Embed Size (px)
Citation preview

[CANCER RESEARCH 47, 1098-1104, February 15, 1987)
Disialoganglioside GD2 on Human Neuroblastoma Cells: Target Antigen forMonoclonal Antibody-mediated Cytolysis and Suppression of Tumor Growth1
Kalpana Mujoo, David A. Cheresh,2 Hsin Ming Yang, and Ralph A. Reisfeld3
Department of Immunology, Scripps Clinic and Research Foundation, La Julia, California 92037
ABSTRACT
A murine monoclonal antibody 14.18 specifically recognizes disiaUHganglioside (M):,the majorganglioside expressed on the surface of humanneuroblastoma cells. This monoclonal antibody (Mab) is of immunoglob-ulin G3 isotype, has an affinity constant (A,,) of 3.5 x III"M~',and reacts
preferentially with tumorcells and fresh frozen tumor tissues of neuroec-todermal origin in enzyme-linked immunosorbentassay and immunoper-oxidase assays, respectively. Mab 14.18 effectively lyses a number ofhuman neuroblastoma cell lines by two distinct mechanisms, i.e., antibody-dependentcellular cytotoxicity and complement-dependentcytotox-icity. There is a good correlationbetweenthe average numberof antibody-binding sites per neuroblastomacell and the amount of cell lysis observedin complement-dependent cytotoxicity and antibody-dependent cellularcytotoxicity. In addition, Mab 14.18 suppresses establishment as well asgrowth of progressively growing, established human neuroblastoma tumors in nude mice when injected 24 h and 9 days, respectively, after theinitial s.c. inoculation of tumorcells. These data suggest that Mab 14.18can mediate tumor cell killing in vivo and in vitro and may thereby proveuseful for immunotherapyof human neuroblastoma.
INTRODUCTION
Neuroblastoma is a neoplasm of the peripheral autonomienervous system that represents the second most common malignancy of childhood. Fifty-five % of patients have widespreadme tastatic disease at the time of the diagnosis (1). More thanone-half of the children present with the disseminated diseaseand are incurable by currently known methods. There has beenrelatively little improvement in outcome during the last 25years, in contrast to the considerable advances made in mostother childhood cancers. It is apparent that new therapeuticmodalities are needed for the patients with disseminated neuroblastoma (Stage III and IV).
Initial studies by several investigators that evaluated a seriesof Mabs4 for new immunotherapy modalities to be applied to
neuroblastoma were largely unsuccessful since these reagentsfailed to react with all neuroblastoma cells and were unable tokill tumor cells in the presence of human complement evenwhen used as a panel (2). In addition, a number of these Mabslacked specificity since they reacted with megakaryocyte precursors and with some normal blood and bone marrow cells(3). When injected into tumor-bearing animals, these Mabslocalized largely in the organs of the reticuloendothelial system,i.e., liver, spleen, and bone marrow (4). Most importantly, thetarget antigens of most of these antibodies were not extensively
Received 8/25/86; revised 11/13/86; accepted 11/17/86.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 I '.S.C'. Section 1734 solely to indicate this fact.1Supported by NIH Grant CA 42508 and a Fellowship Support Grant from
the Joseph Drown Foundation, Los Angeles, CA. This is Scripps Publication No.4444-Imm.
1Recipient of a J. Ernest Ayre Fellowship from the National Cancer Cytology
Center.3To whom requests for reprints should be addressed.4The abbreviations used are: Mab, monoclonal antibody; G,,2, disialoganglio-
side; PBS, phosphate-buffered saline; ADCC, antibody-dependent cellular cytotoxicity; CDC, complement-dependent cytotoxicity; FACS, fluorescence-activated cell sorter, USA, bovine serum albumin; HBSS, Hanks' buffered saltsolution; ELISA, enzyme-linked immunosorbent assay, TLC, thin-layer chro-matography.
characterized and thus their biological function is unknown (5).However, more progress was made in other studies investi
gating disialoganglioside GD2with both human (6) and murine(7, 8) Mabs directed to this chemically defined target antigenuniformly expressed on human neuroblastoma cells. In thisregard, anti-GD2 murine Mabs of IgG3 isotype were used todemonstrate that Go2 redistributes into microprocesses makingdirect contact with a fibronectin substrate and being substantially involved in attachment of melanoma and neuroblastomacells to this substrate (9, 10). Most recently, another anti-GD2Mab of IgG3 isotype was reported to produce a prolongedremission in a neuroblastoma patient (11).
We report here on the reactivity of anti-GDz Mab 14.18(IgG3) with a variety of normal and neoplastic cells and tissuesas well as on the functional role of this antibody with emphasison its potential for immunotherapy of neuroblastoma.
MATERIALS AND METHODS
The following human cell lines were obtained from the AmericanType Culture Collection, Rockville, MD: MOLT-4; HPB-ALL (lym-phoblastoid); and U-138MG (glioblastoma). The neuroblastoma celllines LAN-1 and LAN-5 were kindly provided by Dr. R. C. Seeger,UCLA. Additional neuroblastoma cell lines SK-N-AS, SK-N-RA, andSK-N-LE were a gift from Dr. Helson, Memorial Sloan-KetteringCancer Center, New York. SMS-KCNR, SMS-KAN, and CHP-134were kindly provided by Dr. P. Reynolds, Naval Medical Center,Bethesda, MD, and the IMR-6 and NMB-7 neuroblastoma cell lineswere a gift from Dr. S. K. Liao, McMaster University, Hamilton,Ontario, Canada. L 14 (lymphoblastoid), M21, and M-14 (melanoma)lines were kindly provided by Dr. D. L. Morton, UCLA. FM8 andFM9-Met melanoma cell lines are available from our laboratory andthe Melur melanoma cell line was kindly provided by Dr. U. Koldovsky,University of Dusseldorf, West Germany. The melanoma A375P lineand its A375 Met mix spontaneous metastatic variant were a gift fromDr. I. J. Fidler, M. D. Anderson Hospital and Tumor Institute, Houston, TX. The LG-2 lymphoblastoid cell line is available in our laboratory. The T293 (small cell lung carcinoma) line was provided by Drs.H. Masuie and G. Sato, University of California, San Diego. The NCIH-69, H-82, and N-417 small cell lung carcinoma were made availableby Drs. J. Minna and A. Gazdar, NIH, Bethesda, MD. The pancreaticcarcinoma cell lines FG and SG were provided by Dr. S. Kajiji at ourinstitution.
Monoclonal Antibodies. Monoclonal antibody W6/32 (IgG2a anti-Hi A) is maintained in our laboratory as are Mabs 126-4 (IgM; anti-GD2);C281 (IgG3; anti-GD3); 5G3 (IgG2a smti-M, 215,000 glycoproteinantigen).
Animals. BALB/c athymic (nu/nu) mice were obtained from the nudemouse colony at the University of California, San Diego, La Jolla, CA.
Production of Monoclonal Antibody, mah 14.18 was produced againstthe human neuroblastoma cell line LAN-1 by standard hybridomatechnology (12). Briefly, 5 x IO6 LAN-1 cells were injected i.p. into
BALB/c mice, once every week for 4 weeks. On the third day after thelast injection, the animals' splenocytes were removed and fused withthe M-5 variant of the nonsecretor murine myeloma cell line SP2/0.Hybridoma 14.18 was selected by growth in Dulbecco's modified Eagle's medium containing 10% horse serum, hypoxanthine, aminopterin,
and thymidine and subcloned by limiting dilutions.Ascites Preparation. BALB/c mice were primed by injecting 0.5 ml
pr¡stane.Two weeks after priming, 10-15 x IO6 hybridoma cells were
1098
on July 13, 2018. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

DISIALOGANGLIOSIDE GD2 ON HUMAN NEUROBLASTOMA CELLS
injected i.p. into the mice and ascites fluid was collected repeatedlyevery 2-3 days. The fluid was centrifuged at 2000 x g for 20 min toremove cells and other debris and quickly frozen at -20°C.
Purification of Mab. The ascites fluid was clarified at 10,000 x g for20 min. The supernatant was then titrated on ELISA and only asciteswith high titer was selected for the purification of Mab. Purificationwas achieved by Protein A-Sepharose affinity chromatography as described previously (13). Antibody was eluted with 0.1 M sodium citratebuffer, pH 4.5, and dialyzed extensively at 4°Cagainst PBS, pH 7.4.The purified antibody was stored in small aliquots at -70°C.
Isotyping. The isotype of the Mab was determined by ELISA, usingaffinity-purified rabbit antisera specific for different murine light chains(Southern Biotechnology Associates, Birmingham, AL) that were driedinto 96-well microtiter plates (Dynatech, Alexandria, VA). The murinemonoclonal antibody 14.18 is of IgG3 isotype as determined by analysiswith these murine isotype standards.
ELISA. Screening of Mab by ELISA was done essentially as described previously (14). Briefly, 5x10* target cells/well were plated in
polyvinyl microtiter plates (Dynatech). Prior to ELISA, the dried plateswere rehydrated by washing them twice with 10 HIM PBS, pH 7.4,containing 0.1% Tween 20 and 0.02% thimerosal (washing buffer).Supernatants were diluted 1:2 in washing buffer containing 0.1% BSAas diluent. Diluted test supernatant (SO/¿I)was added to each well andplates were incubated for l h at 4°C.Following three washes, SO/J of
horseradish perioxidase-conjugated goat anti-mouse IgG (Bio-Rad,Richmond, CA) were added to each well and incubated for l h at 4°C.
After two final washes, 50 /il of substrate solution (400 Mg/ml o-phenylenediamine in 0.12% H2O2) were added to each well. The reaction was stopped after 15 min by addition of 25 /il of 4 N H2SO4 toeach well. Absorbance at 492 nm was measured with a microtiter platereader.
Tissues. Fresh frozen normal and malignant tissue were obtainedfrom the surgical pathology department of the Ida M. Green Hospitalof Scripps Clinic, La Jolla, CA. Additional tumor specimens werekindly provided by Drs. F. Kung and A. Yu, Department of Pediatrics,University of California at San Diego. Specimens were embedded intissue Tek-II O.C.T. (Miles, Naperville, IL) frozen in blocks in isopen-tene at -176°C and stored at -70"C.
Immunoperoxidase Staining of Frozen Tissues. Two- to 4-¿im-thick
sections were cut from frozen tissue blocks on a cryostat/microtome(Damon Biotech, Needham Heights, MA). These sections weremounted on gelatin-coated glass slides, air dried briefly, and testedimmediately in an indirect immunoperoxidase assay essentially as described earlier (15). Briefly, after washing them twice in HBSS andonce in PBS, we preincubated the sections for 15 min with PBScontaining 10% goat serum and 0.1% BSA. Excess buffer was thenpipeted off and sections were overlaid with appropriately diluted hy-bridoma supernatant»and incubated for l h at room temperature. Aftertwo washes in HBSS and one wash in PBS, the tissue sections wereoverlaid with horseradish peroxidase-conjugated goat anti-mouse im-munoglobulin (Bio-rad), diluted 1:50 with dilution buffer (PBS containing 10% goat serum and 0.1% BSA), and incubated for l h at roomtemperature. Finally, the tissue sections were washed twice in HBSSand once in PBS and incubated for 15 min at room temperature withimmunoperoxidase substrate buffer (10 mM Tris, pH 7.6, containing0.6 mg/ml 3,3'-diaminobenzidine and 0.015% H2O2). The sections
were counterstained briefly in l % méthylèneblue, dehydrated throughgraded ethanol, washed in Histoclear (National Diagnostics Somerville,NJ), mounted with Pro-Tex (Lerner Laboratories, New Haven, CT),and examined by microscopy.
Ganglioside Extraction. Packed neuroblastoma cells (1 ml) werewashed extensively with PBS and homogenized in chloroform:methanol(2:1). The residue was reextracted with chloroform:methanol (1:1) andpassed through a sintered glass funnel, and the remaining residue wasagain extracted with this same solvent. The extracts were combined,dried under nitrogen, and partitioned in diisopropyl ethenn-butyl alcohol (6:4) and 50 HIMNaCl as described by Ladish and Ciliare! (16).
TLC. Plastic-backed silica gel TLC plates (E. M. Merck, Darmstadt,Germany) were activated by heating at 110°Cfor 1 h. Glycolipids were
dissolved in chloroform:methanol (1:1) and spotted 1.5 cm from the
bottom of the TLC plates. These plates were then placed in a developingtank presaturated with 100 ml of chloroform:methanol:H2O (55:45:10)containing 0.02% CaCl2. Chromatograms were developed for 1.5 h atroom temperature, after which the TLC plates were allowed to dry.The chromatograms were then sprayed with resorcinol reagent tovisualize the gangliosides.
Immunostaining of Gangliosides Separated by TLC. The reactivity ofMab 14.18 with gangliosides separated by TLC was determined byusing the immunostaining method described by Cheresh et al. (17).
Lipid ELISA. The lipid ELISA was performed essentially as described earlier (17), except that the plates were incubated for l h withPBS containing 5% BSA before the addition of monoclonal antibody.The remaining assay was essentially the same as that described for theregular ELISA, except washing buffer consisted of PBS containing 1%BSA. For preparation of lipid plates, a total lipid extract was preparedfrom 2 ml of packed M21 melanoma cells which are known to extensively express Gm ganglioside. These cells were homogenized in 40 mlchloroform:methanol (2:1), followed by filtration through a sinteredglass filter. The residue was reextracted with chloroform:methanol (1:1)and refiltered. The combined filtrates were then dried down. Finally,the glycolipids were resuspended in methanol. Routinely, glycolipidsfrom 50 ¡i\packed cell volume were plated per well in flat-bottomed,polyvinyl microtiter plates (Dynatech).
I25l-Labeling of Mab 14.18. The antibody was labeled with 125Iby thechloramine-T method, as described previously by us (18). Briefly, 100n\ of antibody (1 mg/ml) were mixed with 400 //I of 0.2 M sodiumphosphate buffer, pH 7.5. 125I(carrier-free, 17 Ci/mg) (0.5 mCi) andchloramine-T (10 Mg;Sigma) were added to this solution. The samplewas incubated for 5 min and the reaction was stopped by the additionof 10 jugof sodium metabisulfite (1 mg/ml in H2O) and 200 n\ of PBScontaining 3% Trasylol (Sigma). After completion of the reaction, aSephadex G-25 column (Pharmacia), was used to separate the bound,radiolabe led protein from free I25I.This column was prewashed with
0.5% BSA/PBS and fractions were collected with the same buffer. Thepercentage of incorporation of radioiodine was determined by trichlo-roacetic acid precipitation.
Scali-hard Plot Analysis. The specific activity of the radiolabeledantibody was 2 XÌ104cpm/ng. The association constant (KA)of Mab
14.18 was determined from Scatch ard plot analysis of saturation binding data (19). Briefly, NMB-7 neuroblastoma cells (106/0.1 ml of RPMI
1640 containing 2% fetal calf serum and 0.01% NaN.,) were incubatedfor l h at 4°Cwith increasing amounts of radiolabeled antibody (5-
600 ng), diluted in the same media. Fractions containing bound andfree antibody were separated by a dibutyl phthalate and dinonyl phthal-ate (1:1) oil layer as described by Beaumier et al. (20). Radioactivity ineach fraction was determined in an LKB 1270 gamma counter. Theamount of nonspecific binding of Mab 14.18 was estimated by addinga 100-fold excess of cold antibody to a cell suspension and subtractingbound cpm from total cpm. The data were subjected to linear regressionanalysis.
FACS Analysis. Viable cells were stained at 4'C and used for quan-
titation of cell surface expression of antigens by FACS as describedearlier (21). Briefly, the cells were washed twice with HBSS containing1% BSA before the addition of primary antibody. SP2/0 culture supernatant served as negative control for the primary antibody bindingassay. After l h of incubation at 4°C,the cells were washed twice with
HBSS/1% BSA and allowed to react with fluuresecin isothiocyanate-conjugated goat anti-mouse sera for l h after which the percentage ofreactive cells and the mean intensity of fluorescence was determined byFACS analysis.
CDC. Neuroblastoma cells suspended in 1 ml of growth media werelabeled with 100 ¿tCiof (Na)251CrO4(New England Nuclear, 1 Ci = 37GBq) for 2 h at 37°C.After 2 h of incubation, the cells were washed
twice with tissue culture medium RPMI 1640 and 25-ni aliquotscontaining 10* cells were plated in 96-well microtiter plates (Costar,
Cambridge, MA) with various concentrations of Mab 14.18. Each platewas incubated for l h at 31°C after which 150 n\ of fresh human serum
diluted 1:3 were added as the source of complement. Each plate wasallowed to incubate for an additional 90 min at 37°Cand was then
centrifuged at 400 x g for 5 min and the radioactivity in the supernatant
1099
on July 13, 2018. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

DISIALOGANGLIOSIDE GDJ ON HUMAN NEUROBLASTOMA CELLS
was measured in a gamma counter. The total count were assessed byadding 1% Nonidet P-40. Percentage of lysis was calculated as
(Experimental s'Cr release) - (Spontaneous s'Cr release)(Maximal "Cr release) - (Spontaneous "Cr release) x 100
ADCC. Neuroblastoma cells (SK-N-AS, IMR-6, SMS-KCNR) werelabeled in 1 ml of growth media with 300 nd of (Na)251CrO4for 2 h at37*C. After the cells were washed twice with RPMI 1640, 5 x IO3cells
in 25 ill were plated in 96-well microtiter plates. Mab 14.18 (5 fig inSO fi\) was added to each well containing target cells. Mononuclearhuman effector cells were isolated by subjecting heparinized blood ofhealthy individuals to centrifugation on a Ficoll-Hypaque gradient.These cells were added to microtiter wells at the indicated target reflectorcell ratios and the plates were incubated for 4 h at 37°C.The plates
were then centrifuged at 1000 x g for 5 min and radioactivity wasmeasured in a 100 ^1 aliquot of each supernatant. Spontaneous s'Cr
release was determined in wells that contained only labeled target cells.Total cpm release was determined by lysing tumor cells with 1%Nonidet P-40. The percentage of lysis was calculated by the sameformula as that used to compute complement-dependent cytotoxicity.The percentage of specific lysis was calculated by subtracting thepercentage of lysis obtained with effector cells in the absence of antibody, i.e., natural killer cell lysis, from each value obtained.
RESULTS
ELISA Reactivity of Mab 14.18 with Human Tumor CellLines. Table 1 summarizes the ELISA reaction patterns of Mab14.18 with various human tumor cell lines. This antibody is ofIgG3 isotype and has an association constant of 3.5 x IO8M~l,
as determined by Scalchimi plot analysis. The antibody reactedvery strongly with all eight neuroblastoma cell lines tested. In
Table 1 ELISA reactivity of Mab 14.18 with cultured cell linesAbsorbance"
NeuroblastomaLAN-1 ++++LAN-5 Duke ++++SMS-KCNR ++++CHP-134 ++++SMS-KAN ++++NMB-7 ++++IMR-6 ++++SK-N-AS +++
MelanomaMelur +A375 P +++A375 Met +++FM8 +++FM9 Met +++M-14 ++++M-21 ++++
GliomaU138M6 ++++
Small cell lung carcinomaT293 ++++NIH-N417 ++++NIH-H-82 ++NIH-H-69 ++
Other tumor cell linesPancreatic carcinoma (fast growing) -Pancreatic carcinoma (slow growing) -Pancreatic carcinoma 1320 (metastatici -Adenocarcinoma of lung (UCLA-P3) -
Lymphoblastoid cell linesL-U(B-cell)LG-2 (B-ceil)MOLT-4 (T-cell)HPB-ALL (T-cell) -
•Absorbance at 492 nm: 0.15-0.3 (+); 0.3-0.6 (++); 0.6-0.9 (+++); 0.9->2.0(++++); 0-0.050 (-).
addition, Mab 14.18 also reacted to varying degrees of intensitywith a number of melanoma and small cell lung carcinoma celllines. This is not too surprising since neuroblastoma, melanoma, and possiby tumor cells of small cell lung carcinoma areof neuroectodermal origin. Mab 14.18 also reacts with oneglioma cell line. However, the antibody failed to react with celllines derived from pancreatic carcinoma and adenocarcinomaof lung and with lymphoblastoid cell lines of either T- or B-cellorigin.
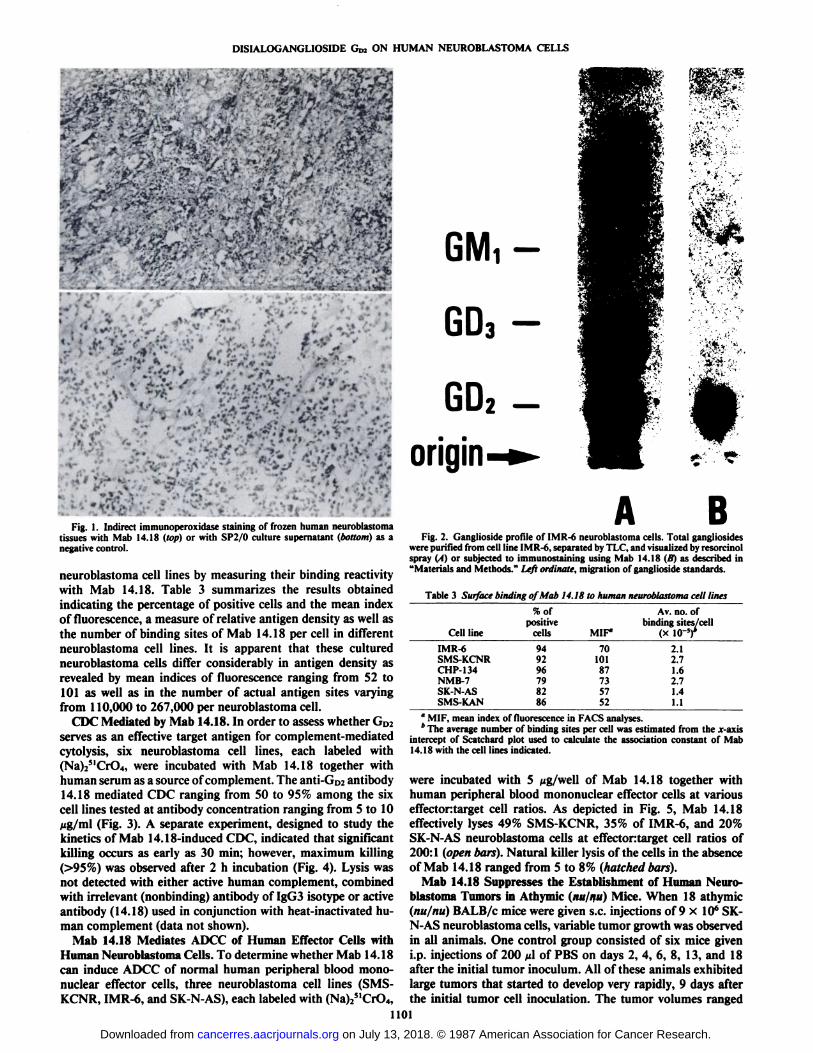
Tissue Specificity of Mab 14.18. When Mab 14.18 was testedagainst a number of frozen normal and tumor tissues, theantibody reacted very strongly with neuroblastoma, melanoma,glioblastoma, and small cell lung carcinoma tissues (Table 2).Since reactivity was particularly pronounced with these tissuesections obtained from tumors of neuroectodermal origin, it isnot surprising that this anti-Go? antibody also reacted withnormal adult cerebellum and fetal brain. A variety of othertumor tissues showed negative staining. All normal tissuestested, with the exception of cerebellum and fetal brain, failedto react with Mab 14.18. Fig. IA illustrates the strong positivestaining obtained when Mab 14.18 was reacted with fresh frozenneuroblastoma tissue and contrasts with the lack of stainingobserved when the same tissue was reacted with a negativecontrol, i.e., the myeloma fusion partner SP2/0 culture supernatant (Fig. IB).
Immunochemical Identification of the Target Antigen of Mab14.18. The pattern of gangliosides in an extract obtained fromneuroblastoma cell line, IMR-6 is revealed by TLC. Go2 is themajor ganglioside present in these cells as demonstrated by theresorcinol sprayed total ganglioside profile (Fig. 2A). By im-munostaining, Mab 14.18 was clearly shown to bind to a singlecomponent that comigrated with authentic GD2 on the TLCplate (Fig. 21}}.In addition, results from lipid ELISA indicatethat Mab 14.18 binds only to GD2and not to other gangliosidestandards tested, i.e., GD3, GM,, and GM3(26).
Binding of Mab 14.18 to Viable Human Tumor Cells. The GD2antigen was detected by FACS analysis on viable cells of six
Table 2 Tissue reactivity of Mab 14.18Reactivity"
Tumor tissuesNeuroblastomaMelanomaGlioblastomaSmall cell lung carcinomaEwing's sarcoma
Lung adenocarcinomaStomach carcinomaSquamous lung carcinomaBreast carcinomaFibroblastoidColon cancerOral squamous cancer
Normal tissuesLungColonLiverKidneySpleenPancreasThyroidCerebellum
Fetal tissuesLungSpleenLiverColonKidneyBrain
" +++, strongly positive; ++, positive; +, weak/positive; -, negative.
1100
on July 13, 2018. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
DISIALOGANGLIOSIDE GD2 ON HUMAN NEUROBLASTOMA CELLS
»ft.'• '- • V ' . . i \ ' , .~ . . , . •t.' A"»'. •V '"'•* >•*•*-4V^ V' . {^.:
- +-*uwj* «...»._ * » r.¡Fig. 1. Indirect immunoperoxidase staining of frozen human neuroblastoma
tissues with Mab 14.18 (top) or with SP2/0 culture supernatant (bottom) as anegative control.
neuroblastoma cell lines by measuring their binding reactivitywith Mab 14.18. Table 3 summarizes the results obtainedindicating the percentage of positive cells and the mean indexof fluorescence, a measure of relative antigen density as well asthe number of binding sites of Mab 14.18 per cell in differentneuroblastoma cell lines. It is apparent that these culturedneuroblastoma cells differ considerably in antigen density asrevealed by mean indices of fluorescence ranging from 52 to101 as well as in the number of actual antigen sites varyingfrom 110,000 to 267,000 per neuroblastoma cell.
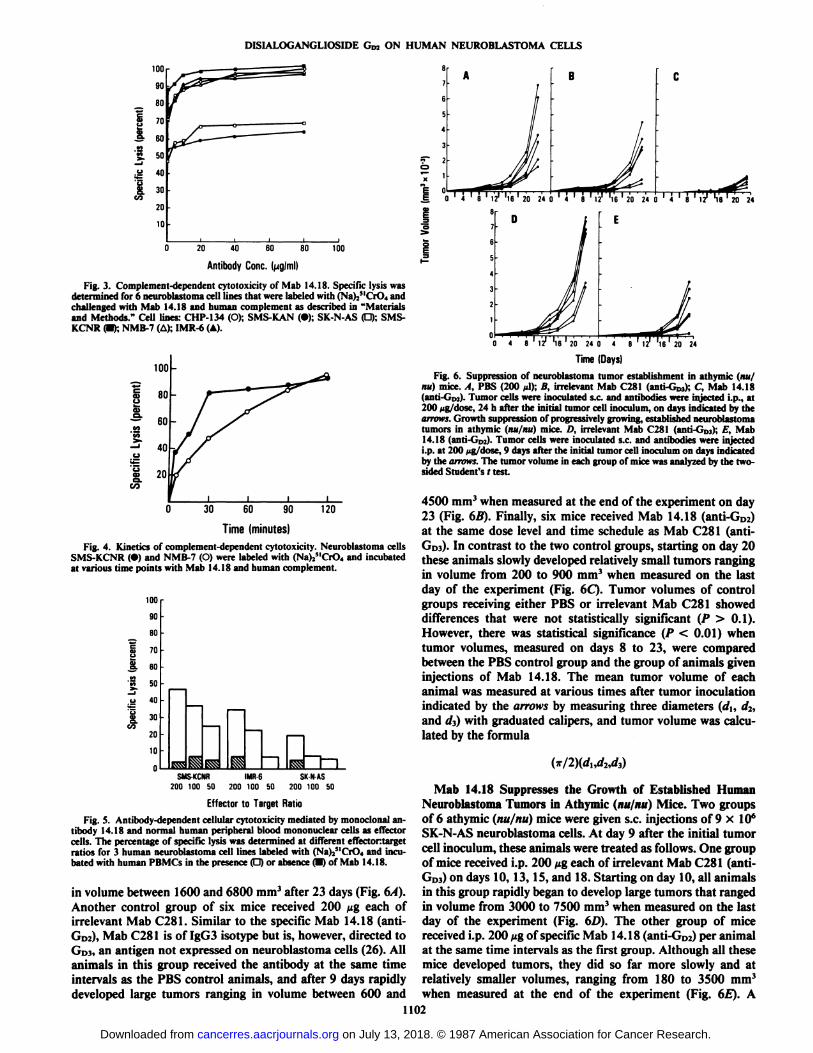
CDC Mediated by Mab 14.18. In order to assess whether GDzserves as an effective target antigen for complement-mediatedcytolysis, six neuroblastoma cell lines, each labeled with(Na)25lCrO4, were incubated with Mab 14.18 together withhuman serum as a source of complement. The anti-GD2 antibody14.18 mediated CDC ranging from 50 to 95% among the sixcell lines tested at antibody concentration ranging from 5 to 10fig/ml (Fig. 3). A separate experiment, designed to study thekinetics of Mab 14.18-induced CDC, indicated that significantkilling occurs as early as 30 min; however, maximum killing(>95%) was observed after 2 h incubation (Fig. 4). Lysis wasnot detected with either active human complement, combinedwith irrelevant (nonbinding) antibody of IgG3 isotype or activeantibody (14.18) used in conjunction with heat-inactivated human complement (data not shown).
Mab 14.18 Mediates ADCC of Human Effector Cells withHuman Neuroblastoma Cells. To determine whether Mab 14.18can induce ADCC of normal human peripheral blood mono-nuclear effector cells, three neuroblastoma cell lines (SMS-KCNR, IMR-6, and SK-N-AS), each labeled with (Na)25lCrO4,
GMi-
GD3-
GD2_
origin
BFig. 2. Ganglioside profile of IMR-6 neuroblastoma cells. Total gangliosides
were purified from cell line IMR-6, separated by TLC, and visualized by resorcinolspray (A) or subjected to immunostaining using Mab 14.18 (B) as described in"Materials and Methods." Left ordinate, migration of ganglioside standards.
Table 3 Surface binding of Mab 14.18 to human neuroblastoma cell lines
CelllineIMR-6SMS-KCNR
CHP-134NMB-7SK-N-ASSMS-KAN%of
positivecells94
9296798286MIF*70
10187735752Av.
no. ofbinding sites/cell(x 10-y2.1
2.71.62.71.41.1
" MIF, mean index of fluorescence in I-ACS analyses.* The average number of binding sites per cell was estimated from the jr-axis
intercept of Scatchard plot used to calculate the association constant of Mab14.18 with the cell lines indicated.
were incubated with 5 /¿g/wellof Mab 14.18 together withhuman peripheral blood mononuclear effector cells at variouseffectontarget cell ratios. As depicted in Fig. 5, Mab 14.18effectively lyses 49% SMS-KCNR, 35% of IMR-6, and 20%SK-N-AS neuroblastoma cells at effectontarget cell ratios of200:1 (open bars). Natural killer lysis of the cells in the absenceof Mab 14.18 ranged from 5 to 8% (hatched bars).
Mab 14.18 Suppresses the Establishment of Human Neuroblastoma Tumors in Athymic (nujnu) Mice. When 18 at hymìe(nu/nu) BALB/c mice were given s.c. injections of 9 x 10e SK-N-AS neuroblastoma cells, variable tumor growth was observedin all animals. One control group consisted of six mice giveni.p. injections of 200 n\ of PBS on days 2, 4, 6, 8, 13, and 18after the initial tumor inoculum. All of these animals exhibitedlarge tumors that started to develop very rapidly, 9 days afterthe initial tumor cell inoculation. The tumor volumes ranged
1101
on July 13, 2018. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
DISIALOGANGLIOSIDE GDJ ON HUMAN NEUROBLASTOMA CELLS
100
90
80
Î-& 60'§. 50
£ 40
S. 30
20
10
20 40 60 BO
Antibody Conc. (jjgiml)
100
Fig. 3. Complement-dependent cytotoxicity of Mab 14.18. Specific lysis wasdetermined for 6 neuroblastoma cell lines that were labeled with (Na)2MCrO4 andchallenged with Mab 14.18 and human complement as described in "Materialsand Methods." Cell lines: CHP-134 (O); SMS-KAN (•);SK-N-AS (O); SMS-KCNR (•);NMB-7 (A); IMR-6 (A).
'55
uI30 60 90 120
Time(minutes)Fig. 4. Kinetics of complement-dependent cytotoxicity. Neuroblastoma cells
SMS-KCNR (•)and NMB-7 (O) were labeled with (Na)Y"( 'r( >4and incubated
at various time points with Mab 14.18 and human complement.
100
90
80I
7°
t60f
50o
40
130GO2010
0SSHESS
^ hIs3sr~l~~iSMS-KCNR
IMR6 SK-N-AS200 100 50 200 100 50 200 100 50
Effector to Target Ratio
Fig. 5. Antibody-dependent cellular cytotoxicity mediated by monoclonal antibody 14.18 and normal human peripheral blood mominuclear cells as effectorcells. The percentage of specific lysis was determined at different effectortargetratios for 3 human neuroblastoma cell lines labeled with (Na)/'("r( ), and incu
bated with human PBMCs in the presence O or absence (D) of Mab 14.18.
in volume between 1600 and 6800 mm3after 23 days (Fig. 6/4).
Another control group of six mice received 200 /¿geach ofirrelevant Mab C281. Similar to the specific Mab 14.18 (anti-Go:), Mab C281 is of IgG3 isotype but is, however, directed toGDI,an antigen not expressed on neuroblastoma cells (26). Allanimals in this group received the antibody at the same timeintervals as the PBS control animals, and after 9 days rapidlydeveloped large tumors ranging in volume between 600 and
.§ 0 ' 4 ' 8 ' 12*''l6 ' 20 24 0 ' 4 ' è ' 121 Ve ' 20 ' 24 Ó' 4 ' I ' 121 "l6 ' 20 24
D Ã \ E
5
Time (Days)
Fig. 6. Suppression of neuroblastoma tumor establishment in athymic (nu/nu) mice. A, PBS (200 »il);B, irrelevant Mab C281 (anti-GD]); C, Mab 14.18(anti-Cìnj).Tumor cells were inoculated s.c. and antibodies were injected i.p., at200 fig/dose, 24 h after the initial tumor cell inoculum, on days indicated by thearrows. Growth suppression of progressively growing, established neuroblastomatumors in athymic (nu/nu) mice. D, irrelevant Mab C281 (anti-Ci,,,); K. Mab14.18 (ant i G 02)- Tumor cells were inoculated s.c. and antibodies were injectedi.p. at 200 fig/dose, 9 days after the initial tumor cell inoculum on days indicatedby the arrows. The tumor volume in each group of mice was analyzed by the two-sided Student's r test
4500 mm3when measured at the end of the experiment on day23 (Fig. 6B). Finally, six mice received Mab 14.18 (anti-Goa)at the same dose level and time schedule as Mab C281 (ami-GDI)-In contrast to the two control groups, starting on day 20these animals slowly developed relatively small tumors rangingin volume from 200 to 900 mm3 when measured on the last
day of the experiment (Fig. 6C). Tumor volumes of controlgroups receiving either PBS or irrelevant Mab C281 showeddifferences that were not statistically significant (P > 0.1).However, there was statistical significance (P < 0.01) whentumor volumes, measured on days 8 to 23, were comparedbetween the PBS control group and the group of animals giveninjections of Mab 14.18. The mean tumor volume of eachanimal was measured at various times after tumor inoculationindicated by the arrows by measuring three diameters (d¡,di,and Ã/0with graduated calipers, and tumor volume was calculated by the formula
Mab 14.18 Suppresses the Growth of Established HumanNeuroblastoma Tumors in Athymic (nu/nu) Mice. Two groupsof 6 athymic (nu/nu) mice were given s.c. injections of 9 x IO6SK-N-AS neuroblastoma cells. At day 9 after the initial tumorcell inoculum, these animals were treated as follows. One groupof mice received i.p. 200 /tg each of irrelevant Mab C281 (anti-GDJ)on days 10, 13, 15, and 18. Starting on day 10, all animalsin this group rapidly began to develop large tumors that rangedin volume from 3000 to 7500 mm3 when measured on the lastday of the experiment (Fig. 6£>).The other group of micereceived i.p. 200 jugof specificMab 14.18 (anti-Go:) per animalat the same time intervals as the first group. Although all thesemice developed tumors, they did so far more slowly and atrelatively smaller volumes, ranging from 180 to 3500 mm3
when measured at the end of the experiment (Fig. 6£).A1102
on July 13, 2018. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

DISIALOGANGLIOSIDE GD2 ON HUMAN NEUROBLASTOMA CELLS
comparison between tumor volumes of the control group receiving irrelevant Mab C281 with those of the group of animalsgiven Mab 14.18 injections showed statistical significance (P<0.01) when tumor volumes were compared between days 10 and23.
DISCUSSION
There has been an ever increasing interest in evaluating theefficacyof monoclonal antibodies to a variety of human tumor-associated antigens for diagnosis and therapy of cancer. Thesereagents have been extensively used as effective molecularprobes to delineate structural and functional properties of theirtarget antigens and in some cases as agents mediating cancertherapy. During the last few years, Mabs directed to the carbohydrate moieties of glycolipids, particularly those containingone or more sialic acid residues, i.e., gangliosides, have becomeavailable to define these antigens on melanoma, neuroblastoma,small cell lung carcinoma, and colon carcinoma as summarizedin several recent reviews (22-26). Furthermore, Mabs of IgG3isotype directed to disialoganglioside GD3were shown previously by this laboratory (27) and others (28-30) to mediateboth complement-dependent cytotoxicity and antibody-dependent cellular cytotoxicity as well as to suppress tumor growth inathymic mice (27) and in melanoma patients (30). As far asdisialoganglioside Go2 is concerned, a murine Mab of IgMisotype was initially found to be useful in our laboratory for thedetection of GD2in serum and monitoring of tumor progressionin neuroblastoma patients (7). More recently, this same antibody bound to immunobeads was used successfully for bonemarrow clearance of such patients.5 Others reported several
murine Mabs to GD2,i.e., three of IgM and one of IgG3 isotype(8). One of the IgM antibodies was used successfully togetherwith complement to purge tumor cells from autologous bonemarrow of neuroblastoma patients (31) while in a recent preliminary study, an anti-GD2Mab of IgG3 isotype was reported toproduce a complete tumor regression in one of four neuroblastoma patients (11).
Although these results are encouraging because they suggestthat GD2 may be an effective target for immunotherapy ofneuroblastoma, it is apparent that additional trials involvingmore patients and possibly also more sophisticated treatmentmodalities making use of this target antigen will be required tooptimize therapy regimens and to render them more uniformlysuccessful. To achieve this goal, a necessary first step is todetermine the specificity and functional properties of monoclonal antibodies directed to G1):.
Following this general line of reasoning, we determined firstthe specificity and second some of the functional properties ofour anti-GD2 murine Mab 14.18 of IgG3 isotype. In initialexperiments, we observed a lack of expression of GD3on neuroblastoma cells in spite of evidence indicating that Gl)} is aprecursor of GD2(26). This may be due to the fact that GD2issynthesized very rapidly in these cells so that it is difficult todetect reasonable amounts of GDs on their surface. It is alsoapparent from the results reported here that Mab 14.18 reactsstrongly on ELISA with all cell lines of neuroectodermal origin,i.e., neuroblastoma, melanoma, glioma, and possibly small cellcancer of the lung. The results from immunoperoxidase assaysof fresh frozen normal and tumor tissues corroborate theseresults as Mab 14.18 reacted solely with tissues of neuroectodermal origin, including among normal tissues only cerebellum
* R. C. Seeger, personal communication.
and fetal brain. However, it is relatively unlikely that this Mabwill actually react to any extent with brain tissues in vivobecauseof the blood brain barrier. The complete lack of reactivity ofMab 14.18 with other normal adult and fetal tissues is advantageous, particularly when one considers using this antibodyfor immunotherapy. It is likely that this antibody may alsoprove useful for adjuvant type therapy of melanoma and smallcell lung carcinoma. In this regard, another advantage of GD2is that this ganglioside, when shed into the circulation, doesnot form immune complexes with antibody. In fact, GD2presentin serum must be extracted with organic solvents and drieddown before it can react with anti-GD2antibody (7).
As far as the functional properties of Mab 14.18 are concerned, it is evident from the data presented here that thisantibody effectively mediates CDC in the presence of humancomplement (Fig. 3) and that this activity varies to some degreeamong different neuroblastoma cell lines. This is probablycaused by the difference in average number of binding sites percell for Mab 14.18 as indicated in Table 3. In 5 of 6 neuroblas-toma cell lines analyzed, there is good correlation between thenumber of binding sites per cell and the percentage of CDCshown in Fig. 3. Mab 14.18 (IgG3) is also effectivein mediatingADCC with human peripheral blood mononuclear cells aseffector cells against several neuroblastoma cell targets. Againthere is some variability in this reactivity among the neuroblastoma cell lines tested. Similar to CDC, the degree of ADCC-mediated tumor cell lysis also correlates with the average number of binding sites expressed per neuroblastoma cell (Table 3).Taken together, data on the functional properties of Mab 14.18indicate that this Mab can effectively mediate neuroblastomacell lysis in vitro by two distinct mechanisms, i.e., CDC andADCC.
Results from our in vivo studies in athymic (nu/nu) mice,inoculated s.c. with SK-N-AS human neuroblastoma cells, demonstrated that repeated i.p. injections of Mab 14.18 clearlyproduced a measurable suppression of tumor growth. This wasevident when a definite suppression of tumor establishment wasobserved that was statistically significant (/••< 0.01) wheneverMab 14.18 was repeatedly injected i.p. into mice, starting 24 hafter they were inoculated s.c. with human neuroblastoma cells.It is apparent from the data that this suppression of tumorestablishment is specifically achieved with Mab 14.18 (anti-Grt;) since injection of animals with Mab C281 (anti-GixO,anirrelevant antibody of identical IgG3 isotype, is ineffective sinceits GÌ,,target is not expressed to any measurable extent on SK-N-AS human neuroblastoma cells. In terms of a possible clinicalapplicability of Mab 14.18, it is relevant that in comparison tothe irrelevant Mab C281 (anti-G,,,)- Mab 14.18 (ant¡-G,,,)atleast partially suppressed the growth of progressively growinghuman neuroblastoma tumors that were allowed to establishthemselves as palpable 40- to 100-mm' lesions for 9 days before
any antibody was injected into the nude mice. This partialsuppression of tumor growth was statistically significant (/' <0.01). It would seem reasonable to assume that in vivo, bothCDC- and/or ADCC-like mechanisms may be involved in thesuppression of tumor growth, especially in view of data published on other human tumor systems (31-33).
One can only speculate at this time whether monoclonalantibodies are required to evoke CDC and/or ADCC to beuseful in the treatment of neuroblastoma; however, from therelatively few data available from phase I clinical trials withmelanoma (30) and neuroblastoma (11) patients, one wouldassume this to be the case, especially when the injection ofmonoclonal antibody per se is the sole therapy regimen. In this
1103on July 13, 2018. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

DISIALOGANGLIOSIDE GD2 ON HUMAN NEUROBLASTOMA CELLS
regard, additional modalities may prove to be useful for thetreatment of neuroblastoma, such as those recently reportedfrom nude mouse model studies for the treatment of melanoma,where human peripheral blood mononuclear cells, augmentedex vivo with recombinant interleukin 2, and then admixed andtargeted with anti-GD3 antibodies, proved effective in suppressing the growth of established, progressively growing humanmelanoma tumors in nude mice (34). One may predict thatfuture treatment of human neuroblastoma may involve this typeof regimen combined with modalities involving conjugates ofchemotherapeutic drugs and radionuclides with monoclonalantibodies directed to an effective tumor target on neuroblastoma cells such as disialoganglioside GD2-
ACKNOWLEDGMENTS
The authors wish to thank Regina A. Derango, Marci Allen, LillianRamos, and David Laufer for their expert technical assistance. Thepreparation of the manuscript by Bonnie Pratt Filiault is gratefullyacknowledged.
REFERENCES
1. Evans, A. E. Natural history of neuroblastoma: In: A. E. Evans (ed.),Advances in Neuroblastoma Research, pp. 3-12. New York: Raven Press,1980.
2. Kemshead, T. J., Greaves, M. F., Walsh, F., Chayen, A., and Parkhouse, M.Monoclonal antibodies to human neuroblastoma reveal a heterogeneity inantigenic expression within the tumor. Proc. Am. Assoc. Cancer Res., 21:399, 1981.
3. Allan, P. M., Garson, J. A., Harper, E. I., Asser, U., Coakham, H. B.,Brownell, B., and Kemshead, J. T. Biological characterization and clinicalapplications of monoclonal antibody recognizing an antigen restricted toneuroectodermal tissues. Int. J. Cancer, 31: 591-598,1983.
4. Trelewen, J. (... Gibson, F. M., Ugelstad, J., Rembaum, A., Phillips, T.,Caine, G. D., and Kemshead, J. T. Removal of neuroblastoma cells frombone marrow: monoclonal antibodies conjugated to magnetic microspheres.Lancet, 1: 70-73, 1984.
5. Gomibuchi, M., Katano, M., Saxton, R. E., Lake, R., Ine, K., Morton, D.L., and Irie, R. F. Human monoclonal antibody to tumor associated ganglio-side GDI' i»vivo reactivity to human melanoma. Proc. Am. Assoc. CancerRes., 25: 248, 1984.
6. Cahan, L. D., Irie, R. F., Singh, R., (assidenti. A., and Paulson, J. C.Identification of a human neuroectodermal tumor antigen (OFA-1-2) asganglioside GDJ. Proc. Nati. Acad. Sci. USA, 79: 7629-7633, 1982.
7. Schulz, G., Cheresh, D. A., Varki, N. M., Yu, A., Staffileno, L. K., andReisfeld, R. A. Detection of ganglioside ( •,,;in tumor tissues and sera ofneuroblastoma patients. Cancer Res., 44:5914-5920, 1984.
8. Cheung, N. V., Saarinen, U. M., Neely, J. E., Landmeier, B., Donovan, D.,and Coccia, P. F. Monoclonal antibodies to a glycolipid antigen on humanneuroblastoma cells. Cancer Res., 45: 2642-2649, 1985.
9. Cheresh, D. A., and Klier, F. G. Disialoganglioside (.,,; distributes preferentially into substrate associated microprocesses on human melanoma cellsduring their attachment to ftbronectin. J. Cell Biol., 102: 1887-1897,1986.
10. Cheresh, D. A., Pierschbacher, M. D., Herzig, M. A., and Mujoo, K.Disialogangliosides <.'•„•and (.,,, are involved in the attachment of human
melanoma and neuroblastoma cells to extracellular matrix proteins. J. CellBiol., 102:688-696, 1986.
11. Cheung, N. V., Berger, N., Coccia, P., Kallick, S., Lazarus, H., Miraldi, F.,Saarinen, U., and Strandjord, S. Murine monoclonal antibody (Mab) specificfor GD2 ganglioside. A Phase I trial in patients with neuroblastoma, melanoma, and osteogenic sarcoma. Proc. Am. Assoc. Cancer Res., 27:318,1986.
12. Kohler, G., and Milstein, C. Continuous cultures of fused cells secretingantibody of pre-defined specificity. Nature (Lond.), 256:495-497, 1975.
13. Ey, P. L., Prowse, S. J., and Jenkins, C. R. Isolation of pure IgGl, IgG2aand IgG2b immunoglobulins from mouse serum using Protein-A Sepharose.Immunochemistry, 15:429-436, 1978.
14. Harper, J. R., and Orengo, A. The preparation of an enzyme-amyloglucosi-dase conjugate and its quantitation by an enzyme cycling assay. Anal.Biochem., m- 51-57, 1980.
15. Taylor, C. R. Immunoperoxidase techniques: practical and theoretical aspect.Pathol. Lab. Med., 102:113-121, 1978.
16. Ladish, S., and Gillard, B. A solvent partition method for microscale ganglioside purification. Anal. Biochem., 146: 220-231, 1985.
17. Cheresh, D. A., Varki, A. P., Varki, N. M., Stallcup, W. B., Levine, J., andReisfeld, R. A. A monoclonal antibody recognizes an O-acylated siálicacidin a human melanoma-associated ganglioside. J. Biol. Chem., 259: 7453-7459, 1984.
18. Mujoo, K., Spiro, R. C., and Reisfeld, R. A. Characterization of a uniqueglycoprotein antigen expressed on the surface of human neuroblastoma cells.J. Biol. Chem., 261: 10299-10305,1986.
19. Scatenarti. O. The attraction of proteins for small molecules and ions. Ann.NY Acad. Sci., 51:660-669, 1949.
20. Beaumier, P. L., Neuzil, D. Yang, H. M. Noll, E. A., Kishore, R., Eary, J.F., Krohn, A., Nelp, Q. V., Hellström,K. E., and Hellström,I. Immunoreac-tivity assay for anti-melanoma monoclonal antibodies. J. NucÃ.Med., 27:824-826, 1986.
21. Wormsley, S. B., Collins, M. L., and Royston, I. R. Comparative density ofhuman T cell antigen 165 on normal peripheral blood T cells and chroniclymphocytic leukemia cells. Blood, 57:657-662,1981.
22. Hakomori, S. Aberrant glycosylation in cancer cell membranes as focused onglycolipids: overview and perspective. Cancer Res., 45: 2405-2414, 1985.
23. Feizi, T. Demonstration by monoclonal antibodies that carbohydrate structures of glycoproteins and glycolipids are onco developmental antigens.Nature (Lond.), 314:53-57, 1985.
24. Cheresh, D. A. Structural and functional properties of ganglioside antigenson human tumors of neuroectodermal origin. Sun. Synth. Pathol. Res., 4:97-109, 1985.
25. Reisfeld, R. A., and Cheresh, D. A. Human tumor antigens. In: F. J. Dixon,(ed.). Advances in Immunology, in press, 1987.
26. Cheresh, D. A., Rosenberg, J., Mujoo, K., Hirschowitz, L., and Reisfeld, R.A, Biosynthesis and expression of the disialoganglioside GDI, a relevanttarget antigen on small cell lung carcinoma for monoclonal antibody-mediated cytolysis. Cancer Res., 46: 5112-5118,1986.
27. Cheresh, D. A., Honsik, C. J., Staffileno, L. K., Jung, G., and Reisfeld, R.A. Disialoganglioside GD3 on human melanoma serves as a relevant targetantigen for monoclonal antibody-mediated tumor cytolysis. Proc. Nati. Acad.Sci. USA, 82:5155-5195, 1985.
28. Hellström, K. E., Hellström, I., Goodman, G. E., and Brankovan, V. Antibody-dependent cellular cytotoxiciry to human melanoma antigens. In: R. A.Reisfeld and S. Sell (eds.). Monoclonal Antibodies and Cancer Therapy, pp.149-164. New York: Alan R. Liss, Inc., 1985.
29. Herberman, R. B., Morgan, A. C., Reisfeld, R. A., Cheresh, D. A., andOrtaldo, J. R. Antibody-dependent cellular cytotoxicity (ADCC) againsthuman melanoma by human effector cells in cooperation with mouse monoclonal antibodies. In: R. A. Reisfeld and S. Sell (eds.), Monoclonal Antibodiesand Cancer Therapy, pp. 193-203. New York: Alan R. Liss, Inc., 1985.
30. Houghton, A. N., Mintzer, D., Cordon-Cardo, C., Welt, S., Fliegl, B.,Vadhan, S., Carswell, E., Melamed, M. R., Oettgen, H. F., and Old, L. J.Mouse monoclonal IgG3 antibody detecting (.,,, ganglioside: a Phase I trialin patients with malignant melanoma. Proc. Nati. Acad. Sci. USA, 82:1242-1246, 1985.
31. Saarinen, U. M., Coccia, P. F., Gerson, S. L., Pelley, R., and Cheung, N. V.Eradication of neuroblastoma cells in vitro by monoclonal antibody andhuman complement: method for purging autologous bone marrow. CancerRes., 45: 5969-5975, 1985.
32. Herlyn, D. M., Steplewski, /.. Herlyn, M. F., and Koprowski, H. Inhibitionof growth of colorectal carcinoma in nude mice by monoclonal antibody.Cancer Res., 40:717-721, 1980.
33. Steplewski, /... Spria, G., Blaszcyzyk, M., Lübeck,M. D., Radbruch, A.,Illges, H., Herlyn, D., Rajcwsky. K., and Sellarli. M. Isolation and characterization of anti-monosialoganglioside monoclonal antibody 19-9 class-switch variants. Proc. Nati. Acad. Sci. USA, «2:8653-8657, 1985.
34. Honsik, C. J., Jung, G., and Reisfeld, R. A. Lymphokine-activated killer(LAK) cells targeted by monoclonal antibodies to disialoganglioside GM and( ¡inspecifically lyse human tumor cells of neuroectodermal origin. Proc.Nati. Acad. Sci. USA, 83:7893-7897, 1986.
1104
on July 13, 2018. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1987;47:1098-1104. Cancer Res Kalpana Mujoo, David A. Cheresh, Hsin Ming Yang, et al. Suppression of Tumor GrowthAntigen for Monoclonal Antibody-mediated Cytolysis and
on Human Neuroblastoma Cells: TargetD2Disialoganglioside G
Updated version
http://cancerres.aacrjournals.org/content/47/4/1098
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/47/4/1098To request permission to re-use all or part of this article, use this link
on July 13, 2018. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from


![Thymoquinoneloadedinnanostructured ... · SH-SY5Y human neuroblastoma cells [10], SW 626 human colon cancer cells [11], ES-2 human ovarian cancer cells [12], HeLa human cervical carcinoma](https://img.dokumen.tips/doc/110x75/604dba5a6c0cbc4067664fea/thymoquinoneloadedinnanostructured-sh-sy5y-human-neuroblastoma-cells-10-sw.jpg)







![INDEX [jpet.aspetjournals.org] · 2006. 1. 27. · sal,neuroblastoma cells,368 cyclicAMP accumulation, bicarbonate-induced sensitization, astrocytoma cells ... phosphodiesterase inhibitors](https://img.dokumen.tips/doc/110x75/60ed80783a2b603b9b2594f9/index-jpet-2006-1-27-salneuroblastoma-cells368-cyclicamp-accumulation.jpg)


















