Embed Size (px)
Citation preview

COMMENTARY
Cryo-EM of viruses and vaccine designLesley A. Earla and Sriram Subramaniama,1
Knowledge of the atomic resolution structures ofviruses can be a powerful tool for vaccine discoveryand design. X-ray crystallography has long served asan invaluable method for virus structure determinationat high resolution, but over the past decade, cryo-electron microscopy (cryo-EM) has begun to emergeas a complementary method that can also provide thiskind of information. Crystallographic approaches re-quire significant amounts of purified virus that isstructurally homogeneous. In instances where produc-tion of adequate amounts of purified virus can bechallenging, or where homogeneity is difficult to achieve,the use of cryo-EM and image averagingmay be the onlyway to obtain structural information at high resolution. InPNAS, Rossmann and co-workers (1) use cryo-EM tech-niques to present elegant results on the structure deter-mination of rhinovirus C, a strain of rhinovirus that isassociated with severe disease in children with asthma(2). Rhinoviruses of A, B, and C categories are the leadingcause of common colds; there are well over 100 of thesetypes of viruses. Efforts to understand the molecularmechanism of cell receptor binding and differencesbetween rhinovirus strains have been hampered by thelack of structural information, especially for those strainsthat are difficult to propagate in cell culture (3).
It was almost two decades ago that the firstbreakthroughs were made in the use of cryo-EM forvirus structure determination, with the report ofstructures at ∼7 Å resolution of the hepatitis B capsid(4, 5). A decade later, the first near-atomic resolu-tion maps of icosahedral viruses, derived solely fromcryo-EM, were published (6, 7). Since that time, as meth-ods for high-resolution cryo-EM have become morewidely used, dozens of near-atomic resolution struc-tures of icosahedral viruses have been deposited inthe Electron Microscopy Data Bank (www.ebi.ac.uk/pdbe/emdb). In addition to the present study (1),the same group reported earlier this year (8) thenear-atomic resolution structure of Zika virus, a path-ogen that has recently been emerging as a major pub-lic health threat (9). Icosahedral viruses, including thepicornavirus and flavivirus families (to which rhinovirusand Zika virus belong, respectively), are uniquely
suited to rapid structure determination by cryo-EM,as explained below.
Three-dimensional structure determination bycryo-EM involves averaging the information presentin 2D projection images of multiple copies of individ-ual particles, which are oriented variably with respectto the incident electron beam. In the case of icosahedralviruses, althougheach individual virioncanbeconsideredaparticle, the symmetry of these structures is such thatmanyidentical copies of the asymmetrical unit are includedin each particle, increasing by many fold the effectivenumber of units averaged to determine the structure. Theintermolecular interactionsbetween the asymmetrical unitsof the icosahedral capsid provide a level of order thatincreases the accuracy of alignment, a feature also relevantfor viruses with helical capsids (10, 11). In contrast, theouter membranes of enveloped viruses, such as influenza,Ebola, and HIV, display their envelope proteins in a highlydisordered arrangement, and structure determination ofenvelope glycoproteins requires the use of subvolumeaveraging methods, which have only produced low-reso-lution structures so far (12, 13).
The study by Liu et al. (1) on rhinovirus C offers aparticularly intriguing look at the potential future ofstructure determination for vaccine design for emerg-ing pathogens. Even after overcoming the initial hur-dle of virus propagation in tissue culture, the team stillhad to work with a relatively low concentration of thevirus purified only through sucrose cushion sedimenta-tion. This relatively quick preparation yielded prepara-tions with a modest number of virus particles as well as amixture of both normal infectious virions and empty par-ticles (which contained only the capsid shell without viralRNA). From this mixed sample, the authors were able tosort the full and empty particles computationally and todetermine the structures of both to near-atomic reso-lution. A notable technical aspect of the work is that theauthors compensated for the lower concentration byworking with lower electron optical magnifications thanare typically used, so that more virions were present in agiven field of view. The use of lower magnifications canintroduce significant image distortions, but the authorswere successful in compensating for these defects. Not
aLaboratory of Cell Biology, Center for Cancer Research, National Cancer Institute, NIH, Bethesda, MD 20892Author contributions: L.A.E. and S.S. wrote the paper.The authors declare no conflict of interest.See companion article on page 8997.1To whom correspondence should be addressed. Email: [email protected].
www.pnas.org/cgi/doi/10.1073/pnas.1609721113 PNAS | August 9, 2016 | vol. 113 | no. 32 | 8903–8905
CO
MM
ENTARY
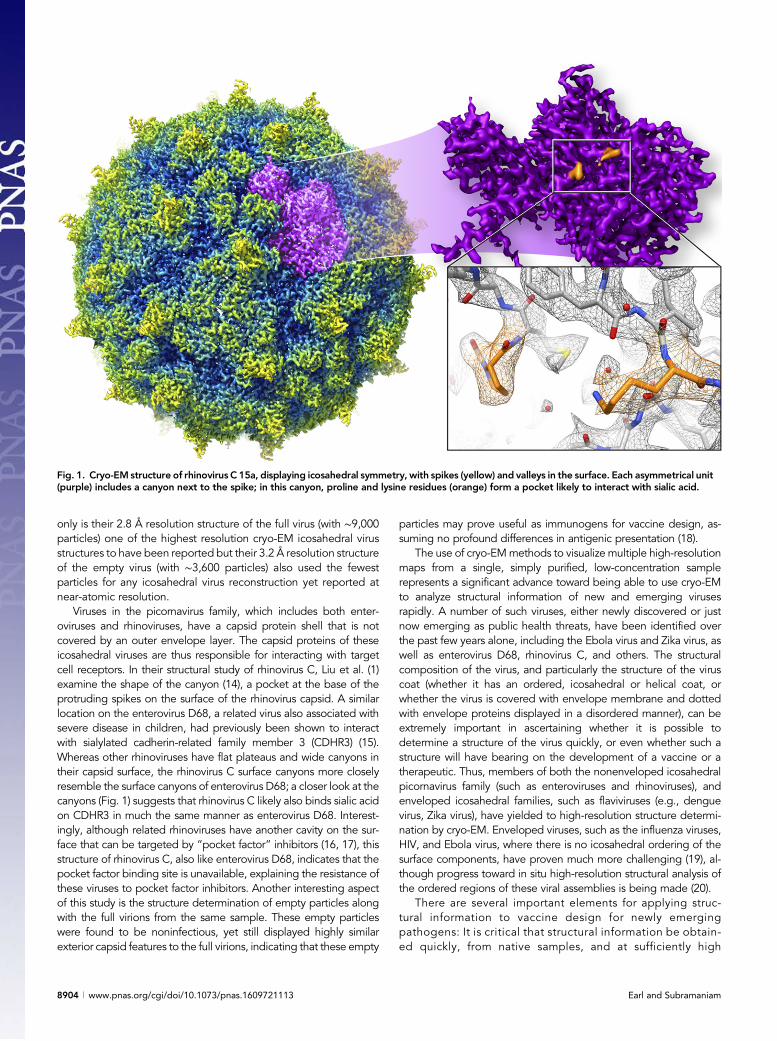
only is their 2.8 Å resolution structure of the full virus (with ∼9,000particles) one of the highest resolution cryo-EM icosahedral virusstructures to have been reported but their 3.2 Å resolution structureof the empty virus (with ∼3,600 particles) also used the fewestparticles for any icosahedral virus reconstruction yet reported atnear-atomic resolution.
Viruses in the picornavirus family, which includes both enter-oviruses and rhinoviruses, have a capsid protein shell that is notcovered by an outer envelope layer. The capsid proteins of theseicosahedral viruses are thus responsible for interacting with targetcell receptors. In their structural study of rhinovirus C, Liu et al. (1)examine the shape of the canyon (14), a pocket at the base of theprotruding spikes on the surface of the rhinovirus capsid. A similarlocation on the enterovirus D68, a related virus also associated withsevere disease in children, had previously been shown to interactwith sialylated cadherin-related family member 3 (CDHR3) (15).Whereas other rhinoviruses have flat plateaus and wide canyons intheir capsid surface, the rhinovirus C surface canyons more closelyresemble the surface canyons of enterovirus D68; a closer look at thecanyons (Fig. 1) suggests that rhinovirus C likely also binds sialic acidon CDHR3 in much the same manner as enterovirus D68. Interest-ingly, although related rhinoviruses have another cavity on the sur-face that can be targeted by “pocket factor” inhibitors (16, 17), thisstructure of rhinovirus C, also like enterovirus D68, indicates that thepocket factor binding site is unavailable, explaining the resistance ofthese viruses to pocket factor inhibitors. Another interesting aspectof this study is the structure determination of empty particles alongwith the full virions from the same sample. These empty particleswere found to be noninfectious, yet still displayed highly similarexterior capsid features to the full virions, indicating that these empty
particles may prove useful as immunogens for vaccine design, as-suming no profound differences in antigenic presentation (18).
The use of cryo-EMmethods to visualize multiple high-resolutionmaps from a single, simply purified, low-concentration samplerepresents a significant advance toward being able to use cryo-EMto analyze structural information of new and emerging virusesrapidly. A number of such viruses, either newly discovered or justnow emerging as public health threats, have been identified overthe past few years alone, including the Ebola virus and Zika virus, aswell as enterovirus D68, rhinovirus C, and others. The structuralcomposition of the virus, and particularly the structure of the viruscoat (whether it has an ordered, icosahedral or helical coat, orwhether the virus is covered with envelope membrane and dottedwith envelope proteins displayed in a disordered manner), can beextremely important in ascertaining whether it is possible todetermine a structure of the virus quickly, or even whether such astructure will have bearing on the development of a vaccine or atherapeutic. Thus, members of both the nonenveloped icosahedralpicornavirus family (such as enteroviruses and rhinoviruses), andenveloped icosahedral families, such as flaviviruses (e.g., denguevirus, Zika virus), have yielded to high-resolution structure determi-nation by cryo-EM. Enveloped viruses, such as the influenza viruses,HIV, and Ebola virus, where there is no icosahedral ordering of thesurface components, have proven much more challenging (19), al-though progress toward in situ high-resolution structural analysis ofthe ordered regions of these viral assemblies is being made (20).
There are several important elements for applying struc-tural information to vaccine design for newly emergingpathogens: It is critical that structural information be obtain-ed quickly, from native samples, and at sufficiently high
Fig. 1. Cryo-EM structure of rhinovirus C 15a, displaying icosahedral symmetry, with spikes (yellow) and valleys in the surface. Each asymmetrical unit(purple) includes a canyon next to the spike; in this canyon, proline and lysine residues (orange) form a pocket likely to interact with sialic acid.
8904 | www.pnas.org/cgi/doi/10.1073/pnas.1609721113 Earl and Subramaniam

resolution to shed light on immunogenic surfaces of the virus.With this study, Liu et al. (1) have shown that cryo-EM can beused to determine a structure quickly, even when the bio-chemistry involved in virus preparation can be challenging.Although these methods may not be generally applicable toevery emerging pathogen, cryo-EM is likely to increasingly
become a routine tool to aid rapid vaccine design, especiallyfor icoshedrally symmetric viruses.
AcknowledgmentsWe thank Veronica Falconieri for assistance with production of the figure. Theauthors’ research is supported by funds from the Center for Cancer Research,National Cancer Institute, at the National Institutes of Health.
1 Liu Y, et al. (2016) Atomic structure of a rhinovirus C, a virus species linked to severe childhood asthma. Proc Natl Acad Sci USA 113:8997–9002.2 Bizzintino J, et al. (2011) Association between human rhinovirus C and severity of acute asthma in children. Eur Respir J 37(5):1037–1042.3 Bochkov YA, et al. (2011) Molecular modeling, organ culture and reverse genetics for a newly identified human rhinovirus C. Nat Med 17(5):627–632.4 Böttcher B, Wynne SA, Crowther RA (1997) Determination of the fold of the core protein of hepatitis B virus by electron cryomicroscopy.Nature 386(6620):88–91.5 Conway JF, et al. (1997) Visualization of a 4-helix bundle in the hepatitis B virus capsid by cryo-electron microscopy. Nature 386(6620):91–94.6 Yu X, Jin L, Zhou ZH (2008) 3.88 A structure of cytoplasmic polyhedrosis virus by cryo-electron microscopy. Nature 453(7193):415–419.7 Zhang X, et al. (2008) Near-atomic resolution using electron cryomicroscopy and single-particle reconstruction. Proc Natl Acad Sci USA 105(6):1867–1872.8 Sirohi D, et al. (2016) The 3.8 Å resolution cryo-EM structure of Zika virus. Science 352(6284):467–470.9 Fauci AS, Morens DM (2016) Zika virus in the Americas–yet another arbovirus threat. N Engl J Med 374(7):601–604.
10 DiMaio F, et al. (2015) Virology. A virus that infects a hyperthermophile encapsidates A-form DNA. Science 348(6237):914–917.11 DiMaio F, et al. (2015) The molecular basis for flexibility in the flexible filamentous plant viruses. Nat Struct Mol Biol 22(8):642–644.12 Liu J, Bartesaghi A, Borgnia MJ, Sapiro G, Subramaniam S (2008) Molecular architecture of native HIV-1 gp120 trimers. Nature 455(7209):109–113.13 Huiskonen JT, et al. (2014) Averaging of viral envelope glycoprotein spikes from electron cryotomography reconstructions using Jsubtomo. J Vis Exp (92):e51714.14 Smith TJ, Chase ES, Schmidt TJ, Olson NH, Baker TS (1996) Neutralizing antibody to human rhinovirus 14 penetrates the receptor-binding canyon. Nature
383(6598):350–354.15 Liu Y, et al. (2015) Sialic acid-dependent cell entry of human enterovirus D68. Nat Commun 6:8865.16 Liu Y, et al. (2015) Structure and inhibition of EV-D68, a virus that causes respiratory illness in children. Science 347(6217):71–74.17 De Colibus L, et al. (2014) More-powerful virus inhibitors from structure-based analysis of HEV71 capsid-binding molecules. Nat Struct Mol Biol 21(3):282–288.18 Kotecha A, et al. (2015) Structure-based energetics of protein interfaces guides foot-and-mouth disease virus vaccine design. Nat Struct Mol Biol 22(10):788–794.19 Tran EE, et al. (2016) Cryo-electron microscopy structures of chimeric hemagglutinin displayed on a universal influenza vaccine candidate. MBio 7(2):e00257.20 Schur FK, et al. (July 14, 2016) An atomic model of HIV-1 capsid-SP1 reveals structures regulating assembly and maturation. Science, 10.1126/science.aaf9620.
Earl and Subramaniam PNAS | August 9, 2016 | vol. 113 | no. 32 | 8905





























