Embed Size (px)
Citation preview

Le fonctionnement normal detout organisme multicellulai-re dépend de la coordinationdes cellules qui le composent.Cette coordination implique
un système de communication grâceauquel chaque cellule est informéede l’activité des cellules voisines etrègle en conséquence son propreniveau de fonctionnement. Les cel-lules des vertébrés communiquententre elles par de nombreux méca-nismes. Plusieurs de ceux-ci fontintervenir des signaux qui sont trans-mis à des cellules équipées de récep-teurs, canaux, enzymes ou voiesmétaboliques appropriés. Dans tous
ces cas, le transfert d’informationd’une cellule à l’autre se fait indirec-tement via la diffusion de l’ion ou dela molécule signal dans l’espaceintercellulaire. Cependant, le fonc-tionnement de nombreux tissus estpréservé dans des conditions qui per-turbent ou même abolissent le fluxde signaux intercellulaires, indiquantque d’autres mécanismes de commu-nication intercellulaire sont au moinsaussi importants. La perte du fonc-tionnement normal de certains tissusaprès isolement de leurs cellules sug-gère que ces mécanismes dépendentd’interactions directes entre cellulesvoisines.
909
Connexines, canauxjonctionnelset communicationscellulaires
La plupart des cellules animales synthétisent des protéinesintégrales de membrane, appelées connexines, dont la sé-quence et la topographie ont été très conservées au cours del’évolution. Ces protéines forment des canaux transmem-branaires, regroupés au niveau des jonctions intercellulairesde type gap, qui permettent à des cellules voisines d’échan-ger directement de nombreux ions et métabolites cytoplas-miques. Ce mécanisme de communication intercellulaire,aussi appelé couplage jonctionnel, a été impliqué dans denombreuses fonctions cellulaires, au cours du développe-ment embryonnaire et dans les tissus adultes. Des études ré-centes indiquent que des altérations des connexines pour-raient jouer un rôle considérable dans diverses maladies, enparticulier la neuropathie périphérique héréditaire de Char-cot-Marie-Tooth (mutations de la connexine 32) et l’hétéro-taxie viscéroatriale (mutations de la connexine 43).
ADRESSEP. Meda : docteur en médecine, professeur ad-joint. Département de Morphologie, Univer-sité de Genève, Centre Médical Universitai-re, 1, rue Michel-Servet, CH-1211 Genève4, Suisse.
SYNTHÈSEmédecine/sciences 1996 ; 12 : 909-20
m/s n° 8-9, vol. 12, août-septembre 96
Paolo Meda

La plupart des cellules animales encontact peuvent, en effet, échangerdes informations par le biais d’inter-actions entre molécules de surfaceappartenant à la famille desconnexines [1], et qui forment lesjonctions de type gap. Les connexinessont des protéines intégrales de lamembrane plasmique qui sont syn-thétisées pratiquement par chaquetype cellulaire, quelle que soit la posi-tion d’un organisme multicellulairedans la phylogenèse du monde ani-mal [2]. Chez les vertébrés, les rarescellules ne produisant pas deconnexines sont les cellules muscu-laires striées adultes, quelques neu-rones, les spermatozoïdes et les cel-lules circulantes du sang. A ce jour,au moins treize connexines (Cx) dis-tinctes [3, 4] ont été identifiées chezles mammifères (Tableau I).
Les gènes des connexines
Chez les mammifères, chaqueconnexine est codée par un gène dis-tinct [3, 4]. Cependant, l’existenced’une liaison génétique entreconnexines de structure primaire dif-férente, suggère que les treize gènesdes connexines dérivent d’une seuleséquence originale, encore indéter-minée, qui a subi au moins deuxduplications [2, 3] juste avant ou autout début de l’évolution des verté-brés (figure 1). Ces duplications ontpermis la divergence de deuxgroupes de protéines. Le groupe αcomprend les connexines compor-tant une boucle cytoplasmique cour-te, alors que le groupe β comprendles connexines comportant uneboucle cytoplasmique longue. Tous les gènes des connexines étu-diés à ce jour montrent une structuresimilaire [5, 6] comportant deuxexons séparés par un intron (figure 1).Le premier exon est petit et codepour une séquence 5’ non transcrite.L’intron varie en longueur d’un gèneà l’autre mais, dans tous les cas,contient un site accepteur d’épissageen amont du codon de début de tra-duction. Le deuxième exon est plusgrand et contient en un bloc toute laséquence codant pour la protéine.Les éléments régulateurs des régionspromotrices varient d’un gène àl’autre, expliquant sans doutel’expression différentielle de diversesconnexines. La région promotrice de
la Cx43 contient une boîte TATA, dessites AP-1 et AP-2 et des séquences deliaison d’œstrogènes [7]. La régionpromotrice de la Cx32 comporte aumoins deux promoteurs (figure 1).L’un, localisé juste en amont du pre-mier exon, est fonctionnel dans lesépithéliums glandulaires et le systèmenerveux central ; l’autre, localisé àl’extrémité 3’ du long intron, estfonctionnel dans les systèmes nerveuxcentral et périphérique [8]. L’exis-tence de ces deux promoteurs et d’unsite unique accepteur d’épissage per-met d’envisager la production dedeux transcrits distincts et donc uneexpression différentielle de cetteconnexine dans divers tissus, souscontrôle de divers facteurs de trans-cription, encore non identifiés.La plupart des gènes des connexinessont dispersés sur des chromosomesdifférents [5, 6]. Dans l’espècehumaine, les gènes codant pour lesCx 26, Cx 32 et Cx 43 se trouvent res-pectivement sur les chromosomes 13,X, et 6, et il y a un pseudogène pourla Cx43 sur le chromosome 5. Cepen-dant, les gènes codant pour les Cx37et Cx40 se trouvent tous les deux surle chromosome 1, et ceux codantpour les Cx26 et Cx46 sur le chromo-some 13, suggérant un partage pos-sible de séquences régulatrices com-munes. Il n’y a pas de lien immédiatentre ce regroupement chromoso-mique et l’expression des connexines.Ainsi, alors que la Cx37 et la Cx40sont généralement produites dans lesmêmes tissus, les Cx26 et Cx46 ne lesont pas.
Les connexines
Toutes les connexines ont leurs ex-trémités amino- et carboxyterminalesdans le cytoplasme et comportentquatre portions hydrophobes reliéesles unes aux autres par une bouclecytoplasmique et deux boucles inter-cellulaires (figure 2) [1-3]. Les quatreportions hydrophobes ont une topo-logie d’hélices α arrangées parallèle-ment. La troisième portion intra-membranaire comporte des résiduspolaires, acides et basiques, qui en-tourent le canal hydrophile de cha-que connexon [3]. Les connexinesde différentes espèces montrent unestructure primaire particulièrementbien conservée, surtout au niveau del’extrémité aminoterminale, des seg-
910 m/s n° 8-9, vol. 12, août-septembre 96
RÉFÉRENCES1. Beyer EC, Paul DL, Goodenough DA.Connexin family of gap junction proteins. JMembr Biol 1990 ; 116 : 187-94.
2. Bennett MVL, Barrio LC, Bargiello TA,Spray DC, Hertzberg E, Saez JC. Gap junc-tions : new tools, new answers, new ques-tions. Neuron 1991 ; 6 : 305-20.
3. Kumar NM, Gilula NB. Molecular biologyand genetics of gap junction channels.Semin Cell Biol 1992 ; I3 : 3-16.
4. Willecke K, Hennemann H, Dahl E, Jung-bluth S, Heynkes R. The diversity ofconnexin genes encoding gap junctionalproteins. Eur J Cell Biol 1991 ; 56 : 1-7.
5. Fishman GI, Eddy RL, Shows TB, Rosen-thal L, Leinwand LA. The human connexingene family of gap junction proteins : dis-tinct chromosomal locations but similarstructure. Genomics 1991 ; 10 : 250-6.
6. Hennemann H, Kozjek G, Dahl E,Nicholson B, Willecke K. Molecular cloningof mouse connexins26 and -32 : similargenomic organization but distinct promotersequences of two gap junction genes. Eur JCell Biol 1992 ; 58 : 81-9.
7. Yu W, Dahl G, Werner R. The con-nexin43 gene is responsive to estrogen. ProcR Soc Lond B Biol Sci 1994 ; 255 : 125-32.
8. Neuhaus IM, Dahl G, Werner R. Use ofalternate promoters for tissue-specificexpression of the gene coding forconnexin32. Gene 1995 ; 158 : 257-62.
9. Sáez JC, Spray DC, Nairn AC, HertzbergEL, Greengard P, Bennett MVL. cAMPincreases junctional conductance and sti-mulates phosphorylation of the 27-kDaprincipal gap junction polypeptide. ProcNatl Acad Sci USA 1986 ; 83 : 2473-7.
10. Meda P, Pepper M, Traub O, WilleckeK, Gros D, Beyer E, Nicholson B, Paul D,Orci L. Differential expression of gap junc-tion connexins in endocrine and exocrineglands. Endocrinology 1993 ; 133 : 2371-8.
11. Musil LS, Goodenough DA. Multisubu-nit assembly of an integral plasma membra-ne channel protein, gap junction con-nexin43, occurs after exit from the ER. Cell1993 ; 74 : 1065-77.
911m/s n° 8-9, vol. 12, août-septembre 96
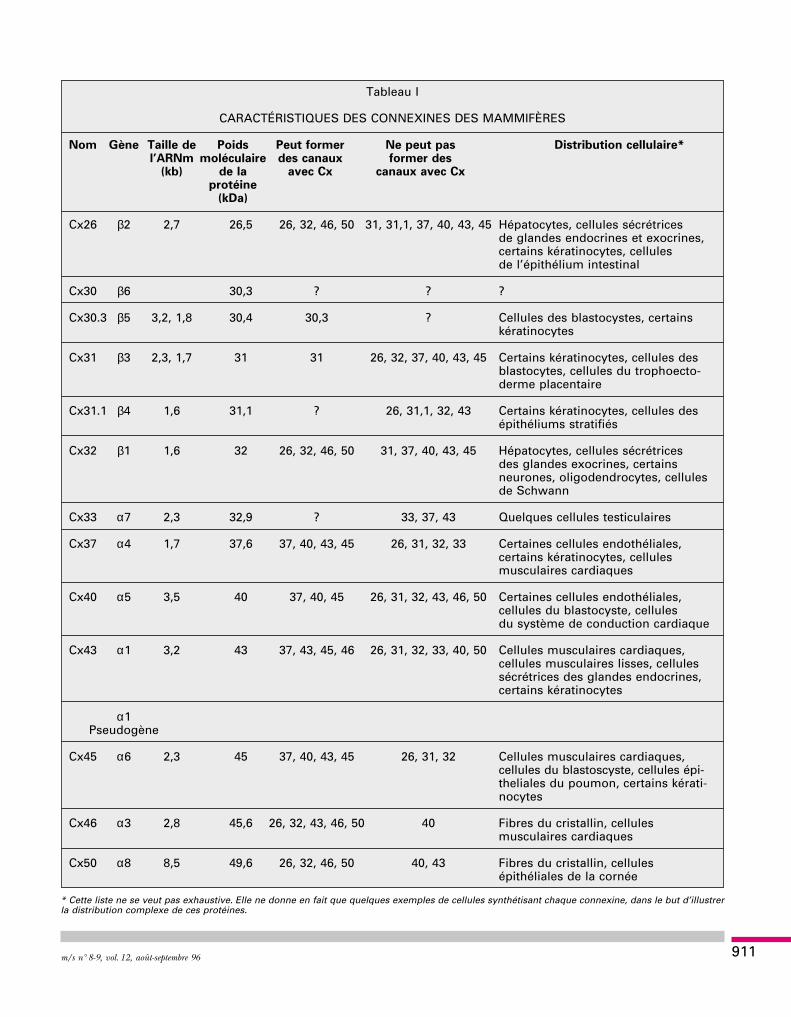
Tableau I
CARACTÉRISTIQUES DES CONNEXINES DES MAMMIFÈRES
Nom Gène Taille de Poids Peut former Ne peut pas Distribution cellulaire*l’ARNm moléculaire des canaux former des
(kb) de la avec Cx canaux avec Cxprotéine
(kDa)
Cx26 β2 2,7 26,5 26, 32, 46, 50 31, 31,1, 37, 40, 43, 45 Hépatocytes, cellules sécrétricesde glandes endocrines et exocrines,certains kératinocytes, cellulesde l’épithélium intestinal
Cx30 β6 30,3 ? ? ?
Cx30.3 β5 3,2, 1,8 30,4 30,3 ? Cellules des blastocystes, certainskératinocytes
Cx31 β3 2,3, 1,7 31 31 26, 32, 37, 40, 43, 45 Certains kératinocytes, cellules desblastocytes, cellules du trophoecto-derme placentaire
Cx31.1 β4 1,6 31,1 ? 26, 31,1, 32, 43 Certains kératinocytes, cellules desépithéliums stratifiés
Cx32 β1 1,6 32 26, 32, 46, 50 31, 37, 40, 43, 45 Hépatocytes, cellules sécrétricesdes glandes exocrines, certainsneurones, oligodendrocytes, cellulesde Schwann
Cx33 α7 2,3 32,9 ? 33, 37, 43 Quelques cellules testiculaires
Cx37 α4 1,7 37,6 37, 40, 43, 45 26, 31, 32, 33 Certaines cellules endothéliales,certains kératinocytes, cellulesmusculaires cardiaques
Cx40 α5 3,5 40 37, 40, 45 26, 31, 32, 43, 46, 50 Certaines cellules endothéliales,cellules du blastocyste, cellulesdu système de conduction cardiaque
Cx43 α1 3,2 43 37, 43, 45, 46 26, 31, 32, 33, 40, 50 Cellules musculaires cardiaques,cellules musculaires lisses, cellulessécrétrices des glandes endocrines,certains kératinocytes
α1Pseudogène
Cx45 α6 2,3 45 37, 40, 43, 45 26, 31, 32 Cellules musculaires cardiaques,cellules du blastoscyste, cellules épi-theliales du poumon, certains kérati-nocytes
Cx46 α3 2,8 45,6 26, 32, 43, 46, 50 40 Fibres du cristallin, cellulesmusculaires cardiaques
Cx50 α8 8,5 49,6 26, 32, 46, 50 40, 43 Fibres du cristallin, cellulesépithéliales de la cornée
* Cette liste ne se veut pas exhaustive. Elle ne donne en fait que quelques exemples de cellules synthétisant chaque connexine, dans le but d’illustrerla distribution complexe de ces protéines.

ments transmembranaires et deuxboucles intercellulaires (30 % à 50 %d’analogie). Chacune de ces bouclescontient, notamment, trois résiduscystéine qui ont conservé la mêmeposition au cours de l’évolution [2,3]. Ces résidus forment des liaisonsdisulfure intramoléculaires qui pour-
raient être nécessaires pour stabiliserla structure tertiaire des portionsintercellulaires, permettant l’aligne-ment et l’interaction de connexons(voir figure 3) se faisant face.Les treize connexines décelées chezles mammifères diffèrent considéra-blement entre elles en ce qui concer-ne la boucle cytoplasmique et l’extré-mité carboxyterminale [1-3]. Cettedernière région comporte 18 acidesaminés dans la Cx26 et 191 acidesaminés dans la Cx46, expliquant queles poids moléculaires des con-nexines varient entre 26 et 50 kDa(Tableau I). La variabilité de l’extré-mité carboxyterminale a permis laproduction d’anticorps spécifiquesqui sont utilisés pour l’immunolocali-sation des différentes isoformes (figu-re 2). Alors que l’on considère que lesrégions conservées des connexinessont importantes pour la formationet le fonctionnement des canauxjonctionnels, les régions variablessont probablement neutres. Cer-taines de ces régions pourraientcependant déterminer des propriétésspécifiques. Ainsi l’extrémité car-boxyterminale de certaines con-nexines comporte des résidus phos-phorylables absents dans d’autresisoformes [1, 3]. La phosphorylation de résidus sérineou tyrosine est la seule modificationpost-traductionnelle qui ait été détec-tée dans les connexines [9]. Contrai-rement à de nombreuses autres pro-téines intégrales de la membranecellulaire, les connexines ont unetrès courte demi-vie (entre 3 et6 heures) [1-3], elles ne sont pas gly-cosylées et n’ont pas d’activité enzy-matique. Plusieurs connexines com-portent cependant des sites pouvantlier la calmoduline, suggérant unepossible régulation par le Ca2+ cyto-plasmique libre [2].Pratiquement chaque type deconnexine est présent dans plusieurstissus, et, à de rares exceptions près,la plupart des cellules synthétisentplusieurs connexines [2, 10]. Cer-taines connexines sont produites parde très nombreux types cellulaires,alors que d’autres ont une distribu-tion beaucoup plus restreinte(Tableau I). Au vu des propriétés spé-cifiques des canaux jonctionnels for-més par différentes connexines, il estvraisemblable que la présence demultiples connexines soit nécessaire
912 m/s n° 8-9, vol. 12, août-septembre 96
RÉFÉRENCES12. Musil LS, Goodenough DA. Biochemicalanalysis of connexin43 intracellular trans-port, phosphorylation, and assembly intogap junctional plaques. J Cell Biol 1991 ; 115 :1357-74.
13. Makowski L, Caspar DL, Phillips WC,Goodenough DA. Gap junction structures.II. Analysis of the X-ray diffraction data. JCell Biol 1977 ; 74 : 629-45.
14. White TW, Paul DL, Goodenough DA,Bruzzone R. Functional analysis of selectiveinteractions among rodent connexins. MolBiol Cell 1995 ; 6 : 459-70.
15. Veenstra RD, Wang HZ, Westphale EM,Beyer EC. Multiple connexins confer dis-tinct regulatory and conductance propertiesof gap junctions in developing heart. CircRes 1992 ; 71 : 1277-83.
16. Hooper ML, Subak-Sharpe JH. Metabo-lic co-operation between cells. Int Rev Cytol1981 ; 69 : 46-104.
17. Kohen E, Kohen C, Thorell B, MintzDH, Rabinovitch A. Intercellular communi-cation in pancreatic islet monolayer culture:a microfluorometric study. Science 1979 ;204 : 862-5.
18. Lawrence TS, Beers WH, Gilula NB.Transmission of hormonal stimulation bycell-to-cell communication. Nature 1978 ;272 : 501-6.
19. Sáez JC, Connor JA, Spray DC, BennettMVL. Hepatocyte gap junctions are per-meable to the second messenger, inositol1.4.5-triphosphate, and to calcium ions. ProcNatl Acad Sci USA 1989 ; 86 : 2708-12.
20. Meda P, Spray DC. Physiological roles ofgap junctions. In : Hertzberg EL, ed. Gapjunctions advances in cellular and molecular bio-logy, vol. 18. Greewich CT : JAI Press 1996(sous presse).
21. Salomon D, Meda P. Heterogeneity andcontact-dependent regulation of hormonesecretion by individual B cells. Exp Cell Res1986 ; 162 : 507-20.
22. Bosco D, Soriano JV, Chanson M, MedaP. Heterogeneity and contact-dependentregulation of amylase release by individualacinar cells. J Cell Physiol 1994 ; 160 : 378-88.
23. Mears D, Sheppard NF Jr, Atwater I,Rojas E. Magnitude and modulation of pan-creatic β-cell gap junction electrical conduc-tance in situ. J Membr Biol 1995 ; 146 : 163-76.
24. Miller SM, Garfield RE, Daniel EE.Improved propagation in myometrium asso-ciated with gap junctions during parturi-tion. Am J Physiol 1989 ; 256 : C130-41.
Exon 1A Exon 2
5' 3'
Régioncodante
P1
5' 3'
Cx 32
BP2
CGroupe 1 (β)
Groupe 2 (α)
Cx 26Cx 30Cx 32Cx 30,3Cx 31Cx 31,1
Cx 37Cx 33Cx 43Cx 40Cx 46Cx 50Cx 45
?
Vertébrés
Figure 1. Structure et phylogénie desgènes des connexines. A. Tous lesgènes des connexines étudiés jus-qu’à présent ont une structure simi-laire. Ils comportent deux exons sé-parés par un intron. La régioncodante est entièrement contenuedans le deuxième exon. B. Le gènecodant pour la Cx32 comporte aumoins deux promoteurs, l’un (P1)fonctionnel au niveau des épithé-liums glandulaires et du systèmenerveux central, l’autre (P2) au ni-veau des systèmes nerveux centralet périphérique. Les lignes obliquesnoires indiquent que deux transcritspeuvent être synthétisés à partir dece gène. C. La reconstitution d’unarbre phylogénétique suggère quel’ensemble des connexines dérived’un ancêtre commun non encoreidentifié. Le groupe des connexines αs’est vraisemblablement distancé dugroupe des connexines β au momentde l’apparition des vertébrés.
pour assurer différentes fonctionscellulaires, une régulation différen-tielle de la communication intercel-lulaire et/ou pour établir des limitesfonctionnelles entre divers territoiresde cellules communicantes. Au seind’un même tissu, l’expression nativedes gènes codant pour les conne-xines peut être modifiée dans desconditions très diverses, notammentau cours du développement, dechangements du fonctionnement
physiologique ou pathologique. Deplus, diverses connexines peuventêtre réglées différemment dans unemême cellule et, à l’inverse, unemême connexine peut être régléedifféremment dans différents typescellulaires [2].
Les connexons
Avant d’atteindre la membrane cellu-laire, les connexines s’assemblent par
groupes de six molécules pour for-mer des structures tubulaires creuses,appelées connexons (figure 3).Contrairement à ce qui se passe pourla plupart des autres protéines inté-grales de la membrane, cette oligo-mérisation survient au niveau d’uncompartiment distal de l’appareil deGolgi [11].Les connexons sont ensuite incorpo-rés, après la fusion de vésicules gol-giennes avec la membrane plasmique,
913m/s n° 8-9, vol. 12, août-septembre 96
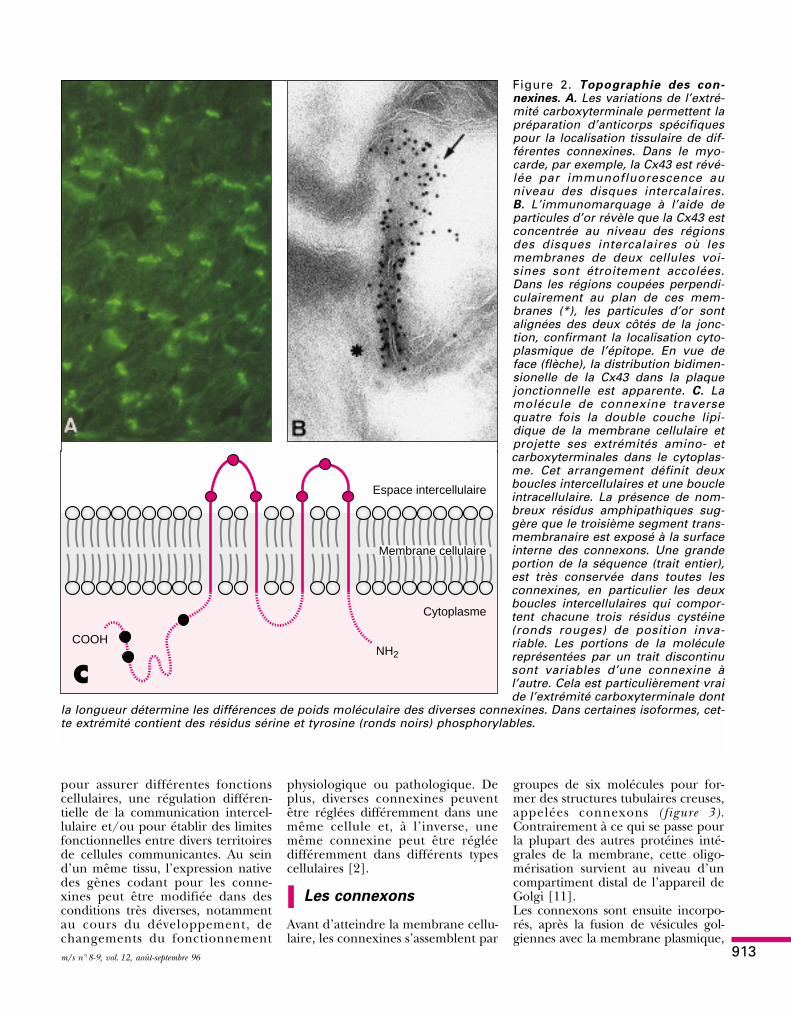
Figure 2. Topographie des con-nexines. A. Les variations de l’extré-mité carboxyterminale permettent lapréparation d’anticorps spécifiquespour la localisation tissulaire de dif-férentes connexines. Dans le myo-carde, par exemple, la Cx43 est révé-lée par immunofluorescence auniveau des disques intercalaires.B. L’immunomarquage à l’aide departicules d’or révèle que la Cx43 estconcentrée au niveau des régionsdes disques intercalaires où lesmembranes de deux cellules voi-sines sont étroitement accolées.Dans les régions coupées perpendi-culairement au plan de ces mem-branes (*), les particules d’or sontalignées des deux côtés de la jonc-tion, confirmant la localisation cyto-plasmique de l’épitope. En vue deface (flèche), la distribution bidimen-sionelle de la Cx43 dans la plaquejonctionnelle est apparente. C. Lamolécule de connexine traversequatre fois la double couche lipi-dique de la membrane cellulaire etprojette ses extrémités amino- etcarboxyterminales dans le cytoplas-me. Cet arrangement définit deuxboucles intercellulaires et une boucleintracellulaire. La présence de nom-breux résidus amphipathiques sug-gère que le troisième segment trans-membranaire est exposé à la surfaceinterne des connexons. Une grandeportion de la séquence (trait entier),est très conservée dans toutes lesconnexines, en particulier les deuxboucles intercellulaires qui compor-tent chacune trois résidus cystéine(ronds rouges) de position inva-riable. Les portions de la moléculereprésentées par un trait discontinusont variables d’une connexine àl’autre. Cela est particulièrement vraide l’extrémité carboxyterminale dont
la longueur détermine les différences de poids moléculaire des diverses connexines. Dans certaines isoformes, cet-te extrémité contient des résidus sérine et tyrosine (ronds noirs) phosphorylables.
Espace intercellulaire
Cytoplasme
COOHNH2
C
Membrane cellulaire

dans des régions membranaires quine participent pas à des contacts inter-cellulaires [12]. Chaque connexontraverse alors complètement la doublecouche lipidique de la membrane,exposant une de ses extrémités dansle cytoplasme et l’autre dans l’espaceintercellulaire. La portion centrale dechaque connexon comporte un espa-ce hydrophile de 2 à 4 nm de dia-mètre (figure 3) qui constitue la moitiéd’un canal jonctionnel [13]. Vraisem-blablement, ce demi-canal est fonc-tionnellement fermé jusqu’à ce quel’extrémité intercellulaire des con-nexons soit isolée de l’espace environ-nant. Dans le cas contraire, une rapi-de dialyse cytoplasmique s’établiraitinévitablement.
Les canaux jonctionnels
Lorsqu’un contact intercellulaire estétabli, les connexons d’une cellules’alignent bout à bout avec ceux de lacellule voisine, établissant un canalhydrophile continu long d’environ10 nm [13]. Ce canal jonctionnelmet en communication directe lecytoplasme des deux cellules encontact, par delà l’espace intercellu-laire (figure 3). La reconnaissance entre connexonsopposés est déterminée par la struc-ture de leurs domaines intercellu-laires et par le type de connexine.Toutes les connexines natives sontcapables de former des canaux jonc-tionnels en interagissant avec desmolécules du même type, mais ellesne le font pas systématiquement [14]lorsqu’elles interagissent avec desmolécules d’un type différent(Tableau I). La compatibilité de diffé-rentes connexines semble spécifiéepar la structure du deuxième domai-ne intercellulaire [14] et par leurenvironnement. En effet, les molé-cules d’adhérence cellulaire, l’état deglycosylation d’autres protéines desurface, la composition en lipides dela double couche membranaire et laconcentration cytoplasmique desions Ca2+ et H+ influencent l’assem-blage des canaux intercellulaires [2].La liaison entre connexons est main-tenue par des interactions hydro-phobes [12] qui doivent être suffi-samment fortes pour isoler le canaljonctionnel de l’espace intercellulai-re. Lorsque cet isolement est établi,les canaux jonctionnels s’ouvrent
progressivement avant d’atteindreleur conductance et perméabilitémaximales [2].Les canaux jonctionnels se différen-cient des autres canaux membra-naires par plusieurs caractéristiquesparticulières. Ils sont formés obliga-toirement par deux cellules, chacunecontribuant pour une moitié ducanal. Malgré une conductanceionique d’amplitude comparable àcelle d’autres canaux membranaireset variant de 35 à 200 pS selon le typede connexine, ils sont perméables àpresque tous les ions, particulière-ment aux cations [2]. Ils sont égale-ment perméables à de très nom-breuses molécules d’un poidsmoléculaire inférieur à 900 Da etd’un diamètre maximal d’environ1,5 nm [2]. Ces propriétés varientsensiblement selon le type deconnexine et son éventuelle modifi-cation post-traductionnelle [2, 15].Généralement, les canaux formés pardes connexines hétérologues ont uneconductance et une perméabilitémoindres que celles des canaux for-més par des connexines du mêmetype. De plus, dans ce dernier cas, laconductance électrique et sa régula-tion par la différence de potentielpeuvent ne pas être symétriques [14],établissant éventuellement une com-munication jonctionnelle préféren-tielle dans une direction.La régulation des canaux jonctionnelsest multifactorielle et diffère selon letype de connexine, le type de celluleet son état d’activité [2]. Expérimen-talement, elle peut être obtenue pardes anticorps, des alcanols (alcanescomportant un groupe hydroxyle),des anesthésiques et des oligonucléo-tides antisens. Dans des conditionsphysiologiques, elle pourrait être assu-rée par le niveau de Ca2+ intracellulai-re libre et/ou la calmoduline, le pHcytosolique, plusieurs hormones, neu-romédiateurs et facteurs de croissan-ce, des métabolites de l’acide arachi-donique, les protéines kinases A et C,des phosphatases, des oncogènes etcertains morphogènes [2].
Les jonctions de type gap
Le contact entre deux cellules entraî-ne, en quelques minutes, la concen-tration de canaux jonctionnels dansdes régions où les membranes plas-miques sont étroitement juxtaposées.
914 m/s n° 8-9, vol. 12, août-septembre 96
RÉFÉRENCES25. Warner A. Gap junctions in develop-ment – a perspective. Semin Cell Biol 1992 ;3 : 81-91.
26. Pepper MS, Montesano R, El AoumariA, Gros D, Orci L, Meda P. Coupling andconnexin 43 expression in microvascularand large vessel endothelial cells. Am J Phy-siol 1992 ; 262 : C1246-57.
27. Vozzi C, Ullrich S, Charollais A, PhilippeJ, Orci L, Meda P. Adequate connexin-mediated coupling is required for properinsulin production. J Cell Biol 1995 ; 131 :1561-72.
28. Stutenkemper R, Geisse S, Schwarz HJ,Look J, Traub O, Nicholson BJ, Willecke K.The hepatocyte-specific phenotype of muri-ne liver cells correlates with high expressionof connexin32 and connexin26 but very lowexpression of connexin43. Exp Cell Res1992 ; 201 : 43-54.
29. Meda P. Junctional coupling of pancrea-tic β-cells. In : Huizinga JD, ed. Pacemakeractivity and intercellular communication. BocaRaton : CRC Press, 1995 : 275-91.
30. Meda P, Perrelet A, Orci L. Increase ofgap junctions between pancreatic B-cellsduring stimulation of insulin secretion. JCell Biol 1979 ; 82 : 441-8.
31. Meda P, Bosco D, Chanson M, GiordanoE, Vallar L, Wollheim C, Orci L. Rapid andreversible secretion changes during uncou-pling of rat insulin-producing cells. J ClinInvest 1990 ; 86 : 759-68.
32. Chanson M, Orci L, Meda P. Extent andmodulation of junctional communicationbetween pancreatic acinar cells in vivo. Am JPhysiol 1991 ; 261 : G28-36.
33. Reaume AG, de Sousa PA, Kulkarni S,Lowell Langille B, Zhu D, Davies TC, JunejaSC, Kidder GM, Rossant J. Cardiac malforma-tion in neonatal mice lacking connexin43.Science 1995 ; 267 : 1831-4.
34. Berghoffen J, Scherer SS, Wang S,Oronzi-Scott M, Bone LJ, Paul DL, Chen K,Lensch MW, Chance PF, Fischbeck KH.Connexin mutations in X-linked Charcot-Marie-Tooth disease. Science 1993 ; 262 :2039-42.

Dans ces régions, l’espace extracellu-laire est réduit à une fine fente de 1 à2 nm [13], le gap anglo-saxon qui adéterminé la dénomination désor-mais usuelle de ces régions (figure 4).Il est maintenant établi que la synthè-se de connexines est nécessaire etsuffisante pour établir les jonctionsde type gap et qu’il existe une rela-tion obligatoire entre la présence deces structures membranaires et celle
d’échanges intercellulaires directsd’ions et de molécules cytoplas-miques [1-3]. Ce type de communica-tion cellulaire est aussi appelé cou-plage cellulaire ou jonctionnel.
Le couplage jonctionnel
Le passage d’ions d’une cellule àl’autre au travers de canaux jonction-nels est suggéré par le couplage élec-
trique de cellules voisines et parl’enregistrement d’un potentiel élec-trotonique, c’est-à-dire induit parune injection de courant dans unecellule, à distance du site d’injection.Le couplage est démontré par unemesure directe de conductance àl’interface de deux cellules dans desconditions de potentiel et de compo-sition ionique intracellulaires impo-sées pour réduire les transitionsd’ouverture-fermeture des autrestypes de canaux membranaires [2].La conductance des canaux jonction-nels peut aussi être différenciée decelle des autres canaux membra-naires par le fait que l’ouverture descanaux jonctionnels se fait simultané-ment dans deux cellules voisines etest bloquée par des alkanols [2]. Pra-tiquement tous les ions passent autravers des canaux jonctionnels [2].La conductance anionique de cescanaux est plus ou moins constante.En revanche, leur conductance catio-nique varie selon le type de connexi-ne [15].L’échange intercellulaire de molé-cules cytoplasmiques au travers descanaux jonctionnels est le plus aisé-ment démontré après injection dansune cellule de traceurs exogènes [2]ne pouvant franchir la membranecellulaire (figure 5). D’autres appro-ches ont montré que les canaux jonc-tionnnels sont aussi perméables à ungrand nombre de molécules endo-gènes, comprenant des nucléotides(dont l’AMPc), des acides aminés,des cofacteurs vitaminiques, desmétabolites des phospho-inositides(IP3...) et des sucres (glucose 6-phos-phate...) [16-19]. Comme la plupartde ces molécules sont chargées néga-tivement, leur passage au travers descanaux jonctionnels peut varier selonle type de connexine [15].Dans tous les cas, le passage intercel-lulaire d’ions et de molécules au tra-vers des canaux jonctionnels se faitpar diffusion le long d’un gradientélectrochimique [2]. Ce passage peutêtre très étendu. On évalue, parexemple, qu’environ 106 moléculesde la taille de l’AMPc pourraient êtreéchangées en une seconde entre unecentaine de cellules couplées. Deplus, la spécificité d’appariement etde perméabilité de différentesconnexines permet d’envisager deséchanges jonctionnels sélectifs, soitse faisant dans une direction préfé-
915m/s n° 8-9, vol. 12, août-septembre 96
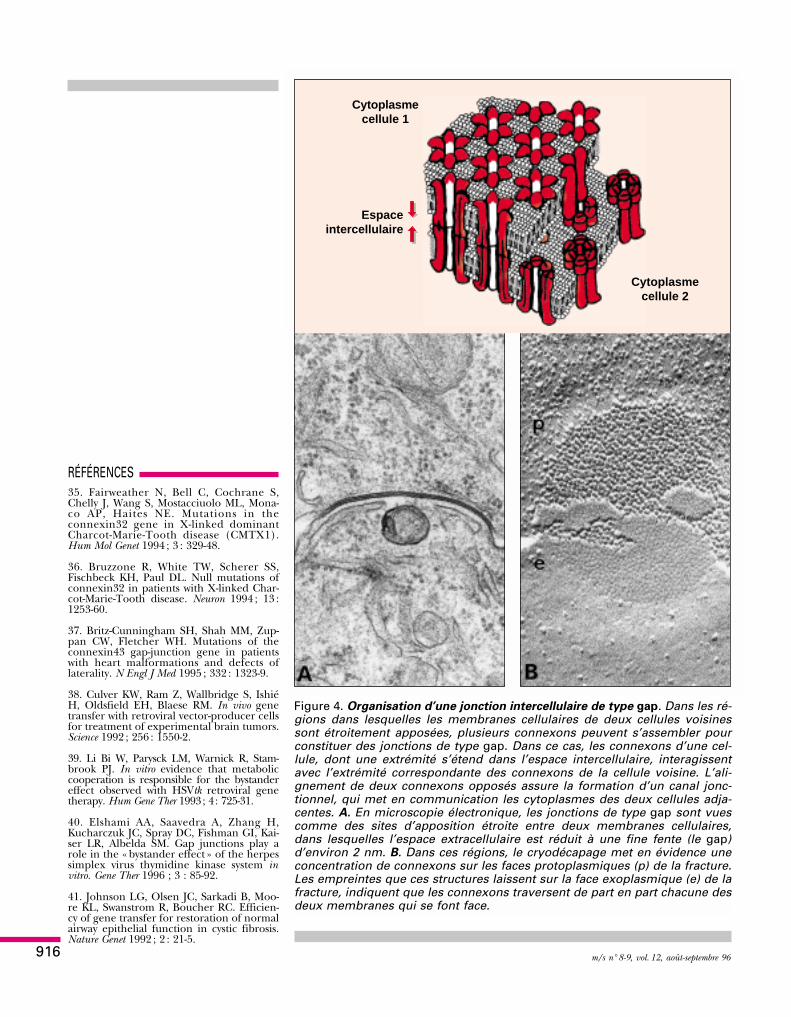
Figure 3. Organisation d’un hémicanal jonctionnel. Haut. Au cours de leurtransit vers la membrane cellulaire, les connexines s’assemblent entre ellespar groupes de six pour former une structure tubulaire creuse, appeléeconnexon. Au centre de chaque connexon, les connexines délimitent un es-pace hydrophile qui constitue la moitié d’un canal jonctionnel. Bas. A fortgrossissement, les sous-unités formant la paroi des connexons ainsi quel’orifice du canal hydrophile central sont visibles dans des répliques de cryo-décapage.
Domaine intercellulaire
Domaine cytoplasmique
Canal hydrophile
NH2COOH
Domaine membranaire
916 m/s n° 8-9, vol. 12, août-septembre 96
RÉFÉRENCES35. Fairweather N, Bell C, Cochrane S,Chelly J, Wang S, Mostacciuolo ML, Mona-co AP, Haites NE. Mutations in theconnexin32 gene in X-linked dominantCharcot-Marie-Tooth disease (CMTX1).Hum Mol Genet 1994 ; 3 : 329-48.
36. Bruzzone R, White TW, Scherer SS,Fischbeck KH, Paul DL. Null mutations ofconnexin32 in patients with X-linked Char-cot-Marie-Tooth disease. Neuron 1994 ; 13 :1253-60.
37. Britz-Cunningham SH, Shah MM, Zup-pan CW, Fletcher WH. Mutations of theconnexin43 gap-junction gene in patientswith heart malformations and defects oflaterality. N Engl J Med 1995 ; 332 : 1323-9.
38. Culver KW, Ram Z, Wallbridge S, IshiéH, Oldsfield EH, Blaese RM. In vivo genetransfer with retroviral vector-producer cellsfor treatment of experimental brain tumors.Science 1992 ; 256 : 1550-2.
39. Li Bi W, Parysck LM, Warnick R, Stam-brook PJ. In vitro evidence that metaboliccooperation is responsible for the bystandereffect observed with HSVtk retroviral genetherapy. Hum Gene Ther 1993; 4 : 725-31.
40. Elshami AA, Saavedra A, Zhang H,Kucharczuk JC, Spray DC, Fishman GI, Kai-ser LR, Albelda SM. Gap junctions play arole in the « bystander effect » of the herpessimplex virus thymidine kinase system invitro. Gene Ther 1996 ; 3 : 85-92.
41. Johnson LG, Olsen JC, Sarkadi B, Moo-re KL, Swanstrom R, Boucher RC. Efficien-cy of gene transfer for restoration of normalairway epithelial function in cystic fibrosis.Nature Genet 1992 ; 2 : 21-5.
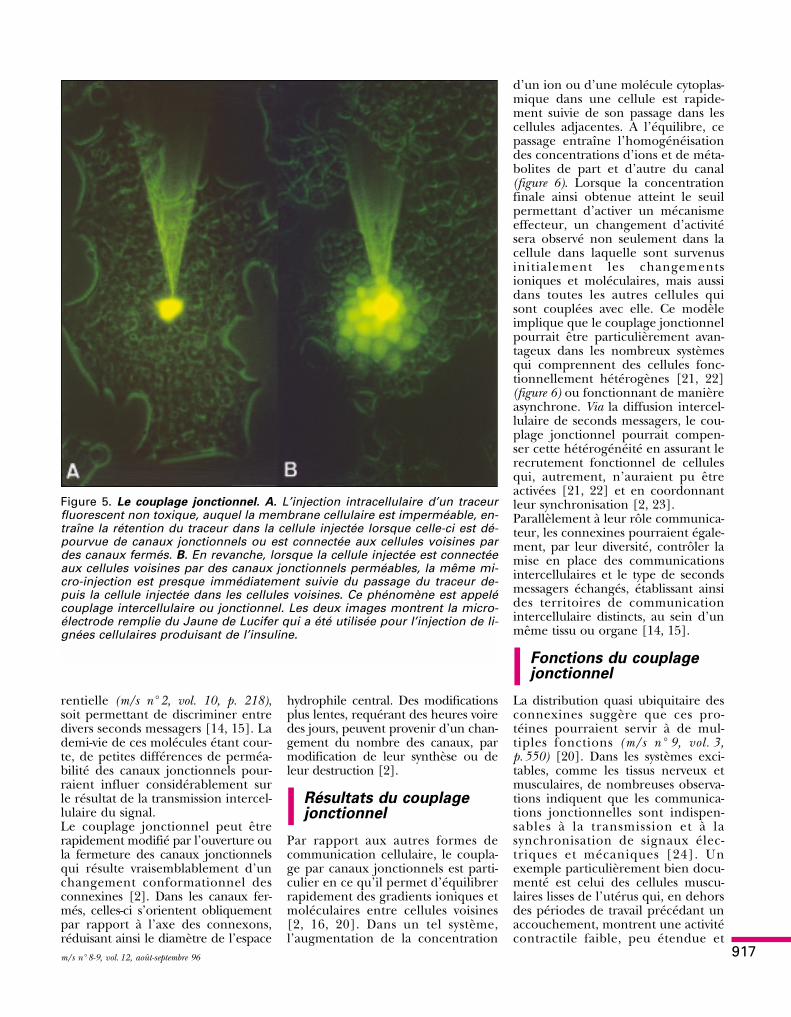
Figure 4. Organisation d’une jonction intercellulaire de type gap. Dans les ré-gions dans lesquelles les membranes cellulaires de deux cellules voisinessont étroitement apposées, plusieurs connexons peuvent s’assembler pourconstituer des jonctions de type gap. Dans ce cas, les connexons d’une cel-lule, dont une extrémité s’étend dans l’espace intercellulaire, interagissentavec l’extrémité correspondante des connexons de la cellule voisine. L’ali-gnement de deux connexons opposés assure la formation d’un canal jonc-tionnel, qui met en communication les cytoplasmes des deux cellules adja-centes. A. En microscopie électronique, les jonctions de type gap sont vuescomme des sites d’apposition étroite entre deux membranes cellulaires,dans lesquelles l’espace extracellulaire est réduit à une fine fente (le gap)d’environ 2 nm. B. Dans ces régions, le cryodécapage met en évidence uneconcentration de connexons sur les faces protoplasmiques (p) de la fracture.Les empreintes que ces structures laissent sur la face exoplasmique (e) de lafracture, indiquent que les connexons traversent de part en part chacune desdeux membranes qui se font face.
Cytoplasmecellule 2
Cytoplasmecellule 1
Espaceintercellulaire
rentielle (m/s n° 2, vol. 10, p. 218),soit permettant de discriminer entredivers seconds messagers [14, 15]. Lademi-vie de ces molécules étant cour-te, de petites différences de perméa-bilité des canaux jonctionnels pour-raient influer considérablement surle résultat de la transmission intercel-lulaire du signal.Le couplage jonctionnel peut êtrerapidement modifié par l’ouverture oula fermeture des canaux jonctionnelsqui résulte vraisemblablement d’unchangement conformationnel desconnexines [2]. Dans les canaux fer-més, celles-ci s’orientent obliquementpar rapport à l’axe des connexons,réduisant ainsi le diamètre de l’espace
hydrophile central. Des modificationsplus lentes, requérant des heures voiredes jours, peuvent provenir d’un chan-gement du nombre des canaux, parmodification de leur synthèse ou deleur destruction [2].
Résultats du couplagejonctionnel
Par rapport aux autres formes decommunication cellulaire, le coupla-ge par canaux jonctionnels est parti-culier en ce qu’il permet d’équilibrerrapidement des gradients ioniques etmoléculaires entre cellules voisines[2, 16, 20]. Dans un tel système,l’augmentation de la concentration
d’un ion ou d’une molécule cytoplas-mique dans une cellule est rapide-ment suivie de son passage dans lescellules adjacentes. A l’équilibre, cepassage entraîne l’homogénéisationdes concentrations d’ions et de méta-bolites de part et d’autre du canal(figure 6). Lorsque la concentrationfinale ainsi obtenue atteint le seuilpermettant d’activer un mécanismeeffecteur, un changement d’activitésera observé non seulement dans lacellule dans laquelle sont survenusinitialement les changementsioniques et moléculaires, mais aussidans toutes les autres cellules quisont couplées avec elle. Ce modèleimplique que le couplage jonctionnelpourrait être particulièrement avan-tageux dans les nombreux systèmesqui comprennent des cellules fonc-tionnellement hétérogènes [21, 22](figure 6) ou fonctionnant de manièreasynchrone. Via la diffusion intercel-lulaire de seconds messagers, le cou-plage jonctionnel pourrait compen-ser cette hétérogénéité en assurant lerecrutement fonctionnel de cellulesqui, autrement, n’auraient pu êtreactivées [21, 22] et en coordonnantleur synchronisation [2, 23].Parallèlement à leur rôle communica-teur, les connexines pourraient égale-ment, par leur diversité, contrôler lamise en place des communicationsintercellulaires et le type de secondsmessagers échangés, établissant ainsides territoires de communicationintercellulaire distincts, au sein d’unmême tissu ou organe [14, 15].
Fonctions du couplagejonctionnel
La distribution quasi ubiquitaire desconnexines suggère que ces pro-téines pourraient servir à de mul-tiples fonctions (m/s n° 9, vol. 3,p. 550) [20]. Dans les systèmes exci-tables, comme les tissus nerveux etmusculaires, de nombreuses observa-tions indiquent que les communica-tions jonctionnelles sont indispen-sables à la transmission et à lasynchronisation de signaux élec-triques et mécaniques [24]. Unexemple particulièrement bien docu-menté est celui des cellules muscu-laires lisses de l’utérus qui, en dehorsdes périodes de travail précédant unaccouchement, montrent une activitécontractile faible, peu étendue et
917m/s n° 8-9, vol. 12, août-septembre 96
Figure 5. Le couplage jonctionnel. A. L’injection intracellulaire d’un traceurfluorescent non toxique, auquel la membrane cellulaire est imperméable, en-traîne la rétention du traceur dans la cellule injectée lorsque celle-ci est dé-pourvue de canaux jonctionnels ou est connectée aux cellules voisines pardes canaux fermés. B. En revanche, lorsque la cellule injectée est connectéeaux cellules voisines par des canaux jonctionnels perméables, la même mi-cro-injection est presque immédiatement suivie du passage du traceur de-puis la cellule injectée dans les cellules voisines. Ce phénomène est appelécouplage intercellulaire ou jonctionnel. Les deux images montrent la micro-électrode remplie du Jaune de Lucifer qui a été utilisée pour l’injection de li-gnées cellulaires produisant de l’insuline.

asynchrone dans diverses régions dumyomètre. Dans cette situation, lesconnexines et le couplage jonction-nel ne sont pratiquement pas détec-tables. En revanche, juste avantl’accouchement, les contractions etles potentiels d’action s’intensifient,se transmettent sur de grandes dis-tances et deviennent synchrones surune vaste étendue du muscle utérin,en même temps qu’augmentent lasynthèse des connexines et le coupla-
ge jonctionnel. L’association obliga-toire de ces changements et leurcontrôle par des taux d’œstrogènes,de progestérone et de prostaglan-dines qui sont observés au cours desphases terminales de la grossesseindique que les événements méca-niques et jonctionnels du muscle uté-rin sont vraisemblablement liés demanière causale et ont une impor-tance physiologique [24].Dans les systèmes en continuel remo-delage, comme la plupart des tissusembryonnaires, mais aussi l’épider-me ou l’endothélium vasculaireadultes, les changements de synthèsedes connexines lors d’étapes détermi-nantes de la différenciation cellulai-re, ainsi que dans des domaines fonc-tionnellement distincts d’un mêmetissu, suggèrent que les communica-tions jonctionnelles participent auxmécanismes de différenciation, demigration et de croissance cellulaires,dont dépend la morphogenèse destissus et des organes [25, 26]. Ce rôlea pu être confirmé par l’observationqu’une synthèse normale de conne-xines est nécessaire au maintien decaractères spécifiques de plusieurstypes cellulaires [27, 28]. Il pourraitégalement expliquer les malforma-tions observées in vivo après bloqua-ge expérimental des canaux jonction-nels. Des molécules morphogéné-tiques peuvent, en effet, affecter lecouplage jonctionnel [25].Dans les systèmes qui ne sont pas spé-cialisés dans la conduction électriqueou mécanique et qui fonctionnent,dans la plupart des situations physio-logiques, sans changement de la pro-lifération et de la différenciation cel-lulaire, les fonctions du couplagejonctionnel sont moins claires. Dansplusieurs épithéliums sécréteurs, lecouplage semble important pour lasynthèse, le stockage et la libérationde produits de sécrétion spécifiques(m/s n° 9, vol. 3, p. 550 et n° 9, vol. 11,p. 1358) [29, 30]. Par exemple,l’expression du gène de l’insuline, labiosynthèse de cette hormone et sasécrétion par les cellules β du pan-créas endocrine augmentent lorsqueces cellules forment des canaux jonc-tionnels [29, 31]. A l’inverse, denombreuses lignées de cellules tumo-rales qui synthétisent et sécrètentl’insuline de façon anormale ne syn-thétisent pas de connexine permet-tant leur couplage jonctionnel [27].
La correction de ces défauts partransfection stable du gène codantpour la connexine native du pan-créas endocrine améliore leurs carac-téristiques sécrétrices, pour autantque le couplage intercellulaire soitrestauré aux niveaux observés in vivo[27]. La stimulation de la sécrétiond’insuline in vivo est également asso-ciée à une augmentation du coupla-ge jonctionnel [30] alors que deschangements inverses sont observésdans des conditions inhibant la sécré-tion d’insuline [31]. Le rôle favo-rable du couplage jonctionnel estvraisemblablement lié à l’hétérogé-néité fonctionnelle des cellules β qui,individuellement, montrent une trèsgrande variabilité sécrétoire. Desobservations analogues ont été faitesà propos de la sécrétion d’amylasepar les cellules acineuses du pancréasexocrine [32]. Dans ce système,cependant, les sécrétagogues natu-rels induisent une diminution ducouplage et le blocage pharmacolo-gique des canaux jonctionnelsentraîne rapidement une augmenta-tion de la sécrétion d’amylase. Cecomportement, opposé à celui descellules β, pourrait être lié à la syn-thèse d’une connexine différente.Les cellules insulaires et acineuses dupancréas, comme d’ailleurs celles denombreuses autres glandes endo-crines et exocrines, synthétisent dif-férentiellement deux isoformes dis-tinctes de connexines [10], quiparaissent fonctionnellement incom-patibles [14].A ce jour, la démonstration rigoureu-se qu’une fonction physiologique estspécifiquement assurée par le coupla-ge jonctionnel n’a pu encore êtrefournie, entre autres parce que lesobservations à ce propos n’ont étéfaites qu’exceptionellement in vivo[32]. A première vue, la grande varié-té des fonctions proposées pour lesconnexines est aussi surprenante.Cependant, l’échange intercellulairede la plupart des seconds messagersau travers des canaux jonctionnelspermet de concevoir que le couplagepourrait compenser aussi bien desdéficits ioniques, métaboliques oudes systèmes effecteurs de certainescellules que la distribution hétérogè-ne de récepteurs ou d’innervation ausein d’un tissu. Le récent développe-ment de souris transgéniques chezlesquelles l’expression génique d’une
918 m/s n° 8-9, vol. 12, août-septembre 96
Cellules découplées
Stimulus
Effet
Cellules couplées
Stimulus
Effet
Cellules isolées
Stimulus
Effet
Figure 6. Résultat du couplage jonc-tionnel. Dans les systèmes compor-tant, soit plusieurs types cellulairesdifférents, soit des cellules fonction-nellement hétérogènes d’un seultype, seules certaines cellules répon-dent à un stimulus, parce qu’ellescontiennent une quantité suffisantede seconds messagers (points noirs).L’établissement de canaux jonction-nels permet à ces cellules de transfé-rer ces seconds messagers aux cel-lules voisines qui en sont déficientes.Ce transfert entraîne le recrutementfonctionnel des cellules receveuseset synchronise la réponse de l’en-semble des cellules couplées. La fer-meture des canaux jonctionnels pré-vient ces effets, en empêchant lescommunications intercellulaires di-rectes.

connexine est altérée suggère queces fonctions sont fondamentales,voire vitales pour l’organisme. Ainsi,l’inactivation biallélique d’un de cesgènes entraîne la mort des animauxau début de l’embryogenèse (Cx26)ou immédiatement après la naissance(Cx43) et, dans ce dernier cas, lesanimaux naissent avec d’importantesmalformations [33].
Couplage jonctionnelet clinique
Au vu des importantes fonctions cel-lulaires attribuées aux connexines, ilest concevable que des altérations ducouplage jonctionnel puissent inter-venir dans la pathogénie de diversesmaladies. Des changements desconnexines, des jonctions de type gapou du couplage jonctionnel ont étéobservés dans des situations cliniquestrès diverses [20], bien que le rôlepathogénique de ces changementsn’ait pas été démontré. Des observa-tions récentes indiquent cependantque des mutations des connexinessont liées à au moins deux maladieshumaines.La première est la maladie de Char-cot-Marie et Tooth, la plus communedes neuropathies périphériques héré-ditaires, qui est caractérisée par desatteintes motrices et sensitives dis-tales, débutant le plus souvent dansl’enfance ou l’adolescence, et pro-gressant lentement vers une atrophiemusculaire, la faiblesse des membres,et une perte de sensibilité [34, 35].Sous cette symptomatologie commu-ne, on regroupe un ensemble de syn-dromes hétérogènes tant du point devue physiopathologique que géné-tique. Le type 1 de la maladie est dûà une dégénerescence de la myélinedes nerfs périphériques et dans l’unede ses formes (CMTX1), le locusgénétique a été localisé sur unerégion du chromosome X, où setrouve, entre autres, le gène codantpour la Cx32 (m/s n° 2, vol. 10,p. 222). Les malades atteints par cetteforme de la maladie présentent desmutations ponctuelles diverses decette connexine pouvant survenirtout au long de la molécule [34, 35]et qui, pour le moins dans certainscas, ne permettent pas l’établisse-ment de canaux jonctionnels compé-tents [36]. Ce qui est surprenant,c’est que la Cx32 des nerfs périphé-
riques ne relie pas des cellules adja-centes, mais relie entre elles desrégions membranaires d’une mêmecellule de Schwann. La distributionde la connexine au niveau desrégions où la myéline n’est pas com-pacte et laisse persister un peu decytoplasme (nœuds de Ranvier etincisures de Schmitt-Lantermann),suggère que la Cx32 pourrait faciliterla diffusion de métabolites etseconds messagers qui transitententre les régions périnucléaires etpériaxonales d’une cellule deSchwann. La deuxième série d’observations aété faite sur des enfants atteintsd’hétérotaxie viscéroatriale (syndro-me d’Ivemark), un ensemble clini-quement et génétiquement hétérogè-ne associant le plus souvent desévères malformations cardiaques àl’absence de rate et à des formes plusou moins complètes de situs inversusthoracique et abdominal. Ces enfantsprésentent des mutations ponctuellesde la Cx43, entraînant le plus sou-vent la substitution du résidu sérine364 par un résidu proline [37]. Dansplusieurs de ces cas, une deuxièmemutation est également détectable auniveau d’autres résidus phosphory-lables de l’extrémité carboxytermina-le de la connexine. Des expériencesde transfection d’une connexinemutée en position 364 ont montréque cette protéine peut établir unecommunication jonctionnelle qui est,cependant, modulée anormalementen présence de protéines kinasesdépendant de l’AMPc. L’impact invivo de cette dysrégulation et ses rap-ports avec la morphogenèse anorma-le qui caractérise l’hétérotaxie doi-vent encore être définis. Il estintéressant de noter que tous lesenfants hétérotaxiques présententune sténose ou une atrésie de l’artèrepulmonaire, une malformationobservée aussi chez les souris transgé-niques dont l’expression du gènecodant pour la Cx43 est supprimée[37].
Couplage jonctionnelet thérapeutique
Les observations précédentes souli-gnent l’intérêt d’envisager desmoyens thérapeutiques permettant,soit d’agir de façon ciblée sur lesjonctions de type gap, soit de profiter
de leur présence pour améliorerl’efficacité de traitements pharmaco-logiques ou génétiques. La première stratégie vise à modifierle fonctionnement et/ou le nombredes canaux jonctionnels, dans le butd’influencer une fonction cellulairequi en serait dépendante. Un grandnombre de molécules endogènes(hormones, facteurs de croissance,neurotransmetteurs, AMPc...) quipeuvent, soit modifier rapidement laconductance ou la perméabilité descanaux jonctionnels, soit induire deschangements à plus long terme deleur formation ou dégradation, pour-raient être utiles dans ce but [2]. Ace jour, cependant, l’utilisation deces molécules est compliquée parleurs effets pléiotropes qui varientselon le type de connexine, de tissuet de situation physiologique, et parla distribution tissulaire complexedes connexines. Toutefois, l’existen-ce de plusieurs promoteurs et de sitesrégulateurs distincts sur les gènes desconnexines [6-8] devrait permettrel’utilisation de facteurs de transcrip-tion spécifiques capables de modulersélectivement l’expression d’iso-formes au niveau de cellules ciblesparticulières.La deuxième stratégie vise à utiliserles canaux jonctionnels natifs pourfaciliter la diffusion de facteurs(médicaments, par exemple) quiauraient pu n’atteindre que quelquesrégions d’un tissu, ou n’être corrigés(ions endogènes, par exemple) quedans quelques cellules. Ce mécanis-me, dénommé effet bystander, a étémis en évidence au cours de traite-ments antitumoraux combinant duganciclovir, un médicament antiher-pétique, à un transfert génique (m/sn° 6, vol. 11, p. 919) [38]. La stratégiede cette approche est d’infecter descellules tumorales par des vecteursrétroviraux recombinants porteursdu gène de la thymidine kinase duvirus de l’herpès (HSV-tk). Les cel-lules infectées qui synthétisent cetteenzyme sont tuées en présence deganciclovir, un analogue de la gua-nosine qui est métabolisé par la thy-midine kinase virale mais pas par cel-le des mammifères. Le métabolitephosphorylé d’environ 300 Da qui enrésulte est incorporé dans les molé-cules d’ADN en formation, bloquantleur synthèse dans les cellules enmitose. Lors des premiers essais, on
919m/s n° 8-9, vol. 12, août-septembre 96

s’est rendu compte que seulement50 % des cellules tumorales pou-vaient être infectées par les rétrovi-rus, mais que cela suffisait pour obte-nir un effet sur la totalité de latumeur [38]. Plusieurs études indi-quent que ces résultats favorablessont vraisemblablement dus à la per-sistance de canaux jonctionnels grâ-ce auxquels les cellules tumoralesinfectées transfèrent aux cellulesadjacentes le métabolite phosphory-lé du ganciclovir et ainsi, également,la sensibilité à ce médicament [39,40]. Un mécanisme analogue peut êtreenvisagé pour amplifier au sein d’untissu l’effet bénéfique d’une correc-tion par thérapie génique de seule-ment quelques cellules. Ainsi, moinsde 10 % des cellules épithéliales res-piratoires dérivées de patientsatteints de mucoviscidose peuventêtre transfectées en culture avec unADNc codant pour le CFTR normal,la protéine membranaire qui règle letransport épithélial de Cl– [41]. Bienqu’affectant directement peu de cel-lules, cette correction suffit à norma-liser le transport ionique au niveaud’un feuillet épithélial. L’interpréta-tion de ce phénomène, considérée laplus vraisemblable par les auteurs dece travail, est que les cellules dont letransport de Cl– a été normalisé sontcapables d’échanger cet ion avec lescellules voisines, au travers descanaux jonctionnels [41]. En équili-brant les concentrations de Cl– entreles cellules couplées, cet échangepermet la correction fonctionnelledu transport épithélial au niveau dutissu entier, malgré la persistance dela mutation du CFTR dans la majori-té des cellules des malades.
Conclusionset perspectives
Les trois decénnies qui ont suivil’identification des jonctions de typegap ont permis d’établir que ces struc-tures représentent des concentrationsde canaux membranaires pour lacommunication cellulaire, d’identifierles connexines constituant ces canauxet d’établir que la séquence et la topo-graphie de ces protéines ont été pré-servées par la majorité des types cellu-laires, tout au long de l’évolution desorganismes pluricellulaires. Noussavons aujourd’hui également que les
canaux jonctionnels permettent deséchanges sélectifs d’ions et de molé-cules cytoplasmiques qui sont contrô-lés de façons diverses dans différentstissus. Parallèlement, nous commen-çons à apprécier les variations de ceschéma général, qui ont été acquisesau cours de l’évolution, de la morpho-genèse des tissus et de la différencia-tion de cellules spécifiques. L’analysedes séquences génomiques et des fac-teurs de transcription qui contrôlentl’expression de connexines spéci-fiques devrait permettre de com-prendre les raisons de cette diversité.A ce jour, les fonctions physiologiquesdes communications jonctionnelles etles mécanismes moléculaires qui lientles changements de ces communica-tions à ceux de diverses fonctions cel-lulaires sont mal compris. De même,nous savons encore peu de choses surles rapports hiérarchiques qui existententre les communications jonction-nelles et les nombreux autres méca-nismes qui coordonnent le fonctionne-ment des systèmes pluricellulaires. Lesobservations récentes réalisées sur desanimaux transgéniques et dans cer-taines maladies humaines suggèrentcependant déjà qu’une synthèse adé-quate de connexines et du couplagejonctionnel est fondamentale, sinonvitale, et pourrait, lorsqu’elle est per-turbée, contribuer à la pathogénie dedysfonctionnements importants. Cesconsidérations soulignent l’intérêt dereconsidérer un certain nombre desituations cliniques, dont la pathogénieest encore peu comprise, sous l’angled’une désorganisation du réseau decommunications cellulaires normales.Elles lancent également le défi d’élabo-rer des stratégies thérapeutiques utili-sant la synthèse des connexines et lescommunications jonctionnellesqu’elles permettent, pour agir sélecti-vement sur certaines populations cellu-laires. Des essais encourageants enoncologie et en thérapie génique per-mettent d’envisager qu’il s’agit làd’une possibilité réalisable dans unavenir pas trop éloigné ■
TIRÉS À PARTP. Meda.
920 m/s n° 8-9, vol. 12, août-septembre 96
SummaryConnexins, junctional channelsand cell-to-cell communication
The majority of cells of multicellularanimals, from mesozoa to man,form gap junctions. These speciali-sed domains of the cell membraneare assemblies of channels for directcell-to-cell exchanges of cytoplasmicions and molecules. Recent studieshave shown that these channels aremade of a multigene family of inte-gral membrane proteins, whichhave been highly conserved throu-ghout evolution and which arereferred to as connexins. We havenow a fair appreciation of the struc-ture, topographic arrangement andtissue distribution of these proteins,as well as of the biophysical charac-teristics of the junctional channelsthey form. Also, the mechanismswhereby the intercellular communi-cations (junctional coupling) whichthese channels mediate are beingunravelled. These recent develop-ments have not completely elucida-ted the physiological function(s) ofjunctional coupling in most cell sys-tems. Interest in this question hasbeen fostered by the recent observa-tions that knockout of two distinctconnexin genes leads to lethalabnormalities in transgenic mice,and that at least two human diseasesare linked to connexin mutations.
Remerciements
Les recherches de mon groupe sont financéespar le Fonds National Suisse de la RechercheScientifique (subside 32-043086.95), l’UnionEuropéenne (subside BMH4-CT96-1427) et laJuvenile Diabetes Foundation International(subside 195077).
ANNIVERSAIRE ENVIDÉO-CASSETTES
.





























