Embed Size (px)
Citation preview

Palaeodiversity 2: 67–95; Stuttgart, 30.12.2009. 67
Die Molluskenfauna der Oberen Meeresmolasse (Untermiozän) von Ermingen und Ursendorf (SW-Deutschland)
The mollusc fauna of the “Obere Meeresmolasse” (Lower Miocene) from Ermingen and Ursendorf (SW Germany)
OLAF HÖLTKE
A b s t r a c t
The localities Ermingen and Ursendorf are located in the Lower Miocene “Obere Meeresmolasse” (OMM). Ermingen is famous for the so called „Erminger Turritellenplatte“, which represents a mass occurrence of Turri-tella shells. Ursendorf is historically well known for its rich bryozoan fauna, but also for fossil molluscs and shark teeth. Both localities were best exposed at the end of the 19th century/beginning of the 20th century, when also the first taxonomic descriptions of the fauna originate, but today they are very poorly exposed. This study is a rede-scription of the mollusc fauna from these two localities.
The Ermingen fauna includes at least 12 bivalve and 10 gastropod taxa. It is totally dominated by the turritellid gastropod Turritella cf. eryna communiformis VOORTHUYSEN, 1944 and ? Tapes suevica QUENSTEDT, 1867. Formerly these species are named Turritella turris BASTEROT, 1825 and Tapes helvetica (MAYER, 1853), respectively. Subordi-nate are members of the Arcidae, Glycymerididae, Mytilidae, Pectinidae, Chamidae, Ostreidae, Carditidae, Car-diidae among the bivalves and members of the Naticidae, Fissurellidae, Xenophoridae, Epitoniidae, Olividae, and Fasciolariidae among the gastropods. The aragonitic molluscan shells are preserved as calcite pseudomorphoses.
The Ursendorf fauna includes at least 11 bivalve and 8 gastropod taxa. It is noticeable different in taxonomic composition. The fauna includes members of the Arcidae, Glycymerididae, Mytilidae, Pectinidae, Anomiidae, Limidae, Ostreidae, Carditidae, and Pholadidae among the bivalves and the gastropod families Trochidae, Epi-toniidae, Calyptraeidae, Naticidae, Cypraeidae, Ficidae, Fasciolariidae, and Conidae. No single faunal element dominates and turritellids and venerids which predominate in Ermingen are missing. The aragonitic fauna is only preserved as internal moulds.
Both localities represent shallow water environments with coarse sandy substrates. The Ermingen locality rep-resents a higher energy environment with mass occurrences of transported shells. Most of the mollusks of both lo-calities were soft substrate inhabitants. Although the sediments of Ermingen and Ursendorf represented soft sub-strate, the fauna is of allochthon orgin. Differences in the faunal composition of both localities are discussed with respect to substrate, food resources, other faunal elements (for example bryozoans and sharks) as well as taphono-my.
K e y w o r d s : Upper Marine Molasse, Lower Miocene, Ermingen, Ursendorf, molluscs, bivalves, gastropods, Turritella, Tapes.
Z u s a m m e n f a s s u n gDie Lokalitäten Ermingen und Ursendorf liegen in der Unter-Miozänen Oberen Meeresmolasse (OMM). Er-
mingen ist berühmt für die sog. „Erminger Turritellenplatte“, eine massenhafte Ansammlung von Turritella-Schalen. Ursendorf ist bekannt für eine reiche Bryozoen-Fauna, aber auch für Mollusken und Haifischzähne. Beide Fundstätten waren Ende des 19. Jahrhunderts bis Anfang des 20. Jahrhunderts sehr gut aufgeschlossen. Aus dieser Zeit stammen auch die ersten taxonomischen Arbeiten über die Faunen der beiden Fundstellen. Die Schichten sind heute nur noch in geringem Maße aufgeschlossen.
Die Fauna von Ermingen umfasst mindestens 12 Bivalven- und 10 Gastropoden-Taxa. Sie wird stark dominiert von der Gastropode Turritella cf. eryna communiformis VOORTHUYSEN, 1944 und ? Tapes suevica QUENSTEDT, 1867. Untergeordnet finden sich Arten der Bivalven-Familien Arcidae, Glycymerididae, Mytilidae, Pectinidae, Chami-dae, Ostreidae, Carditidae, Cardiidae sowie der Gastropoden-Familien Naticidae, Fissurellidae, Xenophoridae, Epitoniidae, Olividae und Fasciolariidae. Die ursprünglich aragonitischen Schalen sind in calcitischer Form er-halten geblieben.
Die Fauna von Ursendorf umfasst mindestens 11 Bivalven- und 8 Gastropoden-Taxa. Es gibt wenige Gemein-samkeiten mit der Fauna von Ermingen. Die Fauna besteht aus Arten der Bivalven-Familien Arcidae, Glycymeridi-dae, Mytilidae, Pectinidae, Anomiidae, Limidae, Ostreidae, Carditidae und Pholadidae sowie den Gastropoden-Familien Trochidae, Epitoniidae, Calyptraeidae, Naticidae, Cypraeidae, Ficidae, Fasciolariidae und Conidae. In Ursendorf ist kein Faunenelement dominierend. Die in Ermingen vorherrschenden Veneriden und Turritelliden konnten in Ursendorf nicht eindeutig nachgewiesen werden. Mollusken mit aragonitischen Schalen sind hier nur als Steinkerne erhalten.
Beide Lokalitäten stellen eine Flachwasser-Fazies dar mit grobsandigem Substrat. Ermingen war allerdings ein höher energetischer Lebensraum, daher kam es hier auch zu der Massenansammlung von transportierten Schalen.

68 PALAEODIVERSITY 2, 2009
1. Einleitung
Die beide Fossilfundstätten Ermingen und Ursendorf, in denen untermiozäne Sedimente der Oberen Meeresmo-lasse (OMM) anstehen, sind schon seit Mitte des 19. Jahr-hunderts bekannt. Trotzdem gibt es über beide Lokalitäten nur sehr wenig Literatur und auch allgemein ist über die Mollusken-Fauna der oberschwäbischen OMM bisher nur sehr wenig publiziert. Abgesehen von einigen Erwähnun-gen in diversen geologischen Führern waren MAYER (1853, 1871), PROBST (1871) und als letztes LUTZEIER (1922) die einzigen, die sich mit diesem Thema auseinandergesetzt haben. Außer bei MAYER (1871) finden sich in den anderen Publikationen ausschließlich reine Fossillisten. Die darin genannten Artnamen sind oft von unklarem nomenklato-rischem Status. So stellte sich insbesondere die Bestim-mung der für die Erminger Turritellenplatte namengeben-den Turritellen-Spezies als falsch heraus. Dies macht deut-lich, wie notwendig eine umfassende Untersuchung der Molluskenfauna der oberschwäbischen OMM ist.
In dieser Arbeit werden zum ersten Mal die Mollusken von Ermingen und Ursendorf beschrieben und abgebildet sowie ein aktueller Überblick über die Mollusken-Fauna der beiden Lokalitäten gegeben. Anschließend werden aufgrund der paläontologischen und sedimentologischen Befunde die paläoökologischen Bedingungen an beiden Lokalitäten rekonstruiert.
Diese Arbeit kann als Grundlage dienen für weitere Untersuchungen der Mollusken der oberschwäbischen OMM, um so das Bild der Diversität und Verbreitung der miozänen Mollusken zu vervollständigen.
D a n k
Herrn Prof. Dr. JAMES NEBELSICK, Universität Tübingen, danke ich für die Vergabe und die hervorragende Betreuung der Arbeit, sowie für die Übersetzung des „Abstracts“. Herrn Dr. MICHAEL W. RASSER und Herrn Dr. RONALD BÖTTCHER vom Staatlichen Museum für Naturkunde Stuttgart danke ich für die außerordentlich kompetente Betreuung und Unterstützung meiner Untersuchungen. Mein Dank gilt ebenfalls Herrn Dr. MATTHIAS HARZHAUSER, Naturhistorisches Museum Wien, für
Die meisten Mollusken der beiden Lokalitäten waren Weichbodenbewohner. Obwohl die Sedimente von Ermingen und Ursendorf Weichböden darstellten, handelt es sich bei beiden um ein allochthones Vorkommen. Die Unter-schiede in der Faunenzusammensetzung werden erörtert unter Berücksichtigung von Substrat, Ernährung, Ta-phonomie sowie dem Vorhandensein anderer Faunenelemente (z. B. Bryozoen und Haifischen).
I n h a l t1. Einleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 682. Geologischer Überblick . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 693. Lokalitäten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 694. Material und Methoden . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 715. Systematik . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71 5.1. Bivalven . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71 Familie Arcidae LAMARCK, 1809. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71 Familie Glycymerididae NEWTON, 1922 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72 Familie Mytilidae RAFINESQUE, 1815 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72 Familie Limidae RAFINESQUE, 1815 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73 Familie Ostreidae RAFINESQUE, 1815 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74 Familie Pectinidae WILKES, 1810 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75 Familie Anomiidae RAFINESQUE, 1815 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77 Familie Veneridae RAFINESQUE, 1815 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77 Familie Cardiidae LAMARCK, 1809 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 79 Familie Carditidae J. FLEMING, 1828 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81 Familie Chamidae LAMARCK, 1809 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81 Familie Corbulidae LAMARCK, 1818 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82 Familie Pholadidae LAMARCK, 1809 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82 5.2. Gastropoden . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83 Familie Fissurellidae FLEMMING, 1822 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83 Familie Trochidae RAFINESQUE, 1815 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83 Familie Turritellidae LOVÉN, 1847 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83 Familie Calyptraeidae LAMARCK, 1809 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 85 Familie Xenophoridae TROSCHEL, 1852 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86 Familie Cypraeidae RAFINESQUE, 1815 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86 Familie Naticidae FORBES, 1838 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87 Familie Ficidae MEEK, 1864 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88 Familie Epitoniidae BERRY, 1910 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88 Familie Fasciolariidae J. E. GRAY, 1853 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89 Familie Olividae LATREILLE, 1825 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 89 Familie Conidae RAFINESQUE, 1815 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 906. Paläoökologie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 907. Literaturverzeichnis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 93

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 69
die äußerst wertvolle Hilfe bei der Bestimmung der Objekte und die Begutachtung des Manuskripts. Herrn Dipl. Geol. OLIVER VONDERSCHMIDT und Herrn ACHIM LEHMKUHL danke ich für fruchtbare Diskussionen und Anregungen. Den Mitgliedern des Vereins der Mineralien- und Fossilienfreunde Ulm/Neu-Ulm danke ich für die wertvolle Unterstützung und Kooperation bei der Geländearbeit und der Untersuchung des Fossilmaterials. Herrn WOLFGANG GERBER, Universität Tübingen, danke ich sehr für die Anfertigung der Fotos. Meinen Eltern danke ich für ihre jahrelange Unterstützung meiner Studien, welche ohne sie nicht möglich gewesen wären.
2. Geologischer Überblick
Ermingen und Ursendorf liegt im nördlichen Alpen-vorland-Becken, dem so genannten Molasse-Becken. Die Entstehung des Beckens begann während des Ober-Eozäns (BIEG et al. 2007) und wurde ausgelöst durch die alpine Gebirgsbildung. Auf eine Flysch-Fazies folgte beginnend im frühen Oligozän die so genannte „Molasse-Phase“, welche ein flacheres Ablagerungsmilieu darstellte (BIEG et al. 2007). Sie wird in vier Einheiten unterteilt: Untere Meeresmolasse (UMM), Untere Süßwassermolasse (USM), Obere Meeresmolasse (OMM), Obere Süßwas-sermolase (OSM). Regional ist zwischen OMM und OSM noch die Brackwassermolasse zwischengeschaltet. Für die Molasse wird die Stufengliederung der Paratethys ange-wendet.
Im Rupelium kam es zu einem ersten Meereseinbruch. Die UMM tritt nach GEYER et al. (2003) nur in der Nähe des Alpenrandes, im Bodenseegebiet sowie im Hegau zutage. Während der USM, welche das Oberchattium und das Aquitanium umfasst, herrschten limnisch-fluviatile Ablagerungsbedingungen (GEYER & GWINNER 1991). Zu Beginn der OMM vor ca. 20 Millionen Jahren konnte das Meer von Südwesten aus dem Rhône-Becken in das nord-alpine Becken vordringen. Dieser so genannte „Burdi-galian Seaway“ verband die Tethys mit der Paratethys (KUHLEMANN & KEMPF 2002). Die OMM wird in zwei Sedimentationszyklen gegliedert (SCHREINER 1966). Laut GALL (1975) soll auch noch ein dritter Zyklus existiert ha-ben. Eine ausführliche Untersuchung zur Stratigraphie der OMM findet sich bei SCHREINER (1966), zur Paläogeogra-phie während der OMM siehe BIEG (2005). Zu welchem Zyklus die in dieser Arbeit untersuchten Schichten ge-hören, ist nicht eindeutig geklärt. Der erste Zyklus beginnt laut WENGER (1987) im unteren Ottnangium mit den Hei-denlöcher Schichten, welche bei Überlingen anstehen. Den Abschluss der OMM bilden die so genannten „Deck-schichten“, welche aus feinkörnigen Sanden bestehen (SCHREINER & LUTERBACHER 1999). SCHREINER (1966) teilt die OMM in fünf Fazieszonen ein. An der nordwestlichen Klifflinie des Molassemeeres, welche von Donauwörth in südwestlicher Richtung über Heldenfingen, Magolsheim, Stetten am kalten Markt bis nach Geisingen verläuft
(ROSENDAHL 2000), finden sich angebohrte Brandungs-schorren (BEURLEN et al. 1981). Die Klifflinie entspricht der Brandungsküste der größten Ausdehnung der OMM während des Ottnangiums (SCHREINER & LUTERBACHER 1999). Die Hauptmasse der fein- bis mittelkörnigen Sand-massen des Beckeninnern wurden aus den Alpen, besonders durch den Napfschuttfächer herbeigeführt und in der Beckenachse nach Osten transportiert (HOFMANN 1957). Im Ottnang, nach der Ablagerung der OMM, führte ein Meeresspiegelabfall zum Einschneiden des Tales der so genannten „Graupensandrinne“, die sich im Laufe des Ottnangiums mit den ästuarinen Ablagerungen der Grim-melfinger und Kirchberger Schichten füllte (SCHREINER & LUTERBACHER 1999). Diese als „Brackwasser-Molasse“ bezeichneten Schichten gehen über in die OSM, in der erneut limnisch-fluviatlie Ablagerungsbedingungen herrschten wie in der USM (GEYER & GWINNER 1991).
Die Lokalitäten Ermingen und Ursendorf sind auf-grund des reichlich vorhandenen Fossilmaterials die wich-tigsten Fundpunkte von Mollusken in der untermiozänen Oberen Meeresmolasse in Oberschwaben. Die „Erminger Turritellenplatte“ besteht zum einen aus den harten Turri-tellenkalken, zum anderen aus sandigen Kalken (Abb. 2), Sanden und Sandsteinen, welche ebenfalls reich an Turri-tellen sind (RASSER & NEBELSICK 2006). Aus diesen Schich-ten stammen auch die übrigen in Kapitel 5 beschriebenen Molluskenarten sowie Haizähne und Gerölle. Die ur-sprünglich aragonitischen Schalen der Mollusken sind zu Calcit umkristallisiert. Zwischengeschaltet finden sich auch fossilfreie Sande und Tone bis Silte (RASSER & NEBEL-SICK 2006). Das Liegende der „Erminger Turritellenplatte“ besteht aus Schichten von weißem bis hellgrauem und bläulichem Lehm (RASSER & NEBELSICK 2006).
In der Sandgrube von Ursendorf ist eine ca. 10 m hohe Wand der OMM aufgeschlossen, an deren Basis sich gut sortierte Grobsande mit den typischen alpinen Geröllen befinden. Darauf folgen weitere Grobsandschichten, welche eine großförmige Schrägschichtung aufweisen. Abgeschlossen wird die Schichtenfolge von feinkörnigen, leicht siltigen Sanden. Ein aktuelles Profil findet sich bei BIEG et al. (2007). Die Schichten, aus welchen die in jün-gerer Zeit (1970er Jahre) gesammelten Mollusken stam-men, sind nicht mehr aufgeschlossen und befinden sich im Liegenden dieser Wand. Wie man allerdings an den Ob-jekten erkennen kann, ist das Gestein von gleicher Grob-körnigkeit wie im heute aufgeschlossenen Teil, aber mit einem mehr oder weniger hohen Anteil an etwas feinerem Material.
3. Lokalitäten
Die Ortschaft Ermingen liegt einige Kilometer westlich von Ulm (Abb.1), die „Erminger Turritellenplatte“ nord-

70 PALAEODIVERSITY 2, 2009
westlich des Ortes (GK 25: 7625 Ulm-Südwest, R: 3566190; H: 5361200). Die kalkigen Sandsteinplatten, die massen-haft Turritellen enthalten (Abb. 2), wurden im 19. Jahrhun-dert in kleinen Steinbrüchen abgebaut und in der Umge-bung als Baustein genutzt. Noch heute sieht man im Waldgebiet der „Erminger Turritellenplatte“ Spuren des ehemaligen Abbaus. Die Lokalität wurde schon von PROBST (1871), ENGEL (1908, 1911), FRAAS (1910) und sehr aus-führlich von LUTZEIER (1922) behandelt. Danach geriet die Fundstätte ein wenig in Vergessenheit, bis 1980 das Ge- biet der „Erminger Turritellenplatte“ zum größten Teil eingezäunt und zum geologischen Naturdenkmal erklärt wurde. 1999 wurde anlässlich des 700-jährigen Bestehens der Gemeinde Ermingen von den zuständigen Behörden zusammen mit dem örtlichen Mineralien- und Fossilien-sammler-Verein ein geologischer Lehrpfad mit Schautafeln errichtet, welche über die Fundlokalität informieren. Erst 2004 erschien dann von BAIER et al. wieder eine spezielle
Veröffentlichung über Ermingen, in der die Hai- und Rochenfauna abgebildet und diskutiert wird. Zudem wurde von GEYER (2001) eine Broschüre über die Geologie im Raum Ulm veröffentlicht. Vom 12.–22.9.2005 wurden vom Staatlichen Museum für Naturkunde Stuttgart (SMNS) in Zusammenarbeit mit der Universität Tübingen einige Me-ter westlich des eingezäunten Bereichs Grabungen durch-geführt. Dabei wurde ein ca. 3,5 m mächtiges Profil der „Turritellenplatte“ freigelegt, aufgenommen und beprobt. Nach Abschluss der Grabung musste dieser Aufschluss wieder verfüllt werden. Die Resultate aus dieser Grabung wurden von RASSER & NEBELSICK (2006) publiziert.
Die Ortschaft Ursendorf liegt ca. 3 km südlich von Mengen im Kreis Sigmaringen (Abb. 1). Wie ENGEL (1908) zu entnehmen ist, existierten Ende des 19. Jahrhunderts/Anfang des 20. Jahrhunderts eine Reihe von Sandgruben, welche aber bis auf eine nicht mehr vorhanden sind. Somit lässt sich bei vielen Exemplaren im SMNS nicht sagen,
Abb. 1. Geographische Lage von Ermingen und Ursendorf in Süddeutschland.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 71
woher genau aus Ursendorf diese stammen. Heute gibt es nur noch eine Sandgrube der Firma STEIDLE am westlichen Ortsende von Ursendorf (GK 25: 7922 Saulgau, ca. bei R: 3527320, H: 5318750). Der Sand, der hier abgebaut wird, wird für Tennisplätze etc. verwendet. Aus dieser Sandgrube stammen auch die Mollusken im SMNS, welche in jün-gerer Zeit (1970er Jahre) gesammelt wurden. Allerdings sind die entsprechenden Schichten dort nicht mehr aufge-schlossen. In den derzeitig aufgeschlossenen Schichten finden sich hauptsächlich Haizähne und Bruchstücke von Molluskenschalen. Die Fauna von Ursendorf wurde schon von MILLER (1875) und ENGEL (1908) erwähnt. MILLER be-schrieb 1875 die reichhaltige Bryozoen-Fauna von Ursen-dorf und benannte auch eine Seeigelart nach der Ortschaft (Fibularia ursendorfensis MILLER). Seine Bryozoen-Samm lung ist im Staatlichen Museum für Naturkunde Stuttgart untergebracht. SCHÜTZE (1904) beschrieb die Schwäm me und Echinodermen von Ursendorf.
4. Material und Methoden
In dieser Arbeit werden die Bivalven- und Gas-tropoden-Arten der beiden Lokalitäten systematisch neu erfasst, untersucht und abgebildet. Aufgrund des Erhal-tungszustandes einzelner Exemplare ist eine eindeutige Bestimmung aber nicht immer möglich gewesen. Alle Originale befinden sich in der Sammlung des Staatlichen Museums für Naturkunde Stuttgart (SMNS), lediglich das Original zu Abb. 2 stammt aus dem Museum des Instituts für Geologie und Paläontologie der Universität Tübingen. Leider war das Originalmaterial von LUTZEIER (1922), das in Tübingen hinterlegt worden ist, nicht mehr auffindbar, so dass dessen Fossilliste nicht überprüft werden konnte.
Die Taxonomie der Mollusken richtet sich nach LIND-NER (1999) und POPPE & GOTO (1991, 1993). Auf die Nen-nung von Untergattungen wird verzichtet. Die Merkmale der Bivalven- und Gastropodengehäuse sind nach LINDNER (1999) benannt, die der Familie Cypraeidae nach LORENZ & HUBERT (1993). Die stratigraphischen Einheiten sind MENNIG & HENDRICH (2005) entnommen.
5. Systematik5.1. Bivalven
Familie Arcidae LAMARCK, 1809Unterfamilie Anadarinae REINHART, 1935
Gattung Anadara J. E. GRAY, 1847
B e m e r k u n g e n . – In der älteren Literatur, wie zum Beispiel bei HÖLZL (1958), wird die Gattung Anadara oft-mals nur als Untergattung von Arca angesehen.
H a b i t a t u n d L e b e n s w e i s e . – Anadara lebt als Filtrierer grabend in Sand oder Schlamm im Gezeiten-bereich und flacheren Sublitoral der wärmeren und tro-pischen Gewässer (LINDNER 1999).
Anadara fichteli (DESHAYES, 1852) Abb. 3
2001 Anadara fichteli (DESHAYES, 1852). – SCHULTZ, S. 54–56, Taf. 1, Fig. 2. − [hier Synonymie-Liste]
M a t e r i a l : 6 Exemplare (Schalenerhaltung).
B e s c h r e i b u n g . – Die Exemplare sind bis zu 50 mm lang und bis zu 35 mm hoch. Die Schalen sind un-gleichseitig und zum Teil sehr massiv. Auf der Oberfläche befinden sich zwischen 25 und 28 stark ausgeprägte Rip-pen.
D i s k u s s i o n . – Die Spezies ist sehr variabel in der Umrissform, daher wurden einige Unterarten abgespalten und es existieren unzählige Synonyme. Eine ähnliche Art
Abb. 2. „Erminger Turritellenplatte“; Ermingen; GPIT, ohne Nummer. – Maßstab 30 mm.

72 palaeodiversity 2, 2009
ist Anadara diluvii (lamarck, 1805), welche sich aber durch weniger kräftige Rippen von A. fichteli unterscheidet.
V o r k o m m e n : Eggenburgium bis ?Pliozän (schultz 2001).
V e r b r e i t u n g : Ermingen; sonstige Vorkommen außerhalb der oberschwäbischen OMM siehe schultz (2001).
Von lutzeier (1922) zusätzlich angeführte AnadaraArt für Ermingen: Anadara turonica (Dujardin, 1837).
Familie Glycymerididae newton, 1922
Gattung Glycymeris da costa, 1778
H a b i t a t u n d L e b e n s w e i s e . – Glycymeris lebt hauptsächlich in warmen und tropischen Gewässern. Sie ist allerdings auch mit einer Art im Ärmelkanal und in der Nordsee vertreten (entrop 1977; lindner 2000). Gly-cymeris bewohnt das Sublitoral und lebt oberflächennah eingegraben in Schlamm und Sandböden (poppe & Goto 1993; lindner 1999).
Glycymeris cf. fichteli (deshayes, 1852) Abb. 4
1994 Glycymeris fichteli (deshayes, 1852). – pfister & weG-müller, s. 912–914, Taf. 1, Fig. 9, 11. – [hier SynonymieListe]
M a t e r i a l : 3 Exemplare (Steinkerne).
B e s c h r e i b u n g . – Die Steinkerne haben einen Durchmesser von bis zu 60 mm und einen runden Umriss. Die Klappen sind stark gewölbt. Die Wirbelregion ist auffallend stark verlängert und hat ein spitzes Ende.
D i s k u s s i o n . – pfister & weGmüller (1994) geben für G. fichteli einen stark verlängerten und zugespitzten Wirbel an, allerdings bei Schalenerhaltung, sowie eine Durchschnittshöhe von ca. 85 mm. Da an den Exemplaren im SMNS sonst keine weitern bestimmungsrelevanten Merkmale zu sehen sind, ist eine sichere Artbestimmung nicht möglich.
V o r k o m m e n : Mittleres Burdigalium (pfister & weG-müller 1994), Eggenburgium (steininGer et al. 1971).
V e r b r e i t u n g : Ursendorf; sonstige Vorkommen außerhalb der oberschwäbischen OMM siehe bei pfister & weGmül-ler (1994).
Glycymeris sp. Abb. 5
M a t e r i a l : 1 Exemplar (linke? Klappe, Steinkern).
B e s c h r e i b u n g . – Die Schale hat einen Durchmesser von 33 mm, einen fast kreisrunden Umriss und ist mäßig gewölbt. Auf der Schalenoberfläche sind konzentrische und radiale Rippen vorhanden. Die Unterseite ist nicht mehr zu erkennen, weshalb eine genaue Ermittlung der Spezies nicht möglich ist.
V e r b r e i t u n g : Ursendorf.
Von lutzeier (1922) erwähnte GlycymerisArt für Ermingen: Glycymeris pilosus (linnaeus).
Familie Mytilidae rafinesque, 1815 Unterfamilie Mytilinae rafinesque, 1815
Gattung Perna philippson, 1788
H a b i t a t u n d L e b e n s w e i s e . – Perna kommt mit einigen Arten in gemäßigten und tropischen Gewässern vor (kilias 1997). Sie lebt wie alle anderen Vertreter der Mytilinae mit Byssus an Hartsubstanzen befestigt.
Perna aquitanica mayer, 1858 Abb. 6
2001 Mytilus (Mytilus) aquitanicus mayer, 1858. – schultz, s. 94–96, Taf. 7, Fig. 8. – [hier SynonymieListe]
Abb. 3. Anadara fichteli (deshayes, 1852), rechte Klappe; Ermingen; SMNS 68601. – a. Innenansicht. b. Außenansicht. – Maßstab 10 mm.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 73
2001 Perna aquitanica (MAYER, 1858). – WESSELINGH et al., S. 121–128, Fig. 2. – [hier Synonymie-Liste]
M a t e r i a l : 1 Exemplar (rechte Klappe, Schalenerhal-tung).
B e s c h r e i b u n g . – Die Schale ist 74 mm lang und hat die typische „Miesmuschel-Form“. Der Wirbel liegt fast ganz am Vorderende und ist leicht nach unten gebo-gen. Die Ventralseite ist schwach konkav. Die Dorsalseite ist leicht gewinkelt. Schließmuskeleindrücke sowie das Schloss sind bei dem vorhandenen Exemplar nicht zu erkennen.
D i s k u s s i o n . – Die Spezies wurde früher als zur Gattung Mytilus gehörig betrachtet. Diskussion dazu siehe WESSELINGH et al. (2001). P. aquitanica ist die einzige Art der Mytilinae, die von PROBST (1871) und LUTZEIER (1922) für die oberschwäbische OMM genannt wird.
V o r k o m m e n : Mittleres Kiscellium bis eventuell Pliozän (SCHULTZ 2001).
V e r b r e i t u n g : Ermingen, Baltringen und nähere Umge-bung (PROBST 1871); sonstige Vorkommen außerhalb der ober-schwäbischen OMM siehe SCHULTZ (2001).
Familie Limidae RAFINESQUE, 1815 Gattung Lima BRUGUIÈRE, 1797
Lima lima (LINNAEUS, 1758) Abb. 7
2001 Lima (Lima) lima (LINNÉ, 1758). – SCHULTZ, S. 296–298, Taf. 47, Fig. 1. − [hier Synonymie-Liste]
M a t e r i a l : 3 Exemplare (Steinkerne).
B e s c h r e i b u n g . – Die Schalen sind bis zu 55 mm hoch, stark ungleichseitig mit schief ovalem Umriss und leicht gewölbt. Auf jeder Klappe befinden sich ca. 20 Ra-dialrippen. Die Rippen sind abgeplattet, mit leicht gerun-detem Querschnitt. Die Abstände zwischen den Rippen entsprechen der Rippenbreite. Die vorliegenden Exem-plare sind nicht ganz vollständig erhalten, so dass die Ohren am Schlossrand nicht mehr genau zu erkennen sind.
D i s k u s s i o n . – POPPE & GOTO (1991) geben für diese Spezies eine Rippenzahl zwischen 18 und 24 an.
H a b i t a t u n d L e b e n s w e i s e . – L. lima lebt zwischen 3 und 100 m Tiefe mit Byssus zwischen Steinen (POPPE & GOTO 1993) oder oft auch an Schwämmen (RIEDL 1963) festgeheftet. Sie ist in der Mediterran-Lusitanischen Provinz sowie in der Karibischen Region beheimatet (LINDNER 1999).
V o r k o m m e n : Seit dem unteren Ottnangium (SCHULTZ 2001).
Abb. 4–5. Glycymeridae. – 4. Glycymeris cf. fichteli (DESHAYES, 1852), rechte Klappe; Ursendorf; SMNS 68600. a. Außenansicht. b. Lateralansicht. – Maßstab 10 mm. 5. Glycymeris sp., linke? Klappe, Außenansicht; Ursendorf; SMNS 68602. – Maßstab 10 mm.
Abb. 6. Perna aquitanica MAYER, 1858, rechte Klappe, Außen-ansicht; Ermingen; SMNS 68603. – Maßstab 10 mm.
74 PALAEODIVERSITY 2, 2009
V e r b r e i t u n g : Ursendorf; sonstige Vorkommen außer-halb der OMM siehe SCHULTZ (2001).
Familie Ostreidae RAFINESQUE, 1815 Unterfamilie Ostreinae RAFINESQUE, 1815
Gattung Ostrea LINNAEUS, 1758
H a b i t a t u n d L e b e n s w e i s e . – Ostrea kommt weltweit vor, allerdings nicht in der Polarregion oder in der Ostsee (FECHTER et al. 1985; MÜLLER 1993). Alle Ver-treter dieser Gattung vertragen kein Brackwasser (KILIAS 1997). Ostrea lebt mit einer Klappe festgekittet auf Felsen, Schillbänken oder sonstigem Hartsubstrat. Sie kommt von der Brandungszone bis in größere Tiefen vor (FECHTER et al. 1985; KILIAS 1997).
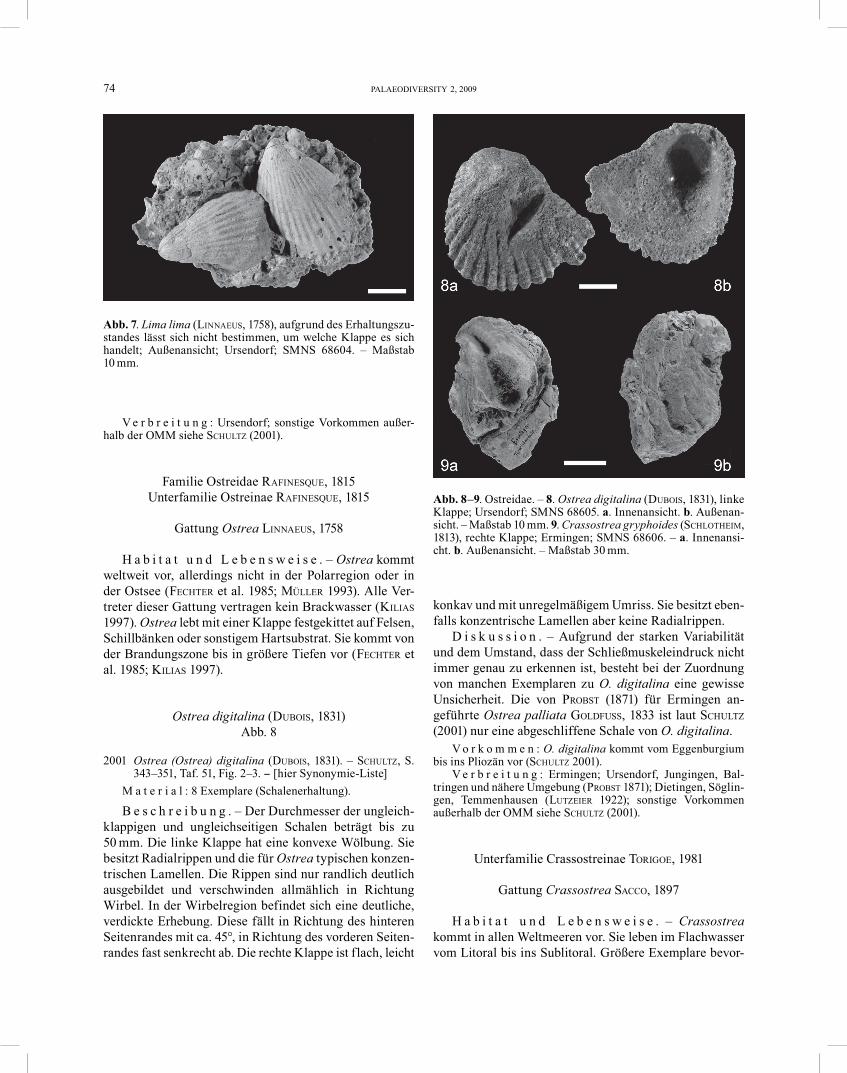
Ostrea digitalina (DUBOIS, 1831) Abb. 8
2001 Ostrea (Ostrea) digitalina (DUBOIS, 1831). – SCHULTZ, S. 343–351, Taf. 51, Fig. 2–3. − [hier Synonymie-Liste]
M a t e r i a l : 8 Exemplare (Schalenerhaltung).
B e s c h r e i b u n g . – Der Durchmesser der ungleich-klappigen und ungleichseitigen Schalen beträgt bis zu 50 mm. Die linke Klappe hat eine konvexe Wölbung. Sie besitzt Radialrippen und die für Ostrea typischen konzen-trischen Lamellen. Die Rippen sind nur randlich deutlich ausgebildet und verschwinden allmählich in Richtung Wirbel. In der Wirbelregion befindet sich eine deutliche, verdickte Erhebung. Diese fällt in Richtung des hinteren Seitenrandes mit ca. 45°, in Richtung des vorderen Seiten-randes fast senkrecht ab. Die rechte Klappe ist flach, leicht
konkav und mit unregelmäßigem Umriss. Sie besitzt eben-falls konzentrische Lamellen aber keine Radialrippen.
D i s k u s s i o n . – Aufgrund der starken Variabilität und dem Umstand, dass der Schließmuskeleindruck nicht immer genau zu erkennen ist, besteht bei der Zuordnung von manchen Exemplaren zu O. digitalina eine gewisse Unsicherheit. Die von PROBST (1871) für Ermingen an-geführte Ostrea palliata GOLDFUSS, 1833 ist laut SCHULTZ (2001) nur eine abgeschliffene Schale von O. digitalina.
V o r k o m m e n : O. digitalina kommt vom Eggenburgium bis ins Pliozän vor (SCHULTZ 2001).
V e r b r e i t u n g : Ermingen; Ursendorf, Jungingen, Bal-tringen und nähere Umgebung (PROBST 1871); Dietingen, Söglin-gen, Temmenhausen (LUTZEIER 1922); sonstige Vorkommen außerhalb der OMM siehe SCHULTZ (2001).
Unterfamilie Crassostreinae TORIGOE, 1981
Gattung Crassostrea SACCO, 1897
H a b i t a t u n d L e b e n s w e i s e . – Crassostrea kommt in allen Weltmeeren vor. Sie leben im Flachwasser vom Litoral bis ins Sublitoral. Größere Exemplare bevor-
Abb. 7. Lima lima (LINNAEUS, 1758), aufgrund des Erhaltungszu-standes lässt sich nicht bestimmen, um welche Klappe es sich handelt; Außenansicht; Ursendorf; SMNS 68604. – Maßstab 10 mm.
Abb. 8–9. Ostreidae. – 8. Ostrea digitalina (DUBOIS, 1831), linke Klappe; Ursendorf; SMNS 68605. a. Innenansicht. b. Außenan-sicht. – Maßstab 10 mm. 9. Crassostrea gryphoides (SCHLOTHEIM, 1813), rechte Klappe; Ermingen; SMNS 68606. – a. Innenansi-cht. b. Außenansicht. – Maßstab 30 mm.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 75
zugen Tiefen zwischen 20 und 40 m (POPPE & GOTO 1993). Sie leben festgekittet auf Hartboden (LINDNER 2000).
Crassostrea gryphoides (SCHLOTHEIM, 1813) Abb. 9
2001 Crassostrea gryphoides (SCHLOTHEIM, 1813). – SCHULTZ, S. 326–339, Taf. 49, Fig. 1. – [hier Synonymie-Liste]
M a t e r i a l : >20 Exemplare (Schalenerhaltung).
B e s c h r e i b u n g . – Die aus konzentrischen Lamel-len aufgebaute dicke Schale weist in ihrer Form und Masse eine große Variabilität auf. Die linke Klappe ist stark gewölbt, während die rechte eher flach ist, aber zum Teil leicht konkave oder konvexe Partien aufweist. Der Wirbel ist verlängert und leicht nach vorne gekrümmt. Die Bandgrube, die bei einzelnen Exemplaren unterschiedlich lang sein kann, wird zu beiden Seiten von je einem Wulst begrenzt. In der Grube sind manchmal Anwachsstreifen zu erkennen. Der Schließmuskeleindruck ist mond- bis leicht nierenförmig. Die Enden sind immer gerundet. Es liegen aus Ermingen nur unvollständig erhaltene Exem-plare vor. Aus einem Vergleich mit bis zu 20 cm hohen vollständigen Individuen von Heldenfingen (SMNS) lässt sich erkennen, dass die Erminger Exemplare eine ähnliche Größendimension erreicht hatten.
D i s k u s s i o n . – C. gryphoides war nach HÖLZL (1958) ein ausgesprochener Litoralbewohner. An Stellen, wo die Art Bänke bildet, sind auch doppelklappige Exem-plare zu finden (PFISTER & WEGMÜLLER 1998). Diese sind z. B. in größerer Anzahl aus der Umgebung des Helden-finger Kliffs im SMNS vorhanden.
V o r k o m m e n : Die Art kommt vom Eggenburgium bis ins Pliozän vor (SCHULTZ 2001).
V e r b r e i t u n g : Ermingen; Oberer Eselsberg bei Ulm; Heldenfingen; Rammingen (GEYER & GWINNER 1979); Altheim, Bissingen, Hülbe bei Dettingen, Söglingen, Weidenstetten (LUTZEIER 1922); sonstige Vorkommen außerhalb der OMM sie-he SCHULTZ (2001).
Familie Pectinidae WILKES, 1810 Unterfamilie Chlamydinae VON TEPPNER, 1922
Gattung Aequipecten FISCHER, 1886
H a b i t a t u n d L e b e n s w e i s e . – ABBOT & DANCE (1998) bilden einige Arten von Aequipecten ab, welche vom Südosten der USA bis südlich nach Uruguay vorkom-men. Die einzige Art in Europa ist Aequipecten opercu-laris (LINNAEUS, 1758). Diese kommt nach WAGNER (1990) von Island bis zu den Kapverdischen Inseln sowie in der Nordsee und im Mittelmeer vor. Sie lebt frei beweglich auf Sand- oder Geröllboden.
Aequipecten macrotis (SOWERBY, 1847) Abb. 10
2001 Aequipecten macrotis macrotis (SOWERBY, 1847). – SCHULTZ, S. 192–194, Taf. 18, Fig. 2. – [hier Synonymie-Liste].
M a t e r i a l : 8 Exemplare (Schalenerhaltung).
B e s c h r e i b u n g . – Die Schalen sind bis zu 25 mm hoch, ungleichseitig und mehr oder weniger gleich lang wie hoch. Das vordere Ohr ist länger als das hintere. Auf der Schalenoberfläche verlaufen ca. 20 flach gerundete radiale Rippen. Der Abstand zwischen den Rippen ist un-gefähr gleich groß wie die Rippenbreite. Der Schlossrand ist gerade. Der Seitenrand des hinteren Ohrs ist distal ab-gestutzt und verläuft mit fast 90° nach unten.
D i s k u s s i o n . – SCHULTZ (2001) teilt Aequipecten macrotis in zwei Unterarten auf: A. macrotis macrotis (SOWERBY, 1847) und A. macrotis miotransversus (SCHAF-FER, 1910). Zu welcher Unterart die untersuchten Exem-plare gehören ist nicht mit Sicherheit festzustellen.
V o r k o m m e n : Eggenburgium bis Badenium (SCHULTZ 2001).
V e r b r e i t u n g : Ermingen; Ursendorf; sonstige Vorkom-men außerhalb der oberschwäbischen OMM siehe SCHULTZ (2001).
Gattung Crassadoma BERNARD, 1986
Crassadoma ? multistriata (POLI, 1795) Abb. 11
2001 Crassadoma ? multistriata (POLI, 1795). – SCHULTZ, S. 176–184, Taf. 16, Fig. 6, 9–12; Taf. 17, Fig. 1–4. – [hier Synonymie-Liste]
M a t e r i a l : 5 Exemplare (Schalenerhaltung).
B e s c h r e i b u n g . – Die Schalen haben eine durch-schnittliche Höhe von ca. 35 mm. Sie sind ungleichseitig und haben einen länglichen, ovalen Umriss. Das vordere Ohr ist verlängert. Beide Ohren sind schräg abgestutzt. Die Schale hat auf der Oberfläche ca. 22 Radialrippen mit darauf sitzenden Schuppen. Dazwischen sind noch kleine, schwache Radialrippen eingeschaltet. Der Übergang vom Seitenrand zum Ventralrand bildet auf beiden Seiten einen schwach ausgeprägten stumpfen Winkel. Das verlängerte vordere Ohr trägt ca. 10 Radialrippen.
D i s k u s s i o n . – Nach WALLER (zitiert in SCHULTZ 2001) wird die Spezies multistriata in der neueren Litera-tur der Gattung Crassadoma zugeordnet. Die Spezies wird nach POPPE & GOTO (1993) rezent bis zu 45 mm hoch.
H a b i t a t u n d L e b e n s w e i s e . – Crassadoma multistriata kommt von den Britischen Inseln bis nach Westafrika und im Mittelmeer vor und lebt mit Byssus befestigt auf Kiesböden und Grobsand zwischen 10 und 180 m Tiefe (POPPE & GOTO 1993).

76 PALAEODIVERSITY 2, 2009
V o r k o m m e n : Ab dem oberen Kiscellium (SCHULTZ 2001).
V e r b r e i t u n g : Ursendorf; Jungingen (PROBST 1871); sonstige Vorkommen außerhalb der OMM siehe SCHULTZ (2001).
Gattung Amussiopecten SACCO, 1897
B e m e r k u n g . – Amussiopecten wird häufig nur als Untergattung von Pecten angesehen. Lebensweise, Habi-tat und Verbreitung ähnlich Crassadoma.
Amussiopecten burdigalensis (LAMARCK, 1806) Abb. 12
1971 Flabellipecten burdigalensis (LAMARCK, 1806). – STEI-NINGER et al., S. 420–421, Taf. 20, Fig. 1.
2001 Amussiopecten (Amussiopecten) burdigalensis (LAMARCK, 1806). – SCHULTZ, S. 223–225. − [hier Synonymie-Liste]
M a t e r i a l : 1 Exemplar (linke Klappe, Schalenerhal-tung).
B e s c h r e i b u n g . – Die Schale hat eine Höhe von 60 mm und ist 62 mm lang. Auf der Schalenoberfläche finden sich ca. 14 bis 16 flache Rippen. Der Raum zwi-schen den Rippen entspricht ungefähr der anderthalb-fachen Rippenbreite. Die Rippen sind in der Wirbelregion nur noch schwach zu erkennen. Die Wirbelregion ist zu-dem leicht konkav. Vorderer und hinterer Dorsalrand sind leicht verdickt. Die beiden Ohren sind leicht nach innen gewölbt. Auf den Ohren sind schwache konzentrische Streifen vorhanden.
V o r k o m m e n : Egerium bis ?Badenium (SCHULTZ 2001).V e r b r e i t u n g : Ermingen; sonstige Vorkommen außer-
halb der oberschwäbischen OMM siehe SCHULTZ (2001).
Abb. 10–13. Pectinidae. – 10. Aequipecten macrotis (SOWERBY, 1847), linke Klappe; Ursendorf; SMNS 68607. a. Außenansicht. b. In-nenansicht. 11. Crassodama multistriata (POLI, 1795), linke Klappe; Ursendorf; SMNS 68608. a. Außenansicht. b. Innenansicht. 12. Amussiopecten burdigalensis (LAMARCK, 1806), linke Klappe; Ermingen; SMNS 68609. a. Außenansicht. b. Innenansicht. 13. Pecten sp., linke Klappe, Außenansicht; Ursendorf; SMNS 68610. – Maßstab 10 mm.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 77
Unterfamilie Pectininae WILKES, 1810
Gattung Pecten O. F. MÜLLER, 1776
H a b i t a t u n d L e b e n s w e i s e . – Pecten kommt in allen Weltmeeren vor. Sie leben frei beweglich auf Sand und Geröll.
Pecten sp. Abb. 13
M a t e r i a l : 1 Exemplar (Schalenerhaltung).
B e s c h r e i b u n g . – Die Schale ist 40 mm hoch und 40 mm lang. Die Umrissform ist leicht oval. Die Schale ist kaum gewölbt. Die Ohren sind unvollständig erhalten, las-sen aber erahnen, dass das vordere Ohr ein wenig größer ist. Beide Seitenränder gehen in einem stumpfen Winkel in den Ventralrand über. Auf der Schalenoberfläche befin-den sich etwa 10 Radialrippen, welche am Ventralrand bis zu 5 mm breit sind. Die Abstände zwischen den Rippen betragen ca. 2,5 mm. Die Rippen sind stark abgeplattet und haben einen leicht eckigen Querschnitt. Die Innen-seite der Schale ist mit einer grauen Substanz bestrichen wurde, daher sind keine Merkmale dieser Seite zu erken-nen.
D i s k u s s i o n . – Diese Schale lässt sich keiner Art eindeutig zuordnen.
V e r b r e i t u n g : Ursendorf.
Von LUTZEIER (1922) zusätzlich angeführte Arten der Familie Pectinidae für Ermingen: Aequipecten opercu-laris (LINNAEUS, 1758), Flabellipecten herrmannseni (DUNKER, 1848), Pecten ventilabrum GOLDFUSS, 1833.
Familie Anomiidae RAFINESQUE, 1815 Unterfamilie Anomiinae RAFINESQUE, 1815
Gattung Anomia LINNAEUS, 1758
Anomia ephippium (LINNAEUS, 1758) Abb. 14
2001 Anomia (Anomia) ephippium LINNÉ, 1758. – SCHULTZ, S. 282–288, Taf. 45, Fig. 1. – [hier Synonymie-Liste]
M a t e r i a l : 10 Exemplare (linke Klappen, Schalenerhal-tung).
B e s c h r e i b u n g . – Es sind nur linke Klappen vorhanden. Diese sind bis zu 60 mm breit, stark gewölbt mit einem unregelmäßigen Umriss. Die dünne Schale ist gewellt.
D i s k u s s i o n . – Die rechte Klappe von Anomia ist flach und zeigt eine tiefe Bucht für den Byssus, der aber vom Ventralrand meist derart umwachsen ist, dass ein
Loch entsteht (MÜLLER 1993). Sie ist sehr dünn und daher meist nicht erhalten. Die Umrissform ist dem jeweiligen Untergrund angepasst, auf welchem das Tier sich angesie-delt hat (LINDNER 2000).
H a b i t a t u n d L e b e n s w e i s e . – Diese Spezies kommt in der Mediterran-Lusitanischen Provinz sowie auch bei Island und den Britischen Inseln vor. Sie lebt auf Hartgrund von der Gezeitenzone bis in 150 m Tiefe (POPPE & GOTO 1993).
V o r k o m m e n : Seit dem Eggenburgium (SCHULTZ 2001).V e r b r e i t u n g : Ursendorf, Rammingen; sonstige
Vorkommen außerhalb der oberschwäbischen OMM siehe SCHULTZ (2001).
Familie Veneridae RAFINESQUE, 1815
H a b i t a t u n d L e b e n s w e i s e . – Alle Vertreter der Veneridae leben im Sediment und sind Filtrierer. Die Familie kommt in allen warmen und tropischen Gewässern vor (KILIAS 1997).
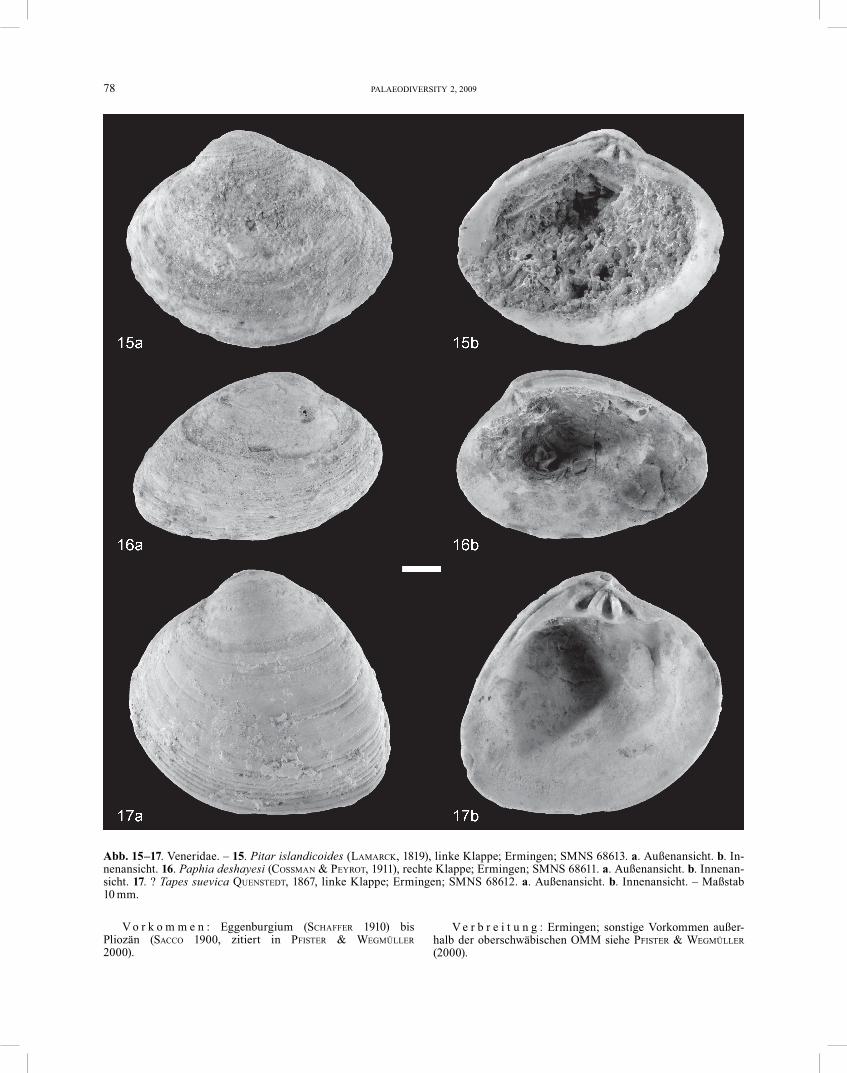
Pitar islandicoides (LAMARCK, 1819) Abb. 15
2000 Pitar islandicoides (LAMARCK, 1819). – PFISTER & WEG-MÜLLER, S. 453–455, Taf. 6, Fig. 1–9. – [hier Synonymie-Liste]
M a t e r i a l : 1 Exemplar (rechte Klappe, Schalenerhal-tung).
B e s c h r e i b u n g . – Die im Umriss eiförmige, el-liptische Schale ist hinten etwas stärker verlängert als vorn. Sie ist mäßig gewölbt, 68 mm lang und 54 mm hoch. Der Wirbel ist nach vorne gebogen. Der vordere Seiten-rand ist stärker gerundet als der hintere. Es sind zwei Kardinalzähnevorhanden, Seitenzähne sind nicht ein-deutig erkennbar.
Abb. 14. Anomia ephippium (LINNAEUS, 1758), linke Klappe; Ursendorf; SMNS 68618. – a. Außenansicht. b. Innenansicht. – Maßstab 10 mm.
78 PALAEODIVERSITY 2, 2009
V o r k o m m e n : Eggenburgium (SCHAFFER 1910) bis Pliozän (SACCO 1900, zitiert in PFISTER & WEGMÜLLER 2000).
V e r b r e i t u n g : Ermingen; sonstige Vorkommen außer-halb der oberschwäbischen OMM siehe PFISTER & WEGMÜLLER (2000).
Abb. 15–17. Veneridae. – 15. Pitar islandicoides (LAMARCK, 1819), linke Klappe; Ermingen; SMNS 68613. a. Außenansicht. b. In-nenansicht. 16. Paphia deshayesi (COSSMAN & PEYROT, 1911), rechte Klappe; Ermingen; SMNS 68611. a. Außenansicht. b. Innenan-sicht. 17. ? Tapes suevica QUENSTEDT, 1867, linke Klappe; Ermingen; SMNS 68612. a. Außenansicht. b. Innenansicht. – Maßstab 10 mm.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 79
Unterfamilie Tapetinae H. ADAMS & A. ADAMS, 1857 Gattung Paphia RÖDING, 1798
Paphia deshayesi (COSSMAN & PEYROT, 1911) Abb. 16
2000 Paphia deshayesi (COSSMAN & PEYROT, 1911). – PFISTER & WEGMÜLLER, S. 466–467, Taf. 12, Fig. 1–11. − [hier Syno-nymie-Liste]
M a t e r i a l : 1 Exemplar (rechte Klappe, Schalenerhal-tung).
B e s c h r e i b u n g . – Die Schale ist 65 mm lang und 40 mm hoch, hat einen länglichen, elliptischen Umriss und ist stark nach hinten verlängert. Sie ist schwach gewölbt und ungleichseitig. Der Wirbel liegt vor der Mitte und ist nach vorne gebogen. Die Schale weist konzen-trische Rippen in unterschiedlicher Stärke auf. Das Schloss ist nur unvollständig erhalten, die Zähne 1, 3a und 3b sind ausgebildet.
V o r k o m m e n : Helvetikum (= Ottnangium) bis Burdiga-lium (PFISTER & WEGMÜLLER 2000).
V e r b r e i t u n g : Ermingen; sonstige Vorkommen außer-halb der oberschwäbischen OMM siehe PFISTER & WEGMÜLLER (2000).
? Tapes suevica QUENSTEDT, 1867 Abb. 17
1853 Cytherea helvetica MAYER. – MAYER, S. 83.1867 Tapes suevica. – QUENSTEDT, S. 650, Taf. 60, Fig. 16.1871 Tapes helvetica. – MAYER, Taf. 9, Fig. 1.1871 Tapes ulmensis. – MAYER, Taf. 9, Fig. 3.1882 Tapes suevica. – QUENSTEDT, S. 833, Taf. 66, Fig. 7.1910 Tapes helvetica C. MAY. – FRAAS, S. 213, Taf. 64, Fig. 8.1981 Tapes ulmensis. – BEURLEN et al., S. 199, Fig. 11.2005 ? Pelecyora (Cordiopsis) helvetica (MAYER, 1871). –
SCHULTZ, S. 908. – [hier weitere Syanonyme]M a t e r i a l : >100 Exemplare (Schalenerhaltung).
B e s c h r e i b u n g . – Diese in Ermingen am häufig-sten vorkommende Bivalven-Art hat einen rundlichen, leicht elliptischen Umriss. Sie wird ca. 50 mm lang und ca. 40 mm hoch. Die Schale ist schwach gewölbt und un-gleichseitig. Der Wirbel liegt vor der Mitte und ist nach vorne gebogen. Die Lunula verläuft leicht bogenförmig nach unten. Die Area ist vergleichbar groß und konkav. Der hintere Dorsalrand geht unmerklich in den hinteren Seitenrand über. Dieser verläuft schräg abgestutzt nach unten, wo er durch eine schwache Rundung auf den Ven-tralrand trifft. Der vordere Dorsalrand bildet zusammen mit dem vorderen Seitenrand eine leicht eiförmige Wöl-bung. Der vordere Seitenrand geht dann bogenförmig in den Ventralrand über. Der Ventralrand ist gerundet. Die Mantelbucht ist tief und gerundet. In jeder Klappe sind 3 Kardinalzähne vorhanden. Der 3b ist gekerbt und auch der 2b weist eine leichte Kerbung auf.
D i s k u s s i o n . – Diese in Ermingen dominierende Bivalven-Spezies ist allgemein unter dem Namen Tapes helvetica (MAYER, 1853 oder 1871) bekannt. Die älteste Erwähnung dieser Spezies findet sich in MAYER (1853). Sie wird hier als Cytherea helvetica bezeichnet, ohne jedoch beschrieben oder abgebildet zu werden. Ermingen wurde noch nicht als Fundort erwähnt sondern nur Tour la Molière bei Estavayer im Kanton Freiburg, Schweiz. 1867 führte QUENSTEDT für die Art von Ermingen den Namen Tapes suevica ein. 1871 veröffentlichte MAYER eine Be-schreibung und eine Zeichnung der Art, stellte sie zur Gattung Tapes und fasste T. suevica als Synonym auf. MAYER (1871, Taf. 9, Fig. 3) beschrieb für Ermingen des Weiteren die Spezies Tapes ulmensis. Als Synonym führte er ebenfalls T. suevica an. Die neue Art soll sie sich von der vorhergehenden Art durch folgende Merkmale unter-scheiden: 1. Geringere Größe; 2. Eine konvexe, gleich-mäßig ovale Form; 3. Ein weniger gut entwickeltes Schloss; 4. Eine flache Lunula. Laut MAYER (1871) soll diese Spezies in Ermingen seltener sei als T. helvetica. Wegen der großen Variabilität der Muscheln kommt in Ermingen jedoch höchstwahrscheinlich nur eine Art vor. In PROBST (1871) gab MAYER drei Arten für die OMM an. Danach kommen T. helvetica und T. ulmensis in Ermingen vor, T. suevica in Baltringen und Umgebung. Von QUENSTEDT (1882) wurde die Art von Ermingen weiterhin als Tapes suevica be-zeichnet und abgebildet. Sollte es sich bei T. helvetica und T. suevica wirklich um eine Art handeln, wäre Tapes suevica QUENSTEDT, 1867 der gültige Name. Nach SCHULTZ (2005) gehört die Art helvetica möglicherweise zur Gat-tung Pelecyora (Cordiopsis). Eventuell könnte es sich bei der Erminger Art auch um Cordiopsis incrassata (NYST, 1836) handeln (HARZHAUSER, persönl. Mitteilung).
Aufgrund der hohen Anzahl an morphologisch sehr ähnlichen Veneriden im Tertiär wäre zur genauen Bestim-mung ein umfangreicher Vergleich mit Orignalmaterial notwendig. Dies würde aber den Rahmen dieser Arbeit sprengen. Hier wird die Art daher vorläufig als ? Tapes suevica geführt.
V o r k o m m e n : Unter-Miozän, Unteres Ottnagium (SCHULTZ 2005).
V e r b r e i t u n g : Ermingen; Rammingen, Süßen (SCHULTZ 2005); weitere Vorkommen der beiden Arten siehe SCHULTZ (2005).
Weitere von LUTZEIER (1922) erwähnte Arten der Ve-neridae für Ermingen: Venus multilamella LAMARCK, 1818, Venus haidingeri (HOERNES, 1848).
Familie Cardiidae LAMARCK, 1809
B e m e r k u n g . – Oftmals dient die Gattung Cardium als Sammelgattung für nicht eindeutig bestimmbare fos-sile Exemplare der Cardiidae.
80 PALAEODIVERSITY 2, 2009
H a b i t a t u n d L e b e n s w e i s e . – Alle Vertreter der Cardiidae sind Flachwasserbewohner. Sie leben als Filtrierer dicht unter der Sedimentoberfläche.
Unterfamilie Protocardiinae KEEN, 1851
Gattung Nemocardium MEEK, 1876
H e u t i g e V e r b r e i t u n g . – Nemocardium lebt mit einigen Arten im Pazifik, besonders von Japan bis Australien und an der Küste von Zentralamerika (KILIAS 1997).
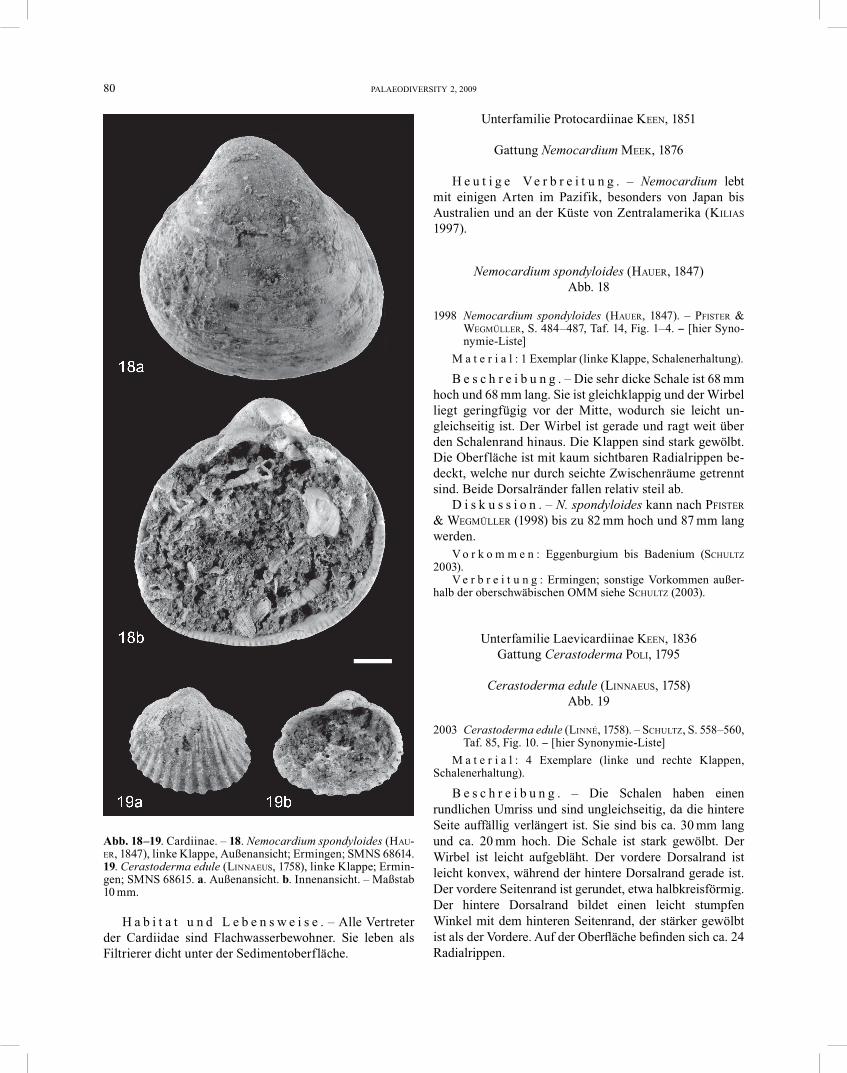
Nemocardium spondyloides (HAUER, 1847) Abb. 18
1998 Nemocardium spondyloides (HAUER, 1847). – PFISTER & WEGMÜLLER, S. 484–487, Taf. 14, Fig. 1–4. − [hier Syno-nymie-Liste]
M a t e r i a l : 1 Exemplar (linke Klappe, Schalenerhaltung).
B e s c h r e i b u n g . – Die sehr dicke Schale ist 68 mm hoch und 68 mm lang. Sie ist gleichklappig und der Wirbel liegt geringfügig vor der Mitte, wodurch sie leicht un-gleich seitig ist. Der Wirbel ist gerade und ragt weit über den Schalenrand hinaus. Die Klappen sind stark gewölbt. Die Oberfläche ist mit kaum sichtbaren Radialrippen be-deckt, welche nur durch seichte Zwischenräume getrennt sind. Beide Dorsalränder fallen relativ steil ab.
D i s k u s s i o n . – N. spondyloides kann nach PFISTER & WEGMÜLLER (1998) bis zu 82 mm hoch und 87 mm lang werden.
V o r k o m m e n : Eggenburgium bis Badenium (SCHULTZ 2003).
V e r b r e i t u n g : Ermingen; sonstige Vorkommen außer-halb der oberschwäbischen OMM siehe SCHULTZ (2003).
Unterfamilie Laevicardiinae KEEN, 1836 Gattung Cerastoderma POLI, 1795
Cerastoderma edule (LINNAEUS, 1758) Abb. 19
2003 Cerastoderma edule (LINNÉ, 1758). – SCHULTZ, S. 558–560, Taf. 85, Fig. 10. − [hier Synonymie-Liste]
M a t e r i a l : 4 Exemplare (linke und rechte Klappen, Schalenerhaltung).
B e s c h r e i b u n g . – Die Schalen haben einen rundlichen Umriss und sind ungleichseitig, da die hintere Seite auffällig verlängert ist. Sie sind bis ca. 30 mm lang und ca. 20 mm hoch. Die Schale ist stark gewölbt. Der Wirbel ist leicht aufgebläht. Der vordere Dorsalrand ist leicht konvex, während der hintere Dorsalrand gerade ist. Der vordere Seitenrand ist gerundet, etwa halbkreisförmig. Der hintere Dorsalrand bildet einen leicht stumpfen Winkel mit dem hinteren Seitenrand, der stärker gewölbt ist als der Vordere. Auf der Oberfläche befinden sich ca. 24 Radialrippen.
Abb. 18–19. Cardiinae. – 18. Nemocardium spondyloides (HAU-ER, 1847), linke Klappe, Außenansicht; Ermingen; SMNS 68614. 19. Cerastoderma edule (LINNAEUS, 1758), linke Klappe; Ermin-gen; SMNS 68615. a. Außenansicht. b. Innenansicht. – Maßstab 10 mm.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 81
D i s k u s s i o n . – Laut WILLMANN (1989) gibt es bei C. edule eine erhebliche Variation in der äußeren Form.
H a b i t a t u n d L e b e n s w e i s e . – Die Art lebt bis in 10 m Wassertiefe in Sand oder Schlamm und kann in sehr großer Dichte vorkommen (WILLMANN 1989; POPPE & GOTO 1993).
V o r k o m m e n : Unter-Miozän bis rezent (SCHULTZ 2003).Ve r b r e i t u n g : Ermingen; sonstige Vorkommen außerhalb
der oberschwäbischen OMM siehe SCHULTZ (2003).
Weitere von LUTZEIER (1922) erwähnte Arten der Car-diidae für Ermingen: Cardium hians BROCCHI, 1814, Acan-thocardia multicostata (BROCCHI, 1814).
Familie Carditidae J. FLEMING, 1828 Unterfamilie Venericardiinae CHAVAN, 1899
Gattung Megacardita SACCO, 1899
H a b i t a t u n d L e b e n s w e i s e . – Megacardita ist rezent laut KILIAS (1997) mit wenigen Arten im West-pazifik (Japan bis NW-Australien) vertreten. Sie leben im Flachwasser (WYE 2003) mit Byssus verankert auf Hart-boden (KILIAS 1997). Alle Mitglieder der Carditidae sind Filtrierer.
Megacardita guenterti PFISTER & WEGMÜLLER, 1998 Abb. 20
1998 Megacardita guenterti n. sp – PFISTER & WEGMÜLLER, S. 471–472, Taf. 8, Fig. 1–5. – [hier Synonymie-Liste]
M a t e r i a l : 3 Exemplare (Schalenerhaltung).
B e s c h r e i b u n g . – Die Schalen sind bis zu 60 mm lang und 35 mm hoch. Sie sind stark nach hinten verlän-gert, wodurch sie einen leicht eiförmigen Umriss haben. Der Wirbel ist nach vorne geneigt. Auf der Oberfläche der gewölbten Schale verlaufen ca. 16 radiale Rippen.
D i s k u s s i o n . – Megacardita jouanetti hat laut PFIS-TER & WEGMÜLLER (1998) mehr Rippen und auch geringere Abstände zwischen diesen als M. guenterti. Schon SUESS (1891) erkannte, dass die bei Ermingen gesammelten Exemplare nicht ganz zu den Beschreibungen für die als Cardita jouanetti bezeichnete Form aus dem Helvetium von St. Gallen passten. Seiner Meinung nach ähnelten die Stücke aus Ermingen eher C. crassicosta oder C. scabri-costa, zu welchen die Erminger Formen aber auch nicht eindeutig zugeordnet werden konnten. Erst 1998 wurde das Problem durch die Beschreibung einer neuen Art durch PFISTER & WEGMÜLLER gelöst (SCHULTZ 2003).
V o r k o m m e n : Burdigalium (PFISTER & WEGMÜLLER 1998); unteres Ottnangium (SCHULTZ 2003).
V e r b r e i t u n g : Ermingen, Ursendorf; sonstige Vorkom-men außerhalb der oberschwäbischen OMM siehe SCHULTZ (2003).
Weitere von LUTZEIER (1922) erwähnte Arten der Carditidae für Ermingen: Cardita crassicosta LAMARCK, 1819, Cardita scabricosta (MICHELOTTI).
Familie Chamidae LAMARCK, 1809
Gattung Pseudochama ODHNER, 1917
B e m e r k u n g . – Nach LINDNER (1999) wird Pseudo-chama nur als Untergattung von Chama angesehen.
Pseudochama gryphina (LAMARCK, 1819) Abb. 21
2003 Pseudochama (Pseudochama) gryphina gryphina (LA- MARCK, 1819). – SCHULTZ, S. 450–453, Taf. 63, Fig. 6, 7, 9. – [hier Synonymie-Liste]
M a t e r i a l : 2 Exemplare (linke Klappen, Schalenerhal-tung).
B e s c h r e i b u n g . – Die Schalen sind bis zu 30 mm hoch. Sie sind stark gewölbt und haben eine unregelmäßige Umrissform. Der Wirbel springt weit vor und ist nach links eingerollt. Auf der Oberfläche befinden sich konzen-trische Lamellen.
Abb. 20. Megacardita guenterti PFISTER & WEGMÜLLER, 1998, linke Klappe; Ermingen; SMNS 68616. – a. Außenansicht. b. Innenansicht. – Maßstab 10 mm.

82 PALAEODIVERSITY 2, 2009
H a b i t a t u n d L e b e n s w e i s e . – Pseudochama gryphina kommt im Mittelmeer sowie angrenzendem At-lantik (Mediterran-Lusitanische Provinz) bis nach Angola vor (POPPE & GOTO 1993). Dort lebt sie mit der rechten Klappe auf Hartsubstrat festzementiert in einer Tiefe zwischen 10 und 60 m.
V o r k o m m e n : P. gryphina kommt seit dem Eggenbur-gium vor (SCHULTZ 2003).
V e r b r e i t u n g : Ermingen; sonstige Vorkommen außer-halb der oberschwäbischen OMM siehe SCHULTZ (2003).
Familie Corbulidae LAMARCK, 1818 Unterfamilie Corbulinae LAMARCK, 1818
Gattung Corbula BRUGUIÈRE, 1797
H a b i t a t u n d L e b e n s w e i s e : Corbula lebt im Atlantik, Mittelmeer und Pazifik (LINDNER 1999) in sandi-gem Schlick und ernährt sich von Diatomeen, Bakterien etc., die sie von der Oberfläche aufnimmt (KILIAS 1997).
Corbula sp. Abb. 22
M a t e r i a l : 2 Handstücke mit Steinkernen.
B e s c h r e i b u n g . – Die bis zu 4 mm langen Exem-plare haben eine leicht dreieckige Form. Die Steinkerne sind zum Teil stark gewölbt und eine Aufblähung des Wir-bels lässt sich oftmals erahnen.
D i s k u s s i o n . – Aufgrund der Steinkern-Erhaltung ist die exakte Bestimmung der Art nicht möglich. Nach der dreieckigen Form könnte es sich um Corbula gibba (OLIVI, 1792) handeln, welche rezent im Ostatlantik und im Mittelmeer (KILIAS 1997) sowie in der Nordsee vor-kommt (WILLMANN 1989).
V e r b r e i t u n g : Ursendorf.
Familie Pholadidae LAMARCK, 1809 Unterfamilie Pholadinae LAMARCK, 1809
Gattung Pholas LINNAEUS, 1758
H a b i t a t u n d L e b e n s w e i s e . – Die Familie ist weltweit verbreitet. Die Arten der Pholadidae bohren mechanisch in Holz, Korallen und Gestein. Sie sitzen dann zeitlebens in den Hohlräumen fest (LINDNER 1999).
Pholas desmoulinsii BENOIST, 1876 Abb. 23
2001 Pholas desmoulinsii BENOIST, 1876. – PFISTER & WEGMÜL-LER, S. 403–404, Taf. 3, Fig. 1–7. − [hier Synonymie-Liste]
M a t e r i a l : 1 Exemplar (linke Klappe, Steinkern).
B e s c h r e i b u n g . – Die Schale ist 50 mm lang und 20 mm hoch. Sie ist stark ungleichseitig und nach hinten stark verlängert. Die hintere Hälfte nimmt 4/5 der Gesamt-länge ein. Der hintere Seitenrand ist zungenförmig verlän-gert. Der Vorderrand endet in einem zugespitzten Schna-bel. Oberflächenskulpturen sind an dem Steinkern nicht zu erkennen.
D i s k u s s i o n . – HÖLZL (1958) gibt für Pholas des-moulinsii diesselben Proportionen an. Nach PFISTER & WEGMÜLLER (2001) ist P. dactylus LINNAEUS, 1758 mit die-ser Art vergleichbar.
V o r k o m m e n : Unter-Miozän, siehe PFISTER & WEGMÜL-LER (2001).
V e r b r e i t u n g : Ursendorf; LUTZEIER (1922) gibt für di-verse Örtlichkeiten in der oberschwäbischen OMM das Vorkom-
Abb. 21. Pseudochama gryphina (LAMARCK, 1819), linke Klappe; Ursendorf; SMNS 68619. – a. Außenansicht. b. Innenansicht. – Maßstab 10 mm.
Abb. 22. Corbula sp., rechte Klappe, Außenansicht; Ursendorf; SMNS 68620. – Maßstab 5 mm.
Abb. 23. Pholas desmoulinsii BENOIST, 1876, linke Klappe, Außenansicht; Ursendorf; SMNS 68617. – Maßstab 10 mm.
HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 83
men von Pholas sp. an. Sonstige Vorkommen außerhalb der oberschwäbischen OMM siehe PFISTER & WEGMÜLLER (2001).
5.2. Gastropoden
Familie Fissurellidae FLEMMING, 1822 Unterfamilie Diodorinae WENZ, 1938
Gattung Diodora J. E. GRAY, 1834
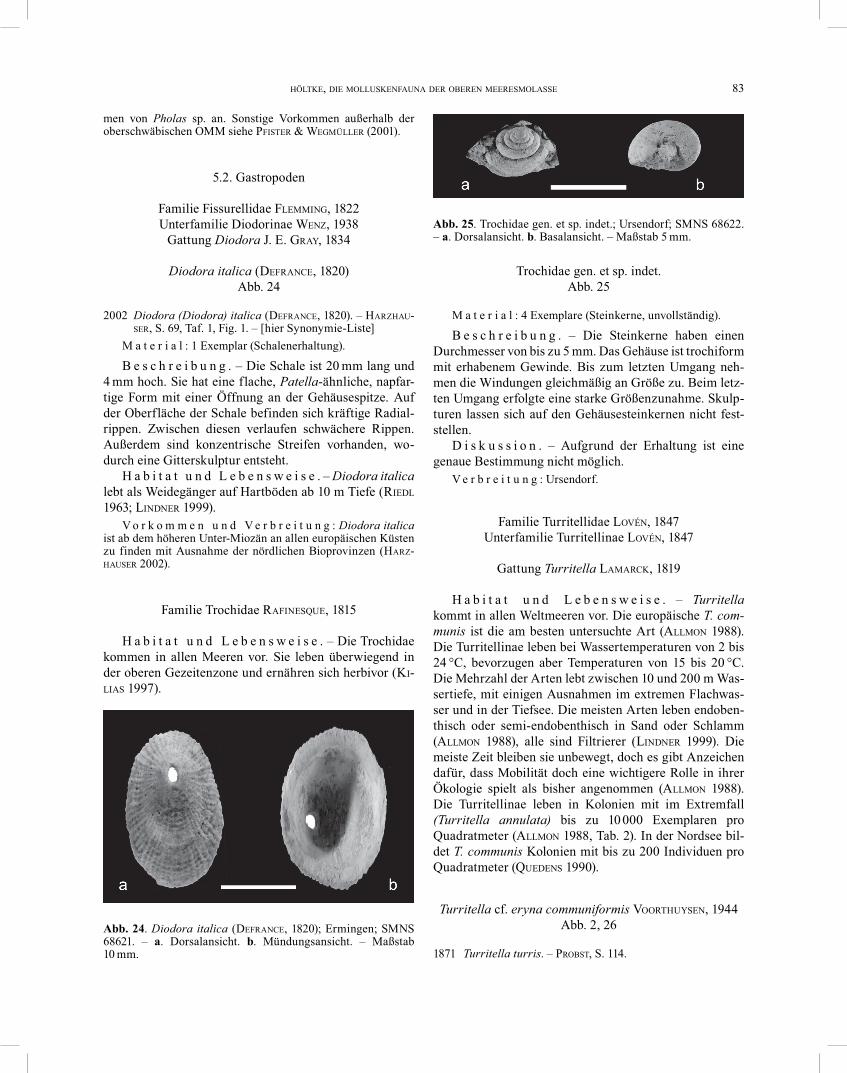
Diodora italica (DEFRANCE, 1820) Abb. 24
2002 Diodora (Diodora) italica (DEFRANCE, 1820). – HARZHAU-SER, S. 69, Taf. 1, Fig. 1. – [hier Synonymie-Liste]
M a t e r i a l : 1 Exemplar (Schalenerhaltung).
B e s c h r e i b u n g . – Die Schale ist 20 mm lang und 4 mm hoch. Sie hat eine flache, Patella-ähnliche, napfar-tige Form mit einer Öffnung an der Gehäusespitze. Auf der Oberfläche der Schale befinden sich kräftige Radial-rippen. Zwischen diesen verlaufen schwächere Rippen. Außerdem sind konzentrische Streifen vorhanden, wo-durch eine Gitterskulptur entsteht.
H a b i t a t u n d L e b e n s w e i s e . – Diodora italica lebt als Weidegänger auf Hartböden ab 10 m Tiefe (RIEDL 1963; LINDNER 1999).
V o r k o m m e n u n d V e r b r e i t u n g : Diodora italica ist ab dem höheren Unter-Miozän an allen europäischen Küsten zu finden mit Ausnahme der nördlichen Bioprovinzen (HARZ-HAUSER 2002).
Familie Trochidae RAFINESQUE, 1815
H a b i t a t u n d L e b e n s w e i s e . – Die Trochidae kommen in allen Meeren vor. Sie leben überwiegend in der oberen Gezeitenzone und ernähren sich herbivor (KI-LIAS 1997).
Trochidae gen. et sp. indet. Abb. 25
M a t e r i a l : 4 Exemplare (Steinkerne, unvollständig).
B e s c h r e i b u n g . – Die Steinkerne haben einen Durch messer von bis zu 5 mm. Das Gehäuse ist trochiform mit erhabenem Gewinde. Bis zum letzten Umgang neh-men die Windungen gleichmäßig an Größe zu. Beim letz-ten Umgang erfolgte eine starke Größenzunahme. Skulp-turen lassen sich auf den Gehäusesteinkernen nicht fest-stellen.
D i s k u s s i o n . – Aufgrund der Erhaltung ist eine genaue Bestimmung nicht möglich.
V e r b r e i t u n g : Ursendorf.
Familie Turritellidae LOVÉN, 1847 Unterfamilie Turritellinae LOVÉN, 1847
Gattung Turritella LAMARCK, 1819
H a b i t a t u n d L e b e n s w e i s e . – Turritella kommt in allen Weltmeeren vor. Die europäische T. com-munis ist die am besten untersuchte Art (ALLMON 1988). Die Turritellinae leben bei Wassertemperaturen von 2 bis 24 °C, bevorzugen aber Temperaturen von 15 bis 20 °C. Die Mehrzahl der Arten lebt zwischen 10 und 200 m Was-sertiefe, mit einigen Ausnahmen im extremen Flachwas-ser und in der Tiefsee. Die meisten Arten leben endoben-thisch oder semi-endobenthisch in Sand oder Schlamm (ALLMON 1988), alle sind Filtrierer (LINDNER 1999). Die meiste Zeit bleiben sie unbewegt, doch es gibt Anzeichen dafür, dass Mobilität doch eine wichtigere Rolle in ihrer Ökologie spielt als bisher angenommen (ALLMON 1988). Die Turritellinae leben in Kolonien mit im Extremfall (Turritella annulata) bis zu 10 000 Exemplaren pro Quadratmeter (ALLMON 1988, Tab. 2). In der Nordsee bil-det T. communis Kolonien mit bis zu 200 Individuen pro Quadratmeter (QUEDENS 1990).
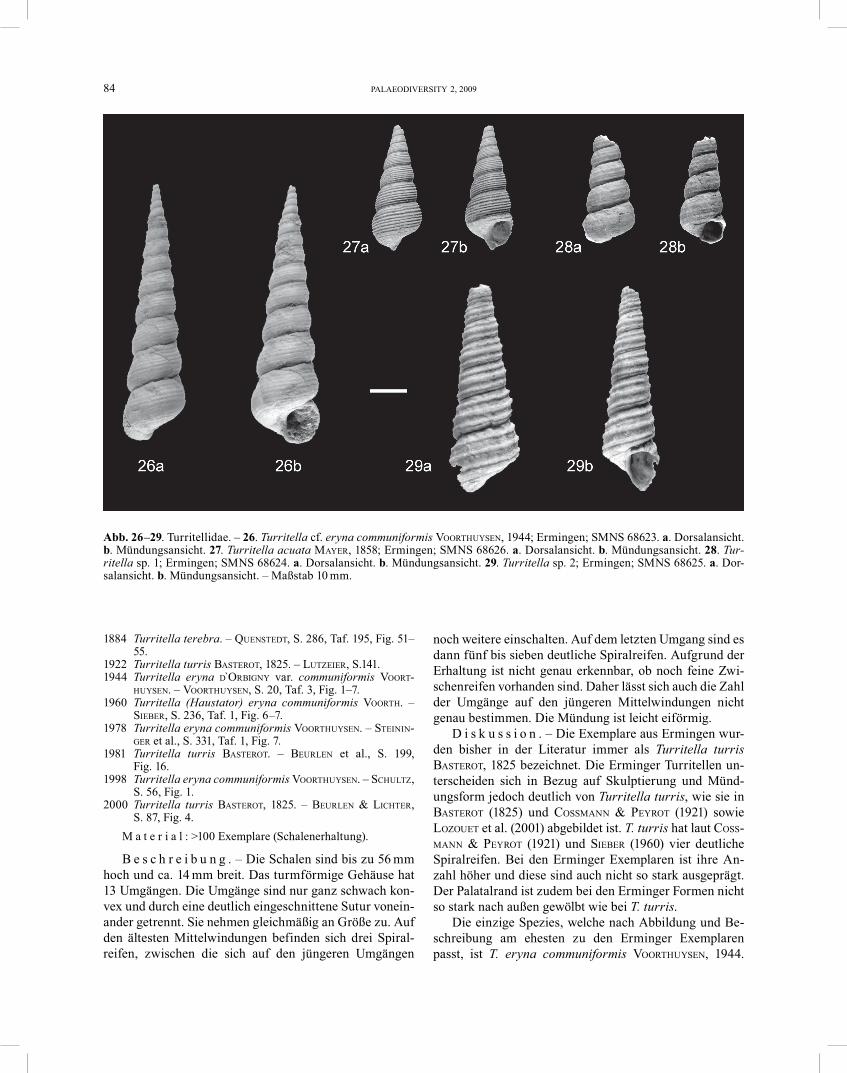
Turritella cf. eryna communiformis VOORTHUYSEN, 1944 Abb. 2, 26
1871 Turritella turris. – PROBST, S. 114.
Abb. 24. Diodora italica (DEFRANCE, 1820); Ermingen; SMNS 68621. – a. Dorsalansicht. b. Mündungsansicht. – Maßstab 10 mm.
Abb. 25. Trochidae gen. et sp. indet.; Ursendorf; SMNS 68622. – a. Dorsalansicht. b. Basalansicht. – Maßstab 5 mm.
84 PALAEODIVERSITY 2, 2009
1884 Turritella terebra. – QUENSTEDT, S. 286, Taf. 195, Fig. 51–55.
1922 Turritella turris BASTEROT, 1825. – LUTZEIER, S.141.1944 Turritella eryna D ORBIGNY var. communiformis VOO RT-
HUYSEN. – VOORTHUYSEN, S. 20, Taf. 3, Fig. 1–7.1960 Turritella (Haustator) eryna communiformis VOORTH. –
SIEBER, S. 236, Taf. 1, Fig. 6–7.1978 Turritella eryna communiformis VOORTHUYSEN. – STEININ-
GER et al., S. 331, Taf. 1, Fig. 7.1981 Turritella turris BASTEROT. – BEURLEN et al., S. 199,
Fig. 16.1998 Turritella eryna communiformis VOORTHUYSEN. – SCHULTZ,
S. 56, Fig. 1.2000 Turritella turris BASTEROT, 1825. – BEURLEN & LICHTER,
S. 87, Fig. 4.M a t e r i a l : >100 Exemplare (Schalenerhaltung).
B e s c h r e i b u n g . – Die Schalen sind bis zu 56 mm hoch und ca. 14 mm breit. Das turmförmige Gehäuse hat 13 Umgängen. Die Umgänge sind nur ganz schwach kon-vex und durch eine deutlich eingeschnittene Sutur vonein-ander getrennt. Sie nehmen gleichmäßig an Größe zu. Auf den ältesten Mittelwindungen befinden sich drei Spiral-reifen, zwischen die sich auf den jüngeren Umgängen
noch weitere einschalten. Auf dem letzten Umgang sind es dann fünf bis sieben deutliche Spiralreifen. Aufgrund der Erhaltung ist nicht genau erkennbar, ob noch feine Zwi-schenreifen vorhanden sind. Daher lässt sich auch die Zahl der Umgänge auf den jüngeren Mittelwindungen nicht genau bestimmen. Die Mündung ist leicht eiförmig.
D i s k u s s i o n . – Die Exemplare aus Ermingen wur-den bisher in der Literatur immer als Turritella turris BASTEROT, 1825 bezeichnet. Die Erminger Turritellen un-terscheiden sich in Bezug auf Skulptierung und Münd-ungsform jedoch deutlich von Turritella turris, wie sie in BASTEROT (1825) und COSSMANN & PEYROT (1921) sowie LOZOUET et al. (2001) abgebildet ist. T. turris hat laut COSS-MANN & PEYROT (1921) und SIEBER (1960) vier deutliche Spiralreifen. Bei den Erminger Exemplaren ist ihre An-zahl höher und diese sind auch nicht so stark ausgeprägt. Der Palatalrand ist zudem bei den Erminger Formen nicht so stark nach außen gewölbt wie bei T. turris.
Die einzige Spezies, welche nach Abbildung und Be-schreibung am ehesten zu den Erminger Exemplaren passt, ist T. eryna communiformis VOORTHUYSEN, 1944.
Abb. 26–29. Turritellidae. – 26. Turritella cf. eryna communiformis VOORTHUYSEN, 1944; Ermingen; SMNS 68623. a. Dorsalansicht. b. Mündungsansicht. 27. Turritella acuata MAYER, 1858; Ermingen; SMNS 68626. a. Dorsalansicht. b. Mündungsansicht. 28. Tur-ritella sp. 1; Ermingen; SMNS 68624. a. Dorsalansicht. b. Mündungsansicht. 29. Turritella sp. 2; Ermingen; SMNS 68625. a. Dor-salansicht. b. Mündungsansicht. – Maßstab 10 mm.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 85
Nach COSSMANN & PEYROT (1921) finden sich in der Litera-tur einige Formen, welche fälschlicherweise als T. turris bezeichnet werden. Auch SIEBER (1960) führte an, dass die Behandlung der T. turris bei SCHAFFER (1910) nicht mehr aktuell ist. Die Bestimmung von miozänen Turritellen ist aufgrund der Formenfülle ohne Vergleich mit Original-material äußerst problematisch. Somit ist auch die hier gegebene Identifizierung der Art mit einem Unsicher-heitsfaktor behaftet.
Aufgrund des massenhaften Vorkommens in Ermin-gen wurde diese Art namensgebend für die „Erminger Turritellenplatte“. Allerdings findet sich weder bei VOORT-HUYSEN (1944) noch bei SIEBER (1960) ein Hinweis auf dieses Massenvorkommen.
In der Fossilliste von MAYER (1853) ist T. turris nicht aufgeführt. PROBST (1871) nannte T. turris als einzige Tur-ritellide für Ermingen. QUENSTEDT (1884) bezeichnete die Erminger Formen als T. terebra und führte an, dass ähnli-che Formen in Saucats/Frankreich als T. turris bezeichnet werden. Ferner verwies QUENSTEDT (1884) auf eine Abbil-dung in VON ZIETEN (1833). Daher nahm LUTZEIER (1922) an, VON ZIETEN (1833) hätte die “Erminger Turritellen-platte“ als erster beschrieben. In Wirklichkeit bildete aber VON ZIETEN (1833) eine T. terebra aus der Umgebung von Baltringen ab. Wer den Begriff „Erminger Turritellen-platte“ als erster verwendet hat, ist unklar (RASSER & NEBELSICK 2006).
V o r k o m m e n : Aquitanium, Burdigalium (VOORTHUYSEN 1944; SIEBER 1960).
V e r b r e i t u n g : Ermingen; Baltringen und Umgebung (PROBST 1871); Ballendorf, Hochsträß, Rammingen, Temmen-hausen (LUTZEIER 1922); Juckenberg im Klettgau (GEYER et al. 2003); Niederlande (VOORTHUYSEN 1944); weitere Vorkommen siehe SIEBER (1960).
Turritella acuta MAYER, 1858 Abb. 27
1921 Turritella acuta MAYER, 1858. – COSSMANN & PEYROT, S. 38, Taf. 2, Fig. 46–47.
M a t e r i a l : 8 Exemplare (Schalenerhaltung, unvollstän-dig).
B e s c h r e i b u n g . – Die unvollständigen Gehäuse sind bis 34 mm hoch und 13 mm breit. Die Umgänge neh-men schnell an Größe zu und sind bauchiger als bei den vorherigen Formen. T. acuta hat ca. acht deutlich aus-geprägte Spiralreifen auf den Windungen.
D i s k u s s i o n . – Die Form und Skulpturierung der Umgänge unterscheiden die Art deutlich von den anderen Turritellen.
V o r k o m m e n : Burdigalium (COSSMANN & PEYROT 1921).V e r b r e i t u n g : Ermingen; Vorkommen in Frankreich
siehe COSSMANN & PEYROT (1921).
Turritella sp. 1 Abb. 28
M a t e r i a l : 2 Exemplare (Schalenerhaltung, unvollstän-dig).
B e s c h r e i b u n g . – Die Umgänge sind flach konvex und durch eine tiefe Naht voneinander getrennt. Auf den Umgängen sind jeweils drei Spiralreifen erkennbar. Zwi-schen diesen verlaufen weitere, sehr schwach ausgebildete Spiralreifen.
D i s k u s s i o n . – Die Anzahl der Spiralreifen sowie die Tiefe der Naht unterscheiden die Exemplare von den anderen Turritellen von Ermingen. Aufgrund der unvoll-ständigen Erhaltung ist aber eine genaue Art-Ermittlung nicht möglich.
V e r b r e i t u n g : Ermingen.
Turritella sp. 2 Abb. 29
M a t e r i a l : 1 Exemplar (Schalenerhaltung, unvollstän-dig).
B e s c h r e i b u n g . – Das Gehäuse ist mittelgroß mit bis zu 12 Umgängen. Die Umgänge nehmen gleichmäßig an Größe zu und sind flach. Sie werden durch eine deutli-che Sutur voneinander getrennt. Auf den Umgängen befin-den sich jeweils drei relativ hohe und schmale Spiralrip-pen. Zwischen diesen verlaufen noch weitere, sehr feine, Spiralrippen. Von der Mündungsunterseite an verlaufen zwei von diesen Spiralrippen entlang der Basis nach oben bis zur Parietalregion. Parallel zu diesen verlaufen auch noch zwei sehr feine Spiralrippen.
D i s k u s s i o n . – Durch die Skulptierung unter-scheidet sich das Gehäuse deutlich von den oben beschrie-benen Formen. Aufgrund der unvollständigen Erhaltung ist aber eine genauere Bestimmung nicht möglich.
V e r b r e i t u n g : Ermingen.
Von LUTZEIER (1922) zusätzlich angeführte Turritella-Art für Ermingen: Turritella vermicularis (BROCCHI, 1814).
Familie Calyptraeidae LAMARCK, 1809
Gattung Calyptraea LAMARCK, 1809
H a b i t a t u n d L e b e n s w e i s e . – Die Gattung kommt im Atlantik und Pazifik vor (KILIAS 1997) sowie mit einer Art auch im Mittelmeer. Die Tiere sitzen auf Hartsubstrat (DANCE 1977) oder auch auf anderen Scha-lentieren (LINDNER 1999). Sie leben unterhalb der Gezei-

86 PALAEODIVERSITY 2, 2009
tenlinie bis in 260 m Tiefe (DANCE 1977; ABBOT & DANCE 1998). Die Vertreter dieser Gattung sind Filtrierer.
Calyptraea sp. Abb. 30
M a t e r i a l : 1 Exemplar (Steinkern).
B e s c h r e i b u n g . – Der Gehäuseumriss ist rundlich oval. Es hat einen Durchmesser von ca. 10 mm und ist ca. 6 mm hoch. Die Schale hat eine hohe und zugespitzte, kappenförmige Morphologie. Die Gehäusespitze liegt genau in der Mitte.
V e r b r e i t u n g : Ursendorf.
Familie Xenophoridae TROSCHEL, 1852
Gattung Xenophora FISCHER VON WALDHEIM, 1807
H a b i t a t u n d L e b e n s w e i s e . – Die Gattung kommt ausschließlich in warmen Meeren vor. Die meisten Arten bewohnen tieferes Wasser, wo sie auf Schlamm-boden leben (ABBOT & DANCE 1998; WYE 2003). Die Xeno-phoridae befestigen an ihrem Gehäuse Muschelschalen, kleine Steine oder sonstige Hartteile, wodurch sie verhin-dern, in das schlammige Substrat einzusinken (WYE 2003). Die Mitglieder der Familie sind Detritusfresser (KILIAS 1997).
Xenophora cf. deshayesi (MICHELOTTI, 1847) Abb. 31
1973 Xenophora deshayesi (MICHELOTTI, 1847). – HÖLZL in STEININGER et al., S. 409, Taf. 4, Fig. 10. – [hier Synony-mie-Liste]
1998 Xenophora deshayesi (MICHELOTTI, 1847). – SCHULTZ, S. 60, Taf. 23, Fig. 5.
M a t e r i a l : 1 Exemplar (unvollständiges Gewinde, Schalenerhaltung).
B e s c h r e i b u n g . – Das coeloconoide Gehäuse und die Umgänge sind flach. Es lassen sich auf dem Gehäuse
noch die Abdrücke bzw. Spuren der agglutinierten Hart-teile erkennen.
D i s k u s s i o n . – Aufgrund der unvollständigen Er-haltung ist eine eindeutige Bestimmung nicht möglich.
V o r k o m m e n : Xenophora deshayesi kommt vom Egeri-um bis zum Badenium vor (HÖLZL in STEININGER et al. 1973).
V e r b r e i t u n g : Ermingen; weitere Vorkommen außer-halb der OMM siehe HÖLZL in STEININGER et al. (1973).
Familie Cypraeidae RAFINESQUE, 1815
Gattung Cypraea LINNAEUS, 1758
H a b i t a t u n d L e b e n s w e i s e . – Cypraea ist mit über 500 Arten in allen warmen und tropischen Meeren verbreitet. Die meisten Arten kommen im Indopazifik vor, nur vier in der Mediterran-Lusitanischen Provinz (LIND-NER 2000). Einige Arten leben unter Felsen und in Höhlun-gen von Riffen in unterschiedlicher Tiefe (ALLEN & STEENE 1995), andere auf steinigen, sandigen oder sandig-schlam-migen Böden (LINDNER 2000). Die Tiere fressen Korallen-tiere, Polypen, Gorgonien und tierischen Detritus (LIND-NER 1999).
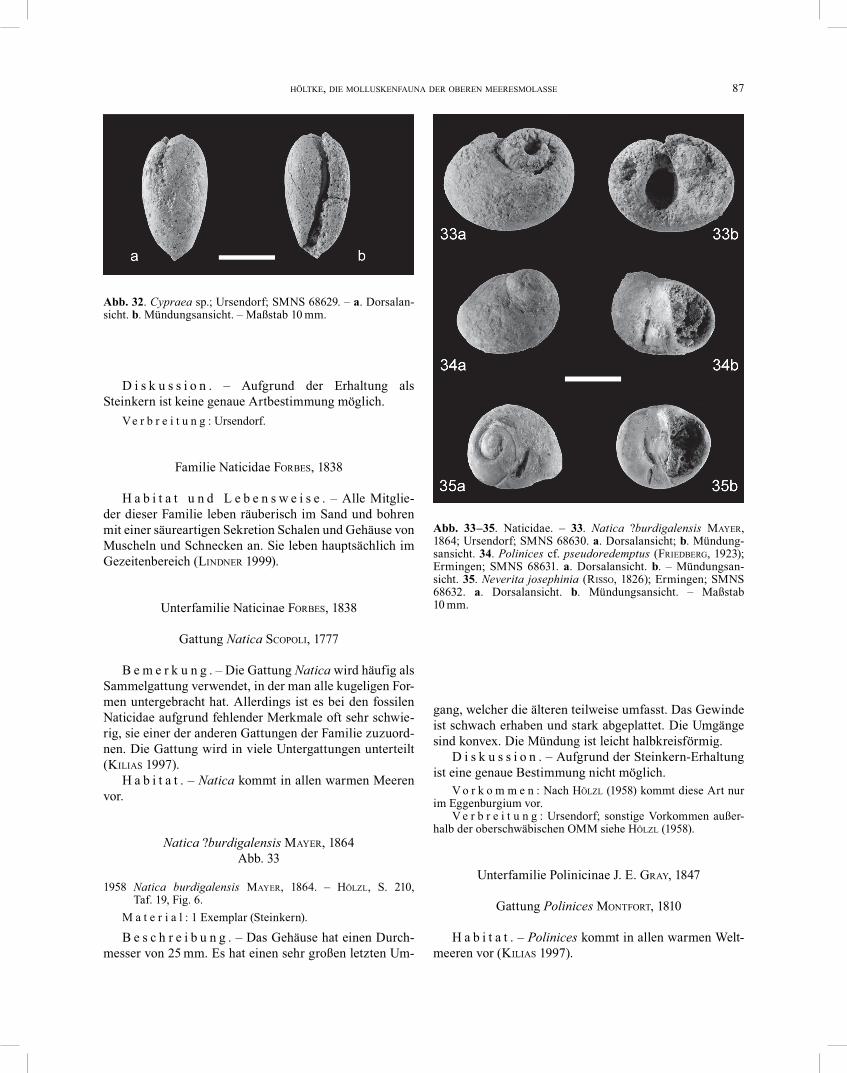
Cypraea sp. Abb. 32
M a t e r i a l : 5 Exemplare (Steinkern).
B e s c h r e i b u n g . – Das Gehäuse ist 25 mm hoch und ca. 13 mm breit. Es ist involut und hat einen ovalen Umriss. Dorsal ist das kleine, eingesunkene Gewinde sicht bar. Die Basis ist sehr schwach konvex. Die Columel-larlippe ist leicht konkav. Die Basis ist fast doppelt so breit wie das Labrum.
Abb. 30. Calyptraea sp.; Ursendorf; SMNS 68627. – a. Dor-salansicht. b. Lateralansicht. – Maßstab 10 mm.
Abb. 31. Xenophora cf. deshayesi (MICHELOTTI, 1847); Ermin-gen; SMNS 68628. – a. Lateralansicht. b. Dorsalansicht. – Maßstab 10 mm.
HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 87
D i s k u s s i o n . – Aufgrund der Erhaltung als Steinkern ist keine genaue Artbestimmung möglich.
Ve r b r e i t u n g : Ursendorf.
Familie Naticidae FORBES, 1838
H a b i t a t u n d L e b e n s w e i s e . – Alle Mitglie-der dieser Familie leben räuberisch im Sand und bohren mit einer säureartigen Sekretion Schalen und Gehäuse von Muscheln und Schnecken an. Sie leben hauptsächlich im Gezeitenbereich (LINDNER 1999).
Unterfamilie Naticinae FORBES, 1838
Gattung Natica SCOPOLI, 1777
B e m e r k u n g . – Die Gattung Natica wird häufig als Sammelgattung verwendet, in der man alle kugeligen For-men untergebracht hat. Allerdings ist es bei den fossilen Naticidae aufgrund fehlender Merkmale oft sehr schwie-rig, sie einer der anderen Gattungen der Familie zuzuord-nen. Die Gattung wird in viele Untergattungen unterteilt (KILIAS 1997).
H a b i t a t . – Natica kommt in allen warmen Meeren vor.
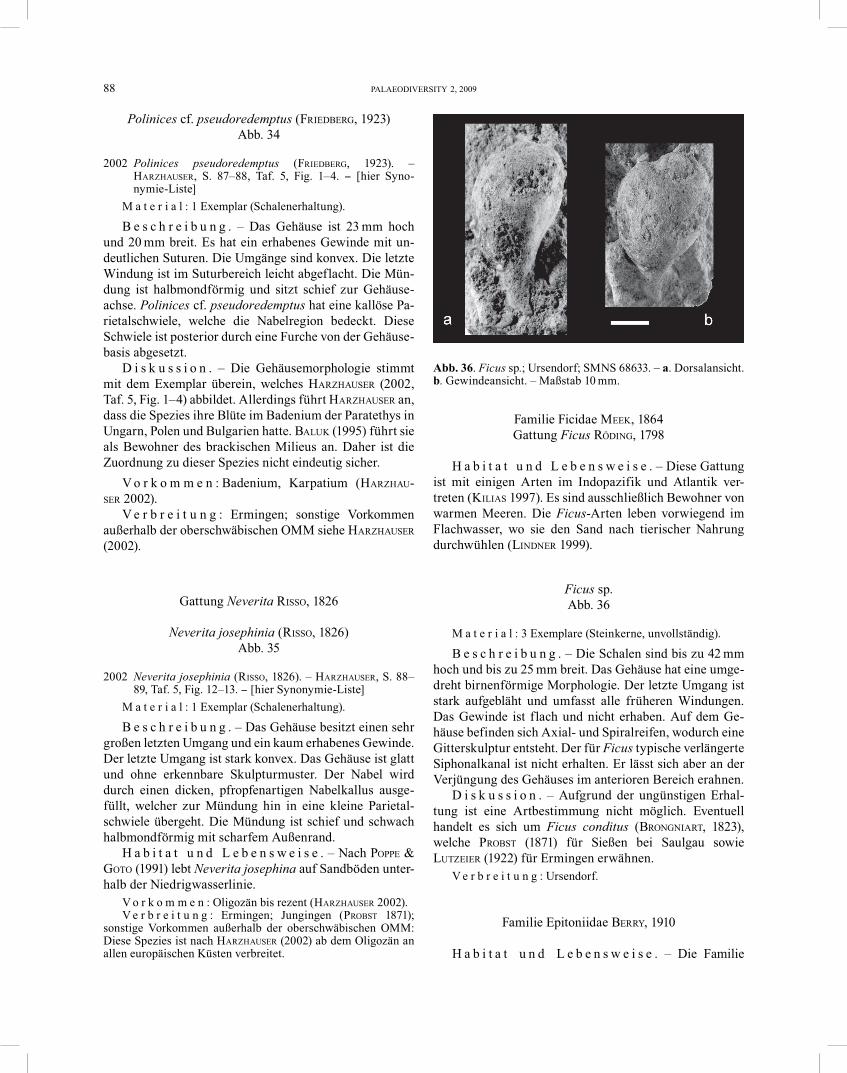
Natica ?burdigalensis MAYER, 1864 Abb. 33
1958 Natica burdigalensis MAYER, 1864. – HÖLZL, S. 210, Taf. 19, Fig. 6.
M a t e r i a l : 1 Exemplar (Steinkern).
B e s c h r e i b u n g . – Das Gehäuse hat einen Durch-messer von 25 mm. Es hat einen sehr großen letzten Um-
gang, welcher die älteren teilweise umfasst. Das Gewinde ist schwach erhaben und stark abgeplattet. Die Umgänge sind konvex. Die Mündung ist leicht halbkreisförmig.
D i s k u s s i o n . – Aufgrund der Steinkern-Erhaltung ist eine genaue Bestimmung nicht möglich.
V o r k o m m e n : Nach HÖLZL (1958) kommt diese Art nur im Eggenburgium vor.
V e r b r e i t u n g : Ursendorf; sonstige Vorkommen außer-halb der oberschwäbischen OMM siehe HÖLZL (1958).
Unterfamilie Polinicinae J. E. GRAY, 1847
Gattung Polinices MONTFORT, 1810
H a b i t a t . – Polinices kommt in allen warmen Welt-meeren vor (KILIAS 1997).
Abb. 32. Cypraea sp.; Ursendorf; SMNS 68629. – a. Dorsalan-sicht. b. Mündungsansicht. – Maßstab 10 mm.
Abb. 33–35. Naticidae. – 33. Natica ?burdigalensis MAYER, 1864; Ursendorf; SMNS 68630. a. Dorsalansicht; b. Mündung-sansicht. 34. Polinices cf. pseudoredemptus (FRIEDBERG, 1923); Ermingen; SMNS 68631. a. Dorsalansicht. b. – Mündungsan-sicht. 35. Neverita josephinia (RISSO, 1826); Ermingen; SMNS 68632. a. Dorsalansicht. b. Mündungsansicht. – Maßstab 10 mm.
88 PALAEODIVERSITY 2, 2009
Polinices cf. pseudoredemptus (FRIEDBERG, 1923) Abb. 34
2002 Polinices pseudoredemptus (FRIEDBERG, 1923). – HARZHAUSER, S. 87–88, Taf. 5, Fig. 1–4. − [hier Syno-nymie-Liste]
M a t e r i a l : 1 Exemplar (Schalenerhaltung).
B e s c h r e i b u n g . – Das Gehäuse ist 23 mm hoch und 20 mm breit. Es hat ein erhabenes Gewinde mit un-deutlichen Suturen. Die Umgänge sind konvex. Die letzte Windung ist im Suturbereich leicht abgeflacht. Die Mün-dung ist halbmondförmig und sitzt schief zur Gehäuse-achse. Polinices cf. pseudoredemptus hat eine kallöse Pa-rietalschwiele, welche die Nabelregion bedeckt. Diese Schwiele ist posterior durch eine Furche von der Gehäuse-basis abgesetzt.
D i s k u s s i o n . – Die Gehäusemorphologie stimmt mit dem Exemplar überein, welches HARZHAUSER (2002, Taf. 5, Fig. 1–4) abbildet. Allerdings führt HARZHAUSER an, dass die Spezies ihre Blüte im Badenium der Paratethys in Ungarn, Polen und Bulgarien hatte. BALUK (1995) führt sie als Bewohner des brackischen Milieus an. Daher ist die Zuordnung zu dieser Spezies nicht eindeutig sicher.
V o r k o m m e n : Badenium, Karpatium (HARZHAU-SER 2002).
V e r b r e i t u n g : Ermingen; sonstige Vorkommen außerhalb der oberschwäbischen OMM siehe HARZ HAUSER (2002).
Gattung Neverita RISSO, 1826
Neverita josephinia (RISSO, 1826) Abb. 35
2002 Neverita josephinia (RISSO, 1826). – HARZHAUSER, S. 88–89, Taf. 5, Fig. 12–13. − [hier Synonymie-Liste]
M a t e r i a l : 1 Exemplar (Schalenerhaltung).
B e s c h r e i b u n g . – Das Gehäuse besitzt einen sehr großen letzten Umgang und ein kaum erhabenes Gewinde. Der letzte Umgang ist stark konvex. Das Gehäuse ist glatt und ohne erkennbare Skulpturmuster. Der Nabel wird durch einen dicken, pfropfenartigen Nabelkallus ausge-füllt, welcher zur Mündung hin in eine kleine Parietal-schwiele übergeht. Die Mündung ist schief und schwach halbmondförmig mit scharfem Außenrand.
H a b i t a t u n d L e b e n s w e i s e . – Nach POPPE & GOTO (1991) lebt Neverita josephina auf Sandböden unter-halb der Niedrigwasserlinie.
V o r k o m m e n : Oligozän bis rezent (HARZHAUSER 2002).V e r b r e i t u n g : Ermingen; Jungingen (PROBST 1871);
sonstige Vorkommen außerhalb der oberschwäbischen OMM: Diese Spezies ist nach HARZHAUSER (2002) ab dem Oligozän an allen europäischen Küsten verbreitet.
Familie Ficidae MEEK, 1864 Gattung Ficus RÖDING, 1798
H a b i t a t u n d L e b e n s w e i s e . – Diese Gattung ist mit einigen Arten im Indopazifik und Atlantik ver-treten (KILIAS 1997). Es sind ausschließlich Bewohner von warmen Meeren. Die Ficus-Arten leben vorwiegend im Flachwasser, wo sie den Sand nach tierischer Nahrung durchwühlen (LINDNER 1999).
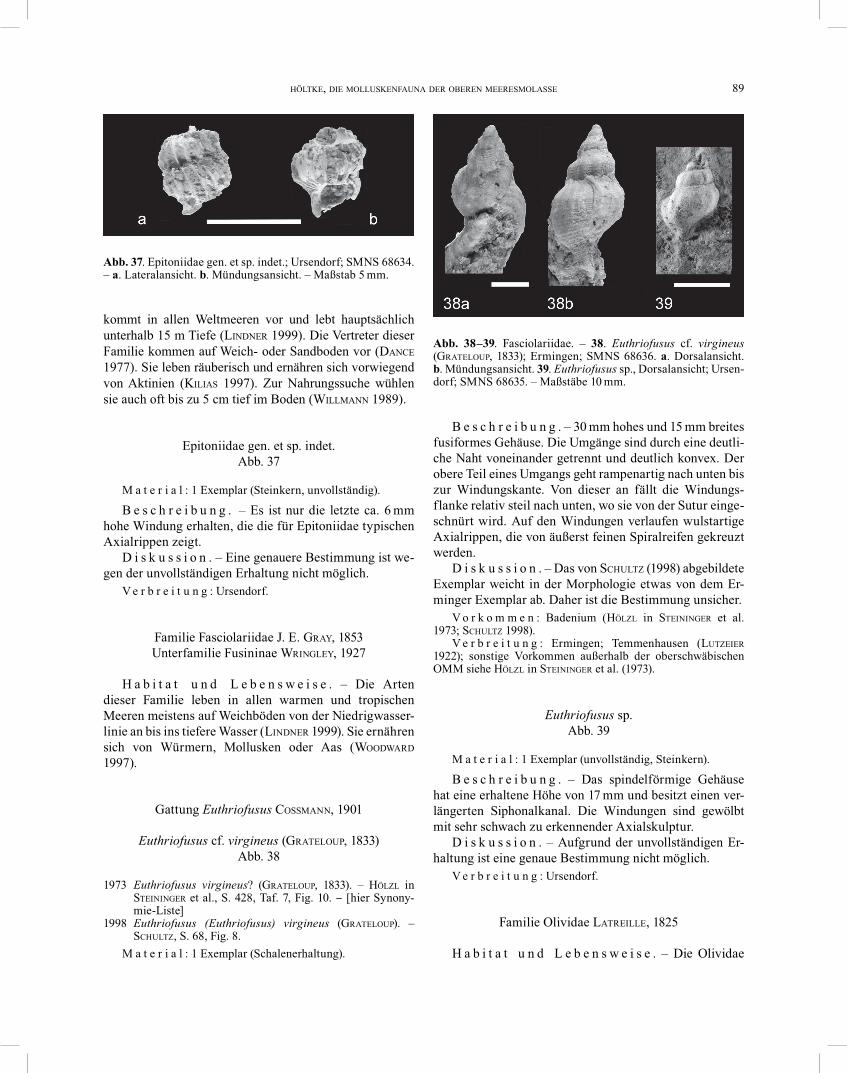
Ficus sp. Abb. 36
M a t e r i a l : 3 Exemplare (Steinkerne, unvollständig).
B e s c h r e i b u n g . – Die Schalen sind bis zu 42 mm hoch und bis zu 25 mm breit. Das Gehäuse hat eine umge-dreht birnenförmige Morphologie. Der letzte Umgang ist stark aufgebläht und umfasst alle früheren Windungen. Das Gewinde ist flach und nicht erhaben. Auf dem Ge-häuse befinden sich Axial- und Spiralreifen, wodurch eine Gitterskulptur entsteht. Der für Ficus typische verlängerte Siphonalkanal ist nicht erhalten. Er lässt sich aber an der Verjüngung des Gehäuses im anterioren Bereich erahnen.
D i s k u s s i o n . – Aufgrund der ungünstigen Erhal-tung ist eine Artbestimmung nicht möglich. Eventuell handelt es sich um Ficus conditus (BRONGNIART, 1823), welche PROBST (1871) für Sießen bei Saulgau sowie LUTZEIER (1922) für Ermingen erwähnen.
V e r b r e i t u n g : Ursendorf.
Familie Epitoniidae BERRY, 1910
H a b i t a t u n d L e b e n s w e i s e . – Die Familie
Abb. 36. Ficus sp.; Ursendorf; SMNS 68633. – a. Dorsalansicht. b. Gewindeansicht. – Maßstab 10 mm.
HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 89
kommt in allen Weltmeeren vor und lebt hauptsächlich unterhalb 15 m Tiefe (LINDNER 1999). Die Vertreter dieser Familie kommen auf Weich- oder Sandboden vor (DANCE 1977). Sie leben räuberisch und ernähren sich vorwiegend von Aktinien (KILIAS 1997). Zur Nahrungssuche wühlen sie auch oft bis zu 5 cm tief im Boden (WILLMANN 1989).
Epitoniidae gen. et sp. indet. Abb. 37
M a t e r i a l : 1 Exemplar (Steinkern, unvollständig).
B e s c h r e i b u n g . – Es ist nur die letzte ca. 6 mm hohe Windung erhalten, die die für Epitoniidae typischen Axialrippen zeigt.
D i s k u s s i o n . – Eine genauere Bestimmung ist we-gen der unvollständigen Erhaltung nicht möglich.
V e r b r e i t u n g : Ursendorf.
Familie Fasciolariidae J. E. GRAY, 1853 Unterfamilie Fusininae WRINGLEY, 1927
H a b i t a t u n d L e b e n s w e i s e . – Die Arten dieser Familie leben in allen warmen und tropischen Meeren meistens auf Weichböden von der Niedrigwasser-linie an bis ins tiefere Wasser (LINDNER 1999). Sie ernähren sich von Würmern, Mollusken oder Aas (WOODWARD 1997).
Gattung Euthriofusus COSSMANN, 1901
Euthriofusus cf. virgineus (GRATELOUP, 1833) Abb. 38
1973 Euthriofusus virgineus? (GRATELOUP, 1833). – HÖLZL in STEININGER et al., S. 428, Taf. 7, Fig. 10. − [hier Synony-mie-Liste]
1998 Euthriofusus (Euthriofusus) virgineus (GRATELOUP). – SCHULTZ, S. 68, Fig. 8.
M a t e r i a l : 1 Exemplar (Schalenerhaltung).
B e s c h r e i b u n g . – 30 mm hohes und 15 mm breites fusiformes Gehäuse. Die Umgänge sind durch eine deutli-che Naht voneinander getrennt und deutlich konvex. Der obere Teil eines Umgangs geht rampenartig nach unten bis zur Windungskante. Von dieser an fällt die Windungs-flanke relativ steil nach unten, wo sie von der Sutur einge-schnürt wird. Auf den Windungen verlaufen wulstartige Axialrippen, die von äußerst feinen Spiralreifen gekreuzt werden.
D i s k u s s i o n . – Das von SCHULTZ (1998) abgebildete Exemplar weicht in der Morphologie etwas von dem Er-minger Exemplar ab. Daher ist die Bestimmung unsicher.
V o r k o m m e n : Badenium (HÖLZL in STEININGER et al. 1973; SCHULTZ 1998).
V e r b r e i t u n g : Ermingen; Temmenhausen (LUTZEIER 1922); sonstige Vorkommen außerhalb der oberschwäbischen OMM siehe HÖLZL in STEININGER et al. (1973).
Euthriofusus sp. Abb. 39
M a t e r i a l : 1 Exemplar (unvollständig, Steinkern).
B e s c h r e i b u n g . – Das spindelförmige Gehäuse hat eine erhaltene Höhe von 17 mm und besitzt einen ver-längerten Siphonalkanal. Die Windungen sind gewölbt mit sehr schwach zu erkennender Axialskulptur.
D i s k u s s i o n . – Aufgrund der unvollständigen Er-haltung ist eine genaue Bestimmung nicht möglich.
V e r b r e i t u n g : Ursendorf.
Familie Olividae LATREILLE, 1825
H a b i t a t u n d L e b e n s w e i s e . – Die Olividae
Abb. 37. Epitoniidae gen. et sp. indet.; Ursendorf; SMNS 68634. – a. Lateralansicht. b. Mündungsansicht. – Maßstab 5 mm.
Abb. 38–39. Fasciolariidae. – 38. Euthriofusus cf. virgineus (GRATELOUP, 1833); Ermingen; SMNS 68636. a. Dorsalansicht. b. Mündungsansicht. 39. Euthriofusus sp., Dorsalansicht; Ursen-dorf; SMNS 68635. – Maßstäbe 10 mm.

90 PALAEODIVERSITY 2, 2009
sind rezent sehr artenreich in allen tropischen Meeren vertreten (MÜLLER 1994). Sie leben gewöhnlich in flachem Wasser, halten sich tagsüber im Sand verborgen und gehen nachts dicht unter der Sandoberfläche auf Nahrungssuche (LINDNER 1999). Sie fressen Aas und andere Weichtiere (ALLEN & STEENE 1995).
Olividae gen. et sp. indet.
M a t e r i a l : 2 Exemplare (Schalenerhaltung, unvollstän-dig).
B e s c h r e i b u n g . – Die Gehäuse haben ein zu-gespitztes Gewinde und sind bis ca. 20 mm hoch und 15 mm breit.
D i s k u s s i o n . – Aufgrund der Erhaltung ist eine genaue Bestimmung nicht möglich. Eventuell handelt es sich um Amalda glandiformis (LAMARCK, 1810), welche laut HARZHAUSER (2002) bereits ab dem Eggenburgium in der Paratethys vorkommt. Auch LUTZEIER (1922) nennt das Vorkommen dieser Art (unter der Bezeichnung Ancillaria glandiformis LAMARCK, 1810) in Ermingen.
V e r b r e i t u n g : Ermingen.
Familie Conidae RAFINESQUE, 1815
Gattung Conus LINNAEUS, 1758
H a b i t a t u n d L e b e n s w e i s e . – Die Gattung kommt in allen warmen Meeren vom Gezeitenbereich bis in eine Tiefe von mehreren 100 m vor (LINDNER 1999). Die Tiere leben auf Korallenriffen, Weich- und Hartböden, unter Steinen und in Löchern und Riffspalten. Conus hat eine toxoglosse Radula mit darin enthaltenen harpunenar-tigen „Giftzähnen“. Beutetiere sind Würmer, Mollusken und sogar kleine Fische (LINDNER 1999).
Conus sp. Abb. 40
M a t e r i a l : 3 Exemplare (Steinkerne, unvollständig).
B e s c h r e i b u n g . – Die Gehäuse sind bis zu 19 mm hoch. Das Gewinde ist erhaben. Die einzelnen Umgänge sind stufenförmig voneinander getrennt.
D i s k u s s i o n . – Aufgrund der ungünstigen Erhal-tung ist eine Bestimmung der Art nicht möglich.
V e r b r e i t u n g : Ursendorf; Heldenfingen, Temmenhau-sen (LUTZEIER 1922).
6. Paläoökologie 6.1. Ermingen
In Ermingen dominieren bei den Gastropoden ein-deutig die Weichbodenbewohner (Tab. 2). Diodora ist die einzige hier vorkommende Gattung, welche auf Hart grund lebt. Die Mehrzahl der Gastropoden-Taxa hat eine endo-benthische Lebensweise. Die dominierende Turritella ist wie auch Xenophora Suspensionsfresser. Drei der vor-kommenden sechs Gastropoden-Gattungen leben als reine Carnivore, während die Olividae als Carnivore sich auch als Aasfresser ernähren. Nur eine Gattung (Diodora) ge-hört zu den Weidegängern.
? Tapes suevica, welche bisher als Tapes helvetica bezeichnet wurde, ist die mit Abstand häufigste Bivalven-Art in Ermingen. Es sind ausschließlich Suspensionsfres-ser unter den Bivalven und keine so genannten Pipettierer (Tab. 1). Auffallend ist auch hier das relativ seltene Vorkommen von reinen Hartbodenbewohnern (Perna, Pseudochama).
Es handelt sich in Ermingen wohl um ein allochthones Vorkommen, da die Bivalven nie doppelklappig erhalten sind und bei den Turritellen eine eindeutige Einregelung zu beobachten ist (RASSER & NEBELSICK 2006). Der Ablagerungsraum war dominiert von Quarzsediment. Erst spätdiagenetisch kam es nach Lösung von Kalkschalen zur Ausfällung von Kalk und Bildung verhärteter Bänke und Platten (RASSER & NEBELSICK 2006). Außer Mollusken und Haizähnen finden sich u. a. Balaniden-Reste (RASSER & NEBELSICK 2006), Rochenzähne (LUTZEIER 1922; BAIER et al. 2004), Knochenfischreste sowie Reste von Trionyx sp. (LUTZEIER 1922).
Die rezenten Vertreter der Gattung Turritella leben, wie schon in Kap. 3 erwähnt, in Kolonien, so z. B. am Golf von Kalifornien, in Neuseeland, Venezuela, Brasilen, etc. (ALLMON 1988). Laut MÜLLER (1993: 60, Fig. 57a, b) findet man am Strand von Vung Tau (Süd-Vietnam) massenhaft Turritellen am Strand, welche seewärts im Litoral ihren Lebensraum haben. Ähnlich könnte es auch in Ermingen der Fall gewesen sein. Wahrscheinlich hat es nahe der Fundstelle eine große Kolonie von Turritelliden gegeben.
Abb. 40. Conus sp.; Ursendorf; SMNS 68638. – a. Mündungsan-sicht. b. Dorsalansicht. – Maßstab 10 mm.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 91
Da das Einfallen der Schichten auf eine Schüttung von Ost nach West hindeutet (RASSER & NEBELSICK 2006), siedelte diese Kolonie vermutlich östlich der Fundstelle. Wahr-scheinlich lebten die Turritellen dort in etwas tieferem Wasser auf Weichsubstrat, da laut ALLMON (1988) die meisten der rezenten Turritellinae auf Weichböden zwi-schen 10 und 200 m Wassertiefe leben. Solche Kolonien kommen häufig in coastal-upwelling-Gebieten vor, da diese reich an Nährstoffen sind (ALLMON 1988). Aufgrund der paläogeographischen und ozeanographischen Situa-tion ist es aber unwahrscheinlich, dass es im miozänen Molassemeer derartige Erscheinungen gab (RASSER & NEBELSICK 2006). Nach RASSER & NEBELSICK (2006) wäre ein durch verstärkten Eintrag vom Festland lokal erhöhter Nährstoffgehalt, der ein erhöhtes Nahrungsaufkommen im Wasser zur Folge hatte, ein geeignetes Erklärungsmo-
dell für dieses Massenvorkommen. Die Ablagerung der Turritellen-Massenakkumulation fand dann aber nicht an einem Strand sondern in flachem Wasser statt, da es keine Hinweise auf eine Strandbildung gibt (RASSER & NEBEL-SICK 2006). Auch das Vorhandensein von vollständig er-haltenen Haifischzähnen (BAIER et al. 2004) spricht eher gegen eine Zusammenschwemmung am Ufer.
Auf einigen Turritellen lassen sich Bohrlöcher beobachten, welche von Naticiden herrühren. Die Nati-ciden lebten offensichtlich ebenfalls im Gebiet der Turri-tellen-Kolonie. Ihre Schalen finden sich allerdings in Er-mingen sehr selten.
Die Mollusken von Ermingen sind rezent hauptsächlich Bewohner von warmen und tropischen Meeren. Es sind keine reinen Kaltwasserarten zu finden. Allerdings hat es laut NORDSIECK (1972) im Miozän oft nicht so eine strenge
Taxon Habitat Lebensweise Ermingen Ursendorf
Anadara Endobenthisch, in Sand oder Schlamm Suspensionsfresser X
Glycymeris Endobenthisch, oberflächennah in Sand oder Schlamm Suspensionsfresser X
Perna Epibenthisch, mit Byssus an Hartsubstrat befestigt Suspensionsfresser X
Lima Epibenthisch, mit Byssus an Steinen festgeheftet Suspensionsfresser X
Ostrea Epibenthisch, festgekittet auf Hartsubstrat Suspensionsfresser X X
Crassostrea Epibenthisch, festgekittet auf Hartsubstrat sowie auch auf Schlammboden Suspensionsfresser X
Aequipecten Epibenthisch, juvenil mit Byssus auf Hartgrund befestigt, adult frei beweglich auf Sand- oder Geröllboden Suspensionsfresser X X
Crassadoma Epibenthisch, juvenil mit Byssus auf Hartgrund befestigt, adult frei beweglich auf Sand- oder Geröllboden Suspensionsfresser X
Amussiopecten Epibenthisch, auf Sand- oder Geröllboden Suspensionsfresser X
Pecten Epibenthisch, auf Sand- oder Geröllboden Suspensionsfresser X
Anomia Epibenthisch, mit Byssus auf Hartgrund befestigt Suspensionsfresser X
Pitar Endobenthisch, oberflächennah Suspensionsfresser X
Paphia Endobenthisch, oberflächennah Suspensionsfresser X
Tapes Endobenthisch, oberflächennah Suspensionsfresser X
Nemocardium Endobenthisch, oberflächennah Suspensionsfresser X
Cerastoderma Endobenthisch, oberflächennah Suspensionsfresser X
Megacardita Epibenthisch, mit Byssus auf Hartgrund befestigt Suspensionsfresser X X
Pseudochama Epibenthisch, auf Hartgrund festzementiert Suspensionsfresser X
Corbula Endobenthisch, in sandigem SchlickNimmt Diatomeen und Bakterien von der Oberfläche auf
X
Pholas Bohrend in Holz, Korallen und weichem Kalkstein Suspensionsfresser X
Tab. 1. Habitat und Lebensweise der Bivalven von Eremingen und Ursendorf. X = Gattung oder Familie eindeutig nachgewiesen.
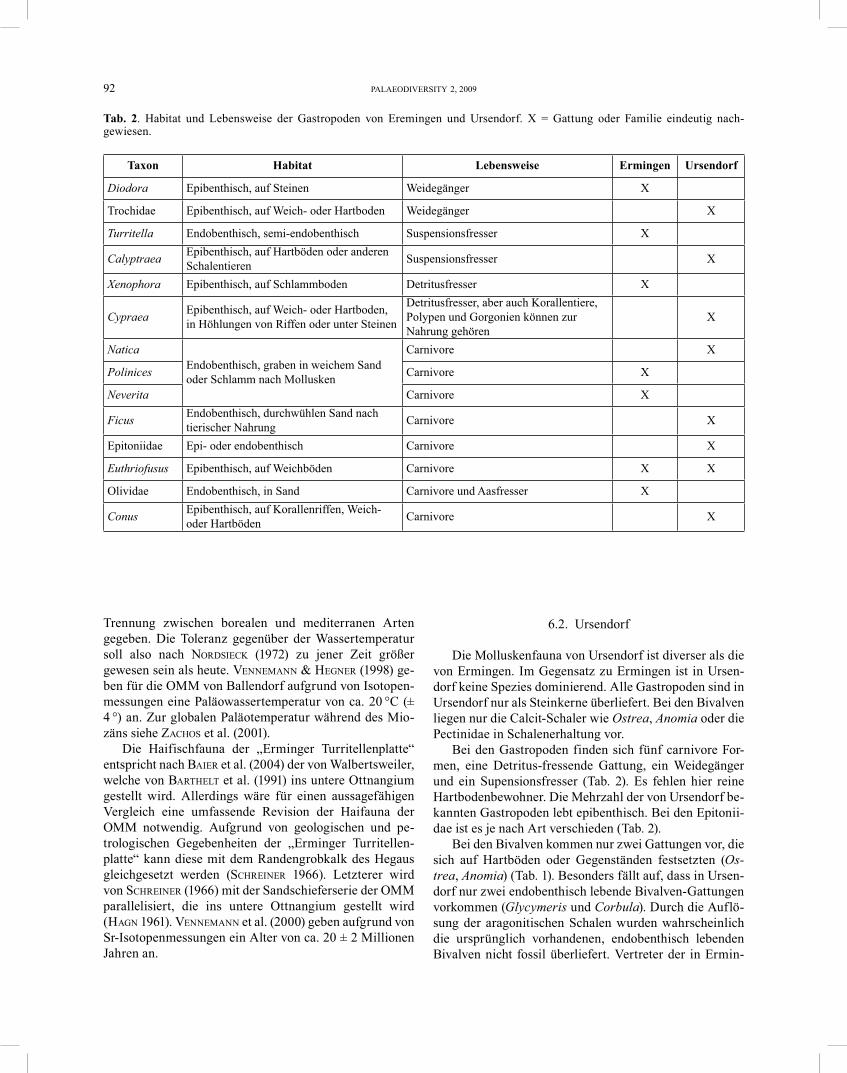
92 PALAEODIVERSITY 2, 2009
Trennung zwischen borealen und mediterranen Arten gegeben. Die Toleranz gegenüber der Wassertemperatur soll also nach NORDSIECK (1972) zu jener Zeit größer gewesen sein als heute. VENNEMANN & HEGNER (1998) ge-ben für die OMM von Ballendorf aufgrund von Isotopen-messungen eine Paläowassertemperatur von ca. 20 °C (± 4 °) an. Zur globalen Paläotemperatur während des Mio-zäns siehe ZACHOS et al. (2001).
Die Haifischfauna der „Erminger Turritellenplatte“ entspricht nach BAIER et al. (2004) der von Walbertsweiler, welche von BARTHELT et al. (1991) ins untere Ottnangium gestellt wird. Allerdings wäre für einen aussagefähigen Vergleich eine umfassende Revision der Haifauna der OMM notwendig. Aufgrund von geologischen und pe-trologischen Gegebenheiten der „Erminger Turritellen-platte“ kann diese mit dem Randengrobkalk des Hegaus gleichgesetzt werden (SCHREINER 1966). Letzterer wird von SCHREINER (1966) mit der Sandschieferserie der OMM parallelisiert, die ins untere Ottnangium gestellt wird (HAGN 1961). VENNEMANN et al. (2000) geben aufgrund von Sr-Isotopenmessungen ein Alter von ca. 20 ± 2 Millionen Jahren an.
6.2. Ursendorf
Die Molluskenfauna von Ursendorf ist diverser als die von Ermingen. Im Gegensatz zu Ermingen ist in Ursen-dorf keine Spezies dominierend. Alle Gastropoden sind in Ursendorf nur als Steinkerne überliefert. Bei den Bivalven liegen nur die Calcit-Schaler wie Ostrea, Anomia oder die Pectinidae in Schalenerhaltung vor.
Bei den Gastropoden finden sich fünf carnivore For-men, eine Detritus-fressende Gattung, ein Weidegänger und ein Supensionsfresser (Tab. 2). Es fehlen hier reine Hartbodenbewohner. Die Mehrzahl der von Ursendorf be-kannten Gastropoden lebt epibenthisch. Bei den Epitonii-dae ist es je nach Art verschieden (Tab. 2).
Bei den Bivalven kommen nur zwei Gattungen vor, die sich auf Hartböden oder Gegenständen festsetzten (Os-trea, Anomia) (Tab. 1). Besonders fällt auf, dass in Ursen-dorf nur zwei endobenthisch lebende Bivalven-Gattungen vorkommen (Glycymeris und Corbula). Durch die Auflö-sung der aragonitischen Schalen wurden wahrscheinlich die ursprünglich vorhandenen, endobenthisch lebenden Bivalven nicht fossil überliefert. Vertreter der in Ermin-
Taxon Habitat Lebensweise Ermingen Ursendorf
Diodora Epibenthisch, auf Steinen Weidegänger X
Trochidae Epibenthisch, auf Weich- oder Hartboden Weidegänger X
Turritella Endobenthisch, semi-endobenthisch Suspensionsfresser X
Calyptraea Epibenthisch, auf Hartböden oder anderen Schalentieren Suspensionsfresser X
Xenophora Epibenthisch, auf Schlammboden Detritusfresser X
Cypraea Epibenthisch, auf Weich- oder Hartboden, in Höhlungen von Riffen oder unter Steinen
Detritusfresser, aber auch Korallentiere, Polypen und Gorgonien können zur Nahrung gehören
X
NaticaEndobenthisch, graben in weichem Sand oder Schlamm nach Mollusken
Carnivore X
Polinices Carnivore X
Neverita Carnivore X
Ficus Endobenthisch, durchwühlen Sand nach tierischer Nahrung Carnivore X
Epitoniidae Epi- oder endobenthisch Carnivore X
Euthriofusus Epibenthisch, auf Weichböden Carnivore X X
Olividae Endobenthisch, in Sand Carnivore und Aasfresser X
Conus Epibenthisch, auf Korallenriffen, Weich- oder Hartböden Carnivore X
Tab. 2. Habitat und Lebensweise der Gastropoden von Eremingen und Ursendorf. X = Gattung oder Familie eindeutig nach-gewiesen.

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 93
gen so häufig vorkommenden Turritelliden und Veneriden konnten bisher nicht eindeutig nachgewiesen werden.
Bekannt ist Ursendorf für seine reichhaltige Bryo-zoenfauna, welche in den ehemaligen Sandgruben gesam-melt und schon von MILLER (1875) beschrieben wurde. Wahrscheinlich hat es in der näheren Umgebung eine aus-gedehnte Bryozoen-Kolonie gegeben. Die Gesamtfauna von Ursendorf ist recht divers, neben den Bryozoen kom-men auch noch irreguläre Seeigel und Balaniden vor. Laut MILLER (1875) gibt es in Ursendorf insgesamt sechs ver-schieden Seeigel-Arten. Zudem gab MILLER (1875) auch den Fund einer Koralle der Gattung Ballanophyllia an. Einige der von MILLER gesammelten Exemplare befinden sich im SMNS. SCHÜTZE (1904) nannte ebenfalls sechs Seeigel-Arten, darunter vier endobenthische und zwei epibenthische. Es sind außerdem Brachiopoden (Terebra-tula sp.) und Reste einer Seekuh gefunden worden (eigene Beobachtungen). Daneben finden sich auch Haizähne in großer Zahl. Laut PFEIL (in BARTHELT et al. 1991) kommt in Ursendorf die Art Carcharoides catticus (PHILIPPI, 1846) auffallend häufig vor. Des Weiteren finden sich noch Odontaspiden, Galeocerdo sp. sowie auch Carcharodon megalodon (AGASSIZ, 1848) (eigene Beobachtungen).
Die großmaßstäblichen Schrägschichtung deutet auf Flachwasserablagerungen im hochenergetischen Milieu hin (BIEG et al. 2007). Eventuell hat es in der näheren Um-gebung auch Bereiche mit geringerer Wasserenergie gege-ben, da MILLER (1875) das Vorkommen von Bryozoen mit verästelten dünnen Zweigen beschreibt. Anhand der Mol-lusken ist es schwer, etwas über die damals herrschende Wasserenergie zu sagen. In der Literatur über die rezenten Vertreter finden sich dazu keine expliziten Angaben. Es handelt sich in Ursendorf wohl ebenfalls um ein rein al-lochthones Vorkommen, da sich zum einen weder dop-pelklappig erhaltene Bivalven finden zum anderen alle Organismenreste mehr oder weniger ausgeprägte Abra-sionsspuren zeigen. So sind die Haifischzähne oftmals stak abgerollt (eigene Beobachtungen). Die Anzahl der Organismen wie Seeigel, Bryozoen, Bivalven und einige Gastropoden-Arten, welche von Schwebstoffen bezie-hungs weise Mikroorganismen leben, lässt auf sehr nähr-stoff haltiges Wasser schließen. Zudem ist von einem voll-marinen Milieu auszugehen, da Hinweise auf brackische Verhältnisse fehlen (eigene Beobachtungen). Über die Knochenfischfauna von Ursendorf gibt es bisher noch keine Untersuchungen.
Stratigraphisch lässt sich Ursendorf wohl ab dem un-teren Ottnangium einstufen, denn die Transgression des oberen Eggenburgium ist in diesem Gebiet nicht bis in den Bereich der ungefalteten Vorlandmolasse vorgedrungen. Dies erfolgte erst im unteren Ottnangium mit den Heiden-löcher Schichten, mit denen der erst Zyklus der OMM be-ginnt (WENGER 1987).
7. Literatur
ABBOT, R. T. & DANCE, S. (1998): Compendium of Seashells. 411 S.; El Cajon Californien (Odyssey Publishing).
ALLEN, G. R. & STEENE, R. (1995): Riff-Führer. 378 S.; Wies-baden (Christa Hemmen).
ALLMON, W. D. (1988): Ecology of Recent Turritellinae Gastro-pods (Prosobranchia, Turritellidae): Current Knowledge and Palaeontological Implications. – PALAIOS, 3: 259–284.
BAIER, J., SCHMITT, K.-H. & MICK, R. (2004): Notizen zur unter-miozänen Hai- und Rochenfauna der Erminger Turritellen-platte (Mittlere Schwäbische Alb, SW-Deutschland). – Jah-resbericht und Mitteilungen des oberrheinischen geologi-schen Vereins, Neue Folge, 86: 361–371.
BALUK, W. (1995): Middle Miocene (Badenian) Gastropods from Korytnica, Poland. Part 2. – Acta Geologica Polonica, 45 (3–4): 153–255.
BARTHELT, D., FEJFAR, O., PFEIL, F. H. & UNGER, E. (1991): Noti-zen zu einem Profil der Selachier-Fundstelle Walbertsweiler im Bereich der miozänen Oberen Meeresmolasse Süd-deutschlands. – Münchner Geowissenschaftliche Abhand-lungen, 19: 195–208.
BASTEROT, B. DE (1825): Mémoire de géologique sur les environs de Bordeaux. Première partie comprenant les observations générales sur les mollusques fossiles, et la description parti-culière de ceux qu on rencontre dans ce bassin. – Mémoires de la Société d´Histoire Naturelle de Paris, 2: 1–100.
BEURLEN, K. & LICHTER, G. (2000): Fossilien. 287 S.; München (Orbis).
BEURLEN, K., GALL, H. & SCHAIRER, G. (1981): Die Alb und ihre Fossilien. 224 S.; Stuttgart (Franckh-Kosmos).
BIEG, U. (2005): Palaeooceanographic modeling in global and regional scale: An example from the Burdigalian Seaway Upper Marine Molasse (Early Miocene). 108 S.; Dissertation an der Geowissenschaftlichen Fakultät der Eberhard-Karls-Universität Tübingen.
BIEG, U., NEBELSICK, J. & RASSER, M. (2007): North Alpine Fore-land Basin (Upper Marine Molasse) of Southwest Germany: Sedimentology, Stratigraphy and Palaeontology. – Geo Alp, 4: 149–158.
COSSMANN, M. & PEYROT, A. (1921): Cochologie Neogenique de l Aquitaine. – Actes de la Société Linnéenne de Bordeaux, 73: 5–321.
DANCE, P. (1977): Das große Buch der Meeresmuscheln. 304 S.; Stuttgart (Eugen Ulmer).
DANCE, P. (1998): Muscheln und Schnecken. 256 S.; Berlin (Ura-nia-Ravensburger).
ENGEL, T. (1908): Geognostischer Wegweiser durch Württem-berg. 645 S.; Stuttgart (E. Schweizerbart sche Verlagshand-lung).
ENGEL, T. (1911): Geologischer Exkursionsführer durch Würt-temberg. 182 S.; Stuttgart (E. Schweizerbart sche Verlags-handlung).
ENTROP, B. (1977): Muscheln und Schnecken an Europas Küsten. 72 S.; Stuttgart (Franckh-Kosmos).
ETTER, W. (1994): Palökologie. 294 S.; Basel (Birkhäuser).FECHTER, R., GRAU, J. & REICHHOLF, J. (1985): Lebensraum Küste.
287 S.; München (Mosaik).FRAAS, E. (1910): Der Petrefaktensammler. 249 S.; Stuttgart (K.
G. Lutz).GALL, H. (1975): Der III. Zyklus der Oberen Meeresmolasse
(Helvet) am Südrand der Schwäbisch-Fränkischen Alb. – Mitteilungen der Bayerischen Staatssammlung für Paläon-tologie und historische Geolgie, 15: 179–205.
GEYER, M. (2001): Geologisch-naturkundliche Streifzüge im Raum Ulm. 48 S., 1 Wanderkarte 1 : 50 000; Ulm (Natur- kundliches Bildungszentrum).

94 PALAEODIVERSITY 2, 2009
GEYER, O. F. & GWINNER, M. P. (1979): Die schwäbische Alb und ihr Vorland. – Sammlung Geologische Führer, 67: 271 S.
GEYER, O. F. & GWINNER, M. P. (1991): Geologie von Baden-Württemberg. 482 S.; Stuttgart (Schweizerbart sche Ver-lagsbuchhandlung).
GEYER, O. F., SCHOBER, T. & GEYER, M. (2003): Die Hochrhein-Regionen zwischen Bodensee und Basel. – Sammlung Geo-logische Führer, 94: 526 S.
HAGN, H. (1961): Die Gliederung der Oberen Meeresmolasse nördlich vom Überlinger See (Bodensee) in mikropaläon-tologischer Sicht. – Jahreshefte des Geologischen Landes-amtes in Baden-Württemberg, 5: 293–321.
HARZHAUSER, M. (2002): Marine und brachyhaline Gastropoden aus dem Karpatium des Korneuburger Beckens und der Kreuzstettener Bucht (Österreich, Untermiozän). – Beiträge zur Paläontologie, 27: 61–159.
HÖLZL, O. (1958): Die Mollusken-Fauna des oberbayrischen Bur-digals. – Geologica Bavarica, 38: 1–348.
HOFMANN, F. (1957): Untersuchungen in der subalpinen und mit-telländischen Molasse der Ostschweiz. – Eclogae Geologi-cae Helvetiae, 51: 371–377.
KILIAS, R. (1997): Lexikon marine Muscheln und Schnecken. 340 S.; Stuttgart (Eugen Ulmer).
KUHLEMANN, J. & KEMPF, O. (2002): Post-Eocene evolution of the North Alpine Foreland Basin and its response to Alpine tec-tonics. – Sedimentary Geology, 152: 45–78.
LINDNER, G. (1999): Muscheln und Schnecken der Weltmeere. 319 S.; München (BLV).
LINDNER, G. (2000): Muscheln und Schnecken. 95 S.; München (BLV).
LORENZ, F. & HUBERT, A. (1993): A guide to worldwide Cowries. 571 S.; Wiesbaden (Christa Hemmen).
LOZOUET, P., LESPORT, J.-F. & RENARD, P. (2001): Révision des Gastropode (Mollusca) du Stratotype de l Aquitanien (Mio-céne inf.): site de Saucats „Lariey“, Gironde, France. – Coss-manniana, h. s. 3: 1–189.
LUTERBACHER, H. (1997): Stratigraphy and facies evolution of a typical foreland basin – the Tertiary Molasse Basin (Lake Constance Area and Allgäu). – Gaea heidelbergensis, 4: 123–140.
LUTZEIER, H. (1922): Beiträge zur Kenntnis der Meeresmolasse in der Ulmer Gegend. – Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Beilagen-Band, Abt. B, 46: 117–180.
MAYER, K. (1853): Verzeichnis der in der marinen Molasse der schweizerisch-schwäbischen Hochfläche enthaltenen fossi-len Mollusken. – Mitteilungen der naturforschenden Gesell-schaft Bern, 1853: 76–106.
MAYER, K. (1871): Description de coquilles fossiles des terrains tertiaires superieurs. – Journal Conchylia, 24: 168–180.
MENNIG, M. & HENDRICH, A. (Hrsg.) (2005): Erläuterungen zur Stratigraphischen Tabelle von Deutschland 2005 (ESTD 2005). – Newsletter Stratigraphy, 41 (1/3): 405 S.
MILLER, K. (1875): Die Bryozoenfauna von Ursendorf. – Jah-reshefte des Vereins für vaterländische Naturkunde, 31: 82–84.
MÜLLER, A. H. (1993): Handbuch der Paläozoologie, Band 2, Teil 1. 685 S.; Jena (Gustav Fischer).
MÜLLER, A. H. (1994): Lehrbuch der Paläozoologie, Band 2, Teil 2. 618 S.; Jena (Gustav Fischer).
NORDSIECK, F. (1972): Die miozäne Molluskenfauna von Miste-Winterswijk (Hemmoor). 187 S.; Stuttgart (Gustav Fischer).
PFEIL, F. H. (1991): Haie und Rochen aus Walbertsweiler. – In: BARTHELT, D., FEJFAR, O., PFEIL, F. H. & UNGER, E.: Notizen zu einem Profil der Selachier-Fundstelle Walbertsweiler im Bereich der miozänen Oberen Meeresmolasse Süd-
deutschlands. – Münchner Geowissenschaftliche Abhand-lungen, 19: 195–208.
PFISTER, T. & WEGMÜLLER, U. (1994): Bivalven aus der Oberen Meeresmolasse bei Bern. – Beschreibung, Vergleich und Verbreitung der Bivalven-Arten aus den Belpbergschichten (Obere Meeresmolasse, mittleres Burdigalien) in der Umge-bung von Bern, Schweiz. 1. Teil. – Eclogae Geologicae Hel-vetiae, 87: 895–973.
PFISTER, T. & WEGMÜLLER, U. (1998): Bivalven aus der Oberen Meeresmolasse bei Bern. – Beschreibung, Vergleich und Verbreitung der Bivalven-Arten aus den Belpbergschichten (Obere Meeresmolasse, mittleres Burdigalien) in der Umge-bung von Bern, Schweiz. 2. Teil. – Eclogae Geologicae Hel-vetiae, 91: 457–491.
PFISTER, T. & WEGMÜLLER, U. (2000): Bivalven aus der Oberen Meeresmolasse bei Bern. – Beschreibung, Vergleich und Verbreitung der Bivalven-Arten aus den Belpbergschichten (Obere Meeresmolasse, mittleres Burdigalien) in der Umge-bung von Bern, Schweiz. 4. Teil. – Eclogae Geologicae Hel-vetiae, 93: 445–470.
PFISTER, T. & WEGMÜLLER, U. (2001): Bivalven aus der Oberen Meeresmolasse bei Bern. – Beschreibung, Vergleich und Verbreitung der Bivalven-Arten aus den Belpbergschichten (Obere Meeresmolasse, mittleres Burdigalien) in der Umge-bung von Bern, Schweiz. 5. Teil. – Eclogae Geologicae Hel-vetiae, 94: 399–426.
POPPE, G. T. & GOTO, Y. (1991): European Seashells, Vol. 1. 352 S.; Wiesbaden (Christa Hemmen).
POPPE, G. T. & GOTO, Y. (1993): European Seashells, Vol. 2. 221 S.; Wiesbaden (Christa Hemmen).
PROBST, J. (1871): Fossile Meeres- und Brackwasserconchylien aus der Gegend von Biberach. – Jahreshefte des Vereins für vaterländische Naturkunde, 27: 111–118.
QUEDENS, G. (1990): Strand und Wattenmeer. 127 S.; München (BLV).
QUENSTEDT, F. A. (1867): Handbuch der Petrefaktenkunde. 981 S.; Tübingen (Laupp).
QUENSTEDT, F. A. (1882): Handbuch der Petrefaktenkunde. Teil 2: 609–1239; Tübingen (Laupp).
QUENSTEDT, F. A. (1884): Petrefactenkunde Deutschlands. Gas-teropoden. Teil 1. 440 S.; Leipzig (Fues).
RASSER, M. & NEBELSICK, J. (2006): Die Erminger Turritellen-platte. – Fossilien, 23 (4): 220–224.
RIEDL, R. (1963): Fauna und Flora der Adria. 640 S.; Hamburg und Berlin (Paul Parey).
ROSENDAHL, W. (2000): Im Wellenschlag des Tertiärs. – Das Heldenfinger Kliff. – Fossilien, 17 (3): 177–180.
SCHAFFER, F. X. (1910): Das Miozän von Eggenburg. Die Fauna der 1. Mediterranstufe des Wiener Beckens und die geolo-gischen Verhältnisse des Manhartsberges in Niederöster-reich. – Abhandlungen der Kaiserlich-königlichen geologi-schen Reichsanstalt, 22: 129–183.
SCHREINER, A. (1966): Zur Stratigraphie der Oberen Meeresmo-lasse zwischen der Oberen Donau und dem Überlinger See (Baden-Württemberg). – Jahresbericht und Mitteilungen des oberrheinischen geologischen Vereins, Neue Folge, 48: 91–104.
SCHREINER, A. & LUTERBACHER, H. (1999): Die Molasse zwischen Blumberg und Überlingen (Exkursion am 9. April 1999). – Jahresberichte und Mitteilungen des oberrheinischen geolo-gischen Vereins, Neue Folge, 81: 171–181.
SCHÜTZE, E. (1904): Die Fauna der schwäbischen Meeresmolas-se. I. Teil: Spongien und Echinodermen. – Jahreshefte des Vereins für vaterländische Naturkunde, 60: 147–188.
SCHULTZ, O. (1998): Tertiärfossilien Österreichs. 159 S.; Freiburg (Goldschneck).
SCHULTZ, O. (2001): Bivalvia neogenica (Nuculacea-Unionacea).

HÖLTKE, DIE MOLLUSKENFAUNA DER OBEREN MEERESMOLASSE 95
– In: PILLER, E. (Hrsg.): Catalogus Fossilium Austriae. Band 1/Teil 1: 1–379; Wien (Österreichische Akademie der Wis-senschaften).
SCHULTZ, O. (2003): Bivalvia neogenica (Lucinoidea-Mactroi-dea). – In: PILLER, E. (Hrsg.): Catalogus Fossilium Austriae. Band 1/Teil 2: 381–690; Wien (Österreichische Akademie der Wissenschaften).
SCHULTZ, O. (2005): Bivalvia neogenica (Solenoidea-Cla-vagelloidea). – In: PILLER, E. (Hrsg.): Catalogus Fossilium Austriae. Band 1/Teil 3: 691–1212; Wien (Österreichische Akademie der Wissenschaften).
SIEBER, R. (1960): Die miozänen Turritellidae und Mathildidae Österreichs. – Mitteilungen der Geologischen Gesellschaft Wien, 51: 229–281.
STEININGER, F., CTYROKY, P., ONDREJICKOVA, A. & SENES, J. (1971): Die Mollusken der Eggenburger Schichtengruppe. – In: STEININGER, F. & SENES, J. (Hrsg.): Chronostratigraphie und Neostratotypen, 2: M1 Eggenburgien. Die Eggenburger Schichtengruppe und ihr Stratotypus: 356–591; Bratislava (Vydavatel stvo Slovenskej akadémie vied).
STEININGER, F., CTYROKY, P., ONDREJICKOVA, A. & SENES, J. (1973): Die Molluskenfauna des Ottnangien. – In: PAPP, A., RÖGL, F. & SENES, J. (Hrsg.): Chronostratigraphie und Neostrato-typen. Miozän der zentralen Paratethys, 3: M2 Ottnangien. Die Innviertler, Salgótarjáner, Bántapusztaer Schichtgruppe und die Rzehakia Formation: 380–615; Bratislava (Vydavatel stvo Slovenskej akadémie vied).
STEININGER, F., CTYROKY, P., ONDREJICKOVA, A. & SENES, J. (1978): Die Molluskenfauna des Badenien. – In: PAPP, A., RÖGL, F. & SENES, J. (Hrsg.): Chronostratigraphie und Neostratotypen. Miozän der zentralen Paratethys, 4: M4 Badenien. Die Inn-viertler, Salgótarjáner, Bántapusztaer Schichtgruppeund die Rzehakia Formation: 329–360; Bratislava (Vydavatel stvo Slovenskej akadémie vied).
SUESS, F. E. (1891): Beobachtungen über den Schlier in Ober-österreich und Bayern. – Annalen des naturhistorischen Hofmuseums, 6: 407–429.
VENNEMANN, T. & HEGNER, E. (1998): Oxygen, strontium, and neodymium isotope composition of fossil shark teeth as a proxy for the palaeoceanography and palaeoclimatolo-gy of the Miocene Northern Alpine Paratethys. – Palaeo-geography, Palaeoclimatology, Palaeoecology, 142: 107–121.
VENNEMANN, T., HEGNER, H. & REIF, W.-E. (2000): Ökosystem-, Meerwasser- und Klimaentwicklung des nordalpinen Mo-lassebeckens in Abhängigkeit der alpinen Hebung. – In: MOSBRUGGER, V. (Hrsg.): Sonderforschungsbereich (SFB) 275, Bericht 1998–2000, Band 1: 177–199; Tübingen.
VOORTHUYSEN, J. H. VAN (1944): Miozäne Gastropoden aus dem Peelgebiet (Niederlande) (Rissoidae-Muricidae, nach Zittels Einteilung, 1924). – Mededeelingen van de geologische Stichting, Serie C-IV-1, 5: 65 S.
WAGNER, H. P. (1990): Review of the European Pectinidae. – Vita Marina, 41 (1): 1–48.
WENGER, W. F. (1987): Die Basis der Oberen Meeresmolasse im westlichen Oberbayern, am Überlinger See, in Vorarlberg und St. Gallen. – Mitteilungen der bayrischen Staats samm-lung für Paläontologie und historische Geologie, 27: 159–174.
WESSELINGH, F. P., VERVOENEN, M. & VAN NIEULANDE, F. A. D. (2001): On the identity of Mytilus edulis forma giganteus WOOD, 1874 (Pliocene, North Sea Basin), with implications for the generic assignment of other European Cainozoic mytilid bivalves. – Cainozoic Research, 1–2: 121–128.
WILLMANN, R. (1989): Muscheln und Schnecken der Nord- und Ostsee. 310 S.; Melsungen (Neumann-Neudamm).
WOODWARD, F. (1997): Muscheln. 80 S.; Köln (Könemann).WYE, K. R. (2003): The shell handbook. 256 S.; Devon (D & S
Books Ltd).ZACHOS, J., PAGANI, M., SLOAN, L., THOMAS, E. & BILLUPS, K.
(2001): Trendes, Rhythms, and Aberrations in Global Cli-mate 65 Ma to Present. – Science, 292: 686–693.
ZIETEN, C. H. VON (1833): Die Versteinerungen Württembergs. 12. Heft: 89–102; Stuttgart (Schweizerbart).
Anschrift des Verfassers:Dipl. Geol. OLAF HÖLTKE, Staatliches Museum für Naturkunde, Rosenstein 1, 70191 Stuttgart, DeutschlandE-Mail: [email protected]
Manuskript erhalten: 26.5.2006, angenommen: 19.3.2009.
























![Die Physiologische Inkongruenz des oberen Sprunggelenkes ... · Sprunggelenk, ebenfalls eine Inkongruenz der Kontaktflächen [90] besteht. In dieser Arbeit soll über die Darstellung](https://img.dokumen.tips/doc/110x75/605bea98c07e933b233aee94/die-physiologische-inkongruenz-des-oberen-sprunggelenkes-sprunggelenk-ebenfalls.jpg)





