Embed Size (px)
Citation preview

422
Combinatorial peptide libraries in drug design: lessons from
venomous cone snails Baldomero M. Olivera, David R. Hillyard, Maren Marsh and Doju Yoshikami
Many present-day drugs are derived from compounds that are natural products, a
traditional source of which is fermentation broths of microorganisms. The venoms
of cone snails are a new natural resource of peptides that may have a
pharmaceutical potential equivalent to those from traditional sources, particularly
for developing drugs that target cell-surface receptors or ion channels. In effect, cone
snails have used a combinatorial library strategy to evolve their small, highly bioactive
venom peptides. The methods by which the snails have generated thousands of
peptides with remarkable specificity and high affinity for their targets may provide
important lessons in designing combinatorial libraries for drug development.
The cone snails (Conus; Fig. la) are a remarkable group of marine predators that have evolved a phar- macology that uses powerful peptide drugs to elicit specific physiological responses from their prey, preda- tors and competitors lm3. They appear to exploit a combinatorial peptide library strategy to develop ven- oms, the primary purpose of which is to incapacitate prey Many peptide sequences are apparently explored through hypermutation in the evolution of the peptide- rich venom of each Conus species. This strategy is probably a major factor in the notable evolutionary success of this group: Conus is among the largest gen- era of living marine invertebrates, comprising about 500 different species.
In this article, we focus on insights that have been gained from this 50 million-year history of successful drug development by cone snails that are clearly rel- evant to present-day efforts to use combinatorial pep- tide libraries for drug design. In addition, specific venom components may have a variety of direct medi- cal applications; indeed, one particular Conus peptide is currently undergoing clinical trials as a potent anal- gesic for intractable pain (G. Miljanich, pers. commun.).
Conus biology: how to succeed in evolution by really diversifling
Cone snails are carnivorous predators that practise biochemical warfare and use venom as their major weapon for capturing prey. Their efficient drug-deliv-
B. M. Olivera and D. Yoshikami are ivr the Department ofBiology, University of Utah, Salt Lake City, UT 84112, USA. D. R. Hillyard and M. Marsh are in the Department ofPathology, University of Utah, Salt Luke City, UT 842 12, USA.
TIBTECH OCTOBER 1995 NOL 131
ery system, which they appear to have developed more than 50 million years ago, involves injecting their venom by means of a disposable hypodermic needle made out of chitin (Fig. lb).
The bioactive components of Conus venom are mostly small peptides that are typically lo-30 amino acid residues in length4, and each of the 500 species of cone snails has its own distinctive complement of venom peptides. In the more complex venoms, there may be over 100 different peptides’,‘, each of which is found only in the venom of a given species. There- fore, it seems probable that in toto, tens of thousands of different pharmacologically active peptides are present in the venoms of Conus species.
All peptides that are present in the venoms of cone snails are initially made as larger precursors, usually 60-90 amino acid residues in length1s5-’ (Fig. 2), from which the mature peptide (lo-30 amino acid residues), which is located at the C-terminus, is ex- cised by proteolytic cleavage. A comparison of peptide precursor sequences reveals a remarkable dichotomy; within each precursor, two sectors can be readily identified - a constant region at the N-terminus, and extremely hypervariable regions towards the C-termi- nus. As Conus species have diverged during the course of their evolution, hypermutation of the C-terminal variable regions encoding the toxins has taken place; this has led to sequence diversification of the mature peptides. Thus, when peptides from two different Conus species are compared, even in instances where almost no amino acid sequence homology is observed in the mature peptide regions, very few base-pair and amino acid changes are seen in the constant N-ter- minal regions of the peptide precursors (Fig. 2).
0 1995, Elsevler Science Ltd 0167 - 7799/95/$9.50
423
reviews
Even when they are geographically widely separ- ated, individuals of the same species do not exhibit unusual peptide polymorphism - apparently, hyper- mutation occurs primarily when Contrs species diverge. Interspecies comparisons suggest unprec- edented divergence of peptide sequences within this single genus. A comparison of peptides from different species targeting the same receptor sites has revealed that the amino acids in the mature peptide are hyper- variable, except for Cys residues, which are conserved. The disulfide bridges formed by Cys residues are prob- ably responsible for this conservation,
Our current understanding of how this peptide diversification occurred is represented diagrammati- cally in Fig. 3. In effect, the scheme shown is a com- binatorial peptide library strategy2,Y. A very large num- ber of peptide sequences were presumably generated, and mostly discarded by natural selection as new Contrs species evolved. In the venoms of extant Conus species, only the pharmacologically useful structures are prob- ably present - the ‘winners’ in natural selection. Alter- natively, considerable ‘noise’ (e.g. peptides without any function) might have been retained in Conus venoms. However, the high fraction of biologically active Conus peptides makes this unlikely; peptides with no appar- ent biological activity in an initial screen are often sub- sequently shown to be active with a different bioassay.
The Contrs peptides provide an insight into a successful endpoint of a combinatorial peptide library approach to drug development, and in the sections below, we summarize some general features of these peptide ‘winners’.
Lesson one: stifleen yourframework The structures of Conus peptides suggest that it may
more be productive to use a combinatorial library of conformationally constrained peptides for drug devel- opment, rather than a library of linear peptides. In contrast to most peptides 10-30 amino acid residues in length, which would be expected to have flexible conformations in solution, the striking conclusion is that COWS peptides are invariably conformationally constrained. Most COWS peptides fall into three major classes, each with a characteristic disulfide-bonded framework’,4 (Table 1). NMR analyses of all three classes of peptides in solution indicate that the multiple disulfide bonds confer considerable rigidity to the con- formation of the peptide’@i*.
The three types of disulfide-bonded structures shown in Table 1 provide the cone snails with a range of different-sized frameworks. Thus, the smallest major framework is found in Contrs peptides that target the acetylcholine binding site of the nicotinic acetyl- choline receptor, while peptides that inhibit calcium channels have the largest framework. Even Con~r pep- tides that have no disulfide bonds at all, the conan- tokinsi’, adopt an a-helical conformation in the presence of calcium ions; these peptides contain y-carboxyglutamate, whose negatively charged car- boxy1 groups chelate Ca2+ and confer structural rigid- ity to the peptide”. This exception reinforces the
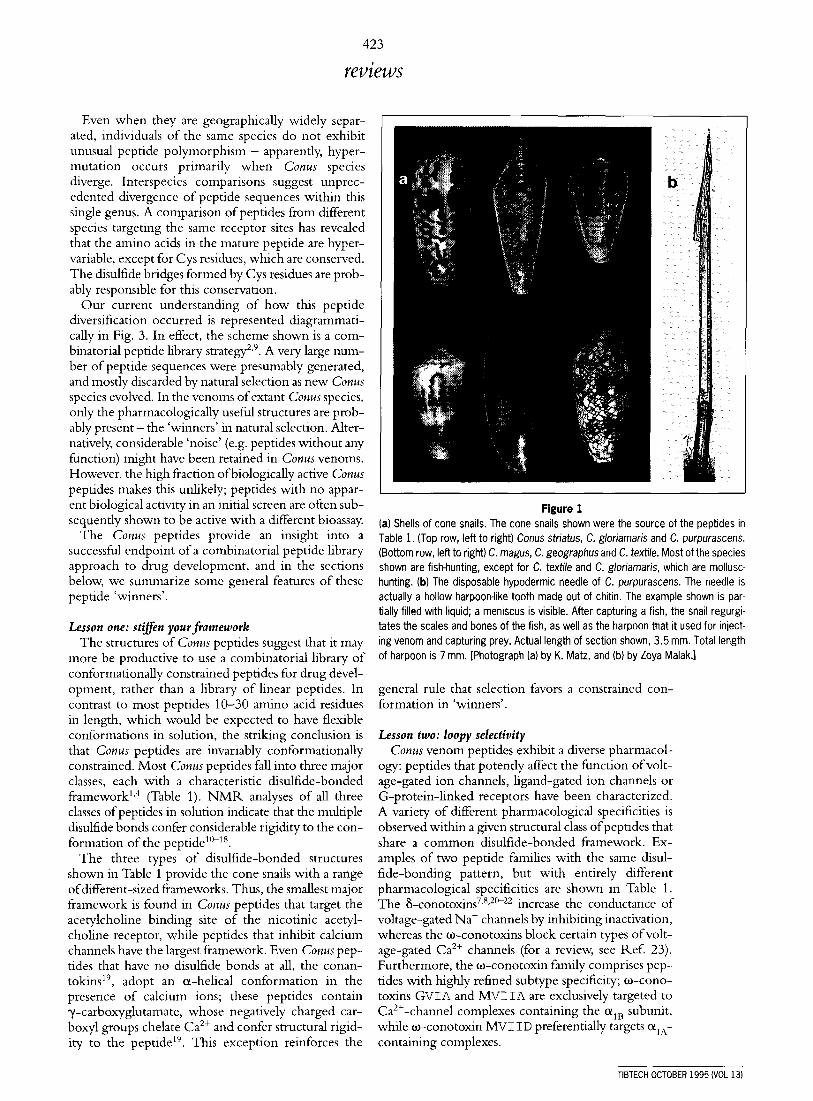
Figure 1 (a) Shells of cone snails. The cone snails shown were the source of the peptides in Table 1. (Top row, left to right) Conus striatus, C. gloriamaris and C. purpurascens. (Bottom row, left to right) C. magus, C. geographus and C. textile. Most of the species shown are fish-hunting, except for C. textile and C. gloriamaris, which are mollusc- hunting, (bl The disposable hypodermic needle of C. purpurascens. The needle is actually a hollow harpoon-like tooth made out of chitin. The example shown is par- tially filled with liquid; a meniscus is visible. After capturing a fish, the snail regurgi- tates the scales and bones of the fish, as well as the harpoon that it used for inject- ing venom and capturing prey. Actual length of section shown, 3.5 mm. Total length of harpoon is 7 mm. [Photograph (a) by K. Matz, and (b) by Zoya Malak.1
general rule that selection favors a constrained con- formation in ‘winners’.
Lesson two: loopy selectivity Contls venom peptides exhibit a diverse pharmacol-
ogy: peptides that potently affect the function of volt- age-gated ion channels, ligand-gated ion channels or G-protein-linked receptors have been characterized. A variety of different pharmacological specificities is observed within a given structural class of peptides that share a common disulfide-bonded framework. Ex- amples of two peptide families with the same disul- fide-bonding pattern, but with entirely different pharmacological specificities are shown in Table 1. The 6-conotoxins7s,2@22 increase the conductance of voltage-gated Na+ channels by inhibiting inactivation, whereas the o-conotoxins block certain types of volt- age-gated Ca2+ channels (for a review, see Ref. 23). Furthermore, the o-conotoxin family comprises pep- tides with highly refined subtype specificity; o-cono- toxins GVIA and MVI IA are exclusively targeted to Ca2+-channel complexes containing the aiB subunit, while o-conotoxin MVI ID preferentially targets oiA- containing complexes.
TIBTECH OCTOBER 1995 (VOL 13)

424
reviews
&TXVIA S-GmVIA &PVIA
10 20 30 40 MKLTCMMIVAVLFLTAWTFATADDPRNGLGNLFSNAHHEMKNP MKLTCMMIVAVLFLTAWTFVTADDSGNGMEILFPKAGHEMENL MKLTCVMIVAVLFLTAWTFVTADDSKNGLENHFWKARDEMKNR
L
EASKLN5KOR--WCrcgsc~D*~CDG~~IVL~T EVS--N-RVKFCRKEGQLCDPIFQNCCRGWhC-VLFCV EASKLDKK-EAC WAPGTFCGIKPGLCCSEF-CLPGVC:FGG
Figure 2 The amino acid sequences of the precursors of &conotoxins from three different Conus species were determined from cDNA clones, The numbers refer to the amino acids in the &TXVIA precursor sequence. The mature peptide consists of amino acids 52-78. Amino actds in the hypervariable regions are italicized. While 60% of all amino acids in the N-terminal prepro sequence (numbers l-51) are identical in all three sequences, in the hypervariable regions of the mature peptide ~5% are identical. By contrast, 67% of amino acids are different In all three peptides at the hypervariable IOCI; in the constant region, the corresponding frequency is ~8%. The conserved Cys residues of the mature peptide involved in disulfide bonding are depicted in bold.
Mature peptides I I cm+mrcmqq
, , I
Figure 3
Peptide diversification during Conus speciation. The ‘cartoon’ depicts how Conus pep- trde precursor sequences may change during the evolution of a new species. The mature peptides, found at the C-terminal end of the precursors shown, are produced by proteolytic cleavage (small arrow). During spectation, the N-terminal region, as well as the Cys residues in the mature peptide, remains conserved, whereas areas rep- resented by assorted symbols are selectively hypermutated. The result is a set of mature peptrdes that have a conserved disulfide-bonded framework wtth hypermutated amino acrds between Cys residues (two examples are shown at the bottom of the fig ure). The generation of novel sequences during Conus speciation may be non-random (in this cartoon, this is depicted by ‘loaded’ dice). Peptides that are represented by this cartoon are the w- and &conotoxins that are shown in Fig. 1 and Table 1.
The pharmacological selectivity can be remarkable: w-conotoxins GVIA and MVI IA, each from a dif- ferent Conus species, discriminate among various sub- types of voltage-gated Ca2+ channels with a selectivity ratio of 1:lO’. Both peptides are specifically targeted to the same Ca’+-channel subtype, and have become the defining pharmacological reagents for that subtype. Despite this pharmacological similarity, the two o-conotoxins have strikingly divergent primary sequences - 76% of amino acids at the hypervariable loci are different”.
Functionally, it is envisaged that many proteins use a relatively rigid structural framework from which
loop amino acids emanate. It has been proposed that such looped regions provide much of the biological specificity for antibodies, ion channels and the active sites of enzynles24. In essence, a Conus peptide may represent the ultimate reduction of a DNA-encoded protein structure that has been designed for high biological specificity, comprising a minimal structural framework (defined by disulfides), while maximizing the proportion of loops that confer high selectivity. Hypermutation of such loops could rapidly lead to new biological function, making the system particu- larly appropriate for diversification through a combi- natorial library selection strategy
Lesson three: selectivity + synergy = success Understanding the functional biology of Conus
peptides has lagged behind their identification and biochemical characterization. Nevertheless, the data strongly indicate that the combinatorial peptide library approach can result in sufficient selectivity to make multiple-drug synergy feasible. The underlying mecha- nisms of action of conotoxins on prey physiology are best understood for studies of several species of fish- hunting cone snails.
Prey capture can have two distinct phases: an initial stage, during which the major fish fin musculature is rapidly incapacitated (possibly by tetanic paralysis), fol- lowed by an irreversible stage of paralysis, in which neuromuscular transmission becomes nonfunctional. For the latter stage, multiple peptidic components from a single venom act synergistically, and each pep- tide acts at a distinct target site to interfere with neuro- muscular transmission. However, by acting together, a physiological effect that is far more potent than the simple sum of individual activities is produced. Thus, in this case, the peptide library strategy may have pro- duced an interacting ensemble of drugs, each with a different target.
The reason why multiple drug ensembles make the high specificity of target subtypes a biological neces- sity for each individual drug is illustrated by the following example: a single Contrs venom contains not only a peptide that blocks the Na+ channel, but also a peptide that increases Na+ conductance by inhibiting Na+-channel inactivation. It appears that the peptidic component that increases Na+-channel conductance - a 8-conotoxin7 - plays a role in the early knock-out of the major fin musculature of the fish, while the Na+- channel blocker (a p=conotoxin) contributes to the final inhibition of muscle contraction. If the two peptides were to act simultaneously on the same set of Na+ channels, they would antagonize each other’s action. However, by acting on different molecular forms of NaC channels, each peptidic drug can help to elicit a different physiological phase of prey immobi- lization. This multiple-drug synergy, which maximizes the snail’s ability to capture fish, would be far less effective if each individual peptide drug were not subtype-specific.
The continuing success of the cone snail strategy for drug development underlines the great potential of a
TIBTECH OCTOBER 1995 NOL 13)

425
reviews
Table 1. Mature Conus peptide sequencesa
Toxin Sequenceb Conus species Ref.
6-conotoxinsc TXVIA WCKQSGEtMCNLL~NCCW-CIVLVCT C. textile 20 GmvIA VKPCRKEGQLCDPIFQNCCRGWNC~VLFCV C. gloriamaris PVIA EACWAPGTFCGIKPGLCCSEF-CLPGVCFG* C. purpurascens ;
w-conotoxinsC 23 GVIA CKSCGSSCSOTSYNCCR-SCNOYTKRCY" C. geographus SVIA CRSSGSOCGVTSI-CC-GRC--YRGKCT* C. striatus MVIIA CKGKGAKCSRLMYDCCTGSCRS--GKC* C. magus MVIID CQGRGASCRKTMYNCCSGSCNP--GRC* C. magus
Disulfide I
c------c------cc---c------c bonding -1
cw-conotoxins 4 GI ECCNPACGRHYSC* C. geographus SIA YCCHPACGKNFDC* C. striatus
Disulfide I ~---C-----C
bonding L’
p,-conotoxins 4 GIIIA RDCCTCOKKCKDRQCKCQRCCA* C. geographus
I Disulfide
I cc+----c----c----(yJ bonding L-
aAll peptides shown have been chemically synthesized. b. represents 4-Pans-hydroxyproline; * denotes peptides that are amidated at the C-terminus. c&conotoxins and oconotoxins have different pharmacological specificities, despite sharing the same disulfide-bonding framework.
future pharmacology that routinely uses synergistic multiple-drug ensembles. Following the snails’ lead, this would involve the use of combinatorial peptide library strategies to develop drugs with much more highly refined target selectivity than that afforded by present-day pharmaceuticals.
Concluding remarks What we have learned so far about the Conus pep-
tide system has clear implications for the use of com- binatorial library strategies in drug design.
In many ways, the shared structural frameworks of the conotoxins are analogous to the conserved struc- tural framework of antibodies. In both systems, the underlying biology requires the generation of ligands of extremely high specificity and affinity. In effect, the immune system uses a combinatorial library strategy as well, except that the structural framework is much larger. The insights that have been gained from the Conus peptide system, together with what is known about the immune system, suggest that it may be difficult to generate a set of diverse ligands with highly refined target specificity through a combinatorial library search if a stable structure had to be searched for simultaneously. In both systems, the search pattern is restricted to members of a set of ‘pre-constrained structures. Cone snails primarily use three structural frameworks of different sizes, which seem to fulfill their biological needs.
When a peptide with a given framework is an effec- tive ligand against a particular subtype of receptor or
ion channel, it seems reasonable to assume that other peptides sharing the same structural framework could be specific for different subtypes of the same receptor or ion channel family (i.e. structurally related ligands for structurally related targets). Therefore, it may be efficient to use the same framework for evolving ligands of different specificity, which is apparently what cone snails have done. This seems good advice for people using a combinatorial library strategy to design drugs; a given structural framework that works for one subtype of a receptor or ion channel could be a good starting point for developing new ligands that affect different subtypes. It may be too much to search for a framework that works, while at the same time search- ing for the loopy amino acids that confer the requisite ability to discriminate between closely related sub- types. Consequently, a framework that is already known to work for the ion channel or receptor family may be an important prerequisite for a success- ful combinatorial library search for peptides targeting a similar functional site in other subtypes.
Clearly, however, more is involved than just struc- tural frameworks. Enormous sequence differences exist between the members of a family; indeed, two &conotoxins do not obviously share more sequence identity with each other than with an w-conotoxin. Nevertheless, it is clear that other general characteristics of the peptides in these two conotoxin families play a role in conferring specificity for very different targets. The w-conotoxins always have a high net positive charge, and very few strongly hydrophobic amino
TIBTECH OCTOBER 1995 (VOL 13)

426
reviews
acids. By contrast, although the total number of charged amino acids in &conotoxins is roughly equivalent, the net charge is much closer to neutral, and these peptides always have a high proportion of strongly hydrophobic amino acids. Clearly, such features must be important to allow the peptides to access their target site; it has been hypothesized that, in view of the large number of hydrophobic amino acids in &conotoxins, the binding site is at least partially in the lipid bilayer.
These ‘general’ characteristics must also be consid- ered when designing effective libraries. Although it would be desirable to examine hypermutated variants that target either the o-conotoxin or &-conotoxin sites of a particular channel subtype, peptide libraries could be biased to produce peptides with the ‘general’ desired characteristics; this would presumably improve the chances for success.
Acknowledgements The research of the authors has been supported by
grant PO1 48677 from the National Institute of General Medical Sciences.
References 1 Olivera, B. M. et al. (1990) Science 249, 257-263 2 Ohvera, B. M., Rivier, J., Scott, J, K., Hillyard, D. R. and Cruz,
L. J. (1991)]. Biul. Chem. 266, 22067-22070 3 Myers, R. A., Cruz, L. J., Rivier, J. and Olivera, B. M. (1993)
Chem. Rev. 93. 1923-1936
4 5
6
7
8 9
10
11 12
13 14
15
16
17
18 19
Olivera, B. M. ef al. (1985) Science 230, 1338-1343 Woodward, S. R., Cmz, L. J., Olivera, B. M. and Hillyard, D. R.
(1990) EMBO]. 1. 1015-1020 Colledge, C. J., Hunsperger, J. P., Imperial, J. S. and Hillyard, D. R. (1992) Tvxiun 30, 1111-1116 Shon, K-J., Hasson, A., Spira, M. E., Cruz, L. J., Gray, W. R. and Olivera, B. M. (1994) Biochemistry 33, 11420-l 1425 Shon, K. el al. (1995) Biochemistry 34, 4913-4918
Scott, J. K. and Smith, G. P. (1990) S&w 249, 386-390 Kobayashi, Y., Ohkubo, T., Kyogoku, Y., Nishmchi, Y. and Sakakibara, S. (1988) in Peptide Chemistry (Shiba, T. and Sakakibara, S., eds), p. 163, Protein Research Foundation, Osaka, Japan
Kobayashi, Y. et al. (1989) Biuchemistry 28, 4853-4860 Pardi, A., Goldes, A., Florance, J. and Maniconte, D. (1989) Biochemistry 28, 5494-5501 Law&n, J-M. et al. (1991) Biudzemisrry 30, 6908-6916 Ott, K-H., Becker, S., Gordon, R. D. and Rutejans, H. (1991)
FEBS Len. 278, 16&166 Davis, J. H., Bradley, E. K., Miljamch, G. P., Nadasdl, L., Ramachandran, J. and Basus, V. (1993) Biochemistry 32, 7396-7405 Pallaghy, P. K., Duggan, B. M., Pennington, M. W and Norton, R. S. (1993)]. Ma!. Bioi. 234, 405-420 Sevllla, P., Bnnx. M., Satoro, J., Gago, F. and Garcia, A. G. (1993) Biochem. Biuphys. Res. Comma. 192, 1238-1244 Nemoto, N. et al. (1995) Bkxhem. Biophys. Ref. Cummun. 207,695-700 McIntosh, J. M., Olivera, B. M., Cmz, L. J. and Gray, W. R. (1984)
/. Biol. Gem. 259, 14343-14346 20 Hillyard, D. R. ef al. (1989) Biochemisiry 28, 358-361 21 Fainzilber, M., Kofman, O., Zlotkin. E. and Gordon, D. (1994)
1, Biol. Chem. 269, 2574-2580 22 McIntosh, J. M. et al. (1995)J. Biul. C&em. 270, 16796-16802 23 Olivera, B. M., Miljamch, G., Ramachandran, J. and Adams, M. E.
(1994) Annu. Rev. Biochem. 63, 823-867 24 MacKinnon, R. (1995) Neuron 14,889-892
Constrained peptides as binding entities Robert C. Ladner
Displaying proteins and peptides on genetic packages and selecting packages that
display high-affinity binders allows large numbers of peptidyl compounds to be
tested for binding to targets. Phage-displayed libraries of unstructured peptides
(UPS) have yielded binders for some targets, but not for many others. Therefore,
more attention is being paid to peptidyl compounds that have varied regions that
are subject to conformational constraint.
Genetic packages (i.e. replicable entities such as cells or viruses - especially phage - see Glossary) can dis- play a wide variety of peptidyl compounds’-3. Although large numbers of linear unstructured pep- tides (UP) have been displayed, there are many targets for which no high-affinity binders can be found4s5. Imposing constraints on the displayed peptidyl mol- ecules can confer a number of important advantages
R. C. Ludner is at Protein Engineering Corporatiovt, 765 Concord Avenue, Cambridge, MA 02138, USA.
that overcome at least some of the shortcomings of unstructured peptides2v3.
The term ‘constrained peptide’ (CP) is used to describe a variety of forms of peptide, from a disul- fide-closed loop @CL) of amino acids, to one or more peptide segments embedded in a protein3. The residues of a DCL are only partially constrained by the disulfide - the larger the loop the less the constraint - and the amino acid sequence within the loop can impose additional constraint3,6; for example, Pro and, to a lesser extent, Val and Ile, limit the freedom of the
TlBTECHOCTOBER1995&'OL 13) 01995, Elsewier Science Ltd 0167- 7799/95/$9.50





























