Embed Size (px)
Citation preview

Ecologia de campo
2013
Foto: Luciana Zago da Silva

Os Programas de Pós-Graduação em Ecologia e Conservação daUniversidade Federal do Paraná e em Ecologia de Ecossistemas Aquáticosda Universidade Estadual de Maringá se uniram neste ano, para arealização de um curso conjunto de Ecologia de Campo. O curso ocorreuno período de 24 de junho a 4 de julho de 2013, na Base de Estudos doNúcleo de Pesquisas em Liminologia (NUPÉLIA), em Porto Rico, PR. Tendocomo pano de fundo o belo entardecer das margens do Rio Paraná, ocurso foi uma oportunidade única para troca de experiências deprofessores e estudantes de Curitiba e de Maringá. Os mini-projetosdesenvolvidos durante o período do curso despertaram para questõesecológicas inéditas e interessantes, demonstrando as potencialidadesdeste sistema para testar teorias ecológicas. Num clima de amizade etrabalho em grupo, foram produzidos vários relatórios, que aquiencontram-se compilados.
Porto Rico, 4 de julho de 2013.

Docentes PPGECO - UFPR
André Padial (Ecologia de macrófitas aquáticas)
Fernando Passos (Ecologia de mamíferos)
Márcia Marques (Ecologia vegetal)
Docentes PPGE - UEM
Luiz Gomes (Ecologia de peixes)
Sidnei M Thomaz (Ecologia de macrófitas aquáticas)
Roger Mormul (PDJ - Ecologia de macrófitas aquáticas)
Juliana Dias (PDJ - Zooplâncton)
Docentes colaboradores externos:
Marcos Barbeitos - (Ecologia Aquática) ZOO-UFPR
Discentes PPGECO - UFPR
Bianca Possamai
Emanuel Luis Razzolini
Fernanda Ceschin
Flávio Miranda Marteleto
Francisco W. von Hartenthal
Juliana Wojciechowski
Luciana Zago da Silva
Monica Vasques Mangas Pereira
Rafael Antunes Baggio
Renata Gabriela Noguchi
Sabrina Loise de Morais Calado
Sandra Marisa Nobre de Andrade
Simone Satsuki Takeuchi
Vinícius Marcílio da Silva
Discentes PPGE - UEM
Daniel Alves dos Santos
Diego Corrêa Alves
Érika Neif
Fernando Garcia de Oliveira
Hugo José Message
Vivan Cionek
Letícia Cucolo Karling
Lilian Paula Vasconcelos
Natália C. Lacerda dos Santos
Priscilla Guedes Gambale
Taíse Miranda Lopes
Guilherme Okuda

4
FernandoPassos Luiz
Gomes
Márcia

5
ConteúdoFATORES LOCAIS E REGIONAIS INFLUENCIANDO A ESTRUTURA DAS COMUNIDADES
VEGETAIS NA PLANÍCIE DE INUNDAÇÃO DO RIO PARANÁ........................................................... 7
INFLUÊNCIA DA DIVERSIDADE MORFOLÓGICA NA SEDIMENTAÇÃO DO FITOPLÂNCTON:
QUEM VENCE ESSA CORRIDA? ............................................................................................................. 13
RIQUEZA DE FORMIGAS EM AMBIENTES COM DIFERENTES GRAUS DE PERTURBAÇÃO 17
AÇÃO DE METABÓLITOS SECUNDÁRIOS DE PLANTAS NA PREDAÇÃO DE FORMIGAS
SOLENOPSIS SP. EM DIFERENTES ITENS ALIMENTARES ............................................................. 22
CONTRIBUIÇÃO DE ESPÉCIES ZOOPLANCTÔNICAS EM RIACHOS TRIBUTÁRIOS SOB
INFLUÊNCIA DE UM RIO DE INUNDAÇÃO.......................................................................................... 26
A DENSIDADE DE INVERTEBRADOS ASSOCIADOS À SERAPILHEIRA EM FRAGMENTOS
FLORESTAIS................................................................................................................................................ 31
ESCONDENDO A UTRICULARIA FOLIOSA: SERÁ QUE ENGANAMOS O ZOOPLÂNCTON? .. 36
PARTIÇÃO DE DIVERSIDADE EM MACROINVERTEBRADOS DE SERAPILHEIRA EM ILHA E
CONTINENTE .............................................................................................................................................. 41
EFEITO DA ALTERAÇÃO DA TURBIDEZ DA ÁGUA NAS COMUNIDADES DE
FITOPLÂNCTON E ZOOPLÂNCTON ..................................................................................................... 45
PREDAÇÃO SOBRE MODELOS APOSEMÁTICOS E CRÍPTICOS EM FRAGMENTOS ABERTOS E
FECHADOS ................................................................................................................................................... 50
DISPERSÃO DE FORMIGAS MEDIADA PELOS RECURSOS ALIMENTARES: THE ANTMATE
FIGHTING..................................................................................................................................................... 54
FOCO NA ESCALA: IMPORTÂNCIA DA HIERARQUIA DE ESCALA NA PARTIÇÃO DA
DIVERSIDADE BETA EM AUTÓTROFOS AQUÁTICOS..................................................................... 59
COMPOSIÇÃO SEXUAL DE GRUPOS DE ALOUATTA SPP.: DE ESTUDOS PONTUAIS À
PADRÕES...................................................................................................................................................... 65
RELAÇÃO DA COBERTURA VEGETAL DE EICCHORNIA AZUREA (KUNTH) COM AS
COMUNIDADES ÍCTICA E FITOPLANCTÔNICA................................................................................ 70
COMPLEXIDADE ESTRUTURAL E VOLUME DE RAÍZES DE MACROFITAS AQUÁTICAS
COMO PROMOTORAS DO ENRIQUECIMENTO DA COMUNIDADEDE INVERTEBRADOS
AQUÁTICOS................................................................................................................................................. 75
MIGRAÇÃO HORIZONTAL DE ZOOPLÂNCTON SOB ESTÍMULO DE PREDAÇÃO: UMA
ABORDAGEM EXPERIMENTAL............................................................................................................. 80

6
BIOGEOGRAFIA DE ILHAS: UMA ABORDAGEM UTILIZANDO MACRÓFITAS E
MACROINVERTEBRADOS AQUÁTICOS.............................................................................................. 85
EFEITO DA DISTÂNCIA DA MARGEM NA HERBIVORIA EM EICHHORNIA AZUREA.............. 89
ASSOCIAÇÃO SIMBIÓTICA ENTRE A TRIPLARIS AMERICANA E FORMIGAS
PSEUDOMYRMEX: DA OCORRÊNCIA À RESPOSTA A ESTÍMULOS ............................................. 94
INUNDAÇÕES COMO REGULADORAS DA DIVERSIDADE ZOOPLANCTÔNICA EM
PLANÍCIE DE INUNDAÇÃO ..................................................................................................................... 99
FORRAGEIO ÓTIMO: DETERMINAÇÃO DO TAMANHO DA CARGA EM FORMIGAS-
CORTADEIRAS (ATTA SP) ...................................................................................................................... 104
CONSPICUIDADE DE CORES: A VISUALIZAÇÃO NO PONTO DE VISTA DE MAMÍFEROS
PRONATOPIOS.......................................................................................................................................... 109
GRADIENTE DE LUZ INFLUENCIA A DIVERSIDADE FUNCIONAL DE PLANTAS? ................ 114
INFLUÊNCIA DO TAMANHO DA INFLORESCÊNCIA NA RIQUEZA E VISITA DE INSETOS EM
UMA LEGUMINOSA TROPICAL ........................................................................................................... 118
EXISTEM EFEITOS DA DISTÂNCIA DA BORDA DE BANCOS DE MACRÓFITAS AQUÁTICAS
NA DIVERSIDADE DE GRUPOS FUNCIONAIS DE INVERTEBRADOS?....................................... 124
VARIAÇÃO DE RIQUEZA E COMPOSIÇÃO DE ESPÉCIES DE MACRÓFITAS AO LONGO DE
UM GRADIENTE AMBIENTAL.............................................................................................................. 130
EFEITO DA TURBIDEZ DA ÁGUA NA PREDAÇÃO DO ZOOPLÂNCTON POR PEIXES ........... 135
EFEITO DA INUNDAÇÃO NA CO-OCORRÊNCIA DE ESPÉCIES EM COMUNIDADES
VEGETAIS DA PLANÍCIE DE INUNDAÇÃO DO RIO PARANÁ....................................................... 140
HETEROGENEIDADE AMBIENTAL COMO PREDITOR DA DIVERSIDADE BETA EM LAGOAS
DE PLANÍCIE DE INUNDAÇÃO ............................................................................................................. 144
DIVERSIFICAÇÃO ECOMORFOLÓGICA EM ASSEMBLÉIAS DE PEIXES DO ALTO RIO
PARANÁ ...................................................................................................................................................... 149

7
FATORES LOCAIS E REGIONAIS INFLUENCIANDO A ESTRUTURA DAS COMUNIDADES VEGETAIS NA
PLANÍCIE DE INUNDAÇÃO DO RIO PARANÁ
Vinicius Marcilio da Silva
Projeto Individual
Resumo
O modo como comunidades ecológicas são estruturadas é uma temática central em ecologia.
Filtros bióticos, abióticos e a história evolutiva das espécies permite compreender como ocorre essa
estruturação. Esse trabalho avaliou o efeito de variações ambientais locais e a diversidade
filogenética e funcional em duas escalas distintas, local e regional. O local de estudo foi a planície
de inundação do alto rio Paraná. Para avaliar tal estrutura, utilizaram-se listas de 9 trabalhos
fitossociológicos na região. A estrutura mais fina foi avaliada com coletas em campo em três pontos
distintos. Os resultados indicaram que a variação ambiental em escala local, influencia a estrutura
mais específica dessas comunidades, e não se relaciona, de forma direta, com os padrões gerais
dessa estrutura.
Palavras-chave: regras de montagem, filogenia, ecologia funcional
Introdução
A estruturação das comunidades no espaço e no tempo é uma temática central em ecologia
de comunidades (Pillar & Duarte 2010). Essa estruturação, em termos gerais, é relacionada aos
filtros ambientais e interações bióticas (Cornwell & Ackerly 2009). Além disso, o efeito cumulativo
dessas relações ao longo do tempo também influencia a forma como ocorre essa estruturação
(Cavender-Bares et al. 2009). Para se entender melhor a estruturação de comunidades ecológicas, é
importante identificar os maiores gradientes filogenéticos entre variações ambientais e/ou
geográficas (Duarte et al. 2012).
A relação filogenética e funcional entre as espécies, ao se utilizar listas de
presença/ausência de espécies e valores médios de estados de atributos por espécie, refletem
padrões mais gerais da estrutura das comunidades. Já analises que consideram a abundância dos
indivíduos e os estados dos atributos de cada um (i.e. variação intra-especifica), refletem padrões
mais finos da estrutura das comunidades (Violle et al. 2012).
A diversidade funcional, combinada com as diversidades filogenética e taxonômica,
potencialmente revelam mais amplamente os processos que estruturam comunidades biológicas.
Juntas, essas medidas podem ajudar a incrementar o conhecimento de como a biodiversidade

8
interage com processos ecossistêmicos e limitações ambientais (Mouchet et al. 2010) num contexto
histórico (Cavander-Bares et al. 2009). Além disso, os padrões de correlação entre essas métricas de
biodiversidade podem ajudar a entender processos ecológicos mais complexos, tais como aqueles
que determinam a distribuição de toda uma biota numa escala ampla (Díaz & Cabido 2001,
Carvender-Bares et al. 2009, Safi et al. 2011).
Neste trabalho, foram analisadas as diversidades funcional e filogenética, em diferentes
escalas, para verificar a influência de variações locais sobre a estruturação das comunidades.
Material e métodos
Área de estudos:
A área de estudo compreendeu remanescentes de florestas ripárias localizadas na planície de
inundação do alto Rio Paraná, no estado do Mato Grosso do Sul e do Paraná (Fig.1).
Figura 1 – Localização do local de estudo, das áreas dos levantamentos compilados (1 = 3 listas, 2 = 2 listas,3 = 1 lista, 4 = 3 listas) e dos pontos coletados neste trabalho (3 transecções em cada ponto coletado), naplanície de inundação do alto rio Paraná (Modificado de Slusarski & Souza 2012)
Compilação de dados:
Foram compiladas informações sobre a ocorrência de espécies lenhosas em 9 listas de
espécies, apresentadas por Slusarski & Souza(2012), de trabalhos de florística e fitossociologia que
abrangem margem esquerda, direita e ilha, na planície de inundação do alto Rio Paraná, próximo ao
município paranaense de Porto Rico (Fig.1). Cada trabalho compilado foi considerado um ponto de
amostragem e, além do número de espécies, foram mensurados índices de diversidade filogenética e
funcional para cada um.

9
A diversidade filogenética foi gerada a partir da relação filogenética entre as espécies,
proposta pela APG III (2009), considerando como medida de semelhança entre pares de espécies o
número de nós da árvore filogenética entre elas. Apesar de não expressar as distâncias evolutivas
reais entre as espécies, o método em questão fornece uma medida útil para avaliar as relações
filogenéticas em plantas (Webb 2000, Webb et al. 2002, Pillar & Duarte 2010, Swenson 2011).
A diversidade funcional foi baseada em atributos funcionais compilados da literatura. A
partir da distância funcional entre as espécies, baseada no estado de seus atributos, foi utilizada a
entropia quadrática de Rao Q (Rao 1982) como medida de diversidade funcional.
Coleta em campo:
A coleta de atributos funcionais em campo foi feita nas duas margens e na ilha, nos
mesmos locais dos trabalhos compilados. Em cada local (margem esquerda, direita e ilha) foram
feitos 3 transecções de 21m paralelas ao Rio Paraná, distando 15m, 30m e 45m da margem do rio.
Em cada transecção foram alocadas quatro parcelas quadradas de 2x2m ao longo doa transecção,
equidistando 5m cada uma da mais próxima. Dentro de cada parcela foram amostrados todos os
indivíduos lenhosos maiores de 50 cm de altura.
Estrutura filogenética da comunidade vegetal:
A estrutura filogenética das espécies vegetais compiladas para a região foi analisada
baseando-se no método descrito por Duarte (2011), para a Análise dos Componentes Filogenéticos
Principais (PCPS). Este método permite verificar os principais gradientes de variação da estrutura
filogenética entre as comunidades. A associação entre os métodos de amostragem e a estrutura
filogenética dos pontos, foi verificada pelo teste de Mantel. Para isso, foi construída uma matriz de
distância entre os pontos baseado nos métodos de amostragem. Essa análise demonstrou a ausência
de vieses causados pelas diferenças de métodos de amostragem na estrutura filogenética dos pontos
(rM=0,205, P=0,245).
A relação entre a distância geográfica e a estrutura filogenética das comunidades foi
verificada analisando a relação entre as coordenadas geográficas (decimais) e cada PCPS gerado. A
disposição geográfica dos pontos manteve alta correlação entre latitude e longitude (r²=0,97,
P<0,005). Por este motivo, cada PCPS foi relacionado com cada eixo geográfico, controlado pelo
outro eixo geográfico, através do teste de Mantel parcial. Isto permitiu se verificar a relação entre
cada PCPS e cada eixo geográfico de maneira exclusiva.
Resultados

10
A diversidade filogenética e a diversidade funcional baseada em dados da literatura não
diferiram entre as situações (P>0,005).
A análise dos componentes filogenéticos principais gerou 7 eixos da PCPS. Os dois
primeiros eixos PCPS continham, respectivamente 76% e 13% da variação total na relação
filogenética da comunidade; todos os outros PCPS representaram <3% cada.
O PCPS1 não teve relação com as coordenadas geográficas (rM(PCPS1,Lat-Long)=0,097,
P=0,236; rM(PCPS1, Long-Lat)= -0,075, P=0,645). O PCPS2 teve relação com latitude (rM(PCPS2,Lat-
Long)=0,48, P=0,021), mas não com a longitude (rM(PCPS1,Lat-Long)=-0,019, P=0,911).
A diversidade funcional, baseada nos atributos levantados em campo, foi maior na margem
esquerda do que na ilha, enquanto a margem direita não diferiu nem da ilha nem da margem
esquerda (Fig. 2).
Local
me il md
Rao
0.12
0.14
0.16
0.18
0.20
0.22
0.24
0.26
0.28
0.30
0.32
Figura 2 – Variação da diversidade funcional (Rao) entre os locais ( me = Margem esquerda, Il = Ilha e md =
Margem direita. A ANOVA indicou diferenças apenas entre me e Il.
Discussão

11
O fato da maior riqueza de espécies na margem esquerda não refletir em maiores
diversidades filogenética e funcional, indica que a adição de cada espécie na comunidade pode não
gerar diretamente a adição de maiores variações filogenéticas e funcionais na mesma. A
heterogeneidade ambiental na margem esquerda, por diferentes influências dos eventos de
inundação do Rio Paraná na região estudada (Slusarski & Souza 2012), não é suficiente para alterar
a estrutura filogenética dessa margem em relação às comunidades da ilha e da margem direita.
Deste mesmo modo, essa heterogeneidade ambiental também não reflete em um aumento na
variação dos estados dos atributos (levantados na literatura) para a comunidade vegetal da margem
esquerda em relação a esta mesma variação nas comunidades da ilha e da margem direita.
Possivelmente, as variações ambientais locais não refletem o padrão geral da estrutura dessas
comunidades.
A análise dos componentes filogenéticos principais corrobora a ideia de que a variação
encontrada na composição de espécies entre as margens e a ilha não reflete nos padrões mais gerais
da estruturação dessas comunidades. O PCPS1 descreve um gradiente relacionado com os nós mais
profundos da árvore filogenética enquanto o PCPS2 descreve um gradiente filogenético mais fino
(e.g. gêneros, espécies) (Duarte et al. 2012). A ausência de relação entre as variações geográficas
locais e o PCPS1 corrobora a ideia de que os padrões mais gerais estão estruturados por fatores
ambientais e bióticos em escala regional. A relação entre o PCPS2 e a latitude, sem influência da
longitude, indica que a variação filogenética mais fina está relacionada com a localização das
comunidades em relação ao rio Paraná (margens direita, esquerda ou ilha), uma vez que a principal
diferença entre as áreas e este vetor geográfico é justamente sua localização relativa ao rio.
A variação ambiental (diferentes influências do alagamento entre as áreas) se torna
importante para a variação da diversidade funcional mais fina. O alagamento pode ser o fator que
homogeniíza os estados dos atributos nos indivíduos que estão na ilha, agindo como um forte filtro
ambiental a essa escala.
Os padrões das diversidades filogenéticas e funcionais mais gerais para essas comunidades
estão estruturadas regionalmente, não variando localmente. Já os padrões mais finos dessas
diversidades se relacionam com a posição relativa ao rio Paraná, tendo este uma grande influência
na estrutura fina dessas comunidades.
Referências
APG III. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and
families of flowering plants: APG III. - Botanical Journal of Linnean Society 161: 105-121.
Cavender-Bares J, Kozak KH, Fine PVA & Kembe, SW. 2009. The merging of community ecology

12
and phylogenetic biology. Ecology Letters 12: 693–715.
Cornwell, WK & Ackerly, DD. 2009. Community assembly and shifts in plant trait distributions
across an environmental gradient in coastal California. Ecol. Monogr., 79, 109–126.
Díaz, S & Cabido, M 2001 Vive la diference: plant functional diversity matters to ecosystem
processes. TRENDS in Ecology & Evolution 16: 646-655.
Duarte, LS 2011. Phylogenetic habitat filtering influences forest nucleation in grasslands. Oikos
120: 208–215.
Duarte, LS, Pietro, PV, and Pillar, VD 2012. Assessing spatial and environmental drivers of
phylogenetic structure in Brazilian Araucaria forests. Ecography 35: 001–009.
Mouchet, MA, Villeger, S, Mason, NWH, Mouillot, D2010. Functional diversity measures: an
overview of their redundancy and their ability to discriminate community assembly rules.
Funct. Ecol. 24 (4), 867–876.
Pillar, VD & Duarte, LDS. 2010. A framework for metacommunity analysis of phylogenetic
structure. - Ecology Letters 13: 587-596.
Rao, CR. 1982. Diversity and dissimilarity coefficients: a unified approach. Theoretical Population
Biology 21: 24-43.
Safi, K, Cianciaruso, MV, Loyola, RD, Brito, D, Armour-Marshall, K & Diniz-Filho, JAF. 2012.
Understanding global patterns of mammalian functional and phylogenetic diversity. Phil.
Trans. R. Soc. B 366, 2536–2544.
Slusarski, S R & Souza, M C 2012. Analysis of floristic similarity between forest remnants from the
upper Paraná river floodplain, Brazil. Acta Scientiarum 34: 343-352.
Swenson, NG. 2011. Phylogenetic beta diversity metrics, trait evolution and inferring the functional
beta diversity of communities. PLoS One 6: e21264.
Violle, C, Enquist, B J, McGill, BJ, Jianq, L, Albert, C H, Hulshof, C, Junq, V, & Messier, J 2012.
The return of the variance: intraespecific variability in community ecology. Trends Ecol Evol.
27:244-52.
Webb, CO, Ackerly, DD, McPeek, MA & Donoghue, MJ. 2002. Phylogenies and community
ecology. Annual Review of Ecology and Systematics 33: 475–505.
Webb, CO. 2000. Exploring the phylogenetic structure of ecological communities: an example of
rain forests trees. American Naturalist 156: 145-155.

13
INFLUÊNCIA DA DIVERSIDADE MORFOLÓGICA NA SEDIMENTAÇÃO DO FITOPLÂNCTON: QUEM
VENCE ESSA CORRIDA?
Resumo
As algas fitoplanctônicas apresentam uma série de formas complexas que permitem a manutenção
da sua posição na zona fótica. O objetivo desse trabalho foi verificar a taxa de sedimentação de
diferentes formas do fitoplâncton, a fim de testar a hipótese de que algas com maior relação
superfície/volume apresentam maior resistência à sedimentação. Nossos resultados indicam que a
forma das algas influencia na velocidade de sedimentação e corroboram a hipótese inicial do
trabalho.
Palavras-chave: Morfologia, relação superfície/volume, microalga.
Introdução
O fitoplâncton é composto por microalgas autotróficas que não apresentam locomoção
suficiente para vencer os movimentos da água (Reynolds, 1984). Por conta disso, ao longo do
tempo evolutivo, surgiram alguns mecanismos que aumentam a permanência das células na zona
iluminada da coluna de água, já que, em geral, elas não são capazes de sobreviver no escuro. A
principal adaptação morfológica relacionada a redução da sedimentação foi a formação de colônias,
que aumenta a relação superfície/volume do indivíduo. Essa característica otimiza a fotossíntese e
causa um efeito de “sombreamento” nas outras espécies (Fernandes et al., 2005), evitando a
também a competição inter-específica.
O objetivo desse trabalho foi verificar a taxa de sedimentação de diferentes modelos
baseados em algumas formas comuns do fitoplâncton, com a hipótese de que formas com maior
relação superfície/volume são mais resistentes a sedimentação na coluna de água.
Material e Métodos
Os modelos das formas do fitoplâncton (Fig. 1) foram confeccionados a partir de uma esfera
de massa de modelar com volume padrão (5 g para as algas menores e 10 g para as coloniais
maiores). Setas, feitas com linha de pesca, foram incluídas nos modelos para mimetizar algas com
esses ornamentos. A densidade das células do fitoplâncton é ligeiramente superior a da água
(Reynolds, 1984), já a densidade da massa de modelar utilizada no experimento é superior a da
água, por esse motivo, escolhemos detergente comercial como um líquido mais viscoso para
determinar a taxa de sedimentação dos modelos. A taxa de sedimentação foi calculada a partir da
velocidade de deslocamento dos modelos em um aquário contendo 30 litros de detergente. A
relação superfície/volume foi obtida dividindo-se a área de cada um dos modelos, calculada no
programa ImageJ, pelo volume padrão. Uma correlação foi realizada entre a relação

14
superfície/volume e o tempo de sedimentação de cada modelo. Além disso, uma ANCOVA foi
usada para verificar como a relação superfície/volume e o tempo variam, controlando a variável
presença de setas nos modelos. Para verificar se as taxas de sedimentação dos modelos foram
significativamente diferentes do controle foram realizados teste-t, exceto para Cylindrospermopsis e
Desmodesmus que possuem dois morfotipos, para esses modelos foi utilizado ANOVA seguido de
Tukey. As análises estatísticas foram realizadas em ambiente R.
Figura 1. Modelos das formas dos gêneros do fitoplâncton confeccionados com massa de modelar.
ResultadosAs formas com setas apareceram como outliers e foram excluídas das análises para evitar
a influência dessa característica (Fig. 2A). A relação superfície/volume influenciou
significativamente a taxa de sedimentação dos modelos (R²=0,85; F3,3=12,69; P=0,03; Fig. 2B).
Figura 2. Correlação entre o tempo de sedimentação e a relação superfície/volume dos modelos testados.
As diferenças entre as taxas de sedimentação das formas do fitoplâncton e o controle
foram significativas, exceto para Desmodesmus sem setas (Tab.1; Fig. 3). O teste post-hoc
demonstrou que o tempo de sedimentação de ambas as formas de Cylindrospermopsis (reta e
A B

15
espiralada) diferiram do controle (p<0,001). Apenas o tempo de sedimentação de Desmodesmus
com setas diferiu do controle (p<0,001).
Tabela1. Comparação entre o tempo de sedimentação dos modelos e o controle.
Modelo Estatística p-valor
Cylindrospermopsis F2,6=117,6 <0,001
Desmodesmus F2,6=39,9 <0,001
Fragilaria t5=4,23 0,008
Golenkinia t5=3,32 0,02
Microcystis t5=5,92 0,001
Pediastrum t5=5,47 0,002
Figura 3. Tempo de sedimentação dos morfotipos de Desmodesmus (esquerda) e de Golenkinia (direita) comparadoscom o controle. Letras diferentes indicam p<0,05 (ANOVA).
DiscussãoComo demonstrado em nossos experimentos, a formação de colônia aumenta a relação
superfície/volume do indivíduo e, consequentemente, leva a uma diminuição na taxa de
sedimentação na coluna de água. Por outro lado, em cultivo, algumas algas coloniais não formam
essas agregações. Esse fato é, muitas vezes, atribuído a uma “economia de energia” com a
manutenção da posição na coluna de água, já que os frascos de cultura estão sob constante
agitamento. Alguns exemplos dessas algas são Microcystis (Sangolkar et al., 2009) e Desmodesmus
(Johnson et al., 2007).
Para o gênero Desmodesmus a presença de setas foi o que determinou a diminuição
significativa da taxa de sedimentação. Essa característica é, normalmente, atribuída a resistência à
herbivoria (Verschoor et al., 2004). Todavia, nosso experimento demonstrou que esse pode ser um
importante fator na manutenção da posição do indivíduo na coluna de água. O gênero Golenkinia,
com maior número de setas, foi ainda mais resistente à sedimentação.
Cylindrospermopsis raciborskii é uma alga colonial filamentosa que possui dois tipos
morfológicos principais: reto e espiralado. De acordo com nosso experimento, o morfotipo reto é
a a
b

16
vantajoso em condições de ausência de turbulência, por possuir taxa de sedimentação menor que o
espiralado. De fato, em períodos secos, quando os reservatórios tendem a estratificação, há um
predomínio do morfotipo reto. Já em períodos chuvosos, que levam a circulação da coluna de água,
ocorre uma dominância do morfotipo espiralado (Bittencourt-Oliveira et al., 2011).
O gênero Fragilaria é uma das poucas diatomáceas penadas com hábito planctônico (Round
et al., 1990) e essa característica pode ser atribuída a formação de colônia. Esse hábito diminui a
taxa de sedimentação desses indivíduos que possuem carapaça de sílica e levariam desvantagem
frente às algas de outros grupos que são menos pesadas.
Esse trabalho evidenciou a importância da evolução da forma das algas planctônicas, como
o aumento da relação superfície/volume e o aparecimento de ornamentações e anexos que, como
demonstrado, são eficientes mecanismos para evitar a sedimentação.
Referências Bibliográficas
Bittencourt-Oliveira MC, Moura AN, Hereman TC, Dantas EW 2011 Increase in Straight and
Coiled Cylindrospermopsis raciborskii (Cyanobacteria) Populations under Conditions of
Thermal De-Stratification in a Shallow Tropical Reservoir. Journal of Water Resource and
Protection, v. 3, p. 245-252.
Fernandes LF, Wosiak AC, Domingues L, Pacheco CV, Lagos PED 2005 Comunidades
Fitoplanctônicas em Ambientes Lênticos. Pages 315-366 in: Andreoli CV, Carneiro C
Gestão Integrada de Mananciais de Abastecimento Eutrofizados. Sanepar Finep, Curitiba.
Johnson JL, Fawley MW, Fawley KP 2007 The diversity of Scenedesmus and Desmodesmus
(Chlorophyceae) in Itasca State Park, Minnesota, USA. Phycologia, v. 46, n. 2, p. 214-229.
Reynolds CS 1984 The Ecology of Freshwater Phytoplankton. Cambridge University Press,
Cambridge.
Round FE, Crawford RM, Mann DG 1990 The Diatoms: Biology and Morphology of the Genera.
Cambridge University Press, Cambridge, 747 p.
Sangolkar LN, Maske SS, Muthal PL, Kashyap SM, Chakrabarti T 2009 Isolation and
characterization of microcystin producing Microcystis from a Central Indian water bloom.
Harmful Algae, v. 8, p. 674-684.
Verschoor AM, van der Stap I, Helmsing NR, Lürling M, van Donk E 2004 Inducible colony
formation within the Scenedesmaceae: adaptive responses to infochemicals from two
different herbivore taxa. Journal of Phycology, v. 40, p. 808-814.

17
RIQUEZA DE FORMIGAS EM AMBIENTES COM DIFERENTES GRAUS DE PERTURBAÇÃO
Diego Corrêa Alves, Fernanda Ceschin, Letícia Cucolo Karling, Lilian Vasconcelos, Luciana Zago,
Monica Vasques Mangas Pereira
“Projeto livre”
Resumo: O objetivo deste estudo foi verificar o efeito do impacto antrópico na riqueza de
comunidades de formigas e comparar diferentes tipos de iscas utilizadas como método de captura.
Apesar de haver uma tendência para a diminuição da riqueza de espécies em locais mais
impactados, não foi significativa a diferença do número de morfotipos de formigas encontrados na
área da mata, pastagem e zona urbana. Quanto às iscas utilizadas, não foi possível testar as
diferenças devido a limitações nas amostragens.
Palavras-chave: Formigas; isca; riqueza; perturbações ambientais.
Introdução
Os artrópodes compreendem mais de um milhão de espécies e ocupam uma grande
variedade de nichos nos ecossistemas (Aquino et al, 2006). Esta fauna apresenta uma tendência em
acompanhar a riqueza vegetal, pois muitos animais dependem desta riqueza para sua sobrevivência.
Modificações de paisagens naturais podem influenciar a diversidade de espécies de artrópodes
associada a essas áreas, já que a baixa diversidade vegetal acarreta diferenças na disponibilidade de
recursos em tais ambientes (Battirola et al., 2007).
A estrutura da comunidade de formigas é influenciada por perturbações de origem humana,
tal como alterações nos habitats como pastagens, urbanização e outras mudanças na forma do uso
dos ecossistemas terrestres. Segundo Delabie et al. (2006), as comunidades de formigas possuem
um grande potencial para serem utilizadas como bioindicadores de impactos ambientais, já que a
riqueza das comunidades é relacionada negativamente com o grau de antropização.
Alguns autores enfatizam a importância da utilização de mais de uma técnica de amostragem
para conseguir abranger diferentes espécies em um mesmo estudo (Silva et al. 2011). Iscas em geral
são utilizadas para aumentar a eficiência das armadilhas (Aquino et al. 2006), no entanto, mesmo
estudos que utilizam de técnicas de amostragem diferentes não costumam testar diferentes iscas
(p.ex.Lopes & Vasconcelos 2008).
Sob esse contexto, o presente trabalho visa testar as seguintes hipóteses: i) localidades com
diferentes níveis de impactos influenciam a riqueza de morfotipos de formigas; ii) diferentes iscas
atraem diferentes composições de formigas. Visto que a fauna de invertebrados apresenta uma

18
tendência em acompanhar a diversidade vegetal, espera-se que a riqueza de formigas diminua
conforme aumenta o nível de impacto do local.
Material e métodos
Coleta de dados
As amostragens dos artrópodes foram realizadas através de armadilhas com iscas em três
diferentes localidades: área urbana (dentro da cidade de Porto Rico), área rural (campo de pastagem,
ao lado da base de estudos do Nupélia) e mata (fragmento de mata ciliar do rio Paraná). Em cada
localidade foram determinados quatro pontos aleatórios de coleta. Para todas as localidades foi
utilizado doce de banana pastoso como isca. No fragmento de mata ciliar, foi realizada amostragem
adicional com sardinha em lata. As iscas foram colocadas no solo, em cima de uma folha de papel,
onde ficaram expostas por 15 minutos. Em seguida, as formigas aderidas as iscas foram recolhidas
com o papel e armazenadas em sacos plásticos para posterior identificação dos morfotipos em
laboratório.
Análise dos dados
A primeira hipótese foi testada através de uma ANOVA unifatorial, considerando a
distribuição de Poisson para a estrutura dos resíduos, tendo a riqueza de morfotipos como variável
resposta e as diferentes localidades como o fator. Para testar a segunda hipótese a intenção era
realizar uma PERMANOVA; no entanto, como houve captura de formigas apenas em um dos
quatro pontos amostrados com a isca de sardinha, não foi possível calcular a média das
dissimilaridades para esta isca. Assim, tentou-se identificar o padrão visualmente através de um
gráfico exploratório.
Resultados
Nas amostragens com isca de banana nas três localidades, foram encontradas nove
morfotipos de formigas. Os resultados da ANOVA demonstraram que a riqueza de morfotipos de
formigas nas três localidades não foi significativamente diferente, embora seja possível observar
que a mediana da riqueza na mata foi superior às demais (Fig. 1).

19
Fig. 1. Mediana e quartis 1 e 3 da riqueza de morfotipos de formigas por localidade.
Foi possível observar que houve dominância de um morfotipo de formigas na amostragem
com isca de sardinha, enquanto que com a isca de banana foram capturados mais morfotipos, porém
menos abundantes. Além disso, o único morfotipo capturado com a isca de sardinha não foi
amostrado com isca de banana.
mata pasto cidade
Localidade
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Riq
ueza
Mediana 25%-75%
p=0,278
19
Fig. 1. Mediana e quartis 1 e 3 da riqueza de morfotipos de formigas por localidade.
Foi possível observar que houve dominância de um morfotipo de formigas na amostragem
com isca de sardinha, enquanto que com a isca de banana foram capturados mais morfotipos, porém
menos abundantes. Além disso, o único morfotipo capturado com a isca de sardinha não foi
amostrado com isca de banana.
mata pasto cidade
Localidade
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Riq
ueza
Mediana 25%-75%
p=0,278
19
Fig. 1. Mediana e quartis 1 e 3 da riqueza de morfotipos de formigas por localidade.
Foi possível observar que houve dominância de um morfotipo de formigas na amostragem
com isca de sardinha, enquanto que com a isca de banana foram capturados mais morfotipos, porém
menos abundantes. Além disso, o único morfotipo capturado com a isca de sardinha não foi
amostrado com isca de banana.
mata pasto cidade
Localidade
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Riq
ueza
Mediana 25%-75%
p=0,278

20
Fig.2. Número de indivíduos (bolas maiores = maior número de indivíduos) capturados de cada
morfotipo de formigas por ponto amostral de cada isca utilizada.
Discussão
Embora os resultados não tenham confirmado as diferenças entre os três fragmentos
amostrados no estudo, sabe-se que a fauna do solo é afetada por fatores ambientais e bióticos que
podem gerar alterações na composição das comunidades (Copatti & Daudt 2009). Regiões mais
impactadas tendem a suportar riquezas menores de espécies (p. ex. Lopes et al. 2010 ), e esta
tendência, apesar de não significativa, pode ser observada no estudo. No entanto, o espaço e o
tempo amostral possivelmente foram insuficientes para representar a composição das comunidades.
Para a análise de comparação de iscas, a hipótese não foi confirmada. Entretanto, foi
possível observar que as duas iscas atraíram morfotipos distintos, ressaltando a importância já
discutida por outros autores da utilização de métodos diferentes (Silva et al. 2011, Lopes &
Vasconcelos 2008). Frente a estes resultados, fica evidente que a utilização de diferentes métodos
de captura com a inclusão da busca ativa, maior esforço amostral e diferentes iscas podem prover
melhores representações da mirmecofauna de diferentes habitats.
Referências
Battirola LD, Adis J, Marques MI, Silva FHO 2007 Composição da comunidade de artrópodes
associada à copa de Attalea phalerata Mart. (Arecaceae), durante o período de cheia no
Pantanal de Poconé, Mato Grosso, Brasil. Neotropical Entomology 36:640-651.
Copatti CE, Daudt CR 2009 Diversidade de artrópodes na serapilheira em fragmentos de mata
nativa e Pinus elliottii (Engelm. Var elliottii). Ciência e Natura 31 (1): 95 – 113.
De Aquino AM, et al. 2006 Recomendações para coleta de artrópodes terrestres por armadilhas de
queda (" Pitfall-Traps"). Embrapa Agrobiologia.
Delabie JHC, Paim VRLM, Nascimento IC, Campiolo S, Mariano CSF 2006 As formigas como
indicadores biológicos do impacto humano em manguezais da costa sudeste da Bahia.
Neotropical Entomology 35(5):602-615.
Lopes CT, Vasconcelos HL 2008 Evaluation of Three Methods for Sampling Ground-Dwelling
Ants in the Brazilian Cerrado. Neotropical Entomology 37(4): 399-405.

21
Lopes DT, Lopes J, Nascimento IC, Delabie JH 2010 Diversidade de formigas epigéicas
(Hymenoptera, Formicidae) em três ambientes no Parque Estadual Mata dos Godoy,
Londrina, Paraná. Iheringia, Sér. Zool. 100(1): 84-90.

22
AÇÃO DE METABÓLITOS SECUNDÁRIOS DE PLANTAS NA PREDAÇÃO DE FORMIGAS SOLENOPSIS SP.
EM DIFERENTES ITENS ALIMENTARES
Érika Maria Neif, Francisco von Hartenthal, Guilherme Landgraf, Rafael A. Baggio, Sabrina
Calado
Resumo
Nesse estudo, testamos se metabólitos secundários de plantas podem reduzir o consumo de
diferentes itens alimentares de Solenopsis sp. Foram realizados experimentos de curta duração in
situ com extratos de Piper amalago L., Psidium guajava L. e Zanthoxyllum caribaeum sobre maça e
sardinha em três formigueiros. Nossos resultados sugerem que esses metabólitos secundários
possuem potencial para reduzir a herbivoria de Solenopsis sp. sobre esses itens alimentares,
contudo, as análises estatísticas não foram significativas.
Introdução
As plantas produzem uma ampla e diversa ordem de componentes orgânicos divididos em
metabólitos primários e secundários. Os metabólitos primários desempenham função essencial para
o crescimento e desenvolvimento, bem como para a manutenção da célula das plantas. Em
contrapartida, o metabolismo secundário é responsável pelas relações entre o individuo e o
ambiente onde se encontra, essencial para a sobrevivência e continuidade da espécie dentro do
sistema (Taiz & Zeiger, 2006).
As espécies Piper amalago L., Psidium guajava L. e Zanthoxyllum caribaeum Lam, por
exemplo, possuem uma ampla distribuição na região da planície de inundação do alto rio Paraná
(Souza et al, 1997), e são consideradas potencialmente capazes de sintetizar metabólitos
secundários.
Piper amalago tem sido usada na medicina popular como um antiinflamatório, analgésicos,
antipiréticos, terapia para problemas de estômago, vermífugo e ação repelente (Carrara, et al. 2013).
Plantas do gênero Zanthoxylum, têm demonstrado propriedades anti-helmínticas, causando a
redução da ovoposição de nematoides quando expostas a extratos provenientes deste grupo de
plantas (Peneluc, et al. 2009). Vários estudos fitoquímicos e farmacológicos tem sido realizados
com Psidium guajava, evidenciando a inibição de edemas, dores e problemas respiratórios em ratos
(Jaiarj, et al. 1999). É possível, ainda, que compostos secundários dessas plantas tenham efeito
negativo sobre a herbivoria, fenômeno comum entre espécies vegetais (Harborne 1997). Assim,

23
testamos a hipótese que metabólitos secundários de P. amalago, Z. caribaeum e P. guajava podem
reduzir a herbivoria de formigas sobre diferentes itens alimentares.
Material e métodos
O estudo foi realizado na base avançada de pesquisa do Núpelia, em Porto Rico, Paraná,
Brasil. Foram utilizados 2,0 g de folhas de 3 espécies de plantas com compostos secundários
conhecidos: Zanthoxylum caribaeum Lam., Psidium guajava L. e Piper amalago L., as quais foram
coletadas na região. As folhas de cada espécie foram maceradas em solução de etanol 50% em
função da extração dos metabólitos secundários.
Para testar a hipótese, foram utilizados formigueiros da formiga Solenopsis sp, Lava-pés.
Essa espécie foi escolhida por serem amplamente distribuídas no território brasileiro, por terem
hábito alimentar onívoro, sendo altamente atraídas por substâncias oleosas, e por recrutarem um
grande número de operárias ao encontrar alimento (Bueno& Campos-Farinha 1999). Dessa forma, é
possível testar a influência de compostos secundários sobre diferentes itens alimentares (i.e.
sardinha, maçã), com uma forte resposta da colônia.
Foram selecionados 3 formigueiros (i.e. réplicas), nos quais foram colocados 5 tratamentos
com maçã e 5 tratamentos com sardinha. Cada tratamento com o composto secundário de
determinada planta compunha o item alimentar e 1 ml do extrato da planta, totalizando 3
tratamentos, somados a um tratamento com 1 ml de álcool 50% e um controle sem manipulação. Os
experimentos nos formigueiros foram observados por 30 minutos, sendo a quantidade de formigas
forrageando sobre cada item estimado a cada 10 min.
As análises foram realizadas através do método Kruskall-Wallis, implementados no software
Statistica 7.0.
Resultados
Os testes de Kruskal-Wallis não foram significativos para os diferentes itens alimentares
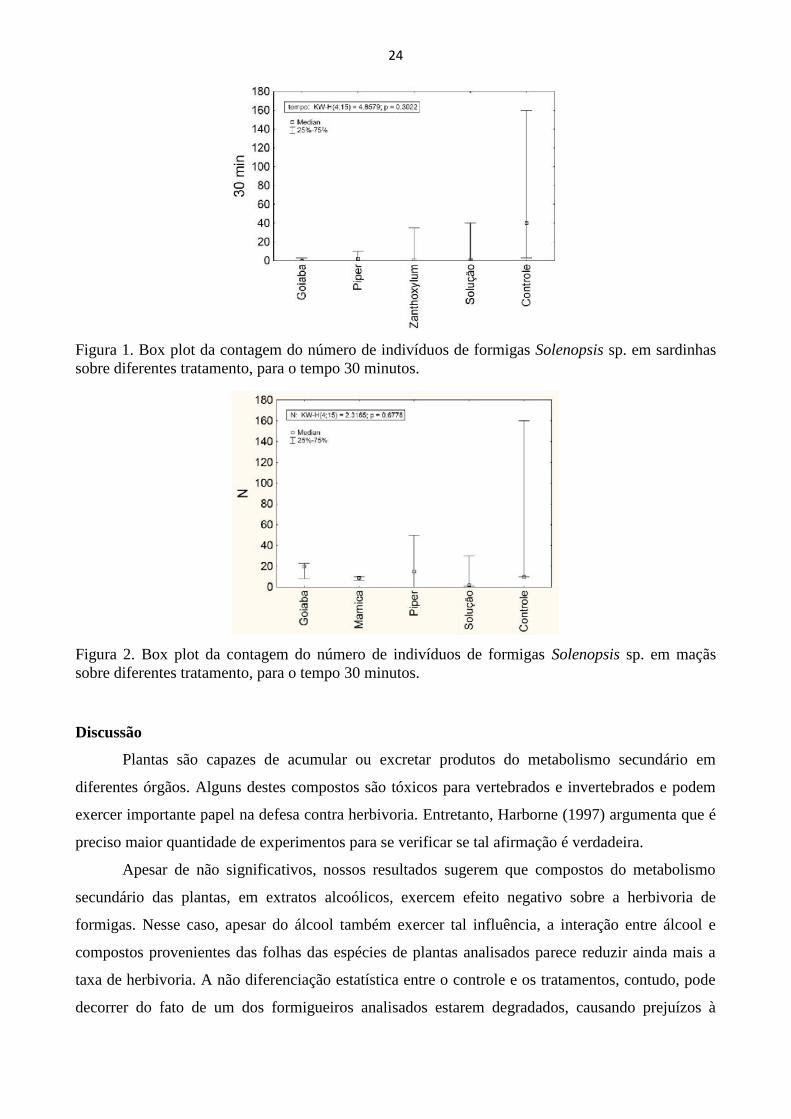
sardinha (H = 4,86; P > 0,05) (Fig 1) e maçã (H = 2,32; P = 0,68) (Fig. 2), portanto não suportaram
a hipótese de que as soluções com compostos das plantas poderiam inibir a herbivoria da formiga
sobre os itens alimentares. Os gráficos também indicam grande variância nos dados, sendo este
motivo, junto com o pequeno número de réplicas, a razão da não significância do teste.
24
Figura 1. Box plot da contagem do número de indivíduos de formigas Solenopsis sp. em sardinhassobre diferentes tratamento, para o tempo 30 minutos.
Figura 2. Box plot da contagem do número de indivíduos de formigas Solenopsis sp. em maçãssobre diferentes tratamento, para o tempo 30 minutos.
Discussão
Plantas são capazes de acumular ou excretar produtos do metabolismo secundário em
diferentes órgãos. Alguns destes compostos são tóxicos para vertebrados e invertebrados e podem
exercer importante papel na defesa contra herbivoria. Entretanto, Harborne (1997) argumenta que é
preciso maior quantidade de experimentos para se verificar se tal afirmação é verdadeira.
Apesar de não significativos, nossos resultados sugerem que compostos do metabolismo
secundário das plantas, em extratos alcoólicos, exercem efeito negativo sobre a herbivoria de
formigas. Nesse caso, apesar do álcool também exercer tal influência, a interação entre álcool e
compostos provenientes das folhas das espécies de plantas analisados parece reduzir ainda mais a
taxa de herbivoria. A não diferenciação estatística entre o controle e os tratamentos, contudo, pode
decorrer do fato de um dos formigueiros analisados estarem degradados, causando prejuízos à

25
colônia, com impacto no forrageamento de suas formigas. O aumento do número amostral,
portanto, é um requesito importante para reduzir a variação decorrente de um ponto amostral com
padrão incomum.
Agradecimentos
Agradecemos aos professores André Padial, Marcos Barbeitos e Roger Mormul pelas contribuições
para as análises estatísticas deste trabalho.
Referências
Carrara VS, Cunha-Júnior EF, Torres-Santos EC, Corrêa AG, Monteiro JL, Demarchi IG,Lonardoni MVC,4 Cortez DAG 2013 Antileishmanial activity of amides from Piper amalagoand synthetic analogs. Brazilian Journal of Pharmacognosy 23: 447-454.
Bueno OC, Campos-Farinha AEC 1999 As Formigas Domésticas, pages 135-180, in MariconiFAM, eds, Insetos e outros invasores de residáncias. Piracicaba: FEALQ.
Harborne JB 1997 Plant Secondary Metabolism, pages 132-155, in Crawley MJ, eds., PlantEcology, 2ª edição. Blackwell Science: Oxford.
Jaiarj P, Khoohaswan P, Wongkrajang P, Peungvicha, P, Suriyawong P, Saraya S M L,Ruangsomboon O 1999. Anticough and antimicrobial activities of Psidium guajava Linn. leafextract. Journal of Ethnopharmacology 67:203-212.
Peneluc T, Domingues LF, Almeida GN, Ayres, MCC Moreira ELT, Cruz ACF, BittencourtTCBST, Almeida MAO; Batatinha MJM 2009. Atividade anti-helmíntica do extrato aquosodas folhas de Zanthoxylum rhoifolium Lam. (Rutaceae). Rev. Bras. Parasitol. Vet., Jaboticabal18: 43-48.
Souza MC de, Cislinski J, Romagnolo MB 1997 Levantamento Florístico, pages 371-394, inVazzoler AEA, Agostinho AA, Hahn NS. A planície de inundação do alto rio Paraná:aspectos físicos, biológicos e socioeconômicos. EDUEM: Maringá.
Souza-Filho APS, Alves SM 2002 Alelopatia: princípios básicos e aspectos gerais. EmbrapaAmazônia Oriental: Belém. 260 p.
Taiz L, Zeiger E 2006 Plant physiology. 4. ed. Sinauer Associates Inc: Massachusetts.

26
CONTRIBUIÇÃO DE ESPÉCIES ZOOPLANCTÔNICAS EM RIACHOS TRIBUTÁRIOS SOB INFLUÊNCIA DE
UM RIO DE INUNDAÇÃO
EMANUEL Razzolini; FERNANDO G. de Oliveira; FLÁVIO M. Marteleto; NATÁLIA C. L. dos
Santos; SANDRA M. N. de Andrade
Projeto livre
Resumo Riachos não são habitats propícios para o desenvolvimento da comunidade zooplanctônica
por apresentarem baixa produtividade primária e dinâmica fluvial instável. Nessa perspectiva, os
objetivos foram analisar a riqueza zooplanctônica no gradiente de influência do rio Paraná sob os
riachos tributários e comparar a riqueza entre regiões com influência dos riachos (montante) e do
rio Paraná (jusante). Os resultados mostraram tendência significativa de redução de espécies
zooplanctônicas à medida que a inundação do rio Paraná diminui o seu efeito a montante dos
riachos tributários.
Palavras-chave: zooplâncton, espécies, variação espacial.
Introdução
Riachos, em particular, não são habitats propícios de desenvolvimento para muitas espécies
de zooplâncton. Allan (1998) descreve uma baixa diversidade de espécies e abundância do
zooplâncton nesses ambientes. Os principais motivos desse cenário são a baixa produtividade
primária e a alta instabilidade da dinâmica fluvial (Ejsmont-Karabin & Kruk 1998). Em riachos de
baixo fluxo, por exemplo, as condições químicas podem exercer um efeito mais sensível sobre a
comunidade zooplanctônica do que em contraste com os riachos de fluxo mais rápido (Czerniawski
2012).
Em macrossistemas fluviais, a dinâmica de planícies de inundação é regida pelos pulsos de
cheia (potamofase) e seca (limnofase) que podem transformar os componentes bióticos e abióticos
de todo o sistema (Neiff 1990). A conectividade durante a cheia promove um maior intercâmbio de
organismos aumentando a similaridade da distribuição de espécies entre os ambientes. Sob essa
perspectiva, os objetivos foram analisar a riqueza zooplanctônica no gradiente de influência do rio
Paraná sob os riachos tributários, e, comparar a riqueza zooplanctônica entre regiões com influência
dos riachos e do rio Paraná. As hipóteses foram: (i) a cheia do rio Paraná contribui com o aporte de
espécies de zooplâncton para riachos tributários; (ii) há diferença quanto as espécies de zooplâncton
entre as regiões a montante e a jusante dos riachos.
Material e Métodos
Área de estudo

27
A área de estudo compreende a região da bacia do Alto rio Paraná situada nas proximidades
do município de Porto Rico, Paraná, Brasil. A fisiografia do rio Paraná encontra-se, notavelmente,
alterada por uma sucessão de aproximadamente 150 barragens a montante da bacia, sendo estas os
principais agentes controladores da vazão do rio (Agostinho & Zalewski 1996). Neste estudo foram
inventariados três riachos da margem esquerda do rio Paraná (São Pedro, Caracu e Araldo) (Fig. 1).
Figura 1. Localização dos riachos (triângulo preto) onde os pontos de coleta foram amostrados. 1. Riacho São Pedro; 2.
RiachoCaracu; 3. Riacho do Araldo.
Para cada riacho, foram estabelecidos 4 pontos de coleta. O primeiro ponto (P0) consistiu
em uma amostragem de zooplâncton no rio Paraná a montante da foz dos riachos (ponto controle).
O segundo ponto (P1) foi amostrado no relento da foz dos riachos conectados com o rio Paraná. O
terceiro ponto (P2) foi estabelecido em uma região mais a montante dos riachos (característico de
água parada), região esta intermediária entre a foz do riacho e o limite da influência (inundação) do
rio Paraná sobre o riacho. O quarto ponto (P3) foi amostrado na região mais a montante, lugar em
que se observava uma pequena corrente de água proveniente do fluxo do riacho. As amostras do
zooplâncton foram coletadas à sub-superfície da região pelágica de cada ponto através de uma rede
de plâncton, com abertura de malha de 68μm. O material coletado foi acondicionado em frascos de
polietileno, sendo etiquetados e fixados em solução de formaldeído a 4%, tamponada com
carbonato de cálcio. Os organismos foram identificados por morfotipos atribuindo ser uma espécie
aqueles diferenciáveis morfologicamente entre um organismo e outro. Para a tabulação dos dados,
foram utilizados a presença e ausência de espécies por riachos e por pontos, e, através de uma
análise de variância (ANOVA), testou-se a significância entre as médias de riqueza dos pontos
amostrados aceitando um nível de significância P < 0,05. O programa estatístico utilizado foi o
Statistic 7.0.
Resultados
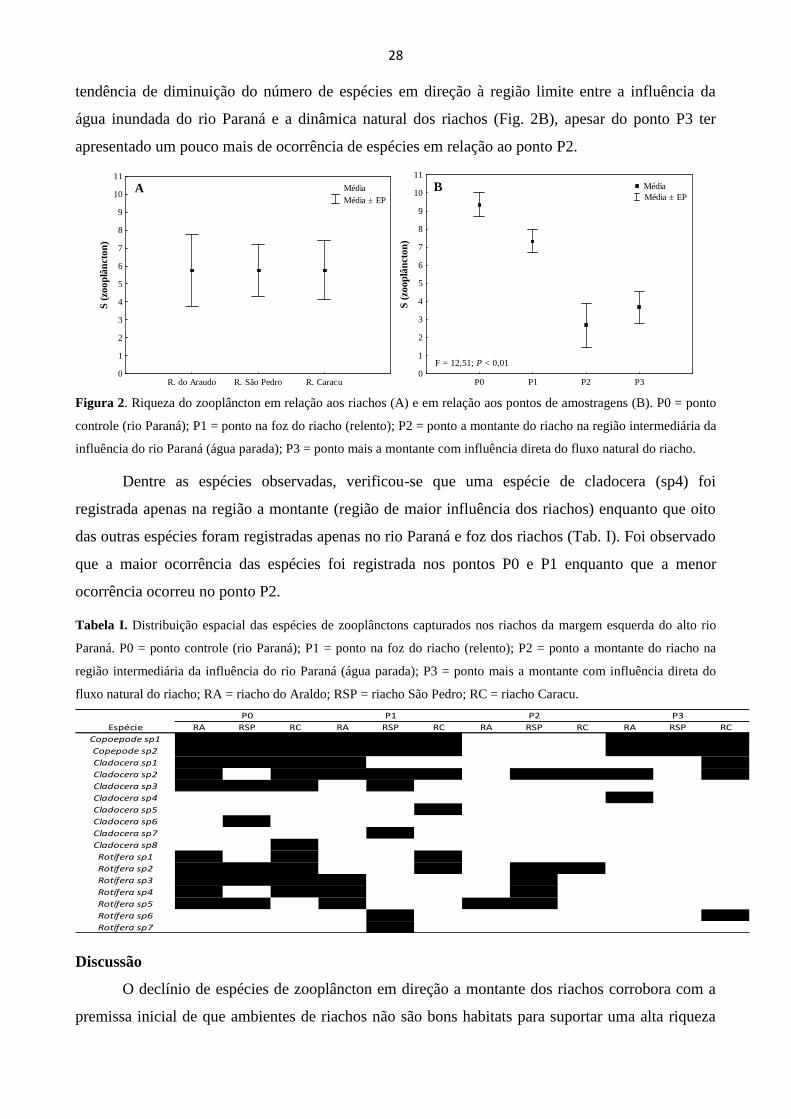
A riqueza de espécies de zooplâncton foi semelhante entre os três riachos (Fig. 2A). Já em
relação à distância espacial entre o rio Paraná e a montante dos riachos, observou-se que há uma
28
tendência de diminuição do número de espécies em direção à região limite entre a influência da
água inundada do rio Paraná e a dinâmica natural dos riachos (Fig. 2B), apesar do ponto P3 ter
apresentado um pouco mais de ocorrência de espécies em relação ao ponto P2.
Figura 2. Riqueza do zooplâncton em relação aos riachos (A) e em relação aos pontos de amostragens (B). P0 = ponto
controle (rio Paraná); P1 = ponto na foz do riacho (relento); P2 = ponto a montante do riacho na região intermediária da
influência do rio Paraná (água parada); P3 = ponto mais a montante com influência direta do fluxo natural do riacho.
Dentre as espécies observadas, verificou-se que uma espécie de cladocera (sp4) foi
registrada apenas na região a montante (região de maior influência dos riachos) enquanto que oito
das outras espécies foram registradas apenas no rio Paraná e foz dos riachos (Tab. I). Foi observado
que a maior ocorrência das espécies foi registrada nos pontos P0 e P1 enquanto que a menor
ocorrência ocorreu no ponto P2.
Tabela I. Distribuição espacial das espécies de zooplânctons capturados nos riachos da margem esquerda do alto rio
Paraná. P0 = ponto controle (rio Paraná); P1 = ponto na foz do riacho (relento); P2 = ponto a montante do riacho na
região intermediária da influência do rio Paraná (água parada); P3 = ponto mais a montante com influência direta do
fluxo natural do riacho; RA = riacho do Araldo; RSP = riacho São Pedro; RC = riacho Caracu.
Discussão
O declínio de espécies de zooplâncton em direção a montante dos riachos corrobora com a
premissa inicial de que ambientes de riachos não são bons habitats para suportar uma alta riqueza
R. do Araudo R. São Pedro R. Caracu0
1
2
3
4
5
6
7
8
9
10
11
S (z
oopl
ânct
on)
Média Média ± EP
A
P0 P1 P2 P30
1
2
3
4
5
6
7
8
9
10
11
S (z
oopl
ânct
on)
Média Média ± EP
F = 12,51; P < 0,01
F = 12,51; P < 0,01
B
Espécie RA RSP RC RA RSP RC RA RSP RC RA RSP RCCopoepode sp1 x x x x x x x x xCopepode sp2 x x x x x x x x xCladocera sp1 x x x x xCladocera sp2 x x x x x x x x xCladocera sp3 x x x xCladocera sp4 xCladocera sp5 xCladocera sp6 xCladocera sp7 xCladocera sp8 xRotífera sp1 x x xRotifera sp2 x x x x x xRotífera sp3 x x x x xRotífera sp4 x x x xRotífera sp5 x x x x xRotífera sp6 x xRotífera sp7 x
P0 P1 P2 P3

29
desses organismos. A hipótese confirmada de que o rio Paraná fornece aporte de espécies para os
ambientes de riacho pode ser explicada pela conectividade entre esses ambientes devido à cheia do
rio. Nessa área, uma grande inundação conecta os diferentes biótopos (lagoas abertas, fechadas,
canais, rios e riachos) (Souza-Filho 2009) configurando vias de dispersão para os organismos
aquáticos entre esses diferentes ambientes. De acordo com Thomaz et al. (2004) e Thomaz et al.
(2007), esses ambientes apresentam uma considerável variabilidade quanto às características
limnológicas, sendo esta associada ao grau de conexão (ou isolamento) com os rios e,
principalmente, ao regime hidrológico do rio Paraná. Nessa área, além das diferenças no que diz
respeito às suas características físico-químicas, outros mecanismos de interação como o influxo e
refluxo da água (Rocha 2002; Comunello et al. 2003) são observados nos ambientes da região,
podendo, então, esta estar sendo a principal característica que esta influenciando na estruturação da
comunidade zooplanctônica.
Agradecimentos
Os autores agradecem ao barqueiro Sebastião, funcionário do Nupélia/UEM, pelo serviço prestado;
e aos programas de pós-graduação da Universidade Federal do Paraná e Universidade Estadual de
Maringá pela disciplina ofertada para o treinamento e aprendizagem das pesquisas ecológicas em
campo.
Referencias bibliográficas
Agostinho AA & Zalewski M 1996 A planície alagável do alto rio Paraná: importância e
preservação. Maringá EDUEM 100p.
Allan JD 1998 Ekologia wod p_lyn˛acych [Stream Ecology. Structure and Function of Running
Waters]. Wydawnictwo Naukowe PWN S.A.,Warszawa, 450 pp. ISBN: 83-01-12535-7 Basu
B.K. & Pick F.R. 1996. Factors regulating phytoplankton and zooplankton biomass in
temperate rivers. Limnology Oceanography. 41 (7): 1572–1577.
Comunello EE, Souza-Filho E, Rocha PC, Nanni MR 2003 Dinâmica de inundação de áreas
sazonalmente alagáveis na planície aluvial do alto rio Paraná: estudo preliminar. Anais XI
SBSR, 2459-2466.
Czerniawski R 2012 Spatial pattern of potamozooplankton community of the slowly flowing
fishless stream in relation to abiotic and biotic factors. Polish journal of ecology 60 (2) 323–
338.
Ejsmont-Karabin J, Kruk M 1998 Effects of contrasting land use on free-swimming rotifer
communities of streams in Masurian Lake District, Poland. Hydrobiologia 241–249.
Neiff JJ 1990 Ideas para la interpretación ecológica del Paraná. Interciencia v 15 n 16 p 424-441.

30
Rocha PC 2002 Dinâmica dos canais no sistema rio-planície fluvial do alto rio Paraná, nas
proximidades de Porto Rico-PR. Unpublished Ph.D. Dissertation. Universidade Estadual de
Maringá, Maringá, 171p.
Souza-Filho EE 2009 Evaluation of the Upper Paraná River discharge controlled by reservoirs.
Brazilian Journal of Biology 69: 707-716.
Thomaz SM, Pagioro TA, Bini LM, Roberto MC & Rocha RRA 2004 Limnological
Characterization of the aquatic environments and the influence of the hydrometric levels.
Pages 75-102 in SM Thomaz, AA Agostinho, NS Hahn (eds.). The upper Paraná River and its
floodplain: physical aspects, ecology and conservation. Leiden, Backhuys Publishers.
Thomaz SM, Bini LM & Bozelli RL 2007 Floods increase similarity among aquatic habitats in
river-floodplain systems. Hydrobiologia 579: 1-13.

31
A DENSIDADE DE INVERTEBRADOS ASSOCIADOS À SERAPILHEIRA EM FRAGMENTOS FLORESTAIS
Hugo José Message, Priscilla Guedes Gambale, Renata Gabriela Noguchi, Simone Satsuki
Takeuchi & Taise Miranda Lopes
Projeto Livre
Resumo: A serapilheira exerce inúmeras funções na dinâmica dos ecossistemas, sendo regulada
pela quantidade de material depositado pela parte aérea das plantas e apresentando uma rica fauna
associada. A hipótese de que florestas mais desenvolvidas apresentam uma menor densidade de
serapilheira devido à maior decomposição foi testada, porém os fragmentos analisados não
diferiram entre si em grau de estruturação. O presente estudo comprovou a hipótese de que a
densidade da serapilheira apresenta relação positiva com a densidade da fauna associada.
Palavras-chave: fauna associada, estruturação vegetal, folhas, densidade.
Introdução
A serapilheira exerce inúmeras funções no equilíbrio e dinâmica dos ecossistemas,
compreendendo a camada mais superficial do solo em ambientes florestais, compostas por folhas,
ramos, órgãos reprodutivos e detritos (Costa et al., 2010). Ela contribui para a intercepção das gotas
de chuva, minimizando os efeitos erosivos no armazenamento de água, funcionando como um
isolante térmico e contribuindo para a manutenção do microclima do solo (Figueiró, 2005).
Também possui influência na germinação de sementes, no estabelecimento (Moraes et al., 1999) e
crescimento de plantas (Garay & Kindel, 2001).
A produção de serrapilheira em florestas tropicais pode ser determinada por vários fatores,
por exemplo, diferenças no microclima, composição das espécies, estágio sucessional de
desenvolvimento (Sundarapadian & Swamy, 1999). A camada orgânica da serapilheira é regulada
pela quantidade de material depositado pela parte aérea das plantas e pela taxa de decomposição. A
taxa de decomposição é afetada pela densidade e composição específica das folhas e gravetos
presentes na matéria orgânica, além da diversidade da fauna associada. Os organismos que habitam
esses ambientes podem influenciar todo o ecossistema local através da composição do solo em
microescala, a capacidade de drenagem do local e, em alguns locais, até a entrada de água em
corpos de água vizinhos por percolação.
O presente estudo testou as hipóteses de que (1) florestas mais desenvolvidas (estruturadas)
apresentam uma menor densidade de serapilheira devido à maior decomposição da mesma pelos

32
microrganismos e (2) a densidade de fauna associada apresenta relação positiva com a densidade da
serapilheira.
Material e métodos
A coleta de dados foi realizada em dois fragmentos florestais próximos à base de estudos da
Universidade Estadual de Maringá, um ao lado do córrego Caracu e outro distante cerca de 800
metros em linha reta do primeiro. Os fragmentos localizam-se na margem do rio Paraná e
encontram-se sujeitos a processos antropogênicos. Em cada fragmento realizaram-se duas
transecções de 20 metros de comprimento, cerca de 15 metros distantes entre si e dispostos
perpendicularmente em relação ao rio. Em cada transecção foram medidos os diâmetros à altura do
peito (DAP) de troncos de árvores escolhidos ao acaso. Aleatoriamente, uma área de 0,25 m² no
solo e dentro da transecção foi escolhida e utilizada para a coleta de serapilheira, a qual foi
armazenada em sacos plásticos. Registros fotográficos do dossel foram realizados exatamente acima
deste ponto, a fim de estimar a porcentagem de cobertura vegetal de cada área, determinadas com o
uso do programa Image J. Tais características foram utilizadas para verificar o grau de estruturação
dos fragmentos florestais coletados.
O material coletado foi analisado em laboratório: com auxílio da balança de precisão foram
medidos o peso total. Os invertebrados foram triados com auxílio de um sistema de duas peneiras
com malhas de 1 mm e 2 mm. Os indivíduos triados na malha 2 mm foram quantificados
visualmente, enquanto os na malha 1 mm foram contabilizados com uso de estereoscópio. Para as
verificações estatísticas foi utilizado Teste-t para averiguar os DAP entre as áreas e regressão linear
para a análise da relação entre a densidade de serapilheira e a abundância de invertebrados.
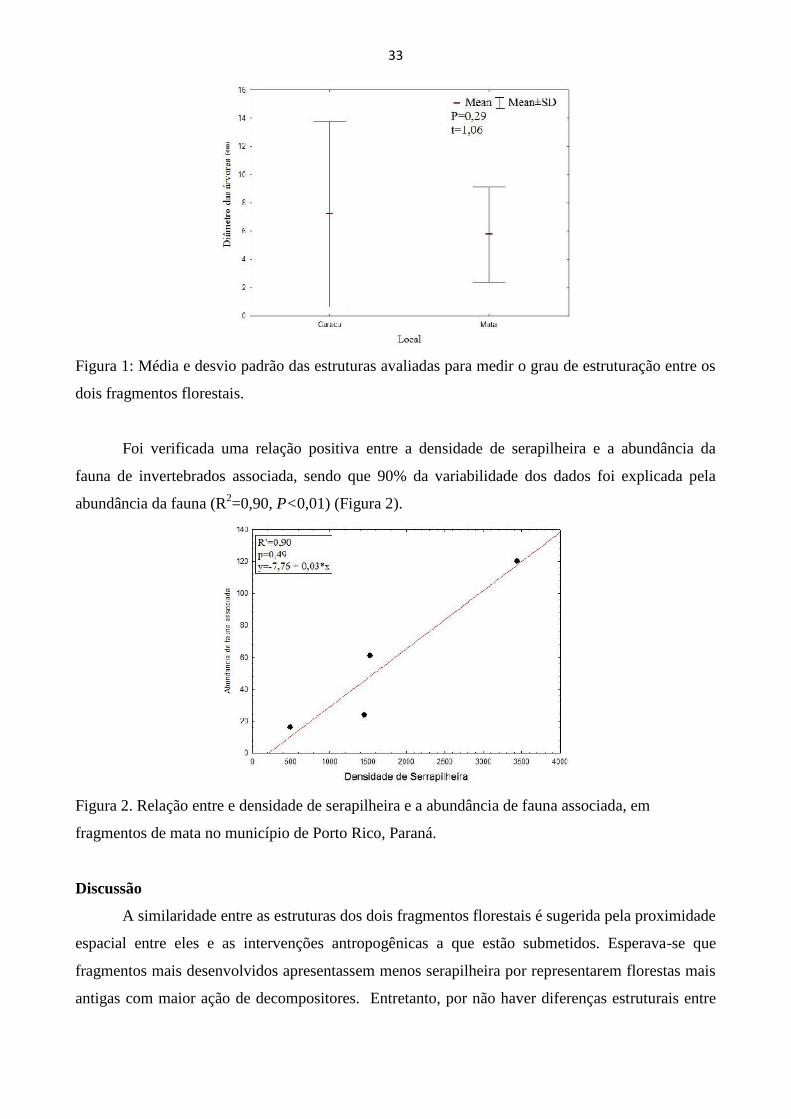
Resultados
O DAP médio das árvores das duas áreas não diferiu entre si (Fig. 1), bem como a
porcentagem de cobertura do dossel, o que impossibilitou o teste da hipótese de que florestas mais
estruturadas apresentam uma menor densidade de serapilheira, porque os fragmentos foram
semelhantes entre si em grau de estruturação.
33
Figura 1: Média e desvio padrão das estruturas avaliadas para medir o grau de estruturação entre os
dois fragmentos florestais.
Foi verificada uma relação positiva entre a densidade de serapilheira e a abundância da
fauna de invertebrados associada, sendo que 90% da variabilidade dos dados foi explicada pela
abundância da fauna (R2=0,90, P<0,01) (Figura 2).
Figura 2. Relação entre e densidade de serapilheira e a abundância de fauna associada, em
fragmentos de mata no município de Porto Rico, Paraná.
Discussão
A similaridade entre as estruturas dos dois fragmentos florestais é sugerida pela proximidade
espacial entre eles e as intervenções antropogênicas a que estão submetidos. Esperava-se que
fragmentos mais desenvolvidos apresentassem menos serapilheira por representarem florestas mais
antigas com maior ação de decompositores. Entretanto, por não haver diferenças estruturais entre

34
os fragmentos estudados, ou seja, os dois fragmentos se encontrarem em estágios de
desenvolvimento semelhantes, a correlação entre estrutura da floresta e densidade de serapilheira
foi nula.
A serapilheira constitui um ambiente adequado para o desenvolvimento da fauna edáfica,
sendo que sua composição pode variar com o clima e a vegetação. Sendo um ambiente adequado,
devido à riqueza de matéria orgânica, maiores densidades de microorganismos associados podem se
alojar em maiores densidades de serapilheira acumulada (Hofer et al., 2001), confirmado no
presente trabalho. Os invertebrados de vários grupos utilizam este ambiente para o forrageamento,
como abrigo e proteção, em especial nos estágios imaturos, para a diapausa, para oviposição,
crescimento, desenvolvimento (Jacot, 1940). Variações da temperatura ou outros fatores abióticos
também podem explicar algumas alterações na densidade dos artrópodes (Basset, 1966), dentre
outros organismos invertebrados terrestres. Adicionalmente pode haver preferências de
invertebrados por componentes estruturais das plantas, devido ao teor de celulose ou baixos teores
de ligninas e taninos (Loranger et al. 2002).
Agradecimentos
Agradecemos ao doutorando Vinícius pela ajuda prestada durante a escrita do trabalho, bem
como ao prof. Marcos pela ajuda durante a elaboração do projeto.
Referências
Basset Y 1966 The seasonality of arboreal arthropods foraging within an Australian rainforest tree.
Ecological entomology 16: 265 – 278.
Costa CCA, Camacho RGV, Macedo ID, Silva PCM 2010 Análise comparativa da produção de
serapilheira: em fragmentos arbóreos e arbustivos em área de caatinga na Flona de Açu -
RN. Revista Árvore 34: 259-265.
Figueiró AS 2005 Mudanças ambientais na interface floresta-cidade e propagação de efeitos de
borda no Maciço da Tijuca – Rio de Janeiro, RJ. Tese (Doutorado em Geografia) –
Programa de Pós-Graduação em Geografia, Universidade Federal do Rio de Janeiro, Rio de
Janeiro.
Garay I, Kindel A 2001 Diversidade funcional em fragmentos de Floresta Atlântica. Valor
indicador das formas de húmus florestais. Pages 350-368 in I Garay, B Dias,
eds. Conservação da biodiversidade em ecossistemas tropicais: avanços conceituais e
revisão de novas metodologias de avaliação e monitoramento. Vol. 1 Vozes, Petrópolis.

35
Hofer H, Hanagarth W, Garcia M, Martius C, Franklin E, Rombke J, Beck L 2001 Structure and
function of soil fauna communities in Amazonian anthropogenic and natural ecosystems.
European Journal of Soil Biology 37: 229 – 235.
Jacot AP 1940 The fauna of soil. The Quaternaly Review of Biology 15 (1): 28 – 58.
Loranger G, Ponge JF, Imbert D, Lavelle P 2002 Leaf decomposition in two semi-evergreen
tropical forests: influence of litter quality. Biology and Fertility of Soils 35 (4): 247–252.
Moraes RM, Delitti WBC, Struffaldi-DE Vuono Y 1999 Litterfall and litter nutrient content in two
Brazilian Tropical Forests. Revista Brasileira de Botânica 22(1):09-16.
Sundarapadian SM, Swamy PS 1999 Litter production and leaf-litter decomposition os selected tree
species in tropical forests at Kodayar in the Western Ghats, India. Forest Ecology and
Management 123: 231 – 244.

36
ESCONDENDO A UTRICULARIA FOLIOSA: SERÁ QUE ENGANAMOS O
ZOOPLÂNCTON?
Ariane Ghelfi, Bianca Possamai, Daniel Santos, Juliana Wojciechowski e Vivian Cionek
Resumo: As plantas aquáticas carnívoras do gênero Utricularia podem consumir diferentes
organismos aquáticos, como o zooplâncton. A fim de testar o efeito da camuflagem da macrófita
Utricularia folisosa na predação do zooplâncton, foi desenvolvido um experimento com
tratamentos em que a planta foi exposta e camuflada. Não foram verificadas diferenças
significativas nas densidades de zooplâncton entre tratamentos, o que reflete a biologia dos
organismos, que não são visuais, e erros experimentais. Conclui-se, portanto, que a camuflagem de
Utricularia foliosa não influencia a predação de zooplâncton.
Palavras-chave: planta carnívora, predação, camuflagem.
Introdução
Utricularia foliosa é uma planta carnívora que pode obter recursos além da fotossíntese a
partir da predação, devido à presença de utrículos capazes de digerir diferentes organismos
aquáticos. Essa estratégia auxilia a planta a superar desvantagens ecológicas associadas a falta de
nutrientes (Guisande et al., 2004). Uma grande diversidade de itens alimentares pode ser encontrada
em utrículos dessa espécie, que podem variar desde algas perifíticas aderidas aos seus próprios talos
(Eskinazi-Sant’Anna et al., 2002), espécies zooplanctônicas (Guisande et al., 2000), até alevinos e
larvas de insetos (Pompêo e Bertuga, 1996). Na natureza, essa planta carnívora ocorre emaranhada
entre outras macrófitas. As espécies zooplanctônicas tendem a explorar a estruturação ambiental
oferecida por macrófitas em busca de abrigo e recursos, como algas fitoplanctônicas e perifíticas,
culminando em maiores taxas de encontro do zoo com a planta carnívora (Pompêo e Bertuga,
1996).
O objetivo do trabalho foi investigar a taxa de predação de zooplâncton por Utricularia
foliosa em condições em que a planta se encontra camuflada por outra espécie de macrófita ou
exposta no ambiente, a fim de testar a hipótese de que a camuflagem aumenta a taxa de predação.
Espera-se que o zooplâncton visite mais a macrófita camuflada por fornecer maior área de abrigo,
culminando em redução na densidade de zooplâncton devido à predação.
Material e Métodos
Em campo foram coletados vários ramos de Utricularia foliosa e mantidos em um aquário
aerado e iluminado por 5 dias. O zooplâncton foi amostrado na manhã do experimento com rede de
plâncton (68 µm), nas proximidades de bancos de macrófitas. Ambos foram amostrados em uma

37
lagoa da planície de inundação do Alto Rio Paraná. Em laboratório a amostra contendo o
zooplâncton foi diluída em 1,4 litros da mesma água que foi usada para encher os aquários, a fim de
contabilizar a possível presença de espécies zooplanctônicas nessa água. Uma alíquota de 2 ml foi
retirada para a contagem do número de espécies zooplanctônicas.
Para o design experimental foram definidos 3 tratamentos: controle, sem camuflagem e com
camuflagem; designados aleatoriamente a aquários de 20 litros, preenchidos com água do rio
Paraná. Em todos os aquários foram depositados 200 ml da amostra de zooplâncton, com
estimativa de densidade de 5 indivíduos por ml. O controle foi estabelecido para contabilizar a
possível perda de indivíduos por morte natural e corrigir essa perda nos demais tratamentos. Nos
demais tratamentos foram acrescentados 50 g de ramos de U. foliosa com pelo menos 90% dos
utrículos vazios. Para o tratamento “com camuflagem” foram confeccionados bancos de macrófitas
artificiais constituídos de 20 fitas de plástico cinza de 33 cm de comprimento, amarradas a uma
base cerâmica. A camuflagem foi gerada com material artificial para não introduzir recurso
perifítico, que inevitavelmente estaria associado a qualquer outra macrófita natural. Com o intuito
de estimular a migração horizontal do zooplâncton em direção às macrófitas, foram introduzidos
dois cubos de gelo com sinal químico de peixes depois de 4h e 16h. O período de incubação foi de
20h, depois do qual toda a água do aquário foi filtrada em rede de plâncton (68 µm). As amostras
obtidas foram fixadas em formol e os indivíduos de 3 lâminas (cada uma com 2,5 ml de amostra)
foram contados para a obtenção da densidade de indivíduos após o experimento. Adicionalmente,
foram abertos 5 utrículos cheios de cada tratamento para a visualização do conteúdo interno. A fim
de comparar a densidade inicial e final dos grupos de zooplâncton foi utilizado um teste-t pareado,
bem como para comparação entre tratamentos (com camuflagem e sem camuflagem) foi realizada
um teste-t.
Resultados
Houve uma diminuição significativa na densidade total do zooplâncton ao final do
experimento (t17 = 5,11; P < 0,001), porém a magnitude da perda não pode ser atribuída somente a
predação, considerando que o tratamento controle (sem macrófitas) apresentou uma redução de 50
ind/L (inicial) para 4 ind/L (final). As densidades inicial e final de todos os grupos em ambos os
tratamentos foram significativamente diferentes (Tab. 1).
Tabela 1. Variação das densidades iniciais e finais dos grupos de zooplâncton nos dois tratamentos.
Tratamento Rotíferos Copépodos Cladóceros
Com camuflagem t5 = 14,74; P < 0,01 t5 = 11,89; P < 0,01 t4 = 71,96; P < 0,01
Sem camuflagem t5 = 20,85; P < 0,01 t5 = 10,37; P < 0,01 t4 = 17,26; P < 0,01

38
Não houve diferença significativa entre as densidades finais de zooplâncton nos tratamentos
com e sem camuflagem (t4 = 1,4 = 0,155; P = 0,714). A densidade final dos grupos também não
diferiu entre esses tratamentos: Copépodos: t4 = -0,46; P = 0,66; Cladóceros: t4 = -1,05, P = 0,83; e
Rotíferos: t4 = 0,44; P = 0,67) (Fig. 1).
Figura 1. Comparação das densidades finais de cada grupo zooplanctônico entre tratamentos.
Discussão
O declínio da biomassa zooplanctônica tem sido atribuído à predação por espécies de
Utricularia (Harms, 2002), no entanto, a camuflagem utilizada no presente estudo não afetou a
movimentação do zooplâncton, culminando em densidades muito similares dos grupos. Na
realidade, a camuflagem teve pouca influência no comportamento do zooplâncton, uma vez que
esses organismos são estimulados quimicamente e não identificam alvos através da visão. Cabe
destacar que na natureza a carnívora não ocorre sozinha, por isso simulamos a macrófita camuflada.
A procura pela área com a macrófita poderia estar mais relacionada com a busca do perifíton
associado aos talos da Utricularia, como demonstram alguns estudos da interação Utricularia x
zooplâncton (Sanabria-Aranda et al., 2006; Gross et al., 2007), se houvesse maior tempo para que
esse processo pudesse ser visualizado. Espécies do gênero Utricularia podem obter recursos através
da fotossíntese e dos utrículos, e o investimento em utrículos aumenta em ambientes com menor
qualidade nutricional (Guisande et al., 2004). As plantas do experimento foram mantidas em
aquário com água do rio Paraná e iluminação, o que pode ter contribuído para a obtenção de
recursos através da fotossíntese, culminando em menores taxas de predação de zooplâncton.
O decréscimo acentuado das densidades do zooplâncton nos tratamentos pode ser atribuída,
primeiramente, à um erro na estimativa inicial da densidade de zoo, que pode ter sido enviesada
devido ao uso de uma pipeta de plástico para a contagem da amostra, uma vez que ela é altamente

39
seletiva para animais maiores, culminando na estimativa de densidade inicial muito maior do que a
real. Cabe destacar que apesar de não contribuir com o decréscimo da densidade, houve predação,
uma vez que foram registrados dois organismos zooplanctônicos no interior de utrículos. Conclui-
se, portanto, que a camuflagem de Utricularia foliosa não influencia a predação de zooplâncton e
que esta interação deve ser muito mais influenciada pela busca de recursos (perifíton) pelo zoo,
culminando em maiores possibilidades de encontro dos organismos com os utrículos.
Referências Bibliográficas
Díaz-Olarte J, Valoyes-Valois V, Guisande C, Torres NN, González-Bermudez A, Sanabri-Aranda
L, Majarrés-Hernádez AM, Duque SR, Marciales LJ, Nuñez-Avellaneda M 2007
Periphyton and phytoplaknton associated with the tropical carnivorous plant Utricularia
foliosa. Aquatic Botany 87: 285-291.
Eskinazi-Sant’anna EM, Maia-Barbosa PM, Braz SM, Faria VR, Barbosa FAR 2002 Conteúdo
alimentar em macrófitas aquáticas carnívoras: Utricularia foliosa L. e U. hydrocarpa Vahl.
(Tubiflorae, Lentibulariaceae) Acta Scientiarum 24(2): 297-301.
Gross EM, Nee Körner H, Lombardo S, Mulderij G 2007 Searching for allelopathhic effects of
submerged macrophytes on phytoplankton – state of the art and open questions.
Hydrobiologia 584: 77-88.
Guisande C, Andrade C, Granado-Larencio C, Duque Sr, Núñez-Avellaneda M 2000 Effects of
zooplâncton and conductivity on tropical Utricularia foliosa investment in carnivory.
Aquatic Ecology 34: 137-142.
Guisande C, Aranguren N, Andrade-Sossa C, Prat N, Granado-Lorencio C, Barrios Ml, Bolivar A,
Núnez-Avellaneda M, Duque Sr 2004 Relative balance of the cost benefit associated with
carnivory in the tropical Utricularia foliosa. Aquatic Botany 80: 271-282.
Harms S, Johansson F 2000 The influence of prey behaviour on prey selection of the carnivorous
plant Utricularia vulgaris. Hydrobiologia, 427, 113–120.
Kosiba P 1992 Studies on the ecology of Utricularia vulgaris L. II. Physical, chemical and biotic
factors and the growth of Utricularia vulgaris L. in cultures in vitro. Ekologia Polska, 40,
193–212.

40
Pompêo MLM, Bertuga M 1996 Captura de organismos planctônicos pelas plantas carnívoras do
gênero Utricularia (Angiospermae, Dicotyledoneae). Revista Brasileira de Biologia
35(14): 697-703.
Sanabria-Aranda L, González-Bermúdez A, Torres N, Guisande C, Manjarrés-Hernández A,
Valoyes-Valois V, Díaz-Olarte V, Andrade-Sossa C, Duque SR 2006 Predation by the
tropical plant Utricularia foliosa. Freshwater Biology 51: 1999-2008.

41
PARTIÇÃO DE DIVERSIDADE EM MACROINVERTEBRADOS DE SERAPILHEIRA EM ILHA E
CONTINENTE
Fernanda Ceschin, Letícia Cucolo Karling, Luciana Zago, Renata Gabriela Noguchi e Simone
Satsuki Takeuchi
Projeto livre
Resumo
Considerando-se que a fauna habitante da serapilheira pode possuir baixas capacidades de
dispersão, visamos testar a hipótese de que menores escalas são mais importantes na determinação
da diversidade de macroinvertebrados edáficos. Os resultados apontaram que maiores escalas são
mais importantes na determinação da diversidade da fauna edáfica, o que pode ser resultante da
existência de uma grande barreira geográfica, de um evento recente de inundação e da amostragem
insuficiente para a representação de toda a comunidade no presente trabalho.
Palavras-chave: Diversidade α, Diversidade β, Ilha Mutum, Rio Paraná.
Introdução
A diversidade de espécies é uma relação entre o número de espécies (riqueza de espécies) e
a distribuição do número de indivíduos entre as espécies (equitabilidade) (Walker 1989). Para
Magurran (2004), diversidade alfa (α) ou diversidade local baseia-se no número total de espécies
em uma localidade, a diversidade gama (γ) ou diversidade regional, na soma do número de espécies
observados nas diferentes localidades, e a diversidade beta (β) é a de mudança de espécies ao longo
dessas localidades.
O sistema solo-serapilheira, composto por um mosaico de condições microclimáticas, é o
habitat natural para uma grande variedade de organismos responsáveis por inúmeras funções (Moço
et al. 2005). A quantidade de serapilheira depositada no solo depende de fatores bióticos, como as
espécies vegetais envolvidas e a estrutura do dossel, e de fatores abióticos, como temperatura e
inundações (Kataguiri 2006). A fauna que habita esses sistemas é composta por organismos
pequenos, de 4 µm até 20 mm, que podem ter uma capacidade de dispersão muito limitada (Moço et
al. 2005). Sob esse contexto, o presente trabalho objetiva testar a hipótese de que menores escalas
são mais importantes na determinação da diversidade de macroinvertebrados em decorrência do

42
baixo potencial de deslocamento dos indivíduos, de modo que eles predominantemente se
apresentariam agregados localmente.
Material e métodos
Os dados foram coletados no mês de junho de 2013, na ilha Mutum e na margem direita do
rio Paraná (continente), nos quais foram amostrados invertebrados da serapilheira. Em cada
localidade foram determinados dois pontos de amostragem nas quais foram dispostas quatro
parcelas de 0,25 x 0,25 m² distantes uma da outra por 10 m de modo a formar um quadrado com as
parcelas em seus vértices. A serapilheira de cada parcela foi recolhida rapidamente e armazenada
em sacos plásticos. Em laboratório, os invertebrados presentes na serapilheira foram triados por
busca ativa e classificados por morfotipos com auxílio de lupas.
Para se testar a hipótese de que a diversidade beta varia principalmente em escala pequenas
foi realizada a partição da diversidade gama em diferentes componentes através da função adipart
para o software R. A contribuição da diversidade em diferentes escalas encontradas na partição
(diversidade média de cada quadrado; variação da diversidade entre parcelas; e variação da
diversidade entre localidades) foram comparados com aqueles esperados em um modelo nulo no
qual o pool de espécies de uma escala é aleatorizado entre suas unidades amostrais. Adicionalmente
foi realizada uma Análise de Coordenadas Principais para visualizar padrões na composição de
espécies.
Resultados
Foi registrado um total de 398 organismos invertebrados associados à serapilheira,
diferenciados em 91 morfotipos pertencentes a diferentes grupos (Platyhelminthes, Oligochaeta,
Insecta, Mollusca). Desses morfotipos 27 foram exclusivos de ilha e 57 exclusivos de continente.
Os demais sete morfotipos ocorreram em ambas às localidades.
A diversidade alfa e beta 1 foram significativamente menores do que esperado pelo modelo
nulo, enquanto que a beta 2 foi não significativamente diferente (Tabela 1 e Figura 1).
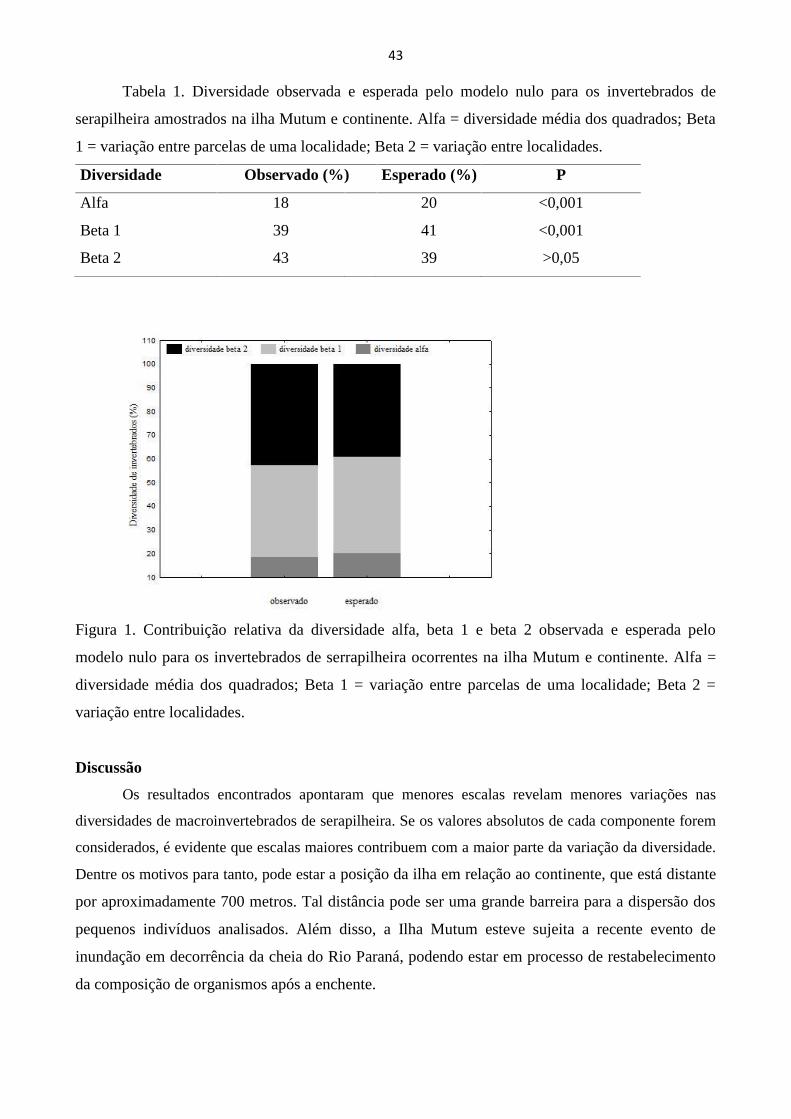
43
Tabela 1. Diversidade observada e esperada pelo modelo nulo para os invertebrados de
serapilheira amostrados na ilha Mutum e continente. Alfa = diversidade média dos quadrados; Beta
1 = variação entre parcelas de uma localidade; Beta 2 = variação entre localidades.
Diversidade Observado (%) Esperado (%) P
Alfa 18 20 <0,001
Beta 1 39 41 <0,001
Beta 2 43 39 >0,05
Figura 1. Contribuição relativa da diversidade alfa, beta 1 e beta 2 observada e esperada pelo
modelo nulo para os invertebrados de serrapilheira ocorrentes na ilha Mutum e continente. Alfa =
diversidade média dos quadrados; Beta 1 = variação entre parcelas de uma localidade; Beta 2 =
variação entre localidades.
Discussão
Os resultados encontrados apontaram que menores escalas revelam menores variações nas
diversidades de macroinvertebrados de serapilheira. Se os valores absolutos de cada componente forem
considerados, é evidente que escalas maiores contribuem com a maior parte da variação da diversidade.
Dentre os motivos para tanto, pode estar a posição da ilha em relação ao continente, que está distante
por aproximadamente 700 metros. Tal distância pode ser uma grande barreira para a dispersão dos
pequenos indivíduos analisados. Além disso, a Ilha Mutum esteve sujeita a recente evento de
inundação em decorrência da cheia do Rio Paraná, podendo estar em processo de restabelecimento
da composição de organismos após a enchente.

44
Boieiro et al. (2013) observou que a baixa diversidade em ilhas pode ser explicada pela sua
idade geológica, as distância da fonte mais próxima de colonização, a estrutura da floresta e o nível
de distúrbio do habitat. Assim, os aspectos estruturais da floresta da ilha, como áreas alagadas,
somado à presença de efeitos antropogênicos (residência no local e inserção de espécies vegetais
exóticas), podem ter refletido na diferença da diversidade dos invertebrados em relação a
diversidade do continente.
As pequenas distâncias entre as parcelas também podem ter influenciado os resultados, já
que abordam distâncias insuficientes para contemplar uma maior heterogeneidade local. Além
disso, os esforços amostrais podem ter sido insuficiente para a representação de ambos os locais,
Ilha e Continente, o que pode ter intensificado as diferenças encontradas entre as mesmas.
Referências bibliográficas
Boieiro M, Carvalho JC, Cardoso P, Aguiar CAS, Rego C 2013 Spatial factors play a major role as
determinants of endemic ground beetle beta diversity of madeira island laurisilva. PLoS
ONE 8(5): e64591. doi:10.1371/journal.pone.0064591.
Kataguiri VS 2006 Restabelecimento da fauna edáfica e a qualidade da serapilheira na Floresta da
USP – área de reflorestamento de Floresta Estacional Semidecidual. Dissertação de
Mestrado do Programa de Pós-Graduação em Entomologia da Universidade de São Paulo,
Ribeirão Preto.
Magurran AF 2004. Measuring Biological diversity. Blackwell, Oxford.
Moço MKS, Gama-Rodrigues EF, Gama-Rodrigues AC, Correia ME 2005 Caracterização da Fauna
Edáfica em Diferentes Coberturas Vegetais na Região Norte Fluminense. Revista Brasileira
de Ciências do Solo 29: 555-564.
Walker D 1989 Diversity and stability. Pages 115-146 in Cherrett JM (ed.). Ecological concepts.
Oxford, Blackwell Scientific Public.

45
EFEITO DA ALTERAÇÃO DA TURBIDEZ DA ÁGUA NAS COMUNIDADES DE FITOPLÂNCTON E
ZOOPLÂNCTON
Érika Maria Neif, Francisco von Hartenthal, Guilherme Landgraf, Rafael A. Baggio, Sabrina
Calado
Resumo
A turbidez pode variar espacialmente e temporalmente nos ecossistemas aquáticos e afetar direta ou
indiretamente as comunidades de fitoplâncton e zooplâncton. Dessa forma, hipotetizamos que
existem diferenças entre a riqueza de espécies de fitoplâncton e zooplâncton no gradiente de
turbidez do rio Paraná, da margem esquerda (afetada pelo Rio Paranapanema) para a margem
direita. Os resultados obtidos não foram significativos, entretanto, mostraram uma tendência de
aumento da riqueza em níveis intermediários de turbidez.
Palavras-chave: comunidades bióticas, turbidez, riqueza de espécies, planície de inundação, Alto
Rio Paraná.
Introdução
A matéria orgânica suspensa é responsável por quase a totalidade da dispersão da luz e
presença de compostos húmicos na coluna d´água (Kirk 1985), sendo, juntamente com a matéria
inorgânica, uma das principais responsáveis pela determinação da turbidez em corpos hídricos
(Esteves 1998). A turbidez pode variar espacialmente e temporalmente em lagos e rios (Scheffer et
al. 1994, Abrams & Kattenfeld 1997) e afetar direta ou indiretamente praticamente todos os grupos
de organismos na coluna d´água (Salonen et al. 2009, Esteves 1998, Abrams & Kattenfeld 1997).
Outros fatores podem alterar a turbidez da água, como afluentes fluviais, que ainda podem
alterar as características nas áreas da foz. No caso do Rio Paraná, existe uma diferença pronunciada
entre as margens esquerda e o resto do rio devido ao grande aporte de águas do Rio Paranapanema,
o que causa uma variação na turbidez de uma margem para a outra.
A turbidez originada por substâncias inorgânicas é facilmente induzidas por distúrbios no
sedimento de fundo, devido à atividade biótica ou resuspensão induzida pelo vento (Horppila &
Nurminen 2003). A turbidez pode atuar como fator limitante nas interações predador-presa em
muitos ecossistemas aquáticos continentais (Abrams & Kattenfeld 1997), pois a habilidade de
presas e predadores em detectar uns aos outros é frequentemente alterada pelas alterações na
turbidez da água (Abrams & Kattenfeld 1997). Nurminem et al 2010 demonstraram que a predação
de Daphnia sp. por Perca fluviatilis e Rutilus rutilus diminui com o aumento da turbidez, podendo

46
levar a um aumento na abundância na comunidade zooplanctônica. Estes padrões se repetem para a
interação predador-presa entre organismos zooplanctônicos e fitoplanctônicos (Salonen et al. 2009).
Dessa forma, hipotetizamos que existem diferenças entre riqueza de fitoplâncton e
zooplâncton ao longo da variação de turbidez do Rio Paraná da margem esquerda (afetada pelo Rio
Paranapanema) para a margem direita (afetada por pela UHE de Porto Primavera), e que a
composição da comunidade de fitoplâncton afeta a composição da comunidade de zooplâncton.
Material e métodos
O estudo foi realizado no Rio Paraná, em frente à Base do Nupélia. Foram selecionados 6
pontos amostrais seguindo o gradiente de transparência da água, da margem esquerda para a direita
do rio. A turbidez foi avaliada a partir da transparência, sendo essa medida com auxilio de disco de
Secchi. Em cada ponto foram coletadas amostras de fitoplâncton e zooplâncton, cada qual em
triplicatas, através de redes de arrasto com malhas de 20 µm e 68 µm, respectivamente, em
lançamentos verticais. As amostras de fitoplâncton foram fixadas com solução de Transeau e
identificadas de forma qualitativa com auxílio de microscópio óptico e literatura especializada
(Bicudo & Menezes, 2006). O zooplâncton foi fixado com solução de formol a 4% e identificado
até o nível de Classe. As métricas utilizadas para testar a hipótese foram riqueza de espécies de
fitoplâncton e de zooplâncton e os gradientes de turbidez.
Para testar a hipótese de que áreas com maior turbidez apresentam maior riqueza de
fitoplâncton, foi utilizado o teste não paramétrico Kruskal-Wallis para as comunidades. Para
visualizar se a composição das espécies dessas comunidades entre as áreas de pequena e grande
turbidez, foi utilizada uma rede bipartida construída no software R (pacote Bipartite). Para testar a
influência da composição da comunidade fitoplanctônica na composição de zooplanctônica, foi
realizado um teste de Mantel (método de distância: Jaccard, 10000 permutações), implementado no
software R (pacote labdsv).
Resultados
A riqueza das comunidades diferiu entre os pontos, apresentando a maior no ponto 3
(Tab.1). O teste Kruskal-Wallis suporta a hipótese de diferenciação para o fitoplâncton (H = 3,9; P
< 0,05). Para o zooplâncton o teste Kruskal-Wallis não foi significante (H=3,13, P > 0,05).
Tab.1. Turbidez da água e abundância das comunidades de fitoplâncton e zooplâncton amostradasno Rio Paraná, por ponto e agrupando pontos com turbidez semelhantes.
Local Turbidez (cm) Fitoplâncton ZooplânctonP1 25 10 10P2 25 10 7P3 45 20 12P4 125 5 4P5 165 6 7

47
P6 165 7 5
Total Alta 26 16Total Baixa 11 10
As diferenças entre as comunidades de fitoplâncton e zooplâncton podem ser vizualizado narede bipartida (Fig. 1). Contudo, a composição da comunidade de fitoplâncton não demonstrou estarrelacionada à composição da comunidade de zooplâncton (r = -0,10; P > 0,05).
Discussão
Os resultados das análises estatísticas não foram significativos para a riqueza e composição
do zooplâncton, mas para o fitoplâncton sim. Desta forma, os resultados, mostraram uma tendência
de que a riqueza do fitoplâncton aumenta com o aumento da turbidez. Contudo, os dados brutos
sugerem que existe uma maior riqueza em níveis intermediários de transparência da água,
possivelmente devido a essas comunidades serem favorecidas com o aumento de matéria orgânica e
inorgânica, porém, quando em excesso, pode ocorrer a predominância de algumas espécies, o que
contribui para a diminuição da riqueza de espécies. A riqueza de zooplâncton deve possuir a mesma
relação de aumentar a riqueza com o aumento da turbidez, corroborando trabalho de Hulot et al.
(2000). Vale a pena ressaltar que nos locais de alta turbidez foram encontradas colônias de uma alga
potencialmente tóxica (Microcystis cf. aeruginosa). As margens do rio Paranapanema apresentam
grande atividade agrícola, o que pode ter contribuído para a presença desta espécie, a qual necessita
de uma alta concentração de nutrientes.

48
Figura 1. Composição das comunidas de zooplâncton e fitoplâncton em ambientes de alta e baixa turbidez no RioParaná.
Agradecimentos: Agradecemos ao ajudante de campo Goiaba, e aos professores André Padial e
Roger Mormul pela contribuição na elaboração deste trabalho.
Referências Bibliográficas
Bicudo CEM, Menezes M 2006. Gêneros de Algas de Águas Continentais do Brasil: Chave para
identificação e descrições. São Carlos: RiMa, 2ed.
Esteves FA 1998. Fundamentos de Limnologia. Editora Interciência, Rio de Janeiro.
Horppila J & Nurminen L. 2003. Effects of submerged macrophytes on sediment resuspensios and
internal phosphorous loading in Lake Hiidenvesi (Southern Finland). Water research 37:4468-
4474.
Hulot FD, Lacroix G, Lescher-Moutoué F, Loreau M. 2000 Functional diversity governs ecosystem
response to nutrient enrichment. Nature 405: 340-344.

49
Kirk JTO 1985. Effects of suspensoids (turbidiy) on penetration of solar radiation in aquatic
ecosystems. Developments in Hydrobiology, 28:195-208.
Nurminen L, Pekcan-Hekim Z, Horppila J 2010. Feeding efficiency of planktivorous perch Perca
fluviatilis and roach Rutilus rutilus in varying turbidity: an individual-based approach.
Salonen, M & Engström-Öst J 2009. Effects of turbidity and zooplankton availability on the
condition and prey selection of pike larvae. Boreal environment research 14:981-989.
Scheffer M, Van den Berg, M, Breukellaar, Breukers, C, Coops H, Doef, R, Meijer, ML. Vegetated
areas with clear water in turbid shallow lakes. Aquatic botany. 49:193-196.

50
PREDAÇÃO SOBRE MODELOS APOSEMÁTICOS E CRÍPTICOS EM FRAGMENTOS ABERTOS E FECHADOS
Emanuel Razzolini, Fernando G. de Oliveira, Flávio M. Marteleto, Monica Vasques M. Pereira,
Natália C. L. dos Santos, Sandra M. N. de Andrade
Projeto Livre
Resumo
Diferentes grupos de animais respondem a diferentes estímulos visuais. Mamíferos em geral
não conseguem enxergar a faixa do vermelho, já aves apresentam uma complexidade visual maior,
possibilitando que os indivíduos enxerguem uma faixa ampla do espectro de luz. Para avaliar o
efeito do contraste de diferentes faixas do espectro de luz visível na predação, foram comparados a
predação em lagartas artificiais de diferentes cores. Tal avaliação também foi comparada em
diferentes ambientes com intuito de determinar a predação em locais onde há baixa e alta incidência
de luz.
Palavras-chave: Espectro da luz, contraste, aposematismo.
Introdução
Padrões crípticos e aposemáticos são mecanismos de defesa encontrados na natureza que se
baseiam em confundir ou alertar predadores. Padrões de coloração crípticos minimizam a
probabilidade de detecção dos organismos no ambiente, enquanto espécies aposemáticas
apresentam cores conspícuas como sinal de advertência a seus predadores, anunciando seu potencial
tóxico e impalatabilidade (Endler 1978).
O padrão de cor dos animais pode mudar de críptico para conspícuo, ou vice-versa, de
acordo com a iluminação do ambiente. Em ambientais florestados, a complexidade estrutural do
dossel determina tanto a quantidade de luz quanto o espectro que atinge estratos inferiores em uma
floresta. A luminosidade destes locais é determinada pela geometria dos raios de luz incidentes,
pelas condições ambientais e pelo período do dia, afetando a capacidade de visão dos animais e
influenciando diretamente nas interações de predação de predadores visuais (Endler 1993).
Além disso, diferentes predadores, como aves e mamíferos, têm percepção distinta das
colorações das presas e acabam tendo limitação da altura em que podem predar organismos que
vivem no tronco doas árvores. A maioria dos mamíferos é dicromata, pois possuem somente dois
tipos de cones, não enxergando a faixa do vermelho. Outros vertebrados, como as aves, possuem

51
maior quantidade de cones, apresentando maior complexidade visual. Assim, a coloração críptica e
aposemática das presas é relativa ao tipo de predador.
O objetivo do presente estudo foi avaliar se há diferença na taxa de predação em lagartas
artificiais de diferentes colorações em dois níveis de cobertura vegetal, tendo como hipóteses: i) o
contraste entre as cores das lagartas artificiais com o fundo é diferente entre ambientes com dossel
fechado e aberto; ii) a predação entre lagartas com cores distintas é diferente entre ambientes com
dossel fechado e aberto.
Material e métodos
As coletas de dados foram realizadas na mata do Caracu, ao lado da base avançada do
Nupélia. Foram utilizadas lagartas artificiais como modelo de presa (mimetizando lagartas de
lepidópteros), feitas com massa de modelar atóxica. Essas lagartas foram fixadas em troncos de
árvores em duas alturas distintas do solo (20 cm e 130 cm), em dois tipos de áreas: áreas
sombreadas, onde existiria maior incidência de luz difusa, e áreas abertas, com maior incidência de
luz direta. Os valores de contraste para cada cor de larva foram calculados a partir de fotos obtidas
sem flash de luz.
As imagens foram desfocadas no Photoshop® para misturar os pixels e foram traçados três
transectos: um sobre a larva e dois adjacentes a ela (à direita e à esquerda). Em cada transecto
traçado, foram analisados 10 pontos. A medida da proporção RGB (vermelho, verde e amarelo) em
cada pixel de cada transecto foi calculada e a eles foi atribuída uma média de cada ponto para a
obtenção de um índice de contraste de cada larva com o respectivo fundo.
Resultados
Em ambientes fechados (69% de cobertura do dossel), a cor branca foi a que apresentou o
maior contraste em relação ao fundo, enquanto a amarela apresentou o menor. Já em regiões abertas
(3% de área coberta), o maior contraste foi encontrado com a cor amarela, e o menor com a cor
preta. Os contrastes das cores (Fig. 1) em ambientes fechados apresentaram maiores valores do que
em ambientes abertos (Fig. 2), apresentando uma correlação negativa entre os contrastes de regiões
abertas e fechadas (R2 = -0.8088, G.L = 4, P = 0,05).
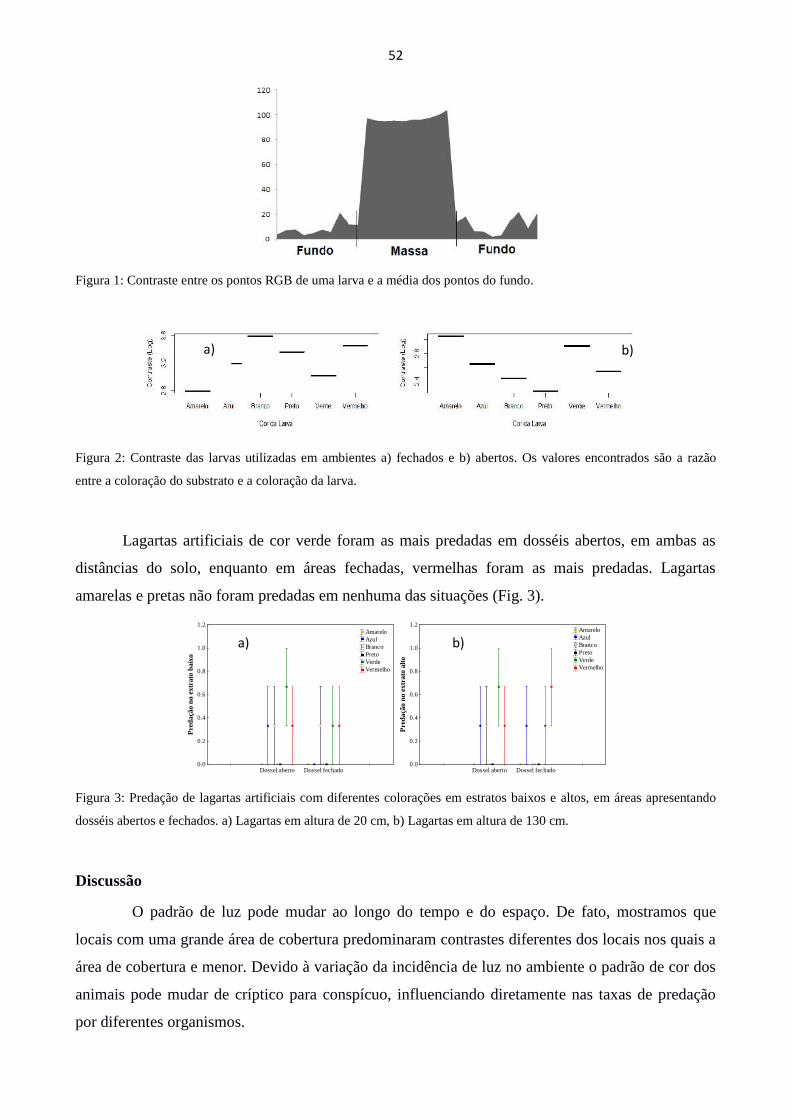
52
Figura 1: Contraste entre os pontos RGB de uma larva e a média dos pontos do fundo.
Figura 2: Contraste das larvas utilizadas em ambientes a) fechados e b) abertos. Os valores encontrados são a razão
entre a coloração do substrato e a coloração da larva.
Lagartas artificiais de cor verde foram as mais predadas em dosséis abertos, em ambas as
distâncias do solo, enquanto em áreas fechadas, vermelhas foram as mais predadas. Lagartas
amarelas e pretas não foram predadas em nenhuma das situações (Fig. 3).
Figura 3: Predação de lagartas artificiais com diferentes colorações em estratos baixos e altos, em áreas apresentando
dosséis abertos e fechados. a) Lagartas em altura de 20 cm, b) Lagartas em altura de 130 cm.
Discussão
O padrão de luz pode mudar ao longo do tempo e do espaço. De fato, mostramos que
locais com uma grande área de cobertura predominaram contrastes diferentes dos locais nos quais a
área de cobertura e menor. Devido à variação da incidência de luz no ambiente o padrão de cor dos
animais pode mudar de críptico para conspícuo, influenciando diretamente nas taxas de predação
por diferentes organismos.
Dossel aberto Dossel fechado0.0
0.2
0.4
0.6
0.8
1.0
1.2
Pre
daçã
o no
ext
rato
bai
xo
Amarelo Azul Branco Preto Verde Vermelho
Dossel aberto Dossel fechado0.0
0.2
0.4
0.6
0.8
1.0
1.2
Pre
daçã
o no
ext
rato
alto
Amarelo Azul Branco Preto Verde Vermelho
a) b)
a) b)

53
De acordo com resultados obtidos a cor mais contrastante em ambientes fechados foi
diferente das cores mais contrastantes em ambientes abertos, contrariando resultados anteriores
(veja PO3G1), que mostraram que a cor mais contrastante para ambientes fechados foram a branca
e a azul. No entanto, os resultados obtidos indicam que na área do estudo do presente trabalho, a cor
menos contrastante para ambientes fechados foi o azul. Vale ressaltar que a abertura de dossel da
área utilizada no estudo PO3G1 foi de 11%, enquanto que a utilizada realizada neste trabalho
apresentava uma área aberta de 31%. Diversos fatores relevantes no estudo devem ser levantados,
durante o período do experimento houve uma intensa precipitação no local amostrado, fazendo com
que o tempo de exposição fosse reduzido para apenas um período do dia.
Apesar dos dados utilizados terem respondido parcialmente nossas hipóteses, a utilização
de equipamentos como espectrofotômetro para medir a faixa de luz refletida poderia permitir uma
comparação mais acurada sobre a diferença de ambientes abertos e fechados. Espectros ultravioleta
e infravermelho poderiam ser incluídos, auxiliando a explicação de eventuais diferenças na
predação de presas com estratégias de defesa crípticas e aposemáticas.
Referências
Endler JA 1978 A predador´s view of animal color patterns. Evolutionary Biology 11:319:364.
Endler JA 1993 The Color of light in forests and its implications. Ecological monographs 63(1):1-
27.

54
DISPERSÃO DE FORMIGAS MEDIADA PELOS RECURSOS ALIMENTARES: THE ANTMATE FIGHTING
Diego Corrêa Alves, Hugo José Message, Lilian Paula Vasconcelos, Priscila Guedes Gambale,
Taise Miranda Lopes
Projeto Livre
Resumo
A competição interespecífica é um fator estruturante das comunidades e também pode influenciar
nos padrões de dispersão destes organismos. Essa teoria pode ser aplicada na dinâmica de dispersão
das colônias de formigas. Neste estudo, a média de abundância de formigas em locais com recurso
foi maior do que em locais sem recurso, corroborando com as hipóteses de que a dispersão das
formigas é determinada por busca de recursos alimentares.
Palavras-chave: Deslocamento, forrageamento, formigas.
Introdução
A competição interespecífica é um fator estruturante das comunidades e também pode
influenciar nos padrões de dispersão destes organismos. O grupo dos Hymenopteras apresentam
acentuados comportamentos dispersivos(Price, 1997), o que influencia a interação entre indivíduos
de populações diferentes, as quais sobrevivem e respondem de forma diferenciada às condições
ambientais e recursos, além de persistirem dentro de um contexto de interações bióticas (Fukami,
2004; Macedo-Soares et al., 2010).
As colônias de formigas são capazes de se auto-organizar com base em regras de
comportamento simples e uso de informações locais e comunicação indireta (Bonabeau et al.,
1997). Nestes sistemas existe uma divisão em castas, formados por rainhas e operários. As rainhas
são férteis e responsáveis pela geração de novos indivíduos, já os operários são inférteis e realizam
uma série de atividades dentro da colônia. Em geral, os operários dedicam-se aos cuidados de
ninhada, manutenção ninho, forrageamento e defesa da colônia. No entanto, os indivíduos não são
comprometidos a uma única tarefa para a vida inteira e trocam de tarefa à medida que amadurecem
(Holldobler et al., 1990).
O comportamento de forrageamento foi apontado por Wheller (1928) como a resposta da
fome da colônia. Esse comportamento faz com que a formiga deixe o ninho à procura de um

55
recurso, recolha o alimento e retorne ao ninho com parte do alimento, seja com material sólido ou
através do regurgito.
Assim, este trabalho foi desenvolvido buscando testar a hipótese de que a dispersão e
competição interespecíficas das formigas são determinadas pela busca de recursos alimentares e
área.
Materiais e Métodos
O estudo foi desenvolvido através de um experimento realizado na Base de Pesquisas
Avançadas do Nupélia/UEM (Porto Rico, Paraná). Para o experimento em laboratório dois
formigueiros distantes 40m entre si foram coletados utilizando enxadas e alocados em baldes de
20L. Foi preparado um doce de maça com açúcar para ser utilizado como fonte de recursos
alimentar.
Em laboratório foi construída uma estrutura composta por 5 áreas conectadas por 4
mangueiras de 20 cm de comprimento e 2,5 cm de diâmetro cada (Fig. 1). A estrutura foi composta
por dois terráreos, uma área de forrageamento e duas áreas vazias (todas de 15 cm de diâmetro).
Cada terráreo continha duas conexões, uma com a área de forrageamento e outra com a área vazia,
assim, a área de forrageamento era conectada aos dois terráreos (Fig. 1). Duas colônias distintas
foram alojadas em terráreos separados (550g cada terráreo), em seguida, foi realizada uma
aclimatação monitorada dos indivíduos por 5 horas, que possibilitou evidenciar o tempo de
organização social de cada colônia. As interações entre as colônias foram observadas e registradas.
Fig. 1. Esquema da estrutura construída para o primeiro experimento demonstrando o local onde
foram colocadas as colônias 1 e 2 (terráreos 2 e 3, respectivamente), local com recurso (terráreo 3) e
locais sem recurso (potes 1 e 5).
O comportamento das formigas foi gravado continuamente com uma câmera de alta
definição (Sony HX-10) durante uma hora durante a noite. A fim de evidenciar a abundância de
indivíduos no recurso foi realizada uma contagem no intervalo de cinco em cinco minutos por um
período de 9 horas. Encontros entre as duas colônias também foram contabilizados bem como o
tempo percorrido para a formiga de cada colônia chegar ao recurso.

56
Com objetivo de testar a primeira hipótese, foi realizado um Kruskal-Wallis para variância
heterogênea, tendo como variável resposta a abundância de indivíduos nas diferentes áreas (com e
sem recurso). Além disso, uma correlação de Spearman foi realizado a fim de verificar se existe
relação positiva entre o tempo de observação do experimento e a abundância de indivíduos
frequentando o local com recurso.
Resultados
O comportamento inicial de ambas as colônias foi de reconhecimento da área do pote e
transporte dos ovos para o interior do ninho, de maneira a se reestruturarem, por um período médio
de 2h. Durante 3 horas e meia, apenas alguns indivíduos da colônia 1 se deslocaram entre os potes.
Aparentemente, os primeiros deslocamentos foram realizados para o reconhecimento da área,
depois foi observado o início da utilização do recurso. Além disso, a colônia 1 passou a reestruturar
seu ninho, recobrindo com terra toda a superfície do pote, diminuindo progressivamente a
visibilidade do interior do ninho.
Por duas ocasiões formigas da colônia 1 que visitaram o terráreo 1 ficaram presas em um
vestígio de fita adesiva, e, durante este período, foi observado o comportamento de vários outros
indivíduos tentando resgatá-las. Por todo o período de observação a colônia 1 se mostrou mais
ativa e fez alguns deslocamentos entre os terráreos 1, 2 e 3; posteriormente também invadindo o
ninho da colônia 2, passando a ocorrer alguns conflitos e um comportamento de fuga de indivíduos
da colônia 2 para o terráreo 5.
A média da abundância de formigas que se deslocaram em direção ao recurso se diferiu
daquelas que se deslocaram aleatoriamente em direção ao compartimento sem recurso (KW-
H1,122=57,35; P<0,05; N=61) (Fig. 2 esquerda). Adicionalmente, a abundância de indivíduos que se
deslocaram em direção ao recurso apresentou uma tendência de aumento em relação ao tempo de
amostragem (R=0,83; P<0,05) (Fig. 2 direita).

57
Fig. 2. Esquerda: Média da abundância de indivíduos nas áreas com e sem recurso. Direita:
Abundância de indivíduos no tempo de observação (ranking).
Discussão
Nossos resultados demonstraram que a dispersão das formigas estudadas é direcionada,
sendo esta direção mediada pela busca do recurso alimentar. Este comportamento é plausível, já que
as formigas são organismos com grande capacidade sensorial (Bonabeau et al., 1997) e podem
utilizar desta característica para otimizar a demanda energética investida neste tipo de movimento.
O delineamento utilizado neste experimento propiciou um modelo para testar hipóteses
relacionadas a padrões de dispersão de populações. Este delineamento pode ser facilmente
expandido, através do aumento de terráreos e com diferentes configurações de conexão, assim
sendo um delineamento com potencial utilidade para estudar a dinâmica entre organismos e
possivelmente até metacomunidades.
Agradecimentos
Aos docentes do Curso de Campo pelas contribuições no delineamento dos experimentos.
Referências
Bonabeau E, Theraulaz G, Deneubourg JL, Aron S, Camazine S 1997 Selforganization in social
insects. Trends Ecol. Evol., 12, 188-193.
Fukami T 2004 Community assembly along a species pool gradient: implications for multiple-scale
patterns of species diversity. Population Ecology 46: 137-147
Holldobler B, Wilson E 1990 The Ants. Harvard University Press, Cambridge, MA.
Leibold MA, Holyak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB,
Law R, Tilman D, Loreu M, Gonzalez A 2004 The metacommunity concept: a framework for
multi-scale community ecology. 7:601-613.
Price, PW 1997 Insect ecology. New York: John Wiley & Sons, Inc.
Macedo-Soares PHM Petry AC Farjalla VF Caramaschi EP 2010 Hydrological connectivity in
coastal inland systems: lessons from a Neotropical fish metacommunity. Ecology of Freshwater
Fish 19: 7-18.
Wheeler WM 1928 The social insects. New York: Harcourt Brace.
Wilson DS 1992 Complex interactions in metacommunities, with implications for biodiversity and
higher levels of selection. Ecology 73:1984–2000.

58

59
FOCO NA ESCALA: IMPORTÂNCIA DA HIERARQUIA DE ESCALA NA PARTIÇÃO DA DIVERSIDADE
BETA EM AUTÓTROFOS AQUÁTICOS.
Ariane Ghelfi, Bianca Possamai, Daniel Santos, Juliana Wojciechowski e Vivian Cionek
Orientador: André Padial
Resumo
A diversidade regional pode ser particionada em componentes alfa e beta em diferentes escalas, as
quais tem seus graus de importâncias relativas investigados.Verificamos em qual escala a
diversidade beta é mais importante em estudos sobre assembleias de macrófitas e fitoplâncton em
lagoas da planície de inundação do Alto Rio Paraná. Para macrófitas observou-se uma maior
variação entre as espécies das diferentes lagoas, enquanto para o fitoplâncton a maior variação foi
entre amostras do mesmo local.
Palavras-chave: Variação de comunidades, turnover, fitoplâncton, macrófitas aquáticas.
Introdução
A diversidade de espécies da planície de inundação do Alto Rio Paraná é alta (Thomaz et al.,
1992) e pode ser particionada em três componentes principais de acordo com a escala de estudo
(Flach, 2012): Diversidade gama (γ), que corresponde ao número total de espécies de uma região;
Diversidade beta (β), que é a variação de espécies observada entre duas ou mais unidades espaciais
ou ainda a variação temporal de espécies em um local e; Diversidade alfa (α), que é definida como o
número de espécies encontradas em um local específico (Magurran, 2004). Em relação à
diversidade β, trabalhos prévios têm demonstrado que a composição de macrófitas e fitoplâncton
varia espacialmente, devido a aspectos limnológicos e morfométricos dentro de uma mesma lagoa,
e entre diferentes lagoas, possivelmente por dispersão e/ou heterogeneidade ambiental (Thomaz e
Bini, 2003; Margalef, 1983). Devido ao curto ciclo de vida, a composição do fitoplâncton pode
variar diariamente em decorrência da dinâmica de circulação da coluna de água (Barbosa e Padisák,
2002) e espacialmente devido à fatores como colonização e adaptação ao ambiente (Reynolds et al.,
2002).
O objetivo do presente estudo foi investigar a principal fonte de variação da diversidade
beta, a fim de testar a hipótese de que a diversidade beta varia principalmente entre lagoas, quando
comparada com as outras fontes de variação em menor escala espacial. Também é esperado que a
variação diária do fitoplâncton seja menos importante para compor a diversidade gama do que a
variação entre lagoas.
Material e Métodos

60
As amostragens foram realizadas na margem esquerda da Lagoa das Garças (22°43’27.18”S;
53°13’4.56”W) e Ressaco do Pau Véio (22°44’76”S; 53°15’11.16”W), ambas pertencente à sub-
bacia do Rio Paraná. As macrófitas foram amostradas aleatoriamente com um quadrado de 50x50
cm, em tréplicas, na região litoral e no meio do banco (6 amostras). Dois bancos foram amostrados
em cada lagoa (entrada e final), totalizando 12 amostras por lagoa. O fitoplâncton foi coletado a
sub-superfície, em tréplicas, na entrada e final de cada lagoa para a investigação da variação
espacial. Nos mesmos locais, foram tomadas amostras em dois períodos (manhã e tarde), a fim de
verificar variação temporal, totalizando 12 amostras por lagoa. Em laboratório, as amostras foram
sub-amostradas em três lâminas, observadas em microscópio e a composição de taxa foi registrada
(presença-ausência).
Com o intuito de verificar a fonte de variação que mais contribui para a diferença nos
componentes da diversidade beta para macrófitas e fitoplâncton foi realizada uma Análise de
Partição de Diversidade, implementada no programa estatístico R® (R Core Team, 2013). As escalas
definidas para investigação foram: a variação entre lagoas (β3), a posição dentro da lagoa (entrada e
final) (β2), posição no banco de macrófitas (regiões litorânea e meio) (β1 – macrófitas), os períodos
matutino e vespertino (β1 – fitoplâncton), dentro do quadrado (α1 – macrófitas) e dentro do pote (α1
– fitoplâncton). Uma NMDS foi aplicada aos dados de riqueza de espécies de macrófitas e
fitoplâncton a fim de visualizar a similaridade da composição entre as fontes de variação definidas
acima.
Resultados
Onze espécies de macrófitas foram identificadas, pertencentes a 8 gêneros e 8 famílias. Para
o fitoplâncton, 89 taxa foram identificados, pertencentes a 9 classes e 40 gêneros. A partição da
diversidade para a assembleia de macrófitas revelou que o valor observado para a diversidade α foi
significativamente menor que o esperado (propexp>obs>0.999). O valor observado do componente β1
foi similar ao esperado (propexp>obs =0.63), já os valores observados para β2 e β3 foram maiores do
que aqueles esperados ao acaso (propexp>obs<0.001) (Fig. 1b). De fato, a ordenação das macrófitas
revelou uma separação clara entre as diferentes lagoas, e também uma separação clara entre o final
e entrada do Ressaco do Pau Véio (Fig. 1a).

61
Figura 1. a. Porcentagem dos componentes alfa e beta observado e esperado da riqueza de espécies de macrófitasaquáticas em duas lagoas da planície de inundação do Alto Rio Paraná e b. Resultado do Escalonamentomultidimensional não-métrico (índice de Jaccard) diferenças nas principais escalas de variação (β2 e β3).
Para algas fitoplanctônicas, a partição da diversidade revelou que os valores observados para
a diversidade alfa (α) foram significativamente maiores que o esperado (propexp>obs<0.001). Os
valores observados dos componentes 1 e β2 variaram pouco menos que o esperado
(propexp>obs>0.999). Já os valores de diversidade observados na maior escala de variação (β3) foram
maiores do que seria esperado ao acaso (propexp>obs<0.001) (Fig. 2b). A importância da variação
local (α) é ressaltada pela ordenação do fitoplâncton, que mostra que as tréplicas nem sempre ficam
agrupadas em relação à outra fonte de variação superior (Fig. 2a). Também é possível observar uma
diferença entre lagoas (Figura 2b).
Observado Esperado30
40
50
60
70
80
90
100
% R
ique
za M
acró
fitas
β3β2β1α
> 0,999
< 0,001
< 0,001
= 0,029
A
-2 -1 0 1 2NMDS Eixo 1
-1.2-1.0-0.8-0.6-0.4-0.20.00.20.40.60.81.01.21.41.6
NM
DS
Eix
o 2
Garças_Final Garças_Entrada Pau Véio_Final Pau Véio_Entrada
B

62
Figura 2. a. Porcentagem dos componentes alfa e beta observado e esperado da riqueza de espécies de algasfitoplanctônicas em duas lagoas da planície de inundação do Alto Rio Paraná e b. Resultado do Escalonamentomultidimensional não-métrico (índice de Jaccard) diferenças nas principais escalas de variação (α). Símbolosrepresentam amostragens, sendo os de cor cinza representando o ressaco do Pau Véio e os preenchidos de preto a lagoadas Garças.
Discussão
Segundo Thomaz (2012) a estrutura das assembleias de macrófitas aquáticas da planície do
Alto Paraná está organizada em macro-escala e a composição de espécies varia de acordo com os
ambientes, o que corrobora nossos resultados. A riqueza de espécies local (α) foi baixa, indicando
que a maior parte da diversidade total é derivada dos componentes beta e que há alta agregação dos
indivíduos das espécies. De fato, macrófitas tipicamente desenvolvem bancos homogêneos. A
variação da composição de espécies entre lagoas pode estar relacionada a limitações de dispersão
das espécies e às características morfométricas e limnológicas de cada lagoa. A maior variação de
macrófitas dentro do ressaco do Pau Véio, ser explicada pelo domínio da espécie P. sagitarium no
trecho final, e a composição mais diversificada observada no trecho entrada.
A diversidade fitoplanctônica entre as amostras foi maior que o esperado, indicando que a
amostragem não representou a flora fitoplanctônica local. A diversidade β1 observada foi menor que
Observado Esperado0
20
40
60
80
100
% R
ique
za F
itopl
ânct
on
β3β2β1α
> 0,999
> 0,999
< 0,001
< 0,001
A
-1.2 -0.8 -0.4 0.0 0.4 0.8 1.2
NMDS Eixo1
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
NM
DS
Eixo
2
2
26
8
8
6 6
4 4
4
2
B

63
a esperada, portanto, a variação diária das espécies não foi tão ampla quanto o esperado ao acaso,
provavelmente devido a perda de alguns táxons por sedimentação ao longo do dia. Porém, os
determinantes da variação diária ainda precisam ser melhor avaliados. A pequena variação diária
observada também pode ser conseqüência de uma pequena variação das condições ambientais ao
longo do dia, que foi caracterizado por alta nebulosidade e pouca variação térmica, provavelmente
diminuindo efeitos de estratificação. Assim como para macrófitas a variação ente lagoas deve ser
devido à heterogeneidade ambiental. Porém, tal variação é pequena se comparada com a variação
das réplicas.
Nosso trabalho evidenciou que grupos biológicos, mesmo que com restrições ambientais
similares, variam em diferentes escalas. Macrófitas aquáticas variam em grandes escalas, pelo
menos no range de escalas estudadas. Porém, não encontramos tal padrão para o fitoplâncton, e a
variação entre réplicas provavelmente reflete a limitação do método para representar corretamente
um ponto de amostragem. De qualquer maneira estudos e mais escalas ainda são necessários para
determinar as principais escalas de variação de diferentes grupos biológicos em planícies de
inundação.
Referências Bibliográficas
Barbosa FA, Padisák J 2002 The Forgotten Lake Stratification Pattern: Atelomixis, And Its
Ecological Importance. Verhandlungen Internationale Vereinigung Für Theoretische Und
Angewandte Limnologie, n. 28, p. 1385-1395.
Cunha ER, Thomaz SM, Evangelista HBA, Carniato J, Souza CF, Fugi R 2011 Small-sized fish
assemblages do not differ between a native and a recently established non-indigenous
macrophyte in a Neotropical ecosystem. Natureza & Conservação, v. 9, n. 1, p. 61-66.
Ferreira FA, Mormul RP, Pedralli G, Potti V.J, Potti A 2010 Estrutura da comunidade de macrófitas
aquáticas em três lagoas do Parque Estadual do Rio Doce, Minas Gerais, Brasil. Hoehnea, v.
37, n. 1, p. 43-52.
Flach PZS, Ozorio CP, Melo AS 2012. Alpha and beta components of diversity of freshwater
nematodes at different spatial scales in subtropical coastal lakes. Fundam. Appl. Limnol., v.
180, n. 3, p. 249-258.
Magurran AF 2004 Measuring Biological diversity. Blackwell, Oxford.
Margalef R 1983 Limnologia. Barcelona: Omega.
Reynolds CS, Huszar V, Kruk, C, Naselli-Flores L, Melo S. Towards a functional classification of
the freshwater phytoplankton. Journal of Plankton Research, v. 24, p. 417-428, 2002.

64
Souza-Filho EE, Stevaux, JC 2001 Locais de amostragem. In: Relatório Final do Projeto de
Pesquisa ecológica de Longa Duração (PELD) – A Planície Alagável do Alto Rio Paraná.
7p.
Thomaz SM et al. 1992 Características limnológicas de uma estação de amostragem do alto rio
Paraná e outra do baixo rio Ivinheima - (PR, MS-Brasil). Acta Limnologica Brasiliensia,
Botucatu, v. 4, p. 32-52.
Thomaz SM et al. 2002 Macrófitas aquáticas da planície de inundação do Alto Paraná: listagem de
espécies e padrões de diversidade em ampla escala, Relatório Peld- UEM.
Thomaz SM, Bini LM 2003 Ecologia e manejo de macrófitas aquáticas. Maringá: EDUEM, 341p.

65
COMPOSIÇÃO SEXUAL DE GRUPOS DE ALOUATTA SPP.: DE ESTUDOS PONTUAIS À PADRÕES.
Diego Correa Alves, Fernanda Ceschin, Letícia Cucolo Karling, Luciana Zago, Monica Vasques
Mangas Pereira
Orientador: Fernando C. Passos
Resumo
Verificamos se a proporção sexual de fêmeas e machos adultos dos grupos de primatas das
espécies Alouatta caraya, Alouatta clamitans e híbridos é diferente da esperada ao acaso. Os testes
foram realizados a partir da meta-análise de 14 artigos relacionados ao tema retirados de bancos de
dados da internet. Os resultados encontrados confirmam a proposta de diferentes autores de que os
grupos das espécies A. caraya e A. clamitans possuem um maior número de fêmeas do que de
machos adultos.
Palavras-chave:
Alouatta caraya; Alouatta clamitans; Híbridos; sex ratio.
Introdução
Espécies do gênero Alouatta formam grupos sociais de 2 a 45 indivíduos (Neville et al.
1988), havendo variações intra e interespecíficas nestes valores (Treves 2001). Tais variações
podem estar relacionadas à ampla distribuição do gênero que se estende desde o México até Sul do
Brasil e o Norte da Argentina, englobando diferentes biomas (Gregorin 2006). Os grupos são
geralmente formados por um macho adulto dominante, podendo haver outros machos subordinados,
uma ou várias fêmeas adultas e indivíduos imaturos (Neville et al. 1988). As composições sexo-
etárias encontradas para o gênero sugerem sistemas sociais poligínicos, onde possivelmente apenas
o macho dominante tem acesso às fêmeas reprodutivas (Glander 1980), com eventos sexuais sendo
raros para machos subordinados (Miranda 2009).
A composição sexo-etária dos grupos permite a avaliação de alterações e viabilidade de
populações de primatas não humanos (Treves 2001, Zucker & Clarke 2003). Para o gênero
Alouatta, por exemplo, o sucesso reprodutivo pode ser influenciado pela razão sexual (Treves
2001). Entretanto, não há consenso quanto aos efeitos de variações na razão sexual. Enquanto Ostro
(2001) argumenta que maiores proporções de machos podem ser vantajosas em conflitos
intergrupais, vigilância e defesa do território, Wang & Milton (2003) argumentam que esse maior
número de machos pode aumentar os custos energéticos envolvidos em interações sociais
intragrupais, como a competição por fêmeas em estro.

66
Diante disto, tivemos como objetivo central testar se a proporção entre fêmeas e machos
adultos de grupos de Alouatta caraya, Alouatta clamitans e grupos mistos em diferentes localidades
é diferente da esperada ao acaso (0,5), partindo da hipótese de que esta proporção é maior do que a
esperada ao acaso.
Material e métodos
Os artigos utilizados nesta pesquisa foram obtidos por meio dos bancos de dados do ISI
(Institute for Scientific Information; www.isiknowledge.com) e do Scielo (Scientific Electronic
Library online; www.scielo.org) durante o mês de junho de 2013.
Para a busca de artigos foram utilizados os seguintes termos: group, composition, Alouatta
caraya, Alouatta clamitans. Dos artigos resultantes da busca, foram utilizados para a análise aqueles
que continham o número de machos adultos, fêmeas adultas, indivíduos imaturos, localização dos
grupos. Artigos que analisaram grupos temporalmente tiveram apenas os dados do primeiro ano de
visualização considerados. Grupos com menos de um casal adulto não foram considerados na
análise.
A partir destes dados foi realizado um teste T pareado para verificar se a média das
diferenças entre fêmeas e machos adultos dentro dos conjuntos de grupos de cada espécie e
localidade era significativa. Além disso, foi calculada a estatística Q, a fim de confirmar a
homogeneidade entre os grupos de primatas (Borenstein et al 2009). Com base nesta
homogeneidade foi utilizada a meta-análise com efeitos fixos através da estatística Risk Difference
(Borenstein et al 2009).
Resultados
Foram registrados 14 artigos relacionados à composição sexual das espécies A. caraya, A.
clamitans e seus híbridos na América do Sul, sendo que no total foram analisados 88 grupos de
primatas. Os estudos resgatados na busca predominaram na região sul do Brasil e norte e nordeste
da Argentina (Tab.1).
De maneira geral, o teste T pareado indicou que os conjuntos de grupos de primatas de
mesma região e espécie, apresentaram diferenças significativas no número de indivíduos adultos
fêmeas e machos. No entanto, esta diferença não foi significativa em alguns conjuntos de grupos de
primatas. Após a meta-análise ficou claro que estes resultados destoantes para algumas das análises
possivelmente são decorrentes do pequeno tamanho amostral ou peculiaridades destes estudos.
Tabela 1: Médias dos números de indivíduos machos e fêmeas adultas, sua diferença (∆), número de
grupos de primatas (N) para cada localidade e espécie. Estatística T, grau de liberdade (DF) e valor
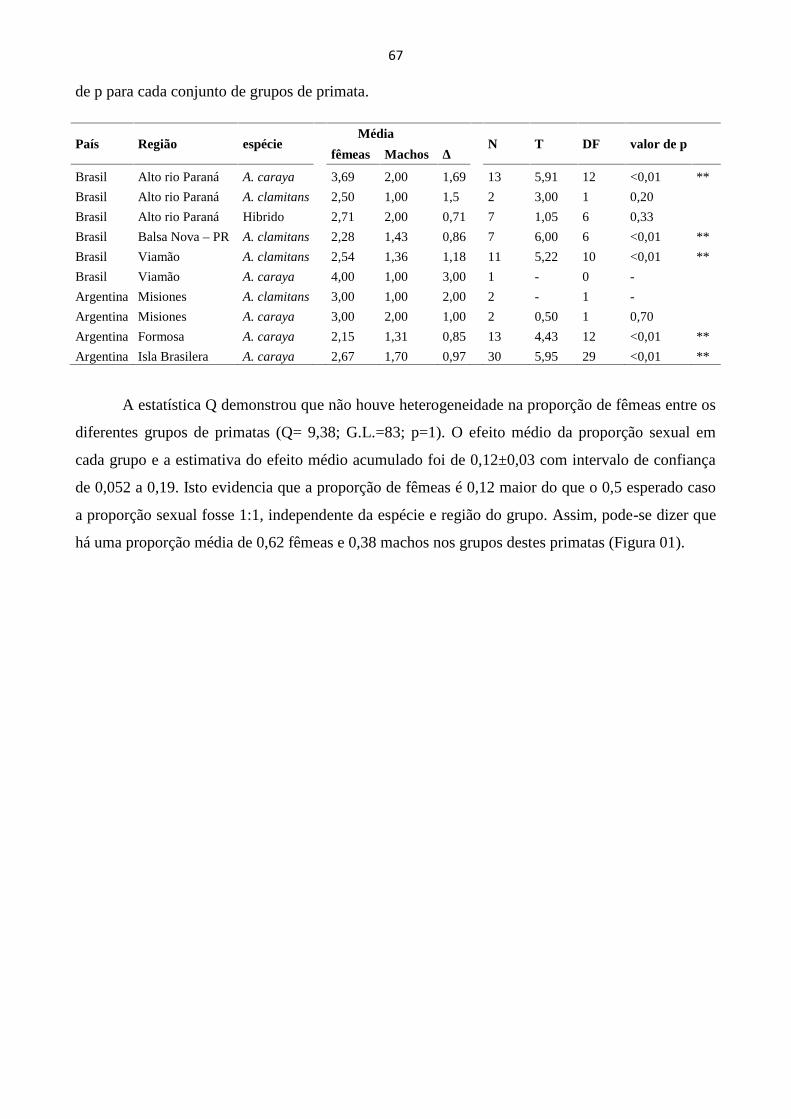
67
de p para cada conjunto de grupos de primata.
País Região espécieMédia
N T DF valor de pfêmeas Machos ∆
Brasil Alto rio Paraná A. caraya 3,69 2,00 1,69 13 5,91 12 <0,01 **
Brasil Alto rio Paraná A. clamitans 2,50 1,00 1,5 2 3,00 1 0,20
Brasil Alto rio Paraná Hibrido 2,71 2,00 0,71 7 1,05 6 0,33
Brasil Balsa Nova – PR A. clamitans 2,28 1,43 0,86 7 6,00 6 <0,01 **
Brasil Viamão A. clamitans 2,54 1,36 1,18 11 5,22 10 <0,01 **
Brasil Viamão A. caraya 4,00 1,00 3,00 1 - 0 -
Argentina Misiones A. clamitans 3,00 1,00 2,00 2 - 1 -
Argentina Misiones A. caraya 3,00 2,00 1,00 2 0,50 1 0,70
Argentina Formosa A. caraya 2,15 1,31 0,85 13 4,43 12 <0,01 **
Argentina Isla Brasilera A. caraya 2,67 1,70 0,97 30 5,95 29 <0,01 **
A estatística Q demonstrou que não houve heterogeneidade na proporção de fêmeas entre os
diferentes grupos de primatas (Q= 9,38; G.L.=83; p=1). O efeito médio da proporção sexual em
cada grupo e a estimativa do efeito médio acumulado foi de 0,12±0,03 com intervalo de confiança
de 0,052 a 0,19. Isto evidencia que a proporção de fêmeas é 0,12 maior do que o 0,5 esperado caso
a proporção sexual fosse 1:1, independente da espécie e região do grupo. Assim, pode-se dizer que
há uma proporção média de 0,62 fêmeas e 0,38 machos nos grupos destes primatas (Figura 01).

68
Figura 1. Efeito médio da diferença da proporção observada para a esperada (0,50; 1:1) para cada grupo de
primatas e o efeito médio acumulado.
Discussão
Os resultados encontrados confirmam a proposta de diferentes autores de que os grupos das
espécies A. caraya e A. clamitans possuem um maior número de fêmeas do que de machos adultos
(Treves 2001, Wang & Milton 2003, Miranda & Passos 2005, Aguiar et al. 2009, Miranda 2009). A
meta-análise realizada permite que esta generalização possa ser aplicada a diferentes locais e a essas
duas espécies e seus híbridos. Tal generalização está em concordância com a proposta de que a
estrutura social das espécies abordadas é baseada na presença de um macho dominante com acesso
a todas as fêmeas em estro e a possível presença de outros machos que raramente conseguem
copular. Embora em estudos pontuais este padrão possa não ser confirmado, a meta-análise
possibilitou a avaliação do efeito médio acumulado, evidenciando esse padrão.
Meta-análises são importantes ferramentas para conectar diferentes trabalhos, permitindo a
realização de generalizações. Diante disso, novas meta-análises que agreguem um maior número de
espécies e preditores como a densidade populacional, gradientes latitudinais e variações ambientais
-1.37 -0,68 0 0,68 1.37

69
podem ser muito úteis para o maior entendimento das relações e padrões encontrados na
primatologia.
Referências
Borenstein M. Hedges, L.V., Higgins JPT, Rothstein HR 2009. Introduction to Meta-Analysis, John
Wiley & Sons, 413 p.
Glander KE 1980 Reproduction and population growth in free-ranging mantled howling monkeys.
American Journal of Physical Anthropology 53:25-36.
Gregorin R 2006 Taxonomia e variação geográfica das espécies do gênero Alouatta Lacépède
(Primates,
Miranda JMD 2009 Comportamentos sociais, relações de dominância e confrontos inter-grupais em
Alouatta caraya (Humboldt, 1812) na Ilha Mutum, Alto Rio Paraná, Brasil. Tese apresentada
no Programa de Pós-Graduação em Zoologia, Universidade Federal do Paraná, Curitiba, PR.
Neville MK, Glander KE, Braza F, Rylands AB 1988 The howling monkeys, genus Alouatta, p.
349-453. In: Mittermeier RA, Rylands AB, Coimbra-Filho A, Fonseca GAB. (Eds.). Ecology
and Behavior of Neotropical Primates V. 2. Washington, DC, World Wildlife Fund, 610p.
Ostro LET, Silver SC, Koontz FW, Horwich RH, Brockett R 2001 Shifts in social structure of
black howler (Alouatta pigra) groups associated with natural and experimental variation in
population density. International Journal of Primatology 22:733-748.
Treves A 2001 Reproductive consequences of variation in the composition of howler monkey
(Alouatta spp.) groups. Behavioral Ecology and Sociobiology 50:61-71.
Wang E, Milton K 2003 Intragroup social relationships of male Alouatta palliata on Barro
Colorado Island, Republic of Panama. International Journal of Primatology 24(6):1227-1243.
Zucker EL, Clarke M 2003 Longitudinal assessment of immatureto-adult ratios in two groups of
Costa Rican Alouatta palliata. Intenational Journal of Primatology 24:87–101.

70
RELAÇÃO DA COBERTURA VEGETAL DE EICCHORNIA AZUREA (KUNTH) COM AS COMUNIDADES
ÍCTICA E FITOPLANCTÔNICA
Érika Maria Neif, Francisco von Hartenthal, Guilherme Landgraf, Lilian Paula Vasconcelos, RafaelA. Baggio, Sabrina Calado
Orientador: Luiz Carlos Gomes
Resumo
Foi testada a hipótese de que a maior porcentagem de cobertura vegetal de Eichhornia azurea
favorece a riqueza e abundância das comunidades fitoplanctônica e íctica. Foram amostrados 12
pontos no Ressaco do Bilé compreendendo diversos graus de cobertura vegetal. Os resultados
refutam a hipótese, visto que as métricas não estiveram correlacionadas às porcentagens de
cobertura vegetal. No entanto, a baixa riqueza e abundância das comunidades devido à cheia
inesperada possivelmente não representa as condições necessárias para validar as conclusões.
Palavras-chave: Macrófitas, Alto Rio Paraná, planície de inundação, ressaco.
Introdução
Em lagos rasos de planícies de inundação diferentes formas biológicas de macrófitas
aquáticas podem constituir grupos mono ou multiespecíficos que contribuem para elevar a
complexidade e a bioarquitetura dos habitats, de maneira que a presença de macrófitas aquáticas
afeta a estrutura das populações, a diversidade e a composição das assembleias de organismos
aquáticos (Thomaz & Cunha 2010).
As macrófitas podem afetar positivamente a comunidade fitoplanctônica ao promover a
heterogeneidade ambiental ou aumentar a disponibilidade de nutrientes (Santos et al. 2013). Por
outro lado, ao competir por recursos (Agostinho et al. 2003), servir de abrigo para predadores
(zooplâncton) ou liberar substâncias alelopáticas (Gross 2003), afetam negativamente a comunidade
fitoplanctônica. O balanço desses efeitos pode ser afetado pela morfologia da macrófita (Santos et
al. 2013).
A presença de macrófitas também afeta a comunidade de peixes, sendo um fator chave na
seleção de habitats - uma vez que podem servir de refúgio contra predadores, sítio de alimentação
ou reprodução, além de afetar as propriedades físicas e químicas do ambiente (Gomes et al. 2012).
Os efeitos das macrófitas sobre a comunidade de peixes dependem das características do sistema
(e.g. disponibilidade de alimento para as presas, aumento da produtividade do sistema devido à
penetração de luz) e das espécies de peixes (e.g. eficiências do forrageamento de predadores e de
refúgio das presas, fase ontogenética, estratégia de vida) (Agostinho et al. 2003).

71
Uma das mais abundantes macrófitas da bacia do Alto Rio Paraná, Eichhornia azurea
(Kunth), provê alta heterogeneidade estrutural devido a suas raízes submersas, oferecendo proteção
a macroinvertebrados contra predação e afetando o uso do habitat por predadores (Padial et al.
2009), além de favorecer as comunidades algais (Rodrigues et al. 2003).
Assim, considerando o papel das macrófitas E. azurea nas comunidades fitoplanctônica e
íctica, este trabalho foi realizado buscando testar a hipótese de que a maior porcentagem de
cobertura vegetal de E. azurea afeta positivamente as comunidades de fitoplâncton e peixes.
Materiais e Métodos
O Ressaco do Bilé (22o45'13.56"S; 53o17' 9.48"W), localizado na planície de inundação do
Alto Rio Paraná, possui conexão direta com o Rio Paraná e apresenta diversos bancos de macrófitas
aquáticas, entre as quais se destacam os de E. azurea.
Os pontos amostrais foram escolhidos ao longo do gradiente longitudinal do ressaco,
compreendendo diversos graus de cobertura vegetal por E. azurea, desde a ausência da vegetação
até a cobertura mais densa encontrada (90%), totalizando 12 amostras. A determinação da cobertura
vegetal foi realizada a partir de inspeção visual da porcentagem de preenchimento de um quadro de
madeira de 50 x 50 cm pelas folhas das macrófitas.
As amostragens da ictiofauna foram realizadas com utilização de um peneirão, constituído
de uma armação metálica de 1,5 x 1,0 m e uma rede de malha de 500 µm. O instrumento foi
empurrado e submergido sob o banco de E. azurea e rapidamente trazido à superfície para a captura
dos peixes. Os peixes foram acondicionados em sacos plásticos, fixados com formol 4% e,
posteriormente, identificados segundo Graça & Pavanelli (2007).
As amostragens do fitoplâncton foram realizadas na sub-superfície com um frasco de vidro
de 150 ml. As amostras foram fixadas com solução de Transeau, e identificadas de forma
qualitativa com auxílio de microscópio óptico e literatura especializada (Bicudo & Menezes 2006).
Os atributos de comunidade utilizados para testar a hipótese foram a riqueza específica e a
abundância (em número de indivíduos) de peixes, e a riqueza de espécies fitoplanctônicas, visto que
a mensuração da abundância foi impossibilitada pela falta de equipamento adequado.
As correlações entre a porcentagem de cobertura vegetal e a abundância de peixes, riqueza
de espécies de peixes e riqueza de espécies fitoplanctônicas foram aplicadas no software Statistica
7, através do método de Pearson.
Resultados

72
No total foram registradas 6 espécies de peixes e 20 espécies de algas, a maioria das quais
pertencentes à comunidade perifítica.
Tanto a correlação de Pearson realizada entre a abundância de peixes (em número total) e a
cobertura vegetal (%), quanto a realizada entre a riqueza de espécies de peixes e a cobertura vegetal
não foram significativas (r = -0,016; P > 0,05; r = -0,007, P > 0,05, respectivamente) (Fig. 21).
0 20 40 60 80 100
Cobertura Vegetal (%)
-2
0
2
4
6
8
10
12
14
16
18
20
Abu
ndân
cia
de p
eixe
s (N
)
A
0 20 40 60 80 100
Cobertura vegetal (%)
0
1
2
3
4
5
Riq
ueza
de
espé
cies
de
peix
es
B
Figura 1. A: Relação entre a abundância de peixes (número de indivíduos) e a cobertura vegetal(%). B: Relação entre a riqueza de espécies de peixes e a cobertura vegetal (%) de E. azurea.
A correlação entre a riqueza de espécies de fitoplâncton e a cobertura vegetal também não foi
significativa (r = -0,064; P > 0,05) (Fig. 2).
0 20 40 60 80 100Cobertura vegetal (%)
0
2
4
6
8
10
12
14
Riq
ueza
de
fito
plân
cton
Figura 2. Relação entre a riqueza de espécies fitoplanctônicas e a cobertura vegetal (%) de E.
azurea.
DiscussãoOs resultados obtidos neste trabalho refutaram a hipótese testada. A cheia inesperada neste
período na planície de inundação do Alto Rio Paraná e, consequentemente no Ressaco do Bilé,
possivelmente iniciou a homogeneização abiótica característica destes períodos (Thomaz et al.
2007), ocasionando também uma grande dispersão dos peixes, principalmente para áreas litorâneas

73
(Gomes et al. 2011), o que possivelmente ocasionou os baixos valores de riqueza e abundância de
peixes na área amostrada.
O aumento do nível hidrométrico do Rio Paraná também pode sido o responsável pelas
baixas riquezas específicas do fitoplâncton e maior número de espécies perifíticas. O evento de
cheia pode ter promovido a diluição e dispersão do fitoplâncton, e o aumento do fluxo da água do
rio em direção à porção final do ressaco potencialmente ocasionou o deslocamento de organismos
aderidos as macrófitas para a coluna d’água, por isso, a maioria das espécies encontradas foi
pertencente à comunidade perifítica. Finalmente, é possível que, na ausência da cheia, a
comunidade de fitoplâncton e peixes seja favorecida pela presença da macrófita que provê abrigo
contra predadores, como sugerido por Barko et al. (1977) e Flöder et al. (2006).
Agradecimentos
Agradecemos aos funcionários do NUPELIA pelo apoio logístico na coleta de dados e aos docentes
do Curso de Campo pelas contribuições no delineamento e análise dos dados.
Referências
Agostinho AA, Gomes LC, Júlio-Jr. HF 2003. Relações entre macrófitas aquáticas e fauna de
peixes, pages 261-279, in Thomaz SM, Bini LM, eds. Ecologia e Manejo de Macrófitas
Aquáticas. EDUEM: Maringá.
Barko J, Murphy P, WETZEL RG 1977 An investigation of primary production and ecosystem
metabolism in a Lake Michigan dune pond. Archiv. für Hydrobiologie 81: 155-187.
Bicudo CEM, Menezes M 2006 Gêneros de Algas de Águas Continentais do Brasil: Chave para
identificação e descrições. RiMa: São Carlos..
Graça WJ, Pavanelli CS 2007 Peixes da planície de inundação do Alto Rio Paraná e áreas
adjacentes. Eduem: Maringá.
Gross EM 2003 Allelopathy of Aquatic Autotrophs. Critical Reviews in Plant Sciences 22: 313-
339.
Gomes LC, Bulla CK, Agostinho AA, Vasconcelos LP, Miranda LE 2012 Fish assemblage
dynamics in a Neotropical floodplain relative to aquatic macrophytes and the homogenizing
effect of a flood pulse. Hydrobiologia, 685: 97-107.
Flöder S, Combüchen A, Pasternack A, Hillebrand H 2006. Competition between pelagic and
benthic microalgae for phosphorus and light. Aquatic Science 68: 425-433.

74
Padial AA, Thomaz SM, Agostinho AA 2009 Effects of structural heterogeneity provided by the
floating macrophyte Eichhornia azurea on the predation efficiency and habitat use of the
small Neotropical fish Moenkhausia sanctaefilomenae. Hydrobiologia 624: 161-170.
Rodrigues L, Bicudo DC, Moschinicarlos V 2003. O papel do perifiton em áreas alagáveis e nos
diagnósticos ambientais, pages 211-230, in:Thomaz SM, Bini LM, eds. Ecologia e manejo de
macrófitas aquáticas. EDUEM: Maringá.
Santos TR, Ferragut C, Bicudo CEM. Does macrophyte architecture influence periphyton?
Relationships among Utricularia foliosa, periphyton assemblage structure and its nutrient (C,
N, P) status. Hydrobiologia 714: 71-83.
Thomaz SM, Cunha ER 2010 The role of macrophytes in habitat structuring in aquatic ecosystems:
methods of measurement, causes and consequences on animal assemblages: composition and
biodiversity. Acta Limnologica Brasiliensia 22: 218-236.

75
COMPLEXIDADE ESTRUTURAL E VOLUME DE RAÍZES DE MACROFITAS AQUÁTICAS COMO
PROMOTORAS DO ENRIQUECIMENTO DA COMUNIDADEDE INVERTEBRADOS AQUÁTICOS
EMANUEL Razzolini; FERNANDO Garcia de Oliveira; FLÁVIO Miranda Marteleto; NATÁLIA
Carneiro Lacerda dos Santos; SANDRA Marisa Nobre de Andrade
Orientador: Prof. Dr. Marcos Barbeitos
O objetivo do trabalho foi verificar a relação entre riqueza de espécies de invertebrados e o volume
e a complexidade estrutural das raízes das macrófitas no rio Paraná. As variáveis medidas foram:
número de indivíduos e riqueza de morfotipos de invertebrados, além do volume e complexidade
das raízes das macrófitas. Com relação à riqueza o volume e complexidade acumulada explicaram
cada um deles separadamente 48% da variação dos invertebrados, enquanto para a abundância, o
volume explicou 13% e a complexidade acumulada 3% da variação.
Palavras-chave: dimensão fractal, riqueza, lagoas, planície de inundação.
Introdução
Comunidades de macrófitas aquáticas são consideradas fundamentais para a composição de
ambientes de água doce, não somente como fonte de carbono, mas funcionalmente como refúgio
para pequenos vertebrados e invertebrados aquáticos (Dibble & Thomaz, 2009). A riqueza
estrutural entre espécies de macrófitas aumenta a complexidade e heterogeneidade dos habitats
afetando vários grupos de organismos, como por exemplo, invertebrados (Thomaz & Cunha, 2010).
Muito embora a complexidade dos habitats seja utilizada para atribuir maior ou menor
riqueza e abundancia aos sistemas biológicos, ainda hoje as formas de estimar esse parâmetro são
controversas (Dibble e Thomaz, 2009). A geometria Fractal surgiu como uma medida eficaz para a
aproximação da complexidade com a forma, sendo assim amplamente utilizada em estudos em larga
escala espacial, tais como estudos com fragmentos florestais (Thomaz & Cunha, 2010)..
Com o objetivo de endereçar esse assunto, o presente estudo aborda o modo como a
complexidade estrutural das raízes afeta a comunidade de invertebrados associados a macrófitas
aquáticas. A hipótese do trabalho é que existe uma relação positiva entre a riqueza de espécies de
invertebrados e a complexidade estrutural e volume das raízes das macrófitas. É predito que quanto
maior a complexidade estrutural e o volume da raiz maior será a riqueza de espécies. Desse modo
temos como objetivo: (i) verificar a relação entre complexidade estrutural das raízes e riqueza de
invertebrados aquáticos; (ii) Verificar a relação entre riqueza de espécies de invertebrados e o
volume das raízes de macrófitas.

76
Material e Métodos
Área de estudo
O rio Paraná é o décimo maior rio em descarga do mundo e nasce no planalto central
brasileiro, passando a receber esse nome a partir da confluência dos rios Paranaíba e Grande. A
planície de inundação do Alto rio Paraná (PIARP) está localizada na margem direita (oeste) do rio
Paraná (23 43’- 25 33’ S e 54 35’-5310’ O) na divisa dos estados do Mato Grosso do Sul e do
Paraná (Oporto, 2008). A área de estudo compreende dois ressacos (Pau Véio e Bilé) e uma lagoa
aberta (Lagoa das Garças), distribuídos na ilha fluvial mutum e margem direita do rio,
respectivamente.
Figura 1: Planície de Inundação do Alto Rio Paraná, com delimitação para os três locais de amostragem:
Ressaco do Pau Véio, Lagoa das Garças e Ressaco do Bilé.
Procedimento
Foram estabelecidos três pontos de amostragens em cada uma das três lagoas. O primeiro
ponto na entrada da lagoa, o segundo na região intermediaria e o terceiro na porção mais interna da
lagoa em relação à conexão com o rio Paraná. Foi utilizado um quadrante de 50 cm2 para a
padronização da área amostrada em cada banco de macrófitas. As plantas foram transportadas para
a o laboratório da base de pesquisas avançadas do Nupélia no município de Porto Rico-Pr. Em
laboratório, as raízes foram separadas lavadas para a retirada de toda a comunidade de
invertebrados. As amostras foram filtradas e posteriormente triadas com o auxilio de
estereomicroscópio e os organismos identificados até o menor nível taxonômico possível. As
variáveis medidas foram: número de indivíduos, volume e complexidade das raízes das macrófitas
aquáticas. O volume das raízes foi medido através do deslocamento de volume equivalente de água,

77
medido em uma proveta com uma quantidade de água conhecida (500ml). Três amostras das raízes
de cada espécie de macrófita foram escaneadas para a análise da dimensão fractal, sendo calculada
através do programa IMAGEJ. Essa análise leva em conta as diversas escalas de aumento em uma
imagem descrevendo como a complexidade de uma estrutura muda em reflexo da mudança da
magnitude da escala de aumento analisada (Sugihara & May, 1990 in Dibble & Nei, 2009). A curva
de rarefação foi calculada através do pacote VEGAN no software R. O método adiciona as espécies
amostradas em cada localidade selecionada aleatoriamente, e somando-se a estas apenas as novas
espécies encontradas na outra localidade pareada. A curva resultante demonstra se o número de
espécies amostradas é uma aproximação razoável da riqueza total. As análises de correlação foram
feitas no Programa STATISTICA 7.0. A variável Complexidade acumulada foi calculada através da
fórmula:
Ca=∑ Vi x Ci.
Onde Ci é o volume da raiz e o Ci é o índice de dimensão fractal (adaptado de Thomaz e
Cunha (2010)).
Resultados
Foram coletados 580 organismos distribuídos em 31 grandes grupos. Nematoda foi o grupo
mais abundante (83 indivíduos), seguido de larvas de quironomídeos (67 indivíduos) e bivalves (55
indivíduos). Com relação à riqueza de invertebrados aquáticos, ambas variáveis analisadas
separadamente (volume e complexidade acumulada) explicaram 48% da variação da riqueza (Figs.
2A e C), enquanto que para a abundância, o volume explicou 13% e a complexidade acumulada 3%
da variação (Figs. 2B e D, respectivamente).
n
i=1
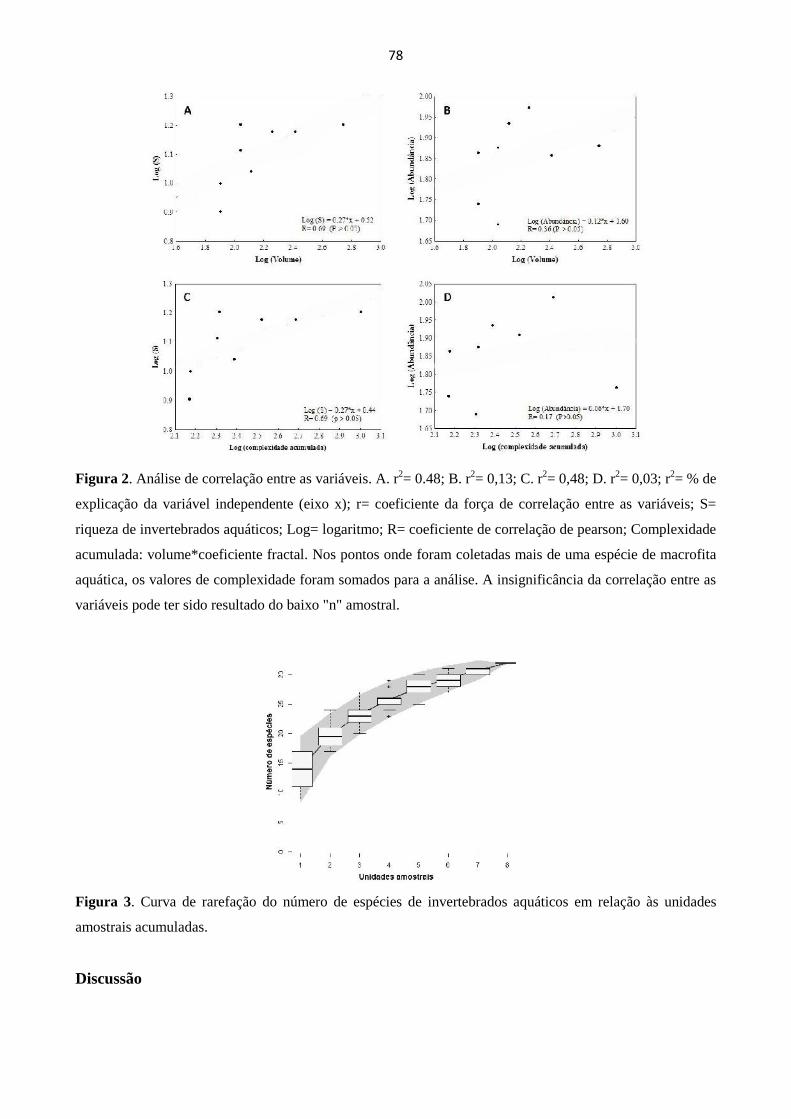
78
Figura 2. Análise de correlação entre as variáveis. A. r2= 0.48; B. r2= 0,13; C. r2= 0,48; D. r2= 0,03; r2= % de
explicação da variável independente (eixo x); r= coeficiente da força de correlação entre as variáveis; S=
riqueza de invertebrados aquáticos; Log= logaritmo; R= coeficiente de correlação de pearson; Complexidade
acumulada: volume*coeficiente fractal. Nos pontos onde foram coletadas mais de uma espécie de macrofita
aquática, os valores de complexidade foram somados para a análise. A insignificância da correlação entre as
variáveis pode ter sido resultado do baixo "n" amostral.
Figura 3. Curva de rarefação do número de espécies de invertebrados aquáticos em relação às unidades
amostrais acumuladas.
Discussão

79
As macrófitas atuam nas comunidades aquáticas promovendo a riqueza das espécies de
animais associadas através da complexidade estrutural dos microhabitats que proporcionam,
(Dibble e Thomaz, 2009). A tendência de correlação positiva observada entre a riqueza de
invertebrados aquáticos e a complexidade das raízes corroboram com o estudo realizado por Dibble
e Thomaz (2009), no qual encontraram uma correlação positiva significativa entre abundância de
invertebrados com a complexidade de estruturas foliares. Em conclusão, admite-se que a ausência
de significância encontrada pode ser associada à baixa amostragem de pontos em cada tratamento.
Entretanto, a ausência de amostragem exaustiva de espécimes menores, residentes nas escalas
intersticiais inferiores das raízes, pode ser o principal fator responsável para que a curva de
rarefação não tenha atingido uma proximidade à assíntota.
Agradecimentos
Os autores agradecem a orientação e ensinamentos do professor Marcos Barbeitos, ao Sebastião,
funcionário do Nupélia, pelo apoio logístico, e aos programas de pós-graduação da Universidade
Federal do Paraná e Universidade Estadual de Maringá.
Referências
Agostinho, A. A., Zalewski, M., 1996. A planície alagável do alto rio Paraná: importância e
preservação. EDUEM, Maringá, PR, BR, 100 pp.
Opoto, L. T. 2008. Modificações a longo prazo na ictiofauna na planície de inundação do Alto rio
Paraná, Brasil. Dissertação de mestrado. Universidade Estadual de Maringá
Dibble, E. D.; Thomaz S.M. (2009) Use of Fractal dimension to assseshabitat complexityand its
influence on dominant invertebrates inhabiting tropical and temperate Macrophytes. Journal of
Freshwater Ecology. 24, 1, 93-104
Thomaz, S. M., T. A. Pagioro, L. M. Bini, M. C. Roberto & R. R. A. Rocha, 2004a. Limnological
characterization of the aquatic environments and the influence of hydrometric levels. In Thomaz,
S. M., A. A. Agostinho & N. S. Hahn (eds) The Upper Paraná River and its floodplain: Physical
aspects, ecology and conservation. Backhuys publishers, Leiden: 75-102.
Thomaz, S. M & Cunha, E. R. 2010. The role of macrophytes in habitats structuring in aquatic
ecosystems: methods of measurements, causes and consequences on animal assemblages’
composition and biodiversity. Acta Limnologica brasiliensia, 22(2): 218-236.

80
MIGRAÇÃO HORIZONTAL DE ZOOPLÂNCTON SOB ESTÍMULO DE PREDAÇÃO: UMA ABORDAGEM
EXPERIMENTAL
Hugo José Message, Priscilla Guedes Gambale, Renata Gabriela Noguchi, Simone Satsuki
Takeuchi, Taise Miranda Lopes & Juliana Dias
Orientadora: Juliana Dias
Resumo: O zooplâncton pode migrar horizontalmente ocasionando variações de sua densidade ao
longo do espaço, sendo a predação um dos maiores estímulos para esse comportamento. Um
experimento foi realizado para testar a hipótese de que o zooplâncton migra da área sem refúgio
(plâncton) para o refúgio (macrófitas aquáticas) na presença de predadores. Não houve diferenças
na densidade média zooplanctônica entre as regiões sem e com refúgio na ausência de predadores.
No entanto, a densidade média de zooplâncton diferiu na presença de predadores, com maiores
densidades na área com refugio. Assim, mostramos que as populações zooplanctônicas buscaram
locais estruturados devido à pressão de predação.
Palavras chave: refúgio, macrófitas aquáticas, plâncton, densidade.
Introdução
A migração horizontal é um comportamento observado nos organismos zooplanctônicos que
faz com que esses organismos apresentem oscilações de suas densidades ao longo do espaço. Vários
fatores ambientais podem promover as migrações horizontais do zooplâncton, como a busca por
alimentos ou estratégias para evitar predadores (White, 1998). Dentre esses fatores, a predação
influencia os padrões de distribuição e abundância desses organismos, através da indução à
mudança de comportamento (Romare & Hansson, 2003). Na presença de um predador a presa pode
ajustar seus padrões de movimentação e de uso de refúgios em resposta à ameaça (Jeffries & Mills,
1994; Romare & Hansson, 2003).
A predação por peixes é um dos fatores mais importantes que influenciam a estrutura e
distribuição espaço-temporal da comunidade de zooplâncton (Brooks & Dodson, 1965). As
estruturas submergidas de macrófitas podem promover refúgio para o zooplâncton (Burks et al.,
2006) e são fontes de alimento para muitas espécies (Henry & Costa, 2003).
O objetivo deste trabalho foi testar experimentalmente a migração horizontal dos
organismos zooplanctônicos. Para isso foi testada a hipótese de que o zooplâncton migra da área
sem refúgio (plâncton) para a área com refúgio (macrófitas) na presença de predadores. A partir

81
dessa hipótese prediz-se que, na ausência de predadores, a densidade de zooplâncton é maior na
área sem refúgio e, que, quando na presença de predadores, a maior densidade é na área com
refúgio.
Material e métodos
Amostragens
O zooplâncton foi coletado com rede de plâncton de 68µm de abertura de malha na
subsuperfície da região limnética do ressaco do Pau Véio. Concomitantemente as amostragens do
zooplâncton foram coletadas macrófitas aquáticas flutuantes da espécie Eichhornia azurea, que
representa abrigo para o zooplâncton na região litorânea.
Os potenciais predadores do zooplâncton (Caracídeos de pequeno porte) foram coletados
com rede de arrasto simples de 10m de comprimento, identificados de acordo com Graça &
Pavanelli (2007) e colocados em um aquário durante 2h para aclimatação.
Delineamento experimental
O experimento foi realizado em três aquários de 200L, representando três réplicas. Em cada
aquário foram delimitadas duas regiões, sendo uma sem refúgio (SR = sem refúgio) e outra com
refúgio (CR =com refúgio - presença de macrófitas aquáticas). A amostra concentrada do
zooplâncton foi adicionada em cada aquário. Após o período de 1 hora foi realizada a primeira
amostragem, afim de que o zooplâncton se estabilize nos aquários. Amostras de 1,5L foram
coletadas nos dois compartimentos de cada aquário e fixadas com solução de formaldeído
tamponado com carbonato de cálcio. Logo em seguida, foram adicionados no aquário 10 peixes
planctívoros isolados em um saco plástico para concentração do estímulo hormonal (5
Bryconamericus stramineus + 3 Serrapinus notomelas + 2 Steindachnerina ‘insculpta’). Após 1h os
peixes foram soltos e 1h depois, o zooplâncton foi novamente amostrado.
O zooplâncton foi quantificado em lupas e microscópios para cálculo e extrapolação da
densidade de indivíduos por litro de água. Para testar a hipótese os dados foram submetidos a um
teste-t (referencia) entre a densidade de zooplâncton na região sem refúgio e com refúgio antes da
presença de predadores e nas mesmas regiões após a presença de predadores.
Resultados
Durante o experimento, a variação da densidade de zooplâncton na ausência de predadores
foi de 43 a 122 indivíduos/L em regiões sem refúgio e de 61 a 112 indivíduos/L em regiões com
refúgio. Após a inserção dos peixes no experimento, foram registradas alterações na densidade de
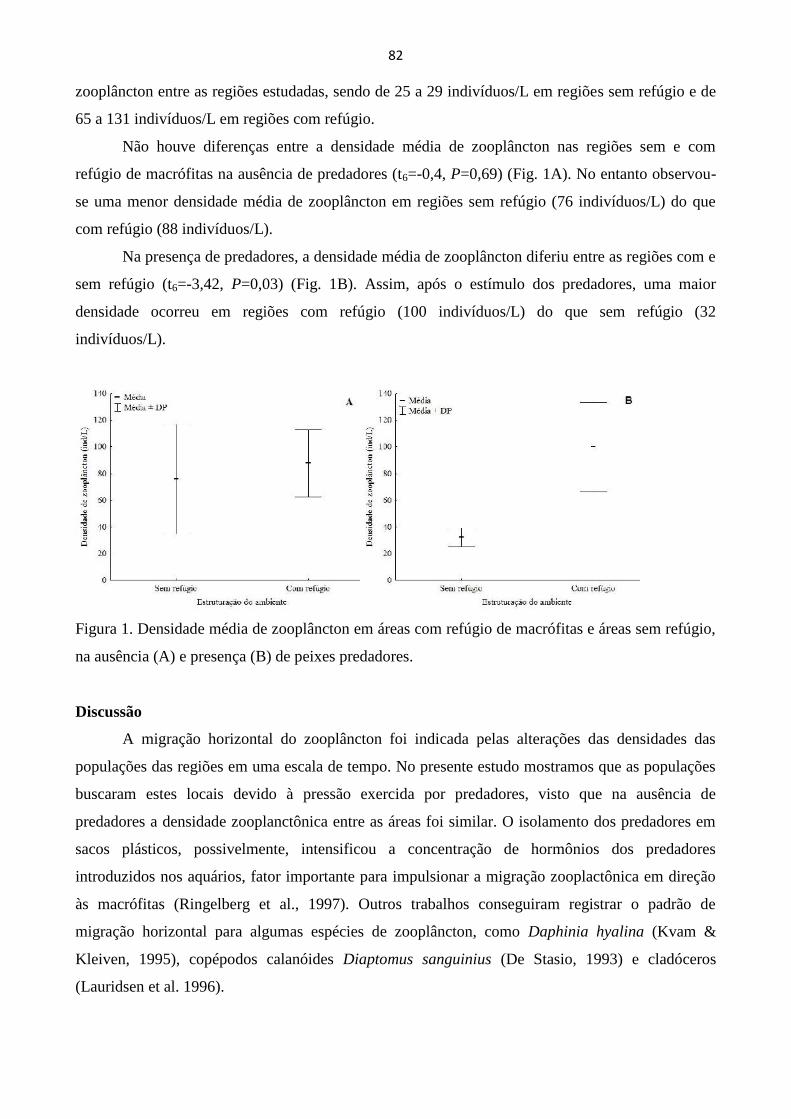
82
zooplâncton entre as regiões estudadas, sendo de 25 a 29 indivíduos/L em regiões sem refúgio e de
65 a 131 indivíduos/L em regiões com refúgio.
Não houve diferenças entre a densidade média de zooplâncton nas regiões sem e com
refúgio de macrófitas na ausência de predadores (t6=-0,4, P=0,69) (Fig. 1A). No entanto observou-
se uma menor densidade média de zooplâncton em regiões sem refúgio (76 indivíduos/L) do que
com refúgio (88 indivíduos/L).
Na presença de predadores, a densidade média de zooplâncton diferiu entre as regiões com e
sem refúgio (t6=-3,42, P=0,03) (Fig. 1B). Assim, após o estímulo dos predadores, uma maior
densidade ocorreu em regiões com refúgio (100 indivíduos/L) do que sem refúgio (32
indivíduos/L).
Figura 1. Densidade média de zooplâncton em áreas com refúgio de macrófitas e áreas sem refúgio,
na ausência (A) e presença (B) de peixes predadores.
Discussão
A migração horizontal do zooplâncton foi indicada pelas alterações das densidades das
populações das regiões em uma escala de tempo. No presente estudo mostramos que as populações
buscaram estes locais devido à pressão exercida por predadores, visto que na ausência de
predadores a densidade zooplanctônica entre as áreas foi similar. O isolamento dos predadores em
sacos plásticos, possivelmente, intensificou a concentração de hormônios dos predadores
introduzidos nos aquários, fator importante para impulsionar a migração zooplactônica em direção
às macrófitas (Ringelberg et al., 1997). Outros trabalhos conseguiram registrar o padrão de
migração horizontal para algumas espécies de zooplâncton, como Daphinia hyalina (Kvam &
Kleiven, 1995), copépodos calanóides Diaptomus sanguinius (De Stasio, 1993) e cladóceros
(Lauridsen et al. 1996).

83
Era esperada maior densidade zooplanctônica na região aberta, devido às condições
propícias para o desenvolvimento do plâncton, antes da introdução do predador. Todavia, foi
registrada uma maior densidade na região com refúgio, apesar da não significância. Esse fato pode
ter sido explicado pela preferência de algumas espécies pela região litorânea, como por exemplo, os
cladóceros. Assim, nossa hipótese foi parcialmente corroborada uma vez que maiores valores de
densidade foram registrados na região de refúgio após a presença do predador, mas na ausência dos
predadores os maiores valores foram registrados nesse mesmo compartimento aquático.
Apesar da existência de poucos estudos realizados na região tropical (Jeppesen et al. 2005),
esse experimento sugere um modelo conceitual para o padrão da migração horizontal de
zooplâncton em diferentes ecossistemas aquáticos. Como vários fatores, tais como mudanças nos
parâmetros físicos e químicos, disponibilidade de recursos alimentares, arquitetura e densidade da
planta, presença de potenciais predadores podem influenciar o efeito de refúgio de plantas aquáticas
(Burks et al. 2006), novas pesquisas utilizando outros refúgios ou outros estímulos para migração
com especificação dos grupos de zooplâncton podem ser realizadas.
Agradecimentos
Agradecemos à professora Juliana Déo Dias pela orientação do projeto, Alfredo pelas ajudas
na coleta. Agradecemos também aos organizadores e professores do Curso de Ecologia de Campo.
Referências bibliográficas
Brooks J L & Dodson S I 1965 Predation, body size and composition of the plankton. Science 150:
28 – 35.
Burks R, Mulderij G, Gross E, Jones I, Jacobsen L, Jeppesen E, van Donk E 2006 Center stage: the
crucial role of macrophytes in regulating trophic interactions in shallow lake wetlands. Pages
37–53 in R Bobbink, B Beltman, JTA Verhoeven, DF Whigham, eds. Functioning,
Biodiversity Conservation, and Restoration, Ecological Studies. Vol. 191, Springer Verlag,
New York, NY.
De Stacio BT 1993 Diel vertical and horizontal migration by zooplankton: population budgets and
the diurnal deficit. Bullein of Marine Science 53: 44 – 64.
Graças W, Pavanelli CS 2007 Peixes da planície de inundação do alto rio Paraná e áreas adjacentes.
Maringá: EDUEM.
Henry R, Costa MLR 2003 As macrófitas como fator de heterogeneidade espacial: um estudo em
três lagoas com diferentes conectividades com o rio Paranapanema. Pages 189 – 210 in SM
Thomas, LM Bini, eds. Ecologia e manejo de macrófitas aquáticas. Maringá: EDUEM.

84
Jeffries M, Mills D 1994 Freshwater ecology: principles and applications. John Wiley & Sons Ltd.
Jeppesen E, Pekcan-Hekim Z, Lauridsen TL 2006 Habitat distribution of fish in late summer:
changes along a nutrient gradient in Danish lakes. Ecology of Freshwater Fish 15: 180–191.
Kvam OV, Kleiven OT 1995 Diel horizontal migration and swarm formation in Daphnia in
response to Chaoborus. Hydrobiologia 307: 177 – 184.
Lauridsen TL, Pedersen L.J., Jeppesen E, Sondergaard M 1996 The importance of macrophyte bed
size for cladoceran composition and horizontal migration in a shallow lake. Journal of
Plankton Research 18:2283-2294.
Ringelberg J, Flik BJG, Aanen D, van Gool E 1997 Amplitude of diel vertical migration (DVM) is
a function of fish biomass, a hypothesis. Archiv für Hydrobiologie–Beiheft Ergebnisse der
Limnologie 49: 71 – 72.
Romare P, Hansson L-A 2003 A behavioral cascade: top-predator induced behavioral shifts in
planktivorous fish and zooplankton. Limnology and Oceanography 48: 1956–1964.
White, MG 1998 Horizontal distribution of pelagic zooplankton in relation to predation gradients.
Ecography 21: 44–62.

85
BIOGEOGRAFIA DE ILHAS: UMA ABORDAGEM UTILIZANDO MACRÓFITAS E
MACROINVERTEBRADOS AQUÁTICOS
Ariane Ghelfi, Bianca Possamai, Daniel Santos, Juliana Wojciechowski e Vivian Cionek
Orientador: Roger Mormul
Resumo
A teoria de Biogeografia de ilhas prediz que a riqueza de espécies em ilhas aumenta com o
tamanho da área amostrada e com a proximidade da fonte de dispersão. Este estudo verificou a relação
espécie-área e relacionou riqueza de macroinvertebrados com distância da fonte (bancos de macrófitas). A
relação espécie-área foi significativa, mas dependeu da diversidade regional e do tamanho amostral. A
distância da fonte de dispersão não foi significativa, possivelmente devido ao grande volume de chuva que
ocorreu nos últimos dias na região.
Palavras-chave: relação espécie-área, área fonte, riqueza de espécies.
Introdução
Uma das principais premissas da teoria de biogeografia de ilhas formulada por MacArthur e Wilson
(1967) é de que o número de espécies presentes em uma ilha é determinado pela sua área e pela
proximidade da fonte de dispersão. Macrófitas aquáticas formam grandes bancos em lagoas e são
conhecidamente locais de abrigo e berçário de espécies (Jeppesen et al., 1997), como por exemplo
macroinvertebrados bentônicos, organismos de grande importância na composição de sedimentos de rios e
lagos (Callisto et al., 1995).
Os objetivos desse trabalho foram (i) investigar se existe relação espécie x área com macrófitas no
ressaco do Pau Véio e (ii) verificar se a riqueza de espécies de macroinvertebrados no sedimento dessa
lagoa é influenciada pela distância da fonte de dispersão (Eichhornia azurea). A primeira hipótese testada
foi de que o número de espécies de macrófitas aumenta com a área amostrada e a segunda hipótese
testada foi de que a riqueza de espécies de macroinvertebrados bentônicos diminui com o aumento da
distância da fonte (banco de macrófitas).
Material e métodos
As amostragens foram realizadas no Ressaco do Pau Véio (22°44’76”S; 53°15’11.16”W),
pertencente à sub-bacia do Rio Paraná. Para analisar a relação espécie-área foram utilizados dois métodos
de amostragem. O primeiro método consistiu em amostrar a riqueza de macrófitas em três escalas

86
aninhadas: pequena, média e grande, com quadrados de 15x15, 50x50 e 100x100 cm, respectivamente, em
três bancos de macrófitas situados na entrada, no meio e no fundo da lagoa. No segundo método a riqueza
foi amostrada nos mesmos bancos a partir de uma transecção utilizando oito quadrados médios (50x50
cm), assim a área amostrada foi sendo aumentada gradativamente em 50 cm a partir de 50X50 até a
transecção atingir 400X50.
Para testar a segunda hipótese foram coletadas amostras de sedimento com Van Veen em locais
próximos e distantes de três bancos de macrófitas (fontes) localizados na entrada, no meio e no fundo do
ressaco. A comunidade de macroinvertebrados dos bancos de macrófitas também foi amostrada (controle)
com auxílio de peneira. Em laboratório, as amostras de sedimentos foram lavadas e os macroinvertebrados
foram identificados em microscópio estereoscópico.
Para verificar a relação espécie-área foram realizadas ANCOVAs e regressões lineares para ambos
os métodos, com alfa = 0,05. A influência da distância foi testada através de ANOVA, com alfa = 0,05. As
análises foram realizadas no R e STATISTICA.
Resultados
Ambos os métodos de amostragem da relação espécie-área foram significativos (Tab. 1). A
regressão linear para o método de transecção foi realizada diferenciando-se os bancos em relação à
diversidade de espécies de cada local, mostrando que em uma baixa diversidade a relação não é observada
(R²=0,40; F1,6=5,78; p>0,05), mas tanto em alta (R²=0,84; F1,6=39,71; p<0,001) como em média diversidade
(R²=0,83; F1,6=36,98; p<0,001) a relação foi significativa (Fig. 1A). Para o aninhamento, todas as escalas de
tamanho foram significativas (Tab. 1, Figura 1B).
Tabela 1. Análise de Covariância da relação espécie-área de macrófitas no ressaco do Pau Véio em dois diferentes métodos
amostrais. β1, β2 e β3 indicam as escalas grande, médio e pequena em aninhado e alta, média e baixa diversidade em transecção.
ANCOVA Aninhado Transecção
Área F=36,46; p<0,01 F=26,09 p<0,01
Escala F = 6,51; p<0,01 F=10,62 p<0,01
Área*Escala F=32,41; p<0,01 F=36,63 p<0,01
β1 0,85 0,53
β2 0,51 0,40
β3 0,22 0,00

87
Figura 1. Regressão linear entre a riqueza de espécies e os tratamentos. (A) método de transecção separado por
diversidade dos bancos: alta y = 0,96+0,53x e média y = 0,17+0,4x; (B) método de aninhamento de escalas.
Com relação à distância dos invertebrados do banco de macrófitas foi encontrada uma diferença
significativa entre a riqueza (F2,9= 30.73; p<0,001) e abundância (F2,9=26.05; p<0,01) de espécies, porém
essa se deu entre o controle e os tratamentos (borda e distante), e não entre os tratamentos (Fig. 2).
Figura 2. Médias de riqueza (esquerda) e abundância (direita) de macroinvertebrados nas diferentes distâncias em relação à
fonte (banco de macrófitas).
Discussão
A riqueza de espécies de macrófitas esteve relacionada com o aumento da área, corroborando
nossa hipótese. Porém, foi possível notar que a relação espécie/área pode ser afetada pelo pool regional de
espécies e pela escala (tamanho da amostra), padrão já relatado em outros estudos (Fattorini e Borges,
2012). Cabe ressaltar que há um viés de área/esforço amostral no caso do uso da escala maior. Conclui-se,
portanto, que o melhor método para analisar a relação espécie-área seria uma abordagem independente
do local, abrangendo a diversidade γ e em escala espacial maior, pois ambas variáveis podem afeta
fortemente a relação.
87
Figura 1. Regressão linear entre a riqueza de espécies e os tratamentos. (A) método de transecção separado por
diversidade dos bancos: alta y = 0,96+0,53x e média y = 0,17+0,4x; (B) método de aninhamento de escalas.
Com relação à distância dos invertebrados do banco de macrófitas foi encontrada uma diferença
significativa entre a riqueza (F2,9= 30.73; p<0,001) e abundância (F2,9=26.05; p<0,01) de espécies, porém
essa se deu entre o controle e os tratamentos (borda e distante), e não entre os tratamentos (Fig. 2).
Figura 2. Médias de riqueza (esquerda) e abundância (direita) de macroinvertebrados nas diferentes distâncias em relação à
fonte (banco de macrófitas).
Discussão
A riqueza de espécies de macrófitas esteve relacionada com o aumento da área, corroborando
nossa hipótese. Porém, foi possível notar que a relação espécie/área pode ser afetada pelo pool regional de
espécies e pela escala (tamanho da amostra), padrão já relatado em outros estudos (Fattorini e Borges,
2012). Cabe ressaltar que há um viés de área/esforço amostral no caso do uso da escala maior. Conclui-se,
portanto, que o melhor método para analisar a relação espécie-área seria uma abordagem independente
do local, abrangendo a diversidade γ e em escala espacial maior, pois ambas variáveis podem afeta
fortemente a relação.
87
Figura 1. Regressão linear entre a riqueza de espécies e os tratamentos. (A) método de transecção separado por
diversidade dos bancos: alta y = 0,96+0,53x e média y = 0,17+0,4x; (B) método de aninhamento de escalas.
Com relação à distância dos invertebrados do banco de macrófitas foi encontrada uma diferença
significativa entre a riqueza (F2,9= 30.73; p<0,001) e abundância (F2,9=26.05; p<0,01) de espécies, porém
essa se deu entre o controle e os tratamentos (borda e distante), e não entre os tratamentos (Fig. 2).
Figura 2. Médias de riqueza (esquerda) e abundância (direita) de macroinvertebrados nas diferentes distâncias em relação à
fonte (banco de macrófitas).
Discussão
A riqueza de espécies de macrófitas esteve relacionada com o aumento da área, corroborando
nossa hipótese. Porém, foi possível notar que a relação espécie/área pode ser afetada pelo pool regional de
espécies e pela escala (tamanho da amostra), padrão já relatado em outros estudos (Fattorini e Borges,
2012). Cabe ressaltar que há um viés de área/esforço amostral no caso do uso da escala maior. Conclui-se,
portanto, que o melhor método para analisar a relação espécie-área seria uma abordagem independente
do local, abrangendo a diversidade γ e em escala espacial maior, pois ambas variáveis podem afeta
fortemente a relação.

88
As macrófitas não responderam como uma fonte de dispersão de macroinvertebrados, rejeitando a
segunda hipótese, porém isso deve ter ocorrido em decorrência das fortes chuvas que lavaram e
resuspenderam o sedimento, gerando a baixa riqueza e abundância encontradas. Em conseqüência disso,
o pool de espécies nas macrófitas foi maior que no sedimento e a identidade das espécies não foi
correspondente entre os locais.
Referências Bibliográficas
Callisto M, Esteves FA 1995 Distribuição da comunidade de macroinvertebrados bentônicos em um lago
amazônico impactado por rejeito de bauxita, Lago Batata (Pará, Brasil). Oecologia Brasiliensis. v. 1.
Estrutura, Funcionamento e Manejo de Ecossistemas Brasileiros. F. A. Esteves (ed.), pp. 281-291,
Programa de Pós- graduação em Ecologia, Instituto de Biologia, Universidade Federal do Rio de
Janeiro, Rio de Janeiro.
Jeppesen E, Søndergaard M, Christofferssen K 1997 The structuring role of submerged macrophytes in
lakes. Ecological studies series, 131 Springerr-Verlag, New York.
MacArthur RH, Wilson EO 1967 The theory of island biogeography. Princepton University Press, Princepton.
Périco E, Cemin G, Lima DFB, Rempel C 2005 Efeitos da fragmentação de hábitats sobre comunidades
animais: utilização de sistemas de informação geográfica e de métricas de paisagem para seleção
de áreas adequadas a testes. Anais XII Simpósio Brasileiro de Sensoriamento Remoto, Goiânia,
Brasil, 16-21 abril 2005, INPE, p. 2339-2346.

89
EFEITO DA DISTÂNCIA DA MARGEM NA HERBIVORIA EM EICHHORNIA AZUREA
Diego Corrêa Alves, Fernanda Ceschin, Letícia Cucolo Karling, Luciana Zago, Monica Vasques
Mangas Pereira
Orientador: André Padial
Resumo
O objetivo deste estudo foi verificar o efeito da distância da fonte de insetos terrestres na
herbivoria da macrófita Eichhornia azurea. A proporção de folhas consumidas diminuiu e houve
uma diminuição da porcentagem de área consumida em regiões mais distantes da margem de uma
lagoa de inundação. Além disso, ocorreu uma redução na diversidade de níveis de herbivoria entre
folhas de uma mesma distância da margem, com maior dominância do menor nível de consumo em
locais distantes da margem.
Palavras-chave: Macrófitas aquáticas; Planície de inundação; rio Paraná; rio Baía.
Introdução
As macrófitas aquáticas desempenham importantes papéis ecológicos, servindo, por
exemplo, como local de abrigo, desova e substrato para outros organismos (Thomaz & Cunha
2010). Por um longo período, a herbivoria de macrófitas vivas foi considerada funcionalmente sem
importância em teias alimentares aquáticas (Hutchinson 1975). No entanto, estudos posteriores
demonstraram que a biomassa, produtividade e composição de espécies de macrófitas são
frequentemente influenciadas por uma variedade de herbívoros vertebrados e invertebrados (Lodge
1991, Newman 1991).
Embora existam espécies de invertebrados que se alimentem exclusivamente de macrófitas
aquáticas, como o Orthoptera Cornops aquaticum (Ferreira 1995, Capello et al. 2012), espécies que
consomem altas porcentagens de plantas terrestres também podem consumir plantas aquáticas
(Capello et al. 2012). Nesse sentido, a herbivoria deve ter relação com a distância da margem dos
corpos aquáticos. Sob esse contexto, o presente trabalho visa testar as hipóteses: i) a proporção de
folhas consumidas diminui com a distância da margem de uma lagoa de inundação; ii) a área foliar
consumida diminui com a distância da margem e iii) a variação na área foliar predada aumenta com
a distância da margem. Tendo-se como predição de que maiores áreas foliares e maiores números
de folhas serão consumidas em macrófitas que estejam a distâncias menores da margem do rio, já
que nesses ambientes o número de herbívoros é maior.
Material e métodos

90
As coletas foram realizadas no mês de junho de 2013, na lagoa Fechada do afluente rio Baía
da planície de inundação do alto rio Paraná. A coleta de dados foi realizada em três bancos de
macrófitas de Eichhornia azurea, seguindo uma transecção da margem ao meio da lagoa. A
transecção foi divida em pontos equidistantes de 0,5m, nos quais foram acondicionados quadrados
de 625 cm². As folhas de E. azurea de cada quadrado foram coletadas, armazenadas em sacos
plásticos e transportadas para o laboratório para posteriores análises.
As folhas foram classificadas com relação ao grau de herbivoria, estabelecendo ranks de 1 a
4 (do menos para o mais consumido). Como pressuposto das hipóteses, foi realizada uma análise de
covariância (ANCOVA) para verificar se havia diferença nos padrões de relação da herbivoria com
a distância entre os três bancos de macrófitas amostrados. A variável resposta da ANCOVA foi a
proporção de folhas consumidas pelas folhas não consumidas e a variável independente foi a
distância da margem e o banco. Não houve efeito do banco de macrófitas (p = 0.37; Efeito da
interação banco*distância: p = 0,95), portanto as análises foram realizadas utilizando os valores dos
três bancos. A primeira hipótese foi testada com a correlação de Pearson entre a proporção de folhas
consumidas (%) e a distância da margem. A correlação de Spearman foi utilizada para testar a
segunda hipótese, utilizando as variáveis: mediana dos níveis de consumo pela distância da
margem. A variação na área foliar predada foi sumarizada em duas variáveis: riqueza e
equitabilidade de ranks, e correlacionada com a distância da margem através de uma correlação de
Spearman.
Resultados
A análise de Pearson indicou que a proporção de folhas consumidas diminuiu com o
aumento da distância da margem (r =-0.67, p<0.01)(Fig. 1. esquerda). A correlação de Spearman,
(ρ=-0.75, p<0.01), verificou a diminuição da porcentagem de área consumida em regiões mais
distantes da margem (Fig. 1. Direita). As correlações entre equitabilidade e distância da margem
(ρ=-0.60, p<0.01) e entre riqueza de ranks e distância da margem (ρ=-0.62, p<0.01) indicaram uma
redução no número de ranks e a dominância do rank 1 com o aumento da distância da margem (Fig.
2).

91
Figura 1. Esquerda: Relação da proporção de folhas consumidas (%) com a distância da margem.
Direita: Relação da mediana dos ranks de folhas consumidas com a distância da margem. Os
valores das correlações de Pearson (r) e Spearman (ρ) estão mostrados junto com suas
significâncias nos gráficos.
Figura 2. Esquerda: Relação da riqueza de ranks de folhas consumidas com a distância da margem.
Direita: Relação da equitabilidade dos ranks de folhas consumidas com a distância da margem. Os
valores das correlações Spearman (ρ) estão mostrados junto com suas significâncias nos gráficos.
Discussão
Os resultados encontrados para E. azurea corroboram com a proposta de que a herbivoria
em macrófitas aquáticas é maior nas proximidades das margens das lagoas. Invertebrados terrestres
podem alimentar-se intensamente de macrófitas aquáticas e os efeitos combinados do consumo por
coleópteros adultos e larvas de dípteros podem causar um grande dano nas populações Neotropicais
destas macrófitas (Thomaz et al. 2008). Diante disso, embora existam consumidores que se
alimentam exclusivamente de macrófitas aquáticas e têm todo seu ciclo de vida sobre elas (Ferreira
1995, Capello et al. 2012), nas margens dos rios podem estar presentes tanto estas espécies quanto
-2 0 2 4 6 8 10
Distância da margem (m)
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
1.1Fo
lhas
con
sum
idas
(%)
r = -0.67; p <0.01-2 0 2 4 6 8 10
Distância da margem (m)
0.81.01.21.41.61.82.02.22.42.62.83.03.2
Med
iana
dos
rank
s
ρ = -0.758; p < 0.01
-2 0 2 4 6 8 10
Distância da margem (m)
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
Riq
ueza
dos
rank
s
ρ = -0.558; p < 0.01-2 0 2 4 6 8 10
Distância da margem (m)
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Equi
tabi
lidad
e do
sra
nks
ρ = -0.605; p < 0.01

92
espécies terrestres de consumidores. Portanto, as maiores intensidades de herbivoria encontradas
nas margens dos rios podem dever-se a sobreposição de consumidores terrestres e aquáticos. Porém,
também há a possibilidade de as folhas distantes da margem serem mais novas e com maiores
conteúdos de compostos fenólicos. Se existirem, esses dois aspectos também podem contribuir para
uma diminuição da herbivoria com a distãncia da margem de lagoas de inundação. Dessa forma, as
razões para os padrões espaciais na hervivoria de plantas ainda deve ser elucidado.
As menores variações nos graus de herbivoria encontradas mais próximas das margens
também podem ser resultantes do maior número de espécies consumidoras nestes locais, visto que
um maior número de espécies pode resultar em uma maior equitabilidade nas formas e graus de
consumo. Além de fatores abióticos, interações bióticas podem ser importantes modeladores da
estrutura de comunidades de macrófitas tropicais e a herbivoria pode afetar suas assembleias
(Thomaz et al. 2008). Partindo-se desse conhecimento, os resultados aqui encontrados somados a
futuros estudos que abordem as interações bióticas existentes em diferentes espécies de macrófitas
aquáticas podem ser ferramentas fundamentais para a compreensão de estruturas e dinâmicas de
comunidades.
Referências
Capello S, Marchese M, Wysiecki ML 2012 Feeding Habitats and Trophic Niche Overlap of
Aquatic Orthoptera Associated with Macrophytes. Zoological Studies 51(1):51-58.
Ferreira SA 1995 Plantas Hospedeiras de Cornops aquaticum (Bruner) (Orthoptera: Acrididae) no
Pantanal de Poconé – MT. Dissertação de Mestrado do Programa de Pós-Graduação em
Ecologia da Universidade Estadual de Campinas, Campinas, SP.
Hutchinson GE 1975 A Treatise on Limnology. Vol. 3. Limnological Botany. Wiley-Inter-
Science, 660 pp, Nova York.
Lodge DM 1991 Herbivory on freshwater macrophytes. Aquatic Botany 41:195-224.
Newman RM 1991 Herbivory and detritivory on freshwater macrophytes by invertebrates: a review.
Journal of the North American Benthological Society 10(2):89-114.
Thomaz SM, Cunha ER 2010 The role of macrophytes in habitat structuring in aquatic ecosystems:
methods of measurement, causes and consequences on animal assemblages' composition and
biodiversity. Acta Limnologica Brasiliensia 22(2):218-236.
Thomaz SM, Esteves FA, Murphy KJ, Santos AM, Caliman A, Guariento RD 2008 Aquatic
Macrophytes in the Tropics: Ecology of Populations and Communities, Impacts of Invasions
and Use by Man. In: Del Claro K, Oliveira PS, Rico-Gray V, Barbosa AAA, Bonet A,

93
Scarano FR, Garzon FJM, Villarnovo GC, Coelho L, Sampaio MV, Quesada M, Morris MR,
Ramirez N. International Commision on Tropical Biology and Natural Resources. Eolss
Publishers, Oxford.

94
ASSOCIAÇÃO SIMBIÓTICA ENTRE A TRIPLARIS AMERICANA E FORMIGAS PSEUDOMYRMEX: DA
OCORRÊNCIA À RESPOSTA A ESTÍMULOS
Érika Maria Neif, Francisco von Hartenthal, Guilherme Landgraf, Lilian Paula Vasconcelos, Rafael
A. Baggio, Sabrina Calado
Orientador: Fernando C. Passos
Resumo
A associação mutualística entre Triplaris americana e Pseudomyrmex é altamente conhecida
pela agressividade dos insetos. Neste trabalho foi investigado se existe diferença na ocorrência de
Pseudomyrmex em Triplaris americana de diferentes tamanhos e o tempo de resposta das formigas
a um estímulo físico na planta. Árvores menores puderam ou não ter ocorrência das formigas, já em
plantas maiores estiveram sempre presentes. O tempo médio de resposta das formigas ao estímulo
físico foi de 32 segundos, com alto coeficiente de variação.
Introdução
Uma das associações mais interessantes entre animais e plantas é a simbiose entre vários
gêneros de árvores e formigas Pseudomyrmex (mirmecofilia) (Junior et al., 2009) e este tipo de
interação biológica é a mais diversificada nas florestas tropicais (Price,1997).
Nesse tipo de relação, as formigas podem proteger as plantas de herbívoros (Bruna et al.,
2008), facilitar o acesso das plantas a nutrientes, dispersar sementes e frutos e atuar na polinização.
As plantas, por sua vez, podem fornecer locais para abrigo e nidificação e até mesmo recursos
alimentares para as formigas (Beattie, 1985, Mello, 2012).
Triplaris americana L. (pau de novato ou pau-formiga) está entre as espécies arbóreas mais
comuns na Floresta Estacional Semidecidual localizada nos fragmentos próximos às margens do rio
Paraná, sobre as ilhas e dispersos em áreas mais secas na planície de inundação do alto rio Paraná
(Gimenes et al., 2007). Pertencente à família Polygonaceae, trata-se de uma espécie heliófita, de
crescimento rápido (Lorenzi, 2000) e que apresenta uma relação simbiótica bastante característica
com as formigas Pseudomyrmex. Vivendo nessas árvores, as Pseudomyrmex se alimentam da
substância produzida na domácia (Blatix et al., 2009), estrutura oca dentro do caule que comumente
abrigam grupo de indivíduos, e, por serem insetos que defendem ativamente seu ninho, atacam
organismos que dele se aproximam (Junior et al., 2009), proporcionando proteção às árvores.

95
A biologia das Pseudomyrmex é ideal para a defesa das Triplaris americana, uma vez que
apresentam mecanismos sensoriais aguçados para detectar distúrbios e estímulos, respondendo
rapidamente a eles (Agrawal & Dubin-Thaler, 1999).
Jaffé et al. (1986) afirmam que Pseudomyrmex nunca ocupam indivíduos pequenos de “paus
de novato”. No entanto, através de comunicação informal, profissionais da área afirmam que já
observaram ocorrência das formigas neste tipo de indivíduos.
Assim, este trabalho foi desenvolvido buscando responder duas perguntas: i) Existe
diferença na ocorrência de Pseudomyrmex em Triplaris americana de diferentes tamanhos? ii) Qual
é o tempo de resposta das formigas Pseudomyrmex a um estímulo físico produzido na Triplaris
americana?
Materiais e Métodos
O estudo foi realizado nas margens do córrego Caracu, na área da base de pesquisa do
Nupélia.
A fim de responder a primeira pergunta proposta foram amostrados 43 indivíduos de T.
americana, distribuídos em diversos tamanhos (altura). A altura das plantas foi determinada
visualmente quando ultrapassavam o tamanho mensurável por fita métrica. Em todos os indivíduos
foi registrada a ocorrência das formigas Pseudomyrmex (presença ou ausência) a partir de um
estímulo físico.
Para analisar o tempo de resposta das formigas ao estímulo físico, 12 árvores de diferentes
tamanhos foram selecionadas. Os estímulos, únicos e de mesma intensidade, foram realizados com
um facão.
Para verificar se a amostragem foi representativa para todas as classes de altura de T.
americana foi gerado um histograma com seis classes de altura. Para determinar a amplitude de
cada classe foi realizado o seguinte cálculo:
Para testar a hipótese de que existe diferença na ocorrência de Pseudomyrmex em indivíduos
de Triplaris americana de diferentes alturas foi utilizado o modelo logit, utilizando como variável
resposta a ocorrência ou não da formiga e variável preditora a altura das árvores.
Finalmente, para avaliar o tempo médio de resposta ao estímulo foi calculada uma média
simples do tempo de resposta da formiga.
Resultados
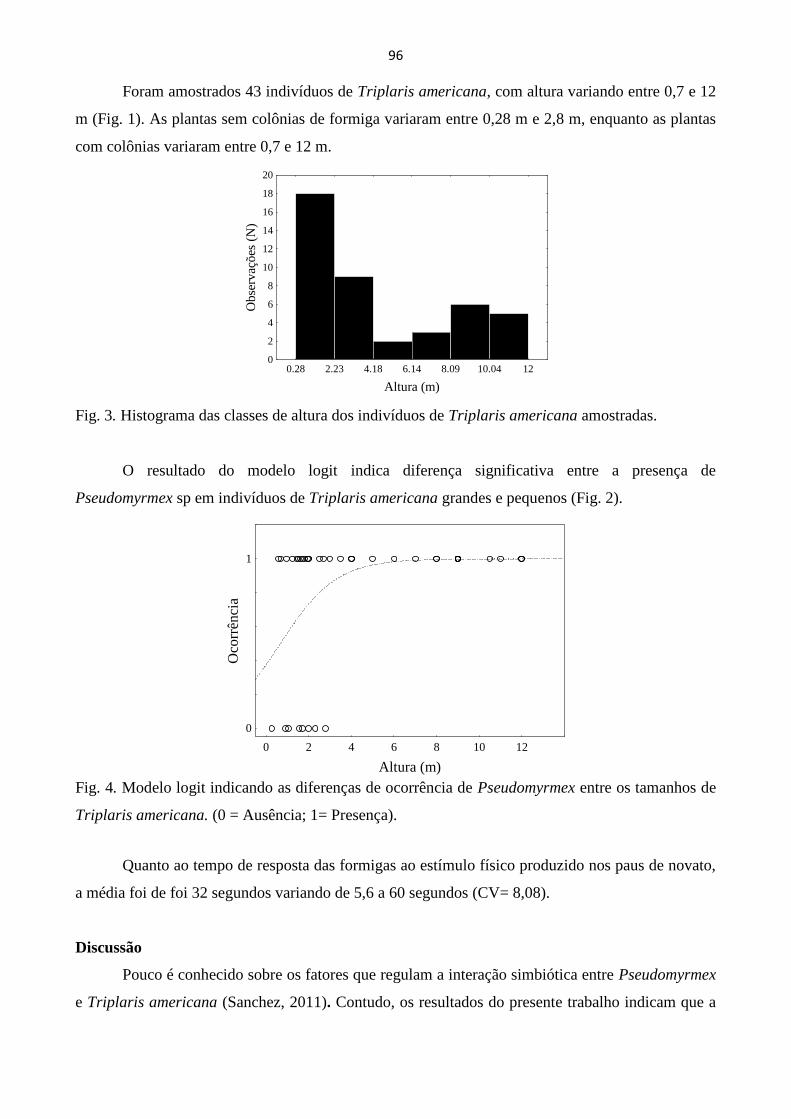
96
Foram amostrados 43 indivíduos de Triplaris americana, com altura variando entre 0,7 e 12
m (Fig. 1). As plantas sem colônias de formiga variaram entre 0,28 m e 2,8 m, enquanto as plantas
com colônias variaram entre 0,7 e 12 m.
0.28 2.23 4.18 6.14 8.09 10.04 12
Altura (m)
0
2
4
6
8
10
12
14
16
18
20
Obs
erva
ções
(N
)
Fig. 3. Histograma das classes de altura dos indivíduos de Triplaris americana amostradas.
O resultado do modelo logit indica diferença significativa entre a presença de
Pseudomyrmex sp em indivíduos de Triplaris americana grandes e pequenos (Fig. 2).
0 2 4 6 8 10 12
Altura (m)
0
1
Oco
rrên
cia
Fig. 4. Modelo logit indicando as diferenças de ocorrência de Pseudomyrmex entre os tamanhos de
Triplaris americana. (0 = Ausência; 1= Presença).
Quanto ao tempo de resposta das formigas ao estímulo físico produzido nos paus de novato,
a média foi de foi 32 segundos variando de 5,6 a 60 segundos (CV= 8,08).
Discussão
Pouco é conhecido sobre os fatores que regulam a interação simbiótica entre Pseudomyrmex
e Triplaris americana (Sanchez, 2011). Contudo, os resultados do presente trabalho indicam que a

97
colonização de Pseudomyrmex em plantas de Triplaris americana ocorre quando a planta atinge
entre 0,7 e 2,8 m de altura. Em plantas maiores do que esses valores, a relação simbiótica sempre
foi observada. Nesse contexto, o tamanho da domária pode ser um fator importante para explicar tal
resultado.
Provavelmente, plantas pequenas de T. americana possuem domária em tamanho
insuficiente para abrigar uma colônia de Pseudomyrmex. A partir de determinado tamanho (i.e.
entre 0,7 e 2,8 m), contudo, a domária se torna grande o suficiente para a colonização, o que tem
grande probabilidade de ocorrer até quando a planta atinge 2,80 m. Os fatores que fazem com que
ocorra essa variação no tempo de colonização devem ser melhores estudados, mas a aleatoriedade
parece ser um fator importante.
Oliveira (1987), através de estudo experimental, mostra que as operárias forrageiam
individualmente e que o maior número de operárias de Pseudomyrmex responde no primeiro minuto
após o estímulo, nossas observações corroboram tal resultado. Contudo, a alta variabilidade nos
valores indica que pode haver diferença de agressividade entre as colônias.
Por fim, o teste das hipóteses levantadas pelo trabalho é essencial para o melhor
entendimento das relações entre Pseudomyrmex e Triplaria americana.
Referências
Agrawal AA, Dubin-Thaler BJ. 1999. Induced responses to herbivory in the Neotropical ant-plant
association between Azteca ants and Cecropia trees: response of ants to potential inducing cues.
Behavior Ecological Sociobiological 45:47–54.
Beattie AJ. 1985. The evolutionary ecology of antplant mutualism. Cambridge University Press,
Cambridge.
Blatix R, Bauamer S, Morand S, Selosse M 2009. Ant-plant mutualisms should be viewed as
symbiotic communities. Plant Signaling & Behavior, 4: 554-556.
Bruna EM, Darrigo MR, Midori AP, Vasconcelos HL 2008. Interspecific variation in the defensive
responses of obligate plant-ants: experimental tests and consequences for herbivory. Oecologia
138: 558-565.
Gimenes MR, Lopes EV, Loures-Ribeiro A, Mendonça LB, Anjos L 2007 Aves da planície
alagável do alto rio Paraná. Maringá, EDUEM.
Jaffé K, Lopez ME, Aragort W 1986 On the communication systems of the ants pseudomyrmex
termitarius and P. triplarinus. Insectes Sociaux Paris 33: 105-117.

98
Junior VH, Bicudo LRH, Fransozo A. 2009. The Triplaria tree (Triplaris spp) and Pseudomyrmex
ants: a symbiotic relationship with risks of attack for humans. Revista da Sociedade Brasileira
de Medicina Tropical 42:727-729.
Lorenzi H 2000 Árvores Brasileiras: manual de identificação de plantas nativas do Brasil. 3 ed.
Nova Odessa: Plantarum.
Price PW 1997. Insect Ecology. John Wiley & Sons: Chinchester.
Oliveira PS, Oliveira-Filho AT, Cintra R 1987 Ant foraging on ant-inhabited Triplaris
(Polygonaceae) in western Brazil: a field experiment using live termite-baits. Journal of
Tropical Ecology 3:193-200.
Sanchez A 2011 Evolutionary relationships in Polygonaceae with emphasis on Triplaris. Phd Tese.
Wake Forest University, Winston-Salen, NC.

99
INUNDAÇÕES COMO REGULADORAS DA DIVERSIDADE ZOOPLANCTÔNICA EM
PLANÍCIE DE INUNDAÇÃO
EMANUEL Razzolini; FERNANDO G. de Oliveira; FLÁVIO M. Marteleto; NATÁLIA C. L. dos
Santos; SANDRA M. N. de Andrade
Orientadora: Prof. Dra. Juliana
Dias
Resumo: O pulso de inundação e a conectividade hidrológica estão entre os fatores mais
importantes para a estruturação da diversidade aquática em planícies de inundação. O objetivo deste
trabalho foi determinar a diversidade de espécies em lagoas abertas e fechadas na planície de
inundação do Alto rio Paraná, pressupondo que lagoas abertas apresentam comunidades mais
homogêneas e lagoas fechadas, mais heterogeneas. As lagoas conectadas apresentaram
comunidades mais homogêneas, apesar dessa diferença não ser significativa, isso se deve a
inundação como promotora de uma maior dispersão das espécies zooplânctonicas pela planície de
inundação.
Palavras-chave: similaridade, conectividade, dispersão, comunidade zooplanctônica, lagoas.
Introdução
A comunidade zooplanctônica em ambientes de planície de inundação apresenta uma
elevada diversidade de espécies e abundância de diferentes grupos (protozoários testáceos,
rotíferos, cladóceros e copépodes), devido a sua heterogeneidade espacial e variabilidade temporal,
governados pela hidrodinâmica ambiental, resultante do pulso de inundação (Lansac-Tôha et al.,
2008; Simões et al., no prelo). Padrões de diversidade de espécies podem ser explicados por vários
mecanismos casuais, incluindo um grupo complexo de interações bióticas e abióticas que definem a
variação das espécies, propondo que espécies defiram ecologicamente e sua variação espacial seja
consequência da resposta das mesmas aos gradientes ambientais (Simões et al, no prelo).
O grau de conectividade juntamente com o pulso de inundação, está entre os fatores mais
importantes para a estruturação da diversidade aquática em planícies de inundação (Santos &
Thomaz, 2002). Assim, neste trabalho nos propusemos avaliar a diversidade zooplanctônica em seis
ambientes da planície de inundação do Alto rio do Paraná com diferentes graus de conexão com o
rio. A hipótese formulada para este estudo é de lagoas abertas apresentam comunidades mais
homogêneas e as lagoas fechadas, comunidades mais heterogêneas..

100
Material e Métodos
Área de estudo
Situada na margem direita do rio Paraná, a Planície de inundação do Alto rio Paraná (22o00’
- 23o00’S; 53o00’ - 53o45’ O; Fig. 1) é caracterizada por elevada diversidade de habitats, como
amplas várzeas, ilhas, lagoas com diferentes graus de conectividade e canais (Thomaz et al., 2004),
exercendo, assim, um papel fundamental na manutenção da diversidade biológica aquática regional
(Agostinho et al., 2001). No presente estudo foram amostradas três lagoas conectadas ao rio e três,
isoladas.
Figura 1: Localização dos pontos de amostragem (círculo preto) na Planície de Inundação do Alto Rio Paraná -. L. das
Garças; 2. L. Pousada das garças; 3. L. do Pau Veio; 4. L. Clara; 5. L. do Bilé; 6. L. do Osmar.
Amostragem e análise de dados
As amostras zooplanctônicas foram coletadas através de arrastos verticais em cada ambiente,
com auxílio de uma rede de plâncton com 68µm de abertura de malha. Dentro de cada lagoa foram
amostrados diferentes pontos e foi obtida uma amostra composta, a fim de se obter uma maior
representatividade das espécies zooplanctônicas presentes na lagoa. O material coletado foi
acondicionado em frascos de polietileno, devidamente etiquetados e fixados em formaldeído
tamponado com carbonato de cálcio.
A composição zooplanctônica foi avaliada utilizando microscópio óptico, lâminas
Sedgwick-Rafter e pipetas de 3ml. Os cladóceros e rotíferos foram identificados a nível de espécie e
os copépodes, de ordem. Para o efeito da análise, considerando os objetivos do trabalho, adotou-se
o método de presença e ausência das espécies nos seis ambientes estudados e os mesmos foram
agrupados em ambientes abertos e fechados. O teste t foi empregue para verificar a diferença entre

101
os ambientes abertos e fechados em termos de diversidade α. A diversidade β foi avaliada por meio
de uma análise multivariada de dispersão (Permidisp, Anderson et al., 2006).
Resultados
Foram identificadas 27 espécies de zooplâncton, das quais 4 espécies de copépodes, 14 de
cladóceros e 9 rotíferos. Verificou-se uma maior diversidade α em lagoas abertas quando
comparadas as lagoas fechadas, apesar desta diferença não ter sido estatisticamente significativa
(t4= 1,60; P˃ 0.05) (Fig.2).
Figura 2: Diversidade α de espécies zooplanctônicas nas lagoas abertas e lagoas fechadas (L. aberta= lagoa aberta; L.
fechada= lagoa fechada).
As lagoas fechadas apresentaram maior diversidade β que as lagoas abertas, embora essa
diferença não tenha sido significativa (pseudo F=0,83; P˃ 0.05) (Fig.3). Este resultado sugere que
lagoas fechadas tendem apresentar comunidades mais heterogêneas que as lagoas abertas.
L. aberta L. fechada0
2
4
6
8
10
12
14
Dive
rsid
ade
α de
zoo
plân
cton
MédiaMédia ± EP
t4 = 1,60; P > 0,05
MédiaMédia ± EP
101
os ambientes abertos e fechados em termos de diversidade α. A diversidade β foi avaliada por meio
de uma análise multivariada de dispersão (Permidisp, Anderson et al., 2006).
Resultados
Foram identificadas 27 espécies de zooplâncton, das quais 4 espécies de copépodes, 14 de
cladóceros e 9 rotíferos. Verificou-se uma maior diversidade α em lagoas abertas quando
comparadas as lagoas fechadas, apesar desta diferença não ter sido estatisticamente significativa
(t4= 1,60; P˃ 0.05) (Fig.2).
Figura 2: Diversidade α de espécies zooplanctônicas nas lagoas abertas e lagoas fechadas (L. aberta= lagoa aberta; L.
fechada= lagoa fechada).
As lagoas fechadas apresentaram maior diversidade β que as lagoas abertas, embora essa
diferença não tenha sido significativa (pseudo F=0,83; P˃ 0.05) (Fig.3). Este resultado sugere que
lagoas fechadas tendem apresentar comunidades mais heterogêneas que as lagoas abertas.
L. aberta L. fechada0
2
4
6
8
10
12
14
Dive
rsid
ade
α de
zoo
plân
cton
MédiaMédia ± EP
t4 = 1,60; P > 0,05
MédiaMédia ± EP
101
os ambientes abertos e fechados em termos de diversidade α. A diversidade β foi avaliada por meio
de uma análise multivariada de dispersão (Permidisp, Anderson et al., 2006).
Resultados
Foram identificadas 27 espécies de zooplâncton, das quais 4 espécies de copépodes, 14 de
cladóceros e 9 rotíferos. Verificou-se uma maior diversidade α em lagoas abertas quando
comparadas as lagoas fechadas, apesar desta diferença não ter sido estatisticamente significativa
(t4= 1,60; P˃ 0.05) (Fig.2).
Figura 2: Diversidade α de espécies zooplanctônicas nas lagoas abertas e lagoas fechadas (L. aberta= lagoa aberta; L.
fechada= lagoa fechada).
As lagoas fechadas apresentaram maior diversidade β que as lagoas abertas, embora essa
diferença não tenha sido significativa (pseudo F=0,83; P˃ 0.05) (Fig.3). Este resultado sugere que
lagoas fechadas tendem apresentar comunidades mais heterogêneas que as lagoas abertas.
L. aberta L. fechada0
2
4
6
8
10
12
14
Dive
rsid
ade
α de
zoo
plân
cton
MédiaMédia ± EP
t4 = 1,60; P > 0,05
MédiaMédia ± EP

102
Figura 3: Diversidade β das lagoas abertas e lagoas fechadas ilustrando que lagoas abertas (representadas por
triângulos) apresentam menor variação de diversidade β quando comparadas as lagoas abertas (representadas por
circulos). O eixo um corresponde à PCoA 1, enquanto que o eixo dois corresponde ao componente PCoA 2.
Discussão
Estudos indicam que a presença de conexões hidrológicas entre lagoas e outros corpos de
água podem prover um corredor para dispersão de organismos, como o zooplâncton. Nesse sentido,
o aumento da similaridade entre múltiplos conjuntos de manchas de habitat pode ser
particularmente evidente quando a dispersão é unidirecional (fluxo do rio), como mostrado em um
modelo onde o número total de espécies em um conjunto de manchas de habitats decresce quando a
dispersão inter-manchas for direcional (Akasaka & Takamura, 2012). Assim, o número total de
espécies em conjuntos de manchas de habitats interconectados (lagoas abertas) pode ser similar ou,
possivelmente, menor que o número total de espécies em manchas sem conectividade (lagoas
fechadas) devido à menor variação na composição das espécies. Porém, isso não foi encontrado no
presente estudo. A maior diversidade α encontrada nas lagoas abertas, possivelmente, está
relacionada a entrada de novas espécies trazidas pelo rio, ou ainda devido a mistura provocada pelo
aumento do nível de água do rio, carreando espécies de regiões litorâneas para a região pelágica. Já
em relação à diversidade β não foi observada diferença significativa, porém houve uma tendência
de maior homogeneidade da comunidade nas lagoas conectadas como pressuposto, Esse resultado
corrobora com a diversidade α, no sentido da importância da dispersão de espécies em função do
aumento do nível de água mediado pela inundação. Esse resultado talvez fosse mais nítido se um
maior número de lagoas conectadas e isoladas tivessem sido amostrados, aumentando assim a
variabilidade dos dados.
Agradecimentos
Os autores agradecem a prof. Dra. Juliana Dias pela orientação dada ao longo do projeto e
ao funcionário do Nupelia pelo apoio logistico.
Referências Bibliográficas
Agostinho, A.A., Gomes, L.C., Zalewski, M., 2001. The importance of floodplains for the dynamics
of fish communities of the upper river Paraná. Ecohydrol. Hydrobiol. 1, 209-217
Akasaka, M. & Takamura, N. 2012. Hydrologic connection between ponds positively affects
macrophyte a and c diversity but negatively affects b diversity. Ecology, 93(5) 967–973

103
Anderson, M.J. 2006. Distance-Based Tests for Homogeneity of Multivariate Dispersions.
Biometrics. 62: 245-253.
Lansac-Tôha, F., Bonecker, C., Velho, L., Takahashi, E., Joko, C., Alves, G., Silva, N., Fulone, L.,
Leal, C.M., Fernandes, A., Costa, D., Santos, R., Bastos, V., Soares, P., Braghin, L., Nedlei,
J., Lospes Filho, D. 2008. Relatório Anual/ PELD – A Planície Alagável do Alto Rio Paraná.
Thomaz, S.M., Pagioro, T.A., Bini, L.M., Roberto, M.C., Rocha, R.R.A. 2004. Limnological
Characterization of the aquatic environments and the influence of the hydrometric levels.In:
Thomaz, S.M., Agostinho, A.A., Hahn, N.S. (eds.), The upper Paraná River and its floodplain.
Backhuys Publishers, Leiden, The Netherlands, 75-102pp
Santos, A. & Thomaz, S.M. 2002. O papel da conectividade na estruturação da assembléia de
macrófitas aquáticas. Relatório Anual/ PELD – A Planície Alagável do Alto Rio Paraná.
Simões, N.R., Dias, J.D., Leal, C.M., Braghin, L.S.M, Lansac-Toha, F.A., Bonecker, C.A. no prelo.
Floods control the influence of environmental gradients on the diversity of zooplankton
communities in a neotropical floodplain. 23pp

104
FORRAGEIO ÓTIMO: DETERMINAÇÃO DO TAMANHO DA CARGA EM FORMIGAS-CORTADEIRAS
(ATTA SP)
Hugo José Message, Priscilla Guedes Gambale, Renata Gabriela Noguchi, Simone Satsuki
Takeuchi & Taise Miranda Lopes
Orientador: Marcos Barbeitos
Resumo: As formigas-cortadeiras são caracterizadas pela coleta de folhas, carregando-as por trilhas
que podem chegar a vários quilômetros. Os padrões de forrageamento ótimo das formigas são
complexos e existem poucos estudos em relação ao tamanho da carga e a estrutura corpórea das
formigas. O estudo comprovou as seguintes hipóteses: i) formigas maiores transportam folhas
maiores (R2 = 0,40, p<0,001), ii) todas as integrantes que saem em busca de folhas retornam à
colônia com as mesmas (apenas um número estatisticamente insignificante de formigas contrariou
esta regra) e iii) há um fluxo equivalente de formigas em ambas as direções (entre o formigueiro e a
planta).
Palavras chave: Colônia, Fluxo, Produtividade.
Introdução
As colônias maduras de formigas-cortadeiras (que incluem as saúvas e as quenquéns) são
capazes de chegarem a milhões de formigas e podem ser encontradas facilmente (Rockwood, 1976).
Esses importantes herbívoros são capazes de percorrer vários quilômetros por dia carregando folhas
(Weber, 1966). O estudo do forrageio desses insetos é relativamente simples, desde que eles se
locomovem ao longo de trilhas bem definidas no chão, sendo possível seguir o caminho para uma
determinada planta sendo atacada e consequentemente, informações do tipo de vegetação cortada, a
taxa de corte e a posição da planta em relação ao ninho podem ser adquiridas (Rockwood, 1976).
Por outro lado, as formigas-cortadeiras apresentam padrões de forrageio complexos que podem
mudar constantemente.
Em um cenário ideal, formigas maiores carregariam folhas maiores e todos os indivíduos
que saem do formigueiro para coletar a planta retornariam carregados com folha. É possível, no
entanto, que nem todas as formigas que acessam a planta tragam folhas ao retornarem. Também não
se sabe se as formigas que coletam folhas maiores apresentam maior tamanho corpóreo na intenção
de aproveitar o uso do seu porte para a realização do trabalho. As formigas que carregam folhas
grandes podem ter velocidade média menor em comparação às formigas que carregam folhas

105
pequenas ou às formigas que não transportam nada. Isso geraria uma diferença entre a quantidade
de indivíduos que transitam no rastro nas direções a favor e contra o formigueiro. Em uma
organização ideal, a quantidade de formigas que trafegam em direção à planta é a mesma que as que
retornam ao formigueiro.
O trabalho objetiva verificar os processos de coleta e transporte de folhas por formigas em
termos de produtividade. Deste modo, testaram-se as hipóteses que (1) formigas de maior porte
transportam folhas maiores, (2) todas que saem do formigueiro para coleta de folhas junto às demais
formigas retornam à colônia trazendo folha e (3) o fluxo de formigas em ambas as direções no
rastro entre o formigueiro e a planta é igual.
Material e Métodos
A coleta de dados foi realizada em um pequeno fragmento de mata próximo a base de
estudos da Universidade Estadual de Maringá, localizada na cidade de Porto Rico-PR. Neste
fragmento foi selecionado uma trilha de formigas saúva (Atta sp), identificando o formigueiro
principal, o ponto da primeira ramificação e a árvore de onde elas retiravam as folhas. A trilha foi
mensurada do formigueiro principal até a primeira ramificação.
Para verificar a relação entre o tamanho médio da formiga e a folha carregada, próximo da
primeira ramificação da trilha foram capturadas as formigas e as folhas carregadas com auxílio de
pinça. Foram realizados registros fotográficos dos dois elementos com auxílio de uma régua para
referência de escala. As folhas e as formigas foram fixadas com álcool 95% e analisadas em
laboratório.
Para verificar a densidade de formigas em uma determinada área foram selecionados cinco
pontos aleatórios (distâncias de 1m, 3m, 4m, 8m e 9m), sendo que em cada ponto foi tirada uma
foto a cada dois minutos, com um total de três fotos por ponto. Os pontos foram filmados durante
um minuto em uma escala de 15 cm.
81 folhas foram pesadas com balança de precisão e sua área e maior comprimento foram
calculadas com auxílio do programa Image J. O peso total das folhas também foi medido e dividido
pela área foliar total para calcular a densidade (g/cm²). Através das fotos foram calculados o
comprimento das formigas com auxílio do programa image J, além de quantificar a densidade de
formigas (ind/m²).
Foi computada uma regressão da área das folhas em relação ao comprimento. A equação da
reta de regressão resultante foi utilizada para estimar o peso das folhas amostradas das formigas.
Uma regressão entre o peso estimado e o comprimento das formigas foi efetuada para testar a
correlação entre estas duas variáveis. Para avaliar a diferença na velocidade de transporte das
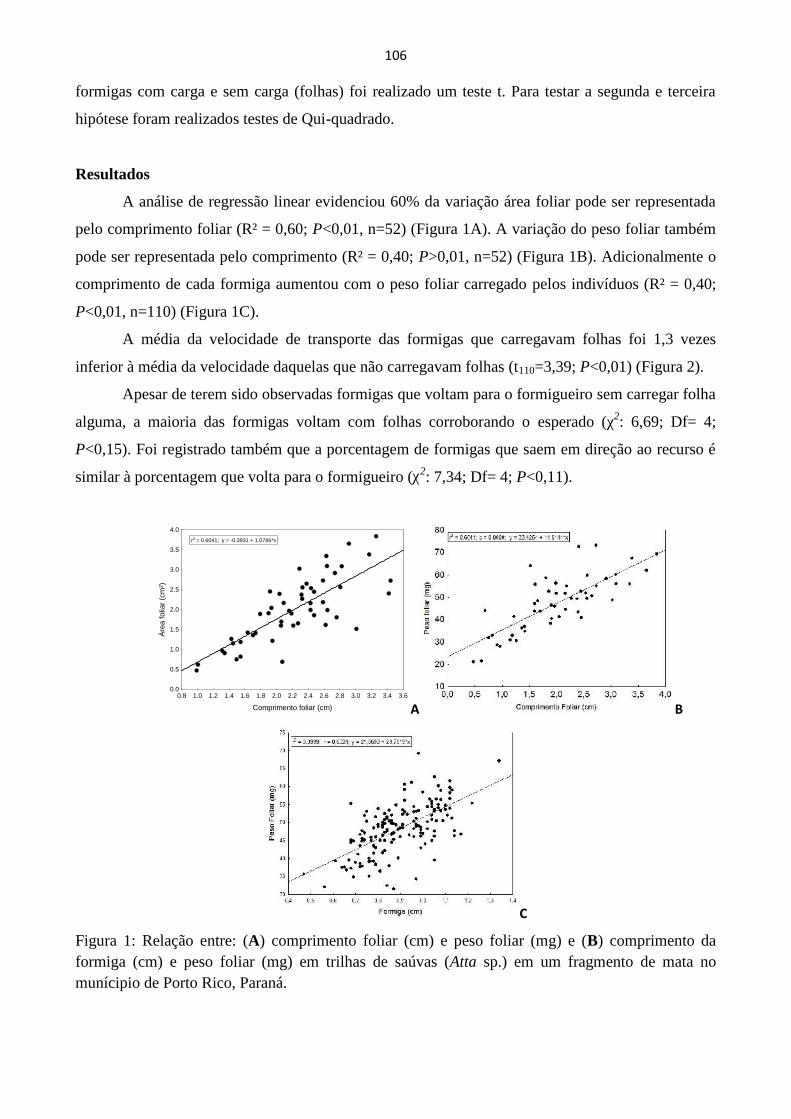
106
formigas com carga e sem carga (folhas) foi realizado um teste t. Para testar a segunda e terceira
hipótese foram realizados testes de Qui-quadrado.
Resultados
A análise de regressão linear evidenciou 60% da variação área foliar pode ser representada
pelo comprimento foliar (R² = 0,60; P<0,01, n=52) (Figura 1A). A variação do peso foliar também
pode ser representada pelo comprimento (R² = 0,40; P>0,01, n=52) (Figura 1B). Adicionalmente o
comprimento de cada formiga aumentou com o peso foliar carregado pelos indivíduos (R² = 0,40;
P<0,01, n=110) (Figura 1C).
A média da velocidade de transporte das formigas que carregavam folhas foi 1,3 vezes
inferior à média da velocidade daquelas que não carregavam folhas (t110=3,39; P<0,01) (Figura 2).
Apesar de terem sido observadas formigas que voltam para o formigueiro sem carregar folha
alguma, a maioria das formigas voltam com folhas corroborando o esperado (χ2: 6,69; Df= 4;
P<0,15). Foi registrado também que a porcentagem de formigas que saem em direção ao recurso é
similar à porcentagem que volta para o formigueiro (χ2: 7,34; Df= 4; P<0,11).
0.8 1.0 1.2 1.4 1.6 1.8 2.0 2.2 2.4 2.6 2.8 3.0 3.2 3.4 3.6
Comprimento foliar (cm)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Área
folia
r (cm
²)
r2 = 0.6041; y = -0.3931 + 1.0786*x
A B
C
Figura 1: Relação entre: (A) comprimento foliar (cm) e peso foliar (mg) e (B) comprimento daformiga (cm) e peso foliar (mg) em trilhas de saúvas (Atta sp.) em um fragmento de mata nomunícipio de Porto Rico, Paraná.

107
Figura 2: Média e desvio padrão da velocidade de formigas que carregavam folhas e formigas quenão carregavam em um fragmento de mata no município de Porto Rico, Paraná.
Discussão
A relação linear positiva entre o comprimento das formigas e o peso foliar da carga
transportada indica que a estrutura corporal destes indivíduos é um importante fator na delimitação
do tamanho das folhas a serem carregadas por elas, corroborando com Weber (1972). Apesar disso
a correlação de 40% demonstra que outros fatores podem estar associados na determinação da
carga. Um dos fatores nessa determinação pode ser a preferência de folhas de menor peso por
formigas de maior porte a fim de otimizar a velocidade de retorno ao formigueiro. Neste estudo
verificamos apenas que a velocidade de formigas sem carga é maior do que aquelas que transportam
algum tipo de carga, sugerindo assim essa provável relação.
Em relação ao deslocamento das formigas ao longo da trilha, apenas alguns indivíduos
foram observados percorrendo o caminho de volta sem nenhuma carga. Não se sabe ao certo a
explicação para este comportamento, mas acredita-se que esteja associado a algum acidente de
percurso, como por exemplo, a perda da folha ao longo do trecho percorrido. Adicionalmente,
existe uma igualdade na densidade de indivíduos que vão em direção ao recurso e as que retornam
ao formigueiro. Estas constatações demonstram a eficácia do trabalho das formigas cortadeiras
quanto ao transporte do recurso, evidenciando um complexo nível organizacional da colônia.
Apesar da velocidade das formigas sem carga ser maior, a porcentagem de indivíduos se
deslocando para o recurso e retornando para o formigueiro foi similar, indicando um possível
paradoxo no estudo. Uma suposição é a de que as formigas regulam esse fluxo, demorando mais
dentro do formigueiro antes de voltar para a árvore. Outra possibilidade é que o pequeno esforço
amostral devido às limitações de tempo podem ter levado a este possível paradoxo.
Agradecimentos

108
Os autores agradecem ao Prof. Dr. Marcos Barbeitos pela orientação e aos professores da
Ecologia de Campo da Universidade Federal do Paraná pela oportunidade.
Referências bibliográficas
Rockwood LL 1976 Plant selection and foraging patterns in two species of leaf-cutting ants (Atta).
Ecology 57: 48 – 61.
Weber NA 1966 Fungus-growing ants. Science 153: 587 – 604.
Weber NA 1972 Garderning ants: the attines American Philosophic Society, Philadelphia.

109
CONSPICUIDADE DE CORES: A VISUALIZAÇÃO NO PONTO DE VISTA DE
MAMÍFEROS PRONATOPIOS.
Ariane Ghelfi, Bianca Possamai, Daniel Santos, Juliana Wojciechowski e Vivian Cionek
Orientador: Marcos Babeitos
Resumo
A luz que incide e é refletida nos diversos ambientes afeta as relações visuais entre os
organismos. Os animais possuem diferentes capacidades de enxergar as cores. A maioria dos mamíferos
não enxerga na faixa do vermelho enquanto as aves enxergam toda a faixa do visível. Neste estudo,
investigamos a diferença do contraste entre cores aposemáticas e o ambiente sob o ponto de vista de um
pronatópio. Verificamos que aqueles com dicromacia não conseguem perceber presas com coloração
aposemática.
Palavras-chave: Aposematismo, comunicação visual, espectro da luz.
Introdução
Sinais visuais são um produto do espectro de refletância e da luz incidente, altamente
dependentes do ambiente. A luz que ocorre nos diversos ambientes do planeta pode ser
caracterizada em dois diferentes tipos: a direta oriunda do sol e as difusas que chegam através de
outras fontes como o céu, nuvens e folhas (Anderson, 1994).
A variação da composição espectral na faixa do visível afeta a visibilidade de animais, flores
e frutos por vertebrados e invertebrados, o que influencia diretamente as interações de predação,
uma vez que a qualidade da luz pode camuflar ou destacar a presença da presa no ambiente (Endler,
1997). Pode-se então concluir que coloração crípitica e aposemática são relativas, uma vez que a luz
varia no espaço e no tempo (Théry, 2010).
O aposematismo é uma característica adaptativa de alguns animais não-palatáveis, tóxicos
ou venenosos que indica impalatabilidade através de coloração de alerta (aposemática). Entre os
animais que possuem essa aparência estão muitas rãs venenosas, salamandras e cobras (Ricklefs,
2010).
A maioria das espécies de mamíferos apresenta dicromacia pronotopica, ou seja, possuem
dois tipos de células sensíveis à cor (cones), não enxergando na faixa do vermelho, diferindo de
macacos e humanos, que possuem tricromacia (três cones), cuja evolução é relativamente recente.
Outros vertebrados, como répteis e pássaros, a complexidade visual é maior, com grande
dominância de cones (Cunha et al, 2006).
O objetivo foi de investigar o contraste de cores entre presas e ambiente simulando a

110
visualização das presas por mamíferos com dicromacia e testar a hipótese de que há diferença entre
as colorações aposemáticas (coloração de advertência) do ponto de vista de predadores com
dicromacia e tricomacia. Espera-se que mamíferos que não enxergam na faixa do vermelho não
consigam perceber presas com coloração aposemática.
Material e Métodos
O trabalho foi realizado na Mata do Araldo, Porto Rico, Paraná. Em campo, foi traçada uma
transecção de 20m paralela ao rio Paraná, com três pontos aleatoriamente definidos. A cada ponto foram
selecionadas as duas árvores mais próximas com diâmetro superior a 12 cm e nelas fixadas discos de massa
de modelar nas cores vermelho, amarelo, azul, preto e branco. A disposição dos discos no tronco das
árvores foi padronizada com um grid e fotografias foram tomadas a 1,2 m do solo e 1 m de distância do
tronco com iluminação branca (flash) a fim de padronizar a luminosidade.
As imagens foram tratadas no software Photoshop®. Um filtro do tipo “ruído” foi aplicado a cada
imagem. Este filtro substitui a coloração de todos os pixels dentro de um raio de 10 pixels pela coloração
correspondente às medianas computadas para três canais de cores (vermelho, verde e azul - RGB). Este
filtro tem o efeito de uniformizar a coloração, impedindo que valores extremos em pixels individuais
introduzam outliers na análise. Em seguida duas transecções (uma horizontal e uma vertical) foram
sobrepostas a cada disco, e os valores dos três canais de cores de cinco pixels do substrato (árvore), cinco
no disco e mais cinco do substrato foram medidos em ambas as direções. Esse processamento foi realizado
para a imagem natural e para a mesma imagem retirando-se a cor vermelha (alterando-se o canal R para -
30% de acordo com a escala da função “Misturador de canais” do Photoshop) a fim de simular a visão de
um mamífero pronatopio (Fig. 1B e 1C).
A média dos valores RGB do fundo de cada árvore ao redor de cada disco foi calculados para dois
tratamentos (com e sem vermelho). Essas médias definem o centro de massa (ou centróide) da coloração
de fundo. A disparidade da cor de cada pixel amostrado do fundo e do disco, e este centróide, foi
computada como distância euclidiana. Os valores destas distâncias foram representados em um gráfico,
contra os respectivos pontos, orientados da esquerda para direita e de cima para baixo, gerando dois picos
correspondentes aos pixels amostrados dos discos. (Fig.1A). O contraste foi estimado como a área sob a
curva pelo método de aproximação por trapezóides. O contraste logaritmizado (devido à homogeneidade
de variâncias) foi utilizado em uma ANOVA bi-fatorial (cor do disco versus tipo de visão) e as diferenças
pareadas entre tratamentos avaliadas através do teste post-hoc de Tukey. As análises estatísticas foram
realizadas no software Statistica® com α=0,05.

111
Figura 1. (A) Contraste entre os pontos RGB de uma imagem e a média dos pontos da árvore (fundo). Linha tracejadademonstra o ponto onde separa a transecção horizontal da vertical.; (B) imagem utilizada para contagem dos pontosem imagem normal (C) imagem utilizada após a subtração do canal vermelho. Notar a mudança de coloraçãoacentuada do disco vermelho.
Resultados
Os tratamentos (com e sem vermelho) diferiram significativamente (F1,50=19,688; p<0,0001), assim
como o contraste entre cores (F4,50=15,153; p< 0,0001). Porém, a interação dos tratamentos com as cores
não foi significativa (F4,100=1,68 p=0,1690).
Com relação às cores, para o tratamento com vermelho as cores azul e branco foram as que mais
contrastaram com o fundo, diferindo das outras (Tukey p<0,05) (Fig.2). Já com a retirada do vermelho
houve uma formação de grupos de cores mais atrativas (amarelo, azul e branco) e outro com cores de
baixo contraste com o fundo e, portanto, crípticas (vermelho e preto) (Tukey p<0,05; Fig.2).
Figura 2. Médias dos logaritmos dos contrastes entre as cores e o fundo das árvores na Mata do Araldo. Em (A)tratamento com vermelho e em (B) sem vermelho. (a) e (b) indicam grupos que diferem (post-hoc Tukey) com α =0,05.
111
Figura 1. (A) Contraste entre os pontos RGB de uma imagem e a média dos pontos da árvore (fundo). Linha tracejadademonstra o ponto onde separa a transecção horizontal da vertical.; (B) imagem utilizada para contagem dos pontosem imagem normal (C) imagem utilizada após a subtração do canal vermelho. Notar a mudança de coloraçãoacentuada do disco vermelho.
Resultados
Os tratamentos (com e sem vermelho) diferiram significativamente (F1,50=19,688; p<0,0001), assim
como o contraste entre cores (F4,50=15,153; p< 0,0001). Porém, a interação dos tratamentos com as cores
não foi significativa (F4,100=1,68 p=0,1690).
Com relação às cores, para o tratamento com vermelho as cores azul e branco foram as que mais
contrastaram com o fundo, diferindo das outras (Tukey p<0,05) (Fig.2). Já com a retirada do vermelho
houve uma formação de grupos de cores mais atrativas (amarelo, azul e branco) e outro com cores de
baixo contraste com o fundo e, portanto, crípticas (vermelho e preto) (Tukey p<0,05; Fig.2).
Figura 2. Médias dos logaritmos dos contrastes entre as cores e o fundo das árvores na Mata do Araldo. Em (A)tratamento com vermelho e em (B) sem vermelho. (a) e (b) indicam grupos que diferem (post-hoc Tukey) com α =0,05.
111
Figura 1. (A) Contraste entre os pontos RGB de uma imagem e a média dos pontos da árvore (fundo). Linha tracejadademonstra o ponto onde separa a transecção horizontal da vertical.; (B) imagem utilizada para contagem dos pontosem imagem normal (C) imagem utilizada após a subtração do canal vermelho. Notar a mudança de coloraçãoacentuada do disco vermelho.
Resultados
Os tratamentos (com e sem vermelho) diferiram significativamente (F1,50=19,688; p<0,0001), assim
como o contraste entre cores (F4,50=15,153; p< 0,0001). Porém, a interação dos tratamentos com as cores
não foi significativa (F4,100=1,68 p=0,1690).
Com relação às cores, para o tratamento com vermelho as cores azul e branco foram as que mais
contrastaram com o fundo, diferindo das outras (Tukey p<0,05) (Fig.2). Já com a retirada do vermelho
houve uma formação de grupos de cores mais atrativas (amarelo, azul e branco) e outro com cores de
baixo contraste com o fundo e, portanto, crípticas (vermelho e preto) (Tukey p<0,05; Fig.2).
Figura 2. Médias dos logaritmos dos contrastes entre as cores e o fundo das árvores na Mata do Araldo. Em (A)tratamento com vermelho e em (B) sem vermelho. (a) e (b) indicam grupos que diferem (post-hoc Tukey) com α =0,05.

112
Discussão
O padrão de luz em uma floresta pode mudar no espaço e no tempo, afetando a capacidade
de visão dos animais. Com a variação da iluminação do ambiente, o padrão de cor de alguns
animais pode mudar de críptico para conspícuo, ou vice-versa, o que influencia diretamente nas
interações de predação de predadores visuais.
De acordo com o observado durante esse estudo, a variação entre o contraste das cores com
a retirada do vermelho é menor do que com a visão do vermelho. Assim, podemos dizer que
mamíferos e aves têm percepção distinta das colorações aposemáticas das presas, corroborando a
hipótese.
Entre os indivíduos que enxergam todo o espectro do visível a cor mais contrastante com o
fundo entre as cores testadas foi o azul. Isso pode ser explicado pelo fato de que as cascas das
árvores apresentam coloração marrom, que estão mais próximas da faixa do vermelho (600 μm) e
mais distantes do espectro azul (400 μm). Além disso, retirando a cor vermelha das imagens não há
uma variação significativa no contraste entre as cores amarelo, azul e branco, porém estas
contrastaram com preto e vermelho, o que vai de encontro à teoria das cores (Gerritsen, 1975), pois
a exclusão da visão do vermelho torna essa cor acinzentada, muito próxima ao preto.
Apesar do método utilizando imagens ter sido eficaz para testar nossa hipótese, seria
importante realizar estudos que utilizassem equipamentos como espectrofotômetro para o registro
da luz refletida, pois assim seria possível considerar comprimentos de onda fora da região do visível
(ultravioleta e infravermelho) sendo que alguns animais podem ter capacidade de enxergar essas
faixas.

113
Anderson MC 1964 Studies of the woodland light climate. I. The photographic
computation of light conditions. Journal of Ecology 52:27-41.
Cunha DCF, Dantas HVM, Moraes AMM 2006 Electroretinographic study of chromatic
vision. Arq Bas Oftalmol. 69(6):857-63.
Ricklefs RE 2010 A Economia da Natureza. 6ª ed. Editora Guanabara Koogan, Rio de
Janeiro.
Théry M 2001 Forest light and its influence on habitat selection. Plant Ecology 153:
251–261.
Gerritsen F 1975 Theory and practice of color.

114
GRADIENTE DE LUZ INFLUENCIA A DIVERSIDADE FUNCIONAL DE PLANTAS?
Diego Corrêa Alves, Fernanda Ceschin, Letícia Cucolo Karling, Luciana Zago, Monica
Vasques Mangas Pereira, Ricardo Augusto Camargo, Vinícus Marcílio da Silva
Orientador: Marcia Marques
Resumo: A estruturação de comunidades ocorre essencialmente através dos filtros
abióticos e bióticos. Neste contexto, o objetivo deste trabalho foi verificar se a riqueza
de morfotipos e a diversidade funcional dos de plantas é influenciada pela cobertura do
dossel. Os resultados encontrados para a riqueza e diversidade funcional não suportam
as hipóteses de que maiores coberturas de dossel resultam em uma maior riqueza e
diversidade funcional. No entanto, eles apontam para um padrão de atuação do filtro
abiótico ao longo do gradiente, não apenas em seu extremo.
Palavras-chave: Diversidade funcional; riqueza; intensidade luminosa.
Introdução
A estruturação de uma comunidade é determinada por processos históricos
biogeográficos (Cavender-Bares et al. 2009), filtros ambientais e interações bióticas
(Dry et al. 2012). A ação de um filtro abiótico deve gerar um padrão de convergência de
atributos funcionais entre as espécies de uma comunidade. Por outro lado, espécies
ecologicamente muito similares não poderiam coexistir em um mesmo local. Assim, a
competição deveria atuar como um filtro biótico e produzir divergência funcional dentro
das comunidades (Tilman 1986).
A fotossíntese das plantas é diretamente dependente da radiação de luz solar
(Mooney & Ehleringer 1997), portanto, esta atua como um filtro abiótico para esses
organismos. A cobertura do dossel, por sua vez, determina a intensidade desse filtro, de
modo que quanto maior for a cobertura do dossel, menor será a intensidade de luz sobre
as plantas. Diante desse contexto, visamos testar as seguintes hipóteses: 1) comunidades
de plantas em estratos inferiores de floresta, sob maiores coberturas de dossel serão
compostas por uma maior diversidade funcional; 2) estratos inferiores com maiores
coberturas de dossel serão compostos por uma maior riqueza de morfoespécies.
Material e métodos

115
Coleta de dados
O estudo foi realizado na ilha Carioca e nas margens do rio Caracu, em Porto
Rico-PR. Em cada uma destas áreas foram determinados 10 pontos amostrais
aleatórios, nos quais a cobertura de dossel foi fotografada e a imagem tratada no
programa Image J, onde calculou-se a porcentagem de abertura de dossel.
Nos pontos amostrais foram delimitadas parcelas circulares com um metro de
raio, nas quais todas as plantas com a altura entre 20 cm e 1 m foram amostradas e
separadas em morfotipos. Cada indivíduo teve os seguintes atributos medidos: textura
da folha (coriácea, cartácea, membranácea); tipo de ápice foliar (acuminado, não
acuminado), número de folhas, largura e comprimento das folhas (a partir dos quais foi
estimada a área foliar, considerando a fórmula da área da elipse); circunferência da base
do caule, altura da planta; área fotossintetizante (calculada a partir da área da folha
multiplicada pelo número de folhas do indivíduo); presença de anexos epidérmicos e
forma de vida.
Para cada parcela, a diversidade funcional foi determinada, através da entropia
quadrática de Rao. Posteriormente estes valores de diversidade, a riqueza e a
abundância das parcelas foram relacionados com a abertura do dossel, através de
análises de regressão.
Resultados
No total foram amostrados 107 indivíduos, compreendendo 12 morfotipos, nas
20 parcelas. A riqueza de morfotipos não apresentou relação com a abertura de dossel
(Fig. 1 A), no entanto, ao aplicar uma relação quadrática entre a riqueza e a abertura de
dossel observou-se um melhor ajuste dos dados (R² = 0,38) (Fig. 1B).
A B
115
Coleta de dados
O estudo foi realizado na ilha Carioca e nas margens do rio Caracu, em Porto
Rico-PR. Em cada uma destas áreas foram determinados 10 pontos amostrais
aleatórios, nos quais a cobertura de dossel foi fotografada e a imagem tratada no
programa Image J, onde calculou-se a porcentagem de abertura de dossel.
Nos pontos amostrais foram delimitadas parcelas circulares com um metro de
raio, nas quais todas as plantas com a altura entre 20 cm e 1 m foram amostradas e
separadas em morfotipos. Cada indivíduo teve os seguintes atributos medidos: textura
da folha (coriácea, cartácea, membranácea); tipo de ápice foliar (acuminado, não
acuminado), número de folhas, largura e comprimento das folhas (a partir dos quais foi
estimada a área foliar, considerando a fórmula da área da elipse); circunferência da base
do caule, altura da planta; área fotossintetizante (calculada a partir da área da folha
multiplicada pelo número de folhas do indivíduo); presença de anexos epidérmicos e
forma de vida.
Para cada parcela, a diversidade funcional foi determinada, através da entropia
quadrática de Rao. Posteriormente estes valores de diversidade, a riqueza e a
abundância das parcelas foram relacionados com a abertura do dossel, através de
análises de regressão.
Resultados
No total foram amostrados 107 indivíduos, compreendendo 12 morfotipos, nas
20 parcelas. A riqueza de morfotipos não apresentou relação com a abertura de dossel
(Fig. 1 A), no entanto, ao aplicar uma relação quadrática entre a riqueza e a abertura de
dossel observou-se um melhor ajuste dos dados (R² = 0,38) (Fig. 1B).
A B
115
Coleta de dados
O estudo foi realizado na ilha Carioca e nas margens do rio Caracu, em Porto
Rico-PR. Em cada uma destas áreas foram determinados 10 pontos amostrais
aleatórios, nos quais a cobertura de dossel foi fotografada e a imagem tratada no
programa Image J, onde calculou-se a porcentagem de abertura de dossel.
Nos pontos amostrais foram delimitadas parcelas circulares com um metro de
raio, nas quais todas as plantas com a altura entre 20 cm e 1 m foram amostradas e
separadas em morfotipos. Cada indivíduo teve os seguintes atributos medidos: textura
da folha (coriácea, cartácea, membranácea); tipo de ápice foliar (acuminado, não
acuminado), número de folhas, largura e comprimento das folhas (a partir dos quais foi
estimada a área foliar, considerando a fórmula da área da elipse); circunferência da base
do caule, altura da planta; área fotossintetizante (calculada a partir da área da folha
multiplicada pelo número de folhas do indivíduo); presença de anexos epidérmicos e
forma de vida.
Para cada parcela, a diversidade funcional foi determinada, através da entropia
quadrática de Rao. Posteriormente estes valores de diversidade, a riqueza e a
abundância das parcelas foram relacionados com a abertura do dossel, através de
análises de regressão.
Resultados
No total foram amostrados 107 indivíduos, compreendendo 12 morfotipos, nas
20 parcelas. A riqueza de morfotipos não apresentou relação com a abertura de dossel
(Fig. 1 A), no entanto, ao aplicar uma relação quadrática entre a riqueza e a abertura de
dossel observou-se um melhor ajuste dos dados (R² = 0,38) (Fig. 1B).
A B

116
Fig. 1. A) Relação linear entre a abertura do dossel e a riqueza de plantas, B) Relação
quadrática entre a abertura do dossel e a riqueza das plantas.
A diversidade funcional de Rao teve relação com a abertura de dossel (Fig. 2), no
entanto, o padrão desta relação não foi linear (R²= 0,05; p>0,05). Contudo, ao aplicar
relação quadrática aos dados observou-se melhor ajuste (R²=0,46)(Fig. 2).
Fig. 1. Relação entre abertura do dossel e diversidade funcional de Rao .
Discussão
y = 0.0008x + 0.2701R² = 0.0535
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
0 20 40 60 80 100
Dive
rsid
ade
func
iona
l de
Rao
Abertura de Dossel
y = -0.0001x2 + 0.0134x + 0.088R² = 0.4641
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0.4
0.45
0.5
0 20 40 60 80 100
Dive
rsid
ade
func
iona
l de
Rao
Abertura de Dossel

117
Os resultados encontrados para a riqueza e diversidade funcional não
suportam as hipóteses de que maiores coberturas de dossel resultam em uma
maior riqueza e diversidade funcional. Entretanto, as relações encontradas refletem
os conhecimentos de que intensidades luminosas extremas desfavorecem a
existência de plantas (Begon et al. 2007), já que baixas iluminações são
insuficientes para atividades fotossintéticas, enquanto iluminações excedentes à
capacidade fotossintética máxima podem estar além das capacidades de dissipação
da energia da planta e gerar estresses térmicos e hídricos (Mooney & Ehleringer
1997).
Frente a isto acreditamos que embora os resultados tenham indicado um
possível padrão com suporte biológico, seriam necessárias novas amostragens para
conseguir confirmá-lo. O estudo deixa evidente a necessidade de maior cautela na
análise de filtros ambientais a partir de um único fator, já que estes filtros não
atuam de maneira isolada. Além disso, é importante ressaltar que os filtros atuam
ao longo de um gradiente, o qual não foi totalmente contemplado no estudo.
Referências
Cavender-Bares J, Kozak KH, Fine PVA, Kembel SW 2009 The merging of community
ecology and phylogenetic biology. Ecological Letters 12:693–715.
Dray S, Pélissier R, Couteron P, Fortin MJ, Legendre P, Peres-Neto PR, Bellier E,
Bivand R, Blanchet FG, De Cáceres M, Dufour A, Heegard E, Jombart T, Munoz
F, Oksanen J, Thioulouse J, Wagner HH 2012 Community ecology in the age of
multivariate multiscale spatial analysis. Ecological Monographs 82(3):257–275.
Mooney HA, Ehleringer JR 1997 Photosynthesis. In: Crawley M (Ed.). Plant Ecology
2nd Ed. Blackwell Publishing.
Tilman D 1986 Plant ecology. Blackwell, Oxford.

118
INFLUÊNCIA DO TAMANHO DA INFLORESCÊNCIA NA RIQUEZA E VISITA DE INSETOS EM
UMA LEGUMINOSA TROPICAL
EMANUEL Razzolini; FERNANDO G. de Oliveira; FLÁVIO M. Marteleto;
NATÁLIA C. L. dos Santos; SANDRA M. N. de Andrade
Orientador:
Fernando
Passos
Resumo
A interação planta-insetos é bem conhecida e estudada na ecologia. A principal
interação nessa relação é a busca por fontes de alimento pelos insetos em contrapartida
com a dispersão de pólens das plantas. Nesse estudo foi verificado uma tendência da
relação positiva entre tamanho de inflorescência e riqueza de morfotipos de insetos;
além de uma relação positiva e significativa entre o número de visitas de insetos com o
aumento do tamanho das inflorescências.
Palavras-chave: Crotalaria, abundância, insetos, riqueza, inflorescência
Introdução
Em ecologia, é bem conhecido que insetos são atraídos por plantas com
inflorescência, devido a sua cor e cheiro. Estes animais procuram néctar e pólen como
fonte de alimento. Grandes áreas de inflorescência podem abrigar mais espécies de
insetos por serem maiores alvos de colonização e ainda por terem mais recursos. Além
do efeito da área, os organismos se comportam de modo a maximizar o ganho
energético melhorando a eficiência do encontro, captura e consumo do alimento. Esse
comportamento é conhecido como forrageamento ótimo proposto por Robert
MacArthur e Eric Pianka em 1966 (Begon et al., 2010; Ricklefs, 2003).
O gênero Crotalaria pertence à família Leguminosae, subfamília Faboideae.
Dentre as 31 espécies nativas e 11 exóticas catalogadas para o Brasil, foram citadas seis
espécies para a Planície de Inundação do Alto Rio Paraná (PIARP). Assim, os objetivos
deste estudo foram (i) analisar a riqueza de espécies de insetos em inflorescências de
Crotalaria sp. de diferentes comprimentos e (ii) avaliar a frequência de visitas de
insetos nas inflorescências de Crotalaria sp com diferentes tamanhos. Sob essa

119
perspectiva, as hipóteses testadas foram: (i) O tamanho de inflorescência de Crotelaria
sp. influencia a riqueza de espécies de insetos. (ii) O tamanho da inflorescência
influencia a frequência de visitas de insetos. Desse modo, é predito que (i)
Inflorescências maiores comportam um maior número de espécies e (ii) Inflorescências
maiores são mais atrativas do ponto de vista visual
Material e Métodos
Área de estudo
O estudo foi desenvolvido próximo à base de pesquisas do Nupélia, município
de Porto Rico, Paraná, Brasil. Situada no trecho superior da Planície de Inundação do
Alto Rio Paraná, a área de estudo está inserida em uma região de clima subtropical, com
temperatura média anual entre 23ºC e 24ºC. A vegetação pertence a uma região do
Bioma Mata Atlântica conhecida como Floresta Estacional Semidecidual, apresentando
ambientes secos e pantanosos, remanescentes florestais ripários, e áreas de Cerrado e de
Campos.
Amostragem e Análise de dados
Em uma primeira etapa, foram coletadas aleatoriamente 40 inflorescências de
Crotalaria sp. com sacos plásticos envolvendo toda a inflorescência, de modo que se
capturasse os organismos presentes na planta. Em laboratório, os sacos plásticos foram
armazenados em ambiente refrigerado para minimizar a atividade dos organismos e,
depois de 30 minutos, as amostras foram examinadas em lupa para a identificação e
contagem dos morfotipos de organismos associados nos diferentes comprimentos de
inflorescências. As inflorescências foram agrupadas em três classes de tamanhos:
pequena (P), com comprimento total (CT) menor ou igual a 7,0 cm; média (M), com
comprimento entre 7,5 e 14,0 cm; e grande (G), com tamanho acima de 14,5 cm. Após
isso, o número de insetos que visitaram cada tamanho de inflorescência foi estimado
durante o tempo de dois minutos e meio. Os organismos foram identificados por
morfotipos diferenciados morfologicamente com características gerais. Os dados foram
analisados através de uma análise de variância (ANOVA) para testar se há diferença
entre as médias de riqueza entre os comprimentos de inflorescência. O programa
estatístico utilizado foi o Statistica 7.0.
Resultados
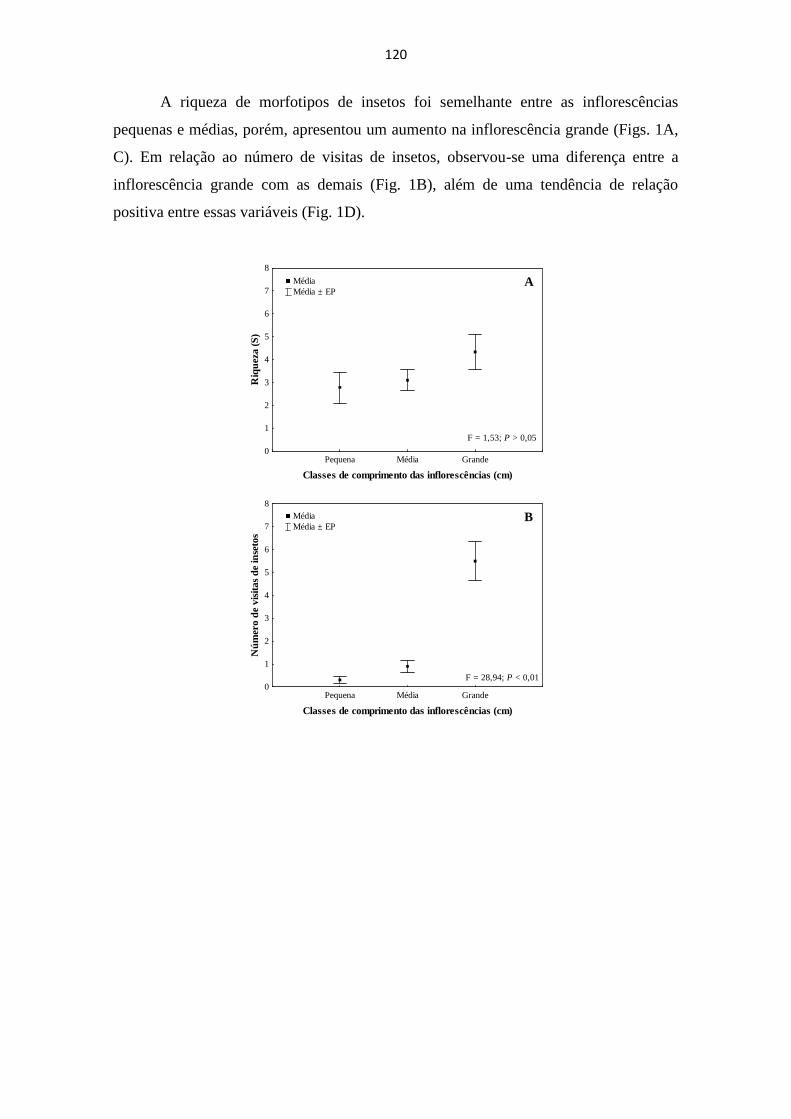
120
A riqueza de morfotipos de insetos foi semelhante entre as inflorescências
pequenas e médias, porém, apresentou um aumento na inflorescência grande (Figs. 1A,
C). Em relação ao número de visitas de insetos, observou-se uma diferença entre a
inflorescência grande com as demais (Fig. 1B), além de uma tendência de relação
positiva entre essas variáveis (Fig. 1D).
Pequena Média Grande
Classes de comprimento das inflorescências (cm)
0
1
2
3
4
5
6
7
8
Riq
ueza
(S)
Média Média ± EP
A
F = 1,53; P > 0,05
Pequena Média Grande
Classes de comprimento das inflorescências (cm)
0
1
2
3
4
5
6
7
8
Núm
ero
de v
isita
s de
inse
tos
Média Média ± EP
F = 28,94; P < 0,01
B

121
Figura 1. Médias da riqueza (A) e número de visitas de insetos (B) em relação às
classes de comprimento das inflorescências. Os gráficos C e D demonstram as relações
entre as respectivas variáveis.
Coleopteras (2, 3 e 4) (Tab. I) foram presentes apenas nas inflorescências
maiores enquanto que alguns outros morfotipos se apresentaram apenas nas classes de
comprimento pequena e média (Tab. I).
Tabela I. Lista de morfotipos de insetos capturados demostrando a presença e ausência
nas diferentes classes de comprimento das inflorescências.
Morfotiposde insetos
Pequena Média Grande
Formiga 1
Formiga 2
Formiga 3
Lepidoptera 1
Lepidoptera 2
0 2 4 6 8 10 12 14 16 18 20 22
Comprimento das inflorescências (cm)
0
2
4
6
8
10
Riq
ueza
(S)
y = 0,10*x + 2.21
P > 0,05 C
2 4 6 8 10 12 14 16 18 20 22 24 26 28
Comprimento das inflorescência (cm)
0
2
4
6
8
10
12
14
Núm
ero
de v
isita
s de
inse
to
y = 0,33*x - 2,1274
P < 0,01 D

122
Ortoptera 1
Ortoptera 2
Vespoidea
Hemiptera 1
Hemiptera 2
Coleoptera 1
Coleoptera 2
Coleoptera 3
Coleoptera 4
Diptera 1
Diptera 2
Diptera 3
Diptera 4
Diptera 5
Aranea
Apidea
Pulgao 1
Pulgao 2
Discussão
Um dos pressupostos clássicos para essa teoria da Hipótese de diversidade
(Williams 1964) é que uma área maior só apresenta mais espécies se houver uma maior
diversidade de habitats a serem ocupados. Desse modo a não diferenciação em termos
de riqueza em relação ao tamanho das inflorescências, está diretamente relacionado ao
fato das mesmas apresentarem estrutura bastante similar na escala estudada. Uma vez
que o padrão das flores é o mesmo para qualquer um dos tamanhos, as mesmas
funcionam de fato como habitats homogêneos e semelhantes entre si. Ainda, baixa
fidelidade quanto ao habitat dos morfotipos encontrados pode ter contribuído de forma
significativa para a não diferenciação dos padrões de riqueza.
A diferença significativa encontrada na comparação de visitas entre os diferentes
tamanhos de inflorescência pode ser explicada pelo fato de que as inflorescências
maiores possuem flores maiores e mais aparentes, efeito que funcionaria como um
atrativo muito maior para os insetos em busca de alimento. Essa predição está

123
condizente com teoria do forrageamento ótimo, no qual a chance de maximização da
obtenção de energia por tempo é muito maior, se um organismo consegue detectar a
uma fonte de alimento mais conspícua (Ricklefs, 2003). Porém, os baixos números de
visitas verificadas, podem estar relacionados ao horário de avaliação das mesmas, uma
vez que fatores abióticos como baixa luminosidade e temperatura, que afetam limitações
fisiológicas dos insetos, podem ter interferido negativamente as taxas de visitas.
Agradecimentos
Os autores agradecem ao professor Fernando Passos pela orientação do projeto
de estudo e aos programas de pós-graduação da Universidade Federal do Paraná e
Universidade Estadual de Maringá pela disciplina ofertada para o treinamento e
aprendizagem das pesquisas ecológicas em campo.
Referencias bibliográficas
Ricklefs, R.E. 2003. A economia da natureza. 5ª Edição, Rio de Janeiro, Brasil.
Townsend, C.R, Begon, M., Harper, J.L. 2010. Fundamentos em ecologia. 3ª Edição,
São Paulo, Brasil.
Williams, C.B. 1964. Patterns in the Balance of nature. Academic Press, London

124
EXISTEM EFEITOS DA DISTÂNCIA DA BORDA DE BANCOS DE MACRÓFITAS AQUÁTICAS
NA DIVERSIDADE DE GRUPOS FUNCIONAIS DE INVERTEBRADOS?
Hugo José Message, Priscilla Guedes Gambale, Renata Gabriela Noguchi, SimoneSatsuki Takeuchi, Taise Miranda Lopes.
Orientador: Roger Mormul
Resumo: As macrófitas aquáticas são responsáveis por promover estruturação aosambientes, ocasionando uma maior diversidade de invertebrados. Assim, testou-se ahipótese de que a distribuição de espécies de invertebrados é determinada pela distânciada posição no banco de macrófitas em relação à borda. De acordo com os resultados daanálise de variância, os atributos riqueza de espécies, abundância, comprimento padrãoe diversidade de Shannon não apresentaram significância, possivelmente devido à baixaheterogeneidade ambiental e de espécies de macrófitas no banco
Palavras-chave: Herbivoria, Interações, Plantas aquáticas, Grupos funcionais.
Introdução
Macrófitas aquáticas são componentes chaves nos ambientes aquáticos porserem fonte de alimento e contribuirem para aumentar a complexidade do habitat,influenciando a variação da diversidade de espécie dos organismos em várias escalasespaciais. Além disso, são responsáveis por influenciar as variáveis climáticas, físicas equímicas da água através da ciclagem de nutrientes, da liberação de substânciasalelopáticas e da geração de detritos e produtos derivados (Walker et al, 2013). Lagoasrasas tropicais geralmente apresentam grande abundância de macrófitas aquáticas.
Os maiores bancos de macrófitas flutuantes encontram-se próximos às margens,região que abriga a maior densidade de raízes, possui maior complexidade de habitats etem influência positiva sobre o número de espécies residentes por oferecer maiornúmero de microhabitats e criação de nichos. As regiões desses bancos mais próximos àzona pelágica são menos densas (Thomaz et al, 2008) e apresentam poucosinvertebrados devido a menor quantidade de perifíton e fitoplâncton para alimentação,além da presença e predação por peixes derivados (Walker et al, 2013).
Testou-se a hipótese de que a distribuição de morfotipos de invertebrados édeterminada pela distância da posição dentro do banco de macrófitas aquáticas em

125
relação à borda. Espera-se que exista uma maior densidade de herbívoros (perfuradores,raspadores e cortadores) próximos a esta localização.
Materiais e Métodos
A coleta foi realizada no ressaco do Bilé, localizado na planície de inundação doalto rio Paraná. Foram amostrados 4 bancos de macrófitas aquáticas, o primeiropróximo a entrada do ressaco e os outros distantes 50m entre si. Em cada banco foramamostrados três pontos em distâncias crescentes da borda (margem) em direção à zonapelágica (regiões: borda, centro e pelágica). Os invertebrados foram coletados compeneira de malha 1mm com duas submersões da peneira por ponto, recolhidos comauxílio de pinça e imediatamente armazenados em potes com álcool.
Em laboratório, as amostras foram triadas em microscópio estereoscópio e osindivíduos foram classificados de acordo com seus grupos funcionais (predador,perfurador, raspador, cortador e detritívoro), morfotipos e mensurados quanto aocomprimento total (cm). Foram quantificadas a riqueza e a abundância de cada grupofuncional e morfotipo.
Para avaliar como os morfotipos dos invertebrados se distribuem no interior dosbancos de macrófitas aquáticas foram selecionados os atributos de riqueza, abundância,comprimento e diversidade de Shannon. Para testar possíveis diferenças na variaçãodestes atributos foi utilizada a análise de variância unifatorial (One-way ANOVA),considerando a localização dos bancos de macrófitas (borda, centro e pelágica) comofator. Essas análises foram feitas no programa Statistic 7.0.
Para sumarizar os padrões de similaridade na composição de morfotipos deinvertebrados em relação ao fator localização (borda, centro e pelágica) foi aplicadouma PCoA (análise de coordenadas principais). Nessa análise de ordenação, aabundância relativa dos morfotipos foi transformada e posteriormente aplicado ocoeficiente de similaridade de Bray-Curtis. Para verificar a diversidade beta daslocalizações foi realizada uma PERMDISP, que compara a dispersão dos dados emrelação à centroide do fator, sendo que quanto maior a dispersão, maior a diversidadebeta. As diferenças entre o fator foi determinada usando uma análise de variânciapermutacional (PERMANOVA), aplicada sobre uma matriz de similaridade comdistâncias de Bray-Curtis. Para testar a significância estatística (p<0,05), foi utilizadopermutações de Monte Carlo (N = 9999). Essas análises foram feitas no programaPRIMER 6 add PermanovaTM.
Resultados
Foram identificados 5 grupos funcionais de invertebrados associados asmacrófitas aquáticas, sendo que a maior abundância foi de predadores e perfuradores na

126
região pelágica (Figura 1). Em geral, a borda apresentou maior abundância deperfuradores e raspadores. Cortadores e detritívoros ocorreram em menor quantidade eos raspadores apresentaram abundância intermediária, ocorrendo de forma homogêneanas três localizações.
Figura 1: Abundância dos grupos funcionais nos três estratos de macrófita (borda,centro e pelágica).
De acordo com a análise de variância unifatorial (one-way ANOVA;localização) nenhum dos atributos da comunidade foram significativamente distintosentre a localização dos bancos de macrofitas. (riqueza de espécies,F=1,09, P=0,38;abundância, F=2,56, P=0,13; comprimento padrão, F=0,73, P=0,51; diversidade deShannon, F=0,69, P=0,52) (Figura 2).
126
região pelágica (Figura 1). Em geral, a borda apresentou maior abundância deperfuradores e raspadores. Cortadores e detritívoros ocorreram em menor quantidade eos raspadores apresentaram abundância intermediária, ocorrendo de forma homogêneanas três localizações.
Figura 1: Abundância dos grupos funcionais nos três estratos de macrófita (borda,centro e pelágica).
De acordo com a análise de variância unifatorial (one-way ANOVA;localização) nenhum dos atributos da comunidade foram significativamente distintosentre a localização dos bancos de macrofitas. (riqueza de espécies,F=1,09, P=0,38;abundância, F=2,56, P=0,13; comprimento padrão, F=0,73, P=0,51; diversidade deShannon, F=0,69, P=0,52) (Figura 2).
126
região pelágica (Figura 1). Em geral, a borda apresentou maior abundância deperfuradores e raspadores. Cortadores e detritívoros ocorreram em menor quantidade eos raspadores apresentaram abundância intermediária, ocorrendo de forma homogêneanas três localizações.
Figura 1: Abundância dos grupos funcionais nos três estratos de macrófita (borda,centro e pelágica).
De acordo com a análise de variância unifatorial (one-way ANOVA;localização) nenhum dos atributos da comunidade foram significativamente distintosentre a localização dos bancos de macrofitas. (riqueza de espécies,F=1,09, P=0,38;abundância, F=2,56, P=0,13; comprimento padrão, F=0,73, P=0,51; diversidade deShannon, F=0,69, P=0,52) (Figura 2).
127
A B
C D
Figura 2: Média e Desvio-Padrão da riqueza total (A), abundância total (B),comprimento (C) e diversidade de Shannon (D) na borda, centro e região pelágica emmacrófitas aquáticas flutuantes.
A análise de coordenadas principais apresentou eixos com baixa representaçãoda variação dos dados (PCoA 1 = 25,9% e PCoA = 17,1%). Além disso, a centroide dasregiões não apresentou uma distância significativa entre si (Figura 3). A PERMDISPnão apresentou significância (F= 2,18; P = 0,24), demonstrando uma homogeneidade dadispersão, e, consequentemente, uma baixa diversidade beta entre os locais nos bancosde macrófitas aquáticas.

128
Figura 3: Gráfico representando a ordenação da composição dos morfotipos deinvertebrados nas três localizações nos bancos de macrófitas (borda, centro e pelágica).
A análise de variância permutacional (PERMANOVA) também não identificoudiferença significativa na composição dos morfotipos de invertebrados em relação àlocalização do banco (Pseudo F = 0,90; p = 0,57).
Discussão
A presença de uma diversidade de morfotipos nas macrófitas aquáticas pode serexplicada pela interação mutualística entre a planta e os invertebrados. A alta produçãode matéria orgânica pelas macrófitas atrai os invertebrados aquáticos, que acabam porse alimentar da camada epífitica das raízes, conferindo maior crescimento a elas(Brönmark, 1985).
A similaridade de morfotipos de invertebrados presentes nos diferentes locaisdos bancos de macrófitas pode ser devido à similaridade na qualidade da água(propriedades químicas) da região pelágica e da borda de bancos de macrófitas emlagoas conectadas ao Alto Rio Paraná (Thomaz, 2008). Considerando que fatoresquímicos da água podem influenciar na distribuição dos invertebrados, a semelhançaentre os pontos também confere semelhança entre os indivíduos ali presentes. Alémdisso, os invertebrados apresentam preferência para determinada espécie de planta, poisessas diferem entre si quanto à palatabilidade e composição química (Elger & Barrat-Segretain, 2004). Desse modo, a homogeneidade de espécies de macrófitas entre ospontos de amostragem (composto predominantemente por Eichhornia azurea) pode terrefletido também na homogeneidade da composição dos morfotipos de invertebradosassociados.

129
Referências bibliográficas
Brönmark C 1985 Interactions between macrophytes, epiphytes and herbivores: anexperimental approach. Oikos 45:26-30.
Elger A, Barra-Segretain MH 2004 Plant palatability can be inferred from a single-datefeeding trial. Functional Ecology 18: 483-488.
Thomaz SM, Dibble ED, Evangelista LR, Higuti J, Bini LM 2008. Influence of aquaticmacrophyte habitat complexity on invertebrate abundance and richness intropical lagoons. Freshwater Biology 53: 358-367.
Walker PD, Wijnhoven S, Velde Gvd 2013 Macrophyte presence and growth forminfluence macroinvertebrate community structure. Aquatic Botany 104: 80-87.

130
VARIAÇÃO DE RIQUEZA E COMPOSIÇÃO DE ESPÉCIES DE MACRÓFITAS AO LONGO DE
UM GRADIENTE AMBIENTAL
Ariane Ghelfi, Bianca Possamai, Daniel Santos, Juliana Wojciechowski e Vivian Cionek
Orientador: Sidinei Thomaz
Resumo: A associação entre fatores ambientais e a riqueza e composição de espécies de
macrófitas de uma lagoa conectada ao rio Paraná foi avaliada. A distância da entrada da lagoa,
juntamente com a redução da luminosidade, determinaram a riqueza de espécies, enquanto a
profundidade foi preditora da composição da assembleia. Nossos resultados indicam que as
macrófitas encontram-se organizadas perpendicular e longitudinalmente ao eixo principal da
lagoa, sendo que a intensidade luminosa e a profundidade podem ser fatores determinantes
dessa zonação.
Palavras-chave: macrófitas, zona eufótica, interação espécie-ambiente.
Introdução
Macrófitas aquáticas desempenham diversos papéis ecológicos importantes, como
ciclagem de nutrientes e estruturação de habitats (Biudes e Camargo, 2008). Essas plantas são
classificadas como emergentes, flutuantes-livres, semi-aquáticas, epífitas, submersas livres e
submersas fixas, podendo formar gradientes de distribuição ao longo de lagos (Matias et al.,
2003; Rotta et al., 2010). Essa distribuição pode ser determinada pela profundidade, pela
concentração de íons e por competição, entre outros fatores (Esteves, 2011). Porém, a
profundidade parece ser o principal determinante, pois esse fator é inversamente
proporcional a quantidade de compostos orgânicos e inorgânicos na coluna d'água
(Preisendorfer, 1986). Entre as diversas interações ecológicas interespecíficas observadas em
macrófitas, é comum observar a diminuição de espécies submersas com o aumento de
espécies que impedem a penetração da luz na coluna d'água (Wetzel, 1983).
Este trabalho objetivou verificar a associação entre variáveis ambientais e a riqueza e
composição de espécies de macrófitas em um gradiente longitudinal de uma lagoa conectada
ao rio Paraná. Também visou analisar a relação entre macrófitas submersas e cobertura da
macrófita flutuante Eichhornia azurea. As seguintes hipóteses foram testadas: i) A riqueza de

131
espécies de macrófitas aumenta e a composição muda ao longo de um gradiente longitudinal
da lagoa; ii) A composição de espécies varia com a profundidade (iii); A radiação luminosa é o
fator determinante da presença e abundância de espécies submersas. Espera-se que ao longo
do gradiente longitudinal a profundidade e transparência diminuam e que a concentração de
nutrientes aumente, culminando em aumento da riqueza e mudança na composição de
espécies. A predição da segunda hipótese é de que espécies emergentes dominem regiões
rasas e espécies flutuantes e submersas dominem regiões profundas. Para a terceira hipótese,
predizemos que locais com mata ciliar e regiões dominadas por E. azurea (uma espécie
enraizada flutuante) sejam colonizados de forma escassa por plantas submersas.
Material e métodos
As amostragens foram realizadas no Ressaco do Pau Véio (22°44’76”S;
53°15’11.16”W), uma lagoa pertencente à sub-bacia do Rio Paraná. Para testar se há
associação entre as variáveis ambientais e a riqueza e composição de espécies de macrófitas
no gradiente longitudinal da lagoa foram amostrados 24 pontos (6 na margem e 18 no centro
da lagoa) em diferentes bancos de macrófitas. A composição e cobertura de espécies foram
amostradas com um quadrado de 100 x 100 cm. Foram mensuradas a profundidade da coluna
de água (régua graduada, cm), a condutividade (condutivímetro, µS/cm), a distância da
entrada do ressaco (GPS, m) e a cobertura de dossel (presença e ausência). Para testar se a
cobertura de E. azurea determina a riqueza de espécies de macrófitas submersas (Cabomba
furcata e Egeria najas), a porcentagem de cobertura da primeira espécie foi estimada dentro
do quadrado de 100 x 100 cm e das submersas a partir de arrasto com garfo no fundo da lagoa
diretamente abaixo do quadrado.
Um nMDS foi realizado para sumarizar a composição de espécies em um eixo para a
geração dos modelos, a fim de investigar a distribuição dos dados no espaço multivariado.
Modelos lineares utilizando as variáveis abióticas como preditoras da composição (valores de
dissimilaridade geradas pelo nMDS) e riqueza de macrófitas foram realizados baseando-se no
critério de AIC. Para analisar diferenças na composição das espécies entre a margem e o meio
do ressaco realizou-se uma PERMANOVA, e a possível diferença entre a riqueza de espécie no
gradiente longitudinal foi testada por ANOVA, ambas análises com α=0,05. Uma correlação de
Pearson foi realizada entre a porcentagem de cobertura de Eichhornea azurea e as espécies
submersas.

132
Resultados
Os modelos revelaram que a variação na riqueza de espécies de macrófitas no ressaco
do Pau Véio é melhor explicada pela distância da entrada e pela cobertura do dossel. A maior
riqueza de espécies ocorreu nas áreas em que a cobertura do dossel foi menor (Fig. 1A). Já a
variação na composição de espécies no gradiente de distância foi predita pela profundidade e
pela condutividade, quando consideradas espécies de margem e meio do ressaco. A
composição de espécies foi diferente entre a margem e o meio do canal (F1,20=7,92; P
=0,0001), e essa diferença é representativa da variação de profundidade. Quando utilizamos
apenas as espécies do meio do ressaco, a distância em relação à entrada foi o que melhor
explicou a composição de espécies (Tab. 1).
Tabela 1. Modelos lineares (LM) para a riqueza e composição de espécies. As variáveisabióticas profundidade (cm), condutividade (µS/cm), distância da entrada (m) do ressaco ecobertura de dossel (aberto/fechado) foram utilizadas como preditoras para os modelos.
Componente Variável Preditora (Valor de importância) AICc
Canal/Margem
Riqueza Distância (0,94); Dossel (0,732) 66,704
Composição (%) Profundidade (1); Condutividade (0,957) 37,607
Composição (P/A) Profundidade (1); Condutividade (0,713) 35,830
Canal
Riqueza Distância (0,92); Dossel (0.83) 50,858
Composição (%) Distância (1) 6,120
Composição (P/A) Distância (1) 18,005
Foi verificada uma fraca correlação do grau de cobertura de E. azurea e a proporção de
macrófitas submersas (R= -0,39; P=0,12), com uma tendência de aumento da cobertura de
submersas com a diminuição da cobertura de E. azurea (Fig. 1B).
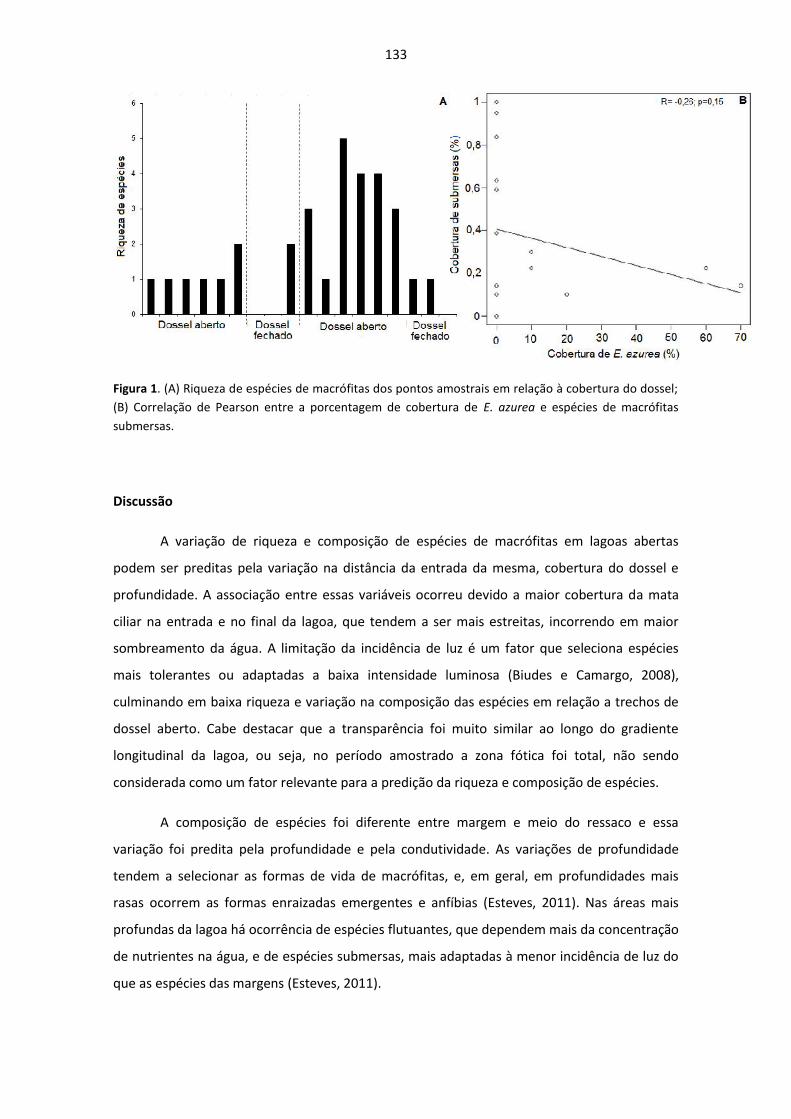
133
Figura 1. (A) Riqueza de espécies de macrófitas dos pontos amostrais em relação à cobertura do dossel;(B) Correlação de Pearson entre a porcentagem de cobertura de E. azurea e espécies de macrófitassubmersas.
Discussão
A variação de riqueza e composição de espécies de macrófitas em lagoas abertas
podem ser preditas pela variação na distância da entrada da mesma, cobertura do dossel e
profundidade. A associação entre essas variáveis ocorreu devido a maior cobertura da mata
ciliar na entrada e no final da lagoa, que tendem a ser mais estreitas, incorrendo em maior
sombreamento da água. A limitação da incidência de luz é um fator que seleciona espécies
mais tolerantes ou adaptadas a baixa intensidade luminosa (Biudes e Camargo, 2008),
culminando em baixa riqueza e variação na composição das espécies em relação a trechos de
dossel aberto. Cabe destacar que a transparência foi muito similar ao longo do gradiente
longitudinal da lagoa, ou seja, no período amostrado a zona fótica foi total, não sendo
considerada como um fator relevante para a predição da riqueza e composição de espécies.
A composição de espécies foi diferente entre margem e meio do ressaco e essa
variação foi predita pela profundidade e pela condutividade. As variações de profundidade
tendem a selecionar as formas de vida de macrófitas, e, em geral, em profundidades mais
rasas ocorrem as formas enraizadas emergentes e anfíbias (Esteves, 2011). Nas áreas mais
profundas da lagoa há ocorrência de espécies flutuantes, que dependem mais da concentração
de nutrientes na água, e de espécies submersas, mais adaptadas à menor incidência de luz do
que as espécies das margens (Esteves, 2011).
133
Figura 1. (A) Riqueza de espécies de macrófitas dos pontos amostrais em relação à cobertura do dossel;(B) Correlação de Pearson entre a porcentagem de cobertura de E. azurea e espécies de macrófitassubmersas.
Discussão
A variação de riqueza e composição de espécies de macrófitas em lagoas abertas
podem ser preditas pela variação na distância da entrada da mesma, cobertura do dossel e
profundidade. A associação entre essas variáveis ocorreu devido a maior cobertura da mata
ciliar na entrada e no final da lagoa, que tendem a ser mais estreitas, incorrendo em maior
sombreamento da água. A limitação da incidência de luz é um fator que seleciona espécies
mais tolerantes ou adaptadas a baixa intensidade luminosa (Biudes e Camargo, 2008),
culminando em baixa riqueza e variação na composição das espécies em relação a trechos de
dossel aberto. Cabe destacar que a transparência foi muito similar ao longo do gradiente
longitudinal da lagoa, ou seja, no período amostrado a zona fótica foi total, não sendo
considerada como um fator relevante para a predição da riqueza e composição de espécies.
A composição de espécies foi diferente entre margem e meio do ressaco e essa
variação foi predita pela profundidade e pela condutividade. As variações de profundidade
tendem a selecionar as formas de vida de macrófitas, e, em geral, em profundidades mais
rasas ocorrem as formas enraizadas emergentes e anfíbias (Esteves, 2011). Nas áreas mais
profundas da lagoa há ocorrência de espécies flutuantes, que dependem mais da concentração
de nutrientes na água, e de espécies submersas, mais adaptadas à menor incidência de luz do
que as espécies das margens (Esteves, 2011).
133
Figura 1. (A) Riqueza de espécies de macrófitas dos pontos amostrais em relação à cobertura do dossel;(B) Correlação de Pearson entre a porcentagem de cobertura de E. azurea e espécies de macrófitassubmersas.
Discussão
A variação de riqueza e composição de espécies de macrófitas em lagoas abertas
podem ser preditas pela variação na distância da entrada da mesma, cobertura do dossel e
profundidade. A associação entre essas variáveis ocorreu devido a maior cobertura da mata
ciliar na entrada e no final da lagoa, que tendem a ser mais estreitas, incorrendo em maior
sombreamento da água. A limitação da incidência de luz é um fator que seleciona espécies
mais tolerantes ou adaptadas a baixa intensidade luminosa (Biudes e Camargo, 2008),
culminando em baixa riqueza e variação na composição das espécies em relação a trechos de
dossel aberto. Cabe destacar que a transparência foi muito similar ao longo do gradiente
longitudinal da lagoa, ou seja, no período amostrado a zona fótica foi total, não sendo
considerada como um fator relevante para a predição da riqueza e composição de espécies.
A composição de espécies foi diferente entre margem e meio do ressaco e essa
variação foi predita pela profundidade e pela condutividade. As variações de profundidade
tendem a selecionar as formas de vida de macrófitas, e, em geral, em profundidades mais
rasas ocorrem as formas enraizadas emergentes e anfíbias (Esteves, 2011). Nas áreas mais
profundas da lagoa há ocorrência de espécies flutuantes, que dependem mais da concentração
de nutrientes na água, e de espécies submersas, mais adaptadas à menor incidência de luz do
que as espécies das margens (Esteves, 2011).

134
A ocorrência de macrófitas submersas em lagoas está condicionada à profundidade da
zona eufótica e à associação com a cobertura formada por espécies flutuantes livres e
emergentes que provocam um grau de sombreamento na coluna d’água (Duarte e Kalff, 1986),
o que pode aumentar a competição por luz entre as espécies submersas (Camargo et al.,
2006). Essa relação não foi verificada no presente estudo, e acredita-se que seja em
decorrência do pequeno tamanho dos bancos de E. azurea, permitindo assim a refração da luz
a partir das bordas (Esteves, 2011). A adição de maior número de réplicas e ambientes poderia
contribuir para verificar essa relação de maneira mais robusta.
Referências Bibliográficas
Biudes JFV, Camargo AFM 2008 Estudos dos fatores limitantes à produção primária por
macrófitas aquáticas no Brasil. Oecol. Bras. 12(1): 7-19.
Calheiros DF, Ferreira CJA 1996 Alterações limnológicas no rio Paraguai (“dequada”) e o
fenômeno natural de mortandade de peixes no Pantanal Mato-Grossense - MS.
Corumbá. MS: EMBRAPA-CPAP, 51p. (EMBRAPA- CPAP. Boletim de Pesquisa, 7).
Camargo AFM, Pesatto MM, Henry-Silva GG, Asumpção AM 2006 Primary production of
Utricularia foliosa L., Egeria densa Planchon and Cabomba furcata Schult & Schult.f
from rivers of the coastalplain of hte State of São Paulo, Brazil. Hydrobiologia 570: 35-
39.
Duarte MC, Kalff J 1986 Littoral slope as a predictor of the maximum biomass of submerged
macrophyte communities. Limnol. Oceanogr 31(5): 1072-1080.
Esteves FA 2011 Fundamentos de Limnologia. 2 ed. Rio de Janeiro: Interciência.
Preisendorfer RW 1986 Secchi disk science: visual optics of natural waters. Limnol. Oceanogr.
31:909-926.
Rotta LHS, Ferreira MS, Imai NN, Tachibana VM 2010 Análise espacial de macrófitas submersas
no reservatório de Porto Colômbia. III Simpósio Brasileiro de Ciências Geodésicas e
Tecnologias da Geoinformação.
Wetzel RG 1983 Limnology. 2 ed. Philadelphia: Saunders College, 767 p.

135
EFEITO DA TURBIDEZ DA ÁGUA NA PREDAÇÃO DO ZOOPLÂNCTON POR PEIXES
Diego Corrêa Alves, Fernanda Ceschin, Letícia Cucolo Karling, Luciana Zago, Monica
Vasques Mangas Pereira
Orientador: Juliana
Resumo
A relação entre a turbidez da água e densidades de organismos zooplanctônicos
é influenciada por interações bióticas. Visamos testar por experimentação se a predação
de peixes em zooplâncton é menor em águas mais túrbidas. Os resultados revelaram
maiores predações em águas claras, não suportando a hipótese proposta. Águas túrbidas
podem reduzir esforços com comportamentos anti-predatórios em peixes, aumentando
seus esforços alimentares, entretanto, a confirmação de que os resultados encontrados
estão relacionados a isto só pode ser dada por novas experimentações.
Palavras-chave: Microcosmos, esforços alimentares, planície de inundação, rio Paraná.
Introdução
A predação exerce forte efeito nas comunidades de presas, este efeito varia de
acordo com a densidade e o tamanho dos predadores, sua preferência alimentar e as
condições abióticas do habitat. (Sih et al. 1985). Além dos efeitos diretos (consumo das
presas) a predação tem efeitos indiretos (redução das taxas de forrageamento, migração,
coexistência de presas) na estrutura das comunidades (Sih 2010 et al., Morin 1999). A
estrutura da comunidade de zooplâncton, por exemplo, pode ser determinada por fatores
bióticos e abióticos, como vêm sendo abordado em muitos trabalhos realizados na
planície de inundação do alto rio Paraná (Bonecker & Lansac-Tôha 1996, Bini et al.
2001, Rossa et al. 2001, Azevedo & Bonecker 2003, Aoyagui & Bonecker 2004,
Bonecker et al. 2005, Leal 2010).
A turbidez da água está entre os fatores abióticos relacionados com as
densidades de alguns organismos zooplanctônicos, entretanto, há suspeitas de que esta
relação é influenciada por interações bióticas como a competição e a predação (Thorp &
Mantovani 2005). Sabe-se que as assembleias de peixes de pequeno porte compõem um
grupo ecológico diverso (Meschiatti et al. 2000, Agostinho et al. 2007) e grande parte
destes peixes tem como principal fonte de alimento o zooplâncton (Silva 2010). Diante
desse contexto, o presente estudo visa através de experimentação em microcosmos

136
testar a hipótese de que a predação de peixes em zooplâncton é menor em águas mais
túrbidas.
Material e métodos
Os experimentos foram realizados em microcosmos (aquários com15 litros de
água do rio filtrada em rede de plâncton) no mês de junho de 2013, utilizando-se dois
tratamentos de turbidez da água, um com baixa turbidez e outro com alta turbidez. Para
os tratamentos com alta turbidez foram adicionadas porções padronizas de argila
dissolvida em água. Foram realizadas três réplicas e um controle para cada tratamento.
Em cada réplica foram dispostos 10 peixes e todas as réplicas e controles foram
adicionadas subamostras padronizadas de zooplâncton (3750 indivíduos por réplica e
controle). Quatro horas depois da adição de zooplâncton, os peixes foram retirados dos
microcosmos e a água de todos os tratamentos e réplicas foi filtrada em rede de plâncton
de abertura de malha de 68 µm, para obtenção do zooplâncton. A predação em
zooplâncton por peixes foi estimada pela diferença inicial e final na densidade do
mesmo, corrigida pela sua mortalidade natural estimada através dos controles.
O zooplâncton foi obtido através de coleta com redes de plâncton de 68 µm no
ressaco do Pau Véio, na planície de inundação do alto rio Paraná. Os peixes também
foram obtidos por meio de coleta, utilizando-se rede de arrasto na margem esquerda do
alto rio Paraná. Antes do experimento os peixes coletados foram mantidos sob
aclimatação em jejum por 18 horas. Ao final do experimento, os peixes foram medidos
com auxílio de paquímetro e foram medidas as taxas de oxigênio dissolvido e
condutividade da água.
A relação entre o tamanho de peixes e a predação em zooplâncton foi verificada
por meio de uma regressão linear. Os resíduos dessa regressão foram utilizados como
variável resposta para se testar se a predação de peixes em zooplâncton é menor em
águas mais túrbidas através de um teste t.
Resultados
O comprimento médio dos peixes predadores foi de 37,2 e 38,0 mm nos
tratamentos com baixa e alta turbidez, respectivamente. O oxigênio dissolvido aferido
nos aquários não variou muito, com exceção do controle com alta turbidez e a
condutividade foi menor no controle com baixa turbidez (Tab. 1).
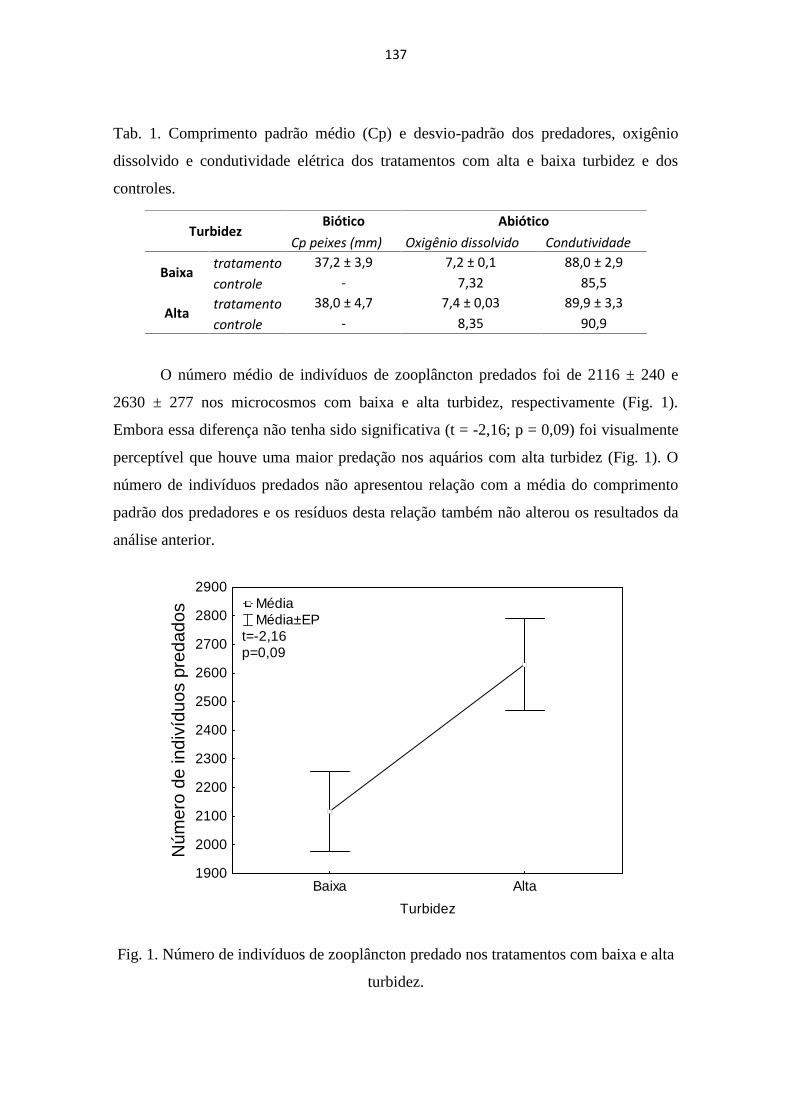
137
Tab. 1. Comprimento padrão médio (Cp) e desvio-padrão dos predadores, oxigênio
dissolvido e condutividade elétrica dos tratamentos com alta e baixa turbidez e dos
controles.
TurbidezBiótico Abiótico
Cp peixes (mm) Oxigênio dissolvido Condutividade
Baixa tratamento 37,2 ± 3,9 7,2 ± 0,1 88,0 ± 2,9controle - 7,32 85,5
Alta tratamento 38,0 ± 4,7 7,4 ± 0,03 89,9 ± 3,3controle - 8,35 90,9
O número médio de indivíduos de zooplâncton predados foi de 2116 ± 240 e
2630 ± 277 nos microcosmos com baixa e alta turbidez, respectivamente (Fig. 1).
Embora essa diferença não tenha sido significativa (t = -2,16; p = 0,09) foi visualmente
perceptível que houve uma maior predação nos aquários com alta turbidez (Fig. 1). O
número de indivíduos predados não apresentou relação com a média do comprimento
padrão dos predadores e os resíduos desta relação também não alterou os resultados da
análise anterior.
Fig. 1. Número de indivíduos de zooplâncton predado nos tratamentos com baixa e alta
turbidez.
Baixa AltaTurbidez
1900
2000
2100
2200
2300
2400
2500
2600
2700
2800
2900
Núm
ero
de in
diví
duos
pre
dado
s Média Média±EP
t=-2,16p=0,09

138
Discussão
Os resultados encontrados revelaram maiores predações nos tratamentos com
alta turbidez, não suportando a hipótese proposta no presente trabalho. Tais resultados
podem dever-se à exclusão do corpo de fundo dos microcosmos e a erros de
identificação, já que os tratamentos com alta turbidez possuíram maiores corpos de
fundo e que mais de um identificador triou as amostras. Apesar disso, os resultados
podem ser reflexos de padrões ainda desconhecidos, pois os efeitos da turbidez podem
envolver múltiplas relações bióticas. Por exemplo, a redução do risco de predação em
peixes em águas mais túrbidas pode levá-los a dispenderem menos esforços com
comportamentos anti-predatórios e, então, maiores esforços em alimentação (Gregory
1993). Considerando-se que nosso objeto de estudo são peixes planctívoros, a redução
da exposição à predação nos ambientes mais túrbidos pode ter levado a maiores taxas de
predação em zooplâncton.
A despeito do conhecimento de que águas túrbidas podem aumentar os esforços
alimentares de peixes, a confirmação de que os resultados encontrados estão
relacionados a isto só pode ser dada por novas experimentações. A criação de uma
padronização na identificação dos organismos zooplanctônicos e a consideração do
corpo de fundo dos microcosmos, também, se fazem necessárias para a confirmação de
resultados e o estabelecimento de padrões.
Referências
Agostinho AA, Gomes LC & Julio Jr. HF 2003. Relações entre macrófitas aquáticas e
fauna de peixes. In: Thomaz SM & Bini LM, eds. Ecologia e manejo de
macrófitas aquáticas. Maringá: EDUEM, pp. 261–277.
Aoyagui A, Bonecker CC 2004 Rotifers in different environments of the Upper Paraná
River floodplain (Brazil): richness, abundance and the relationship with
connectivity. Hydrobiologia 522: 281-290.
Azevedo F, Bonecker CC 2003 Community size structure of zooplanktonic assemblages
in three lakes on the upper River Parana floodplain, PR-MS, Brazil.
Hydrobiologia 505: 147-158.

139
Bini LM, Bonecker CC & Lansac-Tôha FA 2001 Vertical distribution of rotifers on the
upper Paraná River floodplain: the role of thermal stratification and chlorophyll-
a. Studies on Neotropical Fauna and Environment 36(3): 241-246.
Bonecker CC, Lansac-Tôha FA 1996 Community structure of rotifers in two
environments of the upper River Parana floodplain (MS) – Brazil. Hydrobiologia
325, p. 137-150.
Bonecker CC, Aoyagui ASM 2005 Relationships between rotifers, phytoplankyon and
bacterioplankton in the Corumbá reservoir, Goiás State, Brazil. Hydrobiologia
546: 415-421.
Gregory RS 1993 Effect of turbidity on the predator avoidance behavior of juvenile
chinook salmon (Oncorhynchus tshawytscha). Canadian Journal of Fisheries and
Aquatic Sciences 502: 241-246.
Leal CM 2010 Diversidade de espécies zooplanctônicas em uma planície de inundação
neotropical: influência da predação, das variáveis limnológicas ou da
disponibilidade de alimento? Dissertação de Mestrado do Programa de Pós-
Graduação em Ecologia de Ambientes Aquáticos Continentais, Universidade
Estadual de Maringá.
Meschiatti J, Arcifa, MS & Fenerich-Verani N, 2000. Fish communities associated with
macrophytes in Brazilian floodplain lakes. Environmental Biology of Fish 58:
133-143.
Morin P J, 1999. Community Ecology. Blackwell Science, Malden, MA.Rossa DC, Lansac-Tôha FA, Bonecker CC, Velho LFM 2001 Abundance of
cladocerans in the littoral regions of two environments of the upper Parana River
floodplain, Mato Grosso do Sul, Brazil. Revista Brasileira de Biologia 61: 45-53.
Sih, A., P. Crowley, M. McPeek, J. Petranka & K. Strohmeier, 1985. Predation,
competition and prey communities: a review of field experiments. Annual
Review of Ecology and Systematics 16: 269–311.
Silva GC, 2010. Peixes de pequeno porte associados à macrófitas aquáticas em uma
planície de inundação neotropical. Tese de doutorado do Programa de Pós-
Graduação em Ecologia de Ambientes Aquáticos Continentais, Universidade
Estadual de Maringá.
Thorp JH, Mantovani S 2005 Zooplankton of turbid and hydrologically dynamic prairie
rivers. Freshwater Biology 50: 1474–1491.

140
EFEITO DA INUNDAÇÃO NA CO-OCORRÊNCIA DE ESPÉCIES EM COMUNIDADES
VEGETAIS DA PLANÍCIE DE INUNDAÇÃO DO RIO PARANÁ
Érika Maria Neif, Francisco von Hartenthal, Guilherme Landgraf, Lilian Paula
Vasconcelos, Rafael A. Baggio, Sabrina Calado
Orientadora: Márcia Marques
Resumo
Foram testadas as hipóteses de que sob inundação, espécies vegetais apresentam
maior similaridade funcional; e que a distância filogenética entre as espécies vegetais
pode diminuir (atributos são conservados) ou não apresentar padrão (atributos são
distribuídos na filogenia das plantas). Para isso, foi comparada a composição
filogenética e de atributos (i.e. CAP) em 2 comunidades sob efeito ou não do estresse
causado por inundações periódicas. Nossos dados suportaram apenas a primeira
hipótese.
Palavras-chave: Inundações, filtro ambiental, filogenia.
Introdução
A diversidade funcional, definida por Tilman (2001) como "o valor e a variação
dos traços dos organismos e espécies que influenciam no funcionamento do
ecossistema", potencialmente proporciona maior poder explicativo do que a riqueza de
espécies, uma vez que relaciona os organismos às funções que eles mediam (Griffin et
al., 2009). Esta diversidade tem sido proposta como uma importante característica das
assembleias biológicas, permitindo a previsão das taxas dos processos dos ecossistemas
(Mason et al., 2005). Da mesma forma, aspectos filogenéticos têm sido crescentemente
utilizados em estudos de comunidades, visto que propiciam um aparato histórico para
determinar padrões e inferir processos evolutivos e ecológicos (Emerson & Gillespie,
2008).
Acredita-se que filtros ambientais determinem a similaridade funcional e as
relações filogenéticas na co-ocorrência de espécies de plantas (Silva & Batalha, 2010),
na medida que selecionam espécies que conseguem persistir em comunidades com base
na sua tolerância a condições abióticas (Weiher & Keddy, 1995).
A inundação de terras adjacentes em sistemas rio-planícies de inundação pode ser
considerada um filtro ambiental. Em agrupamentos florestais situados em solos que se

141
cobrem de água nesses períodos, desenvolvem-se apenas espécies tolerantes, formando
florestas abertas, constituídas de poucas espécies, quando comparadas a áreas mais
enxutas, que foram florestas mais densas e diversas (Campos & Souza,1997).
Assim, o objetivo deste trabalho foi verificar se a inundação modifica a co-
ocorrência de espécies vegetais na planície de inundação do alto rio Paraná, testando,
especificamente as seguintes hipóteses:
H1: Em locais sujeitos a inundação, espécies vegetais apresentam maior similaridade
funcional; H2: Em locais sujeitos a inundação, a distância filogenética entre as espécies
vegetais diminui (i.e. atributos são conservados) ou não diferem (i.e. atributos
relacionados a inundação são distribuídos na filogenia das plantas).
Materiais e Métodos
O estudo foi realizado na mata do Araldo, à margem esquerda do rio Paraná, no
município de Porto Rico, PR. Foram determinadas duas transecções de 150 metros, uma
na região mais próxima do rio e outra mais para o interior da mata. Em cada transecção
foram amostrados 15 pontos, divididos em 4 quadrantes, com uma distância de 10
metros entre eles. Em cada quadrante foi amostrada a planta que se encontrava mais
próxima, desde que possuísse mais de 20 cm de circunferência a altura do peito (CAP).
Foi calculada a proporção de co-ocorrência de pares de espécies em cada ponto
amostrado. Para se determinar a distância funcional entre as espécies, foi gerada uma
matriz de distância euclidiana da circunferência a altura do peito (CAP) dos pares de co-
ocorrência de espécies. A distância filogenética entre as espécies, baseada na APG 3, foi
calculada a partir do software Phylocom.
Para verificar se existe diferença na relação entre a distância funciona e a distância
filogenética com a co-ocorrência, entre as duas regiões amostradas (perto e longe do
rio), foram realizadas duas Análises de Covariância (ANCOVA). Nestas analises, as
distancias funcionais e as distâncias filogenéticas entre os pares de espécies foram
consideradas como variáveis contínuas independentes, a co-ocorrência dos pares de
espécies como variável resposta, e as duas regiões como variável categórica.
Resultados
As análises de covariância demonstram que o atributo funcional (i.e. CAP) afeta
a co-ocorrência das espécies (F1,378 = 6,06; P < 0,05, Fig. 1), enquanto a filogenia não
(F1,378 = 0,52; P > 0,05, Fig. 2).

142
Fig. 1. Relação entre as distâncias dos atributos (CAP) e co-ocorrência das espécies de plantasestudadas, para as regiões perto, afetada por inundações, e longe do Rio Paraná, não afetada porinundações.
Fig. 2. Relação entre as distâncias filogenéticas das espécies e co-ocorrência das espécies deplantas estudadas, para as regiões perto, afetada por inundações, e longe do Rio Paraná, nãoafetada por inundações.
Discussão
Nossos resultados indicam que a inundação modifica os padrões de co-
ocorrência das plantas. A inundação age como importante filtro ambiental, selecionando
espécies, homogeneizado o CAP entre as espécies que co-ocorrem. Em contrapartida, a
relação filogenética não afeta a co-ocorrência das espécies de plantas.
Dessa forma, os resultados sugerem que a composição florística da região
estudada se insere no modelo de dispersão filogenética e agrupamento fenotípico
apresentado por Emerson & Gillespie (2008). Nesse caso, filtros ambientais têm forte
influência sobre a composição das espécies, selecionando grupos funcionais específicos
(i.e. atributos funcionais em comum), e consequentemente sobre sua co-ocorrência.
-10 0 10 20 30 40 50 60 70 80
Distância de atributos (CAP)
-0,02
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
0,22
0,24
0,26
0,28
Co-
ocor
rênc
ia (L
e P
)
Longe Perto
-20 0 20 40 60 80 100Distância filogenética
-0,02
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
0,20
0,22
0,24
0,26
0,28
Co-o
corrê
ncia
(L e
P)
Longe Perto

143
Entretanto, tais características não demonstraram ser conservativas ao longo da
filogenia (i.e. conservação filogenética de nicho), ocorrendo variação no atributo
analisado (i.e. CAP) ao longo da filogenia, em resposta à ocupação do nicho.
Agradecimentos
Agradecemos ao “Tião” e ao Ricardo Augusto Camargo pelo apoio logístico
fundamental na coleta de dados e ao Vinícius Marcilio da Silva pelas contribuições no
delineamento e análise dos dados.
Referências
Campos JB, Souza MC 1997 Vegetação. Pages 331-342 in AEAM Vazzoler, AAAgostinho, NS Hahan, eds. A planície de inundação do alto rio Paraná: Aspectosfísicos, biológicos e socioeconômicos. Editora da Universidade Estadual deMaringá.
Emerson BC, Gillespie 2008 Phylogenetic analysis of community assembly andstructure over space and time. Trend in Ecology and Evolution 23(11): 619-630.
Griffin JN, Méndez V, Johnson AF, Jenkins SR, Foggo A 2009 Functional diversitypredicts overyielding effect of species combination on primary productivity. Oikos118: 37-44.
Mason NWH, Mouillot D, Lee WG, Wilson B 2005 Functional richness, functionalevenness and functional divergence: the primary components of functionaldiversity. Oikos 111: 112-118.
Silva IA, Batalha MA 2010 Woody plant species co-occurrence in Brazilian savannasunder different fire frequencies. Acta Oecologica 36: 85–91.
Tilman D 2001 Functional diversity. In: Levin, S. A. (ed.), Encyclopedia ofbiodiversity. Academic Press.
Weiher E, Keddy P 1995 The assembly of experi-mental wetland plant communities.Oikos 73:323–335.

144
HETEROGENEIDADE AMBIENTAL COMO PREDITOR DA DIVERSIDADE BETA EM
LAGOAS DE PLANÍCIE DE INUNDAÇÃO
Emanuel Razzolini; Fernando G. de Oliveira; Flávio M. Marteleto; Natália C. L. dos Santos;
Sandra M. N. de Andrade
Orientador: Prof. Dr. André
Padial
Resumo
A diversidade beta é um aspecto chave para entender sistemas naturais. O objetivo foi
identificar o papel da heterogeneidade ambiental na diversidade beta de macrofitas aquáticas
em lagoas de planície de inundação. A variação ambiental, considerando a profundidade das
lagoas, foi a que teve maior influência na diversidade beta. Considerando a variação entre
lagoas de um mesmo subsistema, a distância média do centroide da diversidade beta nas
lagoas do rio Baía é menor do que nas lagoas do rio Paraná, padrão que é acompanhado de
uma maior heterogeneidade ambiental e extensão espacial no rio Paraná.
Palavras-chave: escala espacial; riqueza; variação espacial.
Introdução
O estudo da variação é um aspecto chave para entender sistemas naturais e fazer
predições. Em ecologia de comunidades, a maioria dos estudos foca na diversidade local de
espécies (diversidade alfa), e, apenas recentemente, ecólogos passaram a dar atenção
apropriada a variação na composição de comunidades entre sítios amostrais (diversidade beta)
(Melo et al., 2011). A diversidade beta pode ser conceituada como a mudança na estrutura da
comunidade ao longo de um gradiente (turnover) ou a variação na estrutura da comunidade
entre as unidades de amostragem dentro de uma determinada área, sem referência a um
gradiente particular ou direção (Carvalho et al., 2012). Nesse contexto, o objetivo principal
desse estudo foi identificar os determinantes da diversidade em lagoas de planície de
inundação, tendo como objetivo específico investigar o efeito da heterogeneidade ambiental
de uma lagoa na variação da comunidade de macrófitas aquáticas desse ambiente. Em
decorrência disto, foi testada a hipótese de que há uma relação entre a variação das
macrófitas aquáticas com a heterogeneidade ambiental de lagoas de planície de inundação.

145
Dessa forma espera-se que quanto maior for a variação ambiental da lagoa maior será a
diversidade beta da mesma.
Material e Métodos
Situada na margem direita do rio Paraná, a Planície de inundação do Alto rio Paraná
(22o00’ - 23o00’S; 53o00’ - 53o45’ O) é caracterizada por elevada diversidade de habitats.
Alguns exemplos são amplas várzeas, ilhas, lagoas com diferentes graus de conectividade e
canais (Thomaz et al., 2004), exercendo, estes ambientes, um papel fundamental na e
manutenção da diversidade biológica aquática regional (Agostinho et al., 2001). Mesmo dentre
de um ambiente, como uma lagoa, há grande variação ambiental. Em vista dessa complexa
heterogeidade ambiental, foram amostradas 10 lagoas em dois subsistemas: seis lagoas no
subsistema do rio Baía e quatro no subsistema do rio Paraná.
Para testar a hipótese, foram delineados quatro pontos de amostragem dentro de
cada lagoa para obter uma estimativa da composição de macrofitas e heterogeneidade
ambiental. Desses quatro pontos, dois foram em um banco de macrofitas no fundo e outro na
entrada da lagoa. Em cada banco, no interior e na borda, foi contado o número de espécies
observado dentro de um quadrado de 50 cm2. Na borda dos bancos, acrescentou-se à
contagem as espécies capturadas por 3 lançamentos de garatéia. As variáveis preditoras da
heterogeneidade ambiental foram: profundidade (variação da profundidade média dos bancos
localizados no fundo e na entrada da lagoa), transparência da água, largura da lagoa e
declividade da margem (variação média das profundidades no interior e borda dos bancos). A
extensão espacial de cada subsistema foi obtida medindo a distância longitudinal entre os
locais amostrados mais distantes entre si através do programa Google Earth. A diversidade
beta entre bancos foi calculada através do índice de dissimilaridade de Jacard. A variação na
comunidade entre lagoas de um mesmo subsistema foi calculada através da distância média
para o centróide de uma dispersão de pontos gerada após uma análise de coordenadas
principais (PCoA) utilizando índice de Jacard. Nesse caso, as variáveis ambientais originais
também foram submetidas a uma PCoA com distância euclidiana para calcular a
heterogeneidade ambiental de cada subsitema. A extensão espacial de cada subsistema
também foi estimada ao mensurar a distância entre os dois pontos de coleta mais distantes.
Resultados

146
A correlação entre a diversidade beta dentro das lagoas com os coeficientes de
variação da declividade, largura e transparência não foi significativa (Fig. 1A, B e D). A melhor
correlação, com padrão não linear, foi entre a diversidade beta e a variação da profundidade
das lagoas (Fig. 1C).
Figura 1. Análise de correlação (pearson) entre a diversidade beta e os coeficientes de variação
dos fatores: declividade (A), largura (B) e profundidade da lagoa (C), e transparência da água
(D). r = coeficiente de correlação entre as variáveis; R2 = porcentagem de explicação da
regressão não linear; P = nível de significância; CVP = coeficiente de variação da profundidade;
CVT = coeficiente de variação da transparência; Para a variação da profundidade e
transparência foi gerado e testado o ajuste de um modelo não linear.
Em relação à diversidade beta entre os subsistemas rio Baía e Paraná, observou-se
uma evidente separação do centróide da diversidade beta entre esses subsistemas (Fig. 2) e
uma maior heterogeneidade dessa diversidade nas lagoas do rio Paraná em relação às do rio
Baía. Da mesma forma, a extensão espacial e os valores obtidos para a heterogeneidade
ambiental foram maiores para os ambientes do subsistema Paraná (14 km entre os pontos
mais distantes e distância média para o centróide da ordenação com dados ambientais de
3,02) do que do subsistema Baía (8,1 km entre os pontos mais distantes e distância média para
o centróide da ordenação com dados ambientais de 1,92).
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Coeficiente de variação (declividade)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Div
ersi
dade
bet
a (J
acca
rd)
r = -0.12; P = 0.78
A
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Coeficiente de variação (largura)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Div
ersi
dade
bet
a (J
acca
rd)
r = -0.119; P > 0.05
B
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Coeficiente de variação (profundidade)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Div
ersi
dade
bet
a (J
acca
rd)
r = 0.61; P = 0.05
β = 0.66+0.04*log(CVP) *P < 0.05; R2 = 18%
C
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Coeficiente de variação (transparência)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9D
iver
sida
de b
eta
(Jac
card
)
r = 0.242; P > 0.05
β = 0.81+0.32*log10(CVT) *NS
D

147
Figura 2. Análise de coordenadas principais mostrando a dispersão dos pontos amostrais e
suas e distâncias o centróide dos subsistemas dos rios Paraná e Baía.
Discussão
Devido aos valores insignificantes encontrados na correlação entre a diversidade beta
e as variáveis descritivas de heterogeneidade ambiental, podemos especular que existam
outras variáveis abióticas ou mesmo bióticas que podem explicar a distribuição dos
organismos no gradiente longitudinal das lagoas. De qualquer forma, parece haver um sinal da
heterogeneidade ambiental em termos de variação da profundidade com a diversidade beta
de lagoas de planície de inundação. Além disso, também pode-se especular que a estratégia
amostral podem não estar representando a totalidade das espécies existentes nas lagoas dos
subsistemas estudados.
É conhecido que a distância geográfica e as dissimilaridades ambientais têm efeitos
sobre os padrões globais da diversidade β (Quian & Ricklefs, 2012). A maior variação dessa
diversidade nas lagoas do rio Paraná parece ser explicada pela maior heterogeneidade
ambiental encontrada nesse subsistema. De acordo com Souza-Filho & Stevaux (2004), o rio
Paraná apresenta um padrão de multicanais apresentando uma ampla heterogeneidade de
intensidade de fluxo de água e formas ambientais (ressacos e lagoas de vários formatos e
comprimentos de extensão). Dessa forma, essa heterogeneidade ambiental promoveria uma
maior variação na composição de espécies uma vez que, Shmida & Wilson (1985), descrevem
as condições e disponibilidade de recursos nos diferentes habitats como determinantes da
coexistência e distribuição das espécies. Nesse contexto, era esperado que em ambientes mais
heterogêneos encontraríamos maiores indíces de diversidade beta.
-0.6 -0.4 -0.2 0.0 0.2 0.4
PCoA1
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
PC
oA2
Rio Paraná
Rio Baía

148
Referências Bibliográficas
Agostinho, A. A., L. C. Gomes & M. Zalewski. 2001. The importance of floodplains for the
dynamics of fish communities of the upper Paraná River. Ecohydrology Hydrobiology, 1:
209–217.
Carvalho, J.C.; Cardoso, P. & Gomes, P. 2012. Determining the relative roles of species
replacement and species richness differences in generating beta-diversity patterns. Global
Ecology Biogeography. 21: 760–771.
Melo, A.S., Schneck, F., Hepp, L.U., Simões, N.R., Siqueira, T., Bini, L.M. 2011. Focusing on
variation: methods and applications of the concept of beta diversity in aquatic ecosystems.
Acta Limnologica Brasiliensia. 3 (3): 318-331.
Qian, H. & Ricklefs, R. 2012. Disentangling the effects of geographic distance and
environmental dissimilarity on global patterns of species turnover. Global Ecology and
Biogeography. 21: 341-351.
Shmida, A. & M.V. Wilson. 1985. Biological determinants of species diversity. Journal of
Biogeography, 12:1-20.
Souza-Filho, E. E. & J. C. Stevaux. 2004. Geology and Geomorphology of the Baía-Curutuba-
Ivinheima river Complex. Pp. 75-102. In: Thomaz, S. M., A. A. Agostinho & N. S. Hahn (eds.).
The upper Paraná River and its floodplain: physical aspects, ecology and conservation.
Leiden, Backhuys Publishers.
Thomaz, S. M.; T. A. Pagioro, L. M. Bini, M. C. Roberto & R. R. A. Rocha. 2004. Limnological
Characterization of the aquatic environments and the influence of the hydrometric levels.
Pp. 75-102. In: Thomaz, S. M., A. A. Agostinho, N. S. Hahn (eds.). The upper Paraná River and
its floodplain: physical aspects, ecology and conservation. Leiden, Backhuys Publishers.

149
DIVERSIFICAÇÃO ECOMORFOLÓGICA EM ASSEMBLÉIAS DE PEIXES DO ALTO RIO
PARANÁ
Hugo José Message, Priscilla Guedes Gambale, Renata Gabriela Noguchi, Simone
Satsuki Takeuchi & Taise Miranda Lopes
Orientador: Luiz Carlos Gomes
Resumo: As variações na forma do corpo podem afetar a capacidade de natação, e são
bons indicativos das preferências de habitat. As hipóteses testadas foram 1) ambientes
com diferentes composições de espécies abrigam assembleias de peixes com formatos
diferentes; 2) morfologia de peixes difere entre ambientes. Observou-se que o
morfoespaço não difere claramente entre os ambientes amostrados, apesar de haver uma
tendência de diversificação morfológica para Hemigrammus marginatus e Moenkhausia
sp.
Palavra-chave: Ecomorfologia, macrófitas aquáticas, morfoespaço.
Introdução
As condições hidrológicas e geomorfológicas dos rios são muito variáveis e
dinâmicas, fornecendo diversos habitats para diferentes organismos aquáticos (Chuang
et al., 2006). A arquitetura das macrófitas aquáticas adiciona complexidade estrutural
para a região litorânea, o que possibilita maior diversidade e estabilidade das
comunidades bióticas (Dibble et al., 1996), dentre elas, as assembléias de peixes
(Agostinho et al., 2003). Estudos em ambientes de água doce neotropicais mostram os
peixes associados às macrófitas são tipicamente espécies de pequeno porte (Meschiatti
et al., 2000).
Os peixes apresentam uma das maiores diversidades morfológicas dentro dos
vertebrados. Ademais, a variação morfológica pode ser uma importante fonte de
respostas para indagações a respeito das relações entre morfologia e ecologia (Cunico &
Agostinho, 2006). Para muitas espécies a morfologia pode apresentar uma forte
correlação com a especialização do habitat, sendo que até mesmo uma sutil variação na
forma do corpo pode ser correlacionada com parâmetros de aptidão (Webster et al,
2011).
A forma do corpo, o comprimento do pedúnculo caudal, o tamanho e o formato
da nadadeira caudal, podem afetar o custo energético para o sustento da natação (Webb,
1982). Esses caracteres têm relação com as suas preferências de habitat para águas

150
lênticas ou lóticas (Langerhans et al, 2003). Um corpo cilíndrico que tem uma pequena
proporção de área de superfície para volume corporal com nadadeiras duras e curtas
favorece a existência em rios de rápido fluxo (Webb, 1984).
O objetivo do presente estudo foi comparar se existem diferenças quanto à
assembléia de peixes em duas áreas diferentes (Rio Baía e Ressaco do Bilé), como
também diferenças morfológicas nos peixes em relação aos ambientes. As hipóteses
foram que: 1) locais com diferenças na composição de espécies abrigam assembleias de
peixes com formatos diferentes; 2) morfologia de peixes difere em distintos ambientes.
Espera-se especialmente uma diferença na forma do corpo entre locais com e sem
colonização de plantas aquáticas estruturadoras do habitat.
Material e métodos
As amostragens foram realizadas no Rio Baía (3 pontos – macrófitas; 2 pontos –
sem macrófitas) e no ressaco do Bilé (3 pontos – macrófitas). A coleta dos peixes foi
realizada com o auxílio de uma rede de arrasto simples de 10 m de comprimento. Os
organismos capturados foram acondicionados em sacos plásticos e fixados com
formaldeído diluído.
Em laboratório foram identificadas as espécies e a abundância em cada ponto
amostrado. As variáveis ecomorfológicas medidas foram comprimento total,
comprimento padrão, altura do corpo e espessura com auxílio de paquímetro. Para 2
espécies que coexistem nos ambientes mais afastados do Rio Baia foram medidas
também altura do pedúnculo, comprimento anal e comprimento do pedúnculo.
Para descrever a organização espacial da composição de espécies entre os
ambientes foi realizada uma NMDS, através da medida de distância de Jaccard (Manly,
1994). E para as variáveis ecomorfológicas entre os locais e as espécies foi utilizada a
distância Euclidiana. Após as ordenações foram realizadas PERMANOVAs para
verificar a diferenças entre os fatores. A fim de diminuir a dimensionalidade dos dados
morfológicos de Moenkhausia sp. e Hemigrammus marginatus foi realizada uma análise
de componentes principais (PCA). Os escores da PCA foram testados através do teste-t
de student.
Resultados
A NMDS (Stress=0,0) separou grupos de espécies dos dois locais amostrados
(ressaco do Bilé e ambientes do rio Baía) (Figura 1A). O teste de PERMANOVA

151
confirmou diferenças na composição das assembleias de peixes entre estes ambientes
(Pseudo F = 3,52; P=0,02). Também com uma NMDS baseado na composição de
espécies (Stress=0,0), grupos de espécies distintos foram observados entre três locais
amostrados (lagoas abertas e fechadas no rio Baía, e o ressaco do Bilé no rio Paraná)
(Figura 1B). O teste de PERMANOVA confirmou diferenças na composição das
assembleias de peixes entre estes ambientes (Pseudo F= 2,86; P=0,007).
A
B
Figura 1: Gráfico de ordenação por NMDS baseada no índice de Jaccard (dados de
presença e ausência) mostrando a dissimilaridade da composição das assembleias de
peixes entre os locais do rio Baía e ressaco do Bilé e entre lagoas abertas e fechadas no
rio Baía, e o ressaco do Bilé no rio Paraná (Stress=0,0).
A NMDS considerando variáveis ecomorfológicas das espécies não apresentou a
formação de grupos definidos. No entanto, houve uma tendência de separação entre as
espécies de Hemigrammus marginatus e Moenkhausia sp. do ponto 3 (ambiente do Rio
Baía com colonização de Macrófitas próximo ao rio Paraná) em relação aos pontos 1 e 2
(ambientes do Rio Baía com colonização de Macrófitas distante do rio Paraná). O teste
de PERMANOVA confirmou diferenças na ecomorfologia das espécies para os locais
acima (Pseudo F = 6,91; P<0,01), para as espécies (Pseudo F = 7,57; P<0,01) e para
interação dos fatores (Pseudo F = 5,16; P<0,01).
A ordenação resultante da aplicação da Análise de Componentes Principais
(PCA) para Hemigrammus marginatus, resultou na retenção dos dois primeiros eixos
para a interpretação da variabilidade dos dados, sendo que o eixo 1 representou 47,74%
de explicação e o eixo 2 representou 19,37% (Figura 2A). A ordenação resultante da
aplicação da Análise de Componentes Principais (PCA) para Moenkhausia sp., resultou
na retenção dos dois primeiros eixos para a interpretação da variabilidade dos dados,
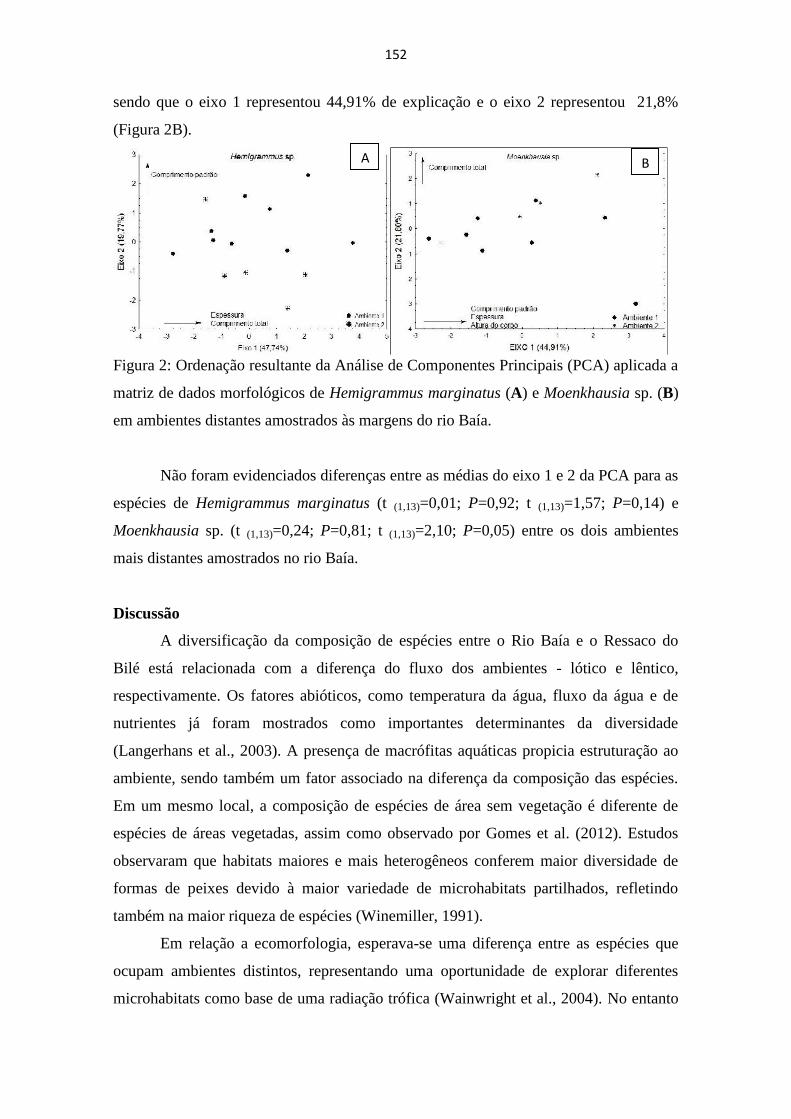
152
sendo que o eixo 1 representou 44,91% de explicação e o eixo 2 representou 21,8%
(Figura 2B).
Figura 2: Ordenação resultante da Análise de Componentes Principais (PCA) aplicada a
matriz de dados morfológicos de Hemigrammus marginatus (A) e Moenkhausia sp. (B)
em ambientes distantes amostrados às margens do rio Baía.
Não foram evidenciados diferenças entre as médias do eixo 1 e 2 da PCA para as
espécies de Hemigrammus marginatus (t (1,13)=0,01; P=0,92; t (1,13)=1,57; P=0,14) e
Moenkhausia sp. (t (1,13)=0,24; P=0,81; t (1,13)=2,10; P=0,05) entre os dois ambientes
mais distantes amostrados no rio Baía.
Discussão
A diversificação da composição de espécies entre o Rio Baía e o Ressaco do
Bilé está relacionada com a diferença do fluxo dos ambientes - lótico e lêntico,
respectivamente. Os fatores abióticos, como temperatura da água, fluxo da água e de
nutrientes já foram mostrados como importantes determinantes da diversidade
(Langerhans et al., 2003). A presença de macrófitas aquáticas propicia estruturação ao
ambiente, sendo também um fator associado na diferença da composição das espécies.
Em um mesmo local, a composição de espécies de área sem vegetação é diferente de
espécies de áreas vegetadas, assim como observado por Gomes et al. (2012). Estudos
observaram que habitats maiores e mais heterogêneos conferem maior diversidade de
formas de peixes devido à maior variedade de microhabitats partilhados, refletindo
também na maior riqueza de espécies (Winemiller, 1991).
Em relação a ecomorfologia, esperava-se uma diferença entre as espécies que
ocupam ambientes distintos, representando uma oportunidade de explorar diferentes
microhabitats como base de uma radiação trófica (Wainwright et al., 2004). No entanto
A B

153
o presente estudo evidenciou apenas uma tendência da diversificação ecomorfológica de
uma mesma espécie (Moenkhausia sp. e Hemigrammus marginatus) em relação
ambientes distintos. Para uma melhor representação destas hipóteses deve ser necessário
um maior esforço amostral e uma maior diferença entre os pontos analisados.
Agradecimentos
Agradecemos ao professor Luiz Carlos Gomes pela orientação do projeto, Alfredo, Tato
e João Carlos pelas ajudas na coleta. Agradecemos também aos organizadores e
professores do Curso de Ecologia de Campo.
Referências bibliográficas
Agostinho AA, Gomes LC, Júlio Jr HF 2003 Relações entre macrófitas aquáticas e
fauna de peixes. Pages: 261–280 in Thomaz SM, Bini LM (eds) Ecologia e
Manejo de Macrófitas Aquáticas. Eduem, Maringá, Brasil.
Chuang L-C, Lin Y-S, Liang S-H 2006 Ecomorphological Comparison and Habitat
Preference of 2 Cyprinid Fishes, Varicorhinus barbatulus and Candidia
barbatus, in Hapen Creek of Northern Taiwan Zoological Studies 45(1): 114-
123.
Cunico AM, Agostinho AA 2006 Morphological patterns of fish and their relationships
with reservoirs hydrodynamics. Braz. arch. biol. technol 49 (1): 125-134.
Dibble ED, Killgore KJ, Harrel SL 1996 Assessment of fish–plant interactions. Pages:
357–372 in Miranda E, DeVries D (eds) Multidimensional approaches to
reservoir fisheries management. American Fisheries Society, Bethesda,
Maryland, USA.
Langerhans RB, Layman CA, Langerhans AK, Dewitt TJ 2003 Habitat-associated
morphological divergence in two Neotropical fish species. Biol. J. Linn. Soc. 80:
689-698.
Meschiatti AJ, Arcifa MS, Fenerich-Verani N 2000 Fish communities associated with
macrophytes in Brazilian floodplain lakes. Environ Biol Fish 58:133–143
Webb PW 1982 Locomotor patterns in the evolution of Actinopterygians fishes. Amer.
Zool. 22: 329–342.
Webb PW 1984 Body form, locomotion and foraging in aquatic vertebrates. American
Zoologist 24:107-120.

154
Webster MM, Atton N, Hart PJB, Ward AJW 2011 Habitat-Specific Morphological
Variation among Threespine Sticklebacks (Gasterosteus aculeatus) within a
Drainage Basin. PLoS ONE 6(6): 1-10.





























