate ofreproduction (13%) com-bined with high rate of mor-tality (43%; fig. 3).
Because of limited obser-vations and the difficulty ofdetecting tracks, our esti-mates of kill rate are fragmen-tary, especially for MiddlePack and Chippewa HarborPack. The East Pack of fivewolves killed six moose in 40days, a kill rate of 0.03kills/wolf/day (fig. 4). This killrate is higher than expectedgiven the rarity of moose andthe lack of snow. We wereable to perform necropsieson 12 wolf-killed moose from
Figure 3. Percent mortality and recruitment for Isle Royale wolves, 1971-2007 (left pan-els), and relationships between wolf abundance and wolf vital rates (right panels). Thedotted lines mark long-term averages. The stars in the right panels indicate abundancein 2007 and the past year’s vital rates. The right-hand panels show that (i) mortalityrates tend to decrease with increasing abundances, and (ii) that recruitment rate islower when wolves are either particularly abundant or rare.
Figure 2. On January 23, the collared wolf from East Pack (left) was seentraveling briefly with a “rope-tailed” wolf (right) whose pack membershipis unknown. The tail appears like a rope because it has short guard hairs.
Perc
ent R
ecru
itmen
t
Wolves
Perc
ent R
ecru
itmen
tPe
rcen
t Mor
talit
y
Perc
ent M
orta
lity
Wolf Population/Mortality/Pup Production1971–2007

all three packs. Typically, wolf consumption wasextremely high (fig. 5). From some adult moose kills wecould find only portions of two or three bones—all thehide, hooves, teeth, and even skulls were completelyconsumed. Such complete consumption is quite rare.
This year, we observed several significant biologicalpatterns among the wolves. First, the turnover in packleadership that began two years ago continued.The alphamale of Middle Pack died sometime between late Sep-tember and mid-January. He was probably at leasteight–ten years old and his pelage had become verywhite. He was last seen on the Feldtmann Ridge Trail lastOctober by Karena Schmidt, who works in our lab. Thealpha female of Middle Pack, wearing a nonfunctioningradio collar (1072), is still alive and at least eight years old.
The alpha male (and a founder) of Chippewa HarborPack was killed by the East Pack in January 2006. Inmid-January of 2007, we found and collected theremains of this pack’s other founder, the radio-collaredalpha female (and founder). She was likely betweennine and eleven years old when she died. She died onthe north shore of Angleworm Lake sometime in mid-December. The location of her death, just outside her
recent territory, strongly suggests that she was killedby East Pack. Moreover, we observed East Pack visit thesite of her death several times. The life of this alphafemale is recalled in the sidebar on page 7.
The East Pack is led by a young female and large andold male (fig. 6). The alpha female began leading Eastpack in fall 2005 and reproduced for the first time inspring 2006.The alpha male of East Pack, radio-collared in2001 and now probably about 10 years old, is notexpected to live more than another year or two (fig. 7).Weobserved the alpha male tolerating courtship advancesby younger males toward two females in winter 2007.
That second significant pattern observed this winterwas a continuing shift in territorial boundaries betweenEast Pack and Chippewa Harbor Pack. In previousyears, this territorial boundary ran from Daisy Farm toMcCargo Cove. Last year, East Pack showed signs ofdepleting the moose in the eastern portion of their ter-ritory, and subsequently challenged Chippewa HarborPack over a significant portion of their territory. Thisyear, East Pack appeared to have complete control overareas formerly used by the Chippewa Harbor Pack onthe north side of the island (fig. 8).
Figure 4. Members of the East Pack feed on an adult cowkilled 20 minutes before. Aside from stomach contents, theentire moose was consumed, including the skull, hooves,leg bones, and almost all the teeth.
Figure 5. All that remained of this wolf-killed adult moosewere a few bone fragments.
Figure 6. East Pack investigates a scent mark depositedby Chippewa Harbor Pack at Baker Point. The large wolvesare the alpha male and the alpha female (third and secondfrom left, respectively).
Figure 7. The old alpha male from the East Pack spent alot of time bedded in 2007, but he was still a key leader inthe pack. He had an infection or injury near his left eye.
5

6
The third significant change we observed this winterwas the possible beginnings of a new pack in the mid-dle of the island. We regularly observed tracks of asmall group of two to three wolves in this area, and twowolves were observed scent-marking at the southwestend of Hatchet Lake in mid-January. This group killed atleast three moose in about ten days in an area justsouthwest of Harvey Lake. One of these kills wasaggressively taken over by the Chippewa Harbor Pack.The very next day, January 29, we observed MiddlePack on Lake Desor with eight wolves (three more thanour observation from several days earlier). For this rea-son, and because this small group used a portion of theisland that had traditionally been Middle Pack terri-tory, it is likely that this group had its origin in the Mid-dle Pack. In time, DNA analysis will confirm the identityof these wolves, and whether they succeed in becom-ing established as a reproducing pack.
We did not observe any copulation in 2007, butestrous blood on the snow or courtship behavior wasobserved in all three packs during February 10-19,(fig. 9). On this basis, we expect pups to be born in allthree packs this year.
Fearless wolves were frequently reported by visitorsduring summer 2007, at a higher rate than in recentyears when the phenomenon first became evident.Wolves killed a cow and calf on Minong Island in TobinHarbor in late June, and to access this island they oftentraveled through heavily peopled Rock Harbor. Visitorswatched wolves chase a fox down the asphalt lane inRock Harbor, and wolves reportedly trailed backpack-ers on trails and confronted people in campgrounds.The number of wolves observed by Vucetich and Peter-son on the ground in summer approximately tripled in2007. Significantly, no aggressive behavior of wolveswas reported, just curiosity and lack of fear. We are notaware that wolves obtained any food from people, andvisitors were routinely warned by park staff that anyfeeding of wolves was illegal and dangerous.
The continuing loss of six long-term pack leadersmay mean that the stable pack organization of 2000-2007 will change as new breeding wolves arerecruited. The food supply for wolves has certainlydeclined in recent years as old moose, born in theearly 1990s, are culled by wolf predation. Continuingdecline in the wolf population is expected.
Figure 9. In the East Pack, a subordinate male courts a consenting alpha female (left) while the alpha male consorts withanother female (right).
Figure 8. Wolf pack territorial boundaries and moose carcasses (all fresh kills) during the winter study in 2007.
Wolf Pack Territories and Kill Locations2007
MP IICHP
EP III

On the first research flight in2007, the radio signal from alphafemale 410 in the Chippewa Har-bor Pack was on mortality mode.The next day we recovered herradio collar and skull from theheavily scavenged site on thenorth side of Angleworm Lake,and in the next week the East Packmade regular forays to the site.The East Pack had probably killed410, as she died on the disputedterritorial border between thepacks, much as the ChippewaHarbor alpha male died in Febru-ary 2006. Early in 2006, laboratorywork confirmed that alpha female410 in the Chippewa Harbor Packwas the same wolf that was chasedinto Lake Superior, attackedseverely, and left for dead by theMiddle Pack in February 2000.When live-captured in 2003, thisalpha female had just borne pupsand was in excellent conditioneven though one of her canineteeth had broken off.
The story of 410 and her mate isone of the most dramatic talescoming from over four decades ofwolf research on Isle Royale. Wethink of it as an unusually com-pelling story, but probably all oldwolves would have an equallyunlikely story, if we only knew.
On February 18, 2000, we fol-lowed the running tracks of theMiddle Pack for six miles as theychased a foreign wolf…
“…We caught up to the 11-member pack at water’s edge justbefore noon, vigorously shakingthemselves dry, while 20 feet outin Lake Superior, standing on asubmerged rock in 10 inches ofwater, stood a bedraggled wolf,cowering, its hindquarters almostunderwater.
“During the next hour we cir-cled overhead. After several min-utes of rolling in the snow to dryoff, Middle Pack wolves either layin the snow, watching the victim inthe lake, or strutting stiffly backand forth along the shore in frontof the hapless wolf. Suddenly, inquick succession, three wolves
jumped into shallow water andleaped for the rock where theirquarry stood quivering. Con-fronted by this snarling trio, itfought for its life, snapping furi-ously toward the lunging packmembers. The lone wolf wasforced backward into neck-deep
7
The End of the Story Rolf Peterson
In 2000, a female wolf sought refuge in Lake Superior during an attack bythe Middle Pack, but was beaten thoroughly and left for dead.
A Middle Pack male roused the female, and they eventually founded theChippewa Harbor Pack.
Continued on page 11

Over the past five years, mooseabundance has declined by abouttwo-thirds (i.e., in 2002 mooseabundance was 1100). The moosecensus in 2007 was severely ham-pered by incomplete snow cover.After evaluating census resultstogether with conditions known toaffect moose abundance, we con-clude that moose probablydeclined further from 2006 to2007.
Last year, in 2006, we counted92 moose on 91 one-km2 plots,and estimated moose abundanceat 450 (with a 80% confidenceinterval of [370, 535]; fig. 1). In2007, we counted 51 moose on thesame 91 plots. These counts anddemographic considerations sug-gest that moose abundance wasbetween 310 and 460. A more pre-cise but less certain estimate formoose abundance is 385 moose(with an 80% confidence intervalof [270, 500]) (fig. 10). The justifi-cation for this estimate requires along explanation.
Typically, we assume we areable to see about 75% of themoose on a census plot, based on aprevious radiotelemetry study con-ducted on Isle Royale under aver-age winter conditions. If we takethis standard assumption and thenumber of moose we counted this
The Moose Population
8
Figure 11. Long-term trends (1959-2007) in percent of the total moose populationthat are eight-month-old calves (upper panel). The 49-year average (13.7%) ismarked by the light dotted line. The linear trend for the past 25 years is marked bythe heavy dotted line. The lower left panel shows that calf production is an impor-tant determinant of moose population dynamics. The lower right panel shows thatwolf abundance is not closely related to calf production. Calf production dependson the combined effects of many factors (predation, food, and climate), the detailscomplex and poorly understood. The stars in the lower panels represent condi-tions during the past year. Number of wolves is the average of the current year andprevious year. The lower panels include data from 1959-2007.
2007 Moose Distribution
Moose Growth Rate and Calf Production1959–2006
Figure 10. Moose distribution on Isle Royale in 2007 was relatively uniform, as it was in 2006. Only two strata were delin-eated, based on habitat types and results of the aerial counts on 91 plots that comprise 17 percent of the main island area.
Moo
se G
row
th R
ate
Perc
ent C
alve
s
Perc
ent C
alve
s
Percent Calves Number of Wolves

9
year, then the estimated moose abundanceis 310, a significant reduction (31%) fromlast year’s estimate of 450. However, condi-tions for counting moose were far belowaverage this winter. There was so littlesnow that some portions of the groundwere bare and brown (same color asmoose) and the conifers had no snow onthem (so the conifers were dark, just likemoose). Without access to radio-collaredmoose in the population, it is impossible toempirically assess sightability of themoose census. However, experience sug-gests that sightability could have been aslow as 50%. If we were to assume thatsightability was 50%, then the best esti-mate of moose abundance would be 460.For the moment, take 310 and 460 to be arange of plausible estimates for abun-dance.
Independent, demographic considera-tions provide additional basis for judgingmoose abundance. Only three of the 51moose we observed on plots in 2007 werecalves. Six percent recruitment is quitelow compared to the long-term average of14% (fig. 11). The explanation for low calfrecruitment is likely twofold: (i) Duringthe summer of 2006 there would havebeen between 21 and 30 wolves, and scatanalyses indicated they preyed primarilyon moose calves; (ii) the summer of 2006was extremely hot, probably causingmoose to forage less and gain weight lessquickly. This is likely the reason whycalves in 2006 (and 2005) appeared to beparticularly small. Small calves may beespecially vulnerable to wolf predation.
In addition to low recruitment, the mor-tality rate of adult moose is likely to havebeen average (i.e., ~14%) or somewhatabove average (fig. 12). Several factorsaffect moose mortality—food supply, win-ter severity, winter ticks, age structure,and wolf predation. During the past year,moose had plenty of food available tothem, although hot weather probablyreduced foraging rates. The winter of2006-2007 was extremely mild (from themoose’s perspective, this means low snowdepths), but moose suffered from heavyloads of moose ticks (figs. 13, 14). The agestructure of moose is probably dominatedby middle-aged moose, which tend to
Winter Predation Rates1974–2007
Figure 13.The extent of moose hair loss in spring, caused by winterticks, remained high for the fourth year in a row. Circles represent indi-vidual moose, heavy bars are annual averages, and smaller bars markinterquartile ranges.
Moose Hair Loss, 2001–2006
Figure 12. Winter predation rates for Isle Royale moose, 1974-2007(upper panel). Predation rate (number of predation events per livingmoose per month) is influenced by average snow depth (lower panel).
Prop
ortio
n of
Los
t or D
amag
ed H
air
Win
ter P
reda
tion
Rate
Mean Snow Depth (cm)
Win
ter P
reda
tion
Rate

10
have relatively high survival rates. Yet,for the past year there have been only15-20 moose per wolf, suggesting highpredation pressure. From this combina-tion of factors, we infer that mortalityrate is likely to have been average or abit higher.
These demographic considerations(low recruitment and normal to highadult mortality) suggest moose arelikely to have declined from last year’s450 moose. If recruitment was low (say6%) and mortality was average (i.e.,14%), then one would expect abun-dance to be 410 moose. If mortality werevery high, say 25%, then one wouldexpect abundance to be 360 moose.
Plausible point estimates for abun-dance are between 310 and 460 moose,based on census data, and between 360and 410 moose, based on demographicconsiderations. The midpoint of bothintervals is 385 moose, which seems areasonable point estimate for abun-dance. The 80% confidence intervalassociated with this point estimate, fromthe observed variance in moose countsamong survey plots and the negativebinomial distribution, is 270 and 500.Ultimately, a less precise, but more reli-able conclusion is that in 2007 mooseabundance probably did not increase,and more likely declined further.
All the dead moose we discovered inwinter 2007 were killed by wolves. Mar-row fat levels in these moose exhibitedthe typical range of values, based onprevious data (fig. 15).Wolves continuedto rely heavily on very old moose, bornin the early 1990s when the moose pop-ulation was large and expanding rap-idly. These old moose, with typicalpathologies (fig. 16), are becoming veryrare, and this change in age structureprobably underlies the decline in thewolf population.
The summertime weather in 2006 pre-sented challenges for moose with its hot temperaturesand continued drought. Many beaver ponds dried upcompletely. Although summertime observations sug-gested that a relatively high proportion of cow moosehad calves, wolf predation was intensive, and few calvessurvived to face their first winter (fig. 17).
The combination of warmer weather and high preda-tion pressure may lead to continued decline in themoose population, now at a historic low. A substantialdecline in the wolf population may be necessary beforethe moose population stabilizes.
Figure 14. In the winter and spring of 2006, most moose suffered substan-tially from moose ticks and the hair loss that they cause.
Figure 15. Long-term trends in bone-marrow fat for moose. The line foradults shows the proportion of adults with >70% fat in their bone marrow.The line for calves shows the mean value of percent fat in bone marrow.
Percent Bone-Marrow Fat of Moose, 1970–2006
Perc
ent
Year of Death

11
Figure 16. This skull is from a bull moose killed by theChippewa Harbor Pack. The palate of this bull decom-posed while the moose was alive, a result of a severe infec-tion beginning along the tooth row. The cavity and nasalpassage was impacted with a softball-sized wad of rottenvegetation.
Figure 17. A single wolf kept this cow moose and hercrippled calf in Lake Superior for at least a half day as theytraveled in tandem along the shoreline for over threemiles. The outcome of this standoff was unknown.
water, but it retained its footingand held the attackers at bay;they retreated to shore to shakeand roll again in the snow…”
The lone wolf lost this battle,and was left for dead on the icyedge of Lake Superior. But a trail-ing male from the Middle Packshowed up and took an interest inthe motionless victim, whichturned out to be a female wolf inheat. Before the day was out thefemale managed to rise andmake it 50m into the forest, whereshe lay all night as her bleedingwounds stabilized. The nextmorning we found that the maleand female had walked out of theimmediate area, and we pickedup a few hairs from her bloodybed. Five days later we found thepair a half-mile from the attacksite, the female standing shakilywhile the male alternately lickedthe wounds on her neck and fre-netically tried to court the female.
The following summer a pupwas born to the pair and theChippewa Harbor Pack wasestablished. Within two years thispack had killed two successive
alpha males in the adjacent EastPack as it systematically tookover a moose-rich portion of theeast end of the island. The saviourmale and courageous femalethrived, reproducing each yearand emerging as a territorialpowerhouse. We radio-collaredthe female in 2003, and DNA fromher blood matched that from thehair recovered from the attacksite in 1999.
The East Pack and ChippewaHarbor Pack both flourished aslong as there were plenty ofmoose. But by 2006, the East Packhad run short, and it began tomake inroads on the edges ofChippewa Pack territory. In Feb-
ruary 2006, the East Pack movedin on a recent kill of the neigh-boring pack. Finding theChippewa alpha male feeding,the East Pack promptly killedhim. Female 410 was then atgreater risk, but she recruited anew alpha male, mated, andraised another litter of pups in2006, her seventh successful sea-son of reproduction. Againstimpossible odds, she survivedand carved out, by wolf stan-dards, a highly successful life. Hercanine teeth were worn to smallstubs (see photo), one caninetooth was badly infected andloose, and in the end life was sim-ply wrung out of her.
The female died late in 2006, her worn teeth a testament to the rigors ofher life.
The End of the Story (continued from page 7)

Other WildlifeThe National Park Service conducts aerial surveys of
known osprey and bald eagle nests each summer. Afterregional declines in historic organochlorine pollutants inthe Lake Superior watershed, both species reestablishedat Isle Royale in the 1980s. Eagles and ospreys are bothpresent at low numbers, and factors limiting further
12
Figure 19. Indices of abundance for red foxes and snow-shoe hares on Isle Royale, 1974-2007. The hare index is thenumber of hares seen per 100 km of summer hiking. Thefox index is the number of foxes seen from the plane dur-ing winter study, the sum of the maximum number seen atkills, and the number seen otherwise per 100 hours offlight time. The hare population appears to cycle (albeitwith a variable amplitude) but the fox population fluctu-ates in a more complex manner and the fox populationappears not to cycle. Rather the fox population appears tobe in long-term decline.
Snowshoe Hare Population Density1974–2007
Forest VegetationThe forests of Isle Royale provide the context and
foundation for long-term fluctuations in wolves andmoose. The eastern, western, and middle regions ofIsle Royale exhibit important differences in their vege-tative communities. The middle of the island, burned in1936, currently provides little winter forage for moose.The west end of Isle Royale supports very old forestswith sparse but well-distributed balsam fir trees thatare either in the forest canopy or less than two meterstall. These shorter trees have been browsed intensivelyfor many years, and are prevented by herbivory fromgrowing into seed-producing trees. The canopy trees onthe island’s west end are gradually dying without beingreplaced. Currently, only about 20% of the canopy bal-sam fir trees that existed in 1988 are still standing (fig.18), and these survivors are projected to disappear by2011. The ultimate implications of this forest change arenot entirely clear. On a decadal time scale, moose arelikely to decline at least somewhat on the west end ofthe island as the young fir trees die without replace-ment. However, a moose decline might be mitigated, atleast partially, if moose switch to some other forage(e.g., cedar). Also, it may be that only a modest declinein moose is necessary to allow enough fir trees to reachinto the canopy. We have already noticed that fir treesin the west end have been growing better in recentyears, due to reduced moose abundance.
By contrast, the east end of Isle Royale is characterizedby dense stands of balsam fir of all size categories(seedlings, samplings, pole-sized trees, and canopy trees).Moreover, east-end balsam fir trees are only lightlybrowsed. Until recently, east-end forests have supportedgreater densities of Isle Royale moose, which in turn havesupported greater densities of Isle Royale wolves. More
specifically, in 2001-2007 two wolf packs (with ~2/3 of thewolf population) shared the eastern one third of the island,with about half of the moose population.
However, in recent years intense wolf predation on theeast end has reduced moose density substantially, out-weighing abundant forage as a moose population deter-minant. The Blake Point peninsula at the northeast endremains largely vacant of moose, and the East Pack hasshifted its attention in midwinter to other areas, includingthe territory formerly held by Chippewa Harbor Pack.
Balsam Fir Tree Abundance and Mortality
Figure 18. Abundance (filled circles) and annual mortalityrate (open circles) of balsam fir trees in the forest canopythat were tagged in 1988. These trees are not beingreplaced by younger fir trees. The remainder of these sam-pled trees is expected to die by approximately 2011, anindicator of a dramatic reduction over 75% of Isle Royalein seed source for this species. The demise of this speciesis ultimately caused by moose herbivory.
Prop
ortio
n of
Tre
es L
ivin
g
Annu
al M
orta
lity
Rate

expansion are poorly understood. In 2007, active eaglenests numbered seven, with six eaglets fledged.The num-ber of osprey nests was five, with three young fledged.
Snowshoe hare observations were still relatively lowin summer 2007, consistent with a cyclical decline fol-lowing a peak at the turn of the decade (fig. 19). Redfox, a major hare predator, have apparently declined toexceptionally low levels (fig. 19), probably a result oflow food from scavenged moose carcasses and a lowhare population (fig 20).
Active beaver colonies were estimated during 1974-2004 during biennial aerial surveys in autumn, con-ducted by Phil Shelton and Doug Smith (fig. 21). Afterreaching a historic peak in 1974, beaver declined, thenrecovered as wolves peaked and crashed. During 1986-2004, beaver numbers have slowly contracted as habitatdeteriorated in successional forests recovering fromfires in the nineteenth and early twentieth century. InOctober 2006, a double-count of beaver sites wasmade, for the first time, using two aircraft, with RolfPeterson and Mark Romanski as observers. The searchpattern was more intensive than in previous aerial sur-veys, and the number of active sites observed was morethan twice as great as in the 2004 survey. The highernumber reflects higher sightability rather than anincreasing population. A followup survey will be con-ducted in 2007 to better clarify the population statusand trend.
River otters continue to thrive in all parts of the island.The recently-established American marten is the sub-ject of a study by park staff during 2006-2008. Althoughno marten were reportedly observed in the past year, ascavenged half-carcass was recovered on a trail lastsummer, and fresh tracks of several individuals wererecorded in winter 2007 at the west end of Isle Royale.
Figure 21. Even with very low water and an aging forest, a few newly established beaver colonies could be found inautumn 2006.
13
Figure 20. Two scavenging foxes compete agonisticallyover the sparse remains of a wolf-killed moose.

14
June 5, 2006—I was awakened to thesound of moose grunts outside theyurt, at Windigo, where we live. Myalarm had not gone off, so it had to besometime before 6:00 AM. I was not pre-pared (mentally or logistically) to makeobservations on the foraging behaviorof this moose. However, I can’t seem toresist the sound of those burdenedgrunts! You cannot make these obser-vations when you want; they have to bemade when a moose decides to shareits time with you.
So I crawled over my comatose hus-band, John Vucetich, and sprung out ofbed. I pulled my baggie field pants onover my sleeping long undies, zippedon a fleece, and slipped on my redCrocs. Later, I realized that I pulled myfield pants on backwards. I dug my binocs out of yes-terday’s pack and grabbed my notebook.
A cow moose was right outside the yurt, toward thePark Service junkyard that we live next to. She was for-aging busily. The early morning light was still dim, somaking observations was difficult. Nevertheless, Irecorded several bites. As she moved deeper into theforest, I realized I was being swarmed with mosquitoesand blackflies, and I had forgotten an essential piece ofequipment—my bug shirt. I noted her position and lineof movement and headed back to the yurt. I changedinto socks, boots, bug shirt, baseball hat, gloves, andcitronella spray and headed back out. Despite theruckus, John showed no signs of awakening.
I listened, but heard nothing. I proceeded carefullyfrom where I had last seen her. Then I heard the crack ofa hoof on a twig, and the methodical munching of leaves.I began to record more foraging observations. As themorning light became brighter, I noticed her pattern ofhair loss (caused by winter ticks).The pattern was differ-ent from those which I had seen this year. This was ourfirst meeting, this cow moose and I. I thought I knew allthe moose who had been foraging in this area. Then Isaw something I recognized—a tear on her left ear. I hadmet this moose before—two years before. In the summerof 2004, she and I had spent quite a bit of time together.That year, she had a calf, and I helped a German filmcrew video-record her at a nearby mud lick.
It was great to see an old friend, and it made mesmile to know she was still doing well. She now carriesa tumor on her right shoulder (these skin tumors arecommon and typically benign). I think that’s new since
we last met (I’ll check my notes).She continued to forage, and I continued to record.
She is a moving browser—no standing in one spot andgorging. She takes a few steps and a few bites and afew steps. I’m sure this works for her, but it does makerecording a challenge. This morning she eats lots ofyellow birch leaves—both from branches on tall treesand from small suckers. Each birch bite is nearlymatched with bites of lady fern and Dryopteris, andsupplemented with hazel leaf here and a maple leafthere. After some time, she continued foraging, butbegan moving a bit more quickly—too quickly for meto handily follow, observe, and write notes. It is amaz-ing how swiftly they can move while still appearing tobe just meandering casually. I was disappointed thatshe was getting too far away from me to note her forag-ing.
Then she stopped suddenly and looked intently intothe forest beyond her. She stood stock still for severalseconds. Then she turned and bolted straight for me.In an instant, I felt that she was not charging me, ordisplaying any type of aggression toward me; but shewas beating a rapid retreat from something she hadheard, seen, or smelled ahead of her. She trottedtoward me—all 800 pounds of brown fur and muscle—and then past me, just an arm’s length to my left. Shespun around and I could feel her looking over myshoulder from just behind me.
There we both stood, both looking intently into theforest. Then I saw it—just a fleeting glimpse of thedark form of a wolf as it ran through the trees just
A Small Safe Haven Leah Vucetich
Continued on page 16
15
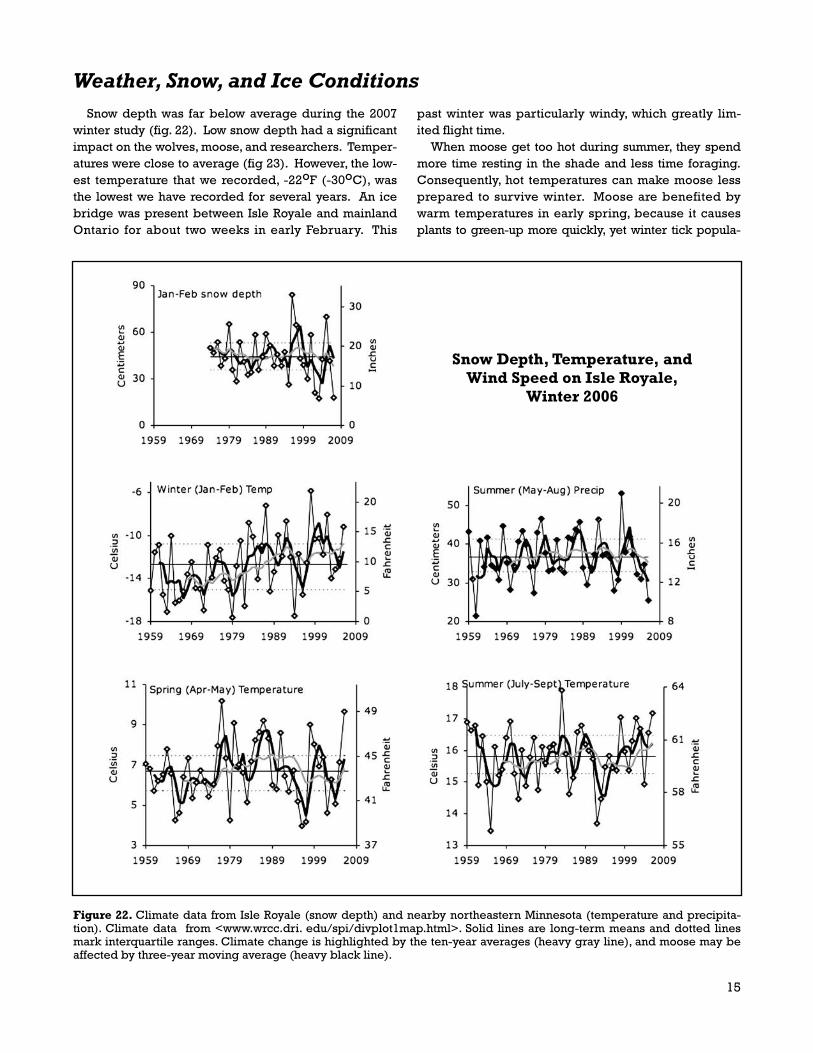
Figure 22. Climate data from Isle Royale (snow depth) and nearby northeastern Minnesota (temperature and precipita-tion). Climate data from <www.wrcc.dri. edu/spi/divplot1map.html>. Solid lines are long-term means and dotted linesmark interquartile ranges. Climate change is highlighted by the ten-year averages (heavy gray line), and moose may beaffected by three-year moving average (heavy black line).
Weather, Snow, and Ice ConditionsSnow depth was far below average during the 2007
winter study (fig. 22). Low snow depth had a significantimpact on the wolves, moose, and researchers. Temper-atures were close to average (fig 23). However, the low-est temperature that we recorded, -22oF (-30oC), wasthe lowest we have recorded for several years. An icebridge was present between Isle Royale and mainlandOntario for about two weeks in early February. This
past winter was particularly windy, which greatly lim-ited flight time.
When moose get too hot during summer, they spendmore time resting in the shade and less time foraging.Consequently, hot temperatures can make moose lessprepared to survive winter. Moose are benefited bywarm temperatures in early spring, because it causesplants to green-up more quickly, yet winter tick popula-
Snow Depth, Temperature, and Wind Speed on Isle Royale,
Winter 2006

16
beyond where she had been standing.After her danger had passed, we
stood near each other for what seemeda long time—perhaps two-three min-utes. I spoke with her and we examinedeach other carefully. I asked if sheremembered me from the mud lick. Sheleaned toward me, sniffing with earscocked forward toward me. I stretchedout my hand and spoke gently to her.She decided to move past me, and for-aged a bit more. She never stoppedlooking where she had seen danger.She was no longer relaxed andleisurely in her foraging. After a coupleof minutes, and just a few more bites,she left the area, walking too quicklyfor me to keep up.
I had been excluded from her mooseworld and was back into my own. Butwhere exactly was I? I had no map, nocompass, no GPS, no glasses or contacts, no sun, andhad been on a moose trajectory through the forest. Idecided to first look for tracks that may have beenleft by the wolf, but found no appropriate substrate torecord the passing. I began to walk in the direction I
thought was home. Eventually, I returned to the yurt,where John was still groggy—hadn’t heard the alarm,didn’t even know that I had been out, and had no ideathat a moose decided that I was the safest thing to putbetween her and a wolf!
Figure 23. Snow depth (daily) and ambient temperature (hourly) during the 2007winter study on Isle Royale.
tions seem to thrive under theseconditions. The effect of summerprecipitation is liable to beimportant, but its effect isunknown. The effect of summerprecipitation would be manifestthrough its impact on foragequality.
With this context, the summerof 2006 was extremely hot anddry. Moreover, seven of the pastnine summers have been hotterthan average, and the past fouryears have been drier thanusual (Fig. 22). This past spring,green-up came earlier thanusual. Although summer andwinter temperatures are warm-ing, there is little indication thatspring green-up is occurringearlier.
Temperature and Snow Depth
A Small Safe Haven (continued from page 15)
Tem
pera
ture
(C)
Tem
pera
ture
(F)
Snow
Dep
th (c
m)
Snow
Dep
th (i
n)































