Embed Size (px)
Citation preview

373
Biochimica et Biophysica Acta, 425 (1976) 373--383 © Elsevier Scientific Publishing Company, Amsterdam -- Printed in The Netherlands
BBA 98550
COMPARISON OF PROTEINS BOUND TO THE DIFFERENT FUNCTIONAL CLASSES OF MESSENGER RNA
J.P. LIAUTARD, B. SETYONO, E. SPINDLER and K. KOHLER
Biologisches lnstitut der Universitiit Stuttgart, D-7000 Stuttgart 60, Ulmer Strasse 227 (G.F.R.)
(Received August 19th, 1975)
Summary
Proteins from nuclear ribonucleoproteins, informosomes, polysomal messen- ger ribonucleoproteins and cytoplasmic "binding factor" are characterized.
1. Nuclear ribonucleoproteins are purified from nuclei disrupted by ultra- sonication. Possible contamination by nucleoplasm, histones or remaining cyto- plasmic structures is controlled.
2. Informosomal proteins are obtained by mild RNAase degradation. This method gives informosomal proteins without appreciable contamination.
3. Polysomal messenger ribonucleoproteins are obtained from cells where the initiation of protein synthesis is arrested in order to release the messenger ribo- nucleoproteins from the polysomes. Their proteins are obtained like the infor- mosomal proteins by mild RNAase digestion. No contamination by informo- somes could be detected by sodium dodecyl sulfate gel electrophoresis.
4. Cytoplasmic "binding factor" proteins are purified by affinity~chromatog- raphy.
5. The four sets of proteins are analysed by sodium dodecylsulfate acrylamide gel electrophoresis.
In spite of the fact that some proteins from one or another kind of messen- ger ribonucleoprotein, have apparently the same molecular weight, the majority of proteins differ.
Introduction
mRNA has been found associated with proteins: in the nucleus (nuclear ribonucleoprotein) * [1,2], in free cytoplasmic particles (informosomes) [3,4],
* Ribonucleoprote in , c o m p l e x be tween RNA and protein; Nuclear r ibonucleoprotein, c o m p l e x f o u n d in the nucleus a n d c o n t a i n i n g h e t e r o g e n e o u s R N A ; P o l y s o m a l m e s s e n g e r r i b o n u c l e o p r o t e i n , n o n - r i b o s o m a l r i b o n u c l e o p r o t e i n r e l eased f r o m t h e p o l y s o m e s ; I n f o r m o s o m e , n o n - r i b o s o m a l r i b o - n u c l e o p r o t e i n f o u n d f ree in t h e c y t o p l a s m ; Binding factor, protein from the c y t o s o l w i t h a f f i n i t y f o r R N A .

374
and in polysomes (polysomal messenger ribonucleoprotein) [ 5,6]. The question has yet to be answered whether the proteins of these different
messenger ribonucleoproteins are totally exchanged or partially substituted either during the transport from the nucleus to the cytoplasm and/or during the activation of the inactive messenger ribonucleoproteins (informosomes)to the translatable polysomal messenger ribonucleoprotein according to the hy- pothesis of Spirin [7]. If the latter is true, one can find the same proteins in the cytoplasm. They should have a great affinity to added RNA and form "in- formosome-like particles" [7,8].
To answer these questions one has to isolate first the protein components of the nuclear ribonucleoproteins, informosomes, polysomal messenger ribonu- cleoproteins and cytoplasmic "binding factor", and then to compare all these proteins from the different sources.
In spite of the large amount of research published about isolation and char- acterization of these messenger ribonucleoproteins it is thus far difficult to make a reasonable comparison of their protein composition, for these particles were isolated from different eukaryotic cells by different methods [9]. We re- port here the characterization of the protein moiety from nuclear ribonucleo- protein, informosome, polysomal messenger ribonucleoprotein and cytoplasmic "binding factor" in HeLa cells which were isolated with minimal cross-con- tamination. We show that they share few, if any, common protein constituents.
Materials and Methods
I. Cell culture and homogenization procedure HeLa cells were grown as suspension culture to a concentration of 5 • l0 s
cells/ml in Eagle's medium supplemented with 5% calf serum. Harvested cells were poured on crushed frozen saline (--80°C), washed once and resuspended in cold buffer A (10 mM Tris • HC1 pH 7.4; 10 mM KC1; 3 mM MgC12; 7 mM mercaptoethanol). After 10 min swelling they were homogenized by 10 strokes in a Dounce homogenizer. Nuclei were sedimented by centrifugation (700 X g, 4 min 4°C), and used to prepare nuclear ribonucleoprotein particles. The super- natant was recentrifuged to remove membranes and mitochondria (Sorvall 30 000 X g, 20 min, 4°C). The postmitochondrial supernatant contained ribo- somal particles, informosomes, and polysomal messenger ribonucleoproteins. The cytosol was prepared independently from postmitochonck, Sal supernatants by centrifuging out all particles above 15S (rotor SW 50, 45 000 rev./min, 2 h).
II. Preparation of nucleoplasm Nuclei obtained as described above were washed once with buffer A contain-
ing 1% Triton X-100 and twice with buffer A in order to remove the detergent. Phase contrast microscopy of the nuclei showed no cytoplasmic contamination. The purified nuclei were disrupted by sonification (4 × 15 s, 50W), the homoge- nate was layered on a 30% sucrose cushion in buffer A and centrifuged (2200 X g, 30 min). The upper opalescent phase, the nucleoplasm, was pipetted off.
IlI. Preparation of informosomal proteins Postmitochondrial supernatant was centrifuged (rotor SW 50, 23 000 rev./

375
min, 16 h) through a cushion of 30% sucrose in buffer B (10 mM Tris • HCI (pH, 7.4) 500 mM KCI; 3 mM MgC12; 7 mM mercaptoethanol), or in buffer A. The pellet was resuspended in buffer B (or in buffer A), and analysed by zonal centrifugation (rotor SW 40, 22 000 rev./min, 17 h) in a 15--30% sucrose gra- dient in buffer B (or in buffer A). Fractions containing 20 S--80 S particles were pooled, sedimented by centrifugation (rotor Ti 50, 35 000 rev./min, 17 h), resuspended in buffer B containing pancreatic RNAase (2 #g/ml) and incubated for 20 min at 20°C. The reaction mixture was layered on a cushion of 15% sucrose in buffer A and centrifuged in a truncated tube (2.2 cm) (rotor SW 50, 45 000 rev./min, 2.15 h). The upper third (0.6 ml) of the tube content was analysed by sodium dodecyl sulfate gel electrophoresis.
IV. Preparation of polysomal messenger ribonucleoproteins In order to stop the protein synthesis at the initiation step, the cells were
shocked by a temperature shift according to Schochetman and Perry [10]. The tenfold concentrated cells were incubated for 20 min at 43°C in a water bath. The cells were harvested and the post-mitochondiral supernatant was prepared, fractionated and treated with a low dose RNAase as described for the informo- somes.
The problem of cross-contamination of polysomal messenger ribonucleopro- teins and informosomes will be discussed later in this paper.
V. Affinity chromatography 4 mg of RNA (prepared from polysome pellet according to Perry et al. [11] )
were allowed to react with 1 g of CNBr-sepharose (Pharmacia) for 24 h at 4°C under constant stirring. The mixture was packed in a column (1 × 3 cm), and non-bound RNA was washed off with buffer B. After blocking unreacted CNBr with ethanolamine, an excess of cytosol was passed through the column. Following extensive rinsing with buffer A, the bound material was eluted with buffer B, then with buffer A containing 0.5% sodium deoxycholate, at a flow rate of 60 ml/h.
As an alternative mRNA was coupled to sepharose, mRNA wasprepared by affinity chromatography on poly(U) sepharose as published by Lindberg and Person [12].
VI. Gel eldctrophoresis and ultracentrifugation Gel electrophoresis of the proteins was performed in 7.5% or 10% acrylamide
with 0.1% sodium dodecyl sulfate using a Shandon apparatus; the gels were stained with Coomassie brillant blue [13].
All ultracentrifugations were performed at 4°C with Beckman centrifuges.
Results and Discussion
I. Proteins from nuclear ribonucleoprotein To exclude possible contamination by other components various control ex-
periments were conducted. The nucleoplasm was analysed in a 15--30% sucrose gradient. Proteins of the
fractions I and II (Fig. la) were analysed by sodium dodecyl sulfate gel elec-
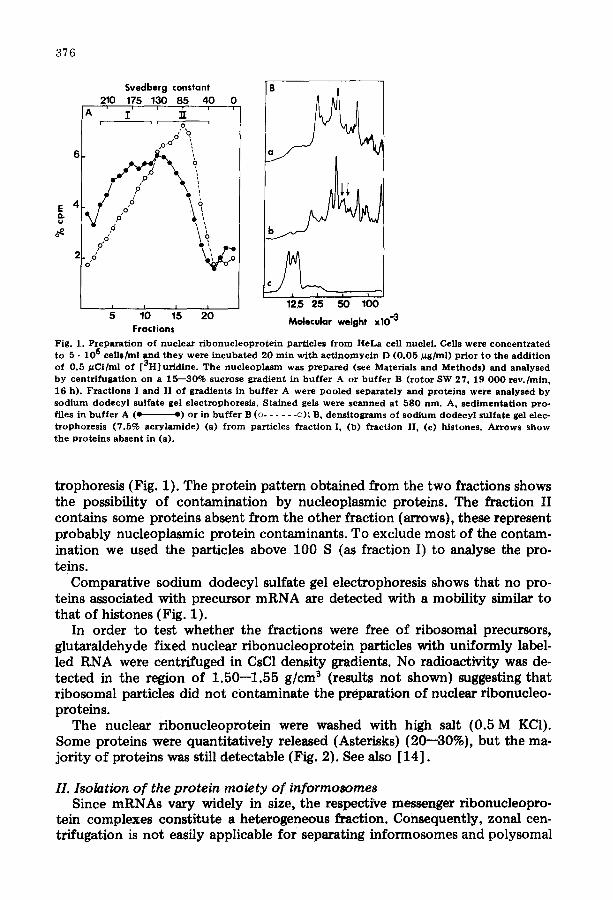
3 7 6
Svedberg constant ~ 0 175 130 55 40 0
IA I' ' ~" ' r - - i , ro "
.o( ~ o
~41 ~oo:"°'O ~el $ Ioo
I I I I I I
5 10 15 20 Fractions
12,5 2 5 5 0 1 0 0
Molecular weight xlO -3
Fig. 1. P z e p a r a t i o n o f n u c l e a r r i b o n u c l e o p r o t e i n pa r t i c l e s f r o m H e L a cell nuc le i . Cells we re c o n c e n t r a t e d to 5 • 106 cellSh~LI a n d t h e y were i n c u b a t e d 2 0 m i n w i t h a c t i n o m y c i n D ( 0 . 0 5 / 1 g / m 1 ) p r i o r t o t h e a d d i t i o n o f 0 . 5 / ~ C i / m l o f [ Z H ] u r i d i n e . T h e n u c l e o p l a s m w a s p r e p a r e d (see Mate r i a l s a n d M e t h o d s ) a n d a n a l y s e d b y c e n t r i f u g a t i o n o n a 1 5 - - 3 0 % s u c r o s e g r a d i e n t in b u f f e r A or b u f f e r B ( r o t o r SW 27 , 19 0 0 0 r e v . / m i n , 16 h) . F r a c t i o n s I a n d II o f g r a d i e n t s in b u f f e r A were p o o l e d s e p a r a t e l y a n d p r o t e i n s were a n a l y s e d b y s o d i u m d o d e c y l su l f a t e gel e l e c t r o p h o r e s i s . S t a i n e d gels we re s c a n n e d a t 5 8 0 rim. A, s e d i m e n t a t i o n p r o - files in b u f f e r A (e ~) o r in b u f f e r B (o . . . . . . o) ; B, d e n s i t o g r a m s of s o d i u m d o d e c y l su l f a t e gel elec- t r o p h o r e s l s ( 7 . 5 % a c r y l a m i d e ) (a) f r o m pa r t i c l e s f r a c t i o n I, (b) f r a c t i o n II, (c) h i s tones . A r r o w s s h o w t h e p r o t e i n s a b s e n t in (a) .
trophoresis (Fig. 1). The protein pattern obtained from the two fractions shows the possibility of contamination by nucleoplasmic proteins. The fraction II contains some proteins absent from the other fraction (arrows), these represent probably nucleoplasmic protein contaminants. To exclude most of the contam- ination we used the particles above 100 S (as fraction I) to analyse the pro- reins.
Comparative sodium dodecyl sulfate gel electrophoresis shows that no pro- teins associated with precursor mRNA are detected with a mobility similar to that of histones (Fig. 1).
In order to test whether the fractions were free of ribosomal precursors, glutaraldehyde fixed nuclear ribonucleoprotein particles with uniformly label- led RNA were centrifuged in CsCl density gradients. No radioactivity was de- tected in the region of 1.50--1.55 g/cm 3 (results not shown) suggesting that ribosomal particles did not contaminate the preparation of nuclear ribonucleo- proteins.
The nuclear ribonucleoprotein were washed with high salt (0.5 M KC1). Some proteins were quantitatively released (Asterisks) (20--30%), but the ma- jority of proteins was still detectable (Fig. 2). See also [14].
II. Isolation of the protein moiety o f informosomes Since mRNAs vary widely in size, the respective messenger ribonucleopro-
tein complexes constitute a heterogeneous fraction. Consequently, zonal cen- trifugation is not easily applicable for separating informosomes and polysomal
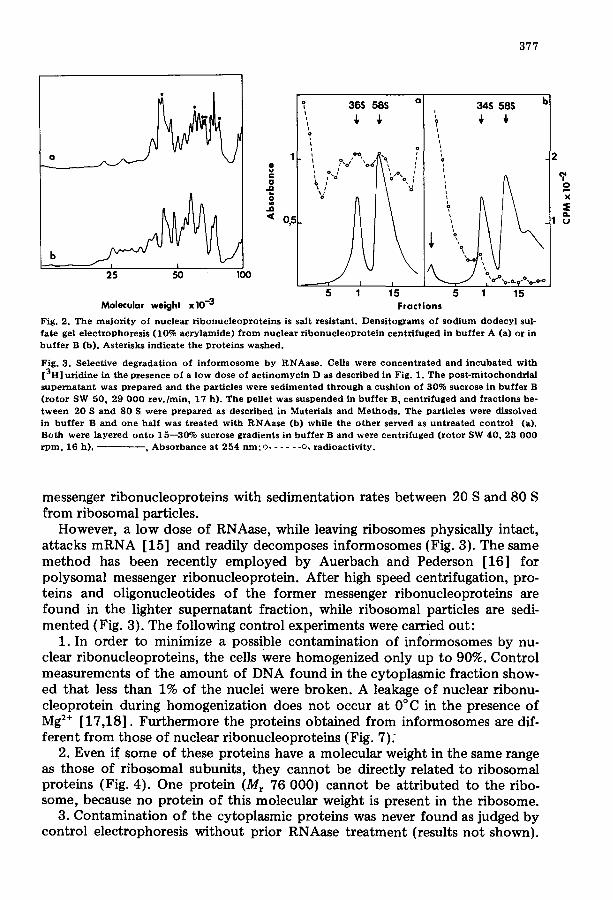
3 7 7
25 50 100
! o
M o l e c u l a r w e i g h t x lO -3
0,~
36S 58S a I , + +
i '~ P'o' o,, 'o~ , ', o / " ' ~, / o o o,,
Z 5 1
34S 58S o ,l, ,i,
I
,,
\ o ,o.O.?-°,o.O-O
115 5 1 15
FracHons
Fig. 2. Th e m a j o r i t y of nuc lea r r i b o n u c l e o p r o t e i n s is salt res is tant . Densi tog~ams of s o d i u m d o d ecy l sul- fa te gel e l ec t rophores i s (10% a c r y l a m i d e ) f r o m nuc lea r r i b o n u c l e o p r o t e i n cen t r i fuged in b u f f e r A (a) or in b u f f e r B (b). Aster isks ind ica te the p ro te ins washed .
Fig. 3. Select ive deg rada t i on of i n f o r m o s o m e by RNAase . Cells were c o n c e n t r a t e d an d i n c u b a t e d wi th [3H] ur id ine in the p resence of a l ow dose o f a c t i n o m y c i n D as descr ibed in Fig. 1. The p o s t - m i t o c h o n d r i a l s u p e r n a t a n t was p r e p a r e d and the par t ic les were s e d i m e n t e d t h r o u g h a cush ion of 30% sucrose in b u f f e r B ( r o t o r SW 50, 29 0 0 0 r e v . / m i n , 17 h) . The pel le t was s u s pe nde d in b u f f e r B, cen t r i fuged and f rac t ions be- t w e e n 20 S and 80 S were p r e p a r e d as descr ibed in Materials and Methods . The par t ic les were dissolved in b u f f e r B and one ha l f was t r e a t e d wi th R N A a s e (b) while the o th e r se rved as u n t r e a t e d co n t ro l (a). Both were l a y e r e d o n t o 15 - -30% sucrose grad ien ts in b u f f e r B an d were cen t r i fuged ( r o t o r SW 40, 23 0 0 0 r p m , 16 h). - - , A b s o r b a n c e a t 254 n m ; o . . . . . . o, r ad ioac t iv i ty .
b 2
? o x
eL 1 u
messenger ribonucleoproteins with sedimentation rates between 20 S and 80 S from ribosomal particles.
However, a low dose of RNAase, while leaving ribosomes physically intact, attacks mRNA [15] and readily decomposes informosomes (Fig. 3). The same method has been recently employed by Auerbach and Pederson [16] for polysomal messenger ribonucleoprotein. After high speed centrifugation, pro- teins and oligonucleotides of the former messenger ribonucleoproteins are found in the lighter supernatant fraction, while ribosomal particles are sedi- mented (Fig. 3). The following control experiments were carried out:
1. In order to minimize a possible contamination of informosomes by nu- clear ribonucleoproteins, the cells were homogenized only up to 90%. Control measurements of the amount of DNA found in the cytoplasmic fraction show- ed that less than 1% of the nuclei were broken. A leakage of nuclear ribonu- cleoprotein during homogenization does not occur at 0°C in the presence of Mg 2÷ [17,18]. Furthermore the proteins obtained from informosomes are dif- ferent from those of nuclear ribonucleoproteins (Fig. 7).
2. Even if some of these proteins have a molecular weight in the same range as those of ribosomal subunits, they cannot be directly related to ribosomal proteins (Fig. 4). One protein (Mr 76 000) cannot b e attributed to the ribo- some, because no protein of this molecular weight is present in the ribosome.
3. Contamination of the cytoplasmic proteins was never found as judged by control electrophoresis without prior RNAase treatment (results not shown).
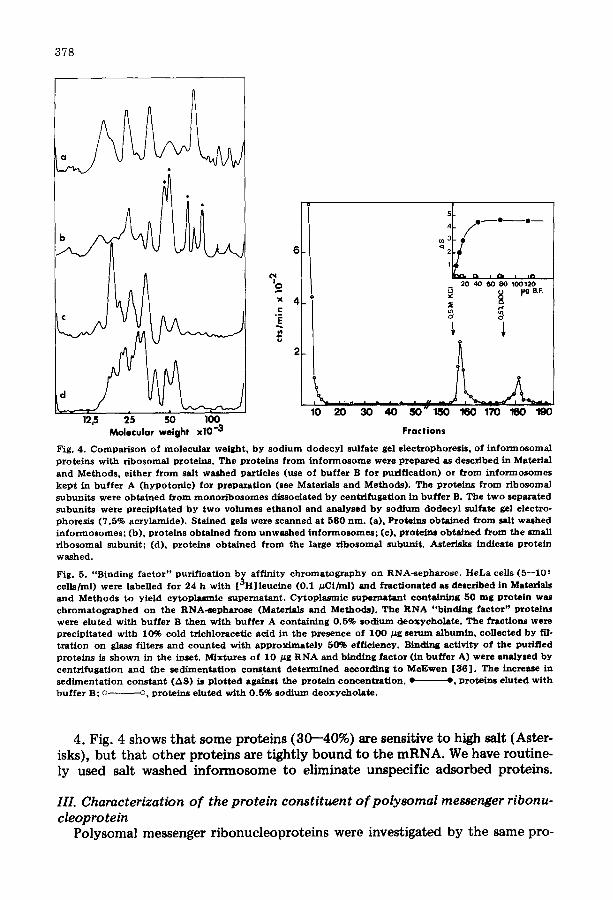
378
b
c
12~ 25 50 100 Molecular weight xlO -3
% x e-
6
10
: t / ' - " " -
20 40 60 80 100120
o
20 30 40 5 0 " 150 160 170 180 190
Fractions
Fig. 4. Comparison of molecular weight, by sodium dodecyl sulfate gel electrophoresls, of informosomal proteins with r ibosomal proteins. The proteins from informosome were prepared as described in Material and Methods, ei ther from salt washed particles (use of buffer B for purif icat ion) or from informosomes kep t in buffer A (hypotonic) for preparat ion (see Materials and Methods). The proteins from ribosomal subuni ts were obtained from monor ibosomes dissociated by centrifugaUon in buffer B. The two separated subunits were precipi ta ted by two volumes e thanol and analysed by sodium dodecyl sulfate gel electro- phoresis (7.5% acrylamide). Stained gels were scanned at 580 rim. (a), Proteins obtained from salt washed informosomes; (b), proteins obtained from unwashed informosomes; (e), proteins obtained from the small r ibosomal subunit ; (d), proteins obtained from the large r ibosomal subunit . Asterisks indicate protein washed.
Fig. 5. "Binding factor" purif icat ion b~ affinity chromatography on RNA-sepharose. HeLa cells (5--105 cells/ml) were labelled for 24 h with [ °H] leuc ine (0.1 #Ci/ml) and fract i0nated as described in Materials and Methods to yield cytoplasmic supernatant . Cytoplasmic supernatant containing S0 mg protein was chromatographed on the RNA-sepharose (Materials and Methods). The RNA "binding factor" proteins were eluted with buffer B then with buffer A containing 0.5% sodium deoxycholate . The fractions were precipi ta ted with 10% cold tr ichloracetic acid in the presence of I 0 0 #g serum albumin, collected by fil- t ra t ion on glass filters and counted with approximate ly 50% efficiency. Binding act ivi ty of the purified proteins is shown in the inset, Mixtures of 10 ~g RNA and binding factor (in buffer A) were analysed by centr i fugat ion and the sedimenta t ion constant determined according to McEwen [36] . The increase in sedimenta t ion constant (AS) is p lo t t ed against the prote in concentrat ion. • -', proteins eluted with buffer B; o o, proteins a luted with 0.5% sodium deoxyeholate .
4. Fig. 4 shows that some proteins (30--40%) are sensitive to high salt (Aster- isks), but that other proteins are tightly bound to the mRNA. We have routine- ly used salt washed informosome to eliminate unspecific adsorbed proteins.
III. Characterization of the protein constituent of polysomal messenger ribonu- cleopro rein
Polysomal messenger ribonucleoproteins were investigated by the same pro-

379
cedure as selected for informosomes, i.e. by RNAase treatment of the particles. These particles were obtained from cells cultured for a short period at 43°C whereupon initiation of protein synthesis was blocked and most of the poly- somes were converted to monoribosomes (Liautard, J.P. and KShler, K., see reference 19) [10]. The 60 S--100 S fraction of the postmitochondrial super- natant consisted of ribosomes and messenger ribonucleoprotein complexes of former polysomes, but also informosomes and native ribosomes.
When treated with RNAase, again ribosomes conserved their physical integri- ty, while polysomal messenger ribonucleoproteins disappeared, and a number of proteins and oligonucleotides were found in the supernatant after high speed centrifugation (results not shown).
Besides polysomal messenger ribonucleoprotein, informosomes were degrad- ed also and contaminated the polysomal messenger ribonucleoproteins; how- ever, informosomes represent a maximum of 10% of the mRNA in HeLa cells (as judged by specific labelling of mRNA), and so their protein concentration is too low to be detected in these experiments. This deduction is confirmed by the comparison of the protein pattern from two kinds of particles (Fig. 7) where informosomal proteins are not detected with polysomal messenger ribo- nucleoproteins. Analysis by sodium dodecyl sulfate gel electrophoresis revealed many bands with molecular weights ranging between 120 000 and 35 000 (Fig. 7); there were however, two major bands with molecular weights of 75 000 and 50 000. As for informosomes, control samples did not carry any visible ribosomal or cytoplasmic contaminants. These proteins are resistant to high ionic strength (0.5 M KC1).
The two major proteins from HeLa cells have the same molecular weights as those previously found in various animals [16, 20--22]. This suggests a rather common structure for these particles and confirms the validity of the method we used.
IV. Isolation of cytosol proteins with affinity to RNA The cytoplasmic proteins which bind to RNA can theoretically be divided
into two classes: first, proteins with affinity to RNA alone; and second, pro- teins which only bind to RNA in cooperation with the first class proteins.
If only separated and purified proteins from the cytosol are tested for their ability to bind to RNA, the proteins of the second class can obviously not be found. By using affinity chromatography of cytosol on RNA-sepharose, how- ever, we were able to isolate both classes of proteins. (For experimental details see Materials and Methods). The bound proteins were eluted with the buffer B (0.5 M KC1) as one sharp peak. A second minor component was eluted with buffer A containing 0.5% desoxycholate (Fig. 5). Sodium dodecyl sulfate is known to remove proteins from ribonucleoproteins [23] which cannot be elut- ed by raising the ionic strength up to 0.5 M KCI alone.
Further characterization showed that the proteins eluted by 0.5 M KC1 were still able to bind to RNA. This was shown by the formation of RNA-protein complexes with increasing sedimentation constants [24] (Fig. 5). In contrast, the proteins eluted by sodium deoxycholate lost their affinity to RNA.
Both protein fractions were then analysed by sodium dodecyl sulfate gel electrophoresis (Fig. 6). Numerous protein bands are present in the fractions
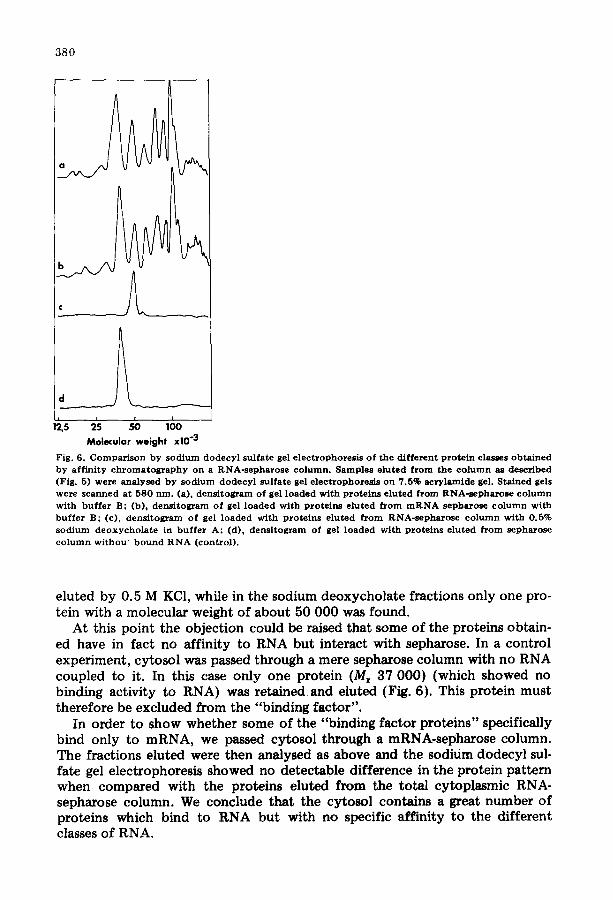
3 8 0
I 5 25 100
Molecular weight x l0 "3
Fig. 6. Comparison by sodium dodecyl sulfate gel electrophoresis of the different protein classes obtained by affinity chromatography on a RNA-sepharose column. Samples eluted from the column as described (Fig. 5) were analysed by sodium dodecyl sulfate gel electrophoresis on 7.5% acrylamide gel. Stained gels were scanned at 580 nm. (a), densitogram of gel loaded with proteins eluted from RNA-sepharose column with buffer B: (b), densitogram of gel loaded with proteins eluted from mRNA sepharose column with buffer B; (c), densitograrn of gel loaded with proteins eluted from RNA-sepharose column with 0.5% sodium deoxychola te in buffer A; (d), densitogram of gel loaded with proteins eluted from sepharose column withou~ bound RNA (control).
eluted by 0.5 M KC1, while in the sodium deoxycholate fractions only one pro- tein with a molecular weight of about 50 000 was found.
At this point the objection could be raised that some of the proteins obtain- ed have in fact no affinity to RNA but interact with sepharose. In a control experiment, cytosol was passed through a mere sepharose column with no RNA coupled to it. In this case only one protein (Mr 37 000) (which showed no binding activity to RNA) was retained and eluted (Fig. 6). This protein must therefore be excluded from the "binding factor".
In order to show whether some of the "binding factor proteins" specifically bind only to mRNA, we passed cytosol through a mRNA-sepharose column. The fractions eluted were then analysed as above and the sodium dodecyl sul- fate gel electrophoresis showed no detectable difference in the protein pattern when compared with the proteins eluted from the total cytoplasmic RNA- sepharose column. We conclude that the cytosol contains a great number of proteins which bind to RNA but with no specific affinity to the different classes of RNA.
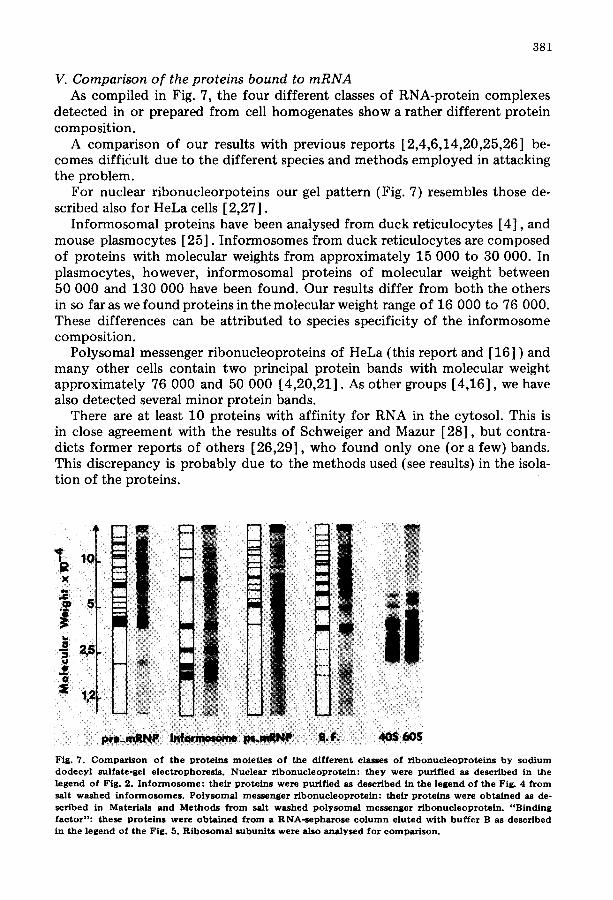
381
V. Comparison o f the proteins bound to mRNA As compiled in Fig. 7, the four different classes of RNA-protein complexes
detected in or prepared from cell homogenates show a rather different protein composition.
A comparison of our results with previous reports [2,4,6,14,20,25,26] be- comes difficult due to the different species and methods employed in attacking the problem.
For nuclear ribonucleorpoteins our gel pattern (Fig. 7) resembles those de- scribed also for HeLa cells [2,27].
Informosomal proteins have been analysed from duck reticulocytes [4], and mouse plasmocytes [25]. Informosomes from duck reticulocytes are composed of proteins with molecular weights from approximately 15 000 to 30 000. In plasmocytes, however, informosomal proteins of molecular weight between 50 000 and 130 000 have been found. Our results differ from both the others in so far as we found proteins in the molecular weight range of 16 000 to 76 000. These differences can be attributed to species specificity of the informosome composition.
Polysomal messenger ribonucleoproteins of HeLa (this report and [16] ) and many other cells contain two principal protein bands with molecular weight approximately 76 000 and 50 000 [4,20,21]. As other groups [4,16], we have also detected several minor protein bands.
There are at least 10 proteins with affinity for RNA in the cytosol. This is in close agreement with the results of Schweiger and Mazur [28], but contra- dicts former reports of others [26,29], who found only one (or a few) bands. This discrepancy is probably due to the methods used (see results) in the isola- tion of the proteins.
_
n
i iiii i n
m
iii i i~iiiii m ......
Fig. 7. C o m p a r i s o n of the prote ins moie t i e s o f the different classes of r i b o n u c l e o p r o t e i n s b y s o d i u m d o d e c y l sulfate-gel e lec t rophores i s . Nucleas r ibonuc leoprote in: they were pur i f ied as descr ibed in the legend of Fig. 2. I n f o r m o s o m e : their prote ins were pur i f ied as descr ibed in the l egend of the Fig. 4 f r o m salt w a s h e d i n f o r m o s o m e s . P o ly s oma l me s s e nge r r i b o n u c l e o p r o t e i n : the i r p ro te ins were o b t a i n e d as de° sc r ibed in Mater ia ls and M e thods f r o m salt washed p o l y s o m a l messenge r r i bonuc l eop ro t e in . " B i n d i n g factor": these prote ins were o b t a i n e d f r o m a RNA-s e pha rosc c o l u m n e luted with b u f f e r B as descr ibed in the legend of the Fig. 5. R i b o s o m a l subun i t s were also ana lysed for co mp a r i so n .

382
Of the many proteins discovered in the four kinds of r ibonucleoprotein par- ticles, two are likely to be of special interest. It has been reported that a pro- tein {molecular weight approximately 78 000) is attached to the poly (A) se- quence at the 3' end of mRNA [30] . As seen from Fig. 7 a protein of this size is present in informosomes, polysomal messenger ribonucleoprotein, nuclear r ibonucleoprotein and among the "binding factors". We have detected poly (A) sequences in the informosome, polysomal messenger r ibonucleoprotein and nuclear r ibonucleoprotein {unpublished results).
Another protein with a molecular weight of approximately 50 000 is on polysomal messenger r ibonucleoprotein and among the "binding factors", but is absent from informosomes. We have recently shown that this protein pro- motes binding of polysomal messenger ribonucleoproteins to small ribosomal subunits {submitted for publication).
The presence of some proteins common to the nuclear particle (nuclear ribo- nucleoprotein) and the polysomal messenger r ibonucleoprotein in the rat liver, was reported [31--33] , but the authors did not characterize further the pro- teins and used either sodium deoxycholate-sensitive proteins [33] or a group of basic proteins related to messenger r ibonucleoproteins [31] . In contrast to ret iculocytes no common proteins have been found [34--36] , (see also [9] for review).
From our results we can conclude that in spite of the fact that some proteins are common to one or another kind of messenger r ibonucleoprotein complexes, it is evident that the majori ty of proteins differs .
The role of the "binding factor" proteins remains unclear but some of them could be a reservoir of proteins for the activation of mRNA. This is suggested by the presence of two proteins in this fraction with a molecular weight close to these found for the two major proteins in the polysomal messenger ribonu- cleoprotein.
Acknowledgements
We thank our colleague J. Kreft for discussion and helping us in the redaction of the manuscript. The help of Mr. and Mrs. B. Smith in translating the manu- script is gratefully acknowledged. This work has been supported by the Deut- sche Forschungsgemeinschaft.
References
1 Samarina, O.]~., Lukanldin, E.M., Molnar, J. and Georgiev, G.P. (1968) J. Mol. Biol. 33, 251--263 2 Pederson, T. (1974) J. Mol. Biol. SS, 163--183 S Spirin, A.S., Belltsina, N.V. and Ajtkhozhin, M.A. (1964) Zhurnal Obschei Biologii (USSR) 25, 321 4 Gander, E., Stewart, A., Morel, C. and Scher~r, K. (1973) Eur. J. Biochem. 38, 443--452 5 Perry, R.P. and Kelley, D.E. (1968) J. Mol. Biol. 35, 37--59 6 Lebleu, B., Marbaix, G., Huez, G., Temmermann, J., Burny, A. and Chantrenne, H. (1971) Eur. J.
B i o c h e m . 19, 264--269 7 Spirin, A.S. (1969) Eur. J. Biochem. 10, 29--35 S Baltimore, D. and Huang, A.S. (1970) J. Mol. Biol. 47, 268--273 9 Williamson, R. (1973) FEBS Lett. 37, 1--6
10 Schochetman, G. and Perry, R ~ . (1972) J. Mol. Biol. 63, 577--566 I I Perry, R.P., La Torre, J., Kelley, D.E. and Greenberg, J. (1972) Biochim. Biophys. Acta 262, 220--
226

383
12 Lindberg, U. and Persson, T. (1972) Eur. J. Biochem. 31, 246--254 13 Weber, K., Pringle, J.R. and Osbom, M. (1972) Method in Enzymology, VoL XXVI, 3--27, Academic
Press, New York 14 Gallinaro-Matringe, H., Stevenin, J. and Jacob, M. (1975) Biochemistry 14, 2547--2554 15 Warner, J.R., Knopf, P. and Rich, A. (1963) Proc. Natl. Acad. Sci. U.S. 49, 122--129 16 Auerbach, S. and Pederson, T. (1975) Biochem. Biophys. Res. Commun. 63, 149--153 17 K~hler, K. and Arends, S. (1968) Eur. J. Biochem. 5, 500--508 18 Chattergee, N.K. and Weissbach, H. (1973) Arch. Biochem. Biophys. 157, 160--167 19 Liautard, J.P. and K~hlel, K. (1976) Biochimie, in press 20 Blobel, G. (1972) Biochem. Biophys. Res. Commun. 47, 88---95 21 Bryan, R.N. and Hayashi, M. (1973) Nat. New Biol. 244, 271--274 22 Lindberg, U. and Sundquist, B. (1974) J. Mol, BioL 86,451--468 23 Olsnes, S. (1971) Eur. J. Biochem. 23, 57--563 24 Girard, M. and Baltimore, D. (1966) Proc. Natl. Acad. Sci. U.S. 56,999--1002 25 Egly, J.M., Krieger, O., Mandal, P. and Kempf, J. (1974) FEBS Lett. 40, 101--105 26 Schweiger, A. and Hannig, K. (1971) Biochim. Biophys. Acta 254, 255---264 27 Ducamp, C. and Jeanteur, P. (1973) Biochimie 55, 1235--1243 28 Schweiger, A. and Mazur, G. (1974) FEBS Lett. 46, 255--259 29 Preobrazhensky, A.A. and Ovchinnikov, L.P. (1974) FEBS Lett. 41, 233--237 30 Blobel, G. (1973) Proc. Natl. Acad. Sci. U.S. 70, 924--928 31 Schweiger, A. and Hannig, K. (1970) Biochim. Biophys. Acta 204, 317--324 32 Tomcsanyi, T. and Tigyi, A. (1971) Acta Biochim. Biophys. Acad. Sci. Hung. 6, 149--154 33 Olsnes, S. (1970) Eur. J. Biochem. 15, 464--471 34 Lukanidin, G.M., Georgiev, G.P. and Williamson, R. (1973) Mol. Biol. U.S.S.R. 7, 264--268 35 Morel, C. Kayibanda, B. and Scherrer, K. (1971) FEBS Lett. 18, 64--88 36 Lukanidin, E.M. and Olsnes, S. and Pihl, A. (1972) Nat. New Biol. 240, 90---92 37 McEwen, C.R. (1967) Anal. Biochem. 20, 114--149





























