Embed Size (px)
Citation preview

Biochemical Systamatics and Ecology, Vol. 21, No. 5, pp. 597-605, 1993. 0305-1978/93 $6.00 + 0.00 Printed in Great Britain. © 1993 Pergamon Press Ltd.
Ecogeographical Distribution and Thermostability of Isocitrate Dehydrogenase (IDH) AIIoenzymes in European Silver Fir (Abies alba)
F. BERGMANN and H. R. GREGORIUS Abteilung fer Forstgenetik und ForstpflanzenzLichtung, Universit~t G~ttingen, B(]sgenweg 2, 3400 G6ttingen,
Germany
Key Word Index--Clinal variation; enzyme polymorphism; thermostability; norm of reaction; climate; adaptation; silver fir.
Abstrect--A latitudinal frequency cline observed for the two alleles B~ and B 2 at the IDH-B enzyme locus in European silver fir suggested the existence of a correlation between the associated north-south temperature gradient and the thermostability of the two alloenzymes. B~ prevailed in the northern and B 2 in the southern populations. Thermostability of an enzyme is defined by the length of time the enzyme can endure a given temperature without suffering damage (denaturation) that reduces its catalytic efficiency below a specified threshold; the threshold is specified by the visibility of activity in the zymogram. Using extracts from endosperms of homozygous trees incubated under various temperatures and followed by electrophoresis, it turned out that the B2 distinctly exceeds the B1 alloenzyme in thermostability in trees from both extremes of the fir transect. Supported by observations made in other species, it is concluded that catalytic activity and thermostability are negatively correlated for this enzyme locus. Based on this relationship, the observed IDH polymorphism is argued to be the result of adaptation to high temperature regimes.
Introduction Forest tree species generally possess a relatively high degree of genetic variation as compared to other plant and animal species, if various isoenzyme-gene systems are considered (Mitton, 1983; Ledig, 1986; EI-Kassaby, 1991). Most of the variation of this particular class of structural gene loci (coding for soluble enzymes) was often found to reside within populations, while only a minor proportion of variation is distributed among populations (for review, see Ledig, 1986; EI-Kassaby, 1991). Although there are few exceptions to this rule, the generally small genetic differentiation among populations has led to a particular viewpoint concerning the generation and maintenance of isoenzyme polymorphisms in tree species (Gregorius and Bergmann, submitted). They concluded that polymorphisms, especially major polymorphisms (with two or more prevalent alleles at an enzyme locus), are maintained by hetero- zygote advantage due to ontogenetic (or possibly stable temporal environmental) differentiation in enzyme function, while differences between populations in the frequencies of their prevalent alleles are due to adaptive differentiation for the corresponding homozygotes. The prevalent alleles are largely the same for all populations as a consequence of evolutionary optimization in function.
Allele frequency clines at enzyme loci along ecogeographical (e.g. climatic) gradients may result from two different processes: (a) gene flow between previously isolated populations possessing different prevalent alleles at a gene locus; or (b) adaptation of enzyme phenotype (genotype) polymorphisms to gradually varying environmental factors. These factors should selectively discriminate between physiologically different phenotypes, which are based on kinetically different alloenzyme composition.
A major finding in the above-cited work (Gregorius and Bergmann, submitted) shows cause (a) to be an unrealistic explanation of alloenzyme clines. Moreover, this
(Received 8 December 1992)
597

598 F. BERGMANN AND H. R. GREGORIUS
paper favours cause (b) with the restriction that such clinal variation patterns should be governed by some type of overdominance where the homozygotes are selected differently across the species' transect depending on the corresponding values of the critical environmental factors. Prerequisites for fitness (viability) differences between the homozygotes at an enzyme locus are differences in biochemical and/or biophysical properties (specific activity, Vma x, thermostability) of the respective alloenzymes. Hence, a causal analysis of allele frequency clines should not be solely based on the description of variation patterns but should, if possible, be substantiated by biochemical/biophysical characterizations of the corresponding alloenzyme types.
The objective of the present study is to interrelate the data obtained on the eco- geographical distribution of alleles with those on the degree of thermostability of their alloenzymes at an isocitrate dehydrogenase (IDH) locus in European silver fir. The relationships observed between the climatic conditions of various fir areas and the degree of thermostability of the prevalent alloenzyme in these areas suggest that natural selection acting on thermostability predominantly governs the eco- geographical variation pattern of this IDH polymorphism.
Materials and Methods Seed-lots were sampled in 45 silver fir populations located in seven countries and covering a great part of the European silver fir range, in order to study the geographic variation pattern of this conifer species at various enzyme gene loci (for further details, see Bergmann et aL, 1990). Haploid endosperm tissue of single seeds (representing individual macrogametes) was used as test material so that the allele frequencies could be estimated from sufficiently large seed samples (ca 100 seeds) per population (Feret and Bergmann, 1976).
For isoenzyme analysis, the endosperm tissue was dissected from single dormant seeds and homogenized with a specific extraction buffer [0.02 M Tris-HCI (pH 7.2) containing 10 mg m1-1 EDTA, a few drops of mercaptoethanol and 5 mg m1-1 Polyclar AT]. The crude homogenates were subjected to horizontal starch gel zone-electrophoresis, using a 0.2 M Tris-citrate buffer system (pH 7.0) for resolution of the isocitrate dehydrogenase (IDH) isoenzymes. Following electrophoretic separation, the IDH activity zones in the gel slabs were stained with a mixture composed of 100 ml 0.02 M Tris-HCI (pH 9.0), 150 mg DL-isocitrate (trisodium salt), 20 mg NADP, 20 mg MTT, 2 mg PMS and a few drops of 10% MgCI 2.
To test the thermostability of individual alloenzymes, aliquots of crude homogenates of single seed endosperms collected from homozygous trees (so that only one electrophorically well-defined alloenzyme underwent the treatment) were subjected to different temperature incubation prior to etectrophoretic separation. In order to exclude physiological effects of the maternal tree and biochemical effects of the genetic background on possible thermostability differences between endosperm homogenates, seeds for testing were collected from each of four trees homozygous for B1 (IDH-BIB 1) and four trees homozygous for B 2 (IDH-B~B2). Two trees of each of the two homozygotes are located in the northern Black Forest and two trees of each in Southern Calabria, so that B1- and B2-alloenzymes from seed endosperms of trees growing at opposite ends of one silver fir transect and hence exposed to completely different climatic conditions were treated in the same manner.
The homogenate probes were heated to 50, 60 or 70°C in an incubator, and each heat treatment was stopped after varying lengths of time (5, 10, 20, 40 min). The enzymes were thus observed under environmental conditions differing for two components, temperature and duration of exposure. Following a short cooling, the homogenates were soaked into filter paper wicks, which were then applied to the starch gel for subsequent electrophoresis.
Results and Discussion The IDH system (NADP-dependent IDH, E.C. 1.1.1.42) electrophoretically analysed in silver fir consists of two activity zones encoded by two distinct gene loci (IDH-A, IDH-B: Moiler, 1986; Bergmann and Kownatzki, 1988; Bergmann et al., in prep.). Both IDH loci are polymorphic in all fir populations studied so far (Bergmann et al., 1990). However, the IDH-B locus, which generally possesses two alleles (B1, B2), was found to show a geographically dependent variation pattern indicating latitudinally varying allele frequencies (Bergmann and Kownatzki, 1988).
Latitudinal variation in allele frequencies In the present study, additional population samples were analysed, especially from
marginal locations of the species' range. The resulting allele frequency distributions
DISTRIBUTION AND THERMOSTABILITY OF ALLOENZYMES IN SILVER FIR 599
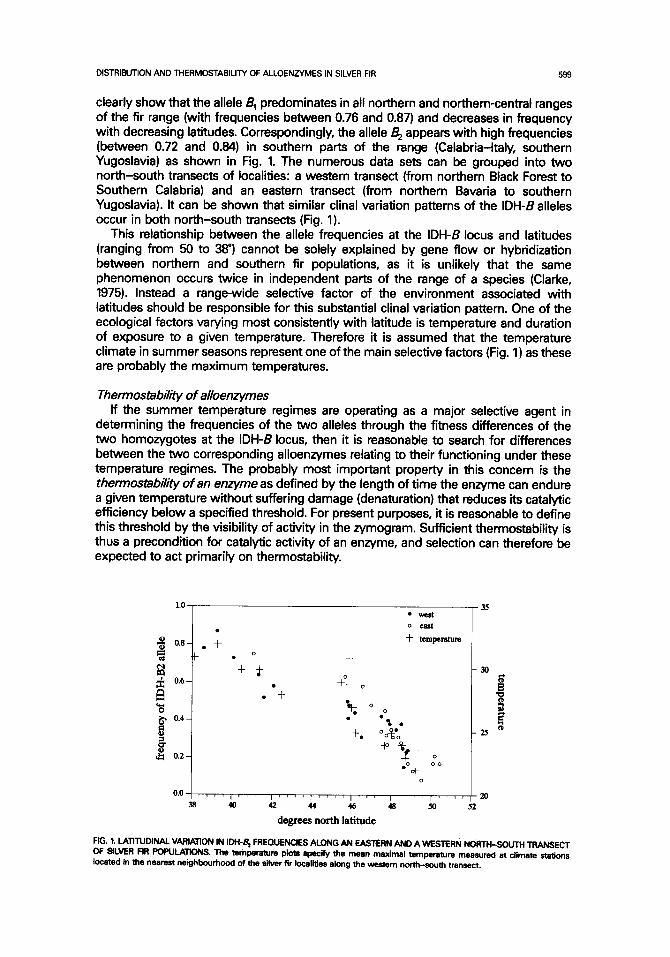
clearly show that the allele B 1 predominates in all northern and northern-central ranges of the fir range (with frequencies between 0.76 and 0.87) and decreases in frequency with decreasing latitudes. Correspondingly, the allele B 2 appears with high frequencies (between 0.72 and 0.84) in southern parts of the range (Calabria-Italy, southern Yugoslavia) as shown in Fig. 1. The numerous data sets can be grouped into two north-south transects of localities: a western transect (from northern Black Forest to Southern Calabria) and an eastern transect (from northern Bavaria to southern Yugoslavia). It can be shown that similar clinal variation patterns of the IDH-B alleles occur in both north-south transects (Fig. 1).
This relationship between the allele frequencies at the IDH-B locus and latitudes (ranging from 50 to 38 °) cannot be solely explained by gene flow or hybridization between northern and southern fir populations, as it is unlikely that the same phenomenon occurs twice in independent parts of the range of a species (Clarke, 1975). Instead a range-wide selective factor of the environment associated with latitudes should be responsible for this substantial clinal variation pattern. One of the ecological factors varying most consistently with latitude is temperature and duration of exposure to a given temperature. Therefore it is assumed that the temperature climate in summer seasons represent one of the main selective factors (Fig. 1) as these are probably the maximum temperatures.
Thermostability of alloenzymes If the summer temperature regimes are operating as a major selective agent in
determining the frequencies of the two alleles through the fitness differences of the two homozygotes at the IDH-B locus, then it is reasonable to search for differences between the two corresponding alloenzymes relating to their functioning under these temperature regimes. The probably most important property in this concern is the thermostability of an enzyme as defined by the length of time the enzyme can endure a given temperature without suffering damage (denaturation) that reduces its catalytic efficiency below a specified threshold. For present purposes, it is reasonable to define this threshold by the visibility of activity in the zymogram. Sufficient thermostability is thus a precondition for catalytic activity of an enzyme, and selection can therefore be expected to act primarily on thermostability.
1.0
• ~ 0 .8 -
¢=
m ~3~ 0.6-
~, 0 .4 -
0.2-
0.0
• west o eBt
• ÷ ÷ temperature
o
• ÷ o
÷,
o e °
o •
"d- o
35
3O
25
. . . . I . . . . I . . . . ~ . . . . I . . . . I . . . . I . . . . 20 38 40 42 44 46 48 50 52
degrees north latitude
FIG. t. LATITUDINAL VARIATION IN IDH-B 2 FREQUENCIES ALONG AN EASTERN AND A WESTERN NORIH-SOUTH TRANSECT OF SILVER FIR POPULATIONS. The temperature plots specify the mean maximal temperature measured at climate stations located in the nearest neighbourhood of the silver fir localities along the western north-south transect.

600 F. BERGMANN AND H. R. GREGORIUS
Thermostability and catalytic activi~ The present measurement of thermostability is based on the observation of catalytic activity of enzymes in terms of colour intensities of bands in zymograms. Clearly, this observation depends on the amount of enzyme entering the homogenates, and this, in turn, is determined by the enzyme content in the endosperm at the time when the seed is harvested. Hence, the measurement of thermostability could be biased by enzyme contents varying between endosperms as a result of environmentally or genetically induced differential regulation of the activity of the gene coding for the enzyme as well as of differing endosperm sizes. At least in the present study this possibility can, however, be considered negligible, since all zymograms obtained under the standard conditions showed no such variation in colour intensity of the respective IDH bands. One possible explanation is the absence of effective genetic variation at the regulatory loci. Another explanation could be stage- specific activity of the structural genes combined with the inability of the environment to modify this activity to an extent discernible under the applied staining method.
Although it encompasses catalytic activity, thermostability in the above sense has to be distinguished from the ordinary measurements of catalytic activity of an enzyme. It may in fact be largely independent of such measurements of catalytic activity. The catalytic activity at lower temperatures may be higher for an enzyme with low (thermosensitive) thermostability than for an enzyme with high (thermoresistant) thermostability. It is indeed easily conceivable that the molecular stability of an enzyme is negatively correlated with its catalytic efficiency. Examples for this negative correlation between thermostability and catalytic efficiency at moderate temperatures are provided by the observations on alcohol dehydrogenase alloenzymes in Drosophila (van Delden, 1982) and on phosphoglucose isomerase alloenzymes in Colias butterflies (Watt, 1977, 1983).
Norms of reaction for thermostabili~ The preceding considerations justify the assumption that our results on thermostability communicated below are characteristic of the respective enzyme and its coding gene. A method of analysis particularly suited to ecologically genetic studies, to which the present study can be counted, is provided by the concept of norm of reaction (or response function). In this concept the enzyme (or its coding gene) is considered as a system that responds to certain stimuli, where response and stimulus are measured in terms of a specified trait (thermostability) and a causal variable (incubation temperature), respectively. Ideally, the enzyme is then characterized by a mapping assigning to each temperature value (stimulus or cause) a thermostability value (response measured as the time until cessation of activity). The resulting graph can generally be expected to represent a decreasing function of temperature with the possibility of very large (probably infinite) values for low temperatures, since practically no denaturation of the enzyme takes place there.
Therefore, for high temperatures where denaturation is accelerated, catalytic activity is predominantly determined by thermostability, so that selective differences between alloenzymes may heavily depend on their degrees of thermostability. In contrast, for low temperatures thermostability is probably irrelevant in this context, since the time until denaturation becomes apparent by its effect on the catalytic activity is too long. Hence, for studies of adaptive significance of alloenzymes it might even be sufficient to concentrate on measurements of thermostability for a single temperature at which one alloenzyme is still active while the other has lost its activity.
Therefore, when trying to relate thermostabilities differing between alloenzymes to the distributions of their corresponding alleles across an ecogeographical gradient, thermostability values for lower temperatures are probably irrelevant, since they are too large to ever become a limiting factor in the catalytic efficiency for any of the involved enzymes.
Observed thermostabilities at the IDH-B locus. Based on the above deliberations, thermostability of the alloenzymes encoded by the alleles B 1 (Bl-alloenzyme) and B 2
601
Control 20 rain heat ing 40 rain heating
B1 B2 B1 B2 B1 B2 B1 B2
BI. Forest Ca labr ia BI. Forest Calabr ia
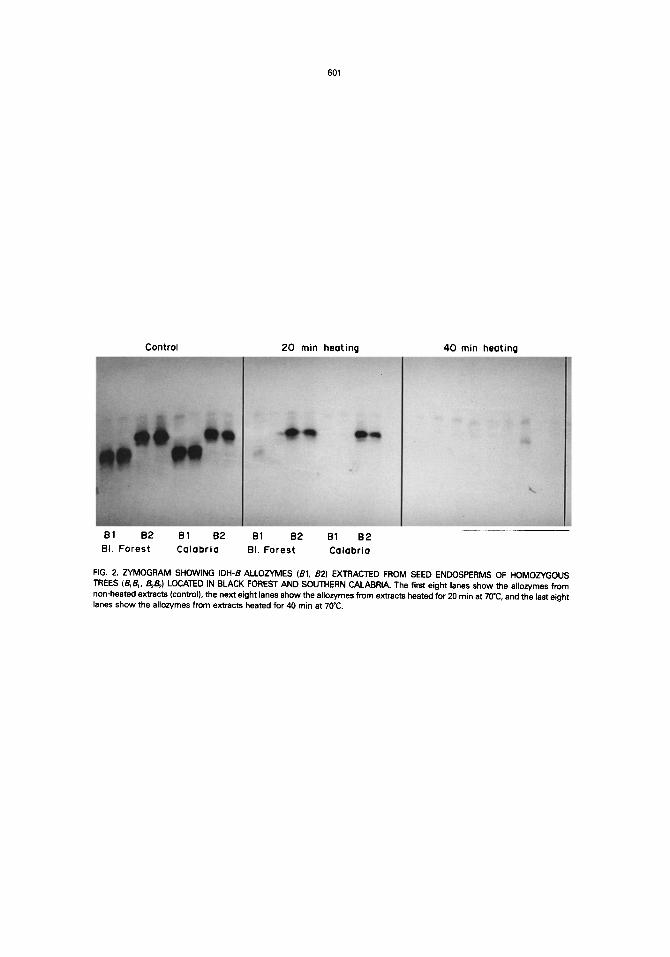
FIG. 2. ZYMOGRAM SHOWING IDH-B ALLOZYMES (81, 82) EXTRACTED FROM SEED ENDOSPERMS OF HOMOZYGOUS TREES (B~B~, B2B2) LOCATED IN BLACK FOREST AND SOUTHERN CALABRIA. The first eight lanes show the allozymes from non-heated extracts (control), the next eight lanes show the allozymes from extracts heated for 20 rain at 70°C, and the last eight lanes show the allozymes from extracts heated for 40 rain at 70°C.

DISTRIBUTION AND THERMOSTABILITY OF ALLOENZYMES IN SILVER FIR 603
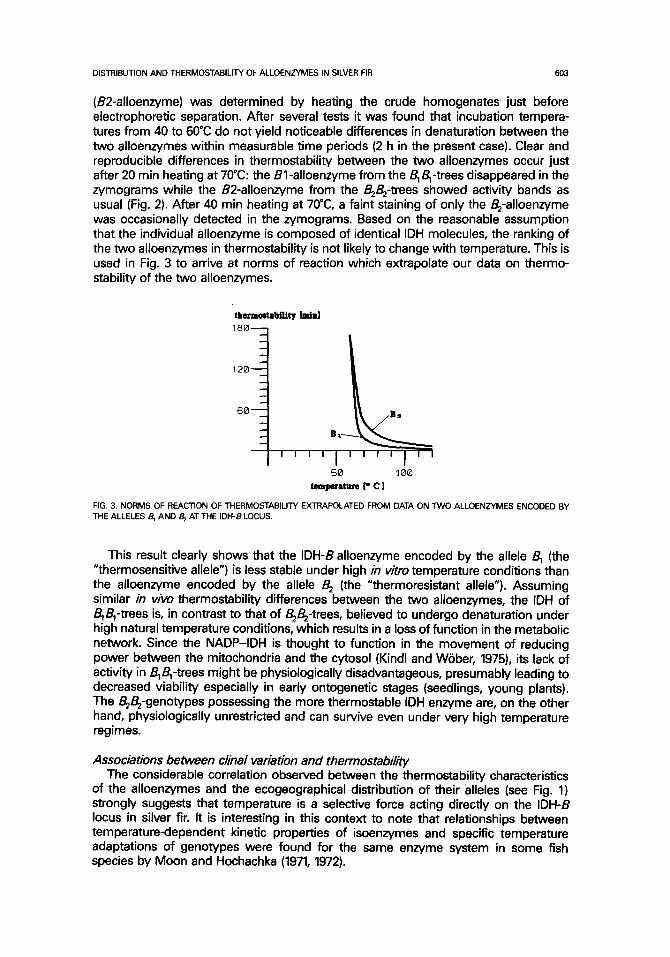
(B2-alloenzyme) was determined by heating the crude homogenates just before electrophoretic separation. After several tests it was found that incubation tempera- tures from 40 to 60°C do not yield noticeable differences in denaturation between the two alloenzymes within measurable time periods (2 h in the present case). Clear and reproducible differences in thermostabiiity between the two alloenzymes occur just after 20 min heating at 70°C: the Bl-alloenzyme from the B~B~-trees disappeared in the zymograms while the B2-alloenzyrne from the B;B2-trees showed activity bands as usual (Fig. 2). After 40 rain heating at 70°C, a faint staining of only the B2-alloenzyme was occasionally detected in the zymograms. Based on the reasonable assumption that the individual alloenzyme is composed of identical IDH molecules, the ranking of the two alloenzymes in thermostability is not likely to change with temperature. This is used in Fig. 3 to arrive at norms of reaction which extrapolate our data on thermo- stability of the two alloenzymes.
t~ermoaabili¢t Imtnl
1 8 0 ~
120.....~-
60"-
I I I I J I I I I J I I
5@ I@@
temlmramte l ~ C ]
FIG. 3. NORMS OF REACTION OF THERMOSTABILrrY EXTRAPOLATED FROM DATA ON TWO ALLOENZYMES ENCODED BY THE ALLELES B, AND B 2 AT THE IDH-B LOCUS.
This result clearly shows that the IDH-B alloenzyme encoded by the allele B~ (the "thermosensitive allele") is less stable under high in vitro temperature conditions than the alloenzyme encoded by the allele B 2 (the "thermoresistant allele"). Assuming similar in vivo thermostability differences between the two alioenzymes, the IDH of BTBl-trees is, in contrast to that of B2B2-trees, believed to undergo denaturation under high natural temperature conditions, which results in a loss of function in the metabolic network. Since the NADP-IDH is thought to function in the movement of reducing power between the mitochondria and the cytosol (Kindl and W0ber, 1975), its lack of activity in B1B,-trees might be physiologically disadvantageous, presumably leading to decreased viability especially in early ontogenetic stages (seedlings, young plants). The B2B2-genotypes possessing the more thermostable IDH enzyme are, on the other hand, physiologically unrestricted and can survive even under very high temperature regimes.
Associations between clinal variation and thermostability The considerable correlation observed between the thermostability characteristics
of the alloenzymes and the ecogeographical distribution of their alleles (see Fig. 1) strongly suggests that temperature is a selective force acting directly on the IDH-B locus in silver fir. It is interesting in this context to note that relationships between temperature-dependent kinetic properties of isoenzymes and specific temperature adaptations of genotypes were found for the same enzyme system in some fish species by Moon and Hochachka (1971, 1972).

604 F. BERGMANN AND H. R. GREGORIUS
The possible causal relationship between higher degrees of thermostability of the B2-alloenzyme in vitro and the higher frequency of occurrence of its allele (IDH-B=) in southern fir ranges does, however, not suffice to explain the lower frequency of occurrence of this allele in northern and central fir populations. If thermoresistance would not imply a disadvantage as compared with thermosensitivity under lower temperatures at least in the northern ranges, allele B 2 would be expected to be fixed in all populations. Hence, to find a plausible cause for the reversed allele frequency distribution in all northern populations, it must additionally be assumed that the thermosensitive alloenzyme (B1) is on the average kinetically superior in these popula- tions to the thermoresistant alloenzyme (B2), as was explicitly described in the above subsection on thermostability and catalytic activity. This higher catalytic efficiency may possibly result in faster seed germination, seedling development or tree growth, so that the IDH-B1B~ genotype has higher fitness than the IDH-B2B 2 genotype (but still below the heterozygote) in the northern ranges.
Based on the relationships between the degree of thermostabilty and catalytic efficiency of alloenzymes on the one hand and the prevailing temperature regime in the respective area on the other hand, the forces balancing the IDH polymorphism across the silver fir range require more consideration. The coexistence of alleles coding for thermosensitive and thermoresistant alloenzymes implies that different temperature regimes may vary both spatially and temporally within the fir populations. In areas or times of high temperature, the thermosensitive alloenzyme would denature more readily and thus be inferior in activity to the thermoresistant alloenzyme. In the reverse situation where temperature is low, the higher catalytic efficiency of the thermosensitive alloenzyme guarantees its superiority over the thermoresistant alloenzyme. Consequently, the frequency of the corresponding alleles could be directly correlated with the spatio-temporal distribution of temperature in the population: the larger the frequency of high temperature conditions in a population, the larger the frequency of the allele coding for the thermoresistant alloenzyme and vice versa.
These correlations may thus be responsible for the clinal variation of the IDH-B alleles from northern to southern ranges of silver fir, since the frequency of high temperature regimes, on the average, gradually increases with decreasing latitude. This general pattern may, of course, be modified in limited geographic areas, if, for instance, populations from higher and lower elevations, from northern or southern slopes of a hill (Bergmann, unpublished data), or from northeastern and southwestern areas (SchrSder, 1989) were compared. In these cases allele frequency changes reflect different frequency distributions of high temperature regimes similar to those of both range-wide north-south transects of our study.
The IDH polymorphism is unlikely to solely determine the overall fitness of fir seedlings or plants under different temperature regimes. Presumably an assembly of many enzymes similarly varying in thermostability should influence the viability of this tree species of which IDH-B alloenzymes may play an indicator role. Furthermore, taking into account the dispersal of silver fir after the latest glacial period, it is conceivable that the mutation event leading to the thermoresistant allele had occurred in later evolutionary epochs of silver fir and has expanded only in those areas which were exposed to extremely high temperature climates. Additional studies on this subject, including determination of kinetic parameters of the two alloenzymes, may uncover further relationships of silver fir adaptation.
References Bergmann, F., Gregorius, H.-R. and Larsen, J. B. (1990) Levels of genetic variation in European silver fir (Abies
alba). Are they related to the species' decline? Genetica 82, 1-10.

DISTRIBUTION AND THERMOSTABILITY OF ALLOENZYMES IN SILVER FIR 605
Bergmann, F. and Kownatzki, D. (1988) The genetic variation pattern of silver fir (Abies alba Mill.) in Europe monitored from enzyme gene loci. In Abhandl. 5. IUFRO Tannensymposium, Zvolen (Paule, L. and Korpel, S., eds), pp. 21-26.
Clarke, B. (1975) The contribution of ecological genetics to evolutionary theory: Detecting the direct effects of natural selection on particular potymorphic loci. GeneEcs 79, 101-113.
EI-Kassaby, Y. A. (1991) Genetic variation within and among conifer populations: Review and evaluation of methods. In Biochemical Markers in the Population Genedcs of Forest Trees (Fineschi, S., Malvolti, M. E., Cannata, F. and Hattemer, H. H., eds), pp. 61-76. SPB Academic Publ. bv The Hague.
Feret, P. P. and Bergmann, F. (1976) Gel electrophoresis of proteins and enzymes. In Modern Methods in Forest Genedcs (Miksche, J. P., ed.), pp. 49-77. Springer-Verlag.
Gregorius, H.-R. and Bergmann, F. (Submitted) Analysis of isoenzyme genetic profiles observed in forest tree populations. Hereditas.
Kindl, H. and WSber, F. (1975) Biochemie derPflanzen. Springer-Verlag. Ledig, F. T. (1986) Heterozygosity, heterosis, and fitness in outbreeding plants. In Conservation Biology (Soul0,
M., ed.), pp. 77-104. Sinauer Associates. Mitton, J. B. (1983) Conifers. In Isozymes in Plant Breeding and Genetics (Tanksley, S. D. and Orton, T. J., eds),
pp. 443-472. Elsevier, Amsterdam. Moiler, K. (1986) Genetische Untersuchengen bei der Tanne mit Hilfe von Enzyme-Genmarkern. AIIg. Forstztg.
97, 60-61. Moon, T. W. and Hochachka, P. W. (1971) Temperature and enzyme activity in poikilotherms: Isocitrate
dehydrogenases in rainbow-trout liver. Biochem. J. 1:)3, 695-705. Moon, T. W. and Hochachka, P. W. (1972) Temperature and the kinetic analysis of trout isocitrate dehydro-
genases. Comp. Biochem. Physiol. 42B, 725-730. SchrSder, S. (1989) Die Weir~tanne in SSddeutschland: Genetische Variation, Kline, Korrelationen. AIIg. Forst-u.
Jagdztg. 160, 100-104. van Delden, W. (1982) The alcohol dehydrogenase polymorphism in Drosophila melanogaster." Selection at an
enzyme locus. Evol, Biol. 15, 187-222. Watt, W. B. (1977) Adaptation at specific loci. I. Natural selection on phosphoglucose isomerase of Colias butter-
flies: Biochemical and population aspects. Genedcs 87, 177-194. Watt, W. B. (1983) Adaptation at specific loci. II. Demographic and biochemical elements in the maintenance of
the Colias PGI polymorphism. Genetics 103, 691-724.





























