Embed Size (px)
Citation preview

European Journal of Clinical Investigation (1980) 1 0 , 4 1-46
Circulating adrenaline and blood pressure: the metabolic effects and kinetics of infused adrenaline in man
G. A. FITZGERALD, P. BARNES, C. A. HAMILTON & C. T. DOLLERY, Department of Clinical Pharmacology, Royal Postgraduate Medical School, Hammersmith Hospital, London
Received 16 March 1979 and in revised form 13 May 1980
Abstract. Six normotensive volunteers were infused with L-adrenaline at 0.01, 0.03, 0.05, 0.075 and 0.10 pg/kg-l min-I, each increment lasted 10 min. Plasma adrenaline rose from 0.27 to 4.61 nmol/l, and there were dose-related increases in plasma renin activity, blood glucose, plasma cyclic AMP and plasma free fatty acids, but not in plasma noradrenaline and cyclic GMP. Levels of circulating adrenaline previously noted in essential hypertensives had minimal cardio- vascular effects. The secretion rate of adrenaline and its rate of clearance from the circulation were calcu- lated from plasma samples taken during an hour-long infusion (0.083 & 0.006 pg kg-' min-I) of L-adrenaline in the same individuals. The secretion rate ranged from 1.40 to 6.01 nmol/min with a mean (+SEM, 6) of 2.82 +O-76 nmol/rnin. Mean clearance ( 2 SEM, 6) was 9.41 k 1.37 I/min and ranged from 4.86 to 14.61 I/min. The decline of plasma adrenaline following the infu- sion was biexponential.
Plasma adrenaline is unlikely to be of primary im- portance in the elevation of blood pressure, either directly, via renin release or by noradrenaline release via presynaptic beta receptors. However, variation in clearance between subjects limits the use of plasma levels as an interindividual index of adrenal release of adrenaline. The relationship between sympatho- adrenal activity and plasma adrenaline may be further perturbed by equilibration between the circulation and sites of tissue uptake. The lower levels of plasma adrenaline than of noradrenaline appear to result from both a slower rate of secretion and a higher rate of clearance from the circulation.
Key words. Adrenaline, blood pressure, kinetics.
Introduction The development of a specific, sensitive, radioenzymic assay has enabled the measurement of picomolar
Cormpondence: Dr G . A. FitzGerald, Department of Medicine and Pharmacology, Vanderbilt University, Nashville, Tennessee 37232. U S A .
a 1980 Blackwell Scientific Publications 00 lc2972/80/ IOOO-040 lf02.00
quantities of adrenaline in plasma [ I ] . Recently, Fran- co-Morselli and co-workers [2] demonstrated a signifi- cant elevation of plasma adrenaline in a group of essential hypertensives compared with nonnotensive controls. The difference persisted when the hyperten- sives were subdivided into 'labile' and 'sustained' hypertensives and a positive correlation between supine basal plasma adrenaline and blood pressure was noted for the group as a whole. Plasma noradrena- line, by contrast with previous work [3], was not higher in the hypertensive group. The increased levels of cir- culating adrenaline might exert a primary pressor effect by stimulating post-synaptic alpha or beta adrenoreceptors causing peripheral vasoconstriction or an increase in cardiac output. Alternatively, adrena- line might act indirectly by stimulating renin release [4] or by modifying adrenergic neuronal release of nora- drenaline via presynaptic receptor stimulation [5-71. The purpose of this study was to determine whether the plasma concentrations noted in hypertensives might result in an elevation of blood pressure by these mechanisms.
The use of the concentration of plasma noradrena- line as an index of sympathetic nerve activity between individuals is limited by wide differences in the rate of clearance from the circulation (81. An additional objec- tive was to establish whether similar limitations apply to plasma adrenaline as an index of sympathoadrenal activity in man.
Methods
Subjects Six normotensive male Caucasian volunteers were
infused with ~-adrenaline. Their mean age (_tSEM) was 29.0+ 1.2 years and their mean weight ( 2 SEM) 78.6 f 5.0 kg. The subjects abstained from all medica- tion for 2 weeks and from food and drink for 12 h prior to the studies. All were non-smokers and had no de- tectable abnormality on clinical or laboratory investi- gation. Blood pressure was measured indirectly every minute in the right arm (Artenosonde 1217, Roche), heart rate was monitored continuously (Grass poly-
401

402 G. A. FITZGERALD P I al.
graph 7D) and the subjects remained supine through- out both studies. The protocol was approved by the Research EthicsCommittee of the Royal Postgraduate Medical School and Hammersmith Hospital.
First s ~ u d ) ? lntravenous cannulae (Venicath- 16. Abbott) were
inserted for infusing and sampling in the right and left brachial veins respectively. Following a 30 rnin control period, L-adrenaline was infused continuously at 0.02, 0.03. 0.05. 0.075 and 0.10 pg kg- ' min-I. These infu- sion rates correspond to mean ( + S E M ) rates of 4.65 0.3, 13.92 0.9. 23.2 1.46. 34.84 +_ 2.2 and 46.39 2 2.9 nmol/min. Each dosage increment lasted 10 min and blood samples were drawn in the ninth minute for the estimation of adrenaline. noradrenaline, blood glucose and plasma renin activity. Plasma free fatty acids were measured in five and cyclic nucleotides in three individuals. All samples were transferred imme- diately to cooled tubes and kept stored in ice prior to centrifugation at 4 C (Sigma 2KD) and storage of plasma at - 20°C. All assays were performed within 2 weeks of sampling.
Second srud.v This took place within 6-12 days of the first study. A
rate of L-adrenaline infusion was selected from the preceding study that would elevate systolic pressure by 15-20 mmHg. Intravenous cannulae were inserted as before and following a 30 min control period, L-adrenaline was infused for 60 min at a mean rate (+SEM, 6 ) of 0.083 C0.06 pg kg-' min-' (38.49k2.3 nmol/min). The pump rate was fixed at 0.44 ml/min. Blood was sampled for adrenaline and noradrenaline at 6 min before, and 1.8.3.75, 7.5, 10, 15,20, 30,45,55, 60. 61-8, 63.75. 67.5, 70, 75 and 90 min after the infusions. which were commenced at 09.00 hours.
Chemical analyses Adrenaline and noradrenaline were measured by a
modification of the method of Da Prada & Zurcher [l]. Blood was drawn into cooled tubes, containing lithium heparin, prior to centrifugation and aspiration of plasma. Plasma (0.21 ml) was incubated at 37°C with 0.1 rnl of 2 mol/l Tris/HCI buffer pH 8.6 containing 1 mg/ml dithiothriotol, 0.1 mol/l MgClz and 8 mol/l of [3H]S-adenosyl-methionine (Sigma). After 1 h the reac- tion was stopped by adding 0.05 ml of 1 mol/l borate buffer pH 8.0 containing 1% sodium tetraphenylbor- ate, followed by 0.05 ml of a 5 nmol/l solution of normetanephrine and metanephrine. The metaneph- rines were extracted into 2 ml of ether, then back- extracted into 75 p1 of 0.01 mol/l HCI. Ethanol (25 pl) was added to the acid and the whole 0.1 ml spotted onto preadsorbant t.1.c. plates (Whatman LK 5DF). The plates were developed in a chlorofonn/rneth- ano1/700/, ethylamine solution (16: 3 : 2, by vol.). The
plates were observed under U V light, the norme- tanephrine and mentanephrine bands marked and the spots scraped into tubes containing 1 ml of 0.05 molil
Finally a periodate oxidation was carried out to convert the metanephrine to vanillin by adding 50 P I of a 3% solution of sodium periodate. After 5 min 1 ml of 0.1 mol/l HCI and 10 ml of toluene were added. The vanillic acid was then extracted into the toluene and the two phases separated. The tritium present in the toluene phase was counted in the presence of 0.4 ml of permaflor. The inorganic and organic phases were separated by freezing the lower aqueous phase in the acetoneisolid carbon dioxide mixture for all extrac- tions. The intra-assay coefficient of variation was 5':(, and the interassay coefficient of variation 7'x.
Plasma glucose was measured by the glucose oxidase method 191, plasma cyclic AMP by using a high affinity. high specificity binding protein (Amersham Kit TRK 432). and plasma cyclic GMP and plasma renin activity by radioimmunoassay (Amersham Kit TRK 500, CEA Sorin Kit respectively). Plasma fatty acids were measured by the method of Dole [ 101.
NHAOH.
Kinr I ic a n d s ICI t is I iccil mi(i1j.s is
Concentrations of circulating adrenaline represent the resultant of two processes: the rate of entry of adrenaline into the circulation and the rate of itsclear- ance from it. Thus:an index of the rate of adrenaline release (AR, e.g. nmol/min) in the basal state may be derived by correcting an individual's basal plasma con- centration of adrenaline (APL. e.g. nmolil) for his clear- ance (CLA, e.g., limin):
AR=APL x CLA (Eq 1)
The applicability of the clearance measurement at high concentrations of adrenaline to the much lower concentration under basal conditions assumes that clearance is independent of the level of plasma adrena- line.
The rate of entry of endogenous adrenaline into the circulation cannot be measured directly. However, clearance (I/min) may be derived by infusing exo- genous adrenaline at a known rate (A infusion rate, e.g. ,umol/min) and measuring the increase in plasma concentration due to the infusion as the difference between plasma concentrations of adrenaline obtained at steady state during the infusion (Ass, e.g. nmol/l) and A ~ L thus:
A infusion rate Ass - APL
CLA = (Eq 2)
This derivation is independent of any particular phar- macokinetic model of adrenaline disposition. How- ever, it does assume that the rate of release of endo- genous adrenaline is not altered by raising the amine concentration in plasma during the infusion.
The turnover time (TA, e.g. min) has been defined

METABOLIC EFFECTS AND KINETICS O F ADRENALINE 403
[ I I ] as the time required for a complete replacement of circulating hormone by a fresh endogenous supply. This is given by the equation:
(Eq 3) (Ass - APL) VPL A infusion rate
T* =
Plasma volume ( VPL) wasestimated for each individual by a regression equation based on body weight [12]. This calculation presumes that adrenaline only has access to plasma volume. Such a hypothetical volume of distribution is employed for the purpose of inter- subject com par is0 ns.
Pairwise comparisons were analysed by a modifica- tion of the 1-test based on ranks [ 13) and correlations by Spearman's method. All mean results are expressed +SEM.
Results
First .stutli*
The mean preinfusion plasma adrenaline was 0,2720.03 (6) nmol/l and rose to 4.61 k0.57 nmol/l during the highest infusion rate. Systolic pressure rose from a mean of 112.1 20.8 mmHg to a mean of 132-3 24.0 mmH3 and heart rate from i1 mean of
65.1 2 2.9 beats/min to 80.3 2 3.3 beats/min. Diastolic pressure fell from a mean of 77.222.3 mmHg to 61.9 23.1 mmHg. The cardiovascular effects were tri- vial during the lowest rate of infusion which produced plasma concentrations of adrenaline similar to those noted by Franco-Morselli et a/ . [2] in essential hyper- tensives (Table I ) . The changes in systolic (r=0.93: P < 0.0 1 ) and diastolic ( r = - 0.92; P < 0.0 I ) pressure and in heart rate (r=0.94; P < O . O I ) correlated with increasing plasma adrenaline. A similar correlation was found with the increase in blood glucose (r=0.93; P<O.OI), plasma renin activity (r=0.93: P<O.Ol)and plasma free fatty acids (r=0.95; P<O.OI). Plasma cyclic GMP and plasma noradrenaline did not differ significantly (P>0.05) from basal values during the infusion (Table 2).
Second ,stud.)*
The mean preinfusion plasma adrenaline was 0.29 20.06 (6) nmol/l. Attainment of new steady states in blood pressure and heart rate required infusion for approximately 30 min. Systolic pressure rose from a mean preinfusion value of 113.9 k2.1 mmHg to 131.023.5 mmHg and heart rate from a mean of 68.8 2 3.6 beatsimin to 78.2 k3.4 beatslmin. Diastolic pressure fell from a mean of 76.4k3.3 mmHg to
Table I . Blood pressure. heart raw and circulaling adrenaline and noradrenaline during incremental infusions of L-adrenaline. Each increment lasted 10 min and blood was sampled in the ninth minute. Blood pressure and heart rille are expressed as changes from baseline values. Values are mean + SEM
(6).
Adrenaline infusion rate kg-' min-')
Basal 0.0 I 0.03 0.05 0,075 0.10
Plasma adrenaline (nmolil)
0.25 2 0.03 0.85 f 0.10 1.46 f 0.24 2.14+2,0 3.78 f 2.24 4.61 f0.57
Plasma noradrenaline (nmolil)
Systolic pressure (mmHg)
Diastolic Heart pressure rate (mmHg) (katsimin)
~~
0.96 f 0.3 1 I .82 2 0.28 I .67 f 0.45 I .50 f 0.26 I .79 f 0.40 I .32 f 0.80
~
I .95 f 0.15 4.83k 1.01 10.65 f 2.06 14.96f2.60 20.2 f 3.75
-4.552 1.3 5,0820.99 -7.02+ 1.3 8,432 1.24 - 12.42 2 1.3 9.78 f 1.39 - 13.03+ 1.6 11.97k 1.69 - 15.3f2.1 15.16f 1.73
Table 2. Mean values o f blood glucose. plasma renin activity, plasma free fatty acids, cyclic AMP and cyclic GMP during incremental infusions of L-adrenaline. Each increment lasted 10 min and blood was
sampled in the ninth minute. Values are mean f SEM.
Adrenaline infusion rate (pg kg-l min-I)
Basal 0.0 1 0.03 0-05 0-075 0.10
Blood glucose (mmol/l) (n=6)
4.24 f 0.36 4,1920.54 4.74 f 0.43 5.3 I f0.37 6.292 0.5 I 7.06 f0.77
Plasma renin activity (ng A1 ml-l h (n=6)
2.00 f 0.6 2.69 k0.5 3-8 f 0.9 6.96 f 1.5 8.97 f 1.8 11.21+3.5
Plasma free fatty acids
(n=5) - '1 ( P O I / l )
457.8 f 133.9 652.0 f 23 1.1
1 107.0 f 323.7 1280.6 f 360.3 1450.0 f 260. I 1555.4 * 291.1
Plasma CAMP
(n=3) (pmolil)
2 1.3 f 3.0 24.7 f 7.9 31.3f 8.4 35.3 f 8.5 43.7f7.1 48.7 f 9.0
Plasma cGMP
(n=3) (pmol/U
4.7 f 0.4 3.3 f0.6 3.4 f 0.4 3.9 f 0. I 4.3 0.9 4.4f 1.1
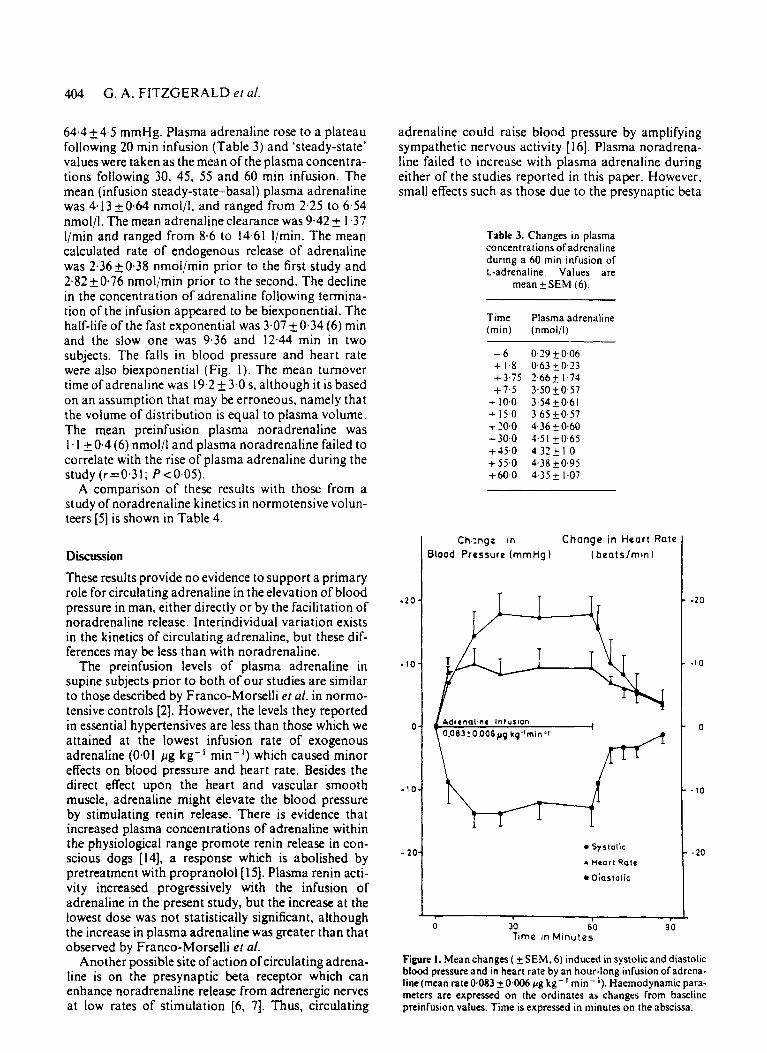
404 G. A. FITZGERALD et al.
64.4 2 4.5 mmHg. Plasma adrenaline rose to a plateau following 20 min infusion (Table 3) and 'steady-state' values were taken as the mean of the plasma concentra- tions following 30, 45, 55 and 60 min infusion. The mean (infusion steady-state-basal) plasma adrenaline was 4.13 kO.64 nmolil, and ranged from 2.25 to 6.54 nmol/l. The mean adrenaline clearance was 9.42 2 1.37 l/min and ranged from 8.6 to 14.61 I/min. The mean calculated rate of endogenous release of adrenaline was 2.36 50.38 nmol/min prior to the first study and 2.82 k 0.76 nmol/min prior to the second. The decline in the concentration of adrenaline following termina- tion of the infusion appeared to be biexponential. The half-life of the fast exponential was 3.07 2 0.34 (6) rnin and the slow one was 9.36 and 12.44 min in two subjects. The falls in blood pressure and heart rate were also biexponential (Fig. I ) . The mean turnover time of adrenaline was 19.2 k 3.0 s. although it is based on an assumption that may be erroneous, namely that the volume of distribution is equal to plasma volume. The mean preinfusion plasma noradrenaline was 1. I & 0.4 (6) nmol/l and plasma noradrenaline failed to correlate with the rise of plasma adrenaline during the study (r=0.31; P ~ 0 . 0 5 ) .
A comparison of these results with those from a study of noradrenaline kinetics in normotensive volun- teers [ 5 ] is shown in Table 4.
Discussion These results provide no evidence to support a primary role for circulating adrenaline in the elevation of blood pressure in man, either directly or by the facilitation of noradrenaline release. Interindividual variation exists in the kinetics of circulating adrenaline, but these dif- ferences may be less than with noradrenaline.
The preinfusion levels of plasma adrenaline in supine subjects prior to both of our studies are similar to those described by Franco-Morselli et al. in normo- tensive controls [2]. However, the levels they reported in essential hypertensives are less than those which we attained at the lowest infusion rate of exogenous adrenaline (0.01 pg kg-' min-I) which caused minor effects on blood pressure and heart rate. Besides the direct effect upon the heart and vascular smooth muscle, adrenaline might elevate the blood pressure by stimulating renin release. There is evidence that increased plasma concentrations of adrenaline within the physiological range promote renin release in con- scious dogs [14], a response which is abolished by pretreatment with propranolol [ 151. Plasma renin acti- vity increased progressively with the infusion of adrenaline in the present study, but the increase at the lowest dose was not statistically significant, although the increase in plasma adrenaline was greater than that observed by Franco-Morselli et al.
Another possible site of action of circulating adrena- line is on the presynaptic beta receptor which can enhance noradrenaline release from adrenergic nerves at low rates of stimulation [6, 1. Thus, circulating
adrenaline could raise blood pressure by amplifying sympathetic nervous activity [ 161. Plasma noradrena- line failed to increase with plasma adrenaline during either of the studies reported in this paper. However, small effects such as those due to the presynaptic beta
Table 3. Changes in plasma concentrations of adrenaline during a 60 rnin infusion of L-adrenaline. Values are
mean 2 S E M (6).
.20
.I0
0
-10
- 20
Time (rnin)
- 6 + 1.8 + 3.75 + 7-5
+ 10.0 + I50 + 20.0 + 30.0 + 35.0 + 55.0 + 60.0
Plasma adrenaline (nmol/l)
0.29 2 0.06 0.63 k 0.23 2.66 2 1.74 3,502 0.57 3.54 2 0.6 I 3.65 2 0.57 4.36 2 0.60 3.51 k0.65 4.32 2 I .o 4.38 20.95 4.352 1.07
Ch.-.ngz in Change in Heart Rate Blood Pressure (mmHg I (beatslmin 1
Adrenaline Infusion I 0.083: 0.006pg kq"min;l R
\
Systolic
A Heart R a t e
rn Diastol ic
.20
. I 0
0
- 10
- 20
I
0 30 60 90 Time in Minutes
Figure 1. Mean changes ( SEM, 6) induced in systolic and diastolic blood pressure and in heart rate by an hour-long infusion of adrena- line (mean rate0.083 +0406 pg kg- l min- l ) . Haemodynamic para- meters are expressed on the ordinates as changes from baseline preinfusion values. Time is expressed in minutes on the abscissa.

METABOLIC EFFECTS AND KINETICS O F ADRENALINE 405
Table 4. Comparative values for plasma concentration. rate of release, clearance and turnover time of adrenaline and noradrenaline. Values are mean kSEM.
Plasma concentration Release rate Clearance Turnover time (nmol/l) (nmol/l) (Ilmin) (s)
Noradrenaline(n=5) 1,12+0.12 5.01 2 1.89 4.46+ 1.02 41.4k 10.2 Adrenaline (n = 6) 0.29 f 0.06 242 f 0.76 9.42 2 1.37 19.2 + 3.0 P < 0.00 I <0.10 <0.01 <0.025
receptor could have been obscured by baroreflex- mediated changes resulting from the fall in diastolic blood pressure induced by adrenaline [ 17, 181.
The increases in blood glucose, plasma renin activity and free fatty acids were of a magnitude that would be anticipated from beta receptor stimulation [ 19-22]. The rise in plasma cyclic AMP provides an indirect measure of the activation of adenylate cyclase. Circu- lating adrenaline may stimulate cyclic GMP produc- tion by activating a adrenoreceptors [23] or via cyclic AMP [24]. However, we observed no rise in plasma cyclic GMP.
The development of sensitive and specific radioenzy- mic assays for catecholamines [ I , 251 enables measure- ments to be made with much greater precision than was possible with fluorimetric methods. The response to sustained elevation of endogenous adrenaline may differ from that evoked by short-term infusions of the exogenous amine. However, these data do not suggest that circulating adrenaline is likely to play an impor- tant part in the direct, or indirect, regulation of blood pressure. There is evidence that the tonic basal secre- tion of adrenaline is regulated by the nervous system and is not just a spillover from the adrenal gland. The concentration of plasma adrenaline falls following the central reduction of sympathetic activity by clondiine [26]. Ariens [27] has suggested that beta 2 receptors may respond mainly to circulating adrenaline and that they should be renamed Beta H (for hormone). The adrenaline infused in this study had profound meta- bolic affects and these are known to be mediated prin- cipally by Beta receptors.
Our data permit only limited conclusions to be drawn about the kinetics of adrenaline in the body. The most striking finding is the very high plasma clear- ance, which must be approximately equal to the car- diac output. There are some difficulties in interpreting full body clearance values from forearm venous sam- ples but our data are directly comparable with those of Franco-Morselli et af. who used the same sampling site. The basal concentration of circulating adrenaline and the clearance derived during infusion permit the total daily adrenaline release to be estimated. Kopin (281 calculated the release in a 70 kg man as being 3-95 pnol/day, using previously published data [ 1 1,29,30]. This agrees well with the mean estimate from our data of 4.06 ,umol/day. Both figures appear high when com- pared with estimates derived from the urinary excre-
tion of unmetabolized adrenaline, but 'more data would be required about the proportion of adrenaline being eliminated by different routes before this ques- tion could be resolved.
The data obtained in this study suggest that the plasma clearance of adrenaline is greater than that of noradrenaline although the result may have been in- fluenced by the differing effects of the two catechola- mines on the resting cardiac output. Further work will be required to define the main sites of adrenaline clear- ance although the very high values obtained must indicate that most organs of the body remove a high proportion of the adrenaline passing through them.
Acknowledgments
We are grateful to Miss B. Edinborough for typing the manuscript. Dr FitzGerald was supported by the Well- come Trust and was in receipt of a Travelling Student- ship from the National University of Ireland during the course of this work.
These data were presented in part at a meeting of the British Pharmacological Society at Leeds (12-14 Sep- tember 1979).
References I Da Prada M. & Zurcher G. (1976) Simultaneous radioenzymatic
deterrninatidn of plasma and tissue adrenaline, noradrenaline and dopamine within the femtomole range. Lije Sci 19. 11161174.
2 Franco-Morselli R., Elghoti J.L.. Joly E., di Giuilio S. & Myer P. (1977) Increased plasma adrenaline concentrations in benign essential hypertension. Br Med J ii, 1251-1254.
3 De Quattro V. & Chan S. (1972) Raised catecholamines in some patients with essential hypertension. Lancer i, 806-809.
4 Winer N., Chokshi D.S. & Walkenhorst W.G. (1971) Effects of cyclic AMP, sympathomimetic amines and adrenergic receptor antagonists on renin secretion. Circ Res 29,23%248.
5 Stjarne L. & Burundin J. (1975) Dual adrenoreceptor mediated control of noradrenaline secretion from human vasoconstrictor nerves: facilitation by beta receptors. Aria Physiol Scund 94, 139-141.
6 Stjarne L. & Burundin J. (1976) Beta2-adrenoreceptors facilitiat- ing noradrenaline secretion from human vasoconstrictor nerves. Acfa Physiol Scand 97,88-93.
7 Celuch S.M., Dubocovich M. & Langer S.Z. (1978) Stimulation of presynaptic beta adrenoreceptors enhances (H')-noradrena- line release during nerve stimulation in the perfused cat spleen. Br J Pharmac 63.97-109.

406 G . A. FITZGERALD r l al.
X FitzGerald G.A.. Hossmann V., Hamilton C.A.. Reid J.L.. Davies D.S. & Dollery C.T. (1979) Interindividual variation in noradrenaline kinetics in man. Clin Plrurmucol Ther 26.669-67s.
9 Fruton J.S. & Simmonds S . (1958) Chemistry of the carbo- hydrates. General Bioc.hrtni.irr.v. 4th edn. pp. 402-429. Wiley. New York.
10 Dole V.P. (1956) A relation between non-esterified fatty acids in . plasma and the metabolism of glucose. J Clin Inrest 35, 150-1 54.
I I Cohen G., Holland B.. Sha J . & Goldenberg ,M. (1959) Plasma concentrations of epinephrine and norepinephrine during intra- venous infusions in man. J Clin Invest 38, 1935-1941.
12 Wennesland R.. Browne Hopper J., Hodges L.. Guttentag O.E., Scott K.G.. Tucker I.N. & Bradley B. (1959) Red cell and blood volume in healthy man: CrS. J Clrn Inrrsr 38. 1065-1077.
I3 Lord E. (1947) The use of range in place of standard deviation in the t test. Biomerriko 34,40-67.
14 Johnson M.D.. Shier D.N. & Clifford Berger A. (1979) Circulat- ing catecholamines and control of plasma renin activity in con- scious dogs. A m J Physrol236, H463-H470.
I S Johnson. M.D.. Fahri E.R.. Troen B.R. & Berger A.C. (1979) Plasma epinephrine and control of plasma renin activity. possible extrarenal mechanisms. An1 J Physiol236, H854-HS59.
16 Starke K. (1977) Regulation of noradrenaline release by pre- synaptic receptor systems. Rer Phviol Biochem Phurtnucol 77. 1-114.
I7 Heymans C. (1929) Le sinus carotidien. zone reflexogene regula- trice du tonus vagal cardiaque. du tonus neurovaculaire et de I’adrenalino-secretion. Arch lnr Phurntacodwr Ther 35. 269-306.
18 Ead H.W.. Green J.H. & Neil E. (1952) A comparison of the efiects of pulsatile and nonpulsatile blood flow through the carotid sinus of the reflexogenic activity of the sinus baroreceptor in the cat. J Phy.riol I IS, 509-5 19.
19 YoungJ.B. & Landsberg L. ( 1978) Catecholamines and interme- diary metabolism. Clrriics €ndocrinol Mt,/ah 6, 599-63 I .
20 Himms-Hagen J. (1972) Effects of catecholamines on metabo- lism. Hantlhook of €.rperinrmral Pharmuc.oloxr.. XXXl I I . Caw- cholamines (ed. by H. Blaschko and E. Muscholl). pp. 363-461. Springer, Berlin.
21 No1lyH.L.. Reid I .A.&Ganong W.F.(1974)Efiectoftheophyl- line and adrenergic blocking drugs on the renin response to norepinephrine in vitro. Circ Rrs 35, 575-579.
22 Cooper B.. Partilla J.S. & Griegerman R.I. (1975) Adenylate cyclase of human fat cells: expression of epinephrine-sensitive activation revealed by 5’-guanylyl-imidodiphosphate. J Clin ln- W S I 56, 1350- 1353.
23 Goldberg N.D.. Haddox M.D.. Nicol S.E.. Glass D.B.. Sanford C.H., Kuehl F.A. & Estensen R. (1975) In: Adiunrr.7 in Cyclic. Nucleoridr R1warc.h (ed. by G. I . Drummond. P. Greengard and G. A. Robinson). Vol. 5. pp. 307-330. Raven Press, New York.
24 Asakawa T.. Ruiz J . & Ho R.-J. (1978) Epinephrine induced elevation of guanosine 3’-5’cyclic monophosphate in isolated fat cells of rat. Proc. Nod Acotl Sci 75, 26842688.
25 Henry D.P.. Starman B.J.. Johnson D.G. & Williams R.H. (1975) A sensitive radioenzymatic assay for norepinephrine in tissues and plasma. Lgi Sci 16. 375-384
26 Brown M.J.. Dollery C.T.. FitzGerald G.A.. Watkins J. & Zam- boulis C. (1980) No evidence for antagonism of clonidine by naloxone in man. Br J Clin Pharnr 9 , 3 0 2 ~ .
27 Aeriens E.J. & Simanis A.M. (1977) Receptors and Receptor Mechanisms. Brra-A(lrnrorec.epror B l i ~ ~ k i t r g A,yeni.v (ed. by P R. Saxena and R. P. Forsyth). pp. 3-27. Elsevier-North Holland, Amsterdam.
78 Kopin I.J. (1979) Biochemical assessment of peripheral adrener- gic activity. The Rrlrurr of Currc~ho/uttriirrs / h i i A i !~mrr~+ Nertronrs (ed. by D. M . Paton). pp. 355-372. Perpamon Press. Oxford.
19 Vendsalu A. (1960) Studies o n adrenaline and noradrenaline in human plasma. Acra Phr.siol Scant1 49. Suppl. 173.
30 Gitlow S.E.. Mendlowitz M.. Smith A,. Gall E.. Wolf R.L. & Naftchi W.E. (1961) The dynamics of norepinephrine metaho- lism. Hypertoision: Rec~c~ir~ Adruniw on Hvptvrcnsir~c~ Di.ri~u.rc~. 2nd Hannrrrrun .S,wiipo.rrisrr (ed. by A. M. Brest and J . H. Moycr). pp. 341 -35 I . Lei & Febiger. Philadelphia.





























