Embed Size (px)
Citation preview

Dominance of Frankia strains in stands of Alnusincana subsp. rugosa and Myrica pensylvanica1
Michael L. Clawson, Jeffrey Gawronski, and David R. Benson
Abstract: To address issues of dominance and diversity ofFrankia spp. strains, we sequenced 16S rRNA genes fromroot nodules and strains collected fromAlnus incanasubsp.rugosa (Du Roi) R.T. Clausen andMyrica pensylvanicaLoisel. stands. Of 22 strains isolated previously fromA. incana, 16 had the same partial rDNA sequence; the remaining6 strains composed five additional groups. The groups identified by 16S rDNA analysis corresponded to phenotypicgroups established previously by one- and two-dimensional polyacrylamide gel analysis, colony and hyphalmorphology, and carbon source utilization patterns. Dominance of one strain was also evident in nodules collected froma singleM. pensylvanicastand. The dominant strain had a partial 16S rDNA sequence identical to that ofFrankia alnistrain CpI1.
Key words: Frankia, Myrica, Alnus, actinorhizal, root nodules, nitrogen fixation, symbiosis, 16S rRNA.
Résumé: Afin d’examiner les questions de dominance et de diversité chez les souches deFrankia, les auteurs ontséquencé le gène 16S rARN à partir de nodules racinaires et de souches récoltées dans des peuplements d’Alnusincana sp. rugosa (Du Roi) R.T. Clausen et deMyrica pensylvanicaLoisel. Sur 22 souches préalablement isolées del’ A. incana, 16 montrent la même séquence partielle, les 6 autres constituant cinq autres groupes additionnels. Lesgroupes identifiés à l’aide de l’analyse du 16 rARN correspondent aux groupes phénotypiques préalablement établis paranalyse en une- et deux-dimensions sur gel de polyacrylamide, par la morphologie des hyphes et des colonies, et parles patrons d’utilisation des sources de carbone. La dominance d’une souche est également évidente chez des nodulesrécoltés dans un même peuplement deM. pensylvanica. La souche dominante montre une séquence partielle du 16SrARN identique à celle de la souche CpI1 duFrankia alni.
Mots clés: Frankia, Myrica, Alnus, actinorhizien, nodules racinaires, fixation de l’azote, symbiose, 16S rARN.
[Traduit par la Rédaction] Clawson et al. 1207
Introduction
Actinorhizal plants occupy soils that include well-drainedsand in dune systems, water-saturated soils bordering bogsand lakes, dry stony soil, glacial till, and chapparal (4).The diversity ofFrankia spp. strains that infect actinorhizalplants and form root nodules in these environments has beena subject of study for the past several years. Many ap-proaches have been taken to assess the biochemical diversityof cultured strains isolated from plants (reviewed in (4)).Relatively fewer attempts have been made to assess directlythe diversity ofFrankia spp. strains in root nodules collectedin the field. The development of rapid methods of PCR(polymerase chain reaction) amplifying microsymbiont DNAfrom nodules permits the simultaneous estimation of diver-sity and relatedness and provides a means to conduct molec-ular ecological studies onFrankia to address issues suchas dominance, competition, distribution, and host range ofstrains (5, 8, 9, 16, 18).
A drawback to using a molecular approach to characterizeunisolated strains is that the relationship between DNA se-quence variability and phenotypic variability is not obvious.We previously isolated severalFrankia spp. strains from astand ofAlnus incanasubsp.rugosa(Du Roi) R.T. Clausenand estimated their diversity using one- and two-dimensionalprotein gel electrophoresis analysis of whole cell proteins,carbon utilization patterns, and hyphal and colony morphol-ogy (1–3). The conclusion from that study was that one setof strains (gel group A) was present in 80% (29 out of 36) ofthe nodules examined. The remaining isolates formed fouradditional gel groups designated B, C, D, and F. Thus, weproposed that in the particular stand of alder under study,one strain comes to dominance and is more competitive inthat environment. At that time it was not possible to assessthe relationships between strains from different gel groups.In this study we have returned to several of the original iso-lates and PCR amplified and sequenced part of the 16SrRNA gene to determine the relationships between many ofthe same strains. In addition, we have used the approach toassess whether one strain ofFrankia sp. becomes dominantin nodules collected from a stand ofMyrica pensylvanicaLoisel.
Methods
Source of strains and nodulesTables 1 and 2 list the sources of strains and root nodules used
in this study.Frankia strains that had been obtained from a 10 ×
Can. J. Bot.77: 1203–1207 (1999) © 1999 NRC Canada
1203
Received August 15, 1998.
M.L. Clawson, J. Gawronski, and D.R. Benson.2
Department of Molecular and Cell Biology, U-44, Universityof Connecticut, Storrs, CT 02629-3044, U.S.A.
1This paper was presented at the 11th InternationalConference onFrankia and Actinorhizal Plants, June 7–11,1998, University of Illinois at Urbana–Champaign.
2Author to whom all correspondence should be addressed.e-mail: [email protected]
J:\cjb\cjb77\cjb-09\B99-070.vpMonday, December 20, 1999 9:22:21 AM
Color profile: DisabledComposite Default screen

10 m study site in 1981 were suspended in 10% glycerol in waterand stored frozen at –80°C for 14 years. ThreeA. incanaroot nod-ules were collected several kilometres from the study site, and nod-ules from M. pensylvanicawere collected immediately adjacent(within a few metres) to the study site.
Isolation of DNA and PCRTo recover DNA from strains, a loopful of the mycelia was
suspended in 100µL TE (10 mM Tris-HCl, 1 mM Na2EDTA(pH 7.6)) and 100µL 0.2 M NaOH – 1% (w/v) SDS. The suspen-sion was boiled for 7 min, and the hyphal debris was removed bycentrifugation. DNA was precipitated from the supernatant by am-monium acetate – ethanol precipitation as previously described (5).
The procedures used for dissecting nodules, isolating DNA, andobtaining PCR product have been described (5). Briefly, noduleswere rinsed on a nylon screen with a stream of cold tap water. Theperiderm of single lobes was removed while viewing with a dis-secting microscope in 0.5-mL of sterile, freshly made TEA buffer(10 mM Tris-HCl, 1 mM Na2EDTA, 20 mM ascorbic acid, pH 7.6)plus solid polyvinylpolypyrrolidone (PVPP; 10 mg/mL). Thepeeled nodule was rinsed in two additional 0.5-mL drops of TEAbuffer with PVPP. The nodule was transferred to a third 0.5-mLdrop of TEA buffer with PVPP, and the contents of infected cellswere released with a sterile scalpel and forceps. Vesicle clusterswere collected and DNA was extracted as previously described (5).
Frankia 16S rDNA was amplified using PCR. To avoid PCR er-rors, each template was amplified in four tubes containing 40µLeach; the contents of the tubes were combined, for a total volumeof 160µL. Each reaction contained the following (final concentra-tions): 12µL DNA template, 0.2µM of the universal primer fD1(17), 0.2µM of the primer rDB1 (5), 200µM each of dATP, dCTP,dGTP, and dTTP, 2.5 mM MgCl2, 3.75 U of AmpliTaq DNA poly-merase (Perkin–Elmer Corp., Norwalk, Conn.), 100µM sterile bo-vine albumin (Sigma Chemical Company, St. Louis, Mo.), 10 mMTris-HCl (pH 8.3), and 50 mM KCl. The primers fD1 and rDB1flank positions 28 to 419 on theEscherichia coli16S rDNA (5).
The PCR was done in a Perkin–Elmer model 2400 thermalcycler using the following protocol: initial denaturation at 95°C for2 min was followed by 35 cycles of denaturation at 94°C for 30 s,primer annealing at 57°C for 30 s, and extension at 72°C for1 min. After 35 cycles, the reactions were held at 72°C for 7 min.Following PCR, the samples were centrifuged for 5 min and thesupernatant was placed in a new microfuge tube. The ampliconswere purified using a Qiagen (Chatsworth, Calif.) PCR purificationkit according to the manufacturer’s protocol, and the DNA wasquantified with a DyNA Quant 200 DNA fluorometer (HoeferPharmacia Biotech Inc., San Francisco, Calif.). The ampliconswere cycle sequenced using an ABI cycle sequencing kit (Perkin–Elmer) with 2 pmol of primer and 100 fmol of dsDNA. Reactionproducts were analyzed on an Applied Biosystems (ABI) Prismsequencer (Perkin–Elmer Norwalk, Conn.). Sequencing was con-ducted in both directions using primers binding in the middle ofthe template, and at both ends (5). Phylogenetic analysis was con-ducted using PHYLIP (11).
Results
Diversity of Frankia from A. incana subsp. rugosaPartial 16S ribosomal RNA gene sequences were obtained
from 22 Frankia strains that had been isolated from separateA. incanaroot nodules collected from a 10 × 10 m section ofan alder stand in 1981 (Table 1). Three additional sequenceswere amplified directly fromA. incana root nodules col-lected several kilometres from the alder stand under study,and three more sequences were amplified from root nodules
of M. pensylvanicacollected immediately adjacent to the al-der stand.
The DNA sequences were aligned with a representativegroup of Frankia spp. strain sequences that define the threemajor clades of strains inhabiting actinorhizal nodules (Ta-ble 1) (15). A dendrogram that establishes the relationship ofthese sequences with each other and with otherFrankia sp.strains is shown in Fig. 1. All sequences (numbered 1–6in Fig. 1) from alder nodules and isolates clustered withingroup 1, sometimes referred to as the “Alnusclade.” Sixteenisolates had the same 16S rDNA sequence (sequence 1 inFig. 1); these isolates correspond to protein gel group Aidentified in previous work as characterizing the dominantnodule inhabitant in the stand (Table 2) (2, 3). Two isolates(Ai21 and Ai23-2; sequence 2 in Fig. 1) had the same se-quence as the type strainFrankia alni strain CpI1 originallyisolated fromComptonia peregrinaL. (Coult.) root nodules(7) and belonged to the same gel group (gel group C) asCpI1 (Table 2) (3). Two other isolates had unique sequencesand corresponded to gel groups D (Ai17-1; sequence 6 inFig. 1) and F (Ai57-2; sequence 3 in Fig. 1); one isolate(Ai58-2; sequence 4 in Fig. 1) that had not been analyzed bySDS–PAGE was closely related to organisms from gel groupD (sequence 6 in Fig. 1).
ThreeA. incananodules collected at some distance fromthe stand under study yielded one sequence identical to thatof gel group A strains and two unique sequences (Ai nod1and Ai nod3 in Fig. 1) not found in any of the strains iso-lated from the stand under study. ThreeM. pensylvanicanodules from plants growing adjacent to the alder stand con-tained sequences identical to the dominant strain isolatedfrom the alder nodules (sequence 1) belonging to gel groupA (Table 2).
Diversity of Frankia in an M. pensylvanicastandMyrica pensylvanicais known to be infected by a diverse
array of Frankia strains in the field (6, 8). To determinewhether local dominance of individual strains occurs withM. pensylvanica, partial 16S rDNA fromFrankia strains wasamplified from 12M. pensylvanicanodules collected from astand growing in Elizabeth Park in Hartford, Conn., with noother actinorhizal plants growing in the vicinity. Nine of the12 sequences were identical (Mp nod4-12), and were thesame as the sequence fromF. alni strain CpI1 and othermembers of gel group C from the alder stand describedabove (sequence 2 in Fig. 1). Two sequences differed fromthe majority at only one nucleotide position (Mp nod13 andMp nod14 in Fig. 1), and one sequence was typical ofFrankia that infect members of the Elaeagnaceae andRhamnaceae (Mp nod15 in Fig. 1).
Discussion
Previous work using SDS–PAGE analysis of isolatedFrankia strains led to the conclusion that the diversity ofstrains in a single established alder stand was low and that asingle strain or set of similar strains dominated the nodulepopulation (2, 3). Using a molecular ecological approach,we have confirmed that conclusion, and have shown furtherthat a different strain or set of closely related strains domi-
© 1999 NRC Canada
1204 Can. J. Bot. Vol. 77, 1999
J:\cjb\cjb77\cjb-09\B99-070.vpMonday, December 20, 1999 9:22:21 AM
Color profile: DisabledComposite Default screen
nates in nodules collected from one particular stand ofM. pensylvanica. Local dominance ofFrankia spp. strainshas also been shown to occur inAlnus viridis(Vill.) Lam. &DC. root nodules collected in France (16), where distinctpatchiness of spore-positive and spore-negative nodules wasnoted and relatively homogeneous 16S rDNA sequenceswere amplified from nodules within patches. It is not clearwhether such dominance is promoted by soil characteristics,plant selection, dissemination capacity, or a combination offactors. However, these issues are amenable to experimentalapproach.
The present study has also demonstrated a clear correla-tion between groups ofFrankia isolates created by SDS–PAGE, morphology, and carbon source utilization compari-sons and 16S rDNA sequence analysis. This correlation vali-dates using either approach for grouping strains or sym-bionts in ecological studies. The 16S rDNA sequencing hasthe advantage of being faster and allowing the placement of
unknown strains in a phylogenetic framework but allowsonly inferences to be made about phenotypic characteristics.Using isolated strains for diversity studies permits pheno-typic characterization but is much slower and depends onthe unpredictable cultivation ofFrankia spp. strains fromnodules.
Two apparent conflicts between the approaches arose inthis work. In the first, Ai20-4 was previously grouped withCpI1, Ai21, and Ai23-2 in gel group C but 16S rDNA se-quence analysis revealed here that it diverged from the otherstrains (Fig. 1). However, Ai20-4 (sequence group 5) clus-ters close to the 16S rDNA group defined by Ai21, Ai23-3,and CpI1 (sequence group 2). In the second conflict, Ai57-2had been previously grouped in gel group F using one-dimensional PAGE (3) and was moved to gel group A basedon two-dimensional PAGE (2). We find here that Ai57-2 hasa different 16S rDNA sequence (sequence group 3) thanother members of gel group A (sequence group 1 in Fig. 1)
© 1999 NRC Canada
Clawson et al. 1205
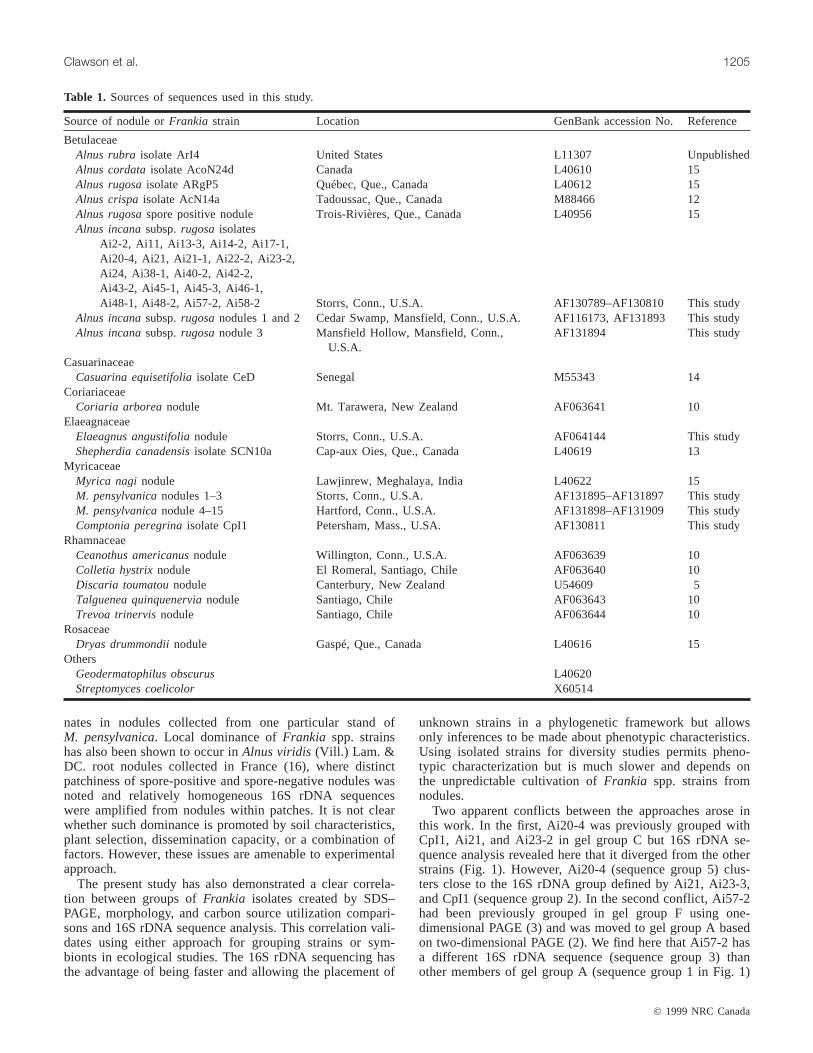
Source of nodule orFrankia strain Location GenBank accession No. Reference
BetulaceaeAlnus rubra isolate ArI4 United States L11307 UnpublishedAlnus cordataisolate AcoN24d Canada L40610 15Alnus rugosaisolate ARgP5 Québec, Que., Canada L40612 15Alnus crispaisolate AcN14a Tadoussac, Que., Canada M88466 12Alnus rugosaspore positive nodule Trois-Rivières, Que., Canada L40956 15Alnus incanasubsp.rugosa isolates
Ai2-2, Ai11, Ai13-3, Ai14-2, Ai17-1,Ai20-4, Ai21, Ai21-1, Ai22-2, Ai23-2,Ai24, Ai38-1, Ai40-2, Ai42-2,Ai43-2, Ai45-1, Ai45-3, Ai46-1,Ai48-1, Ai48-2, Ai57-2, Ai58-2 Storrs, Conn., U.S.A. AF130789–AF130810 This study
Alnus incanasubsp.rugosanodules 1 and 2 Cedar Swamp, Mansfield, Conn., U.S.A. AF116173, AF131893 This studyAlnus incanasubsp.rugosanodule 3 Mansfield Hollow, Mansfield, Conn.,
U.S.A.AF131894 This study
CasuarinaceaeCasuarina equisetifoliaisolate CeD Senegal M55343 14
CoriariaceaeCoriaria arborea nodule Mt. Tarawera, New Zealand AF063641 10
ElaeagnaceaeElaeagnus angustifolianodule Storrs, Conn., U.S.A. AF064144 This studyShepherdia canadensisisolate SCN10a Cap-aux Oies, Que., Canada L40619 13
MyricaceaeMyrica nagi nodule Lawjinrew, Meghalaya, India L40622 15M. pensylvanicanodules 1–3 Storrs, Conn., U.S.A. AF131895–AF131897 This studyM. pensylvanicanodule 4–15 Hartford, Conn., U.S.A. AF131898–AF131909 This studyComptonia peregrinaisolate CpI1 Petersham, Mass., U.SA. AF130811 This study
RhamnaceaeCeanothus americanusnodule Willington, Conn., U.S.A. AF063639 10Colletia hystrixnodule El Romeral, Santiago, Chile AF063640 10Discaria toumatounodule Canterbury, New Zealand U54609 5Talguenea quinquenervianodule Santiago, Chile AF063643 10Trevoa trinervisnodule Santiago, Chile AF063644 10
RosaceaeDryas drummondiinodule Gaspé, Que., Canada L40616 15
OthersGeodermatophilus obscurus L40620Streptomyces coelicolor X60514
Table 1. Sources of sequences used in this study.
J:\cjb\cjb77\cjb-09\B99-070.vpMonday, December 20, 1999 9:22:22 AM
Color profile: DisabledComposite Default screen

© 1999 NRC Canada
1206 Can. J. Bot. Vol. 77, 1999
yces sp.
4
Ai nod1
ACN14a
6
Ai nod3
2, Mp nod4 nod12, Cpl1
Mp nod13,14
5
Ar I4
3
ARgP5
1, A i nod2,Mp nod1-nod3
CeD
M. nagi nod
Col l et i a nod
Elaeagnus nod
SCN14a
Tr ev oa nod
Mp nod4- nod15
Talguenea nod
Discar i a nod
Dryas nod
Ceanothus nod
Cor iar ia nod
Geodermatophilus sp.
Streptom
1
3
2
Fig. 1. Dendrogram of partial 16S rDNA sequences fromFrankia spp. strains in culture and in actinorhizal root nodules. The tree wasconstructed using the UPGMA (unweighted pair–group method using arithmetic averages) program in PHYLIP. Underlined sequencesare from this study. Numbers 1–6 refer to the groups of strains listed in Table 2. Ai nod1-nod3 and Mp nod1-nod15 refer to sequencesfound in A. incanasubsp.rugosaand M. pensylvanicaroot nodules, respectively, listed in Table 1. The numbering of the major groups1–3 follows the convention of Normand et al. (15).
Strains16S rDNAsequencea
Gelgroupb
Hyphalmorphologyb
Colonymorphologyb
Carbon sourcesfor growthb
Ai11, Ai21-1, Ai22-2, 1 A 1 Tan-orange, raised A+Py–S–Tw–Pr+
Ai24, Ai43-2, Ai2-2,Ai13-3, Ai14-2, Ai38-1,Ai40-2, Ai42-2, Ai45-1,Ai45-3, Ai46-1, Ai48-1,Ai48-2
Ai21, Ai23-2, CpI1 2 C 2 White, subsurface A+Py+S+Tw+Pr+
Ai57-2 3 F or A 1 Tan-orange, raised A+Py+S–Tw–Pr+
Ai58-2 4 nd nd nd ndAi20-4 5 C 2 White, subsurface A+Py+S+Tw+Pr+
Ai17-1 6 D 2 Tan-orange, raised A+Py–S–Tw+Pr+
aNumber corresponds to sequence numbers in Fig. 1.bData are from references 1–3. Hyphal morphology of type 1 was highly branched, sinuate; type 2 was smooth, slightly
sinuate in liquid culture (1). Colony morphology was determined on FMC plus acetate agar (3). Carbon sources used forgrowth include acetate (A), pyruvate (Py), succinate (S), Tween 80 (Tw), and propionate (Pr). A plus (+) or minus (–) afterthe carbon source denotes growth or no growth, respectively.
Table 2. Summary ofA. incanastrains grouped by rDNA sequence and phenotypic properties.
J:\cjb\cjb77\cjb-09\B99-070.vpMonday, December 20, 1999 9:22:23 AM
Color profile: DisabledComposite Default screen

© 1999 NRC Canada
but that the two sequence groups cluster closely together inFig. 1. Thus, strains that have very closely related 16S se-quences may be difficult to discriminate on the basis of pro-tein gel analysis alone.
It is noteworthy that the same, or a closely related, strainthat dominates in the alder nodules is also found in threenodules collected from adjacentM. pensylvanicaroots. Thisobservation suggests that the soil population under the stand,as well as the alder nodules, is dominated by a single strain,or group of closely related strains, ofFrankia. It is alsointeresting to note that strains with partial 16S rDNA se-quences identical to theF. alni strain CpI1 are present inboth stands but come to dominance only under the stand ofM. pensylvanicathat we studied. It will be intriguing to de-termine the identity of dominant strains in additional standsof both A. incanasubsp.rugosaandM. pensylvanicagrow-ing in different soil environments to assess whether particu-lar ecotypes have adapted to different soil conditions.
Acknowledgment
This work was supported by grant #95-37305-2230 fromthe United States Department of Agriculture NationalResearch Initiative Competitive Grants Program (USDA–NRICGP).
References
1. Benson, D.R. 1982. Isolation ofFrankia strains from alderactinorhizal root nodules. Appl. Environ. Microbiol.44: 461–465.
2. Benson, D.R., Buchholz, S.E., and Hanna, D.G. 1984. Identifi-cation of Frankia strains by two-dimensional polyacrylamidegel electrophoresis. Appl. Environ. Microbiol.47: 489–494.
3. Benson, D.R., and Hannah, D. 1983.Frankia diversity in analder stand as estimated by sodium dodecyl sulfate – poly-acrylamide gel electrophoresis of whole-cell proteins. Can. J.Bot. 61: 2919–2923.
4. Benson, D.R., and Silvester, W.B. 1993. Biology ofFrankiastrains, actinomycete symbionts of actinorhizal plants.Microbiol. Rev. 57: 293–319.
5. Benson, D.R., Stephens, D.W., Clawson, M.L., and Silvester,W.B. 1996. Amplification of 16S rRNA genes fromFrankia inroot nodules ofCeanothus griseus, Coriaria arborea, Coriariaplumosa, Discaria toumatou, and Purshia tridentata. Appl.Environ. Microbiol. 62: 2904–2909.
6. Bloom, R.A., Mullin, B.C., and Tate, R.L., III. 1989. DNA re-striction patterns and DNA–DNA solution hybridization stud-ies of Frankia isolates fromMyrica pensylvanica(bayberry).Appl. Environ. Microbiol.55: 2155–2160.
7. Callaham, D., DelTredici, P., and Torrey, J.G. 1978. Isolationand cultivation in vitro of the actinomycete causing rootnodulation in Comptonia. Science (Washington, D.C.),199:899–902.
8. Clawson, M.L., and Benson, D.R. 1999. Natural diversity ofFrankia in actinorhizal root nodules from promiscuous hostsin the Myricaceae. Appl. Environ. Microbiol.65. In press.
9. Clawson, M.L., Benson, D.R., Stephens, D.W., Resch, S.C.,and Silvester, W.B. 1997. TypicalFrankia infect actinorhizalplants exotic to New Zealand. N.Z. J. Bot.35: 361–367.
10. Clawson, M.L., Carú, M., and Benson, D.R. 1998. Diversity ofFrankia strains in root nodules of plants from the familiesElaeagnaceae and Rhamnaceae. Appl. Environ. Microbiol.64:3539–3543.
11. Felsenstein, J. 1993. PHYLIP (Phylogeny Inference Package)version 3.5c. Distributed by the author. Department of Genet-ics, University of Washington, Seattle.
12. Johnson, D.A. 1992. Ph.D. thesis, University of Ottawa, Ot-tawa, Ont.
13. Mort, A., Normand, P., and Lalonde, M. 1983. 2-O-methyl-D-mannose, a key sugar in the taxonomy ofFrankia. Can. J.Microbiol. 29: 993–1002.
14. Normand, P., Cournoyer, B., Simonet, P., and Nazaret, S. 1992.Analysis of a ribosomal RNA operon in the actinomyceteFrankia. Gene,111: 119–124.
15. Normand, P., Orso, S., Cournoyer, B., Jeannin, P., Chapelon,C., Dawson, J., Evtushenko, L., and Misra, A.K. 1996. Molec-ular phylogeny of the genusFrankia and related genera andemendation of family Frankiaceae. Int. J. Syst. Bacteriol.46:1–9.
16. Simonet, P., Bosco, M., Chapelon, C., Moiroud, A., andNormand, P. 1994. Molecular characterization ofFrankiamicrosymbionts from spore-positive and spore-negative nod-ules in a natural alder stand. Appl. Environ. Microbiol.60:1335–1341.
17. Weisburg, W.G., Barns, S.M., Pelletier, D.A., and Lane, D.J.1991. 16S ribosomal DNA amplification for phylogeneticstudy. J. Bacteriol.173: 697–703.
18. Wolters, D.J., Van Dijk, C., Zoetendal, E.G., and Akkermans,A.D.L. 1997. Phylogenetic characterization of ineffectiveFrankia in Alnus glutinosa(L.) Gaertn. nodules from wetlandsoil inoculants. Mol. Ecol.6: 971–981.
Clawson et al. 1207
J:\cjb\cjb77\cjb-09\B99-070.vpMonday, December 20, 1999 9:22:24 AM
Color profile: DisabledComposite Default screen





























