Embed Size (px)
Citation preview

© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim 1434-2944/05/102– 0033
SÁRKA AXMANOVÁ and MARTIN RULÍK*
Department of Ecology and Environmental Sciences, Faculty of Science, Palacky University, Slechtitelu 11, CZ – 783 71 Olomouc, Czech Republic; e-mail: [email protected]
DOC Release from Alder Leaves and Catkins duringDecomposition in a Small Lowland Stream
key words: decomposition, POM, DOC, hyporheic zone, surface water
Abstract
Alder leaves and catkins were placed in a small stream, then retrieved in weekly intervals, and ratesof release of dissolved organic carbon (DOC) and substantial DOC characteristics were determined.Decomposition rates of leaves and catkins were 0.026 d–1 and 0.011 d–1 on the streambed, and 0.017 d–1
and 0.009 d–1 in the sediments, respectively, during 5 weeks of decomposition. The rate of DOC release from leaves declined from 18.0% d–1 to 0.7% d–1, and had a higher proportion of low molecu-lar weight compounds, more saturated and contained larger humic molecules than DOC from catkins,which rate of release declined from 10.6% d–1 to 0.1% d–1. In initial stages of the decomposition, leafmaterial produced more biodegradable DOC (BDOC) than catkins. During the period of low leaf litterinput, DOC released from catkins can become an important instream source of labile DOC.
1. Introduction
Allochthonous particulate organic material (POM) is important to the ecology of streamecosystems. About 95% of the annual organic matter inputs to streams may consist of litterinputs, dominated by leaves, followed by wood (MULHOLLAND, 1981; WALLACE et al., 1995).POM that accumulates on the stream bed or is retained within bed sediments is transformedto dissolved organic matter (DOM) through leaching (e.g. PETERSEN and CUMMINS, 1974) or to fine POM by mechanical breakdown and biological processing, i.e. microbial andinvertebrate action (e.g. HIEBER and GESSNER, 2002; HUTCHENS and WALLACE, 2002). Aqua-tic hyphomycetes play a predominant role in the microbial degradation of leaves (e.g. FIND-LAY and ARSUFFI, 1989; MALTBY, 1992; GESSNER and CHAUVET, 1994) and the rate of thedegradation depends on internal (substrate quality, e.g. C/N ratio), and external (nutrientconcentrations in the water, temperature), factors (WEBSTER and BENFIELD, 1986; BÄR-LOCHER, 1990; ROBINSON and GESSNER, 2000).
During autumn, leaf litter leaching contributes 40% of DOC inputs in streams (MEYER
et al., 1998). The initial plant leachate is characterized by a predominance of low molecularweight dissolved organic carbon (DOC) (MCARTHUR and RICHARDSON, 2002); 60% of leafleachate DOC was <10,000 nominal molecular weight (nmw) (KAPLAN and BOTT, 1983;1985). Carbohydrates and phenolics are the most important functional groups (KAPLAN andBOTT, 1983; ENGELHAUPT and BIANCHI, 2001). DOC, which is slowly leached during further decomposition of recalcitrant detrital material, is likely to be rich in fulvic and humicacids. Although the rate at which DOC is leached from recalcitrant material in streams is
Internat. Rev. Hydrobiol. 90 2005 1 33–41
DOI: 10.1002/iroh.200310715
* Corresponding author

very low, the amount of material stored in streams is often so great that this can be a majorsource of DOC (MEYER, 1990).
Although there have been published many studies on transformations and modificationsof leaves in the surface stream water (e.g. GESSNER and CHAUVET, 1994; MEYER et al., 1998;ROBINSON and GESSNER, 2000; HIEBER and GESSNER, 2002) and laboratory leaching studies(e.g. MCARTHUR and RICHARDSON, 2002), studies on decomposition in the hyporheic sediments are limited (CROCKER and MEYER, 1987; RULÍK et al., 2001). The major purposeof the present study was to characterize DOC released during organic matter decompositionin a stream. Our objectives were i) to determine DOC released from alder leaves and catkins, ii) to compare decomposition of material on the streambed and material buried insediments and iii) to determine qualitative and quantitative changes in DOC released duringexposure in the stream.
2. Methods
Investigations were conducted in the Sitka stream, a small, unpolluted lowland stream above the cityof Olomouc in the Czech Republic. Stream width ranges between 4–6 m and substrate is composed ofgravel-sand. Alder is the dominant tree on stream banks.
Fresh leaves and catkins of alder (Alnus glutinosa) were collected at the end of August 2002, andwere air-dried in the laboratory. 1.5 ± 0.1 g of leaves and 0.5 ± 0.1 g of catkins, respectively, were plac-ed into 1-mm-mesh nylon bags. Pairs of mesh bags were put into perforated plastic boxes, which wereinserted into the stream sediment, at a depth of ca. 30–40 cm, using a steel pipe (for detailed descrip-tion see RULÍK et al. (2001). The additional boxes with bags were distributed randomly on the river bedsurface. The boxes were placed in the stream on 2 September 2002 and removed after 2, 7, 14, 21, 28and 35 days. For each sampling period, three boxes were removed from the sediment and from the surface. Each bag was carefully rinsed in distilled water and then placed in a beaker with 250 ml of car-bon-free distilled water for 24 h at in situ temperature with occasional agitation. The resultant leacha-tes were filtered through a 0.4 µm pore size membrane (GF 55, Schleicher & Schuell) to obtain theDOC solution. In order to obtain initial DOC leachates, three litterbags of both leaves and catkins whichhad not been placed in the stream, were leached in carbon-free distilled water for 24 h in the labora-tory. DOC obtained is presented as an initial DOC.
DOC concentration was measured by Pt-catalyzed high temperature combustion on a TOC FOR-MACSHT analyzer with IR detector. For each sample, 200 µl were injected at least twice, resulting in acoefficient of variation <2%.
For molecular weight determinations, a 120-ml stirred cell with membrane ultrafilters (PBGC,44.5 mm diam. Millipore, pressurized with nitrogen) was used to separate DOC < 10,000 nominal mole-cular weight (nmw) from total DOC. High-molecular-weight (HMW) DOC was retained, and low-mole-cular-weight (LMW) DOC was the fraction that passed through the membrane. Before ultrafiltration ofsamples, 240 ml of carbon-free distilled water was passed through the filter to leach residual DOC.
Absorbance was measured at 254 and 400 nm using 5 cm quartz cell in a Cecil Instruments SuperAquarius CE 9000 Double-beam Spectrophotometer against distilled water as a reference. The ab-sorbance A (λ = 254 nm) was converted to the specific absorption a254 = A254/L.DOC, where L is thecell path length in meters and DOC is the DOC concentration in grams of carbon per m3, according tothe method of HEJZLAR et al. (1990). The absorbance at 254 nm is thought to be representative of thedegree of non-saturation of humic molecules (ROSTAN and CELLOT, 1995) that constitute the bulk ofnatural DOC (VOLK et al., 1997). A254/A400 ratios were calculated to obtain evidence for changes inmolecular composition of humic compounds (EDWARDS and CRESSER, 1987).
Biodegradable DOC (BDOC) was determined in batch experiments modified from procedures bySERVAIS et al. (1989). 100 ml of DOC solution was inoculated with 2 ml of stream water filtered througha 2.4 µm pore size membrane (Z6, AMERSIL-FILPAP) and incubated in sterile 100-ml glass bottles at20 ± 0.5 °C in the dark for 30 d with occasional shaking. Subsamples were collected at the beginningand at the end of incubation for DOC determinations. The BDOC values were calculated as the differ-ence between initial and final DOC. Final DOC was considered to be refractory DOC (RDOC).
The material remaining after leaching was oven-dried at 105 °C and combusted at 450 °C for 5 h toobtain ash-free dry mass (AFDM). Carbon was assumed to be 45% of the AFDM (MEYER et al., 1981).
34 S. AXMANOVÁ and M. RULÍK
© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
Breakdown rate (k) was calculated by nonlinear regression analysis using the model AFDMt =AFDM0 e–kt, where AFDMt is the dry mass remaining at the time t, and AFDM0 is the initial dry mass.Analysis of covariance (ANCOVA) was used to test differences in breakdown rates among habitats. Alldata of leachate variables were normally distributed (Kolmogorov-Smirnov test) and were analyzedusing parametric methods. Means of both leaf leachate and catkins leachate variables and weight losses of the material were compared using one-way analyses of variance (ANOVA). All tests were considered significant at the level α ≤ 0.05. Data analyses were performed using NCSS 2000 (HINTZE,2001).
3. Results
3.1. Decomposition of Leaves
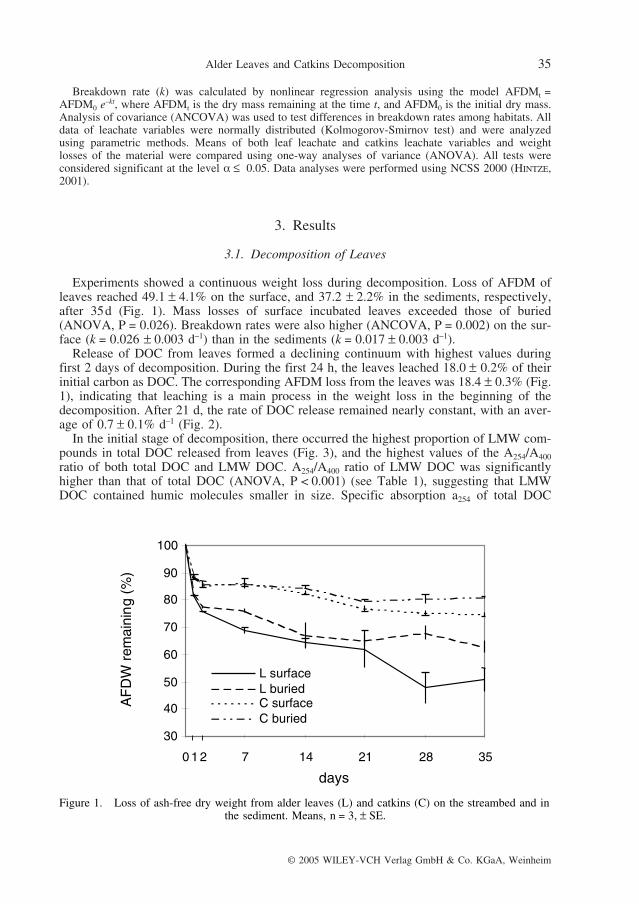
Experiments showed a continuous weight loss during decomposition. Loss of AFDM ofleaves reached 49.1 ± 4.1% on the surface, and 37.2 ± 2.2% in the sediments, respectively,after 35d (Fig. 1). Mass losses of surface incubated leaves exceeded those of buried(ANOVA, P = 0.026). Breakdown rates were also higher (ANCOVA, P = 0.002) on the sur-face (k = 0.026 ± 0.003 d–1) than in the sediments (k = 0.017 ± 0.003 d–1).
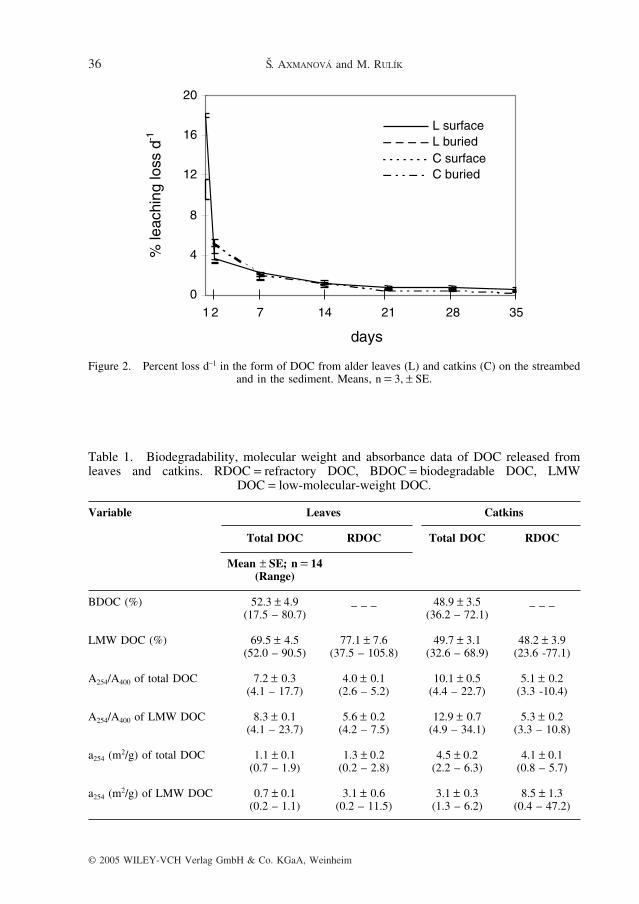
Release of DOC from leaves formed a declining continuum with highest values duringfirst 2 days of decomposition. During the first 24 h, the leaves leached 18.0 ± 0.2% of theirinitial carbon as DOC. The corresponding AFDM loss from the leaves was 18.4 ± 0.3% (Fig.1), indicating that leaching is a main process in the weight loss in the beginning of thedecomposition. After 21 d, the rate of DOC release remained nearly constant, with an aver-age of 0.7 ± 0.1% d–1 (Fig. 2).
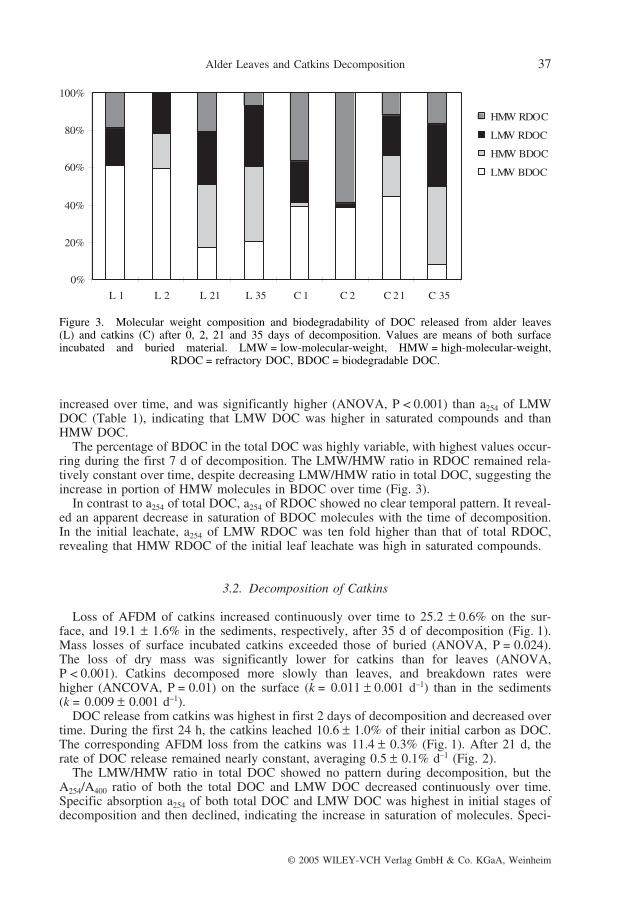
In the initial stage of decomposition, there occurred the highest proportion of LMW com-pounds in total DOC released from leaves (Fig. 3), and the highest values of the A254/A400ratio of both total DOC and LMW DOC. A254/A400 ratio of LMW DOC was significantlyhigher than that of total DOC (ANOVA, P < 0.001) (see Table 1), suggesting that LMWDOC contained humic molecules smaller in size. Specific absorption a254 of total DOC
Alder Leaves and Catkins Decomposition 35
© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
30
40
50
60
70
80
90
100
0 1 2 7 14 21 28 35
days
AF
DW
rem
aini
ng
(%)
L surfaceL buriedC surfaceC buried
Figure 1. Loss of ash-free dry weight from alder leaves (L) and catkins (C) on the streambed and inthe sediment. Means, n = 3, ± SE.
36 S. AXMANOVÁ and M. RULÍK
© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
0
4
8
12
16
20
1 2 7 14 21 28 35
days
%le
achi
nglo
ssd-1
L surfaceL buriedC surfaceC buried
Figure 2. Percent loss d–1 in the form of DOC from alder leaves (L) and catkins (C) on the streambedand in the sediment. Means, n = 3, ± SE.
Table 1. Biodegradability, molecular weight and absorbance data of DOC released fromleaves and catkins. RDOC = refractory DOC, BDOC = biodegradable DOC, LMW
DOC = low-molecular-weight DOC.
Variable Leaves Catkins
Total DOC RDOC Total DOC RDOC
Mean ± SE; n = 14(Range)
BDOC (%) 52.3 ± 4.9 _ _ _ 48.9 ± 3.5 _ _ _(17.5 – 80.7) (36.2 – 72.1)
LMW DOC (%) 69.5 ± 4.5 77.1 ± 7.6 49.7 ± 3.1 48.2 ± 3.9(52.0 – 90.5) (37.5 – 105.8) (32.6 – 68.9) (23.6 -77.1)
A254/A400 of total DOC 7.2 ± 0.3 4.0 ± 0.1 10.1 ± 0.5 5.1 ± 0.2(4.1 – 17.7) (2.6 – 5.2) (4.4 – 22.7) (3.3 -10.4)
A254/A400 of LMW DOC 8.3 ± 0.1 5.6 ± 0.2 12.9 ± 0.7 5.3 ± 0.2(4.1 – 23.7) (4.2 – 7.5) (4.9 – 34.1) (3.3 – 10.8)
a254 (m2/g) of total DOC 1.1 ± 0.1 1.3 ± 0.2 4.5 ± 0.2 4.1 ± 0.1(0.7 – 1.9) (0.2 – 2.8) (2.2 – 6.3) (0.8 – 5.7)
a254 (m2/g) of LMW DOC 0.7 ± 0.1 3.1 ± 0.6 3.1 ± 0.3 8.5 ± 1.3(0.2 – 1.1) (0.2 – 11.5) (1.3 – 6.2) (0.4 – 47.2)
increased over time, and was significantly higher (ANOVA, P < 0.001) than a254 of LMWDOC (Table 1), indicating that LMW DOC was higher in saturated compounds and thanHMW DOC.
The percentage of BDOC in the total DOC was highly variable, with highest values occur-ring during the first 7 d of decomposition. The LMW/HMW ratio in RDOC remained rela-tively constant over time, despite decreasing LMW/HMW ratio in total DOC, suggesting theincrease in portion of HMW molecules in BDOC over time (Fig. 3).
In contrast to a254 of total DOC, a254 of RDOC showed no clear temporal pattern. It reveal-ed an apparent decrease in saturation of BDOC molecules with the time of decomposition.In the initial leachate, a254 of LMW RDOC was ten fold higher than that of total RDOC,revealing that HMW RDOC of the initial leaf leachate was high in saturated compounds.
3.2. Decomposition of Catkins
Loss of AFDM of catkins increased continuously over time to 25.2 ± 0.6% on the sur-face, and 19.1 ± 1.6% in the sediments, respectively, after 35 d of decomposition (Fig. 1).Mass losses of surface incubated catkins exceeded those of buried (ANOVA, P = 0.024). The loss of dry mass was significantly lower for catkins than for leaves (ANOVA,P < 0.001). Catkins decomposed more slowly than leaves, and breakdown rates were higher (ANCOVA, P = 0.01) on the surface (k = 0.011 ± 0.001 d–1) than in the sediments (k = 0.009 ± 0.001 d–1).
DOC release from catkins was highest in first 2 days of decomposition and decreased overtime. During the first 24 h, the catkins leached 10.6 ± 1.0% of their initial carbon as DOC.The corresponding AFDM loss from the catkins was 11.4 ± 0.3% (Fig. 1). After 21 d, therate of DOC release remained nearly constant, averaging 0.5 ± 0.1% d–1 (Fig. 2).
The LMW/HMW ratio in total DOC showed no pattern during decomposition, but theA254/A400 ratio of both the total DOC and LMW DOC decreased continuously over time.Specific absorption a254 of both total DOC and LMW DOC was highest in initial stages ofdecomposition and then declined, indicating the increase in saturation of molecules. Speci-
Alder Leaves and Catkins Decomposition 37
© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim
0%
20%
40%
60%
80%
100%
L 1 L 2 L 21 L 35 C 1 C 2 C 21 C 35
HMW RDOC
LMW RDOC
HMW BDOC
LMW BDOC
Figure 3. Molecular weight composition and biodegradability of DOC released from alder leaves (L) and catkins (C) after 0, 2, 21 and 35 days of decomposition. Values are means of both surface incubated and buried material. LMW = low-molecular-weight, HMW = high-molecular-weight,
RDOC = refractory DOC, BDOC = biodegradable DOC.

fic absorption a254 of LMW DOC was significantly lower (ANOVA, P = 0.001) comparedwith that of total DOC (Table 1), suggesting that LMW DOC was dominated by saturatedcompounds and HMW DOC was of considerably lower degree of saturation.
In contrast to the total DOC, the LMW/HMW ratio in RDOC increased. It suggests thatBDOC of the initial catkins leachate was composed entirely from LMW compounds. Theportion of HMW molecules increased in BDOC over time (Fig. 3).
Compared to a254 of total DOC, a254 of RDOC decreased more sharply over time, reveal-ing an apparent decrease in saturation of BDOC molecules with the time of decomposition.Specific absorption a254 of LMW RDOC was significantly lower (ANOVA, P = 0.001) thana254 of RDOC (Table 1), suggesting that saturated compounds made up the largest percent-age of LMW RDOC, while HMW RDOC was of low degree of saturation.
4. Discussion
Alder leaves and catkins represent the predominant allochthonous source of carbon at thestudy site. The experiment was designed to simulate natural autumnal conditions, althoughit is known that drying of leaves accelerates leaching of substances and mass loss. GESSNER
and SCHWOERBEL (1989) reported no significant mass loss of fresh, senescent leaves of A. glutinosa in 6.5 d of immersion in water, while air-dried leaves lost 20%. BÄRLOCHER
(1991) found 2.5% mass loss from fresh, senescent leaves, while air-dried leaves lost 18.9%.Similarly, colonization by aquatic hyphomycetes proceeds more rapidly on dried leaves thanon fresh leaves (BÄRLOCHER, 1997). Indeed, if leaves on the stream banks dry within thefirst few days after falling, before they are windblown or flushed into stream, no error shouldensue from measuring decomposition using dried leaves. Another potential artifact is thatmesh bags may not allow full access by large invertebrates (ROBINSON and GESSNER, 2000).However, both invertebrates and hyphomycetes are important in POM decomposition parti-cularly in surface water, and they play a minor role within sediments (RULÍK et al., 2001).
Leaching losses of both material types formed a declining continuum, revealing decreaseof the DOC release with increasing age of detritus, and are in general agreement with patterns reported previously; various species of leaves lost from 5% to 33% of their initialdry mass within 28–48 h due to leaching (PETERSEN and CUMMINS, 1974; MULHOLLAND,1981; FINDLAY and ARSUFFI, 1989). Our data indicate that leaching is an important processin the weight loss of organic matter; particularly during the first 24 h of decomposition (Fig. 2), although there is evidence that microbial metabolism of water-soluble material, notsimply leaching, is responsible for the part of mass loss during this phase (PARSONS et al.,1990). BALDY and GESSNER (1997) found that 17% of the microbially mediated leaf massloss was in the form of CO2. Our findings of DOC release from older, more refractory POCalso agree with previous studies; radiolabeled lignocellulose DOC was leached from leaf litter at a rate of 0.5% d–1 after 13 weeks of incubation (MORAN and HODSON, 1989). DOCrelease from the pool of the benthic POC was about 0.1% d–1 (KAPLAN and NEWBOLD, 1993).
Comparing loss of AFDW and decomposition rates revealed significant differences in theprocessing of surface incubated and buried material. Leaf packs incubated on the surfacedecomposed faster than those buried. This is attributed e.g. to low levels of dissolved oxygen, absence of macroinvertebrates and decreased abrasion in sediments (HERBST, 1980;WEBSTER and BENFIELD, 1986). However, except only few instances, there were found nodifferences in DOC characteristics between surface incubated and buried material, revealingsimilar quality of DOC released in both habitats.
KAPLAN and BOTT (1983; 1985) reported that 60% of the initial leaf leachate DOCwas <10,000 nmw. ENGELHAUPT and BIANCHI (2001) and MCARTHUR and RICHARDSON
(2002) also found that a minority of leaf leachate DOC was in the HMW fraction. In ourstudy, LMW DOC reached 90% in the initial leaf leachate DOC. Although the percentage
38 S. AXMANOVÁ and M. RULÍK
© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

of LMW DOC declined continuously over time, it remained a dominant fraction in the DOCpool all the time. MEYER et al. (1987) reported that the fraction of leaf (oak and willow)leachate <1,000 nmw was most available to bacteria and HMW fraction (>10,000 nmw) wasgenerally least available. In the present study, BDOC in the initial leaf leachate was composed entirely of LMW compounds with high degree of saturation, which are formedmainly of carbohydrates (KAPLAN and BOTT, 1983; ENGELHAUPT and BIANCHI, 2001). Withincreasing age of detritus, BDOC released was less saturated and composed rather fromHMW DOC, on account of higher portion of HMW DOC released. As bacteria are unableto assimilate organic molecules much greater than 10,000 nmw (LOCK, 1990), these mustfirst be broken down with extracellular hydrolytic enzymes or through photolysis by UVradiation. A proportion of the cleaved molecules probably would not be transported into the cells, resulting in an increase in percentage of LMW compounds in the RDOC pool.However, biological availability of DOC is controlled not only by molecular weight, but alsoby the presence and abundance of specific functional groups, particularly nitrogenous(MEYER et al., 1987; AMON and BENNER, 1996). This suggests that the biodegradable HMWDOC released after several weeks of decomposition may be richer in organic N than HMWDOC from fresh material.
Specific absorption a254 is a frequently used estimator of the degree of non-saturation ofDOC in ecological studies (ROSTAN and CELLOT, 1995). During the decomposition of leaves, BDOC came to be composed more of HMW molecules, which are rather less saturated, i.e., of humic substances (HS). It is probable that the biodegradable fractions ofHS were actually saturated short chains of DOC, bound chemically to HS and therebyenhancing the availability of HS (MEYER et al., 1987; VOLK et al., 1997), although aroma-tic structures could be also biodegraded (KHAN et al., 1999).
Alder catkins leachates showed no decreasing tendency in biodegradability and no trendin molecular weight composition over time, in contrast to alder leaves. Regardless of DOCconcentration, BDOC made up approximately 50% of the total DOC and LMW/HMW DOCratio was about 1:1 in catkins leachates. On the other hand, a decreasing A254/A400 ratio showed an increasing size of humic molecules and decreasing a254 revealed increasing satu-ration of DOC released. ENGELHAUPT and BIANCHI (2001) have found that aromaticity inplant material was derived from both tannins and lignin. Refractory component of the DOCreleased is possibly made up of lignocellulose oligomers with resistant intermonomericbonds or of phenolic monomers (MORAN and HODSON, 1989). In this study, with increasingage of detritus, non-saturated molecules formed a dominant portion of BDOC. This result isin general agreement with patterns observed in stream water that HS dominate the BDOC(VOLK et al., 1997; MARTIN – MOUSSET et al., 1997).
Allochthonous organic matter have been recognized as an essential component of loticorganic matter dynamics (WALLACE et al., 1995; WEBSTER and MEYER, 1997) and the reten-tion capacity for POM is important for energy flow in streams. Leaves can be considered a seasonal resource, since they enter temperate streams as a pulse during autumn leaf falland decompose within several months (WEBSTER and BENFIELD, 1986). Woody catkins aremostly recalcitrant with a high lignin content, hence, microbial degradation takes severalyears (PUSCH et al., 1998). In spring, most leaf litter has decomposed and wood is the primary POM substrate for heterotrophic microbial colonization. In our study, comparingmass losses and breakdown rates of the plant material confirmed much faster decompositionrate for alder leaves than for catkins. In initial stages of the decomposition, leaf material produced more biodegradable DOC than did catkins. Once the contribution of BDOC leached from leaves decline during the period of low leaf litter input, DOC released fromcatkins can become an important instream source of labile DOC for stream microbial communities, particularly for microorganisms in hyporheic sediments, whose metabolismprofits mainly from allochthonous production (e.g. TRULLEYOVÁ et al., 2003).
Alder Leaves and Catkins Decomposition 39
© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

5. Acknowledgements
The authors are very indebted to Dr. DOUGLAS FRASER, Sheffield Hallam University, for revising theEnglish text. This research was supported by the Ministry of Education, Youth and Sports of the CzechRepublic grant # MSM 153100014.
6. References
AMON, R. M. W. and R. BENNER, 1996: Bacterial utilization of different size classes of dissolved orga-nic matter. – Limnol. Oceanogr. 41: 41–51.
BALDY, V. and M. O. GESSNER, 1997: Towards a budget of leaf litter decomposition in a first-orderwoodland stream. – C. R. Acad. Sci. Paris, Sciences de la vie/Life Sciences 320: 747–758.
BÄRLOCHER, F., 1990: Factors that delay colonization of fresh alder leaves by aquatic hyphomycetes. –Arch. Hydrobiol. 119: 249–255.
BÄRLOCHER, F., 1991: Fungal colonization of fresh and dried leaves in the River Teigh (Devon, Eng-land). – Nova Hedwigia 52: 349–357.
BÄRLOCHER, F., 1997: Pitfalls of traditional techniques when studying decomposition of vascular plantremains in aquatic habitats. – Limnetica 13: 1–11.
CROCKER, M. T. and J. L. MEYER, 1987: Interstitial dissolved organic carbon in sediments of a southernAppalachian headwater stream. – J. N. Am. Benthol. Soc. 6: 159–167.
EDWARDS, A. C. and M. S. CRESSER, 1987: Relationships between ultraviolet absorbance and total orga-nic carbon in two upland catchments. – Wat. Res. 21: 49–56.
ENGELHAUPT, E. and T. S. BIANCHI, 2001: Sources and composition of high-molecular-weight dissolvedorganic carbon in a southern Louisiana tidal stream (Bayou Trepagnier). – Limnol. Oceanogr. 46:917–926.
FINDLAY, S. E. G. and T. L. ARSUFFI, 1989: Microbial growth and detritus transformations duringdecomposition of leaf litter in a stream. – Freshw. Biol. 21: 261–269.
GESSNER, M. O. and K. J. SCHWOERBEL, 1989: Leaching kinetics of fresh leaf-litter with implications forthe current concept of leaf-processing in streams. – Arch. Hydrobiol. 115: 81–90.
GESSNER, M. O. and E. CHAUVET, 1994: Importance of stream microfungi in controlling breakdown ratesof leaf litter. – Ecology 75: 1807–1817.
HEJZLAR, J., P. TRSKA, J. SORM and G. KRUCHNOVÁ, 1990: Analýza rozpuštených organických látekv různých typech vod 13NMR, IR a UV spektroskopií [Analysis of dissolved organic matter in variouswater types by 13NMR, IR and UV spectroscopy]. – Hydrochémia 89: 205–228. (in Czech).
HERBST, G. N., 1980: Effect of burial on food value and consumption of leaf detritus by aquatic in-vertebrates in a lowland forest stream. – Oikos 35: 411–424.
HIEBER, M. and M. O. GESSNER, 2002: Contribution of stream detrivores, fungi, and bacteria to leaf breakdown based on biomass estimates. – Ecology 83: 1026–1038.
HINTZE, J., 2001: NCSS and PASS. Number Cruncher Statistical Systems. Kaysville, Utah.HUTCHENS, J. J. JR. and B. WALLACE, 2002: Ecosystem linkages between southern Appalachian head-
water streams and their banks: leaf litter breakdown and invertebrate assemblages. – Ecosystems 5:80–91.
KAPLAN, L. A. and T. L. BOTT, 1983: Microbial heterotrophic utilization of dissolved organic matter ina piedmont stream. – Freshw. Biol. 13: 363–377.
KAPLAN, L. A. and T. L. BOTT, 1985: Acclimation of stream-bed heterotrophic microflora: metabolicresponses to dissolved organic matter. – Freshw. Biol. 15: 479–492.
KAPLAN, L. A. and J. D. NEWBOLD, 1993: Biochemistry of dissolved organic carbon entering streams.pp. 139–165. – In: FORD, T. E. (ed.). Aquatic microbiology: An ecological approach. Blackwell, Cambridge, Mass, 518 pp.
KHAN, E., S. KING, R. W. Jr. BABCOCK and M. K. STENSTROM, 1999: Factors influencing biodegradabledissolved organic carbon measurement. – J. Env. Engineer. 125: 514–521.
LOCK, M. A., 1990: The dynamics of dissolved and particulate organic material over the substratum ofwater bodies. pp. 117–144. – In: WOTTON, R. S. (ed.). The biology of particles in aquatic systems.CRC Press Inc.
MALTBY, L., 1992: Heterotrophic microbes. pp. 162–194. – In: CALOW, P. and G. E. PETTS (eds.). Therivers handbook. Vol. 1. Blackwell Scientific Publication, Oxford, 526 pp.
40 S. AXMANOVÁ and M. RULÍK
© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim

MARTIN-MOUSSET, B., J. P. CROUE, E. LEFEBVRE and B. LEGUBE, 1997: Distribution et caractérisation de la matiére organique dissoute d’eaux naturelles de surface [Distribution and characterization ofdissolved organic matter of surface waters]. – Wat. Res. 31: 541–553. (in French).
MCARTHUR, M. D. and J. S. RICHARDSON, 2002: Microbial utilization of dissolved organic carbon leached from riparian litterfall. – Can. J. Fish. Aquat. Sci. 59: 1668–1676.
MEYER, J. L., G. E. LIKENS and J. SLOANE, 1981: Phosphorus, nitrogen, and organic carbon flux in aheadwater stream. – Arch. Hydrobiol. 91: 28–44.
MEYER, J. L., R. T. EDWARDS and R. RISLEY, 1987: Bacterial growth on dissolved organic carbon froma blackwater river. – Microb. Ecol. 13: 13–29.
MEYER, J. L., 1990: Production and utilization of dissolved organic carbon in reverine ecosystems. pp. 281–299. – In: PERDUE, E. M. and E. T. GJESSING (eds.). Organic acids in aquatic ecosystems.John Wiley & Sons Ltd, Dahlem Konferenzen, 345 pp.
MEYER, J. L., J. B. WALLACE and S. L. EGGERT, 1998: Leaf litter as a source of dissolved organic carbon in streams. – Ecosystems 1: 240–249.
MORAN, M. A. and R. E. HODSON, 1989: Formation and bacterial utilization of dissolved organic carbon derived from detrital lignocellulose. – Limnol. Oceanogr. 34: 1034–1047.
MULHOLLAND, P. J., 1981: Organic carbon flow in a swamp-stream ecosystem. – Ecological Mono-graphs 51: 307–322.
PARSONS, W. F. J., B. R. TAYLOR and W. F. J. PARSONS, 1990: Decomposition of aspen (Populus tremuloides) leaf litter modified by leaching. – Can. J. For. Res. 20: 934–951.
PETERSEN, R. C. and K. W. CUMMINS, 1974: Leaf processing in a woodland stream. – Freshw. Biol. 4:343–368.
PUSCH, M., D. FIEBIG, I. BRETTAR, H. EISENMANN, B. K. ELLIS, L. A. KAPLAN, M. A. LOCK, M. W. NAE-GELI and W. TRAUNSPURGER, 1998: The role of micro-organisms in the ecological connectivity of running waters. – Freshw. Biol. 40: 453–495.
ROBINSON, CH. T. and M. O. GESSNER, 2000: Nutrient addition accelerates leaf breakdown in an alpinespringbrook. – Oecologia 122: 258–263.
ROSTAN, J. C. and B. CELLOT, 1995: On the use of UV spectrophotometry to assess dissolved organiccarbon origin variations in the Upper Rhone River. – Aquat. Sci. 57: 70–80.
RULÍK, M., P. ZAVRELOVÁ and M. DUCHOSLAV, 2001: Decomposition of two different POM types insurface water and within hyporheic sediments of a small lowland stream (Sitka, Czech Republic). –Internat. Rev. Hydrobiol. 86: 487–500.
SERVAIS, P., A. ANZIL and C. VENTRESQUE, 1989: Simple method for determination of biodegradabledissolved organic carbon in water. – Appl. Environ. Microbiol. 55: 2732–2734.
TRULLEYOVÁ, S., M. RULÍK and J. POPELKA, 2003: Stream and interstitial water DOC of a gravel bar(Sitka stream, Czech Republic): characteristics, dynamics and presumable origin. – Arch. Hydrobiol.158: 407–420.
VOLK, Ch. J., C. B. VOLK and L. A. KAPLAN, 1997: Chemical composition of biodegradable dissolvedorganic matter in streamwater. – Limnol. Oceanogr. 42: 39–44.
WALLACE, J. B., M. R. WHILES, S. EGGERT, T. F. CUFFNEY, G. J. LUGTHART and K. CHUNG, 1995: Long-term dynamics of coarse particulate organic matter in three Appalachian Mountain streams. –J. N. Am. Benthol. Soc. 14: 217–232.
WEBSTER, J. R. and E. F. BENFIELD, 1986: Vascular plant breakdown in freshwater ecosystems. – Ann.Rev. Ecol. Syst. 17: 567–594.
WEBSTER, J. R. and J. L. MEYER, 1997: Stream organic matter budgets. – J. N. Am. Benthol. Soc. 16:3–161.
Manuscript received October 6th, 2003; revised July 26th, 2004; accepted July 28th, 2004
Alder Leaves and Catkins Decomposition 41
© 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim





























