Embed Size (px)
Citation preview

Signal Transduction
Dysregulation of WNT5A/ROR2 SignalingCharacterizes the Progression of Barrett-Associated Esophageal AdenocarcinomaOrestis Lyros1,2, Linghui Nie3, Tami Moore3, Rituparna Medda3, Mary Otterson3,Behnaz Behmaram4, Alexander Mackinnon4, Ines Gockel2, Reza Shaker1, andParvaneh Rafiee3
Abstract
The mechanism underlying the progression of normalesophageal mucosa to esophageal adenocarcinoma remainselusive. WNT5A is a noncanonical WNT, which mainly func-tions via the receptor tyrosine kinase-like orphan receptor 2(ROR2), and has an unclear role in carcinogenesis. In thisstudy, we aimed to determine the role of WNT5A/ROR2signaling in esophageal adenocarcinoma. Analysis of WNT5Aand ROR2 expression patterns in healthy controls, Barrett andesophageal adenocarcinoma patients' esophageal clinical spe-cimens as well as in various esophageal cell lines demonstrateda ROR2 overexpression in esophageal adenocarcinoma tissuescompared with Barrett and healthy mucosa, whereas WNT5Aexpression was found significantly downregulated towardesophageal adenocarcinoma formation. Treatment of esoph-ageal adenocarcinoma OE33 cells with human recombinantWNT5A (rhWNT5A) significantly suppressed proliferation,
survival, and migration in a dose-dependent fashion.rhWNT5A was found to inhibit TOPflash activity in ROR2wild-type cells, whereas increased TOPflash activity in ROR2-knockdown OE33 cells. In addition, ROR2 knockdown aloneabolished cell proliferation and weakened the migration prop-erties of OE33 cells. These findings support an early dysregula-tion of the noncanonical WNT5A/ROR2 pathway in the path-ogenesis of esophageal adenocarcinoma, with the loss ofWNT5A expression together with the ROR2 overexpression tobe consistent with tumor promotion.
Implications: The dysregulation of WNT5A/ROR2 noncanonicalWNT signaling in Barrett-associated esophageal adenocarcinomaintroduces possible prognostic markers and novel targets fortailored therapy of this malignancy. Mol Cancer Res; 14(7); 647–59.�2016 AACR.
IntroductionEsophageal adenocarcinoma is noted by having both the
fastest growing incidence of all cancers in the western worldover the last several decades (1) and a high mortality rate thatexceeds 80% (2). Despite improvements in chemotherapyregimens and the use of multimodal strategies (e.g., surgery,neoadjuvant therapy, and adjuvant therapy), esophageal ade-nocarcinoma remains a therapeutic challenge for oncologists(2) due to high rates of local recurrence and early regional andsystemic metastases. Esophageal adenocarcinoma is oftenstudied in conjunction with Barrett esophagus, the conditionin which the normal squamous epithelium in the distalesophagus is replaced by a metaplastic columnar mucosa inresponse to chronic severe gastroesophageal reflux disease
(GERD; ref. 3). The course of Barrett esophagus is mainlybenign; however, this condition is the main predisposingfactor for esophageal adenocarcinoma and is associated withan increased risk of progression to cancer (4). The molecularmechanisms underlying the development and the aggressivephenotype of esophageal adenocarcinoma remain largelyundefined.
The embryologic WNT signaling pathway is important fordevelopment and tissue homeostasis, and its dysregulation hasbeen associated with tumorigenesis (5). It has been establishedthat WNT proteins regulate at least three distinct intracellularsignaling pathways: b-catenin–dependent (canonical) and -inde-pendent (noncanonical) signaling pathways, the planar cellpolarity/convergent extension pathway, and the Ca2þ pathway(summarized in ref. 6). The canonicalWNT pathway regulates thecell cycle, growth, and proliferation (5), whereas the noncanon-ical WNT signaling is purported to have crucial functions in theregulation of cell migration and polarity (6). Cross-talk betweencanonical and noncanonical WNT pathways still remains elusive,but inhibitory interaction of the canonical by the noncanonicalhas been shown (7, 8).
Aberrant activation of the canonical WNT/b-catenin signal-ing has been associated with the development of esophagealadenocarcinoma (9). However, genetic alterations in compo-nents of the canonical WNT pathway such as b-catenin, APC,and Axin, which are observed in other human cancers and resultin upregulated canonical signaling, are not frequently detected
1Division of Gastroenterology & Hepatology, Medical College of Wis-consin, Milwaukee, Wisconsin. 2Department of Visceral, Transplant,Thoracic and Vascular Surgery, University Hospital, Leipzig, Germany.3Department of Surgery, Medical College of Wisconsin, Milwaukee,Wisconsin. 4Department of Pathology, Medical College of Wisconsin,Milwaukee,Wisconsin.
Corresponding Author: Parvaneh Rafiee, Department of Surgery, MedicalCollege of Wisconsin, 8701 Watertown Plank Road, Milwaukee, WI 53226.Phone: 414-955-4667; Fax: 414-955-6361; E-mail: [email protected]
doi: 10.1158/1541-7786.MCR-15-0484
�2016 American Association for Cancer Research.
MolecularCancerResearch
www.aacrjournals.org 647
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484

in esophageal adenocarcinoma (10, 11). Instead, esophagealadenocarcinoma is marked by changes, such as loss of thenegative regulators of the pathway, Wnt inhibitory factor 1,and secreted frizzled receptor proteins), as well as induction ofWnt-2 expression, which are expected to increase signalingalong the WNT axis (9, 12, 13). Thus, the mechanism ofcanonical WNT activation in esophageal adenocarcinoma isstill unknown, whereas the involvement of the noncanonicalWNT pathway has not been addressed yet.
WNT5A is classified as a nontransformingWNT familymemberthat activates noncanonical WNT signaling by interacting withreceptor tyrosine kinase–like orphan receptors (ROR; ref. 14).ROR2 is amember of the ROR family of receptor tyrosine kinases,acts as a receptor or coreceptor forWNT5A, andhas shown to haveessential functions in developmental morphogenesis (15).WNT5A/ROR2 signaling has been shown to inhibit the canonicalWNT signaling, which is often activated in tumor cells, indicatingthatWNT5Amight act as a tumor suppressor (16). Indeed, loss orlow expression of WNT5A is associated with increased invasionand aggressiveness of many carcinomas (17, 18). Nevertheless,WNT5A has also demonstrated oncogenic properties based onfindings depicting upregulation of WNT5A levels in melanoma,lung, and breast cancers (19–21). Although, the exact functionofWNT5A is highly dependent on the receptor context, the ROR2receptor on the other hand, appears to demonstrate oncogenicproperties regardless the presence of ligand (22). Thus, the roleof WNT5A/ROR2 signaling in carcinogenesis remains stillambiguous.
The current study was undertaken to explore the role of thenoncanonical WNT5A/ROR2 signaling pathway in esophage-al adenocarcinoma with respect to the canonical WNT/b-cate-nin signaling activation. We hypothesized that alteration inthe WNT5A/ROR2 signaling pathway may underlie the aber-rant activation of b-catenin transcriptional activity in esoph-ageal adenocarcinoma cells. By analyzing human esophagealspecimens and esophageal cell lines, we present evidencesuggesting that the progression of esophageal adenocarcino-ma is characterized by deregulation of the noncanonicalWNT5A/ROR2 signaling pathway. The loss of WNT5A expres-sion along with upregulation of ROR2 receptor appears tocontribute to an aggressive phenotype of the esophagealadenocarcinoma cells. Our findings underscore the signifi-cance of the WNT5A/ROR2 signaling pathway in esophagealadenocarcinoma and provide rationale for the developmentof future target therapies along with new insights into themechanisms of noncanonical WNT signaling in the context oftumor progression.
Materials and MethodsReagents and antibodies
Recombinant human WNT5A (rhWNT5A) was obtained fromR&D Systems Inc. Antibodies against ROR2 and WNT5A wereobtained fromAbcam; activeb-catenin antibody (ABC;unphospho-rylated in Ser37 and Thr41 sites) was fromMillipore, total b-cateninand phospo-GSK3b (Ser9) antibodies were obtained from CellSignaling Technology. GSK3b and horseradish peroxidase second-ary antibodies were obtained from Santa Cruz Biotechno-logy. b-Actin antibody, DNA demethylation drug; 5-azacytidine(5-Aza-CR), protease inhibitor cocktail, and all other reagents andchemicals were obtained from Sigma unless otherwise specified.
Human esophageal specimensArchival paraffinized human esophageal specimens of esoph-
ageal adenocarcinoma patients (n¼ 11) who underwent esopha-gectomy in the Department of Thoracic Surgery at FroedtertHospital and Medical College of Wisconsin from 2010 to 2012were retrospectively analyzed (Table 1). Laser microdissectionwas undertaken to isolate RNA from tumor cells from paraffinblocks (purification up to 70%–80% was achieved). In addition,human esophageal biopsy specimens were collected frompatients 18 years or older undergoing endoscopy for Barrettesophagus (n¼ 15) and healthy individuals (n¼ 10) undergoingendoscopy for nonesophageal indications (Table 2). Studies wereapproved by the Human Research Review Committee of theMedical College ofWisconsin, and study participants gavewritteninformed consent prior to their studies.
Cell linesHuman squamous esophageal telomerase-immortalized cells,
EPC1 (EPC1-hTERT) and EPC2 (EPC2-hTERT) with intact p53and p16 genes (23) were the generous gifts of Dr. HiroshiNakagawa (Gastroenterology Division, University of Pennsylva-nia, Philadelphia, PA). CP-A cells are a nondysplastic columnarcell line derived from Barrett esophagus that was obtained fromthe ATCC. OE33 cells are poorly differentiated esophageal ade-nocarcinoma cancer cells (Sigma). Cells have been recently testedand authenticated by immunofluorescence staining for cytoker-atin expression and were grown according to a standard protocolas described previously (24).
Cellular assaysCell proliferation was assessed either by [3H] thymidine
uptake, as described previously (25), or by 3-(4,5-dimethylthia-zol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, as
Table 1. Demographic and histologic findings in esophageal adenocarcinoma patients
Patient Age Sex TNM classificationPresence ofBarrett mucosa Neoadjuvant
#1 77 M pT1a, N0, Mx, (G1) Yes No#2 56 M pT1b, N0, Mx (G2) Yes No#3 62 F pT1b, N0, Mx, (G1) Yes No#4 46 F pT3, pN1a, pMX (G3) No No#5 76 M pT1b, pN0, pMX (G2) Yes No#6 74 M pT1b, pN0, pMX (G3) Yes No#7 60 M pT1b, pN0, pM0 (G2) No No#8 61 M pT1b, pN0, pMX (G3) No No#9 55 M pT3, pN1, pMX (G3) No No#10 68 M pTcis, N0, Mx (G1) Yes No#11 68 M pT3, pN1, Mx (G2) Yes No
Lyros et al.
Mol Cancer Res; 14(7) July 2016 Molecular Cancer Research648
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484

suggested by the manufacturer (Roche Diagnostics). Migration ofOE33 cancer cells was assayed withWound Scratch Assay and celltransmigration by using 24-well HTS FluoroBlok inserts in triplets(8-mm pore size; Becton Dickinson). Later, cells were serumstarved for 24 hours before initiation of the assay. Cells (4 �104) were resuspended in serum-free RPMI and added to the topchamber. Consecutively, RPMI with or without rhWNT5A wasadded to the bottom chamber. Chambers were incubated eitherfor 24 hours or 48 hours at 37�C in 5%CO2. After incubation, thenumber of migrated cells in the bottom chamber was determinedby luminescence assay according to the recommendations of themanufacturer (Celltiter-Glo Cell Viability assay, Promega Corpo-ration) following incubation with Calcein AM Fluorescent Dye(BD Biosciences).
RNA isolation and qRT-PCRRNA was isolated from biopsies and OE33 cell using Arcturus
PicoPure RNA (Life Technologies) and the RNeasy Kit (Qiagen),respectively. cDNA was synthesized from 1 mg of total RNA usingthe iScript cDNA synthesis kit according to the manufacturer'sprotocol (Bio-Rad). qRT-PCR was performed with SsoFast Eva-
Green Supermix (Bio-Rad), with 250 nmol/L primer and 1 mL ofcDNA per 20-mL reaction as described previously (24). 5-Aza-CRwas administrated to the OE33 cells at the concentration of 0.5mmol/L and RNA was collected after 6, 24, 48, and 72 hours andanalyzed as described above. Gene expression was analyzed withiQ5 software (Bio-Rad). The primers used are; ROR2: (F: 50-GGCAGAACCCATCCTCGTG-30, R: 50-CGACTGCGAATCCAGGACC -30),WNT5A: (F: 50-GTGCAATGTCTTCCAAGT TCTT-30, R: 50-ATG AGC CGG ACA CCC CAT GGC A-30), GAPDH: (F: 50- TGCACC ACC AAC TGC TTA GC-30, R: 50-GGC ATG GAC TGT GGTCATGAG-30),b-actin: (F: 50-CACTCTTCCAGCCTTCCTTC-30, R:50-GGT GTA ACG CAA CTA AGT CAT), Axin2: (F: 50-TAC CGGAGGATGCTGAAGGC-30, R: 50-CCACTGGCCGATTCTTCCTT-30), Ki67:(F: 50-ATT ATC GTT CCT TCA GGT ATG-30, R: 50-TCATCA GGG TCA GAA GAG AA-3).
Western blot analysisCell extracts were prepared and subjected to SDS-PAGE and
Western blotting using antibodies against ROR2, WNT5A, b-cate-nin, active b-catenin, phospho GSK3, total GSK3, and b-actin asdescribed previously (26).
Table 2. Demographic, endoscopic, and histologic findings in Barrett patients and healthy controls
Patient Age Sex Indication for upper GI-endoscopy Endoscopy report Pathology report
Healthy Controls#1 61 M Abdominal pain Normal looking squamous mucosa No abnormalities#2 46 F Weight loss, diarrhea Normal looking squamous mucosa No abnormalities#3 23 M Abdominal pain Normal looking squamous mucosa No abnormalities#4 37 M Vitamin B deficiency Normal looking squamous mucosa No abnormalities#5 60 F Abdominal pain Normal looking squamous mucosa No abnormalities#6 24 F Abdominal pain Normal looking squamous mucosa No abnormalities#7 25 M Crohn disease Normal looking squamous mucosa No abnormalities#8 46 F Family history of esophageal cancer Normal looking squamous mucosa No abnormalities#9 39 F Severe iron deficiency anemia Normal looking squamous mucosa No abnormalities#10 54 M Not reported Normal looking squamous mucosa No abnormalities
Barrett Patients#1 66 M History of Barrett, repaired hiatus hernia Barrett esophagus 3 cm BE-Metaplasia, no dysplasia#2 72 F Hx of Barrett, GERD Two tongues of Barrett esophagus <1 cm BE-Metaplasia, no dysplasia#3 60 M Hx of Barrett, GERD Barrett mucosa with scattered patchy
distributionBE-Metaplasia, no dysplasia
#4 45 M Hx of Barrett, GERD Irregular Z-line with possible 1 cm segment ofBarrett esophagus
BE-Metaplasia, no dysplasia
#5 51 M Hx of Barrett Irregular Z-line with islands of salmon coloredBarrett typemucosa in the vicinity of theZ-line.
BE-Metaplasia, no dysplasia
#6 68 F GERD Salmon-colored tongue-shaped extensions of upto 2 cm into distal esophagus
BE-Metaplasia, no dysplasia
#7 64 M Hx of Barrett Area of short-segment Barrett esophagus, visibleon retroflexion within hernia sac
BE-Metaplasia, no dysplasia
#8 60 F Hx of erosive esophagitis Short segment of Barrett extending for �2 cmsegment in two tongues
BE-Metaplasia, no dysplasia
#9 45 F Hx of BE, GERD symptoms Barrettmucosa,with a few islands, extends about1 cm above anatomic GE junction (PragueC0M1).
BE-Metaplasia, no dysplasia
#10 59 M Hx of Barrett Multiple islands Barrett mucosa—short segment BE-Metaplasia, no dysplasia#11 64 F Hx of Barrett Not available BE-Metaplasia, no dysplasia#12 59 M Hx of Barrett Circumferential salmon colored mucosa 34–36
cm without noduleBarrett esophagus with focallow-grade dysplasia
#13 88 M Hx of Barrett 1 cm Barrett esophagus (C0M1) in hiatal hernia BE-Metaplasia, no dysplasia#14 46 F Hx of GERD Barrett appearing mucosa C1, M3 (upper limit
of long segment 33 cm, upper limit ofcircumferential segment 35 cm GE junction36 cm and diaphragmatic indentation 38 cm)
BE-Metaplasia, no dysplasia
#15 74 M Hx of Barrett with LGD Barrett esophagus, Prague classification C8M10 Barrett esophagus with focallow-grade dysplasia
Abbreviations: BE, Barrett esophagus; GE, gastroesophageal; Hx, history; GI, gastrointestinal.
Noncanonical WNT5A-ROR2 Axis in Esophageal Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 14(7) July 2016 649
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484

Immunohistochemical analysisMucosal esophageal biopsy specimens were embedded in
paraffin blocks and cut into 5-mm sections. The sections wereroutinely stained with hematoxylin and eosin for histologicdiagnosis, and additional sequential sections were subjected toIHC using antibodies against WNT5A and ROR2, as describedpreviously (24). Expression of ROR2 immunohistochemicalstaining was quantified by both, determination of the intensityscore of ROR2 expression in all biopsy samples using a 0–4simple scoring system by a specialized pathologist as well asusing an Automated Cellular Imaging System III (ACIS III,DAKO) in a single case as described previously (27). This casewas representative of the entire cohort and contained areas ofnormal squamous mucosa, Barrett mucosa, and adenocarcino-ma, and each of these histologic areas were analyzed indepen-dently and graphed. The percentage of ROR2-positive tissueand the intensity of ROR2 were measured independently andused to create a region score. Obvious artifacts including tissuefolding, edge effect, nonspecific chemical precipitation, anddust or debris artefacts were excluded.
WNT5A and ROR2 gene silencingOE33 and EPC2-hTERT cells were transfected with predesigned
short interfering RNA (siRNA) targeting human WNT5A and/orROR2 gene and a control negative siRNA (Dharmacon) accordingto themanufacturer's protocol and described previously (24). Theratio of siRNA to DharmaFECT reagent was 25 nmol/L siRNA to1.5 mL of transfection reagent.
Luciferase reporter assayThe assay was performed as described previously (24). In
summary, OE33 cells in 60% confluence in 24-well plates weretransfected with DNA plasmids of b-catenin-LEF/TCF-sensitive(TOP-flash) or b-catenin-LEF/TCF-insensitive (FOP-flash) report-er vectors (Addgene) using Lipofectamine 2000 (Invitrogen) for48 hours. Onemicrogram of TOPflash as well as 1 mg of FOPflashwere used perwell of a 24-well plate. phRL-TKplasmid (Promega)was cotransfected as control for transfection efficiency. Reporterassay was performed using dual luciferase reporter system (Pro-mega). Luciferase activity was measured via GLOMAX 20/20Luminometer (Promega). Values for each reporter were normal-ized to phRL-TK values.
Statistical analysisIBMSPSS Statistics 19 softwarewas used for statistical analysis. All
experimentswereperformed in triplicate, andvalues aremeans� SE.Expression patterns between squamous mucosa, Barrett metaplasia,and esophageal adenocarcinoma tissue were compared withinesophageal adenocarcinoma patients and across different patients'groups (healthy controls, Barrett patients, and esophageal adeno-carcinoma patients) by using nonparametric methods followed byWilcoxon signed-rank test and Mann–Whitney test, respectively.Significant differences in cellular assays were evaluated by Studentt test. P � 0.05 was considered statistically significant.
ResultsROR2 is overexpressed in human esophageal adenocarcinoma
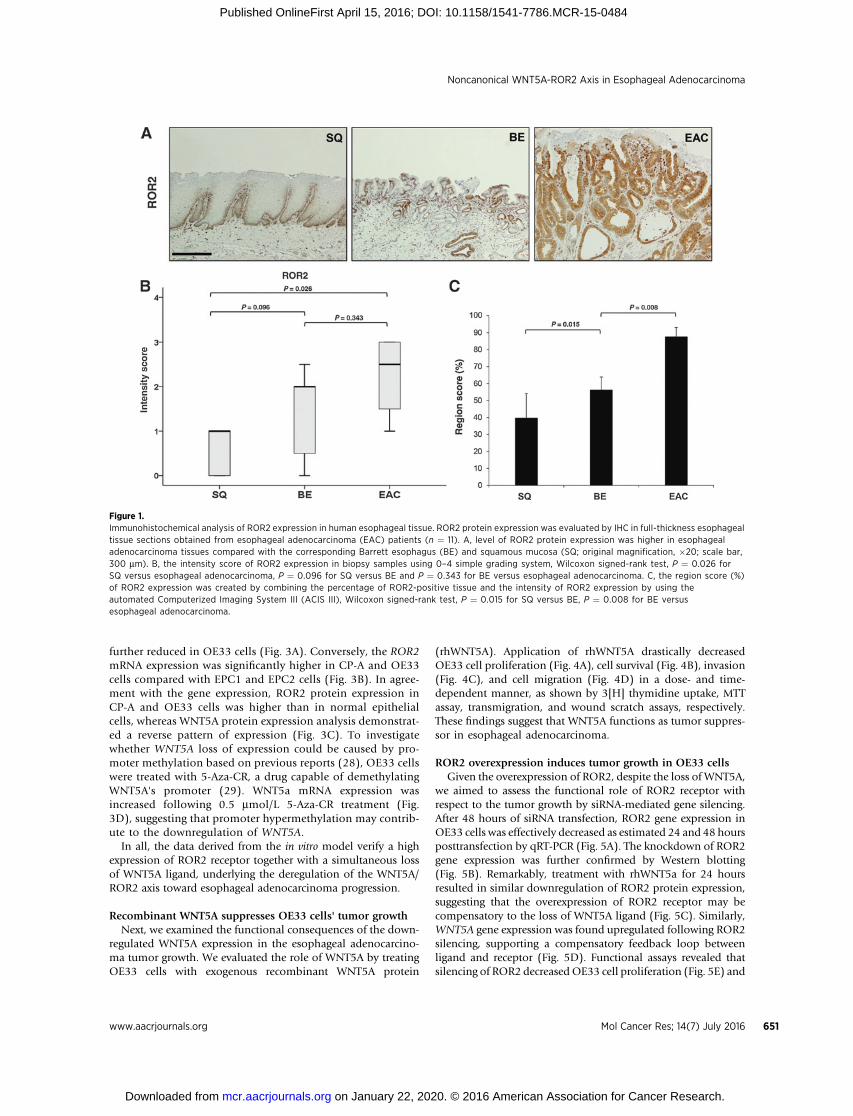
To demonstrate whether ROR2 expression alters in the courseof progression from normal squamous epithelium to esoph-ageal adenocarcinoma, ROR2 expression was evaluated by IHC
in esophageal tissue sections obtained from esophageal adeno-carcinoma patients (n ¼ 11) undergoing esophagectomy, with-out previous treatment. High levels of ROR2 were detected inesophageal adenocarcinoma tissues, whereas ROR2 expressionwas weak in the corresponding Barrett esophagus and evenweaker and isolated in the proliferative basal layer of thecorresponding squamous esophageal mucosa (Fig. 1A). A sim-ple grading system (0–4; Fig. 1B) as well as an image analysissoftware were used to quantify the intensity and region score(%; Fig. 1C) of ROR2 staining in the different tissues asdescribed in Materials and Methods. Statistical analysis, usingFriedman–Kendall W test to analyze the overall results ofintensity score between the three related conditions yielded aKendall coefficient of concordance of 0.363, which approachedstatistical significance (P ¼ 0.079). Finally, significance betweentwo-group comparisons was determined by Wilcoxon signed-rank tests, followed by correction for multiple comparisons.Overall, ROR2 expression was significantly higher in esophagealadenocarcinoma compared with squamous mucosa and Barrettesophagus tissues.
WNT5A expression is downregulated in esophagealadenocarcinoma
Next, we examined the level of WNT5A mRNA expression inesophageal biopsy specimens from three independent patients'groups, healthy controls (n ¼ 10), Barrett (n ¼ 15), and esoph-ageal adenocarcinoma patients (n ¼ 5). To ensure tissue homo-geneity in esophageal adenocarcinoma tissues, laser microdissec-tion was utilized to isolate tumor lesions. qRT-PCR analysisdemonstrated significantly lower level of WNT5A mRNA inesophageal adenocarcinoma samples compared with squamousmucosa and Barrett esophagus biopsy samples (Fig. 2A). Wecompared mRNA expression ofWNT5A gene between groups byusing a global nonparametric Kruskal–Wallis test, whichshowed a significant overall difference (P ¼ 0.0069). Subse-quent comparisons between groups (squamous mucosa vs.Barrett esophagus, squamous mucosa vs. esophageal adenocar-cinoma, and Barrett esophagus vs. esophageal adenocarcino-ma) was done by the nonparametric Mann–Whitney test.Corresponding with the gene expression, Western blot analysisof biopsy samples from esophageal adenocarcinoma patientsshowed a lower level of WNT5A protein in tumor site comparedwith the corresponding healthy esophageal mucosa (Fig. 2B).Conversely, the level of ROR2 protein expression was higher intumor biopsy compared with healthy mucosa. Immunohisto-chemical analysis of WNT5A expression in healthy esophagealmucosa revealed a nuclear staining throughout all mucosallayers, whereas ROR2 expression was detected mainly in cellcytoplasm and membrane of the basal mucosal layer (Fig. 2C).Further immunohistochemical analysis confirmed the negativecorrelation of ROR2 and WNT5A expression in esophagealadenocarcinoma tissue (Fig. 2D).
ROR2/WNT5A expression in esophageal cell linesThe baseline level of WNT5A and ROR2 gene and protein
expression in esophageal cell lines derived from normal esoph-agus (EPC1, EPC2), nondysplastic Barrett esophagus (CP-A),and esophageal adenocarcinoma (OE33) were evaluated byRT-PCR and Western blotting. The level of WNT5A mRNAexpression in EPC1 and EPC2 cells was higher than Barrettesophagus–derived cells (CP-A), and WNT5A expression was
Lyros et al.
Mol Cancer Res; 14(7) July 2016 Molecular Cancer Research650
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484
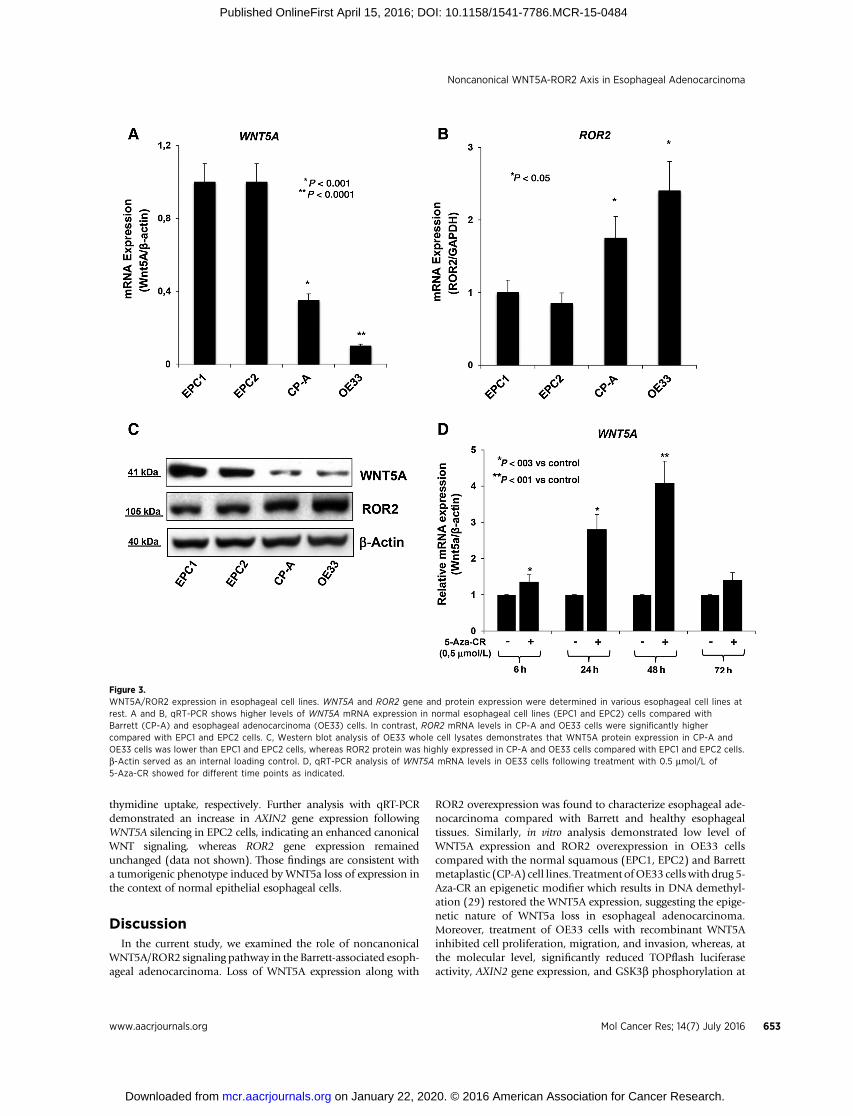
further reduced in OE33 cells (Fig. 3A). Conversely, the ROR2mRNA expression was significantly higher in CP-A and OE33cells compared with EPC1 and EPC2 cells (Fig. 3B). In agree-ment with the gene expression, ROR2 protein expression inCP-A and OE33 cells was higher than in normal epithelialcells, whereas WNT5A protein expression analysis demonstrat-ed a reverse pattern of expression (Fig. 3C). To investigatewhether WNT5A loss of expression could be caused by pro-moter methylation based on previous reports (28), OE33 cellswere treated with 5-Aza-CR, a drug capable of demethylatingWNT5A's promoter (29). WNT5a mRNA expression wasincreased following 0.5 mmol/L 5-Aza-CR treatment (Fig.3D), suggesting that promoter hypermethylation may contrib-ute to the downregulation of WNT5A.
In all, the data derived from the in vitro model verify a highexpression of ROR2 receptor together with a simultaneous lossof WNT5A ligand, underlying the deregulation of the WNT5A/ROR2 axis toward esophageal adenocarcinoma progression.
Recombinant WNT5A suppresses OE33 cells' tumor growthNext, we examined the functional consequences of the down-
regulated WNT5A expression in the esophageal adenocarcino-ma tumor growth. We evaluated the role of WNT5A by treatingOE33 cells with exogenous recombinant WNT5A protein
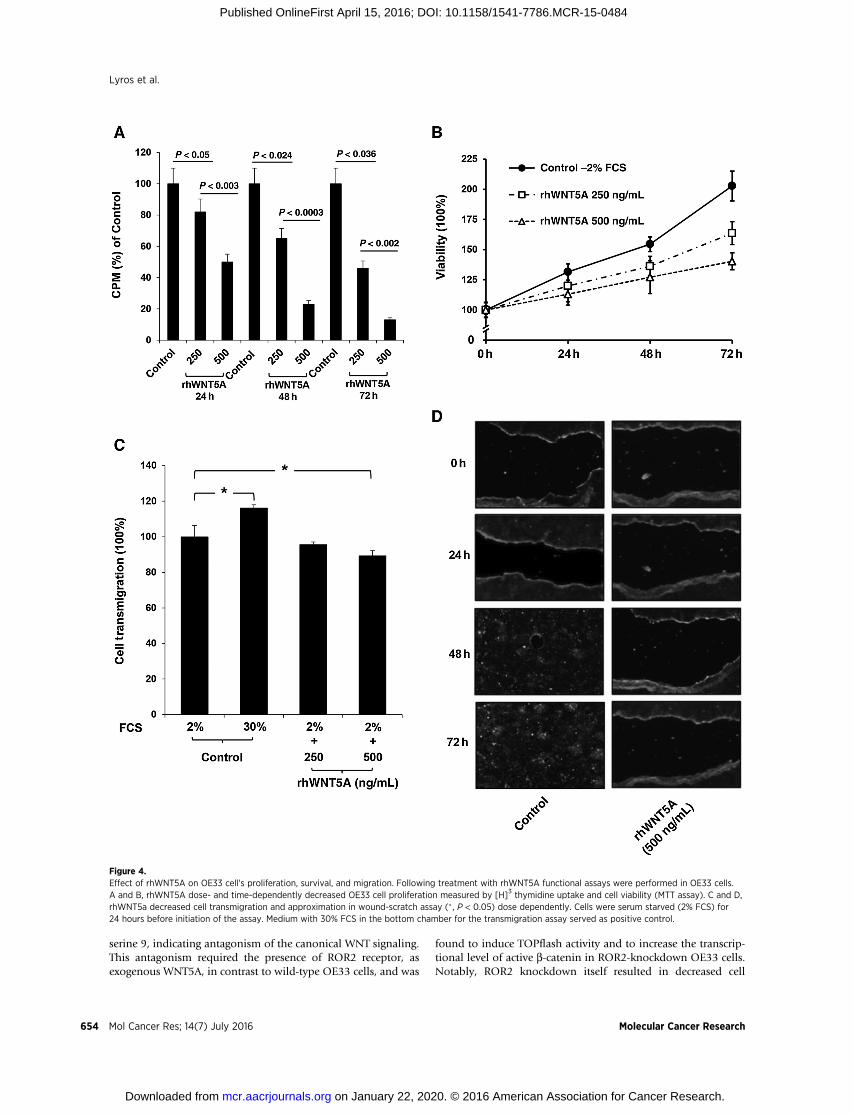
(rhWNT5A). Application of rhWNT5A drastically decreasedOE33 cell proliferation (Fig. 4A), cell survival (Fig. 4B), invasion(Fig. 4C), and cell migration (Fig. 4D) in a dose- and time-dependent manner, as shown by 3[H] thymidine uptake, MTTassay, transmigration, and wound scratch assays, respectively.These findings suggest that WNT5A functions as tumor suppres-sor in esophageal adenocarcinoma.
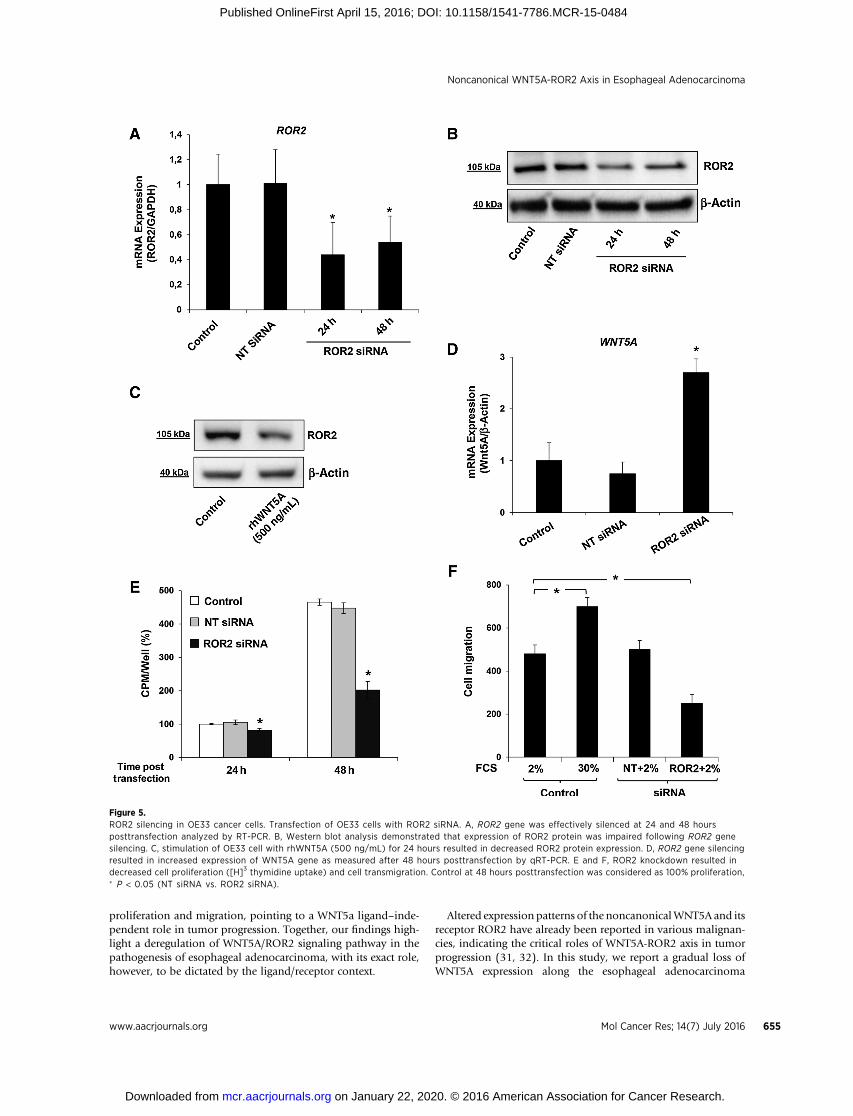
ROR2 overexpression induces tumor growth in OE33 cellsGiven the overexpression of ROR2, despite the loss of WNT5A,
we aimed to assess the functional role of ROR2 receptor withrespect to the tumor growth by siRNA-mediated gene silencing.After 48 hours of siRNA transfection, ROR2 gene expression inOE33 cells was effectively decreased as estimated 24 and 48 hoursposttransfection by qRT-PCR (Fig. 5A). The knockdown of ROR2gene expression was further confirmed by Western blotting(Fig. 5B). Remarkably, treatment with rhWNT5a for 24 hoursresulted in similar downregulation of ROR2 protein expression,suggesting that the overexpression of ROR2 receptor may becompensatory to the loss of WNT5A ligand (Fig. 5C). Similarly,WNT5A gene expression was found upregulated following ROR2silencing, supporting a compensatory feedback loop betweenligand and receptor (Fig. 5D). Functional assays revealed thatsilencing of ROR2 decreasedOE33 cell proliferation (Fig. 5E) and
Figure 1.Immunohistochemical analysis of ROR2 expression in human esophageal tissue. ROR2 protein expression was evaluated by IHC in full-thickness esophagealtissue sections obtained from esophageal adenocarcinoma (EAC) patients (n ¼ 11). A, level of ROR2 protein expression was higher in esophagealadenocarcinoma tissues compared with the corresponding Barrett esophagus (BE) and squamous mucosa (SQ; original magnification, �20; scale bar,300 mm). B, the intensity score of ROR2 expression in biopsy samples using 0–4 simple grading system, Wilcoxon signed-rank test, P ¼ 0.026 forSQ versus esophageal adenocarcinoma, P ¼ 0.096 for SQ versus BE and P ¼ 0.343 for BE versus esophageal adenocarcinoma. C, the region score (%)of ROR2 expression was created by combining the percentage of ROR2-positive tissue and the intensity of ROR2 expression by using theautomated Computerized Imaging System III (ACIS III), Wilcoxon signed-rank test, P ¼ 0.015 for SQ versus BE, P ¼ 0.008 for BE versusesophageal adenocarcinoma.
Noncanonical WNT5A-ROR2 Axis in Esophageal Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 14(7) July 2016 651
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484

transmigration (Fig. 5F), signifying that ROR2 overexpressioninduces tumor growth.
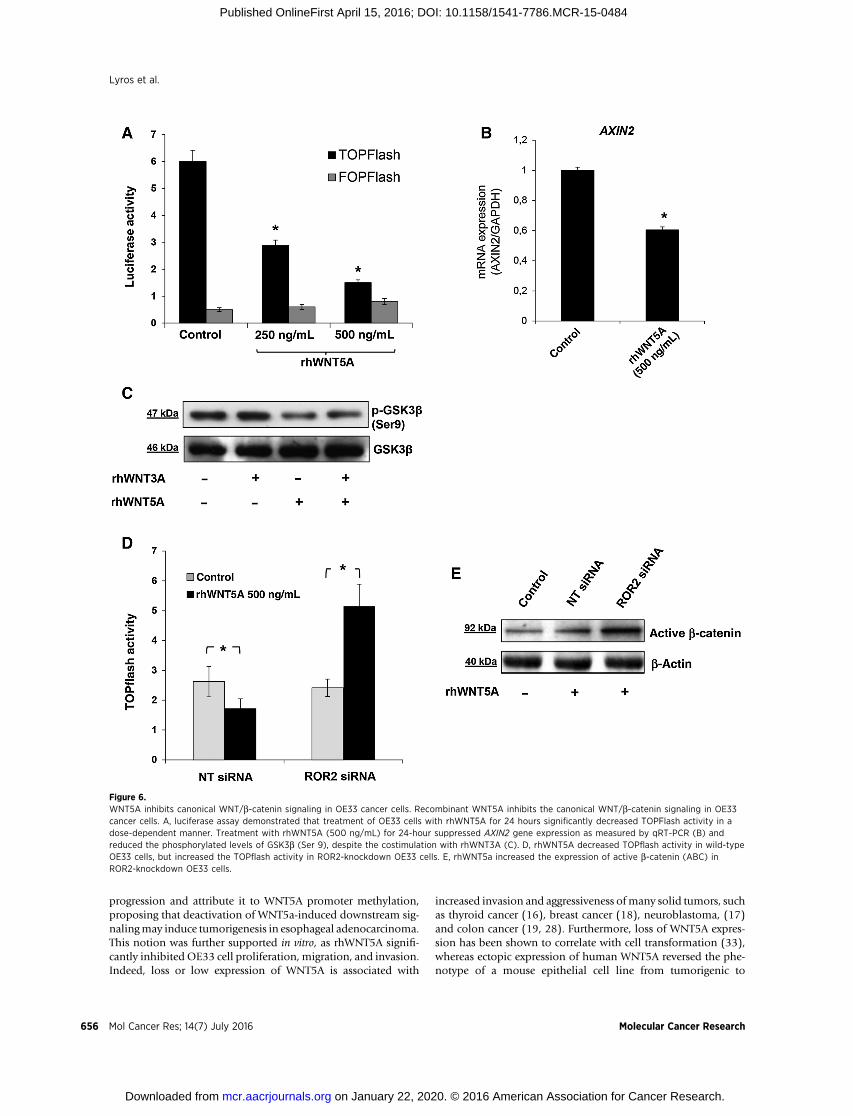
WNT5A suppresses canonical Wnt/b-catenin signalingin OE33 cells
To determine the nature of tumor suppressor properties ofWNT5A, we investigated whether WNT5a inhibits the canonicalWnt/b-catenin signaling in OE33 cells. Treatment with rhWNT5afor 24 hours significantly decreased TOPFlash activity in a dose-dependentmanner (Fig. 6A). In agreementwith a reduced b-catenintranscriptional activity, the gene expression of the b-catenin targetgene, AXIN2, was suppressed by rhWNT5A (Fig. 6B). At the molec-ular level, rhWNT5A was found to reduce the level of phosphor-ylated GSK3b (Ser 9; Fig. 6C), indicating activation of GSK3b andsubsequent inhibition of b-catenin–mediated signaling (30). Ofnote, the WNT5A-mediated dephosphorylation of GSK3b was notrescued by the canonical rhWNT3A, suggesting independent down-stream signaling transduction between the two WNTs.
To further demonstrate the role of the ROR2 receptor in theWNT5A-induced inhibition of the canonical WNT/b-catenin sig-naling, the level of TOPFlash activity was evaluated by treating
OE33 cells with rhWNT5a for 24 hours after sufficient siRNA-mediatedROR2 gene silencing. TOPflash activity was inhibited byrhWNT5A treatment in wild-type OE33 cells; however, it wasmarkedly increased in ROR2-knockdown cells (Fig. 6D). Wefurther confirmed this finding by increased level of ABC byrhWNT5A stimulation in the ROR2-knockdown cells (Fig. 6E).These findings signify that the presence of ROR2 receptor dictatesthe function of WNT5A regarding inhibition or induction of thecanonical WNT/b-catenin signaling.
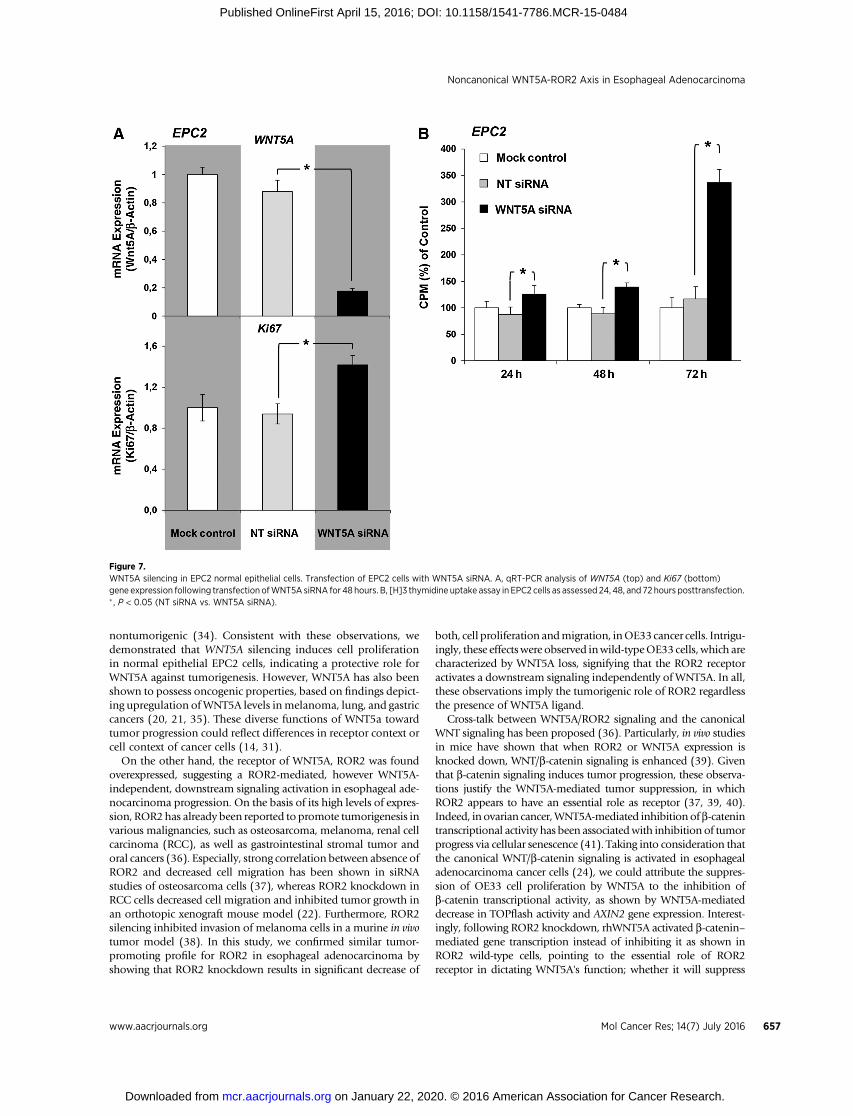
WNT5A knockdown induces proliferation in normal EPC2epithelial cells
Finally, to reversely address,whetherWNT5Asilencing innormalepithelial cells could induce a tumorigenic phenotype, we investi-gated the proliferative properties of EPC2 normal epithelial cellsfollowing by knocking down the WNT5A gene expression usingsiRNA tranfection. Sufficient WNT5A gene silencing in EPC2 cellswas demonstrated after 48 hours of siRNA transfection and wasfound to result in significant increase of Ki67 expression (Fig. 7A)aswell as inmarked inductionof cell proliferation,mainly72hoursposttransfection (Fig. 7B) as shown by qRT-PCR and 3[H]
Figure 2.WNT5A/ROR2 expressions in esophageal tissues. WNT5A gene expression in esophageal biopsy samples was determined by qRT-PCR. A, low levelof WNT5A mRNA expression was found in esophageal adenocarcinoma (EAC) tissues compared with independent squamous mucosa (SQ) (n ¼ 10)and Barrett esophagus (BE; n ¼ 15). Mann–Whitney test, P ¼ 0.002; esophageal adenocarcinoma versus SQ, P ¼ 0.086; BE versus SQ and P ¼ 0.05;esophageal adenocarcinoma versus BE. B, representative Western blot analysis of esophageal human tissues from one esophageal adenocarcinomapatient using antibodies against ROR2 and WNT5A, demonstrates that esophageal adenocarcinoma samples express low level of WNT5A proteinand high level of ROR2 protein. Graph is the summarized data of relative density. b-Actin served as an internal loading control. C, immunohistochemicalanalysis of the cellular and mucosal localization of ROR2 and WNT5A protein expression in healthy esophageal squamous mucosa (originalmagnification, �40; scale bar, 100 mm). D, IHC revealed strong ROR2 expression in absence of WNT5a expression in esophageal adenocarcinomatissue (original magnification, �10; scale bar, 500 mm).
Lyros et al.
Mol Cancer Res; 14(7) July 2016 Molecular Cancer Research652
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484
thymidine uptake, respectively. Further analysis with qRT-PCRdemonstrated an increase in AXIN2 gene expression followingWNT5A silencing in EPC2 cells, indicating an enhanced canonicalWNT signaling, whereas ROR2 gene expression remainedunchanged (data not shown). Those findings are consistent witha tumorigenic phenotype induced by WNT5a loss of expression inthe context of normal epithelial esophageal cells.
DiscussionIn the current study, we examined the role of noncanonical
WNT5A/ROR2 signaling pathway in the Barrett-associated esoph-ageal adenocarcinoma. Loss of WNT5A expression along with
ROR2 overexpression was found to characterize esophageal ade-nocarcinoma compared with Barrett and healthy esophagealtissues. Similarly, in vitro analysis demonstrated low level ofWNT5A expression and ROR2 overexpression in OE33 cellscompared with the normal squamous (EPC1, EPC2) and Barrettmetaplastic (CP-A) cell lines. Treatment ofOE33 cellswith drug 5-Aza-CR an epigenetic modifier which results in DNA demethyl-ation (29) restored the WNT5A expression, suggesting the epige-netic nature of WNT5a loss in esophageal adenocarcinoma.Moreover, treatment of OE33 cells with recombinant WNT5Ainhibited cell proliferation, migration, and invasion, whereas, atthe molecular level, significantly reduced TOPflash luciferaseactivity, AXIN2 gene expression, and GSK3b phosphorylation at
Figure 3.WNT5A/ROR2 expression in esophageal cell lines. WNT5A and ROR2 gene and protein expression were determined in various esophageal cell lines atrest. A and B, qRT-PCR shows higher levels of WNT5A mRNA expression in normal esophageal cell lines (EPC1 and EPC2) cells compared withBarrett (CP-A) and esophageal adenocarcinoma (OE33) cells. In contrast, ROR2 mRNA levels in CP-A and OE33 cells were significantly highercompared with EPC1 and EPC2 cells. C, Western blot analysis of OE33 whole cell lysates demonstrates that WNT5A protein expression in CP-A andOE33 cells was lower than EPC1 and EPC2 cells, whereas ROR2 protein was highly expressed in CP-A and OE33 cells compared with EPC1 and EPC2 cells.b-Actin served as an internal loading control. D, qRT-PCR analysis of WNT5A mRNA levels in OE33 cells following treatment with 0.5 mmol/L of5-Aza-CR showed for different time points as indicated.
Noncanonical WNT5A-ROR2 Axis in Esophageal Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 14(7) July 2016 653
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484
serine 9, indicating antagonism of the canonical WNT signaling.This antagonism required the presence of ROR2 receptor, asexogenous WNT5A, in contrast to wild-type OE33 cells, and was
found to induce TOPflash activity and to increase the transcrip-tional level of active b-catenin in ROR2-knockdown OE33 cells.Notably, ROR2 knockdown itself resulted in decreased cell
Figure 4.Effect of rhWNT5A on OE33 cell's proliferation, survival, and migration. Following treatment with rhWNT5A functional assays were performed in OE33 cells.A and B, rhWNT5A dose- and time-dependently decreased OE33 cell proliferation measured by [H]3 thymidine uptake and cell viability (MTT assay). C and D,rhWNT5a decreased cell transmigration and approximation in wound-scratch assay (� , P < 0.05) dose dependently. Cells were serum starved (2% FCS) for24 hours before initiation of the assay. Medium with 30% FCS in the bottom chamber for the transmigration assay served as positive control.
Lyros et al.
Mol Cancer Res; 14(7) July 2016 Molecular Cancer Research654
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484
proliferation and migration, pointing to a WNT5a ligand–inde-pendent role in tumor progression. Together, our findings high-light a deregulation of WNT5A/ROR2 signaling pathway in thepathogenesis of esophageal adenocarcinoma, with its exact role,however, to be dictated by the ligand/receptor context.
Altered expression patterns of the noncanonicalWNT5A and itsreceptor ROR2 have already been reported in various malignan-cies, indicating the critical roles of WNT5A-ROR2 axis in tumorprogression (31, 32). In this study, we report a gradual loss ofWNT5A expression along the esophageal adenocarcinoma
Figure 5.ROR2 silencing in OE33 cancer cells. Transfection of OE33 cells with ROR2 siRNA. A, ROR2 gene was effectively silenced at 24 and 48 hoursposttransfection analyzed by RT-PCR. B, Western blot analysis demonstrated that expression of ROR2 protein was impaired following ROR2 genesilencing. C, stimulation of OE33 cell with rhWNT5A (500 ng/mL) for 24 hours resulted in decreased ROR2 protein expression. D, ROR2 gene silencingresulted in increased expression of WNT5A gene as measured after 48 hours posttransfection by qRT-PCR. E and F, ROR2 knockdown resulted indecreased cell proliferation ([H]3 thymidine uptake) and cell transmigration. Control at 48 hours posttransfection was considered as 100% proliferation,� P < 0.05 (NT siRNA vs. ROR2 siRNA).
Noncanonical WNT5A-ROR2 Axis in Esophageal Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 14(7) July 2016 655
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484
progression and attribute it to WNT5A promoter methylation,proposing that deactivation of WNT5a-induced downstream sig-nalingmay induce tumorigenesis in esophageal adenocarcinoma.This notion was further supported in vitro, as rhWNT5A signifi-cantly inhibited OE33 cell proliferation, migration, and invasion.Indeed, loss or low expression of WNT5A is associated with
increased invasion and aggressiveness ofmany solid tumors, suchas thyroid cancer (16), breast cancer (18), neuroblastoma, (17)and colon cancer (19, 28). Furthermore, loss of WNT5A expres-sion has been shown to correlate with cell transformation (33),whereas ectopic expression of human WNT5A reversed the phe-notype of a mouse epithelial cell line from tumorigenic to
Figure 6.WNT5A inhibits canonical WNT/b-catenin signaling in OE33 cancer cells. Recombinant WNT5A inhibits the canonical WNT/b-catenin signaling in OE33cancer cells. A, luciferase assay demonstrated that treatment of OE33 cells with rhWNT5A for 24 hours significantly decreased TOPFlash activity in adose-dependent manner. Treatment with rhWNT5A (500 ng/mL) for 24-hour suppressed AXIN2 gene expression as measured by qRT-PCR (B) andreduced the phosphorylated levels of GSK3b (Ser 9), despite the costimulation with rhWNT3A (C). D, rhWNT5A decreased TOPflash activity in wild-typeOE33 cells, but increased the TOPflash activity in ROR2-knockdown OE33 cells. E, rhWNT5a increased the expression of active b-catenin (ABC) inROR2-knockdown OE33 cells.
Lyros et al.
Mol Cancer Res; 14(7) July 2016 Molecular Cancer Research656
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484
nontumorigenic (34). Consistent with these observations, wedemonstrated that WNT5A silencing induces cell proliferationin normal epithelial EPC2 cells, indicating a protective role forWNT5A against tumorigenesis. However, WNT5A has also beenshown to possess oncogenic properties, based on findings depict-ing upregulation ofWNT5A levels inmelanoma, lung, and gastriccancers (20, 21, 35). These diverse functions of WNT5a towardtumor progression could reflect differences in receptor context orcell context of cancer cells (14, 31).
On the other hand, the receptor of WNT5A, ROR2 was foundoverexpressed, suggesting a ROR2-mediated, however WNT5A-independent, downstream signaling activation in esophageal ade-nocarcinoma progression. On the basis of its high levels of expres-sion, ROR2 has already been reported to promote tumorigenesis invarious malignancies, such as osteosarcoma, melanoma, renal cellcarcinoma (RCC), as well as gastrointestinal stromal tumor andoral cancers (36). Especially, strong correlation between absence ofROR2 and decreased cell migration has been shown in siRNAstudies of osteosarcoma cells (37), whereas ROR2 knockdown inRCC cells decreased cell migration and inhibited tumor growth inan orthotopic xenograft mouse model (22). Furthermore, ROR2silencing inhibited invasion of melanoma cells in a murine in vivotumor model (38). In this study, we confirmed similar tumor-promoting profile for ROR2 in esophageal adenocarcinoma byshowing that ROR2 knockdown results in significant decrease of
both, cell proliferation andmigration, inOE33cancer cells. Intrigu-ingly, these effectswere observed inwild-typeOE33 cells,which arecharacterized by WNT5A loss, signifying that the ROR2 receptoractivates a downstream signaling independently of WNT5A. In all,these observations imply the tumorigenic role of ROR2 regardlessthe presence of WNT5A ligand.
Cross-talk between WNT5A/ROR2 signaling and the canonicalWNT signaling has been proposed (36). Particularly, in vivo studiesin mice have shown that when ROR2 or WNT5A expression isknocked down, WNT/b-catenin signaling is enhanced (39). Giventhat b-catenin signaling induces tumor progression, these observa-tions justify the WNT5A-mediated tumor suppression, in whichROR2 appears to have an essential role as receptor (37, 39, 40).Indeed, in ovarian cancer,WNT5A-mediated inhibition ofb-catenintranscriptional activity has been associatedwith inhibition of tumorprogress via cellular senescence (41). Taking into consideration thatthe canonical WNT/b-catenin signaling is activated in esophagealadenocarcinoma cancer cells (24), we could attribute the suppres-sion of OE33 cell proliferation by WNT5A to the inhibition ofb-catenin transcriptional activity, as shown by WNT5A-mediateddecrease in TOPflash activity and AXIN2 gene expression. Interest-ingly, following ROR2 knockdown, rhWNT5A activated b-catenin–mediated gene transcription instead of inhibiting it as shown inROR2 wild-type cells, pointing to the essential role of ROR2receptor in dictating WNT5A's function; whether it will suppress
Figure 7.WNT5A silencing in EPC2 normal epithelial cells. Transfection of EPC2 cells with WNT5A siRNA. A, qRT-PCR analysis of WNT5A (top) and Ki67 (bottom)gene expression following transfection ofWNT5A siRNA for 48 hours. B, [H]3 thymidine uptake assay in EPC2 cells as assessed 24, 48, and 72 hours posttransfection.� , P < 0.05 (NT siRNA vs. WNT5A siRNA).
Noncanonical WNT5A-ROR2 Axis in Esophageal Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 14(7) July 2016 657
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484

the b-catenin axis or enhance it. The most possible explanationfor this notion is that in absence of ROR2, WNT5A activatesb-catenin via Frizzled receptors binding (14). Someone wouldexpect WNT5A to induce the canonical WNT signaling in normalepithelial cells, due to their low ROR2 expression. However,WNT5A silencing in EPC2 cells resulted in increased canonicalWNT signaling as dictated by induction of AXIN2 gene expression,indicating that in normal epithelial cells, WNT5Amight antagonizeother WNTs. Thus, the function of WNT5A regarding the activationof canonical WNT signaling is not only determined by the receptorcontext but also the presence of other WNTs.
It remains questionable, how rhWNT5A impaired the invasiveproperties of OE33 wild-type cells, when WNT5A is known toinduce cell migration and cancer metastasis by promoting epi-thelial–mesenchymal transition and activating transcription ofmatrix metalloproteases (MMP; ref. 42). Notably, exogenousWNT5A decreased ROR2 expression in OE33 cells indicating apossible reduction of receptor levels in the presence of ligand,highly likely via receptor-mediated endocytosis and degradation.Given that cell motility regulation via WNT5A requires ROR2receptor (43), we can speculate at this point that the subsequentROR2 downregulation in OE33 cells following rhWNT5A simu-lation may underlay the impaired invasiveness. In favor of thisnotion, ROR2 knockdown inducedWNT5A gene expression andwas also found to inhibit invasion and migration. However, thecomplex role of ROR2 receptor in esophageal adenocarcinomainvasiveness, in absence of WNT5A ligand, remains unclear andmerits further study.
Our current findings highlight that deregulation of WNT5A/ROR2 signalingmay contribute to the high invasive properties andsubsequently to the poor prognosis of esophageal adenocarcino-ma.Becauseof the limitednumberofpatients in this study,wewereunable to provide a powerful correlation between expressionpatterns and tumor stage or prognosis in this study, which is anacknowledged limitation. However, we observed a stronger ROR2staining together with a further decreased WNT5A gene expressionin the esophageal adenocarcinoma tissues classified in pT3/N1stage compared with those in pT1/N0 stage (data not shown),which allows us, at this point, to speculatively support the prog-nostic potential of ROR2 and WNT5A expressions in esophagealadenocarcinoma. Similar correlations, which allow conclusions onthe prognostic valueofWNT5AandROR2, have beenperformed inother malignancies (31). Intriguingly, in these studies, a pooroutcome was associated with the overexpression of both WNT5AandROR2 (38).Our report, however, indicates that isWNT5A loss-of-expression in combination with ROR2 gain-of-expression thatcontributes to the tumor growth andmetastatic potential of esoph-ageal adenocarcinoma cells.On the basis of our in vitrodata, we canspeculate that high WNT5a levels may be associated with a betterprognosis, whereas ROR2 overexpression with a poor one.Undoubtedly, our data make apparent that in the case of esoph-ageal adenocarcinoma, the prognostic value of WNT5A cannot beevaluated independently of ROR2 and vice versa. With respect toWNT5A's receptors, the expressionROR1 is yetnot addressed in thisstudy. ROR1hasbeen shown tobehighly expressed in several typesof cancer, including mainly hematopoietic malignancies (44) andmight play a role in the esophageal adenocarcinoma tumor pro-gression. However, in contrast to the well-established function ofROR2, it remains unclear whether ROR1 acts as a coreceptor forWNT5A-mediated signal in tumor biology (31) and such aninvestigation was beyond the scopes of this study.
In conclusion, we have examined for the first time that theWNT5A/ROR2 axis in the pathogenesis of esophageal adenocar-cinoma and we have shown that its deregulation is significantlyinvolved to esophageal adenocarcinoma tumor progression. Lossof WNT5A with concurrent activation of WNT5A-independentROR2 signaling appears to mark the progression of esophagealadenocarcinoma (Fig. 8). Although already speculated in othermalignancies, we confirmed thatWNT5A ligand could function intwo discreet pathways in esophageal adenocarcinoma based onROR2 receptor availability. It becomes evident that whetherWNT5A contributes to promote or suppress tumor progressionis partly dependent on the cellular context, the receptor's context,and the balance of the canonical WNT signaling and noncanon-ical WNT signaling. Thus, it is of great importance to understandthe respective molecular mechanisms governing WNT5A-medi-ated tumor-progressive and tumor-suppressive functions, todevelop novel and proper diagnostic and therapeutic strategiestargeting WNT5A signaling for esophageal adenocarcinoma.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: O. Lyros, I. Gockel, R. Shaker, P. RafieeDevelopment of methodology: O. Lyros, L. Nie, T. Moore, A. Mackinnon,P. RafieeAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.):O. Lyros, T. Moore, A. Mackinnon, I. Gockel, P. RafieeAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis):O. Lyros, T.Moore, A.Mackinnon, I. Gockel, P. RafieeWriting, review, and/or revision of the manuscript: O. Lyros, M. Otterson,A. Mackinnon, I. Gockel, R. Shaker, P. RafieeAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): O. Lyros, T. Moore, R. Medda, A. Mackinnon,P. RafieeStudy supervision: O. Lyros, T. Moore, R. Shaker, P. RafieeOther (the pathologist that contributed to the diagnosis and assessments ofthe tissue.): B. Behmaram
AcknowledgmentsThis work was supported by Digestive Disease Center, Medical College of
Wisconsin. The National Center for Advancing Translational Sciences, NIH,through grant number 8UL1TR000055, in part by NIH grant R01DK025731,and Laura Gralton on behalf of Daniel and Laura Gruber Charitable LeadTrust #3.
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received December 15, 2015; revised March 16, 2016; accepted March 30,2016; published OnlineFirst April 15, 2016.
Figure 8.Dysregulation of WNT5A/ROR2 signaling in esophageal adenocarcinoma.Schematics illustrate the speculated deregulation ofWNT5A/ROR2 signalingin esophageal adenocarcinoma. Loss of WNT5A with concurrent ROR2overexpression promotes tumor growth in esophageal adenocarcinoma, highlikely via activation of b-catenin signaling and WNT5A-independentmechanisms downstream of ROR2 receptor.
Lyros et al.
Mol Cancer Res; 14(7) July 2016 Molecular Cancer Research658
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484

References1. Pohl H, Sirovich B, Welch HG. Esophageal adenocarcinoma incidence:
are we reaching the peak? Cancer Epidemiol Biomarkers Prev 2010;19:1468–70.
2. Tougeron D, Richer JP, Silvain C. Management of esophageal adenocar-cinoma. J Visc Surg 2011;148:e161–70.
3. Jankowski JA, Harrison RF, Perry I, Balkwill F, Tselepis C. Barrett's meta-plasia. Lancet 2000;356:2079–85.
4. Hvid-Jensen F, Pedersen L, Drewes AM, Sorensen HT, Funch-Jensen P.Incidence of adenocarcinoma among patients with Barrett's esophagus. NEngl J Med 2011;365:1375–83.
5. Clevers H. Wnt/beta-catenin signaling in development and disease. Cell2006;127:469–80.
6. Moon RT, Kohn AD, De Ferrari GV, Kaykas A. WNT and beta-cateninsignalling: diseases and therapies. Nat Rev Genet 2004;5:691–701.
7. Ishitani T, Kishida S, Hyodo-Miura J, UenoN, Yasuda J, WatermanM, et al.The TAK1-NLK mitogen-activated protein kinase cascade functions in theWnt-5a/Ca(2þ) pathway to antagonize Wnt/beta-catenin signaling. MolCell Biol 2003;23:131–9.
8. Lee JM, Kim IS, Kim H, Lee JS, Kim K, Yim HY, et al. RORalpha attenuatesWnt/beta-catenin signaling by PKCalpha-dependent phosphorylation incolon cancer. Mol Cell 2010;37:183–95.
9. Clement G, Braunschweig R, Pasquier N, Bosman FT, Benhattar J. Altera-tions of the Wnt signaling pathway during the neoplastic progression ofBarrett's esophagus. Oncogene 2006;25:3084–92.
10. Choi YW, Heath EI, Heitmiller R, Forastiere AA, Wu TT. Mutations in beta-catenin and APC genes are uncommon in esophageal and esophagogastricjunction adenocarcinomas. Mod Pathol 2000;13:1055–9.
11. Moyes LH, McEwan H, Radulescu S, Pawlikowski J, Lamm CG, Nixon C,et al. Activation of Wnt signalling promotes development of dysplasia inBarrett's oesophagus. J Pathol 2012;228:99–112.
12. Clement G, Guilleret I, He B, Yagui-Beltran A, Lin YC, You L, et al.Epigenetic alteration of the Wnt inhibitory factor-1 promoter occurs earlyin the carcinogenesis of Barrett's esophagus. Cancer Sci 2008;99:46–53.
13. Zou H, Molina JR, Harrington JJ, Osborn NK, Klatt KK, Romero Y, et al.Aberrant methylation of secreted frizzled-related protein genes in esoph-ageal adenocarcinoma and Barrett's esophagus. Int J Cancer 2005;116:584–91.
14. Kikuchi A, Yamamoto H, Sato A, Matsumoto S. WNT5a: its signalling,functions and implication in diseases. Acta Physiol 2012;204:17–33.
15. Oishi I, Suzuki H, Onishi N, Takada R, Kani S, Ohkawara B, et al. Thereceptor tyrosine kinase Ror2 is involved in non-canonical WNT5a/JNKsignalling pathway. Genes Cells 2003;8:645–54.
16. Kremenevskaja N, von Wasielewski R, Rao AS, Schofl C, Andersson T,Brabant G. Wnt-5a has tumor suppressor activity in thyroid carcinoma.Oncogene 2005;24:2144–54.
17. Blanc E, Roux GL, Benard J, Raguenez G. Low expression of Wnt-5a gene isassociated with high-risk neuroblastoma. Oncogene 2005;24:1277–83.
18. Jonsson M, Dejmek J, Bendahl PO, Andersson T. Loss of Wnt-5a protein isassociated with early relapse in invasive ductal breast carcinomas. CancerRes 2002;62:409–16.
19. Dejmek J, Dejmek A, Safholm A, Sjolander A, Andersson T.Wnt-5a proteinexpression in primary dukes B colon cancers identifies a subgroup ofpatients with good prognosis. Cancer Res 2005;65:9142–6.
20. Iozzo RV, Eichstetter I, Danielson KG. Aberrant expression of the growthfactor Wnt-5A in human malignancy. Cancer Res 1995;55:3495–9.
21. Weeraratna AT, Jiang Y, Hostetter G, Rosenblatt K, Duray P, BittnerM, et al.WNT5a signaling directly affects cell motility and invasion of metastaticmelanoma. Cancer Cell 2002;1:279–88.
22. Wright TM, Brannon AR, Gordan JD, Mikels AJ, Mitchell C, Chen S, et al.Ror2, a developmentally regulated kinase, promotes tumor growth poten-tial in renal cell carcinoma. Oncogene 2009;28:2513–23.
23. Harada H, Nakagawa H, Oyama K, Takaoka M, Andl CD, Jacobmeier B,et al. Telomerase induces immortalization of human esophageal kerati-nocytes without p16INK4a inactivation. Mol Cancer Res 2003;1:729–38.
24. Lyros O, Rafiee P, Nie L, Medda R, Jovanovic N, Otterson MF, et al. Wnt/beta-catenin signaling activation beyond robust nuclear beta-catenin accu-mulation in nondysplastic Barrett's esophagus: regulation via Dickkopf-1.Neoplasia 2015;17:598–611.
25. Medda R, Lyros O, Schmidt JL, Jovanovic N, Nie L, Link BJ, et al. Antiinflammatory and anti angiogenic effect of black raspberry extract onhuman esophageal and intestinal microvascular endothelial cells. Micro-vasc Res 2015;97:167–80.
26. Rafiee P, Johnson CP, Li MS, Ogawa H, Heidemann J, Fisher PJ, et al.Cyclosporine A enhances leukocyte binding by human intestinal micro-vascular endothelial cells through inhibition of p38 MAPK and iNOS.Paradoxical proinflammatory effect on the microvascular endothelium.J Biol Chem 2002;277:35605–15.
27. Ma PC, TretiakovaMS, Mackinnon AC, Ramnath N, Johnson C, Dietrich S,et al. Expression and mutational analysis of MET in human solid cancers.Genes Chromosomes Cancer 2008;47:1025–37.
28. Ying J, Li H, Yu J, Ng KM, Poon FF, Wong SC, et al. WNT5A exhibits tumor-suppressive activity through antagonizing the Wnt/beta-catenin signaling,and is frequently methylated in colorectal cancer. Clin Cancer Res2008;14:55–61.
29. DengG, Li ZQ, ZhaoC, Yuan Y,NiuCC, ZhaoC, et al.WNT5A expression isregulated by the status of its promoter methylation in leukaemia and caninhibit leukemic cell malignant proliferation. Oncol Rep 2011;25:367–76.
30. Katoh M, Katoh M. Cross-talk of WNT and FGF signaling pathways atGSK3beta to regulate beta-catenin and SNAIL signaling cascades. CancerBiol Ther 2006;5:1059–64.
31. Endo M, Nishita M, Fujii M, Minami Y. Insight into the role of WNT5a-induced signaling in normal and cancer cells. Int Rev Cell Mol Biol2015;314:117–48.
32. Kikuchi A, Yamamoto H. Tumor formation due to abnormalities in thebeta-catenin-independent pathway of Wnt signaling. Cancer Sci 2008;99:202–8.
33. Olson DJ, Gibo DM. Antisense wnt-5a mimics wnt-1-mediated C57MGmammary epithelial cell transformation. Exp Cell Res 1998;241:134–41.
34. Olson DJ, Gibo DM, Saggers G, Debinski W, Kumar R. Reversion ofuroepithelial cell tumorigenesis by the ectopic expression of humanwnt-5a. Cell Growth Differ 1997;8:417–23.
35. Kurayoshi M, Oue N, Yamamoto H, Kishida M, Inoue A, Asahara T, et al.Expression of Wnt-5a is correlated with aggressiveness of gastric cancer bystimulating cell migration and invasion. Cancer Res 2006;66:10439–48.
36. Rebagay G, Yan S, Liu C, Cheung NK. ROR1 and ROR2 in humanmalignancies: potentials for targeted therapy. Front Oncol 2012;2:34.
37. Enomoto M, Hayakawa S, Itsukushima S, Ren DY, Matsuo M, Tamada K,et al. Autonomous regulation of osteosarcoma cell invasiveness byWNT5a/Ror2 signaling. Oncogene 2009;28:3197–208.
38. O'Connell MP, Fiori JL, Xu M, Carter AD, Frank BP, Camilli TC, et al. Theorphan tyrosine kinase receptor, ROR2, mediates WNT5a signaling inmetastatic melanoma. Oncogene 2010;29:34–44.
39. Mikels A,Minami Y,Nusse R. Ror2 receptor requires tyrosine kinase activityto mediate WNT5a signaling. J Biol Chem 2009;284:30167–76.
40. Mikels AJ, Nusse R. Purified WNT5a protein activates or inhibits beta-catenin-TCF signaling depending on receptor context. PLoS Biol 2006;4:e115.
41. Bitler BG, Nicodemus JP, Li H, Cai Q, Wu H, Hua X, et al. WNT5asuppresses epithelial ovarian cancer by promoting cellular senescence.Cancer Res 2011;71:6184–94.
42. Ren D, Minami Y, Nishita M. Critical role of WNT5a-Ror2 signaling inmotility and invasiveness of carcinoma cells following Snail-mediatedepithelial-mesenchymal transition. Genes Cells 2011;16:304–15.
43. NishitaM, Yoo SK, Nomachi A, Kani S, SougawaN, Ohta Y, et al. Filopodiaformation mediated by receptor tyrosine kinase Ror2 is required forWNT5a-induced cell migration. J Cell Biol 2006;175:555–62.
44. Barna G, Mihalik R, Timar B, Tombol J, Csende Z, Sebestyen A, et al. ROR1expression is not a uniquemarker of CLL. Hematol Oncol 2011;29:17–21.
www.aacrjournals.org Mol Cancer Res; 14(7) July 2016 659
Noncanonical WNT5A-ROR2 Axis in Esophageal Adenocarcinoma
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484

2016;14:647-659. Published OnlineFirst April 15, 2016.Mol Cancer Res Orestis Lyros, Linghui Nie, Tami Moore, et al. Progression of Barrett-Associated Esophageal AdenocarcinomaDysregulation of WNT5A/ROR2 Signaling Characterizes the
Updated version
10.1158/1541-7786.MCR-15-0484doi:
Access the most recent version of this article at:
Cited articles
http://mcr.aacrjournals.org/content/14/7/647.full#ref-list-1
This article cites 44 articles, 13 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/14/7/647To request permission to re-use all or part of this article, use this link
on January 22, 2020. © 2016 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2016; DOI: 10.1158/1541-7786.MCR-15-0484










![Research Paper The prognostic value of tumor-stromal ratio combined with TNM … · 2020. 12. 12. · cancer[17–21], gastric cancer[22–24], and esophageal adenocarcinoma[25,26]](https://img.dokumen.tips/doc/110x75/60bd58a2f5e1cb6dbc12db8c/research-paper-the-prognostic-value-of-tumor-stromal-ratio-combined-with-tnm-2020.jpg)

![RoR2 functions as a noncanonical Wnt receptor that ... · pocampus of mice (Fig. 1A; Allen Mouse Brain Atlas [21]). RoR2 mRNA is also detected in juvenile rats by using total mRNA](https://img.dokumen.tips/doc/110x75/5f9d6ac1cf40e87f4c668604/ror2-functions-as-a-noncanonical-wnt-receptor-that-pocampus-of-mice-fig-1a.jpg)
















