Embed Size (px)
Citation preview

Cross-bred Cattle Protected against Hyalomma anatolicumanatolicum by Larval Antigens Puri¢ed by Immunoa¤nityChromatography
S. Ghosh1, M.H. Khan1 and N. Ahmed21Entomology Laboratory, Division of Parasitology; 2Protein Chemistry Laboratory,Division of Biochemistry, Indian Veterinary Research Institute, Izatnagar, 243 122, UttarPradesh, India
Ghosh, S., Khan, M.H. and Ahmed, N., 1999. Cross-bred cattle protected against Hyalomma anatolicumanatolicum by larval antigens puri¢ed by immunoa¤nity chromatography. Tropical Animal Health andProduction, 31(5), 263^273
ABSTRACT
Antigens from larvae of Hyalomma anatolicum anatolicum were extracted and puri¢ed by immunoa¤-nity chromatography using immunoglobulin ligands from cross-bred animals immunized with solublelarval antigen. A¤nity-puri¢ed antigen (A¡-TLE) and a total larval extract (TLE) were used toimmunize cross-bred (Bos indicus6Bos taurus) cattle. The group immunized with A¡-TLE rejected71.6% of larvae and 77.3% of nymphs. However, the rejection percentages were lower in the TLE-immunized group. No signi¢cant changes in the feeding period, moulting percentages or moultingperiod of engorged larvae and nymphs were recorded. There was, however, a signi¢cant decrease in thenumber of resultant nymphs p50.01) and adults (p50.01) in the ticks fed on the A¡-TLE-immunizedgroup. The A¡-TLE antigen was 93.3% puri¢ed. SDS-PAGE analysis identi¢ed a 39 kDa protein,reported for the ¢rst time, as the antigen responsible for the induction of resistance in the host.
Keywords: antigen, cattle, Hyalomma anatolicum anatolicum, immunization, immunoa¤nity, larvae,protection, 39 kDa protein
Abbreviations: A¡-TLE, a¤nity-puri¢ed antigen; ELISA, enzyme-linked immunosorbent assay; IFA,incomplete Freund's adjuvant; SDS-PAGE, sodium dodecyl sulphate polyacrylamide gel electrophor-esis; TLE, total larval extract
INTRODUCTION
The development of anti-tick vaccines is an area of research with considerable potentialto minimize acaricidal hazards, resistance and residues and environment pollution.Signi¢cant progress has been made in developing a vaccine against Boophilusmicroplus, a one-host tick, and recombinant vaccines have been released for commer-cial production in Australia (Willadsen et al., 1995) and in Cuba (Canales et al., 1997).However, very little information is available on arti¢cial immunization against the nextimportant tick species to be studied, Hyalomma anatolicum anatolicum, the vector ofTheileria annulata. As reported by Johnston and colleagues (1986), Opdebeeck andcolleagues (1988) and Kimaro and Opdebeeck (1994) for B. microplus, crude antigensof H.a. anatolicum have been found to be protective against the homologous species
Tropical AnimalHealth and Production, 31 (1999) 263^273# 1999 Kluwer Academic Publishers. Printed in the Netherlands
263

(Sran et al., 1996; Ghosh et al., 1998). Nevertheless, no attempts have been directedtowards the puri¢cation of these crude antigens. The immunochemical puri¢cation of alarval antigen was therefore attempted and the isolated fraction was used in animmunization protocol for inducing protection against this tick.
MATERIALS AND METHODS
Animals
Twenty new-born cross-bred (Bos indicus6Bos taurus) male calves with no previousexposure to ticks were procured from the Institute's dairy farm. These calves wereindividually housed on slatted £oors in tick-proof pens. The tick-naive status of thesecalves was con¢rmed by immunodi¡usion and enzyme-linked immunosorbent assaywith larval, nymphal and adult antigens of H.a. anatolicum. At the time of immuniza-tion, the calves were 12^16 months old. They were randomly divided into four groups(A, B, C and D) of 5 calves each.
Ticks
Tick colonies were propagated and maintained at the Entomology laboratory of thedivision by the methods described by Khan and colleagues (1982).
Antigen
Antigen extraction was done in an ice-bath and the antigens were stored at ^708C.Laboratory-reared, unfed larvae of H.a. anatolicum were triturated in 0.15 Mphosphate-bu¡ered saline (PBS), containing 1 mmol/L disodium EDTA and 1 mmol/L phenylmethanesulphonyl £uoride, pH 7.2, in a glass homogenizer, ¢ltered free ofcuticle and debris, and sonicated for 10 min in 60-second bursts interspersed withperiods of cooling (Opdebeeck et al., 1989). The homogenate was centrifuged at15 000g for 20 min at 48C. The supernatant was designated as total larval extract(TLE) and used to immunize the animals in group A.
Immunoa¤nity chromatography
Sera were collected from two animals immunized with a soluble larval antigen of H.a.anatolicum and 67^70% protected against challenge infestation (Ghosh and Khan,1999). Immunoglobulins were precipitated from these sera with 40% ammoniumsulphate (Fey et al., 1976). The IgG was further puri¢ed on a DEAE-sepharose column(Pharmacia Fine Chemicals, Sweden) that had previously been equilibrated withequilibrating bu¡er (Talwar and Gupta, 1992). The puri¢ed IgG was dialysed
264

extensively for 36 h at 48C against 0.1 M NaHCO3, 0.5 M NaCl, pH 8.43 and coupledto cyanogen bromide-activated sepharose 4B as recommended by Pharmacia FineChemicals. The excess reactive sites were blocked by 0.2 M glycine bu¡er, pH 8.0. Theprepared gel was pre-eluted (0.1 M glycine-HCl, pH 2.8) and equilibrated with bu¡ercontaining 10 mmol/L sodium phosphate and 5 mmol/L disodium EDTA, pH 7.2before use. The column was regenerated by using 0.1M Tris-HCl, 0.5MNaCl, pH 8.5;0.1 M sodium acetate, 0.5 MNaCl, pH 4.5 after each use. TLE (200 mg total proteins)was loaded at 15 ml/h on to the a¤nity column, which was then washed withequilibrating bu¡er at a £ow rate of 30 ml/h. Bound antigens were eluted with 0.1 Mglycine-HCl, pH 2.8, into Tris-base to increase the pH to 7.2. The eluted previouslybound antigens were dialysed extensively against phosphate bu¡er, concentrated withpolyethylene glycol 20000 and designated as A¡-TLE. The protein content of theantigen was estimated by the method of Lowry and colleagues (1951).
Immunization
All the calves in group A were immunized with TLE and those in group C wereimmunized with A¡-TLE. The calves in groups B and D were kept as controls forgroups A and C, respectively. The immunization schedule, dose and route of deliveryare given in Table I.
TABLE ISchedule of immunization of cattle with larval extract and a¤nity-puri¢ed antigens
Groupa
Time A B C DImmunization (days) Adjuvant (mg antigen protein/animal) Route
Primary immunization 0 IFA 12.5 Nil 0.8 Nil SubcutaneousFirst booster 14 IFA 12.5 Nil 0.8 Nil SubcutaneousSecond booster 28 None 5.0 Nil 0.4 Nil Intramuscular
aGroup A, calves immunized with TLE; group C, calves immunized with A¡-TLE; groups B and D,unimmunized control calves, which received injections of bu¡er, with or without IFA adjuvant as indicated
265

Challenge infestations
Ten days after the last immunization, all the animals were challenged twice, 10 daysapart, with 2000 larvae and 100 nymphs per infestation. Live larvae and nymphscollected after completing their feeding on these animals were counted. Anti-tickimmunity was assessed by calculating the mean percentage recovery of naturallydropped larvae/nymphs at the end of the feeding period. The percentage recovery ofdropped stages was calculated as:
Number of fed larvae/nymphs droppedööööööööööööööööö 6 100
Number of larvae/nymphs released
The percentage rejection was calculated as 100 ^ percentage recovery. The droppedlarvae/nymphs were grouped as abnormally fed and normally fed. After counting, allthe batches were maintained separately.
Sera samples
Blood was collected from all the animals; sera were separated and diluted 1:800, andthe anti-tick antibodies in themwere detected by ELISA, using tick antigen at 15 mg/mlin carbonate bicarbonate bu¡er, pH 9.6. Peroxidase-conjugated rabbit anti-bovine IgG(Sigma Chemical Company, USA) was used and OD values were measured at 492 nm(Ghosh and Khan, 1997a).
Polyacrylamide gel electrophoresis
All the antigens were resolved by SDS-PAGE on 1.5 mm thick gel using a discontin-uous system (Laemmli, 1970). The stacking gel was 3% acrylamide in 0.5 M Tris, pH6.8 with 0.25% SDS and the resolving gel was 10% acrylamide in 1.5 M Tris, pH 8.8with 0.25% SDS. Gels were stained with Coomassie Brilliant Blue R-250 to identify themarker proteins, which ranged from 97.4 to 29 kDa (Pharmacia Fine Chemicals).
Statistical analysis
Signi¢cant di¡erences in mean values from immunized and control animals weredetermined using Student's t-test (Snedecor and Cochran, 1968).
266

RESULTS
Immunoa¤nity chromatography
IgG was bound at 10 mg per ml of CNBr activated sepharose 4B. A total of 200 mgTLE was loaded in batches and 11.2^12.0 of bound protein was eluted.
Attachment and feeding behaviour of larvae
Table II shows the e¡ect of immunization on the larvae. In all the animals, the larvaehad attached within 24 h of their release. No skin reactions were noted at the tickfeeding sites in the animals in the control groups but small papules and exudation werenoted in the animals in the immunized groups. Larvae were seen dying in situ within 42h of attachment in the calves immunized with TLE antigen, whereas mortality wasnoted within 24 h of attachment in the group immunized with A¡-TLE antigen. Themean percentage rejection in group A was higher than that in the control group B butnot signi¢cantly so. However, the percentage rejection in the animals immunized withA¡-TLE (group C) was signi¢cantly (p50.01) higher than that in the respective controlgroup D. Among the attached larvae, 5^8% of abnormally fed larvae were recoveredfrom the animals in group C. These larvae were white to pale yellow in colour, incontrast to normally fed larvae, which were dark grey in colour. No signi¢cantdi¡erences in the feeding period, moulting percentages or moulting period, or in theoverall percentage decrease in resultant nymphal stages were recorded in the ticks fedon the cattle in group A compared with those fed on animals in group B. However,there was a signi¢cant di¡erence (p50.01) in the overall percentage of resultantnymphs from the ticks fed on the animals in group C compared with those on theanimals in group D.
TABLE IIFeeding and development performance (mean+SE) of larvae of Hyalomma fed on immunizedand control calves
OverallFeeding Moulting percentage
Rejection period Moulting period decrease inGroupsa percentage (days) percentage (days) resultant nymphs
A 38.5+6.7 4.8+0.2 47.2+9.5 13.0+0.3 79.5+9.2B 20.0+10.1 5.2+0.6 54.8+7.3 11.0+0.7 56.2+8.0C 71.6+8.8b 5.4+0.2 12.7+5.2 12.1+0.4 91.3+8.6b
D 11.9+2.2b 3.2+0.4 48.1+18.6 10.5+0.9 57.6+6.5b
aGroups A and C, immunized with TLE and A¡-TLE, respectively; groups B and D, unimmunized controlsbSigni¢cant at p50.01
267
Attachment and feeding behaviour of nymph
The e¡ect on nymphs of immunization of the host is summarized in Table III. In theanimals in group C, a hypersensitivity reaction was observed at the bite sites of manyattached nymphs. The di¡erence in percentage rejection was signi¢cantly higher(p50.05) in group C than in the corresponding control group D. Of the attachednymphs in groups A and C, 4.2%+1.2% and 6.2%+1.0%, respectively, wereabnormally fed, characterized by their white to pale yellow colour. As in the case ofthe larval challenge, immunization of the hosts did not interfere with the feedingperiod, moulting percentages, moulting period of nymphs or percentage decrease inresultant adults except in group C animals, where there was a signi¢cant (p50.01)decrease in resultant adults compared with those in group D.
Antibody response to TLE and A¡-TLE antigens
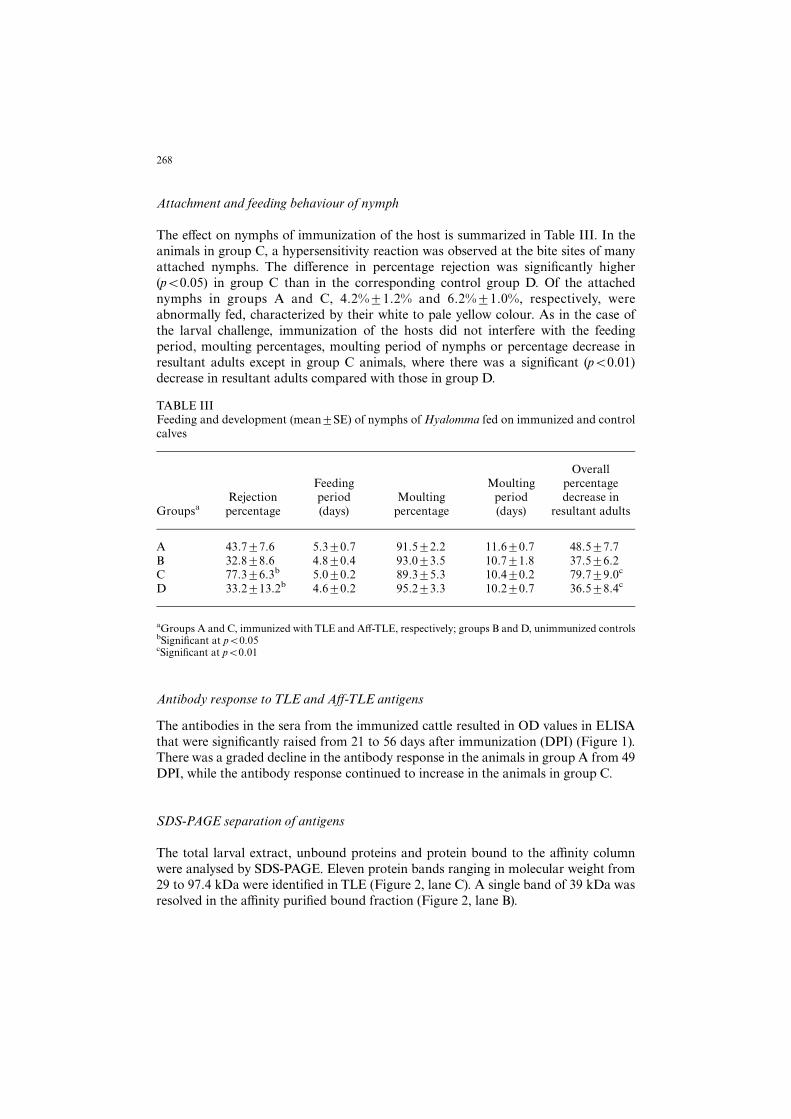
The antibodies in the sera from the immunized cattle resulted in OD values in ELISAthat were signi¢cantly raised from 21 to 56 days after immunization (DPI) (Figure 1).There was a graded decline in the antibody response in the animals in group A from 49DPI, while the antibody response continued to increase in the animals in group C.
SDS-PAGE separation of antigens
The total larval extract, unbound proteins and protein bound to the a¤nity columnwere analysed by SDS-PAGE. Eleven protein bands ranging in molecular weight from29 to 97.4 kDa were identi¢ed in TLE (Figure 2, lane C). A single band of 39 kDa wasresolved in the a¤nity puri¢ed bound fraction (Figure 2, lane B).
TABLE IIIFeeding and development (mean+SE) of nymphs of Hyalomma fed on immunized and controlcalves
OverallFeeding Moulting percentage
Rejection period Moulting period decrease inGroupsa percentage (days) percentage (days) resultant adults
A 43.7+7.6 5.3+0.7 91.5+2.2 11.6+0.7 48.5+7.7B 32.8+8.6 4.8+0.4 93.0+3.5 10.7+1.8 37.5+6.2C 77.3+6.3b 5.0+0.2 89.3+5.3 10.4+0.2 79.7+9.0c
D 33.2+13.2b 4.6+0.2 95.2+3.3 10.2+0.7 36.5+8.4c
aGroups A and C, immunized with TLE and A¡-TLE, respectively; groups B and D, unimmunized controlsbSigni¢cant at p50.05cSigni¢cant at p50.01
268
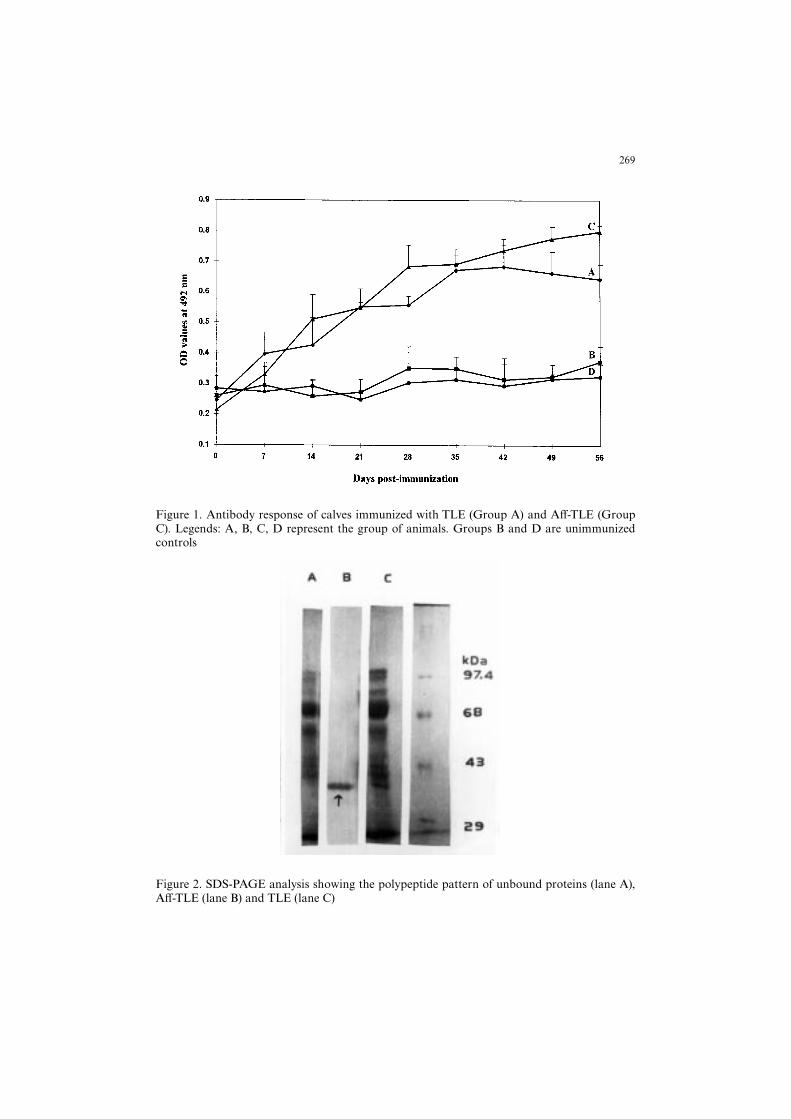
Figure 1. Antibody response of calves immunized with TLE (Group A) and A¡-TLE (GroupC). Legends: A, B, C, D represent the group of animals. Groups B and D are unimmunizedcontrols
Figure 2. SDS-PAGE analysis showing the polypeptide pattern of unbound proteins (lane A),A¡-TLE (lane B) and TLE (lane C)
269

DISCUSSION
Attempts have been made earlier to immunize laboratory animals (Manohar andBanerjee, 1992; Ghosh et al., 1998) and cross-bred calves (Banerjee et al., 1990; Sranet al., 1996; Ghosh and Khan, 1999) against H.a. anatolicum using crude antigensprepared from di¡erent tissues of the tick, with some encouraging results. However,crude homogenates contain a mixture of antigens, most of which may not beresponsible for protective immunity. Identi¢cation of the particular antigen(s) respon-sible for protective immunity to H.a. anatolicum can only be con¢rmed by puri¢cationof the antigens to homogeneity. Although some immunodominant antigens had beenidenti¢ed in salivary gland extracts (Gill et al., 1986; Parmar et al., 1996) and inhomogenates of larvae, nymphs and adults of H.a. anatolicum (Ghosh et al., 1998),these immunodominant antigens have not been further utilized to develop a betterimmunoprotective response. In the present experiment, unfed larval extracts of H.a.anatolicum have been puri¢ed for the ¢rst time by immunoa¤nity chromatography,using immunoglobulin ligands from cattle immunized with soluble larval antigens and65^70% protected against larval challenge (Ghosh and Khan, 1999). Using the puri¢edantigen, signi¢cantly higher percentages of protection against both larvae (71.6%) andnymphs (77.3%) were recorded in the present experiment. In contrast, using unpuri¢edextracts of salivary glands and soluble larval antigen, Sran and colleagues (1996) andGhosh and Khan (1999) obtained 55.8% and 57.25% rejection of larvae, respectively.The percentage rejection of larvae observed using A¡-TLE as antigen was thusconsiderably higher than that with any of the other antigens tested so far. Nosigni¢cant changes were recorded in other parameters in the TLE antigen-immunizedgroup, but a signi¢cantly (p50.01) higher percentage reduction in resultant nymphswas noted in the ticks fed on the animals in group C. Earlier, Essuman and colleagues(1991) reported comparatively low percentages (38%) of moulting by larvae fed oncalves immunized with midgut extracts of Rhipicephalus appendiculatus. In contrast,Sangwan and colleagues (1998) reported unaltered moulting rates in larvae fed oncalves immunized with a nymphal antigen of H.a. anatolicum.
Rejection of nymphs, like that of larvae, was signi¢cantly greater in the immunizedgroups than in the control groups and again the percentage rejection was 33.6% higherin group C than in group A. As reported earlier by Sangwan and colleagues (1998), thefeeding period, moulting period and percentage moulting of nymphs remainedunaltered in all the animals. It is interesting to note that, as with the larvae, theimmunoa¤nity-puri¢ed antigen signi¢cantly interfered with the moulting process ofnymphs. This resulted in an 80% reduction in the number of resultant adults. The whiteto pale yellow colour of the abnormally fed larvae and nymphs may be due to lack ofred blood cells in their feed. This may be of considerable importance in decreasing thetransmission of the parasite, Theileria annulata.
In most experiments with multihost ticks, the challenge has been with same stagefrom which the immunizing extracts were made, although Allen and Humphreys(1979) reported that antigens derived from unfed ticks were ine¡ective as a vaccinecompared to antigens from partly fed adult ticks. However, antigens from unfedimmature stages have been shown to be protective in various tick^host systems
270

(Heller-Haupt et al., 1987; Opdebeeck et al., 1989; Varma et al., 1990; Ghosh andKhan, 1997b, 1999; Ghosh et al., 1998). The present experiment con¢rmed that unfedlarvae of H.a. anatolicum provide a relatively easily available source of antigen forimmunization of cattle against both larvae and nymphs of this species.
Tick antigens have been puri¢ed previously by immunoa¤nity chromatographyusing di¡erent immunoglobulin ligands (Opdebeeck et al., 1989; Wong and Opde-beeck, 1990) and up to 99.9% puri¢cation has been achieved (Brown and Askenase,1986). These a¤nity-puri¢ed antigens were found to be signi¢cantly protective againstchallenge infestations with Amblyomma americanum. In the present experiment usinganti-larval immunoglobulin ligands, the crude extracts were about 93% puri¢ed byimmunoa¤nity chromatography. The study demonstrated for the ¢rst time that a 39kDa immunoa¤nity-puri¢ed protein antigen was responsible for the induction ofimmunity to larvae and nymphs of H.a. anatolicum. Subsequent studies are beingdirected towards characterizing the protein so as to facilitate the use of an mRNAisolation procedure leading to the application of recombinant DNA technology and invitro expression of this protein.
ACKNOWLEDGEMENT
Sincere thanks are due to the Director, Indian Veterinary Research Institute forproviding laboratory facilities.
REFERENCES
Allen, J.R. and Humphreys, S.J., 1979. Immunization of guinea pigs and cattle against ticks. Nature, 280,491^495
Banerjee, D.P., Momin, R.R. and Samantaray, S., 1990. Immunization of cattle (Bos indicus6Bos taurus)against Hyalomma anatolicum anatolicum using antigens derived from tick salivary gland extracts.International Journal for Parasitology, 20, 969^972
Brown, S.J. and Askenase, P.W., 1986. Amblyomma americanum: physiochemical isolation of a proteinderived from the tick salivary gland that is capable of inducing immune resistance in guinea pig.Experimental Parasitology, 62, 40^50
Canales, M., Enriquez, A., Ramos, E., Cabrera, D., Dandie, H., Soto, A., Falcon,V., Rodriguez, M. and dela Fuente, J., 1997. Large-scale production in Pichia pastoris of the recombinant vaccine GavacTM
against cattle tick.Vaccine, 15, 414^422Essuman, S., Dipeolu, O.O. and Odhiamba, T.R., 1991. Immunization of cattle with a semi-puri¢ed
fraction of solubilized membrane bound antigens extracted from the midgut of the tick Rhipicephalusappendiculatus. Experimental and Applied Acarology, 13, 65^73
Fey, H., Phster, H., Messerli, J., Sturzenegger, N. and Grolimund, F., 1976. Methods of isolation,puri¢cation and quantitation of bovine immunoglobulins. A technical review. Zentralblatt fu« r Veterina« rMedizin, 23, 269
Ghosh, S. and Khan, M.H., 1997a. Identi¢cation of tick antigen immunogenic in calves. InternationalJournal of Animal Sciences, 12, 249^252
Ghosh, S. and Khan, M.H., 1997b. Immunization of cattle against tick Boophilus microplus. Indian Journalof Animals Sciences, 67, 183^186
Ghosh, S. and Khan, M.H., 1999. Immunization of cattle against Hyalomma anatolicum anatolicum usinglarval antigens. Indian Journal of Experimental Biology, 37, 203^205
271

Ghosh, S., Khan, M.H. and Gupta, S.C., 1998. Immunization of rabbits against Hyalomma anatolicumanatolicum using homogenates from unfed immature ticks. Indian Journal of Experimental Biology, 26,167^170
Gill, H.S., Boid, R. and Ross, C.A., 1986. Isolation and characterization of salivary antigens fromHyalomma anatolicum anatolicum. Parasite Immunology, 8, 11^25
Heller-Haupt, A., Varma, M.G.R., Rechav, Y., Langi, A.O. and Trinder, P.K.E., 1987. Immunization oflaboratory animals against the tick, Amblyomma variegatum using homogenates from unfed larvalticks. Medical Science Research, 15, 371^372
Johnston, L.A.Y., Kemp, D.H. and Pearson, R.D., 1986. Immunization of cattle against Boophilusmicroplus using extracts derived from female ticks: e¡ects of induced immunity on tick populations.International Journal for Parasitology, 16, 27^34
Khan, M.H., Naithani, R.C. and Singh (Kr.), S., 1982. Study on the bionomics of Nosomma monstrosum(Nuttal and Warburton) (Acari: Ixodidae). Acarologia, 23, 119^123.
Kimaro, E.E. and Opdebeeck, J.P., 1994. Tick infestation on cattle vaccinated with extracts from the eggand the gut of Boophilus microplus.Veterinary Parasitology, 52, 61^70
Laemmli, U.K., 1970. Cleavage of structural proteins during assembly of the head of bacteriophage T4.Nature, 227, 680^685
Lowry, D.H., Rosebrough, N.J., Farr, A.L. and Randall, R.J., 1951. Protein measurement with the Folin-phenol reagent. Journal of Biological Chemistry, 193, 265^275
Manohar, G.S. and Banerjee, D.P., 1992. Immune response in rabbits immunised against the tick,Hyalomma anatolicum anatolicum. Indian Journal of Animal Sciences, 62, 505^512
Opdebeeck, J.P., Wong, J.Y.M., Jackson, L.A. and Dobson, C., 1988. Hereford cattle immunized andprotected against Boophilus microplus with soluble and membrane associated antigens from the midgutof ticks. Parasite Immunology, 10, 405^410
Opdebeeck, J.P., Wong, J.Y.M. and Dobson, C., 1989. Hereford cattle protected against Boophilusmicroplus with antigens puri¢ed by immunoa¤nity chromatography from larval and adult ticks.Immunology, 67, 388^393
Parmar, A., Grewal, A.S. and Dhillon, P., 1996. Immunological cross-reactivity between salivary glandproteins of Hyalomma anatolicum anatolicum and Boophilus microplus. Veterinary Immunology andImmunopathology, 51, 345^352
Sangwan, A.K., Banerjee, D.P. and Sangwan, A.K., 1998. Immunization of cattle with nymphalHyalommaanatolicum anatolicum extracts: e¡ects on tick biology. Tropical Animal Health and Production, 30, 97^106
Snedecor, G.W. and Cochran,W.B., 1968. Statistical Methods, (Oxford and IBH Publishing Co., Calcutta,India)
Sran, H.S., Grewal, A.S. and Kondal, J.K., 1996. Enhanced immunity toHyalomma anatolicum anatolicumticks in cross-bred (Bos indicus6Bos taurus) calves using ascaris extract immunomodulator with ticksalivary gland extract antigens.Veterinary Immunology and Immunopathology, 51, 333^343
Talwar, G.P. and Gupta, S.K., 1992. Handbook of Practical and Clinical Immunology, (CBS Publishers,Delhi, India)
Varma, M.G.R., Heller-Haupt, A., Trinder, P.K.E. and Langi, A.O., 1990. Immunization of guinea pigsagainst Rhipicephalus appendiculatus adult ticks using homogenates from unfed immature ticks.Immunology, 71, 133^138
Willadsen, P., Bird, P., Cobon, G.S. and Hungerford, J., 1995. Commercialisation of a recombinantvaccine against Boophilus microplus. Parasitology, 110, 43^50
Wong, J.Y.M. and Opdebeeck, J.P., 1990. Larval membrane antigens protect Hereford cattle againstinfestation with Boophilus microplus. Parasite Immunology, 12, 75^83
(Accepted: 3 February 1999)
Protection du betail hybride contre la tique Hyalomma anatolicum anatolicum apre© s utilisation d'antige© neslarvaires puri¢es par chromatographie d'a¤nite
Resume ^ Des antige© nes des larves de H. a. anatolicum furent extraits et puri¢es par chromatographied'a¤nite en utilisant des ligands immunoglobuliniques d'animaux hybrides immunises avec des antige© neslarvaires solubles. Des antige© nes puri¢es par chromatographie d'a¤nite (A¡-TLE) et des extraits totaux delarves (TLE) furent utilises pour immuniser du betail hybride (Bos indicus6 Bos taurus).
272

Le groupe immunise avec les antige© nes A¡-TLE rejete© rent 71,6% des larves et 77,3% des nymphes. Lespourcentages de rejet furent plus faibles avec le groupe TLE.Il ne fut pas observe de changement signi¢catif pour le temps de prise de nourriture des tiques, le
pourcentage de tiques se metamorphosant ou pour le temps de la metamorphose pour les larves et lesnymphes engorgees de sang. Il y eut cependant une diminution signi¢cative du nombre de nymphes(p50,01) et des adultes (p50,01) pour les tiques s'etant nourries sur les animaux immunises avec lesantige© nes A¡-TLE. L'antige© ne A¡-TLE fut puri¢e a© 93,3%. Apre© s analyse par SDS-Page (sodium dodecylsulfate en gel de polyacrylamide), une proteine de 39 kDa fut identi¢ee comme l'antige© ne responsable del'induction d'une resistance aux tiques chez l'hoª te.
Ganado vacuno cruzado protegido frente a Hyalomma anatolicum anatolicum mediante ant|genos larvariospuri¢cados por cromatograf|a de inmunoa¢nidad
Resumen ^ Se obtuvieron ant|genos de larvas de Hyalomma anatolicum y se puri¢caron mediantecromatograf|a de inmunoa¢nidad utilizando como ligandos inmunoglobulinas procedentes de animalescruzados inmunizados con ant|genos larvarios solubles. Se utilizo ant|geno de a¢nidad puri¢cado (A¡-TLE) y extracto larvario total (TLE) para inmunizar ganado cruzado (Bos indicus6Bos taurus). El grupoinmunizado con A¡-TLE rechazo el 71,6% de las larvas y el 77,3% de las ninfas. No obstante, losporcentajes de rechazo fueron inferiores en el grupo inmunizado con TLE. No se encontraron diferenciassigni¢cativas en el per|odo de alimentacion, el porcentaje de muda o el per|odo de muda de las larvas y lasninfas. Sin embargo, se encontro una reduccion signi¢cativa en el numero de ninfas producidas (p50,01) yde adultos (p50,01) en las larvas que se alimentaron de animales inmunizados con A¡-TLE. El ant|genoA¡-TLE tuvo una pureza del 93,3%. Mediante un analisis SDS-PAGE se identi¢co una prote|na de 39 kDa(mencionada por primera vez en la literatura) como el ant|geno responsable de la induccion de resistenciaen el hospedador.
273








![Specific histamine binding activity of a new lipocalin ... · Hyalomma asiaticum isoneofthethreehardticks (Ixodidae) widely distributed in northwest China and Central Asia [20]. These](https://img.dokumen.tips/doc/110x75/60624fa973f38b61c170b279/specific-histamine-binding-activity-of-a-new-lipocalin-hyalomma-asiaticum-isoneofthethreehardticks.jpg)












![Lymphangitis-Associated Rickettsiosis, a New Rickettsiosis ...€¦ · jing from Hyalomma asiaticum ticks collected in the Alashian region of Inner Mongolia in 1991 [2]. In 1996,](https://img.dokumen.tips/doc/110x75/60624dd28679b45f690e193e/lymphangitis-associated-rickettsiosis-a-new-rickettsiosis-jing-from-hyalomma.jpg)







