Embed Size (px)
Citation preview

www.theriojournal.com
Theriogenology 67 (2007) 238–248
Cryopreservation of immature and in vitro matured
porcine oocytes by solid surface vitrification
Mukesh Kumar Gupta, Sang Jun Uhm, Hoon Taek Lee *
Department of Animal Biotechnology, Bio-Organ Research Center, Konkuk University,
1 Hwayang-dong, Gwangjin-Gu, Seoul 143 701, South Korea
Received 1 May 2006; accepted 4 July 2006
Abstract
Cryopreservation of normal, lipid-containing porcine oocytes has had limited practical success. This study used solid surface
vitrification (SSV) of immature germinal vesicle (GV) and mature meiosis II (MII) porcine oocytes and evaluated the effects of
pretreatment with cytochalasin B, cryoprotectant type (dimethylsulfoxide (DMSO), ethylene glycol (EG), or both), and warming
method (two-step versus single-step). Oocyte survival (post-thaw) was assessed by morphological appearance, staining (30,60-diacetyl fluorescein), nuclear maturation, and developmental capacity (after in vitro fertilization). Both GV and MII oocytes were
successfully vitrified; following cryopreservation in EG, more than 60% of GV and MII stage porcine oocytes remained intact (no
significant improvement with cytochalasin B pretreatment). Oocytes (GV stage) vitrified in DMSO had lower (P < 0.05) nuclear
maturation rates (31%) than those vitrified in EG (51%) or EG + DMSO (53%). Survival was better with two-step versus single-step
dilution. Despite high survival rates, rates of cleavage (20–26%) and blastocyst formation (3–9%) were significantly lower than for
non-vitrified controls (60 and 20%). In conclusion, SSV was a very simple, rapid, procedure that allowed normal, lipid-containing,
GV or MII porcine oocytes to be fertilized and develop to the blastocyst stage in vitro.
# 2006 Published by Elsevier Inc.
Keywords: Solid surface vitrification; Cryopreservation; Oocyte; Ovary; Porcine
1. Introduction
Cryopreservation of oocytes has important roles in
the preservation and management of genetic resources,
low-cost international movement of selected genetics,
and rapid dissemination of germplasm via in vitro
embryo production, genetic engineering and nuclear
transfer procedures. It has been successfully applied to
livestock including cattle, goats, sheep, and other model
animals, but has met with limited practical success in
pigs, as porcine oocytes are intrinsically very sensitive
* Corresponding author. Tel.: +82 2 4503675; fax: +82 2 4578488.
E-mail address: [email protected] (H.T. Lee).
0093-691X/$ – see front matter # 2006 Published by Elsevier Inc.
doi:10.1016/j.theriogenology.2006.07.015
to cooling [1,2]. The presence of a large number of
intracytoplasmic lipid droplets in the oocytes has been
considered an obstacle to freezing; lipid reduces the
cryotolerance of the oocytes and cause irreversible
damage to membrane structure at temperatures between
+10 and �5 8C [3]. To circumvent this problem,
Nagashima et al. [4] introduced the concept of
delipation before freezing, which could improve the
survival rate of embryos. However, delipation itself
could compromise embryo viability, as intracellular
lipids are a source of oocyte energy and exist as
complexes of ‘‘smooth endoplasmaic reticulum–lipid
globules–mitochondria’’ in cells [5]. In several studies,
in vivo survival rates of dilapidated vitrified embryos
have been fairly low (range, approximately 10–15%),

M.K. Gupta et al. / Theriogenology 67 (2007) 238–248 239
although pregnancy rates of approximately 80% have
been achieved [6,7].
Vitrification is an alternative to traditional freezing
methods to avoid chilling injury and ice crystal
formation [8]. This technique has become the standard
cryopreservation procedure for porcine embryos,
mainly at the blastocyst stage, with several studies
reporting live offspring after the transfer of vitrified
porcine embryos [6–8]. However, it has not given
satisfactory results for porcine oocytes. During the last
several years, important advances in embryo/oocyte
cryobiology optimized vitrification methodology,
including the use of open pulled straws [6], superfine
open pulled straws [10] and microdroplet methods [7].
The key to success of these methods was a reduction in
the amount of vitrification medium surrounding an
oocyte/embryo, which allowed them to rapidly pass
through a critical temperature zone in the presence of a
cryoprotective agent [11,12]. Piglets have been success-
fully produced after transferring rapidly vitrified
embryos to recipients [6,7,9]. More recently, we
reported the successful vitrification of embryos using
simple weighing paper, which also resulted in produc-
tion of live calves ([13,14], Lee et al., unpublished data).
However, even these protocols have only limited
success for porcine oocytes, as demonstrated by low
survival after warming, low rates of in vitro maturation
[15,16] and reduced development to blastocysts after
intracytoplasmic sperm injection or parthenogenetic
activation [17,18]. Park et al. [19] reported nuclear
maturation rate of vitrified GV stage oocytes to be 3.7%
without delipation and 15.0% after delipation. How-
ever, no blastocysts were obtained, although a cleavage
rate of 16% was obtained following intracytoplasmic
sperm injection.
Solid surface vitrification (SSV) is a simple method
that involves placing oocytes/embryos in a small
amount of cryoprotectant solution and cooling this as
a small drop directly on a metal surface cooled by liquid
nitrogen or liquid nitrogen vapor [20]. High rates of
survival and development were reported for in vitro-
matured oocytes derived from cattle [21], goats [22],
and monkeys [23] cryopreserved with SSV. Somfai
et al. [17] recently applied SSV to pig oocytes followed
by parthenogenetic activation, but development was
low; therefore, further improvements are required
before the widespread application of this technology
to pigs. Furthermore, since cryopreservation can alter
the meiotic spindle assembly, microtubules, cortical
granule distribution, zona pellucida characteristics, and
cause chromosomal aberrations [24–26], it is important
to determine the development ability of vitrified oocytes
after IVF. However, no data are available regarding
developmental ability after IVF of frozen–thawed
porcine oocytes cryropreserved with SSV. Therefore,
the objectives of this study were to investigate the
effects of SSV method on viability and in vitro
development of porcine oocytes following IVF. The
study evaluated the effect of: (1) nuclear status; (2)
cytochalasin B treatment; (3) warming procedure; (4)
cryoprotectant solutions (ethylene glycol (EG) alone,
DMSO alone or EG + DMSO) on oocyte viability, in
vitro maturation and subsequent developmental capa-
city (after IVF).
2. Materials and methods
2.1. Oocyte retrieval and in vitro maturation
Ovaries from prepubertal gilts were collected from a
local abattoir and transported to the laboratory in saline
maintained at 30–37 8C. Cumulus oocyte complexes
(COCs) were aspirated from follicles (2–6 mm dia-
meter) using a 10 mL syringe fitted with an 18 gauge
needle. The COCs were washed three times in TL-
HEPES media containing 1 mg/mL BSA [27] and
matured in groups of 50 in 500 mL of Tissue Culture
Medium 199 with Earle’s salts (TCM-199; Gibco BRL,
Paisley, Scotland, UK), supplemented with 25 mM
NaHCO3, 10% (v/v) porcine follicular fluid, 0.57 mM
cysteine, 0.22 mg/mL sodium pyruvate, 25 mg/mL
gentamicin sulfate, 0.5 mg/mL p-FSH (Folltropin V;
Bioniche Animal Health, Belleville, ON, Canada),
1 mg/mL estradiol-17b, and 10 ng/mL epidermal
growth factor (EGF; Sigma cat. no. E-4127) under
mineral oil at 39 8C in a humidified atmosphere of 5%
CO2 in air for 42–44 h, as described [28].
2.2. Vitrification of oocytes
Only morphologically high quality oocytes, as
determined by uniform granular, homogeneously
distributed cytoplasm surrounded by compact layers
of cumulus cells (for immature oocytes) or oocytes
having an intact first polar body and a clear cytoplasm
with uniform texture and homogeneous fine granularity
surrounded by >70% homogeneously spread cumulus
cells (for in vitro matured oocytes), were used for
vitrification [29]. Immature (Germinal Vesicle or GV
stage) or in vitro matured (MII stage) were vitrified by
solid surface vitrification, as described by Dinnyes et al.
[20], with some modifications. Briefly, COCs were
partially denuded using 0.1% hyaluronidase and
washed three times in HEPES buffered TCM 199 with

M.K. Gupta et al. / Theriogenology 67 (2007) 238–248240
Earles salt (HTCM; Gibco BRL) supplemented with
20% FBS (FBS; Hyclone, Gibco BRL) and then
suspended in an equilibration medium consisting of 4%
(v/v) ethylene glycol (EG; Sigma cat. no. E-9129) in
TCM 199 + 20% FBS for 10–15 min at 39 8C. Oocytes
were then briefly rinsed three times (<30 s) in
vitrification solution (pre-warmed to 39 8C) consisting
of 35% EG, 5% polyvinyl pyrrolidone (PVP; Sigma cat.
no. P-0930), and 13.7% (w/v) sucrose (Sigma cat. no.
S9378) in HTCM-199 +20% FBS and placed in groups
of �25 oocytes per �1–2 mL droplet on to aluminum
foil kept directly over the liquid nitrogen (LN2). Upon
visual observation of the droplets, those that were
vitrified (completely transparent) were moved with a
liquid-nitrogen-cooled forceps into 1-mL cryovials
(Nunc cat. no. 343958) and plunged in LN2 for storage.
Vitrified oocytes were held in LN2 for at least 1 week
before warming. Warming was performed by dropping
the vitrified droplets from the cryotube directly into a
Petri dish containing 500 mL of 10.3% (w/v) sucrose
solution at 39 8C for 5 min. All solutions in the
microdrops remained transparent during cooling and
warming, which is one indication that the solutions
vitrified. If droplets appeared opaque, they were
discarded. Warmed oocytes were then washed in
HTCM-199 +20% FBS medium three times and kept
in the last HTCM-199 +20% FBS drop for at least 10–
15 min before transferring them into in vitro maturation
or fertilization medium.
2.3. Evaluation of oocyte viability
Oocyte survival was evaluated morphologically
based on the integrity of the oolemma and zona
pellucida; loss of membrane integrity (lysis) was
obvious upon visual inspection as the sharp demarcation
of the membrane disappeared and the appearance of the
cytoplasm changed. Oocytes were also assessed for
viability based on esterase enzyme activity and
oolemma integrity by FDA (30,60-diacetyl fluorescein)
staining, as described by Noto et al. [30]. Briefly,
oocytes were washed in PBS for 1 min, followed by
incubation with 2.5 mg/mL FDA (Sigma cat. no. F-
7378) stain for 1 min. Stained oocytes were washed
again in PBS and observed under ultraviolet illumina-
tion with an epifluorescence microscope with filters at
460–490 for excitation and 515–550 for emission
(Nikon Co., Tokyo, Japan). Live oocytes had green
fluorescence, whereas dead oocytes were non-fluores-
cent. Surviving oocytes were then further incubated for
at least 2 h at 39 8C in 5% CO2 and re-evaluated for
survival.
2.4. In vitro fertilization (IVF)
In vitro fertilization of surviving oocytes was
conducted as previously described [29]. Briefly, oocytes
were washed three times with the fertilization medium
(modified Tris-buffered medium; [31]) containing
1 mM caffeine sodium benzoate and 0.1% BSA and
were placed into groups of 10–15 oocytes per 50 mL
droplets of fertilization medium. Porcine testis were
collected from a local abbatoir and transported to the
laboratory at 30–35 8C in 0.9% (w/v) saline supple-
mented with 75 mg/mL penicillin G and 50 mg/mL
streptomycin sulfate. Sperm were retrieved from cauda
epididymis in TL-HEPES, and pelleted by centrifuga-
tion at 800 rpm for 5 min. The soft pellet was then
subjected to swim-up in Sp-TALP medium [27] for
10 min. The supernatant was collected and washed
twice by centrifugation at 800 rpm for 5 min. At the end
of the washing procedure, the sperm pellet was
resuspended in the fertilization medium and added to
the fertilization droplet to obtain a final sperm
concentration of 5 � 105 cells/mL. Sperm and oocytes
were co-incubated at 39 8C in a humidified atmosphere
of 5% CO2 in air for 6 h.
2.5. In vitro culture of presumptive zygotes
At the end of co-incubation period, presumptive
zygotes were cultured in NCSU23 medium supplemen-
ted with 0.4% fatty acid free bovine serum albumin
(BSA; Sigma cat. no. A-6033) for 5 days and in NCSU23
medium supplemented with 10% FBS for the next 2 days,
as described [32]. Cleavage rate was assessed on Day 2
and the blastocyst rate on Day 7 of culture.
2.6. Experimental design
The study comprised five experiments to evaluate the
effect of oocyte nuclear status, cytochalasin B treat-
ment, warming procedure and different cryoprotectant
solutions. In the first four experiments, we evaluated the
effect of above treatments on the viability of oocytes
following vitrification; the best combination was chosen
for the fifth experiment in which developmental ability
of vitrified oocytes was evaluated after IVF. At least
three replicates were performed in each experiment.
2.6.1. Experiment 1: effect of nuclear stage of
oocyte
The first experiment was designed to compare the
viability of immature (GV stage; n = 543) and mature
(MII stage; n = 574) porcine oocytes following vitri-
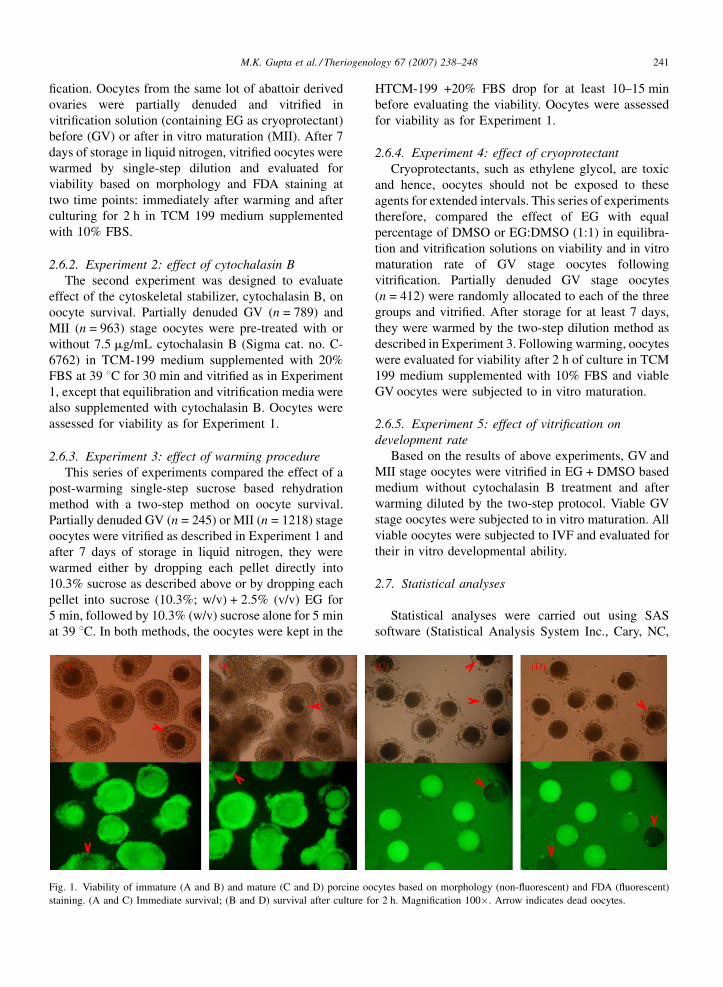
M.K. Gupta et al. / Theriogenology 67 (2007) 238–248 241
fication. Oocytes from the same lot of abattoir derived
ovaries were partially denuded and vitrified in
vitrification solution (containing EG as cryoprotectant)
before (GV) or after in vitro maturation (MII). After 7
days of storage in liquid nitrogen, vitrified oocytes were
warmed by single-step dilution and evaluated for
viability based on morphology and FDA staining at
two time points: immediately after warming and after
culturing for 2 h in TCM 199 medium supplemented
with 10% FBS.
2.6.2. Experiment 2: effect of cytochalasin B
The second experiment was designed to evaluate
effect of the cytoskeletal stabilizer, cytochalasin B, on
oocyte survival. Partially denuded GV (n = 789) and
MII (n = 963) stage oocytes were pre-treated with or
without 7.5 mg/mL cytochalasin B (Sigma cat. no. C-
6762) in TCM-199 medium supplemented with 20%
FBS at 39 8C for 30 min and vitrified as in Experiment
1, except that equilibration and vitrification media were
also supplemented with cytochalasin B. Oocytes were
assessed for viability as for Experiment 1.
2.6.3. Experiment 3: effect of warming procedure
This series of experiments compared the effect of a
post-warming single-step sucrose based rehydration
method with a two-step method on oocyte survival.
Partially denuded GV (n = 245) or MII (n = 1218) stage
oocytes were vitrified as described in Experiment 1 and
after 7 days of storage in liquid nitrogen, they were
warmed either by dropping each pellet directly into
10.3% sucrose as described above or by dropping each
pellet into sucrose (10.3%; w/v) + 2.5% (v/v) EG for
5 min, followed by 10.3% (w/v) sucrose alone for 5 min
at 39 8C. In both methods, the oocytes were kept in the
Fig. 1. Viability of immature (A and B) and mature (C and D) porcine oo
staining. (A and C) Immediate survival; (B and D) survival after culture fo
HTCM-199 +20% FBS drop for at least 10–15 min
before evaluating the viability. Oocytes were assessed
for viability as for Experiment 1.
2.6.4. Experiment 4: effect of cryoprotectant
Cryoprotectants, such as ethylene glycol, are toxic
and hence, oocytes should not be exposed to these
agents for extended intervals. This series of experiments
therefore, compared the effect of EG with equal
percentage of DMSO or EG:DMSO (1:1) in equilibra-
tion and vitrification solutions on viability and in vitro
maturation rate of GV stage oocytes following
vitrification. Partially denuded GV stage oocytes
(n = 412) were randomly allocated to each of the three
groups and vitrified. After storage for at least 7 days,
they were warmed by the two-step dilution method as
described in Experiment 3. Following warming, oocytes
were evaluated for viability after 2 h of culture in TCM
199 medium supplemented with 10% FBS and viable
GV oocytes were subjected to in vitro maturation.
2.6.5. Experiment 5: effect of vitrification on
development rate
Based on the results of above experiments, GV and
MII stage oocytes were vitrified in EG + DMSO based
medium without cytochalasin B treatment and after
warming diluted by the two-step protocol. Viable GV
stage oocytes were subjected to in vitro maturation. All
viable oocytes were subjected to IVF and evaluated for
their in vitro developmental ability.
2.7. Statistical analyses
Statistical analyses were carried out using SAS
software (Statistical Analysis System Inc., Cary, NC,
cytes based on morphology (non-fluorescent) and FDA (fluorescent)
r 2 h. Magnification 100�. Arrow indicates dead oocytes.
M.K. Gupta et al. / Theriogenology 67 (2007) 238–248242
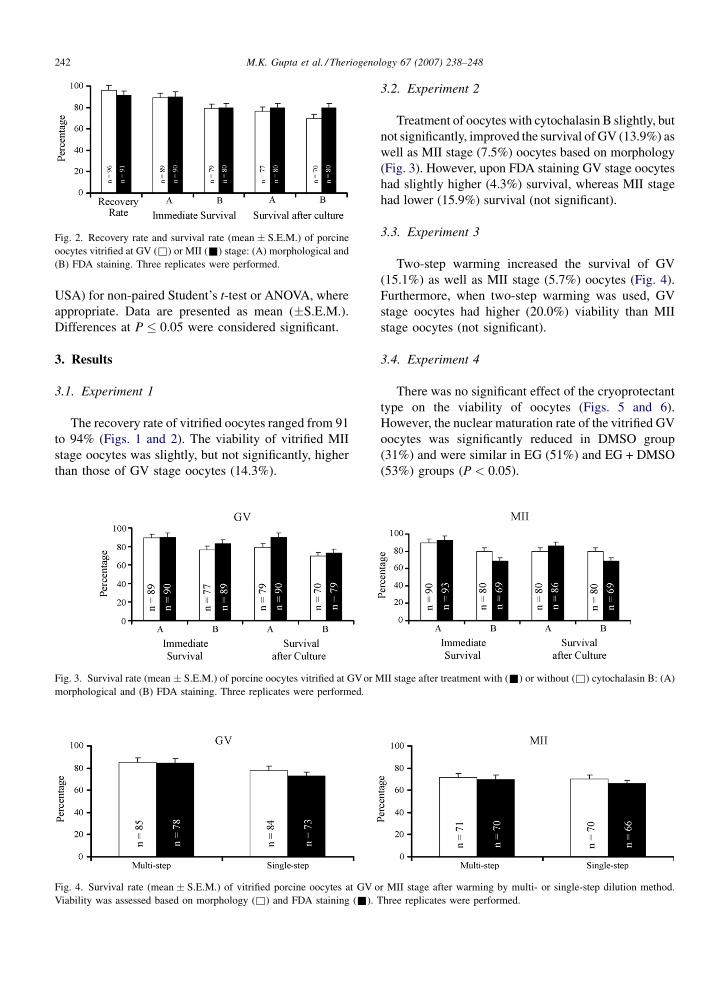
Fig. 2. Recovery rate and survival rate (mean � S.E.M.) of porcine
oocytes vitrified at GV (&) or MII (&) stage: (A) morphological and
(B) FDA staining. Three replicates were performed.
USA) for non-paired Student’s t-test or ANOVA, where
appropriate. Data are presented as mean (�S.E.M.).
Differences at P � 0.05 were considered significant.
3. Results
3.1. Experiment 1
The recovery rate of vitrified oocytes ranged from 91
to 94% (Figs. 1 and 2). The viability of vitrified MII
stage oocytes was slightly, but not significantly, higher
than those of GV stage oocytes (14.3%).
Fig. 3. Survival rate (mean � S.E.M.) of porcine oocytes vitrified at GVor M
morphological and (B) FDA staining. Three replicates were performed.
Fig. 4. Survival rate (mean � S.E.M.) of vitrified porcine oocytes at GV o
Viability was assessed based on morphology (&) and FDA staining (&).
3.2. Experiment 2
Treatment of oocytes with cytochalasin B slightly, but
not significantly, improved the survival of GV (13.9%) as
well as MII stage (7.5%) oocytes based on morphology
(Fig. 3). However, upon FDA staining GV stage oocytes
had slightly higher (4.3%) survival, whereas MII stage
had lower (15.9%) survival (not significant).
3.3. Experiment 3
Two-step warming increased the survival of GV
(15.1%) as well as MII stage (5.7%) oocytes (Fig. 4).
Furthermore, when two-step warming was used, GV
stage oocytes had higher (20.0%) viability than MII
stage oocytes (not significant).
3.4. Experiment 4
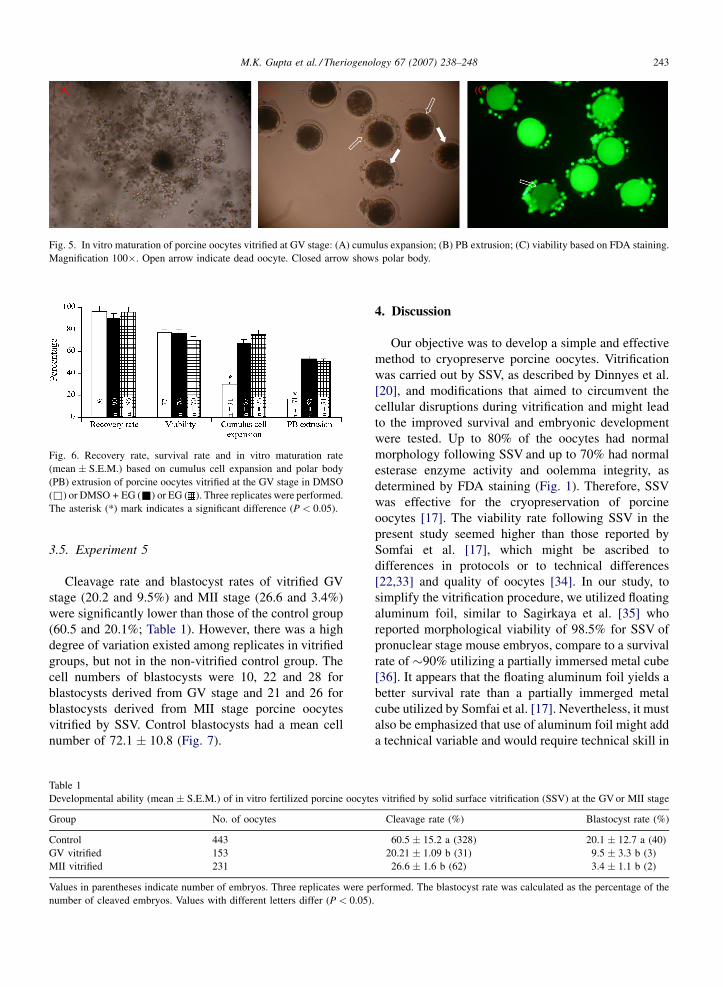
There was no significant effect of the cryoprotectant
type on the viability of oocytes (Figs. 5 and 6).
However, the nuclear maturation rate of the vitrified GV
oocytes was significantly reduced in DMSO group
(31%) and were similar in EG (51%) and EG + DMSO
(53%) groups (P < 0.05).
II stage after treatment with (&) or without (&) cytochalasin B: (A)
r MII stage after warming by multi- or single-step dilution method.
Three replicates were performed.
M.K. Gupta et al. / Theriogenology 67 (2007) 238–248 243
Fig. 5. In vitro maturation of porcine oocytes vitrified at GV stage: (A) cumulus expansion; (B) PB extrusion; (C) viability based on FDA staining.
Magnification 100�. Open arrow indicate dead oocyte. Closed arrow shows polar body.
Fig. 6. Recovery rate, survival rate and in vitro maturation rate
(mean � S.E.M.) based on cumulus cell expansion and polar body
(PB) extrusion of porcine oocytes vitrified at the GV stage in DMSO
(&) or DMSO + EG (&) or EG ( ). Three replicates were performed.
The asterisk (*) mark indicates a significant difference (P < 0.05).
3.5. Experiment 5
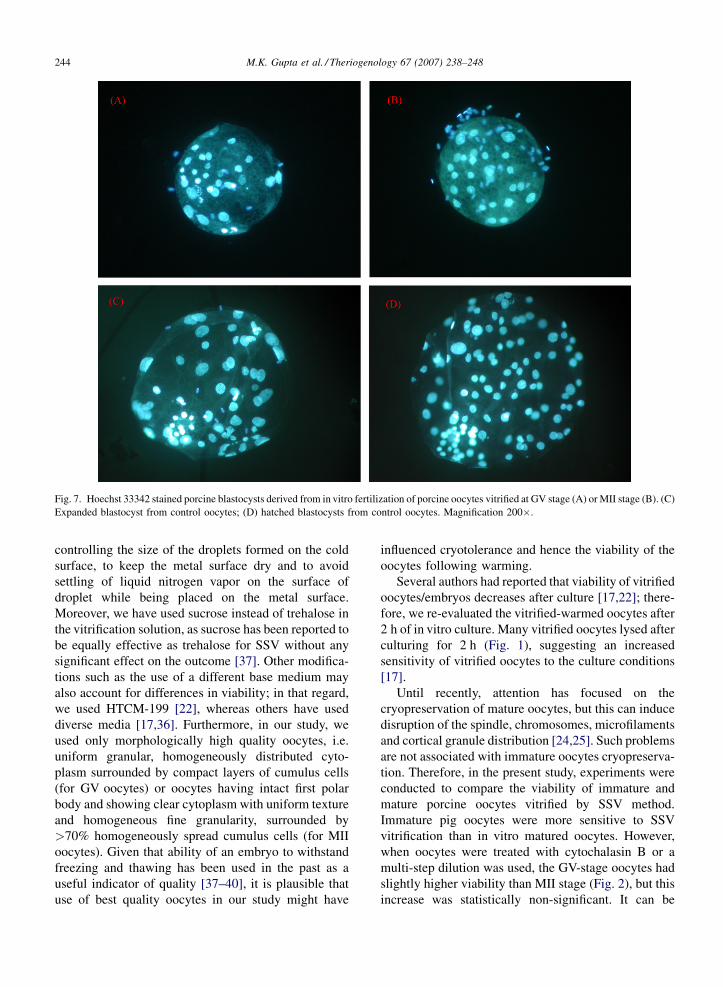
Cleavage rate and blastocyst rates of vitrified GV
stage (20.2 and 9.5%) and MII stage (26.6 and 3.4%)
were significantly lower than those of the control group
(60.5 and 20.1%; Table 1). However, there was a high
degree of variation existed among replicates in vitrified
groups, but not in the non-vitrified control group. The
cell numbers of blastocysts were 10, 22 and 28 for
blastocysts derived from GV stage and 21 and 26 for
blastocysts derived from MII stage porcine oocytes
vitrified by SSV. Control blastocysts had a mean cell
number of 72.1 � 10.8 (Fig. 7).
Table 1
Developmental ability (mean � S.E.M.) of in vitro fertilized porcine oocyte
Group No. of oocytes
Control 443
GV vitrified 153
MII vitrified 231
Values in parentheses indicate number of embryos. Three replicates were p
number of cleaved embryos. Values with different letters differ (P < 0.05)
4. Discussion
Our objective was to develop a simple and effective
method to cryopreserve porcine oocytes. Vitrification
was carried out by SSV, as described by Dinnyes et al.
[20], and modifications that aimed to circumvent the
cellular disruptions during vitrification and might lead
to the improved survival and embryonic development
were tested. Up to 80% of the oocytes had normal
morphology following SSV and up to 70% had normal
esterase enzyme activity and oolemma integrity, as
determined by FDA staining (Fig. 1). Therefore, SSV
was effective for the cryopreservation of porcine
oocytes [17]. The viability rate following SSV in the
present study seemed higher than those reported by
Somfai et al. [17], which might be ascribed to
differences in protocols or to technical differences
[22,33] and quality of oocytes [34]. In our study, to
simplify the vitrification procedure, we utilized floating
aluminum foil, similar to Sagirkaya et al. [35] who
reported morphological viability of 98.5% for SSV of
pronuclear stage mouse embryos, compare to a survival
rate of �90% utilizing a partially immersed metal cube
[36]. It appears that the floating aluminum foil yields a
better survival rate than a partially immerged metal
cube utilized by Somfai et al. [17]. Nevertheless, it must
also be emphasized that use of aluminum foil might add
a technical variable and would require technical skill in
s vitrified by solid surface vitrification (SSV) at the GV or MII stage
Cleavage rate (%) Blastocyst rate (%)
60.5 � 15.2 a (328) 20.1 � 12.7 a (40)
20.21 � 1.09 b (31) 9.5 � 3.3 b (3)
26.6 � 1.6 b (62) 3.4 � 1.1 b (2)
erformed. The blastocyst rate was calculated as the percentage of the
.
M.K. Gupta et al. / Theriogenology 67 (2007) 238–248244
Fig. 7. Hoechst 33342 stained porcine blastocysts derived from in vitro fertilization of porcine oocytes vitrified at GV stage (A) or MII stage (B). (C)
Expanded blastocyst from control oocytes; (D) hatched blastocysts from control oocytes. Magnification 200�.
controlling the size of the droplets formed on the cold
surface, to keep the metal surface dry and to avoid
settling of liquid nitrogen vapor on the surface of
droplet while being placed on the metal surface.
Moreover, we have used sucrose instead of trehalose in
the vitrification solution, as sucrose has been reported to
be equally effective as trehalose for SSV without any
significant effect on the outcome [37]. Other modifica-
tions such as the use of a different base medium may
also account for differences in viability; in that regard,
we used HTCM-199 [22], whereas others have used
diverse media [17,36]. Furthermore, in our study, we
used only morphologically high quality oocytes, i.e.
uniform granular, homogeneously distributed cyto-
plasm surrounded by compact layers of cumulus cells
(for GV oocytes) or oocytes having intact first polar
body and showing clear cytoplasm with uniform texture
and homogeneous fine granularity, surrounded by
>70% homogeneously spread cumulus cells (for MII
oocytes). Given that ability of an embryo to withstand
freezing and thawing has been used in the past as a
useful indicator of quality [37–40], it is plausible that
use of best quality oocytes in our study might have
influenced cryotolerance and hence the viability of the
oocytes following warming.
Several authors had reported that viability of vitrified
oocytes/embryos decreases after culture [17,22]; there-
fore, we re-evaluated the vitrified-warmed oocytes after
2 h of in vitro culture. Many vitrified oocytes lysed after
culturing for 2 h (Fig. 1), suggesting an increased
sensitivity of vitrified oocytes to the culture conditions
[17].
Until recently, attention has focused on the
cryopreservation of mature oocytes, but this can induce
disruption of the spindle, chromosomes, microfilaments
and cortical granule distribution [24,25]. Such problems
are not associated with immature oocytes cryopreserva-
tion. Therefore, in the present study, experiments were
conducted to compare the viability of immature and
mature porcine oocytes vitrified by SSV method.
Immature pig oocytes were more sensitive to SSV
vitrification than in vitro matured oocytes. However,
when oocytes were treated with cytochalasin B or a
multi-step dilution was used, the GV-stage oocytes had
slightly higher viability than MII stage (Fig. 2), but this
increase was statistically non-significant. It can be

M.K. Gupta et al. / Theriogenology 67 (2007) 238–248 245
speculated that the cumulus cells enclosing the oocytes
could have influenced the cryoprotectant or water
permeability and hence affected the survival. There are
however, conflicting reports regarding the effects of
cumulus cells surrounding the oocyte for subsequent
development of matured oocytes after vitrification and
warming [41,42]. Fujihira et al. [16] reported a
significant relationship between the presence of
cumulus cells surrounding the oocytes and the
cryoprotectant permeability. Several earlier studies
have documented the GV stage to be more sensitive
than any other nuclear stages [43,44] for probable
reasons such as impaired intercellular communication
between the oocyte and the cumulus cells and
substantial disruption of microfilaments [43].
Osmotic stress during cryopreservation may alter the
physicochemical properties and integrity of cytoskeletal
elements, leading to abnormal progression of meiotic
division and the retardation of embryo development
[9,43,44]. Stabilizing the cytoskeleton system during
vitrification could be beneficial for improving post-
thawed survival and subsequent development of
vitrified oocytes [15,45]. Cytochalasin B has a specific,
reversible effect on depolymerization of cytoskeletal
elements, making them more flexible and hence, less
susceptible to cryo-damage [46]. However, controver-
sial results have been reported with the effect of
cytochalasin B on oocyte vitrification [15,16,47]. In our
study, pre-treatment of oocytes with cytochalasin B did
not significantly improve the proportion of surviving
oocytes (Fig. 3), consistent with Somfai et al. [17]
during SSV of porcine oocytes and Silvestre et al. [47]
for open pulled straw vitrification of sheep oocytes.
To avoid formation of ice crystals, a high concentra-
tion of cryoprotectants are included in the vitrification
solution, which must be removed after warming. Multi-
step dilution may be beneficial by avoiding sudden
osmolarity changes that may damage the oocyte,
whereas a single-step dilution minimizes the steps
required for oocyte washing, thereby substantially
reducing the time needed for warming and special
manipulations [48]. In the present study, the multi-step
dilution was found to be more effective (Fig. 4),
consistent with a previous report [15].
Different cryoprotectants can have different effects
on the viability and developmental potential of vitrified
oocytes and embryos. Recently, EG has gained
importance as an effective cryoprotectant for vitrifica-
tion of oocytes [17,49] as it is less toxic and is rapidly
permeable (into and out of oocytes) during cryopre-
servation, due to its low molecular weight. A balance
between the toxicity and cryoprotective action of the
cryoprotectant is critical for developing an efficient
cryopreservation method. Inclusion of a second
cryoprotectant can be used to reduce the concentration,
and hence the toxicity, of each cryoprotectant. In that
regard, a combination of EG with DMSO was reported
to give better survivability than EG alone [50], whereas
others have reported no added benefit of the combina-
tion [16]. In the present study, there was no beneficial
effect of the combination of DMSO and EG versus EG
per se on oocyte viability, as well as nuclear maturation
after vitrification of immature porcine oocytes. The EG-
based cryoprotectant medium was superior to DMSO
alone (Fig. 6). Although DMSO is a common
cryoprotectant used for freezing of cells and has
yielded good success in vitrification of human oocytes
by the open pulled straw method [51], we obtained the
worst results when DMSO alone was used. This
outcome was similar to those of Bagis et al. [36]
who compared three different cryoprotectants for SSV
of pronuclear stage mouse embryos; in that study, EG
was better than DMSO or propylene glycol for
embryonic development after vitrification. In a com-
parative study with bovine oocytes, Cestin and Bastan
[52] reported that EG or EG + DMSO was superior to
DMSO alone for vitrification. The exact cause for these
variable results is not clear. Trounson and Kirby [53]
documented that mouse oocytes exposed to DMSO at
temperatures from 4 to 37 8C incurred serious
irreversible changes in microtubules, pericentriolar
material and chromosomes of unfertilized oocytes;
they suggested a relationship between the toxicity of
DMSO with temperature and duration of exposure to
oocytes. Better results were obtained when oocytes
were exposed to DMSO at lower temperatures [53,54].
That the temperature of equilibration and vitrification
solution was 39 8C in the present study might have
contributed to the lower success obtained with DMSO.
Alternatively, DMSO might have caused spindle
polymerization with increased potential for polyploidy
[55] and therefore, had higher adverse effect on spindle
configuration than the combination of EG and DMSO
[56].
The presence of a large number of intracytoplasmic
lipid droplets in the oocytes has been considered an
obstacle to cryopreservation of porcine oocytes as lipid
reduces the cryotolerance of the oocytes and causes
irreversible damage to membrane structure at low
temperatures [3]. To circumvent this problem, Naga-
shima et al. [4] had introduced the concept of delipation
before cryopreservation. However, in the present
experiments using the SSV method to preserve porcine
oocytes that had not undergone delipation, these

M.K. Gupta et al. / Theriogenology 67 (2007) 238–248246
oocytes were not only capable of resuming meiosis, but
also developed in vitro after vitrification (Fig. 5),
consistent with previous reports [17,19].
The EG + DMSO combination allowed both GV and
MII vitrified oocytes to develop to blastocysts following
IVF. However, the development rate and cell number of
blastocysts was much lower than that obtained with fresh
oocytes (Table 1; Fig. 7). Furthermore, there was
individual variation among replicates for formation of
blastocysts, indicating that further improvements in the
technical aspects of the methodology are needed. The
low blastocyst rate and cell number can be ascribed to the
damages caused by cryopreservation, e.g. damage to
microtubules, pericentriolar materials and chromosomes
[53], aberrant spindle polymerization and polyploidy
[54], meiotic spindle disassembly, abnormal cortical
granule distribution and chromosomal aberrations [24–
26] and alterations in zona pellucida characteristics
[55,56]. Larman et al. [26] reported that DMSO as well as
EG used in vitrification solutions caused large transient
increases in intracellular calcium concentration in mouse
metaphase II oocytes, comparable to the initial increases
triggered at fertilization. This rise in calcium level can
negatively affect several physiological processes within
oocytes during oocyte vitrification and thus, might
explain the current poor efficiency of vitrification.
Successful production of blastocysts from vitrified
porcine oocytes has been reported by intracytoplasmic
sperm injection [16], parthenogenetic activation [17] and
following delipation [6]. However, to our knowledge, the
present study is the first to report the IVF blastocyst
production from porcine GV- or MII-stage porcine
oocytes, preserved with SSV, without any micromani-
pulation or delipation procedure.
In conclusion, SSV, a simple and rapid procedure,
allowed normal, lipid-containing, porcine oocytes
vitrified at either the GV or MII stage to be fertilized
and develop into blastocysts in vitro. The study also
demonstrated that: (1) GV and MII stage were equally
tolerant to the vitrification protocol; (2) pre-treatment
with cytochalasin B had no significant effect on the
vitrification of porcine oocytes; (3) two-step warming
was superior to single-step warming; (4) the use of EG or
EG and DMSO as a cryoprotectant were advantageous
for in vitro maturation of vitrified immature porcine
oocytes; (5) vitrified–warmed porcine oocytes matured
after IVM, could develop to the blastocyst stage, albeit at
a low efficiency, following IVF. We do not regard this
vitrification protocol as a definitive protocol; rather, we
regard it as a direction for further investigations into the
cryopreservation of porcine oocytes by SSV, which may
lead to more effective protocols.
Acknowledgment
This work was supported by the Research Project on
the Production of Bio-Organs (no. 200503030201),
Ministry of Agriculture and Forestry, and Biogreen 21,
RDA, Republic of Korea.
References
[1] Didion BA, Pomp D, Martin MJ, Homanics GE, Markert CL.
Observations on the cooling and cryopreservation of pig oocytes
at the germinal vesicle stage. J Anim Sci 1990;68:2803–10.
[2] Polge C, Wilmut I, Rowson LEA. The low temperature pre-
servation of cow, sheep and pig embryos. Cryobiology 1974;
11:560.
[3] Nagashima H, Kashiwazaki N, Ashman RJ, Grupen CG, Sea-
mark RF, Nottle MB. Recent advances in cryopreservation of
porcine embryos. Theriogenology 1994;41:113–8.
[4] Nagashima H, Kashiwazaki N, Ashman RJ, Grupen CG, Sea-
mark RF, Nottle MB. Removal of cytoplasmic lipid enhances the
tolerance of porcine embryos to chilling. Biol Reprod 1994;
51:618–22.
[5] Sathananthan AH. Ultrastructural changes during meiotic
maturation in mammalian oocytes: unique aspects of the human
oocyte. Microbiol Res Technol 1994;27:145–64.
[6] Berthelot F, Marinat-Botte F, Perreau C, Terqui M. Birth of
piglets after OPS vitrification and transfer of compacted morula
stage embryos with intact zona pellucida. Reprod Nutr Dev
2001;41:267–72.
[7] Misumi K, Suzuki M, Sato S, Saito N. Successful production of
piglets from vitrified morulae and early blastocysts using a
microdrop method. Theriogenology 2003;60:253–60.
[8] Rall WF, Fahy GM. Ice-free cryopreservation of mouse embryos
at �196 8C by vitrification. Nature 1985;313:573–5.
[9] Dobrinsky JR, Pursel VG, Long CR, Johnson LA. Birth of piglets
after transfer of embryos cryopreserved by cytoskeletal stabili-
zation and vitrification. Biol Reprod 2000;62:564–70.
[10] Isachenko V, Folch J, Isachenko E, Nawroth F, Krivokharchenko
A, Vajta G, et al. Double vitrification of rat embryos at different
developmental stages using an identical protocol. Theriogenol-
ogy 2003;60:445–52.
[11] Vajta G. Vitrification of the oocytes and embryos of domestic
animals. Anim Reprod Sci 2000;60:357–64.
[12] Arav A, Yavin S, Zeron Y, Natan D, Dekel I, Gacitua H. New
trend in gamete cryopreservation. Mol Cell Endocrinol 2002;
187:77–81.
[13] Kim YM, Ko DH, Uhm SJ, Chung KS, Lee HT. A new paper
container for the vitrification of bovine embryos. Reprod Fertil
Dev 2004;16:173.
[14] Lee HT, Kim YM, Uhm SJ, Ko DH. Development of a new
vitrification container for freezing of bovine embryos. In: Pro-
ceedings of the Ninth Asian Symposium on Animal Biotechnol-
ogy (ASAB); 2004.
[15] Isachenko V, Soler C, Isachenko E, Perez-Sanchez F, Grish-
chenko V. Vitrification of immature porcine oocytes: effects of
lipid droplets, temperature, cytoskeleton, and addition and
removal of cryoprotectant. Cryobiology 1998;36:250–3.
[16] Fujihira T, Kishida R, Fukui Y. Developmental capacity of vitrified
immature porcine oocytes following ICSI: effects of cytochalasin
B and cryoprotectants. Cryobiology 2004;49:286–90.

M.K. Gupta et al. / Theriogenology 67 (2007) 238–248 247
[17] Somfai T, Dinnyes A, Sage D, Marosan M, Carnwath JW, Ozawa
M, et al. Development to the blastocyst stage of parthenogen-
etically activated in vitro matured porcine oocytes after solid
surface vitrification (SSV). Theriogenology 2006 [Epub ahead
of print].
[18] Fujihira T, Nagai H, Fukui Y. Relationship between equilibration
times and the presence of cumulus cells, and effect of Taxol
treatment for vitrification of in vitro matured porcine oocytes.
Cryobiology 2005;51:339–43.
[19] Park KE, Kwon IK, Han MS, Niwa K. Effects of partial removal
of cytoplasmic lipid on survival of vitrified germinal vesicle
stage pig oocytes. J Reprod Dev 2005;51:151–60.
[20] Dinnyes A, Dai Y, Jiang S, Yang X. Somatic-cell nuclear transfer
with vitrified recipient oocytes in cattle. Theriogenology 2000;
53:215.
[21] Dinnyes A, Dai Y, Jiang S, Yang X. High developmental rates of
vitrified bovine oocytes following parthenogenetic activation, in
vitro fertilization, and somatic cell nuclear transfer. Biol Reprod
2000;63:513–8.
[22] Begin I, Bhatia B, Baldassarre H, Dinnyes A, Keefer CL.
Cryopreservation of goat oocytes and in vivo derived 2- to 4-
cell embryos using the cryoloop (CLV) and solid-surface vitri-
fication (SSV) methods. Theriogenology 2003;59:1839–50.
[23] Dinnyes A, Wei S, Li Y, Zheng P, Ji W. First report on cleavage
development following cryopreservation of adult and prepuber-
tal rhesus monkey (Macaca mulatta) oocytes. Reprod Fertil Dev
2004;16:169.
[24] Aman RR, Parks JE. Effects of cooling and rewarming on the
meiotic spindle and chromosomes of in vitro-matured bovine
oocytes. Biol Reprod 1994;50:103–10.
[25] Saunders KM, Parks JE. Effects of cryopreservation procedures
on the cytology and fertilization rate of in vitro-matured bovine
oocytes. Biol Reprod 1999;61:178–87.
[26] Larman MG, Sheehan CB, Gardner DK. Calcium-free vitrifica-
tion reduces cryoprotectant-induced zona pellucida hardening
and increases fertilization rates in mouse oocytes. Reproduction
2006;131:53–61.
[27] Parrish JJ, Susko-Parrish J, Winer MA, First NL. Capacitation of
bovine sperm by heparin. Biol Reprod 1988;38:1171–80.
[28] Uhm SJ, Kim NH, Kim T, Chung HM, Chung KH, Lee HT, et al.
Expression of enhanced green fluorescent protein (EGFP) and
neomycin resistant (Neo(R)) genes in porcine embryos following
nuclear transfer with porcine fetal fibroblasts transfected by
retrovirus vector. Mol Reprod Dev 2000;57:331–7.
[29] Park CY, Uhm SJ, Song SJ, Kim KS, Hong SB, Chung KS, et al.
Increase of ICSI efficiency with hyaluronic acid binding sperm
for low aneuploidy frequency in pig. Theriogenology 2005;64:
1158–69.
[30] Noto V, Campo R, Roziers P, Gordts S. Fluorescein diacetate
assessment of embryo viability after ultrarapid freezing of
human multipronucleate embryos. Fertil Steril 1991;55:1171–5.
[31] Abeydeera LR, Day BN. In vitro penetration of pig oocytes in a
modified Tris-buffered medium: effect of BSA, caffeine and
calcium. Theriogenology 1997;48:537–44.
[32] Koo DB, Kim NH, Lee HT, Chung KS. Effects of fetal calf
serum, amino acids, vitamins and insulin on blastocoel formation
and hatching on in vivo and IVM/IVF-derived porcine embryos
developing in vitro. Theriogenology 1997;48:791–802.
[33] Vajta G, Kuwayama M. Improving cryopreservation systems.
Theriogenology 2006;65:236–44.
[34] Seidel Jr GE. Modifying oocytes and embryos to improve their
cryopreservation. Theriogenology 2006;65:228–35.
[35] Sagirkaya H, Ergin F, Bagis H, Arat A. Vitrification of pro-
nuclear-stage mouse embryos on aluminum foil floating on
liquid nitrogen. Reprod Fertil Dev 2004;16:181.
[36] Bagis H, Mercan HO, Kumtepe Y. Effect of three different
cryoprotectant solutions in solid surface vitrification (SSV)
techniques on the development rate of vitrified pronuclear-stage
mouse embryos. Turk J Vet Anim Sci 2005;29:621–7.
[37] Rizos D, Ward F, Boland MP, Lonergan P. Effect of culture
system on the yield and quality of bovine blastocysts as assessed
by survival after vitrification. Theriogenology 2001;56:1–16.
[38] Rizos D, Gutierrez-Adan A, Perez-Garnelo S, De La Fuente J,
Boland MP, Lonergan P. Bovine embryo culture in the presence
or absence of serum: implications for blastocyst development,
cryotolerance, and messenger RNA expression. Biol Reprod
2003;68:236–43.
[39] Enright BP, Lonergan P, Dinnyes A, Fair T, Ward FA, Yang X,
et al. Culture of in vitro produced bovine zygotes in vitro vs in
vivo: implications for early embryo development and quality.
Theriogenology 2000;54:659–73.
[40] Kaidi S, Donnay I, Van Langendonckt A, Dessy F, Massip A.
Comparison of two co-culture systems to assess the survival of in
vitro produced bovine blastocysts after vitrification. Anim
Reprod Sci 1998;52:39–50.
[41] Gook DA, Osborn SM, Johnston WI. Cryopreservation of mouse
and human oocytes using 1,2-propanediol and the configuration
of the meiotic spindle. Hum Reprod 1993;8:1101–9.
[42] Park SE, Chung HM, Cha KY, Hwang WS, Lee ES, Lim JM.
Cryopreservation of ICR mouse oocytes: improved post-thawed
preimplantation development after vitrification using Taxol, a
cytoskeleton stabilizer. Fertil Steril 2001;75:1177–84.
[43] Rojas C, Palomo MJ, Albarracin JL, Mogas T. Vitrification of
immature and in vitro matured pig oocytes: study of distribution
of chromosomes, microtubules, and actin microfilaments. Cryo-
biology 2004;49:211–20.
[44] Dobrinsky JR, Overstrom EW, Johnson LA. Cytoskeletal ana-
lysis of vitrified porcine embryos. J Anim Sci 1994;72
(Suppl):82.
[45] Shi WQ, Zhu SE, Zhang D, Wang WH, Tang GL, Hou YP, et al.
Improved development by Taxol pretreatment after vitrification
of in vitro matured porcine oocytes. Reproduction 2006;131:
795–804.
[46] Vincent C, Garnier V, Heyman Y, Renard JP. Solvent effects on
cytoskeletal organization and in vivo survival after freezing of
rabbit oocytes. J Reprod Fertil 1989;87:809–20.
[47] Silvestre MA, Yaniz J, Salvador I, Santolaria P, Lopez-Gatius F.
Vitrification of pre-pubertal ovine cumulus-oocyte complexes:
effect of cytochalasin B pre-treatment. Anim Reprod Sci 2006;
93:176–82.
[48] Cuello C, Gil MA, Parrilla I, Tornel J, Vazquez JM, Roca J, et al.
In vitro development following one-step dilution of OPS-vitri-
fied porcine blastocysts. Theriogenology 2004;62:1144–52.
[49] Miyake T, Kasai M, Zhu SE, Sakurai T, Machida T. Vitrification
of mouse oocytes and embryos at various stages of development
in an ethylene glycol-based solution by a simple method.
Theriogenology 1993;40:121–34.
[50] Vicente JS, Garcia-Ximenez F. Osmotic and cryoprotective
effects of a mixture of DMSO and ethylene glycol on rabbit
morulae. Theriogenology 1994;42:1205–15.
[51] Isachenko V, Montag M, Isachenko E, Dessole S, Nawroth F, van
der Ven H. Aseptic vitrification of human germinal vesicle
oocytes using dimethyl sulfoxide as a cryoprotectant. Fertil
Steril 2006;85:741–7.

M.K. Gupta et al. / Theriogenology 67 (2007) 238–248248
[52] Cetin Y, Bastan A. Cryopreservation of immature bovine oocytes
by vitrification in straws. Anim Reprod Sci 2006;92:29–36.
[53] Trounson A, Kirby C. Problems in the cryopreservation of
unfertilized eggs by slow cooling in dimethyl sulfoxide. Fertil
Steril 1989;52:778–86.
[54] Johnson MH, Pickering SJ. The effect of dimethylsulphoxide on
the microtubular system of the mouse oocyte. Development
1987;100:313–24.
[55] Matson PL, Graefling J, Junk SM, Yovich JL, Edirisinghe RW.
Cryopreservation of oocytes and embryos: use of a mouse model
to investigate effects upon zona hardness and formulate treat-
ment strategies in an in vitro fertilization programme. Hum
Reprod 1997;12:1550–3.
[56] Vincent C, Turner K, Pickering SJ, Johnson MH. Zona pellucida
modifications in the mouse in the absence of oocyte activation.
Mol Reprod Dev 1991;28:394–404.





























