Embed Size (px)
Citation preview

news and views
NATURE CELL BIOLOGY VOL 5 JANUARY 2003 www.nature.com/naturecellbiology 9
Programmed cell death (apoptosis) isused to eliminate superfluous, aged,injured or infected cells in diverse bio-
logical settings1. During apoptosis, cells aredismantled from within and display plasma
membrane alterations that attract the atten-tion of phagocytic cells, thereby ensuringremoval of the dying cell with the minimumof disturbance to its neighbours. A key eventin many, if not all, instances of apoptosis is
the activation of caspases, a family of cys-teine proteases. Active caspases orchestrateapoptosis through limited proteolysis ofapproximately 500 cellular proteins2. Amajor pathway to caspase activation and
Defying death: showing Bcl-2 the wayhome
Colin Adrain, Emma M. Creagh and Seamus J. Martin
Bcl-2 and Bcl-xL perform important functions in cell life or death decisions by setting the threshold for acti-vation of the programmed cell death machinery. FKBP38, a member of the immunophilin family, has nowbeen identified as a new Bcl-2/Bcl-xL-binding partner. Furthermore, FKBP38 seems to be crucial in targetingBcl-2/Bcl-xL to the correct cellular locations.
The primary aim of HIV at initial infection of a cell is to rapid-ly deliver its RNA genome to the nucleus for integration into thehost DNA. To get there, its large virion particles must first nav-igate through the cytoplasm towards the nuclear pore. But howdoes it get there before the cell is alerted to its presence? Hopeand colleagues (McDonald, D. et al. J. Cell Biol., 159, 441–452(2002)) have visualized the movement of individual HIV parti-cles in live cells and find that in common with other subvertiveviruses, HIV does this by ‘hitchhiking a ride’ with one of thecell’s own motors, dynein.
To follow the HIV virions’ movement, the authors fused greenfluorescent protein (GFP) to one of the viral accessory proteins,Vpr. They then followed the movement of GFP-labelled virionsthrough the cytoplasm of infected cells and showed that theymove along a curvilinear path, showing significant associationwith the microtubule network, but not actin. To address the sig-nificance of this association, they looked at the consequences ofdisrupting the actin or microtubule networks for particle trans-port. Surprisingly, a complete block in particle transport occurredonly when both the actin and microtubule networks were dis-rupted, suggesting that both are important for HIV movement.
The authors then went on to demonstrate unequivocally thatthe viral particles do track along microtubules (see figure). Theyused real-time imaging to track the movement of particles alongfluorescently labelled tubulin, and they also showed using correla-tive electron microscopy (in which a cell is first imaged on a fluo-rescence microscope and then processed for electron microscopy),that single viral particles are attached to microtubules.
To investigate the functional significance of the microtubuleassociation, they injected antibodies to disrupt the function ofthe microtubule motor, dynein. Importantly, they found thatblocking dynein function alone prevented movement of particlesaway from the periphery of the nucleus.
So how do we resolve this essential role of dynein with theresults of the drug treatments, which suggest that actin and micro-tubules function redundantly during HIV transport? Previouswork has also suggested that both actin and microtubules are
important for HIV infectivity (J. Exp. Med., 188, 2113–2125, 1998).Therefore, the authors favour a model where actin is required forshort-range movement of the HIV complexes at the periphery ofthe cell, and perhaps then to load particles onto microtubules,which mediate the long-range transport of particles towards thenucleus. It will be important to demonstrate association of the HIVparticles with actin at the periphery to support this model.However, this idea is consistent with a growing list of crosstalkbetween the actin and microtubule networks and raises curiosityabout the mechanism by which HIV may be transported alongactin and then handed over to dynein for microtubule transport.Needless to say, these tantalizing first images of the path taken byHIV will spark further studies into these and other key questions.
ALISON SCHULDT
The Hitchhiker’s Guide to HIV
Figure 1 Fluorescent microscopy image of an HIV reverse-tran-scription complex (RTC) on a microtubule (white arrow).Microtubules are shown in blue, incorporated fluorescentnucleotides in red and GFP–Vpr in green. A close-up image ofthe RTC is shown in the inset (lower right).
© 2003 Nature Publishing Group
news and views
NATURE CELL BIOLOGY VOL 5 JANUARY 2003 www.nature.com/naturecellbiology10
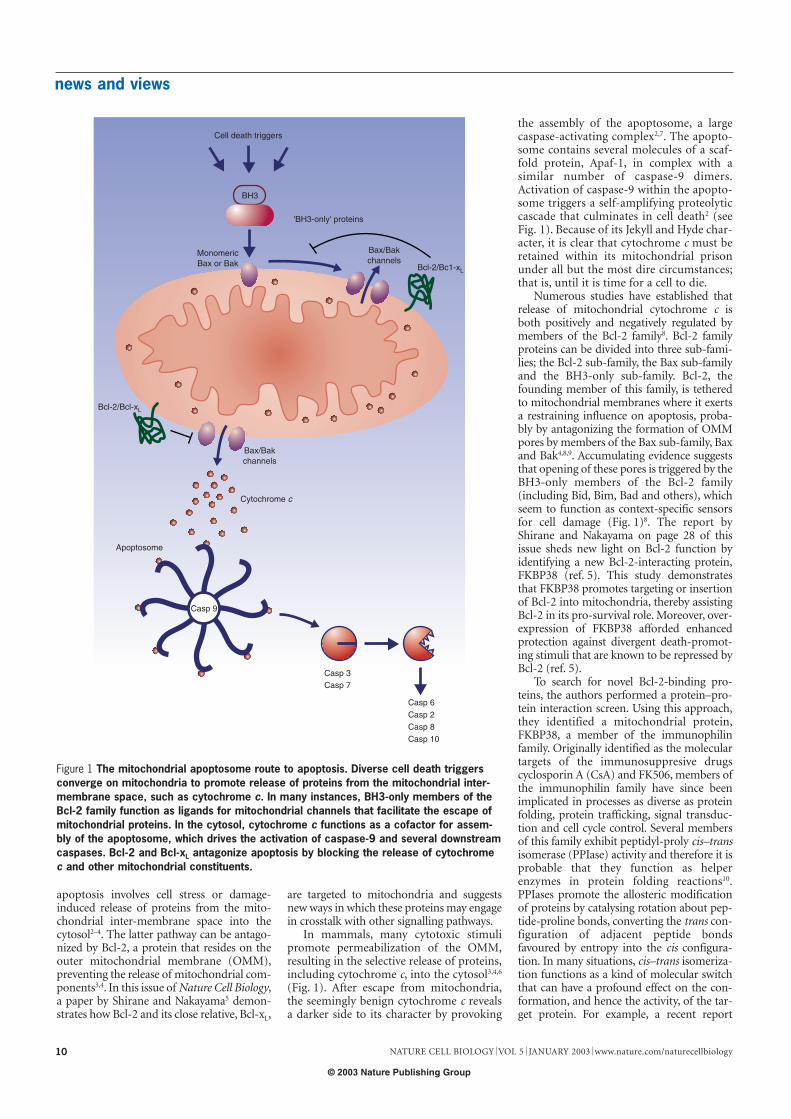
apoptosis involves cell stress or damage-induced release of proteins from the mito-chondrial inter-membrane space into thecytosol2–4. The latter pathway can be antago-nized by Bcl-2, a protein that resides on theouter mitochondrial membrane (OMM),preventing the release of mitochondrial com-ponents3,4. In this issue of Nature Cell Biology,a paper by Shirane and Nakayama5 demon-strates how Bcl-2 and its close relative, Bcl-xL,
are targeted to mitochondria and suggestsnew ways in which these proteins may engagein crosstalk with other signalling pathways.
In mammals, many cytotoxic stimulipromote permeabilization of the OMM,resulting in the selective release of proteins,including cytochrome c, into the cytosol3,4,6
(Fig. 1). After escape from mitochondria,the seemingly benign cytochrome c revealsa darker side to its character by provoking
the assembly of the apoptosome, a largecaspase-activating complex2,7. The apopto-some contains several molecules of a scaf-fold protein, Apaf-1, in complex with asimilar number of caspase-9 dimers.Activation of caspase-9 within the apopto-some triggers a self-amplifying proteolyticcascade that culminates in cell death2 (seeFig. 1). Because of its Jekyll and Hyde char-acter, it is clear that cytochrome c must beretained within its mitochondrial prisonunder all but the most dire circumstances;that is, until it is time for a cell to die.
Numerous studies have established thatrelease of mitochondrial cytochrome c isboth positively and negatively regulated bymembers of the Bcl-2 family8. Bcl-2 familyproteins can be divided into three sub-fami-lies; the Bcl-2 sub-family, the Bax sub-familyand the BH3-only sub-family. Bcl-2, thefounding member of this family, is tetheredto mitochondrial membranes where it exertsa restraining influence on apoptosis, proba-bly by antagonizing the formation of OMMpores by members of the Bax sub-family, Baxand Bak4,8,9. Accumulating evidence suggeststhat opening of these pores is triggered by theBH3-only members of the Bcl-2 family(including Bid, Bim, Bad and others), whichseem to function as context-specific sensorsfor cell damage (Fig. 1)8. The report byShirane and Nakayama on page 28 of thisissue sheds new light on Bcl-2 function byidentifying a new Bcl-2-interacting protein,FKBP38 (ref. 5). This study demonstratesthat FKBP38 promotes targeting or insertionof Bcl-2 into mitochondria, thereby assistingBcl-2 in its pro-survival role. Moreover, over-expression of FKBP38 afforded enhancedprotection against divergent death-promot-ing stimuli that are known to be repressed byBcl-2 (ref. 5).
To search for novel Bcl-2-binding pro-teins, the authors performed a protein–pro-tein interaction screen. Using this approach,they identified a mitochondrial protein,FKBP38, a member of the immunophilinfamily. Originally identified as the moleculartargets of the immunosuppresive drugscyclosporin A (CsA) and FK506, members ofthe immunophilin family have since beenimplicated in processes as diverse as proteinfolding, protein trafficking, signal transduc-tion and cell cycle control. Several membersof this family exhibit peptidyl-proly cis–transisomerase (PPIase) activity and therefore it isprobable that they function as helperenzymes in protein folding reactions10.PPIases promote the allosteric modificationof proteins by catalysing rotation about pep-tide-proline bonds, converting the trans con-figuration of adjacent peptide bondsfavoured by entropy into the cis configura-tion. In many situations, cis–trans isomeriza-tion functions as a kind of molecular switchthat can have a profound effect on the con-formation, and hence the activity, of the tar-get protein. For example, a recent report
Cell death triggers
BH3
'BH3-only' proteins
MonomericBax or Bak
Bax/Bakchannels
Bax/Bakchannels
Bcl-2/Bc1-xL
Bcl-2/Bcl-xL
Cytochrome c
Apoptosome
Casp 3
Casp 6
Casp 2
Casp 8
Casp 10
Casp 7
Casp 9
Figure 1 The mitochondrial apoptosome route to apoptosis. Diverse cell death triggersconverge on mitochondria to promote release of proteins from the mitochondrial inter-membrane space, such as cytochrome c. In many instances, BH3-only members of theBcl-2 family function as ligands for mitochondrial channels that facilitate the escape ofmitochondrial proteins. In the cytosol, cytochrome c functions as a cofactor for assem-bly of the apoptosome, which drives the activation of caspase-9 and several downstreamcaspases. Bcl-2 and Bcl-xL antagonize apoptosis by blocking the release of cytochromec and other mitochondrial constituents.
© 2003 Nature Publishing Group

news and views
NATURE CELL BIOLOGY VOL 5 JANUARY 2003 www.nature.com/naturecellbiology 11
demonstrated that the PPIase Pin-1 can reg-ulate stability of the p53 tumour suppressorprotein by catalysing a conformationalchange that renders p53 incapable of inter-acting with Mdm2 (ref. 11).
Thus, one possibility is that FKBP38 func-tions as a PPIase for Bcl-2 and induces a con-formational change that targets, or promotesinsertion of, Bcl-2 into the OMM (Fig. 2).However, FKBP38, uniquely among theimmunophilin family, seems to lack residueswithin the conserved FK506-binding domainthat may be critically required for PPIaseactivity5. Coupled with a previous report thatfailed to detect any PPIase activity associatedwith recombinant FKBP38, these studies sug-gest that it is improbable that FKBP38 func-tions as a cis–trans-isomerase for Bcl-2,although we cannot fully exclude this possi-bility at present.
Irrespective of whether FKBP38 possessesPPIase activity, it may still function as amolecular chaperone, targeting Bcl-2/Bcl-xL
to mitochondria and/or promoting correctfolding. In support of this, Shirane andNakayama show that expression of anFKBP38 mutant — lacking the carboxy-ter-minal transmembrane domain necessary forinsertion into mitochondria — disrupted thenormal mitochondrial distribution of co-expressed Bcl-2 and Bcl-xL. Moreover, byredirecting FKBP38 from mitochondria tothe plasma membrane, the authors alsoobserved movement of Bcl-2 and Bcl-xL tothe same location5. In the reciprocal experi-ment, overexpressed wild-type FKBP38 redi-rected a plasma membrane-targeted Bcl-xL
mutant to mitochondria. Furthermore,depletion of endogenous FKBP38 by RNAinterference converted the normal mitochon-drial staining pattern of Bcl-2/Bcl-xL to amore cytosolic distribution5. Taken together,these data provide compelling evidence tosuggest that Bcl-2/Bcl-xL cannot sponta-neously insert into the OMM, but require thehelping hand of FKBP38 to get there.
Another twist in the tale is thatFKBP38 was found to function as a con-stitutive inhibitor of the calcium-activat-ed Ser/Thr phosphatase, calcineurin5.Other immunophilins typically functionas calcineurin inhibitors only when incomplex with FK506 or CsA. However,FKBP38 inhibits calcineurin phosphataseactivity in an FK506-independent man-ner, a unique feature among members ofthe immunophilin family5.
Calcineurin has previously been implicat-ed in the regulation of a number of cell deathpathways, most notably, dephosphorylationof the BH3-only molecule, Bad12, as well asthrough suppression of NF-AT-mediatedtranscription of Fas ligand13. Thus, given theability of FKBP38 to function as a calcineurininhibitor, it is possible that FKBP38 may raisethe threshold for entry into apoptosis, notthrough targeting Bcl-2/Bcl-xL to mitochon-dria, but by disabling calcineurin-regulated
signalling pathways (Fig. 2). However, therange of pro-apoptotic stimuli that can berepressed through overexpression of FKBP38is broader than those modulated by cal-cineurin5. In agreement with this, FK506failed to block apoptosis in several contextsthat were clearly repressed by overexpressionof FKBP38. Thus, FKBP38 seems to exert itseffects on apoptosis predominantly throughpromoting insertion of Bcl-2/Bcl-xL into theOMM. However, the possibility remains thatFKBP38 may facilitate crosstalk between Bcl-2/Bcl-xL and the calcineurin/NF-AT sig-nalling pathways, which may be important insome biological contexts.
Interestingly, some immunophilins areknown to be stress-inducible14, although it isunclear whether this is also true for FKBP38.This does, however, raise the possibility thatcell stress may result in increased targeting ofBcl-2 to mitochondria by virtue of elevatedexpression of FKBP38 (Fig. 2). FKBP38 alsocontains additional motifs that may facilitateinteractions with other stress-associated pro-teins. Of particular interest in this regard arethe tetratricopeptide repeats, three of whichare present within the FKBP38 C terminus, asthis region has been implicated in the bindingof Hsp90 to other FKBP proteins. Thus,under conditions of cellular stress,Hsp–FKBP38 interactions may modulateincreased targeting of Bcl-2 to mitochondria,thereby conferring increased protection to themitochondria of stressed cells.
Further studies will be required to teaseout the precise nature of the relationshipbetween FKBP38 and Bcl-2/Bcl-xL. It remainsunclear whether FKBP38 chaperones Bcl-2/Bcl-xL to mitochondria, or promotes theirinsertion into the OMM, or both. It is alsopossible that FKBP38 may remain associatedwith Bcl-2/Bcl-xL within the OMM, promot-ing their role as antagonists of cell death.Notwithstanding these unresolved issues, thecurrent study reinforces the view that theOMM cannot be ignored when makingimportant life or death decisions.Colin Adrain, Emma M. Creagh and Seamus J.Martin are in the Molecular Cell BiologyLaboratory, Department of Genetics, TrinityCollege, Dublin, Ireland.e-mail: [email protected]
1. Jacobson, M. D., Weil M. & Raff, M. C. Cell 88, 347–354 (1997).2. Adrain, C. & Martin, S. J. Trends Biochem. Sci. 26, 319–323
(2001).3. Martinou, J. C. & Green, D. R. Nature Rev. Mol. Cell Biol. 2, 63–67
(2001).4. Kuwana, T. et al. Cell 111, 331–342 (2002).5. Shirane, M. & Nakayama, K. I. Nature Cell Biol. 5, 28–37 (2003).6. Yang, J. et al. Science 275, 1129–1132 (1997).7. Cain, K., Brown, D. G., Langlais, C. & Cohen, G. M. J. Biol. Chem.
274, 22686–22692 (1999).8. Gross, A., McDonnell, J. M. & Korsmeyer, S. J. Genes Dev. 13,
1899–1911 (1999).9. Wei, M. C. et al. Science 292, 727–730 (2001).10. Schiene-Fischer, C. & Yu, C. FEBS Lett. 495, 1–6 (2001).11. Zacchi, P. et al. Nature 419, 853–857 (2002).12. Wang, H. G. et al. Science 284, 339–343 (1999).13. Shibasaki, F., Kondo, E., Akagi, T. & McKeon, F. Nature 386,
728–731 (1997).14. Andreeva, L., Heads, R. & Green, C. J. Int. J. Exp. Path. 80,
305–315 (1999).
FKBP38
FKBP38 Calcineurin
Bcl-2
Bcl-xL
Bcl-xL
Bcl-2Mitochondrion
Mitochondrion
Targeting or insertion ofBcl-2/Bcl-xL
to mitochondrial outer membranes
BadNF-AT
a
b
NF-AT FasL
Nucleus
Nucleartranslocation
Phosphataseactivity
Inhibition of calcineurinphosphatase activity
Figure 2 Possible ways in which FKBP38 may antagonize apoptosis.
© 2003 Nature Publishing Group


























![[Superpartituras.com.Br] Defying Gravity](https://img.dokumen.tips/doc/110x75/55cf9241550346f57b94f8c3/superpartiturascombr-defying-gravity.jpg)


