Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Apr. 2011, p. 1606–1614 Vol. 79, No. 40019-9567/11/$12.00 doi:10.1128/IAI.01187-10Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Differential Requirements for NAIP5 in Activation ofthe NLRC4 Inflammasome!
Karla L. Lightfield,1†‡ Jenny Persson,2†§ Norver J. Trinidad,2 Sky W. Brubaker,2¶ Eric M. Kofoed,2John-Demian Sauer,2 Eric A. Dunipace,2 Sarah E. Warren,3,4
Edward A. Miao,3 and Russell E. Vance2*School of Public Health,1 Division of Immunology & Pathogenesis,2 Department of Molecular & Cell Biology, University of
California, Berkeley, California 94720; Institute for Systems Biology, Seattle, Washington 981033; and Department ofImmunology, University of Washington, Seattle, Washington 981954
Received 8 November 2010/Returned for modification 2 December 2010/Accepted 20 January 2011
Inflammasomes are cytosolic multiprotein complexes that assemble in response to infectious or noxiousstimuli and activate the CASPASE-1 protease. The inflammasome containing the nucleotide binding domain-leucine-rich repeat (NBD-LRR) protein NLRC4 (interleukin-converting enzyme protease-activating factor[IPAF]) responds to the cytosolic presence of bacterial proteins such as flagellin or the inner rod componentof bacterial type III secretion systems (e.g., Salmonella PrgJ). In some instances, such as infection withLegionella pneumophila, the activation of the NLRC4 inflammasome requires the presence of a second NBD-LRR protein, NAIP5. NAIP5 also is required for NLRC4 activation by the minimal C-terminal flagellin peptide,which is sufficient to activate NLRC4. However, NLRC4 activation is not always dependent upon NAIP5. In thisreport, we define the molecular requirements for NAIP5 in the activation of the NLRC4 inflammasome. Wedemonstrate that the N terminus of flagellin can relieve the requirement for NAIP5 during the activation of theNLRC4 inflammasome. We also demonstrate that NLRC4 responds to the Salmonella protein PrgJ indepen-dently of NAIP5. Our results indicate that NAIP5 regulates the apparent specificity of the NLRC4 inflam-masome for distinct bacterial ligands.
The innate immune system initiates defense against infec-tious agents by employing germ line-encoded receptors to de-tect microbial molecules (also called pathogen-associated mo-lecular patterns, or PAMPs) (10). Examples of PAMPs includelipopolysaccharide, cell wall components, bacterial nucleic ac-ids, and flagellin. The mammalian Toll-like receptors (TLRs)are transmembrane receptors that are capable of detectingmicrobial products at the cell surface and within intracellularcompartments (26). Signaling downstream of TLR stimulationresults in the activation of NF-!B and the induction of proin-flammatory cytokines, chemokines, and other antimicrobialdefenses. In addition to TLRs, there are several types of re-ceptors that lack transmembrane domains and function tosense PAMPs within the cytosol (26). For example, the cyto-solic presence of RNA is detected by the MDA5/RIG-I familyof RNA helicases. It is believed that cytosolic immunosurveil-lance permits host cells to make specialized or unique re-sponses to intracellular pathogens and thus to distinguish thesepathogens from extracellular microbes that do not access thehost cell cytosol (27).
The nucleotide binding domain (NBD)-leucine-rich repeat
(LRR)-containing proteins constitute an important family ofcytosolic immunosensors. Certain NBD-LRR proteins, includ-ing NAIP5 and NLRC4, are involved in orchestrating the as-sembly and activation of multiprotein complexes called inflam-masomes (24). The primary function of inflammasomes is toactivate the cysteine protease CASPASE-1 (CASP-1), which isproduced initially as an inactive proprotein and requires re-cruitment to inflammasomes to become activated (16). Onceactivated, CASP-1 is required for the proteolytic processingand secretion of the proinflammatory cytokines interleukin-1"(IL-1") and IL-18. In addition, activated CASP-1 can induce arapid, inflammatory cell death termed pyroptosis (2).
Several distinct NBD-LRR proteins are expressed by hostcells and appear to dictate inflammasome assembly in responseto specific stimuli. For example, the NBD-LRR proteinNLRP3 (NALP3; cryopyrin) orchestrates the assembly of aninflammasome that responds to a wide variety of stimuli, in-cluding crystalline substances such as uric acid, asbestos, andalum (24). In the mouse, the related NBD-LRR proteinNLRP1B (NALP1B) activates a distinct inflammasome in re-sponse to anthrax lethal toxin (3). NBD-LRR proteins are notthe only sensor-scaffold proteins responsible for CASP-1 acti-vation. For example, the PYHIN family member AIM2 bindscytosolic DNA and is responsible for CASP-1 activation inresponse to infection with bacterial pathogens (7, 11, 12, 21,23, 28).
The inflammasome containing the NBD-LRR proteinNLRC4 (interleukin-converting enzyme protease-activatingfactor [IPAF]) is one of the most well-characterized inflam-masomes, and it has been shown to activate CASP-1 specifi-cally in response to a conserved domain within the bacterial
* Corresponding author. Mailing address: 415 Life Science Addi-tion, Department of Molecular and Cell Biology, University of Cali-fornia, Berkeley, CA 94720. Phone: (510) 643-2795. Fax: (510) 642-1386. E-mail: [email protected].
† These authors contributed equally.‡ Present address: Department of Microbiology & Immunology,
Stanford University, Stanford, CA 94305.§ Present address: Department of Microbiology, Tumor and Cell
Biology, Karolinska Institute, Stockholm, Sweden.¶ Present address: Children’s Hospital Boston, Boston, MA 02115.! Published ahead of print on 31 January 2011.
1606

protein flagellin (1, 9, 14, 17), as well as to the conserved innerrod component of the type III secretion system, called PrgJ inSalmonella (18). Interestingly, in some instances, the activationof the NLRC4 inflammasome requires the presence of a sec-ond NBD-LRR protein, NAIP5 (formerly BIRC1E) (14, 19,22, 30). NAIP5 is believed to heterooligomerize with NLRC4(5, 30), but the precise biochemical function of NAIP5 inNLRC4 activation remains enigmatic. It was previously shownthat NAIP5 is required for CASP-1 activation in response tosome, but not all, NLRC4-dependent stimuli (14). For exam-ple, NAIP5 was required for the NLRC4-dependent activationof CASP-1 in response to Legionella pneumophila, whereasNaip5 deficiency had only partial effects on NLRC4-dependentresponses to Salmonella and Pseudomonas (14). NAIP5/NLRC4/CASP-1 activation by L. pneumophila is dependentlargely on amino acids within the C terminus of flagellin (14),and accordingly the retrovirus-mediated expression of a mini-mal C-terminal peptide from flagellin activates CASP-1-medi-ated pyroptotic cell death in a manner completely dependenton NLRC4 and NAIP5 (14). Unexpectedly, however, the ret-rovirus-mediated expression of full-length flagellin activatedNLRC4- and CASP-1-dependent pyroptosis independently ofNAIP5 (14).
The molecular basis for the differential requirements forNAIP5 in NLRC4 activation is not well understood. Legionellaand Salmonella both activate NLRC4 primarily via the trans-location of flagellin to the cytosol of host cells, but the flagellinmolecules translocated exhibit amino acid differences in thecritical C-terminal domain. Moreover, while Salmonella trans-locates flagellin into host cells via its SPI-I type III secretionsystem (T3SS) (25), Legionella lacks a T3SS and instead utilizesthe evolutionarily unrelated Dot/Icm type IV secretion system(T4SS) to translocate flagellin into host cells. T3SSs, but notT4SSs, contain homologs of the bacterial protein PrgJ, anotherknown activator of NLRC4 (18). Thus, differences in flagellinitself, and/or the flagellin-translocating apparatus, may under-lie the differential requirements for NAIP5 in NLRC4 activa-tion.
In this paper, we dissect the molecular features that dictatethe requirement for NAIP5 in the activation of the NLRC4inflammasome. We demonstrate that L. pneumophila engi-neered to express Salmonella flagellin activates NLRC4 in astrictly NAIP5-dependent manner, thus ruling out polymor-phisms within flagellin as an explanation for the differentialrequirement for NAIP5 in the activation of NLRC4 by the twobacterial species. We further demonstrate that the N terminusof flagellin, while not sufficient itself to activate NLRC4, nev-ertheless is able to transform the C terminus of flagellin froman NAIP5-dependent activator of the NLRC4 inflammasometo an NAIP5-independent activator. Lastly, we show that thePrgJ protein can activate NLRC4 without a requirement forNAIP5. Taken together, our results suggest a model in whichNAIP5 functions to dictate the specificity of NLRC4 for dis-tinct stimuli.
MATERIALS AND METHODS
Mice. Wild-type (WT) C57BL/6J (B6) mice were from Jackson Laboratories.Nlrc4#/# mice on a pure B6 background (15) were obtained from S. Mariathasanand V. Dixit (Genentech). Naip5#/# mice on a pure B6 background mice weredescribed previously (14).
Bacterial strains. The Salmonella enterica serovar Typhimurium strain LT2and the flagellin mutant S. Typhimurium LT2FliC/FljB# were gifts from A. VanDer Velden and M. Starnbach. The pLIV2-L.p.FlaA and pLIV2-S.t.PrgJ con-structs were constructed as isopropyl-"-D-thiogalactopyranoside (IPTG)-induc-ible variants of previously described strains. A construct fusing the amino-ter-minal 300 bp of the actA gene with the flaA gene from L.p.FlaA was previouslydescribed and used to facilitate the ectopic secretion of Legionella pneumophilaFlaA (J.-D. Sauer et al., submitted for publication). The actA-L.p.FlaA fusionwas amplified from the pPL2-L.p.FlaA plasmid with the primers AAAAGCGGCCGGTGGGATTAAATAGATTTATGCGTGC and AAAAGTCGACCAGAAATCGAAGTGCAGTTG using Pfu Ultra II (Agilent Technologies, SantaClara, CA), where underlining indicates the restriction sites. The resulting PCRproduct and the IPTG-inducible variant of pPL2, pLIV2 (8), were double di-gested with EagI and SalI (New England Biosystems, Waltham, MA) and ligatedusing the NEB quick ligation kit. The resulting vector, pLIV2-L.p.FlaA, subse-quently was integrated at the tRNAArg locus on the L. monocytogenes 10403schromosome as previously described (13). pLIV2-S.t.PrgJ was constructed sim-ilarly to the previously described actA-prgJ fusion construct (Sauer et al., sub-mitted). The fusion was amplified from the pPL2-S.t.PrgJ plasmid using theprimers AAAAGCGGCCGCAGGAGGGAGTATAAGTGGGATTAAATAGand TTTTGTCGACTCATGAGCGTAATAGCGTTTC, digested, and ligatedinto pLIV2, and then it was integrated into the 10403s chromosome as previouslydescribed. Legionella strain LP02 is a streptomycin-resistant thymidine auxo-troph derivative of Legionella pneumophila strain LP01. LP02$flaA contains anunmarked deletion of flaA and was described previously (22). LP02$flaA::fliCexpresses Salmonella fliC from the chromosome under the control of the endog-enous Legionella flaA promoter. To generate this strain, the flaA promoter fromLP02 was amplified by PCR using primers pflaAFwd (ATAGTCGACTTAATGCCTCTTTCTCTCCTGTCG) and pflaARev (AATGACTTGTGCCATAATTTTAGTCTCCTCAGACCTGAATCC), while fliC was amplified from S. Typhi-murium using primers fliCfwd (GAGGAGACTAAAATTATGGCACAAGTCATTAATACAAACAGC) and fliCrev (ATAGGATCCTTAACGCAGTAAAGAGAGGACGTTTTG). These amplicons were mixed and spliced in a secondround of PCR using pflaAFwd and fliCrev and cloned into the allelic exchangevector pSR47S. This suicide construct was introduced onto the LP02$flaA chro-mosome by a single crossover and selection with kanamycin (50 %g/ml).
Retroviral constructs and transductions. Retroviral constructs were generatedand transductions were performed as previously described (14). Briefly, retrovi-ral particles were generated by the transient transfection of Phoenix-Eco pack-aging cells with MSCV2.2-based retroviral vectors. Bone marrow-derived cells(1 & 106) were cultured for 48 h in a 6-well plate in medium containing macro-phage colony-stimulating factor (M-CSF) and then were transduced with 1 ml ofretrovirus-containing packaging cell supernatant. Macrophages were analyzed 3to 4 days posttransduction and analyzed for GFP expression on a BeckmanCoulter FC-500 flow cytometer. More than 10,000 cells were analyzed per sam-ple. When we simultaneously transduced cells with multiple constructs (see Fig.2), equal volumes of each retroviral supernatant were used. These macrophageswere analyzed on a BD Influx cell sorter for green fluorescent protein (GFP) andmCherry expression. The retroviral constructs used in this paper were generatedas follows: GFP-C65 was constructed by amplifying the C-terminal 65 aminoacids of flagellin using the primers pC65fwd (GACGAGCTGTACAAGTTTGAATCAACGATAG) and pFlaA65rev (TTGCGGCCGCCTATCGACCTAACAAAGATAATACAGATTGCG), while GFP was amplified using the primerspGFPfwd (ATAAGATCTCCACCATGGTGAGCAAGGGCGAGGA) andpGFP65rev (CTATCGTTGATTCAAACTTGTACAGCTCGTC). These ampli-cons were mixed and spliced in a second round of PCR using the primerspGFPfwd and pFlaArev and were cloned into the retroviral expression vectorpMSCV2.2 using the enzymes BglII and NotI. Similarly, the construct N65-GFP-C65 was generated by amplifying GFP-C65 with the primers pN65GFPfwd (GGATGAACCAAGCCGTTATGGTGAGCAAGGGC) and pFlaA65rev, whilethe N-terminal 65 amino acids of flagellin were amplified with the primerspN65fwd (ATAAGATCTCCACCATGGCTCAAGTAATCAACACTAATGTGG) and pN65GFPrev (GCCCTTGCTCACCATAACGGCTTGGTTCATCC).These amplicons were spliced in a second round of PCR using the primers pN65fwdand pFlaA65rev. N65-GFP was generated by amplifying the N-terminal 65 aminoacids of flagellin with the primers pN65fwd and pN65GFPrev while amplifying GFPwith the primers pN65GFPfwd and pGFPrev (TTGCGGCCGCTTACTTGTACAGCTCGTC). These amplicons were mixed and spliced in a second round of PCRusing the primers pN65fwd and pGFPrev. The constructs N65-GFP-C65 L12L32::IIand N65-GFP-C65 were generated using site-directed mutagenesis on the parentalN65-GFP-C65 construct, pL12fwd (CACTAATGTGGCGTCGatcACAGCCCAACGTAATTTGGG), pL12rev (CCCAAATTACGTTGGGCTGTgatCGACGCCACATTAGTG), pL32fwd (CATCGATCCAGCGTatcTCATCGGGATTAAGG),
VOL. 79, 2011 REQUIREMENT FOR NAIP5 IN NLRC4 INFLAMMASOME ACTIVATION 1607

and pL32rev (CCTTAATCCCGATGAgatACGCTGGATCGATG) for L12L32::IIand pL12Afwd (CACTAATGCAGCGTCGGCAACAGCCCAACG), pL12Arev(CGTTGGGCTGTT GCCGACGCTGCATTAGTG), pI5V9Aafwd (GGCTCAAGTAGCAAACACTAATGCAGCGTCGCTCAC), and pI5V9AArev(GTGAGCGACGCTGCATTAGTGTTT GCTACTTGAGCC) for I5V9L12:AAA. mCherry-C65 was generated by amplifying mCherry using pmCherryfwd (ATAAGATCTCCACCATGGTGAGCAAGGGCGAGG) and pmCherry65rev (GGCTATCGTTGATTCAAACTTGTACAGCTCGTCC), while the C-terminal 65 amino acids offlagellin were amplified using the primers pmCherry65fwd (GGACGAGCTGTACAAGTTTGAATCAACGATGCC) and pFlaA65rev. These amplicons were mixedand spliced during a second round of PCR with the primers pmCherryfwd andpFlaA65rev and cloned into pMSCV2.2 with BglII and NotI.
Cytotoxicity assays. Cytotoxicity was measured as described previously (6) bydetermining the activity of lactate dehydrogenase (LDH) released by cells. Bonemarrow-derived macrophages (5 & 104 to 1 & 105) were seeded onto tissueculture-treated 96-well plates and infected with bacteria at the indicated multi-plicity of infection. Plates were centrifuged at 400 & g to ensure the equivalentinfectivity of WT and flagellin mutant strains. At 30 min to 1 h postinfection,medium was removed and replaced with medium containing 10 to 100 %g/mlgentamicin to kill extracellular bacteria. Six hours after infection, plates werecentrifuged at 400 & g, and supernatant was collected and assayed for LDHrelease. Legionella strains used for infection were grown overnight to an opticaldensity of '3.8. Salmonella cultures were grown overnight with shaking at 37°Cand were diluted 3 h prior to infection at 1:40 in 1 ml of LB and regrown withshaking until reaching late-exponential phase. Listeria strains were grown asstanding cultures at 37°C in brain heart infusion (BHI) medium. In the experi-ments with Listeria, infected cells were treated with IPTG to induce the expres-sion of flaA or prgJ. The percent specific lysis of infected cells was calculatedas the following: (amount of LDH released) # (background release by un-infected cells)/(amount of LDH released by detergent lysed cells minus back-ground) & 100.
Growth curves. Bacterial growth was determined as previously described (4).Macrophages were plated at a density of 5 & 105 cells per ml in 24-well plates,allowed to adhere overnight, and infected with stationary-phase Legionella (OD,'3.8) at a multiplicity of infection (MOI) of 0.01. CFU were determined bylysing washed macrophage monolayers with sterile distilled water and platinglysates on buffered charcoal yeast extract plates.
Protein transfection. Salmonella FliC, PrgJ, and SsaI proteins were purified asdescribed previously (18). Macrophages were plated at 5 & 104 per well in a96-well plate, allowed to adhere overnight, and primed with tripalmitoyl cysteinylseryl tetralysine lipopeptide (Pam3CSK4; 0.5 %g/ml) for 3.5 h prior to proteintransfection to induce the expression of pro-interleukin-1" (pro-IL-1"). Protein(0.2 %g/well) was transfected using the Profect P1 (Targeting Systems) transfec-tion reagent according to the manufacturer’s instructions. Supernatants werecollected for analysis 3 h posttransfection and analyzed by enzyme-linked immu-nosorbent assay (ELISA) for IL-1" (R&D Systems).
Statistics. Statistical significance was assessed by the t test or one-way analysisof variance (ANOVA) with Bonferroni posttests. Graph Pad Prism 5 softwarewas used for statistical analyses.
RESULTS
The N terminus of flagellin can relieve the requirement forNAIP5 in flagellin sensing. To better understand the molecu-lar basis for NAIP5/NLRC4 activation, we first sought to de-termine what regions of flagellin enable full-length flagellin tobe toxic to Naip5#/# macrophages in the retroviral lethalityassay (14). In this assay, flagellin is expressed downstream of amammalian promoter in macrophages using a retrovirus-basedconstruct. The transduction of macrophages with a constructexpressing a stimulatory flagellin molecule results in the rapidpyroptotic cell death of the macrophages and thereby preventsthe expression of a coexpressed GFP reporter (14). Con-versely, the transduction of a construct encoding a nonstimu-latory flagellin mutant results in GFP( cells (14). Previously,the retroviral lethality assay was used to establish that NLRC4and NAIP5 both are required for the response to the C-ter-minal domain of flagellin, whereas pyroptotic cell death inresponse to full-length flagellin required NLCR4 but not
NAIP5 (14). Thus, to determine what regions in full-lengthflagellin relieve the requirement for NAIP5, we began by mak-ing a series of retroviral constructs expressing N-terminallydeleted flagellin molecules (Fig. 1A). These constructs weretransduced into primary macrophages. As shown previously(14), retroviral expression constructs encoding full-lengthflagellin (FlaA) did not stably transduce C57BL/6 (WT) orisogenic Naip5#/# macrophages, as evidenced by a lack ofGFP( cells 3 days after transduction (Fig. 1B). However, wefound that a retroviral construct that expresses a flagellin mol-ecule lacking the N-terminal 65 amino acids from flagellin(FlaAN$65) could be transduced into Naip5#/# macrophages(Fig. 1B). Consistently with our previous observation that theC terminus of flagellin is sufficient to activate NLRC4,FlaAN$65 (which retains the normal flagellin C terminus)could not be transduced into WT macrophages. Further, N-terminal deletions of FlaA ($N85, $N100, and $N125) resem-bled $N65 and could be transduced into Naip5#/# macro-phages (Fig. 1B). These results suggest that the N terminus offlagellin is important for the ability of full-length flagellin toactivate NLRC4 independently of NAIP5.
Molecular basis by which the N terminus affects the require-ment for Naip5. We next sought to address whether the Nterminus of flagellin is sufficient to convert the C terminus offlagellin into an NAIP5-independent activator of NLRC4.Consistently with our previous studies (14), we found that theC-terminal 65 amino acids of flagellin, fused to GFP, is anNAIP5-dependent activator of NLRC4 (Fig. 1C). In contrast,full-length flagellin (whether or not it is fused to GFP) is anNAIP5-independent activator of NLRC4 (Fig. 1B and C). Todetermine the role of the N terminus in NAIP5/NLRC4 acti-vation, we created an expression construct that consisted ofGFP flanked by the N-terminal and C-terminal 65 amino acidsof FlaA (Fig. 1A). The resulting N65-GFP-C65 construct be-haved just like full-length flagellin; namely, it was cytotoxic tomacrophages in a manner requiring NLRC4 but independentof the presence or absence of NAIP5 (Fig. 1C). The N termi-nus of flagellin fused to GFP was itself noncytotoxic and couldbe transduced into WT (Nlrc4() macrophages (Fig. 1C). Thus,the N terminus of flagellin is not sufficient to activate NLRC4.However, the N terminus of flagellin appears to enhance oralter NLRC4-dependent responses to the C terminus of flagel-lin in such a way that NAIP5 is no longer required.
Although the C-terminal region of flagellin is thought to beunstructured when flagellin is in its monomeric form, the struc-ture of flagellin within the flagellin filament (29) suggests thatthe N and C termini of flagellin interact weakly via a coiled-coilinteraction. Moreover, the crystal structure of GFP reveals thatits N and C termini are near each other (20), and it thereforeis possible that the appended N and C termini of flagellininteract with each other in the N65-GFP-C65 construct. Thus,we hypothesized that a weak coiled-coil interaction with the Nterminus of flagellin resulted in a stabilized or structurallyaltered C-terminal region that no longer required NAIP5 forthe stimulation of NLRC4. To test this idea, we mutated the Nterminus of flagellin within the N65-GFP-C65 construct todisrupt the putative interaction between the N and C termini.Two leucines (L12 and L32) in the N terminus of flagellin weremutated to more bulky isoleucines, which would not be easilyaccommodated within the core of a coiled coil and thus should
1608 LIGHTFIELD ET AL. INFECT. IMMUN.

result in a disruption of putative coiled-coil interactions. Inaddition, a series of mutations predicted to be even moredisruptive to the putative coiled-coil interaction were made,namely, isoleucine 5, valine 9, and leucine 12 all were mutatedto alanines (I5V9L12:AAA). Neither series of mutations wassufficient to transform the parental N65-GFP-C65 stimulusfrom an NAIP5-independent stimulus (N65-GFP-C65) into anNAIP5-dependent one (Fig. 1D and E). Thus, we were unable
to find evidence that a coiled-coil interaction between the Nand C termini of flagellin is important for the ability of full-length flagellin to activate NLRC4 independently of NAIP5.These results do not rule out the possibility that an interactionbetween the N and C termini of flagellin (that we did notdisrupt with the mutations we made) is important in determin-ing the requirement for NAIP5 in NLRC4 activation. How-ever, a lack of a role for a coiled-coil interaction would not be
FIG. 1. Role for the N terminus of flagellin in NLRC4 activation. (A) Diagram of all retroviral constructs used in transduction experiments.Note that all constructs expressed GFP, either as a direct fusion to L. pneumophila flagellin (FlaA; as shown) or downstream of an internalribosome entry site (IRES) (not shown). C65, C-terminal 65 amino acids of FlaA; C20, C-terminal 20 amino acids of FlaA; N65, N-terminal 65amino acids of FlaA. FlaA$N constructs lack the indicated number of amino acids from the N terminus. (B and C) Wild-type (C57BL/6) or isogenicNaip5#/# or Nlrc4#/# macrophages were transduced with the indicated constructs, and the percentage of GFP( cells was enumerated by flowcytometry 3 to 4 days after transduction. (D and E) Wild-type, Naip5-deficient, or Naip5#/# Nlrc4#/# doubly deficient macrophages weretransduced with the indicated constructs, and the percentage of GFP( cells was enumerated by flow cytometry 3 to 4 days after transduction. Morethan 10,000 cells were analyzed for each experiment. For each panel, data shown are from a single representative experiment of at least three thatproduced similar results.
VOL. 79, 2011 REQUIREMENT FOR NAIP5 IN NLRC4 INFLAMMASOME ACTIVATION 1609

surprising, given that the interaction between the N and Ctermini of flagellin likely is very weak (29) and probably notsignificant when flagellin is in its monomeric form, which is theform that appears to be recognized by NLRC4 (17).
The N and C termini of flagellin can act in trans to activateNLRC4. Because coiled-coil interactions did not appear to berequired for the NAIP5-independent activation of NLRC4 byflagellin, we questioned whether the N and C termini of flagel-lin needed to be expressed on the same molecule or whetherthey could cooperate in trans to activate NLRC4 withoutNAIP5. To address this question, a construct encoding theN-terminal 65 amino acids of flagellin fused to GFP (N65-GFP), and a separate construct encoding the C-terminal 65amino acids of flagellin fused to mCherry (mCherry-C65),were expressed simultaneously in wild-type, Naip5-deficient,and Nlrc4-deficient macrophages via retroviral transduction.As expected, mCherry-C65 was cytotoxic to wild-type macro-phages but not Naip5#/# or Nlrc4#/# macrophages (Fig. 2A toC). While a robust population of brightly double-positive mac-rophages was detected in transduced Nlrc4#/# macrophages,Naip5#/# macrophages harbored only a proportionallysmaller, less bright population of double-positive macrophages(Fig. 2B and C). These results indicate that, at the relativelyhigh expression levels achieved by retroviral transduction, theN terminus of flagellin can act in trans to convert the C termi-nus of flagellin into an NAIP5-independent stimulus. We wereunable to detect a physical interaction between mCherry-C65and N65-GFP by immunoprecipitation (data not shown).Taken together, our data raise the possibility that the N and Ctermini cooperate to activate NLRC4 without physically inter-acting with each other, although a role for a transient or weakphysical interaction cannot be rule out.
NAIP5-independent detection of Salmonella. We previouslyreported that while the activation of the inflammasome byLegionella was strictly flagellin and NAIP5 dependent, the ac-tivation of the inflammasome by Salmonella was partiallyNAIP5 independent. It is difficult to compare infections be-tween two different bacterial species when there may be dif-ferences in infectivity, translocation levels of flagellin, or flagel-
lin molecules themselves. Thus, to begin to determine if thereis in fact a qualitative difference between Salmonella and Le-gionella with respect to the activation of NLRC4, we assayedmacrophage cell death at a wide range of MOIs for bothbacterial species (Fig. 3). We centrifuged bacteria onto themacrophages to minimize differences in infectivity betweenmotile and nonmotile strains and carefully monitored the in-fectivity of the various strains by determining gentamicin-re-sistant cell-associated CFU 1 h after infection. We found thatthe infection of macrophages with Legionella causes macro-phage cell death in a manner dependent upon both the hostexpression of NAIP5 and NLRC4 and the bacterial expressionof flagellin across a broad range of MOIs, from 0.3 to 10 (Fig.3A). In contrast, we found that Salmonella causes a partiallyflagellin-independent cell death at higher bacterial loads (e.g.,MOI, '2) (Fig. 3A), which is consistent with previous results(18). As previously described (14), we also found that at thesehigher MOIs, Salmonella also induces a partially NAIP5-inde-pendent (but NLCR4-dependent) cell death. Some of theNAIP5-independent death appeared to be flagellin dependent,but some appeared to be flagellin independent. The flagellin-dependent activation of NLRC4 could not be explained by areduced infectivity of the $fliC Salmonella mutant, since theCFU per cell were quantified and not found to be significantlydifferent. Thus, the activation of NLRC4 by Salmonella seemsto be only partially NAIP5 and flagellin dependent. This con-trasts with the activation of NLRC4 by Legionella, which wasmore uniformly NAIP5 and flagellin dependent.
To further assess the underlying differences in NLRC4 ac-tivation between Salmonella and Legionella, we asked whetherthe NAIP5-independent death initiated by Salmonella is due toan intrinsic difference between Legionella and Salmonellaflagellin. Thus, we generated a strain of Legionella that ex-presses Salmonella flagellin (fliC) in place of its own flagellin.This strain (LP02 $flaA::fliC) was capable of activating theNLRC4 inflammasome almost as efficiently as wild-type Legion-ella (22), and interestingly, it did so in a fully NAIP5-depen-dent manner (Fig. 3B). In addition, complementing flagellin-deficient Legionella with Salmonella fliC restored the
FIG. 2. N terminus of flagellin can relieve requirement for NAIP5 in flagellin sensing even when expressed on a distinct molecule from the Cterminus. A retroviral construct expressing the N-terminal 65 amino acids of flagellin fused to GFP, and a separate retroviral construct expressingthe C-terminal 65 amino acids of flagellin fused to mCherry, were transduced simultaneously into wild-type (A), Naip5#/# (B), or Nlrc4#/#
(C) macrophages, and the percentage of cells expressing GFP and/or mCherry was enumerated by flow cytometry 4 days posttransduction. Thepercentage of double-positive cells among all transduced cells is indicated in parentheses. The results are representative of two representativeexperiments.
1610 LIGHTFIELD ET AL. INFECT. IMMUN.
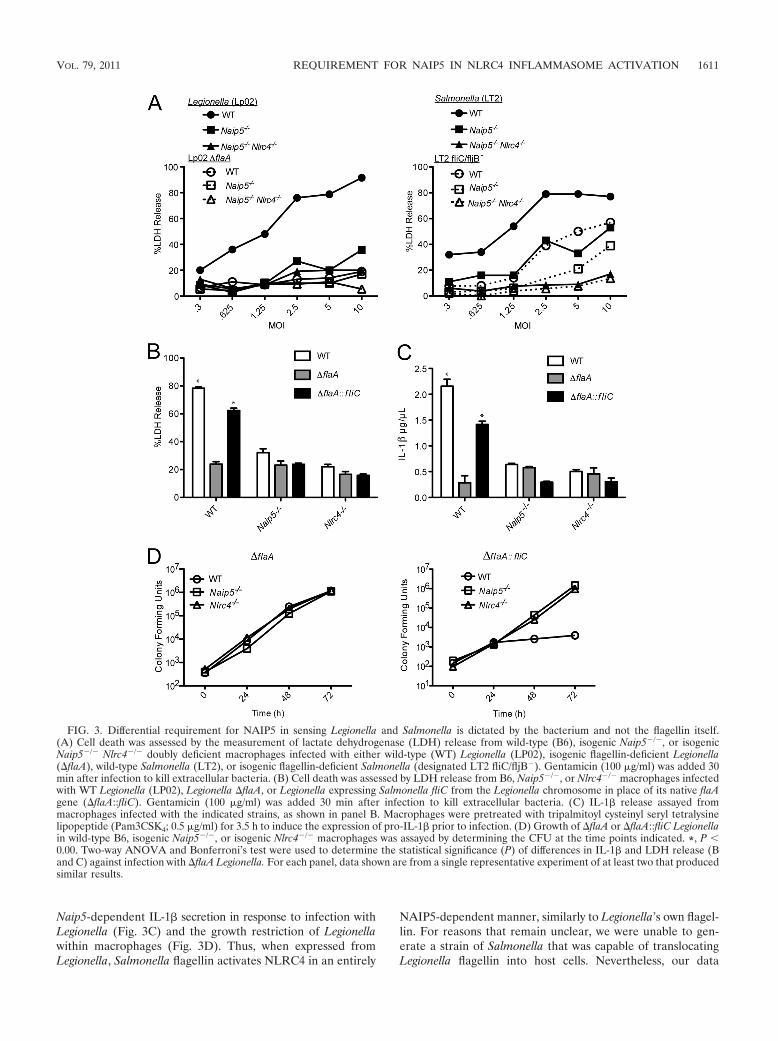
Naip5-dependent IL-1" secretion in response to infection withLegionella (Fig. 3C) and the growth restriction of Legionellawithin macrophages (Fig. 3D). Thus, when expressed fromLegionella, Salmonella flagellin activates NLRC4 in an entirely
NAIP5-dependent manner, similarly to Legionella’s own flagel-lin. For reasons that remain unclear, we were unable to gen-erate a strain of Salmonella that was capable of translocatingLegionella flagellin into host cells. Nevertheless, our data
FIG. 3. Differential requirement for NAIP5 in sensing Legionella and Salmonella is dictated by the bacterium and not the flagellin itself.(A) Cell death was assessed by the measurement of lactate dehydrogenase (LDH) release from wild-type (B6), isogenic Naip5#/#, or isogenicNaip5#/# Nlrc4#/# doubly deficient macrophages infected with either wild-type (WT) Legionella (LP02), isogenic flagellin-deficient Legionella($flaA), wild-type Salmonella (LT2), or isogenic flagellin-deficient Salmonella (designated LT2 fliC/fljB#). Gentamicin (100 %g/ml) was added 30min after infection to kill extracellular bacteria. (B) Cell death was assessed by LDH release from B6, Naip5#/#, or Nlrc4#/# macrophages infectedwith WT Legionella (LP02), Legionella $flaA, or Legionella expressing Salmonella fliC from the Legionella chromosome in place of its native flaAgene ($flaA::fliC). Gentamicin (100 %g/ml) was added 30 min after infection to kill extracellular bacteria. (C) IL-1" release assayed frommacrophages infected with the indicated strains, as shown in panel B. Macrophages were pretreated with tripalmitoyl cysteinyl seryl tetralysinelipopeptide (Pam3CSK4; 0.5 %g/ml) for 3.5 h to induce the expression of pro-IL-1" prior to infection. (D) Growth of $flaA or $flaA::fliC Legionellain wild-type B6, isogenic Naip5#/#, or isogenic Nlrc4#/# macrophages was assayed by determining the CFU at the time points indicated. *, P )0.00. Two-way ANOVA and Bonferroni’s test were used to determine the statistical significance (P) of differences in IL-1" and LDH release (Band C) against infection with $flaA Legionella. For each panel, data shown are from a single representative experiment of at least two that producedsimilar results.
VOL. 79, 2011 REQUIREMENT FOR NAIP5 IN NLRC4 INFLAMMASOME ACTIVATION 1611
strongly suggest that there are not intrinsic differences betweenthe flagellins that underlie the differing abilities of Legionellaand Salmonella to activate the NLRC4 inflammasome inde-pendently of NAIP5. These results are consistent with previousexperiments that demonstrated that the C termini of Legionellaand Salmonella flagellin behaved identically in the retrovirallethality assay (14).
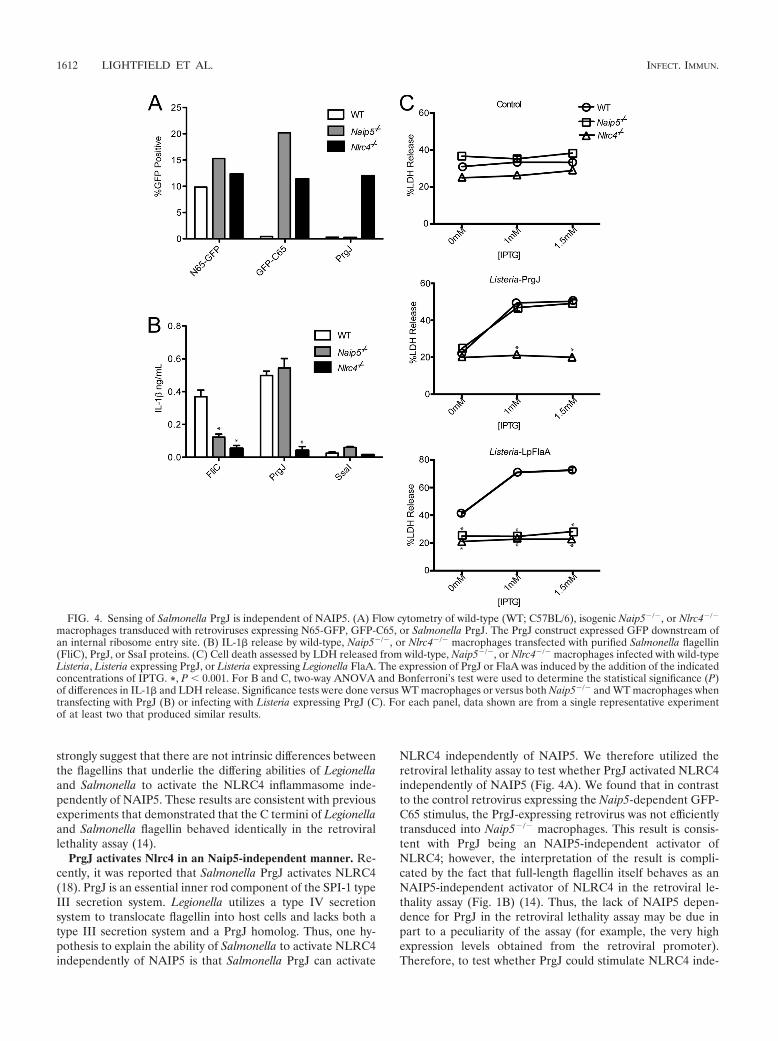
PrgJ activates Nlrc4 in an Naip5-independent manner. Re-cently, it was reported that Salmonella PrgJ activates NLRC4(18). PrgJ is an essential inner rod component of the SPI-1 typeIII secretion system. Legionella utilizes a type IV secretionsystem to translocate flagellin into host cells and lacks both atype III secretion system and a PrgJ homolog. Thus, one hy-pothesis to explain the ability of Salmonella to activate NLRC4independently of NAIP5 is that Salmonella PrgJ can activate
NLRC4 independently of NAIP5. We therefore utilized theretroviral lethality assay to test whether PrgJ activated NLRC4independently of NAIP5 (Fig. 4A). We found that in contrastto the control retrovirus expressing the Naip5-dependent GFP-C65 stimulus, the PrgJ-expressing retrovirus was not efficientlytransduced into Naip5#/# macrophages. This result is consis-tent with PrgJ being an NAIP5-independent activator ofNLRC4; however, the interpretation of the result is compli-cated by the fact that full-length flagellin itself behaves as anNAIP5-independent activator of NLRC4 in the retroviral le-thality assay (Fig. 1B) (14). Thus, the lack of NAIP5 depen-dence for PrgJ in the retroviral lethality assay may be due inpart to a peculiarity of the assay (for example, the very highexpression levels obtained from the retroviral promoter).Therefore, to test whether PrgJ could stimulate NLRC4 inde-
FIG. 4. Sensing of Salmonella PrgJ is independent of NAIP5. (A) Flow cytometry of wild-type (WT; C57BL/6), isogenic Naip5#/#, or Nlrc4#/#
macrophages transduced with retroviruses expressing N65-GFP, GFP-C65, or Salmonella PrgJ. The PrgJ construct expressed GFP downstream ofan internal ribosome entry site. (B) IL-1" release by wild-type, Naip5#/#, or Nlrc4#/# macrophages transfected with purified Salmonella flagellin(FliC), PrgJ, or SsaI proteins. (C) Cell death assessed by LDH released from wild-type, Naip5#/#, or Nlrc4#/# macrophages infected with wild-typeListeria, Listeria expressing PrgJ, or Listeria expressing Legionella FlaA. The expression of PrgJ or FlaA was induced by the addition of the indicatedconcentrations of IPTG. *, P ) 0.001. For B and C, two-way ANOVA and Bonferroni’s test were used to determine the statistical significance (P)of differences in IL-1" and LDH release. Significance tests were done versus WT macrophages or versus both Naip5#/# and WT macrophages whentransfecting with PrgJ (B) or infecting with Listeria expressing PrgJ (C). For each panel, data shown are from a single representative experimentof at least two that produced similar results.
1612 LIGHTFIELD ET AL. INFECT. IMMUN.

pendently of NAIP5, it was necessary to utilize an assay inwhich full-length flagellin behaved as an NAIP5-dependentstimulus. Two such assays were employed. In the first, full-length recombinant FliC and PrgJ proteins were transfectedinto the macrophage cytosol using protein transfection as de-scribed previously (17, 18). The activation of NLRC4 was mon-itored by measuring the NLRC4/CASP-1-dependent release ofIL-1" into the supernatant (Fig. 4B). Although IL-1" releasein response to transfected FliC protein required NAIP5, IL-1"release in response to PrgJ was nearly entirely NAIP5 inde-pendent (Fig. 4B).
In the second assay to assess whether PrgJ is an NAIP5-independent activator of NLRC4, we utilized flagellin- andPrgJ-expressing strains of the Gram-positive bacterium Listeriato activate NLRC4. Listeria strains engineered to express Le-gionella flagellin (FlaA) strongly activate NLRC4-dependentcell death (Sauer et al., submitted). Although the cell deathinduced by Listeria flaA was NAIP5 dependent, cell deathinduced by Listeria prgJ was entirely NAIP5 independent (Fig.4C). Thus, we conclude that PrgJ can activate NLRC4 inde-pendently of NAIP5.
DISCUSSION
Our results demonstrate that NAIP5 is required for theactivation of the NLRC4 inflammasome only in response tocertain stimuli. These stimuli include infection with Legionella(expressing Legionella or Salmonella flagellin) or transductionwith a retrovirus expressing the C terminus of flagellin fused toGFP. Other stimuli, such as constructs expressing both the Nand C termini of flagellin, or PrgJ, appear to activate NLRC4without a requirement for NAIP5. Lastly, infection with Sal-monella activates the NLRC4 inflammasome in a partiallyNAIP5-dependent, partially NAIP5-independent manner.
The NLRC4 inflammasome has yet to be reconstituted orpurified, thus the biochemical role of NAIP5 in the activationof NLRC4 remains poorly understood. Nevertheless, our re-sults indicate that NAIP5 affects the ability of the NLRC4inflammasome to respond to specific stimuli. There are severalpossible models that account for the ability of NAIP5 to reg-ulate the apparent specificity of NLRC4. One model (the po-tentiation model) is that NAIP5 functions as a potentiator ofNLRC4 activation. In this model, the NLRC4 stimuli thatexhibit a requirement for NAIP5 are intrinsically weak activa-tors of NLRC4 and thus require NAIP5 to activate NLRC4.For example, the C terminus of flagellin might be poorly struc-tured in the absence of the N terminus of flagellin and there-fore may be a relatively poor activator of NLRC4. The pres-ence of NAIP5 could stabilize the C terminus of flagellin andthus improve its ability to stimulate NLRC4. NAIP5 also mightpotentiate NLRC4 responses by acting downstream of flagellinrecognition and act to amplify the NLRC4-dependent activa-tion of CASP-1 in response to a weak stimulus. We found thatthe N terminus of flagellin can relieve the requirement forNAIP5 in NLRC4 activation. In the potentiation model, the Nterminus might function to stabilize the C terminus, therebyimproving its signaling capacity and thus relieving the require-ment for NAIP5. A direct biochemical interaction betweenflagellin and NLRC4 or NAIP5 has yet to be demonstrated;thus, these models are difficult to distinguish at present. It also
is not clear whether Legionella is in fact a weaker activator ofNLRC4 than is Salmonella, as would be implied under thepotentiation model. In fact, at similar MOIs, Legionella andSalmonella appear to induce comparable levels of NLRC4-dependent cell death (Fig. 3). It also is not clear whether PrgJ,an NAIP5-independent activator of NLRC4, is a substantiallystronger activator of NLRC4 than is flagellin. In fact, the trans-fection of wild-type macrophages with equivalent amounts ofeach protein activates NLRC4 similarly, but PrgJ appears to bea much better activator of NLRC4 specifically in Naip5#/#
macrophages. Thus, it is not clear whether our results can bereadily explained by a model in which NAIP5 functions pri-marily to potentiate NLRC4 signaling.
Although it is widely assumed that NLRC4 is the proximalsensor of the ligands that lead to its activation, this has neverbeen formally demonstrated. In fact, it is possible that NAIP5,or another host protein, is the true direct cytosolic sensor (orreceptor) of flagellin. Thus, another model to explain our datais that NAIP5 functions not as a general potentiator of NLRC4activation/signaling but as a specific sensor of certain bacterialligands. We call the latter model the specificity model. Forexample, NAIP5 may specifically recognize the C terminus offlagellin, whereas another host protein may specifically recog-nize full-length flagellin or PrgJ. This model could explain thedifferential NAIP5 dependence for NLRC4 activation by Le-gionella versus Salmonella, since Salmonella expresses PrgJ, anNAIP5 independent stimulus, whereas Legionella does not.The specificity model is difficult to address until direct evidenceis obtained that NAIP5, NLRC4, or another host protein is thetrue receptor for flagellin and/or PrgJ.
It was interesting that full-length flagellin delivered by pro-tein transfection or Listeria activated NLRC4 in an NAIP5-dependent manner (Fig. 4B and C), whereas full-length flagel-lin expressed from a retroviral construct activated NLRC4 inan NAIP5-independent manner (Fig. 1B) (14). Although thereare many possible explanations for this result, one plausibleidea is that the very high levels of flagellin expression attainedvia retroviral transduction are responsible for relieving therequirement for NAIP5. Thus, both the expression levels of thestimulus and its molecular characteristics may influence the re-quirement for NAIP5.
Regardless of the underlying molecular mechanism, ourdata illustrate the ability of NLRC4 to respond to distinctbacterial ligands. In future studies, it will be important todetermine the specific molecular role that NAIP5 plays inNLRC4 activation.
ACKNOWLEDGMENTS
This work was supported by Investigator Awards to R.E.V. from theBurroughs Wellcome Fund and the Cancer Research Institute as wellas by NIH grants (AI075039 and AI080749). J.P. was supported byStiftelsen Olle Engkvist Byggmastare through the Swedish ResearchCouncil. J.-D.S. was supported by the American Cancer Society (PF-07-066-01-LIB).
We thank Tom Alber for help with the analysis of the putative coiledcoil in the D0 region of flagellin.
REFERENCES
1. Amer, A., et al. 2006. Regulation of Legionella phagosome maturation andinfection through flagellin and host IPAF. J. Biol. Chem. 281:35217–35223.
2. Bergsbaken, T., S. L. Fink, and B. T. Cookson. 2009. Pyroptosis: host celldeath and inflammation. Nat. Rev. Microbiol. 7:99–109.
VOL. 79, 2011 REQUIREMENT FOR NAIP5 IN NLRC4 INFLAMMASOME ACTIVATION 1613

3. Boyden, E. D., and W. F. Dietrich. 2006. Nalp1b controls mouse macrophagesusceptibility to anthrax lethal toxin. Nat. Genet. 38:240–244.
4. Coers, J., R. E. Vance, M. F. Fontana, and W. F. Dietrich. 2007. Restrictionof Legionella pneumophila growth in macrophages requires the concertedaction of cytokine and Naip5/Ipaf signalling pathways. Cell Microbiol.9:2344–2357.
5. Damiano, J. S., V. Oliveira, K. Welsh, and J. C. Reed. 2004. Heterotypicinteractions among NACHT domains: implications for regulation of innateimmune responses. Biochem. J. 381:213–219.
6. Decker, T., and M. L. Lohmann-Matthes. 1988. A quick and simple methodfor the quantitation of lactate dehydrogenase release in measurements ofcellular cytotoxicity and tumor necrosis factor (TNF) activity. J. Immunol.Methods 115:61–69.
7. Fernandes-Alnemri, T., et al. 2010. The AIM2 inflammasome is critical forinnate immunity to Francisella tularensis. Nat. Immunol. 11:385–393.
8. Fischetti, V. A., and American Society for Microbiology. 2006. Gram-positivepathogens, 2nd ed. ASM Press, Washington, DC.
9. Franchi, L., et al. 2006. Cytosolic flagellin requires Ipaf for activation ofcaspase-1 and interleukin 1beta in salmonella-infected macrophages. Nat.Immunol. 7:576–582.
10. Janeway, C. A., Jr. 1989. Approaching the asymptote? Evolution and revo-lution in immunology. Cold Spring Harb. Symp. Quant. Biol. 54(Pt 1):1–13.
11. Jones, J. W., et al. 2010. Absent in melanoma 2 is required for innateimmune recognition of Francisella tularensis. Proc. Natl. Acad. Sci. U. S. A.107:9771–9776.
12. Kim, S., et al. 2010. Listeria monocytogenes is sensed by the NLRP3 andAIM2 inflammasome. Eur. J. Immunol. 40:1545–1551.
13. Lauer, P., M. Y. Chow, M. J. Loessner, D. A. Portnoy, and R. Calendar. 2002.Construction, characterization, and use of two Listeria monocytogenes site-specific phage integration vectors. J. Bacteriol. 184:4177–4186.
14. Lightfield, K. L., et al. 2008. Critical function for Naip5 in inflammasomeactivation by a conserved carboxy-terminal domain of flagellin. Nat. Immu-nol. 9:1171–1178.
15. Mariathasan, S., et al. 2004. Differential activation of the inflammasome bycaspase-1 adaptors ASC and Ipaf. Nature 430:213–218.
16. Martinon, F., K. Burns, and J. Tschopp. 2002. The inflammasome: a mo-lecular platform triggering activation of inflammatory caspases and process-ing of proIL-beta. Mol. Cell 10:417–426.
17. Miao, E. A., et al. 2006. Cytoplasmic flagellin activates caspase-1 and secre-tion of interleukin 1beta via Ipaf. Nat. Immunol. 7:569–575.
18. Miao, E. A., et al. 2010. Innate immune detection of the type III secretionapparatus through the NLRC4 inflammasome. Proc. Natl. Acad. Sci. U. S. A.107:3076–3080.
19. Molofsky, A. B., et al. 2006. Cytosolic recognition of flagellin by mousemacrophages restricts Legionella pneumophila infection. J. Exp. Med. 203:1093–1104.
20. Ormo, M., et al. 1996. Crystal structure of the Aequorea victoria greenfluorescent protein. Science 273:1392–1395.
21. Rathinam, V. A., et al. 2010. The AIM2 inflammasome is essential for hostdefense against cytosolic bacteria and DNA viruses. Nat. Immunol. 11:395–402.
22. Ren, T., D. S. Zamboni, C. R. Roy, W. F. Dietrich, and R. E. Vance. 2006.Flagellin-deficient Legionella mutants evade caspase-1- and Naip5-mediatedmacrophage immunity. PLoS Pathog. 2:e18.
23. Sauer, J. D., et al. 2010. Listeria monocytogenes triggers AIM2-mediatedpyroptosis upon infrequent bacteriolysis in the macrophage cytosol. CellHost Microbe 7:412–419.
24. Schroder, K., R. Zhou, and J. Tschopp. 2010. The NLRP3 inflammasome: asensor for metabolic danger? Science 327:296–300.
25. Sun, Y. H., H. G. Rolan, and R. M. Tsolis. 2007. Injection of flagellin into thehost cell cytosol by Salmonella enterica serotype typhimurium. J. Biol.Chem. 282:33897–33901.
26. Takeuchi, O., and S. Akira. 2010. Pattern recognition receptors and inflam-mation. Cell 140:805–820.
27. Vance, R. E., R. R. Isberg, and D. A. Portnoy. 2009. Patterns of pathogenesis:discrimination of pathogenic and nonpathogenic microbes by the innateimmune system. Cell Host Microbe 6:10–21.
28. Warren, S. E., et al. 2010. Cutting edge: cytosolic bacterial DNA activates theinflammasome via Aim2. J. Immunol. 185:818–821.
29. Yonekura, K., S. Maki-Yonekura, and K. Namba. 2003. Complete atomicmodel of the bacterial flagellar filament by electron cryomicroscopy. Nature424:643–650.
30. Zamboni, D. S., et al. 2006. The Birc1e cytosolic pattern-recognition recep-tor contributes to the detection and control of Legionella pneumophilainfection. Nat. Immunol. 7:318–325.
Editor: B. A. McCormick
1614 LIGHTFIELD ET AL. INFECT. IMMUN.




























