Embed Size (px)
Citation preview

Symposium contribution / Contribution à un symposium
Climate change: can we predict the impacts onplant pathology and pest management?1
H. Scherm
Abstract: The science of climate change has matured considerably during the past decade, both relative to the strengthof the evidence documenting the ongoing anthropogenic climate change and in terms of the quality of climate modelsprojecting future changes in climate. Concomitantly, modeling studies to project the likely impacts of climate changeon agricultural production also have become more sophisticated. Nonetheless, agricultural impact assessments still donot account for all important factors; for example, potential changes in yield losses due to altered dynamics andintensity of pests (insects, plant pathogens, and weeds) under climate change are generally ignored — an importantomission, given the significant role of plant pests in constraining the production of food and fiber worldwide. Thispaper highlights selected challenges that must be overcome before we can hope to quantify the impacts of a changingclimate on plant disease intensity and yield loss. They pertain both to retrospective analyses seeking to identifyfingerprints related to climate change in long-term plant-disease records, as well as to the use of mathematical modelsto predict likely impacts on plant pathosystems in the future. The use of climate-change fingerprints has been limitedbecause time series containing disease variables collected in a standardized manner are unavailable for most plantpathogens; in cases where such long-term records do exist, trends are often confounded by changes in biological ormanagement factors over time. As for the use of models for impact assessment, key challenges pertain to uncertainty ininput variables, the difficulty in predicting biological responses in the presence of nonlinearities and thresholds, and thehigh likelihood of genetic adaptation to climate change.
Key words: climate change, disease management, epidemiological models, pathogen evolution.
273Résumé : La science des changements climatiques a considérablement évolué durant la dernière décennie, en vertu à lafois de la vigueur des signes de changement climatique anthropique en cours et de la qualité des modèles climatiquesprédisant les futurs changements de climat. En même temps, les études de modélisation visant à faire des projectionssur les impacts vraisemblables des changements climatiques sur la production agricole sont aussi devenues plussophistiquées. Cependant, les évaluations de l’impact sur l’agriculture ne tiennent pas compte de tous les facteursimportants; par exemple, les changements potentiels dans les pertes de rendement dues à une dynamique et une vigueurdifférentes chez les ravageurs (insectes, agents phytopathogènes et mauvaises herbes) en situation de changementclimatique sont généralement ignorés — omission de taille, compte tenu du rôle important des ravageurs dans lalimitation de la production d’aliments et de fibres dans le monde. Le présent exposé met en lumière quelquesdifficultés qui devront être aplanies avant que nous puissions espérer quantifier les effets d’un climat en évolution surl’intensité des maladies des plantes et les pertes de rendement. Elles ont rapport à la fois aux analyses rétrospectivescherchant, dans les anciens relevés de maladies, des traces de changement climatique, et à l’utilisation de modèlesmathématiques pour prédire, dans les années à venir, les effets vraisemblables sur les pathosystèmes végétaux.L’utilisation des traces de changement climatique a été limitée parce que, pour la plupart des agents pathogènes, desséries chronologiques contenant des variables sur les maladies recueillies de manière standard n’existent pas; dans lescas où ces données à long terme existent, les tendances sont souvent confondues avec des changements de naturebiologique ou dans les techniques agronomiques qui ont eu lieu avec le temps. De même que pour l’utilisation demodèles pour études d’impact, les principaux défis concernent l’incertitude en ce qui a trait aux variables utilisées, ladifficulté à prédire les réponses biologiques en présence de non-linéarités et de seuils et la probabilité élevéed’adaptation génétique aux changements climatiques.
Mots clés : changements climatiques, lutte contre les maladies, modèles épidémiologiques, évolution des agentspathogènes.
Can. J. Plant Pathol. 26: 267–273 (2004)
267
Accepted 21 June 2004.
H. Scherm. Department of Plant Pathology, University of Georgia, Athens, GA 30602, USA (e-mail: [email protected]).
1This paper was a contribution to the symposium entitled Climate Change — Impact on Plant Protection, held during The CanadianPhytopathological Society Annual Meeting in Montréal, Quebec, June 2003.

Scherm: climate change / predicting impacts on plant pathologyIntroduction
Since the late 1980s and early 1990s, when studies to as-sess the potential impacts of climate change on agriculturalproduction first entered the scientific mainstream (e.g., Ad-ams et al. 1990; Easterling et al. 1993; Parry et al. 1990;Parry 1992), the science of climate change has maturedconsiderably. For example, there is now much stronger evi-dence that global and regional climates have indeed beenchanging during the recent past, consistent with projectionsmade by climate-change models, and that human activities,primarily via increased greenhouse-gas forcing as a resultof the combustion of fossil fuels, are very likely responsiblefor these changes in climate (Houghton et al. 2001). Indeed,the recorded data show that global mean surface tempera-ture has risen by 0.6 ± 0.2 °C during the 20th century, withthe 1990s having been the hottest decade during that period.Furthermore, statistical analyses of long-term monitoringdata revealed that species across a wide range of ecosys-tems have already responded to the ongoing warming by al-tering their phenology (e.g., earlier timing of first bloom inplants or of spring migration in birds) and (or) their geo-graphical distribution (e.g., poleward range expansion intemperate-zone species) (Hughes 2000; Parmesan and Yohe2003; Root et al. 2003; Walther et al. 2002).
Physically-based climate models have become more so-phisticated, and confidence in their ability to realisticallyproject future changes in climate has increased (Houghtonet al. 2001). This is due, in part, to improved incorporationof processes such as sea-ice dynamics and ocean heat trans-port into these models. As a result, the latest models pro-duce satisfactory large-scale simulations of the currentclimate and of climate changes experienced during the re-cent past; the capability to reproduce interannual climatecycles such as the El Niño – Southern Oscillation (ENSO)has improved as well. However, the ability to model re-gional climate changes or changes in climate variability andextreme events is still limited (Houghton et al. 2001).
Modeling studies to project the likely impacts of climatechange on agricultural productivity and food security alsohave become more refined during the past decade. A recentstudy by the Agriculture Assessment Group within the USGlobal Change Research Program (USGCRP) is a good ex-ample (Reilly et al. 2003). Whereas previous studies uti-lized doubled-CO2 equilibrium scenarios as input for impactassessment, the USGCRP approach employed a more realis-tic transient scenario with gradually increasing greenhouse-gas concentrations. The assessment also included provisionsfor changes in aerosol concentrations (e.g., SO2) and for theincreased climate variability expected to occur in a warmerworld (Easterling et al. 2000; Stenseth et al. 2002). How-ever, although impacts on pesticide use were included in themodel, potential changes in yield losses due to altered dy-namics and intensity of pests (insects, plant pathogens, andweeds) under climate change were not considered byUSGCRP or other recent agricultural impact assessments —an important omission, given the significant role of plantpests in constraining the production of food and fiberworldwide (Oerke et al. 1994). Indeed, given our rudimen-tary level of understanding of how climate change mightimpact plant pathosystems in general and the potential for
yield losses in particular (Chakraborty et al. 2000; Coakleyand Scherm 1996; Coakley et al. 1999; Runion 2003), westill have a long way to go before reliable impact assess-ments for future agricultural production become feasible.
This commentary highlights selected challenges that mustbe overcome before we can hope to quantify the impacts ofa changing climate on plant disease intensity and yield loss.The focus will be on issues related to assessing the impactsof climate change in the strict sense (i.e., greenhouse-gasinduced changes in global and regional temperature andprecipitation patterns); global-change elements in additionto climate (e.g., direct and indirect effects of CO2 and aero-sols, changes in land use, and issues related to invasivepests as a component of global change), which will not bediscussed here, will undoubtedly add further layers of com-plexity to the problem of estimating potential long-term im-pacts. The challenges outlined here and elsewhere(Chakraborty et al. 2000; Coakley et al. 1999; Scherm andCoakley 2003) are not insurmountable, but increased inter-est and collaboration within the plant pathological commu-nity will be needed to overcome them. A concerted andsustained approach is currently lacking.
The search for climate-change fingerprints
For species for which long-term monitoring data areavailable, an empirical approach known as fingerprint anal-ysis has been used to document the species’ response to re-cent changes in climate and to extrapolate the potentialeffects of continued global warming on its abundance,phenology, and (or) geographical distribution. The methodis based on the observation that temperatures in many partsof the world have increased notably during the past quartercentury; for example, annual Northern Hemisphere meansurface temperature was 0.56 °C warmer during the 1990sthan during the 1970s. Given the magnitude and rapid rateof the recent climate warming, it is not surprising that an in-creasing number of fingerprint studies, including severalmeta-analyses, have found significant effects (Hughes 2000;Parmesan and Yohe 2003; Root et al. 2003; Walther et al.2002). These analyses focused on macroscopic species(e.g., birds, butterflies, or plants) that are readily observedand identified and for which population data have been col-lected by scientists and (or) amateur observers over periodsspanning several decades. In higher plants, for example, onestudy revealed that the first flowering date across 385 spe-cies in south-central England advanced by 4.5 days on aver-age during the 1990s compared with the period 1954–1990,and that variations in flowering date among years were sig-nificantly correlated with temperature during the precedingwinter months (Fitter and Fitter 2002). Also in Europe, asurvey of 35 nonmigratory butterfly species revealed thatthe geographical ranges of >60% of the species have shiftednorthward by 35 to 240 km during the 20th century (Parme-san et al. 1999). On a broader geographical and taxonomi-cal scale, a meta-analysis based on long-term records of 61species of invertebrates, amphibians, birds, and plants re-vealed a significant average advance in spring phenology of5.1 days per decade during the latter half of the 20th cen-tury (Root et al. 2003). The results of these and similar fin-gerprint studies clearly show that numerous species in
268 Can. J. Plant Pathol. Vol. 26, 2004

unmanaged ecosystems are responding strongly in the on-going “natural experiment” on climate warming, in a direc-tion consistent with expectations based on basic principlesof physiology and biogeography. However, to what degreethese changes in life cycle timing and geographical distribu-tion will impact ecosystem structure and function is still un-clear.
For reasons discussed in more detail below, pest speciesin agricultural (managed) ecosystems have received very lit-tle attention in fingerprint studies of climate change (Schermand Coakley 2003). A notable exception is an analysis ofhistorical flight phenology records of aphids in Britain,which revealed an advance in the timing of spring migrationby 3 to 6 days as regional temperatures have increased by0.4 °C during the past 25 years (Fleming and Tatchell1995). The earlier arrival of aphids in the spring has obvi-ous implications for the epidemiology and intensity ofaphid-vectored virus diseases in economically importantcrops such as barley and sugar beet (Harrington 2002a,2002b). When a similar analysis was carried out utilizinghistorical records of the time of first appearance of wheatstem rust, caused by Puccinia graminis Pers.:Pers. f. sp.tritici Eriks. & E. Henn., in the wheat production areas ofthe US Great Plains, no significant difference in arrival datewas observed between globally cool (1968–1977) and warm(1993–2002) periods (Scherm and Coakley 2003), indicat-ing the absence of a long-term climate-change fingerprint.However, the same data set showed a significant climate-related signal at shorter time scales, as evidenced by thesignificant association between phases of the ENSO andtime of disease onset at temporal scales typical of theENSO (2–10 years) (Scherm and Yang 1995). For wheatstripe rust caused by Puccinia striiformis Westend. f. sp.tritici Eriks. in the US Pacific Northwest, disease severitywas lower in El Niño years than in non-El Niño years(Scherm and Yang 1995). Since continued global warmingmay alter the frequency and (or) intensity of ENSO events(Fedorov and Philander 2000; Timmermann et al. 1999), abetter understanding of the associations between diseaseand ENSO (and other interannual climate cycles) may leadto improved predictability of plant disease epidemics underclimate change (Scherm and Yang 1998).
Overall, however, the potential for using retrospectivefingerprint studies to assess the impacts of climate changeon plant pathogens is limited. This is primarily becauselong-term series containing population or disease variablescollected in a standardized manner are unavailable for mostpathogens. In cases where such long-term records do exist,trends are often confounded by changes in biological ormanagement factors that have occurred over time. For ex-ample, long-term trends of wheat stem rust in the US GreatPlains were affected greatly by the barberry eradicationcampaign that occurred primarily between the 1920s andthe 1950s (Roelfs 1982). For potato late blight, for whichlong-term disease records are available from Europe, trendsare confounded by the introduction of more aggressivestrains of the A2 mating type of Phytophthora infestans(Mont.) de Bary, presumably during the late 1970s (Fry etal. 1993). Finally, plant pathogens are well known to bemore strongly influenced by localized meteorological vari-ables (primarily moisture-related) than the macroscopic spe-
cies for which most of the fingerprint studies have beenreported. Trends in the 20th century for such moisture-related variables have been considerably more ambiguousthan those for temperature (a variable that is spatially andtemporally much more continuous).
Modeling to assess the impacts of climatechange
Given the limited availability of historical records onplant diseases suitable for fingerprint analysis, it is not sur-prising that plant pathologists have relied largely on the useof mathematical or statistical models for impact assessment.In model-based assessments, climate-change scenarios (e.g.,simple temperature changes in fixed increments or more de-tailed projections based on climate-change models) are usedas inputs for a disease or yield-loss model for thepathosystem of interest, and model runs with these scenar-ios are then compared with control runs under current cli-mate. Several different modeling approaches have beenused within this framework, including climate matching,simple or multiple regression models, population models,and process-based simulation models (reviewed in Coakleyet al. 1999). Each of these modeling approaches has its sup-porters and detractors (Harrington 2002b; Pearson andDawson 2003; Samways et al. 1999; Sutherst 2003;Sutherst et al. 2000; Teng et al. 1996; Thuiller 2003), but itcan be argued that the availability of relevant input parame-ters and the intelligent application of the model are moreimportant than model structure or complexity per se. In-deed, regardless of model type, all model-based impact as-sessments in plant pathology face similar challenges thatlimit their productive application to problems related to cli-mate change. The following discussion will highlight threeof these challenges: uncertainty in key inputs, the difficultyin predicting biological responses in the presence ofnonlinearities and thresholds, and the high likelihood of ge-netic adaptation to climate change.
Uncertainty in model inputsAlthough physically-based climate models have become
more refined, as discussed above, their projections of thefuture climate still contain relatively large uncertainties(Kerr 2001). For example, the suite of emission scenariosand climate models used in the most recent simulations bythe Intergovernmental Panel on Climate Change (Houghtonet al. 2001) produced increases in global temperature rang-ing from 1.4 to 5.8 °C for the period 1990–2100; potentialchanges in precipitation patterns were even less certain interms of their magnitude and direction. When uncertainclimate-change projections are used as inputs in models forimpact assessment, such as disease or yield-loss models, theoutputs generated by these models will also be uncertainbecause of error perpetuation and (or) propagation. There-fore, it is not sufficient to conduct impact assessments usinga single-point estimate of climate change as input; instead,the simulations must be carried out over the full range ofpossible input scenarios to capture the range of possibleoutcomes and their associated probabilities. Methodologyfor incorporating uncertainty into impact models is avail-able in the form of Monte-Carlo simulation or fuzzy arith-
Scherm: climate change / predicting impacts on plant pathology 269
metic, among other approaches (Ferson et al. 1997). Ofthese methods, fuzzy arithmetic is particularly attractive foruse in climate-change applications (Scherm 2000), giventhe fact that very little information is available about proba-bility distributions of climate-change variables and their in-terdependencies; a best estimate of future values of theclimate-change variable and its range (highest and lowestpossible values) is all that can be assumed with some confi-dence.
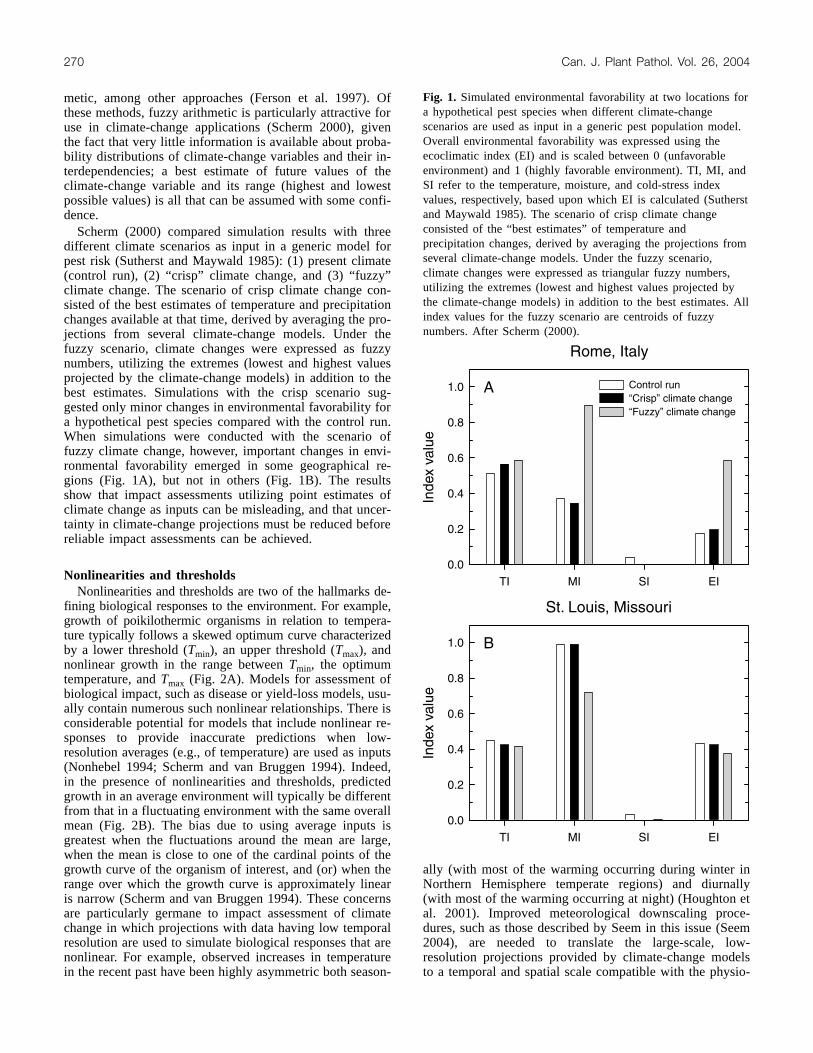
Scherm (2000) compared simulation results with threedifferent climate scenarios as input in a generic model forpest risk (Sutherst and Maywald 1985): (1) present climate(control run), (2) “crisp” climate change, and (3) “fuzzy”climate change. The scenario of crisp climate change con-sisted of the best estimates of temperature and precipitationchanges available at that time, derived by averaging the pro-jections from several climate-change models. Under thefuzzy scenario, climate changes were expressed as fuzzynumbers, utilizing the extremes (lowest and highest valuesprojected by the climate-change models) in addition to thebest estimates. Simulations with the crisp scenario sug-gested only minor changes in environmental favorability fora hypothetical pest species compared with the control run.When simulations were conducted with the scenario offuzzy climate change, however, important changes in envi-ronmental favorability emerged in some geographical re-gions (Fig. 1A), but not in others (Fig. 1B). The resultsshow that impact assessments utilizing point estimates ofclimate change as inputs can be misleading, and that uncer-tainty in climate-change projections must be reduced beforereliable impact assessments can be achieved.
Nonlinearities and thresholdsNonlinearities and thresholds are two of the hallmarks de-
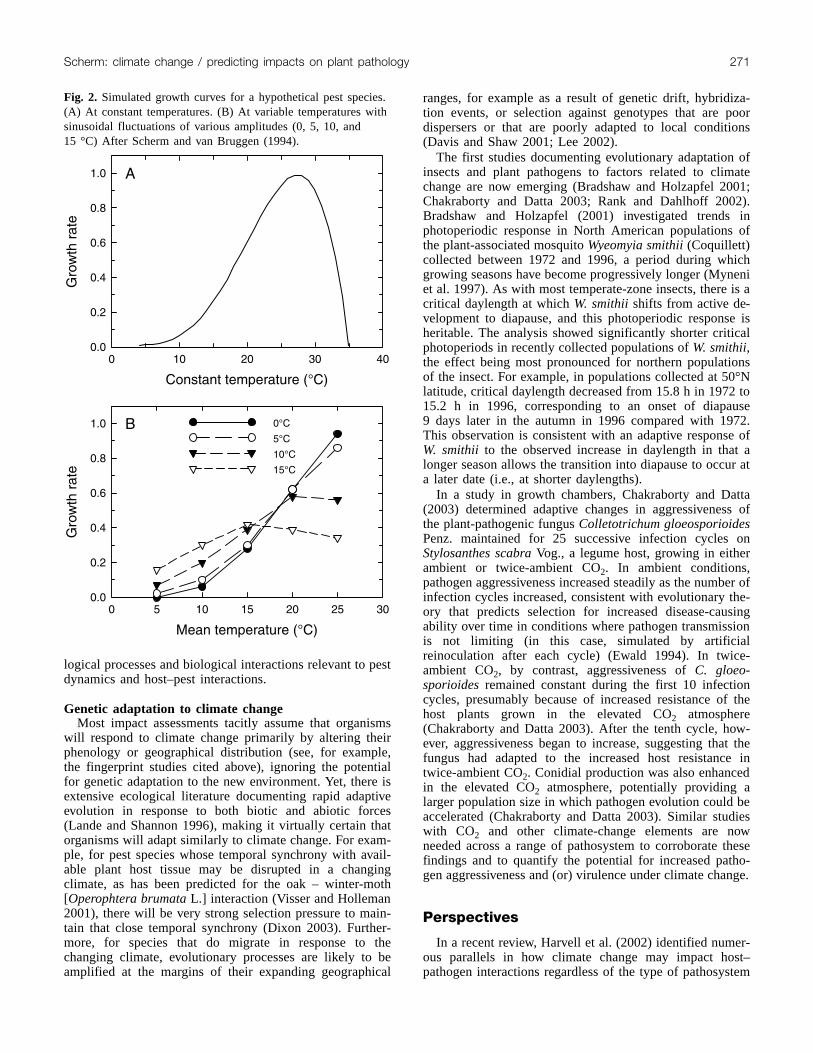
fining biological responses to the environment. For example,growth of poikilothermic organisms in relation to tempera-ture typically follows a skewed optimum curve characterizedby a lower threshold (Tmin), an upper threshold (Tmax), andnonlinear growth in the range between Tmin, the optimumtemperature, and Tmax (Fig. 2A). Models for assessment ofbiological impact, such as disease or yield-loss models, usu-ally contain numerous such nonlinear relationships. There isconsiderable potential for models that include nonlinear re-sponses to provide inaccurate predictions when low-resolution averages (e.g., of temperature) are used as inputs(Nonhebel 1994; Scherm and van Bruggen 1994). Indeed,in the presence of nonlinearities and thresholds, predictedgrowth in an average environment will typically be differentfrom that in a fluctuating environment with the same overallmean (Fig. 2B). The bias due to using average inputs isgreatest when the fluctuations around the mean are large,when the mean is close to one of the cardinal points of thegrowth curve of the organism of interest, and (or) when therange over which the growth curve is approximately linearis narrow (Scherm and van Bruggen 1994). These concernsare particularly germane to impact assessment of climatechange in which projections with data having low temporalresolution are used to simulate biological responses that arenonlinear. For example, observed increases in temperaturein the recent past have been highly asymmetric both season-
ally (with most of the warming occurring during winter inNorthern Hemisphere temperate regions) and diurnally(with most of the warming occurring at night) (Houghton etal. 2001). Improved meteorological downscaling proce-dures, such as those described by Seem in this issue (Seem2004), are needed to translate the large-scale, low-resolution projections provided by climate-change modelsto a temporal and spatial scale compatible with the physio-
270 Can. J. Plant Pathol. Vol. 26, 2004
Fig. 1. Simulated environmental favorability at two locations fora hypothetical pest species when different climate-changescenarios are used as input in a generic pest population model.Overall environmental favorability was expressed using theecoclimatic index (EI) and is scaled between 0 (unfavorableenvironment) and 1 (highly favorable environment). TI, MI, andSI refer to the temperature, moisture, and cold-stress indexvalues, respectively, based upon which EI is calculated (Sutherstand Maywald 1985). The scenario of crisp climate changeconsisted of the “best estimates” of temperature andprecipitation changes, derived by averaging the projections fromseveral climate-change models. Under the fuzzy scenario,climate changes were expressed as triangular fuzzy numbers,utilizing the extremes (lowest and highest values projected bythe climate-change models) in addition to the best estimates. Allindex values for the fuzzy scenario are centroids of fuzzynumbers. After Scherm (2000).
logical processes and biological interactions relevant to pestdynamics and host–pest interactions.
Genetic adaptation to climate changeMost impact assessments tacitly assume that organisms
will respond to climate change primarily by altering theirphenology or geographical distribution (see, for example,the fingerprint studies cited above), ignoring the potentialfor genetic adaptation to the new environment. Yet, there isextensive ecological literature documenting rapid adaptiveevolution in response to both biotic and abiotic forces(Lande and Shannon 1996), making it virtually certain thatorganisms will adapt similarly to climate change. For exam-ple, for pest species whose temporal synchrony with avail-able plant host tissue may be disrupted in a changingclimate, as has been predicted for the oak – winter-moth[Operophtera brumata L.] interaction (Visser and Holleman2001), there will be very strong selection pressure to main-tain that close temporal synchrony (Dixon 2003). Further-more, for species that do migrate in response to thechanging climate, evolutionary processes are likely to beamplified at the margins of their expanding geographical
ranges, for example as a result of genetic drift, hybridiza-tion events, or selection against genotypes that are poordispersers or that are poorly adapted to local conditions(Davis and Shaw 2001; Lee 2002).
The first studies documenting evolutionary adaptation ofinsects and plant pathogens to factors related to climatechange are now emerging (Bradshaw and Holzapfel 2001;Chakraborty and Datta 2003; Rank and Dahlhoff 2002).Bradshaw and Holzapfel (2001) investigated trends inphotoperiodic response in North American populations ofthe plant-associated mosquito Wyeomyia smithii (Coquillett)collected between 1972 and 1996, a period during whichgrowing seasons have become progressively longer (Myneniet al. 1997). As with most temperate-zone insects, there is acritical daylength at which W. smithii shifts from active de-velopment to diapause, and this photoperiodic response isheritable. The analysis showed significantly shorter criticalphotoperiods in recently collected populations of W. smithii,the effect being most pronounced for northern populationsof the insect. For example, in populations collected at 50°Nlatitude, critical daylength decreased from 15.8 h in 1972 to15.2 h in 1996, corresponding to an onset of diapause9 days later in the autumn in 1996 compared with 1972.This observation is consistent with an adaptive response ofW. smithii to the observed increase in daylength in that alonger season allows the transition into diapause to occur ata later date (i.e., at shorter daylengths).
In a study in growth chambers, Chakraborty and Datta(2003) determined adaptive changes in aggressiveness ofthe plant-pathogenic fungus Colletotrichum gloeosporioidesPenz. maintained for 25 successive infection cycles onStylosanthes scabra Vog., a legume host, growing in eitherambient or twice-ambient CO2. In ambient conditions,pathogen aggressiveness increased steadily as the number ofinfection cycles increased, consistent with evolutionary the-ory that predicts selection for increased disease-causingability over time in conditions where pathogen transmissionis not limiting (in this case, simulated by artificialreinoculation after each cycle) (Ewald 1994). In twice-ambient CO2, by contrast, aggressiveness of C. gloeo-sporioides remained constant during the first 10 infectioncycles, presumably because of increased resistance of thehost plants grown in the elevated CO2 atmosphere(Chakraborty and Datta 2003). After the tenth cycle, how-ever, aggressiveness began to increase, suggesting that thefungus had adapted to the increased host resistance intwice-ambient CO2. Conidial production was also enhancedin the elevated CO2 atmosphere, potentially providing alarger population size in which pathogen evolution could beaccelerated (Chakraborty and Datta 2003). Similar studieswith CO2 and other climate-change elements are nowneeded across a range of pathosystem to corroborate thesefindings and to quantify the potential for increased patho-gen aggressiveness and (or) virulence under climate change.
Perspectives
In a recent review, Harvell et al. (2002) identified numer-ous parallels in how climate change may impact host–pathogen interactions regardless of the type of pathosystem
Scherm: climate change / predicting impacts on plant pathology 271
Fig. 2. Simulated growth curves for a hypothetical pest species.(A) At constant temperatures. (B) At variable temperatures withsinusoidal fluctuations of various amplitudes (0, 5, 10, and15 °C) After Scherm and van Bruggen (1994).

(plant or animal, terrestrial or marine). They identified fourresearch priorities to improve the ability to predict impactson disease across a wide range of taxa: (1) collect baselinedata on diseases of natural populations, (2) separate the ef-fects of multiple climate variables on disease, (3) forecastepidemics, and (4) evaluate the role of evolution. From aplant pathologist’s view, the first research area listed byHarvell et al. (disease data collection in unmanaged sys-tems) is especially important, given our preoccupation withpathogens important to production agriculture, despite thepotentially greater impact of climate change in natural com-pared with managed systems. Indeed, without long-termdata on disease dynamics in natural systems, it will be im-possible to document fingerprints related to climate changesfor plant pathogens, given the confounding factors presentin managed systems as discussed above.
By contrast, plant pathologists have been actively pursu-ing research in the second (interactions among multiple cli-mate variables) and third areas (disease prediction) formany years. Indeed, this has been the mainstay of researchrelated to climate change in plant pathology during the pasttwo decades (Chakraborty et al. 2000; Coakley 1995;Coakley and Scherm 1996; Coakley et al. 1999). Nonethe-less, our ability to generalize and predict based on suchstudies is still rather limited; experimentally, this is due topersistent difficulties in extrapolating results from growthchambers or field plots to regional and global scales,whereas for modeling studies, predictive ability is limitedby the various technical challenges discussed above. In myview, therefore, developing solutions to the difficulties asso-ciated with scaling and extrapolation of experimental andmodeling results is more important than the development ofnew models or the execution of additional experiments ingrowth chambers with multiple interacting factors.
The fourth priority item on the list of Harvell et al.(2002), pathogen evolution in response to climate change,has largely been ignored in plant pathology. Yet, more thananything else, processes such as selection against poorlyadapted genotypes, genetic drift, and hybridization amongallopatric species as they alter their geographical distribu-tion in response to climate change have the potential to in-validate experimental or model-based predictions ofclimate-change impacts. Research to understand and antici-pate such evolutionary processes will be among the mostfruitful areas in the interface between climate change andplant pathology in the near future.
References
Adams, R.M., Rosenzweig, C., Peart, R.M., Ritchie, J.T., McCarl,B.A., Glyer, J.D., Curry, R.B., Jones, J.W., Boote, K.J., and Al-len, L.H., Jr. 1990. Global climate change and U.S. agriculture.Nature (London), 345: 219–224.
Bradshaw, W.E., and Holzapfel, C.M. 2001. Genetic shift inphotoperiodic response correlated with global warming. Proc.Natl. Acad. Sci. USA, 98: 14 509 – 14 511.
Chakraborty, S., and Datta, S. 2003. How will plant pathogensadapt to host resistance at elevated CO2 under a changing cli-mate? New Phytol. 159: 733–742.
Chakraborty, S., von Tiedemann, A., and Teng, P.S. 2000. Climatechange: potential impact on plant diseases. Environ. Pollut.108: 317–326.
Coakley, S.M. 1995. Biospheric change: will it matter in plant pa-thology? Can. J. Plant Pathol. 17: 147–153.
Coakley, S.M., and Scherm, H. 1996. Plant disease in a changingglobal environment. Asp. Appl. Biol. 45: 227–238.
Coakley, S.M., Scherm, H., and Chakraborty, S. 1999. Climatechange and plant disease management. Annu. Rev. Phytopathol.37: 399–426.
Davis, M.B., and Shaw, R.G. 2001. Range shifts and adaptive re-sponses to Quarternary climate change. Science (Washington,D.C.), 292: 673–679.
Dixon, A.F.G. 2003. Climate change and phenologicalasynchrony. Ecol. Entomol. 28: 380–381.
Easterling, D.R., Meehl, G.A., Parmesan, C., Changnon, S.A.,Karl, T.R., and Mearns, L.O. 2000. Climate extremes: observa-tions, modeling, and impacts. Science (Washington, D.C.), 289:2068–2074.
Easterling, W.E., III, Crosson, P.R., Rosenberg, N.J., McKenney,M.S., Katz, L.A., and Lemon, K.M. 1993. Agricultural impactsof and responses to climate change in the Missouri–Iowa–Nebraska–Kansas (MINK) region. Clim. Change, 24: 23–62.
Ewald, P.W. 1994. Evolution of infectious disease. Oxford Univer-sity Press, Oxford.
Fedorov, A.V., and Philander, S.G. 2000. Is El Niño changing?Science (Washington, D.C.), 288: 1997–2002.
Ferson, S., Cooper, J.A., Moore, D.R.J., and Lee, R.C. 1997. Be-yond point estimates: risk assessment using interval, fuzzy andprobabilistic arithmetic. Applied Biomathematics, Setauket,N.Y.
Fitter, A.H., and Fitter, R.S.R. 2002. Rapid changes in floweringtime in British plants. Science (Washington, D.C.), 296: 1689–1691.
Fleming, R.A., and Tatchell, G.M. 1995. Shifts in the flight peri-ods of British aphids: a response to climate warming? In In-sects in a changing environment. Edited by R. Harrington andN.E. Stork. Academic Press Ltd., London. pp. 505–508.
Fry, W.E., Goodwin, S.B., Dyer, A.T., Matuszak, J.M., Drenth, A.,Tooley, P.W., Sujkowski, L.S., Koh, Y.J., Cohen, B.A., andSpielman, L.J. 1993. Historical and recent migrations of Phy-tophthora infestans: chronology, pathways, and implications.Plant Dis. 77: 653–661.
Harrington, R. 2002a. BYDV: the heat is on. In Barley yellowdwarf disease: recent advances and future strategies. Edited byM. Henry and A. McNab. Centro Internacional de Mejoramientode Maiz y Trigo (CIMMYT) [International Maize and WheatImprovement Center], Mexico, D.F. pp. 34–39.
Harrington, R. 2002b. Insect pests and global environmentalchange. In Encyclopedia of global environmental change.Vol. 3. Edited by I. Douglas. Wiley, Chichester. pp. 381–386.
Harvell, C.D., Mitchell, C.E., Ward, J.R., Altizer, S., Dobson,A.P., Ostfeld, R.S., and Samuel, M.D. 2002. Climate warmingand disease risks for terrestrial and marine biota. Science(Washington, D.C.), 296: 2158–2162.
Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., van der Lin-den, P.J., Dai, X., Maskell, K., and Johnson, C.A. (Editors).2001. Climate change 2001: the scientific basis. CambridgeUniversity Press, Cambridge.
Hughes, L. 2000. Biological consequences of global warming: isthe signal already apparent? Trends Ecol. Evol. 15: 56–61.
Kerr, R.A. 2001. Rising global temperature, rising uncertainty.Science (Washington, D.C.), 292: 192–194.
Lande, R., and Shannon, S. 1996. The role of genetic variation inadaptation and population persistence in a changing environ-ment. Evolution, 50: 434–437.
272 Can. J. Plant Pathol. Vol. 26, 2004

Scherm: climate change / predicting impacts on plant pathology 273
Lee, C.E. 2002. Evolutionary genetics of invasive species. TrendsEcol. Evol. 17: 386–391.
Myneni, R.B., Keeling, C.D., Tucker, C.J., Asrar, G., and Nemani,R.R. 1997. Increased plant growth in the northern high latitudesfrom 1981 to 1991. Nature (London), 386: 698–701.
Nonhebel, S. 1994. The effects of use of average instead of dailyweather data in crop growth simulation models. Agric. Syst.44: 377–396.
Oerke, E.C., Dehne, H.W., Schönbeck, F., and Weber, A. 1994.Crop production and crop protection: estimated losses in majorfood and cash crops. Elsevier, Amsterdam.
Parmesan, C., and Yohe, G. 2003. A globally coherent fingerprintof climate change impacts across natural systems. Nature (Lon-don), 421: 37–42.
Parmesan, C., Ryrholm, N., Stefanescu, C., Hill, J.K., Thomas,C.D., Descimon, H., Huntley, B., Kalla, L., Kullberg, J.,Tammaru, T., Tennent, W.J., and Thomas, J.A. 1999. Polewardshifts in geographical ranges of butterfly species associatedwith regional warming. Nature (London), 399: 579–583.
Parry, M. 1992. The potential effects of climate changes on agri-culture and land use. Adv. Ecol. Res. 22: 63–91.
Parry, M.L., Porter, J.H., and Carter, T.R. 1990. Agriculture: cli-matic change and its implications. Trends Ecol. Evol. 5: 318–322.
Pearson, R.G., and Dawson, T.P. 2003. Predicting the impacts ofclimate change on the distribution of species: are bioclimate en-velope models useful? Glob. Ecol. Biogeogr. 12: 361–371.
Rank, N.E., and Dahlhoff, E.P. 2002. Allele frequency shifts in re-sponse to climate change and physiological consequences ofallozyme variation in a montane insect. Evolution, 56: 2278–2289.
Reilly, J., Tubiello, F., McCarl, B., Abler, D., Darwin, R., Fuglie,K., Hollinger, S., Izaurralde, C., Jagtap, S., Jones, J.,Mearns, L., Ojima, D., Paul, E., Paustian, K., Riha, S.,Rosenberg, N., and Rosenzweig, C. 2003. U.S. agriculture andclimate change: new results. Clim. Change, 57: 43–69.
Roelfs, A.P. 1982. Effects of barberry eradication on stem rust inthe United States. Plant Dis. 66: 177–181.
Root, T.L., Price, J.T., Hall, K.R., Schneider, S.H., Rosenzweig,C., and Pounds, J.A. 2003. Fingerprints of global warming onwild animals and plants. Nature (London), 421: 57–60.
Runion, G.B. 2003. Climate change and plant pathosystems — fu-ture disease prevention starts here. New Phytol. 159: 531–538.
Samways, M.J., Osborn, R., Hastings, H., and Hattingh, V. 1999.Global climate change and accuracy of prediction of species’geographical ranges: establishment success of introduced lady-birds (Coccinellidae, Chilocorus spp.) worldwide. J. Biogeogr.26: 795–812.
Scherm, H. 2000. Simulating uncertainty in climate–pest modelswith fuzzy numbers. Environ. Pollut. 108: 373–379.
Scherm, H., and Coakley, S.M. 2003. Plant pathogens in a chang-ing world. Australas. Plant Pathol. 32: 157–165.
Scherm, H., and van Bruggen, A.H.C. 1994. Global change andnonlinear growth: how important are changes in average tem-perature? Phytopathology, 84: 1380–1384.
Scherm, H., and Yang, X.B. 1995. Interannual variations in wheatrust development in China and the United States in relation tothe El Niño/Southern Oscillation. Phytopathology, 85: 970–976.
Scherm, H., and Yang, X.B. 1998. Atmospheric teleconnectionpatterns associated with wheat stripe rust disease in NorthChina. Int. J. Biometeorol. 42: 28–33.
Seem, R.C. 2004. Forecasting plant disease in a changing climate:a question of scale. Can. J. Plant Pathol. 26: 274–283.
Stenseth, N.C., Mysterud, A., Ottersen, G., Hurrell, J.W., Chan,K.-S., and Lima, M. 2002. Ecological effects of climate fluctu-ations. Science (Washington, D.C.), 297: 1292–1296.
Sutherst, R.W. 2003. Prediction of species geographical ranges. J.Biogeogr. 30: 805–816.
Sutherst, R.W., and Maywald, G.F. 1985. A computerised systemfor matching climates in ecology. Agric. Ecosyst. Environ. 13:281–299.
Sutherst, R.W., Maywald, G.F., and Russell, B.L. 2000. Esti-mating vulnerability under global change: modular modellingof pests. Agric. Ecosyst. Environ. 82: 303–319.
Teng, P.S., Heong, K.L., Kropff, M.J., Nutter, F.W., and Sutherst,R.W. 1996. Linked pest-crop models under global change. InGlobal change and terrestrial ecosystems. Edited by B. Walkerand W. Steffen. Cambridge University Press, Cambridge.pp. 291–316.
Thuiller, W. 2003. BIOMOD — optimizing predictions of speciesdistributions and projecting potential future shifts under globalchange. Global Change Biol. 9: 1353–1362.
Timmermann, A., Oberhuber, J., Bacher, A., Esch, M., Latif, M.,and Roeckner, E. 1999. Increased El Niño frequency in a cli-mate model forced by future greenhouse warming. Nature(London), 398: 694–697.
Visser, M.E., and Holleman, L.J.M. 2001. Warmer springs disruptthe synchrony of oak and winter moth phenology. Proc. R. Soc.Lond. Ser. B, Biol. Sci. 268: 289–294.
Walther, G.-R., Post, E., Convey, P., Menzel, A., Parmesan, C.,Beebee, T.J.C., Fromentin, J.-M., Hoegh-Guldberg, O., andBairlein, F. 2002. Ecological responses to recent climatechange. Nature (London), 416: 389–395.




























