Embed Size (px)
Citation preview

Chromosomal elementsconferring epigeneticinheritanceFrank Lyko and Renato Paro*
Summary
Epigenetic regulation of transcription can lead to a stable differential expression ofidentical genetic information in the same cell or cell population. There is increasingevidence that higher order chromatin structures, involving specific multiproteincomplexes, constitute one device to establish and maintain epigenetic marks. Inaddition, defined chromosomal elements conferring epigenetic inheritance oftranscriptional expression states have recently been identified. During the periodwhere the difference in expression of identical genes is established, thesesequences appear to be used as switch elements by both negative and positiveregulators. Once the epigenetic mark is ‘‘set’’, the elements maintain either thesilenced or the activated expression state over many cell generations. Here wereview recent data obtained from analyzing epigenetic gene regulation in differentorganisms and show that similarities in the underlying mechanisms appear toexist. BioEssays 21:824–832, 1999. r 1999 John Wiley & Sons, Inc.
IntroductionThe stable and heritable silencing of specific chromosomaldomains is a fundamental aspect of the eukaryotic genome. Itaffects many nuclear processes such as the regulation ofgene expression, DNA replication, and DNA repair, as well aschromosome segregation. Silencing also forms the basis ofepigenetic mechanisms that result in a differential interpreta-tion of identical genetic information in the same cell. Typically,one piece of genetic material is expressed in a given nucleuswhile its homologous counterpart or an equivalent DNAsequence located elsewhere in the genome remains silent.This phenomenon takes place even in unicellular yeast, in theform of the regulation of the mating type loci and it extends tomammalian systems, in the form of X-chromosome inactiva-
tion and genomic imprinting as its most prominent manifesta-tions. Silencing also underlies the process of ‘‘cellular memory’’in fixing a determined state of gene expression by anepigenetic mechanism and thus constituting a memory ofdevelopmental decisions in individual cells. The program ofthe cells is maintained stably over many cell divisions and is,therefore, mitotically heritable.
There is increasing evidence that epigenetically controlledprocesses depend on the intricacies of chromatin structure.Genetic analyses in model organisms like yeast or Drosophilahave revealed many chromatin-associated factors that areinvolved in silencing, as well as other features of epigeneticinheritance. Furthermore, chromosomal aberrations as wellas transgenic experiments have uncovered discrete chromo-somal elements mediating epigenetic control of gene expres-sion. As we show in this review, chromatin-mediated switchesin expression might be the central feature of several distinctepigenetic phenomena.
Silencing in heterochromatinHeterochromatic domains of eukaryotic genomes are gene-poor, but rich in repetitive DNA. The latter appears to becorrelated with the potent silencing abilities of this section ofthe genome, often turning it into a graveyard for transposableelements. The strong repressive functions, considered to becaused by a dense chromatin packaging, become very
ZMBH, University of Heidelberg, Im Neuenheimer Feld 282, 69120Heidelberg, Germany.Funding agencies: Deutsche Forschungsgemeinschaft; Human Fron-tier Science Program; Fonds der Chemischen Industrie.Present address for Frank Lyko: Whitehead Institute for BiomedicalResearch, 9 Cambridge Center, Cambridge, MA 02142, USA.*Correspondence to: Renato Paro, ZMBH, Universitat Heidelberg,Postfach 106249, 69052 Heidelberg, Germany.E-mail: paro@sun0. urz.uni-heidelberg.de
Review articles
824 BioEssays 21.10 BioEssays 21:824–832, r 1999 John Wiley & Sons, Inc.

apparent at junctions between hetero- and euchromatin. Theencounter of a euchromatic gene with nearby heterochroma-tin (by translocation or transgene insertion) results in amosaic expression of the gene, essentially demonstrating acompetition between the silencing structures acting over along range and activating factors counteracting the repres-sion. The patchy nature of the observed gene expressionpatterns seems to indicate that competition takes placeduring a limited window of time in development. At a certainthreshold, the decision (between the active and the inactivestate) becomes fixed and is subsequently clonally inheritedby the progenitor cells. This phenomenon, termed positioneffect variegation (PEV), has been extensively used in Dro-sophila and yeast to identify participating trans-acting factorsby genetic means. Subsequent molecular analyses sup-ported the genetic indications by uncovering many newchromatin- and DNA-associated regulatory factors.(1,2)
The proximity of transcriptionally silent genes to centro-meric heterochromatin has also been observed in mousecells(3) and, similarly, murine transgenes can display varie-gated expression patterns. In cases of insertions close tocentromeres, tissue sections from respective transgenic micerevealed clusters of expressing cells that are surrounded bynonexpressing cells.(4,5) Until now, the specific cis-actingheterochromatic DNA sequences through which the modifiersof PEV exert their silencing function remained elusive. How-ever, an expression pattern highly similar to classical PEVcan be observed when transgenes are integrated into the flygenome as a multicopy array. Repression of these arrays canbe influenced by known modifiers of PEV and by the distanceof the array to centromeric heterochromatin, suggesting afunctional relationship to PEV.(6) Silencing of transgene re-peats has also been observed in the mouse.(7) When thetransgene was integrated as a high copy-number array,expression was barely detectable. Only a significant reduc-tion in copy number allowed for the expression of thetransgene. The mechanism of repeat-induced silencing inmammals is still unknown but it appears to involve heterochro-matin.(7)
The molecular structure of heterochromatin in highereukaryotes is not well understood, but knowledge derivedfrom yeast might serve as a useful paradigm. In yeast,silenced domains can also induce epigenetic control ofneighboring transgenes. The resulting chromatin structuresare very stable and can be maintained through mitosis andmeiosis.(8–10) The silencing process is best characterized attelomeres requiring, among others, the SIR proteins tospecifically interact with the H3 and H4 histone tails (reviewedin Ref. 11). The interaction of SIR proteins with the nucleo-somal backbone seems to generate a particular form ofhigher order chromatin structure. Additionally, the observedclustering of telomeres(12,13) could be a means to furtherstabilize this structure by the formation of larger, more stable
complexes. Indeed, subnuclear compartmentalization of si-lencing complexes appears to be an important part of theprocess, as a similar requirement has also been demon-strated for PEV-inducing heterochromatin structures in Dro-sophila (reviewed in Refs. 14, 15).
Silencing of developmental regulatorsIn a complex organism, cell identities are defined by aspecifically expressed set of controlling factors. The homeoticgenes represent a well-known example whose productsdetermine the fate of cells within the anterior–posterioraxis.(16) In Drosophila their specific expression pattern wasfound to be set early in embryogenesis by transiently actingtranscription factors. Two distinct sets of factors, encoded bythe Polycomb group (PcG) and trithorax group (trxG) ofgenes, subsequently maintain cell fates through growth anddifferentiation. This maintenance function utilizes epigeneticfeatures. After early patterning, the PcG proteins recognize arepressed state of a target gene and translate it into amitotically stable, silent chromatin structure. Conversely,some members of the trxG counteract the silencing effectsand stabilize open chromatin structures such that continuousgene expression can be maintained. Once established earlyin embryogenesis, the respective chromatin states are faith-fully inherited through development. The interplay betweenchromatin complexes containing PcG and trxG proteinsensures that differential expression patterns of regulatorygenes can be maintained.
The function of both groups appears to be conservedthroughout evolution.(17) Homologs have been identified inplants as well as in animals and humans. In mice, recentevidence from transgenic and knockout lines indicate that thistype of regulation is not only necessary during developmentfor regulating the HOX genes but is also important duringadulthood. An aberrant PcG or trxG function is often found incancer, in particular, leukemias. Indeed, PcG-mediated genesilencing is directly involved in certain aspects of cell cyclecontrol and senescence.(18)
Despite the fact that PcG homologs have been identified invarious organisms, we still know very little about the exactmolecular structures these proteins use to influence geneexpression. Genetic as well as molecular parallels betweenPcG silencing and PEV suggested a common basic mecha-nism, however. Although the molecular details are unknownin both cases, it is considered that a dense compaction of thechromatin fiber forms the core of such inert chromosomaldomains and prevents the accessibility of sequence-specificfactors to DNA. Alternatively, PcG proteins might retractrepressed genes to particular nuclear subcompartments thatare transcriptionally inactive. This idea is supported by thefinding that PcG proteins coagulate in nuclear speckles.(19,20)
In addition, it was recently shown that PcG and trxG proteins
Review articles
BioEssays 21.10 825
interact with elements functioning as chromatin insulators,(21)
thereby linking factors regulating chromatin structure to archi-tectural elements like boundaries and matrix attachmentsites.
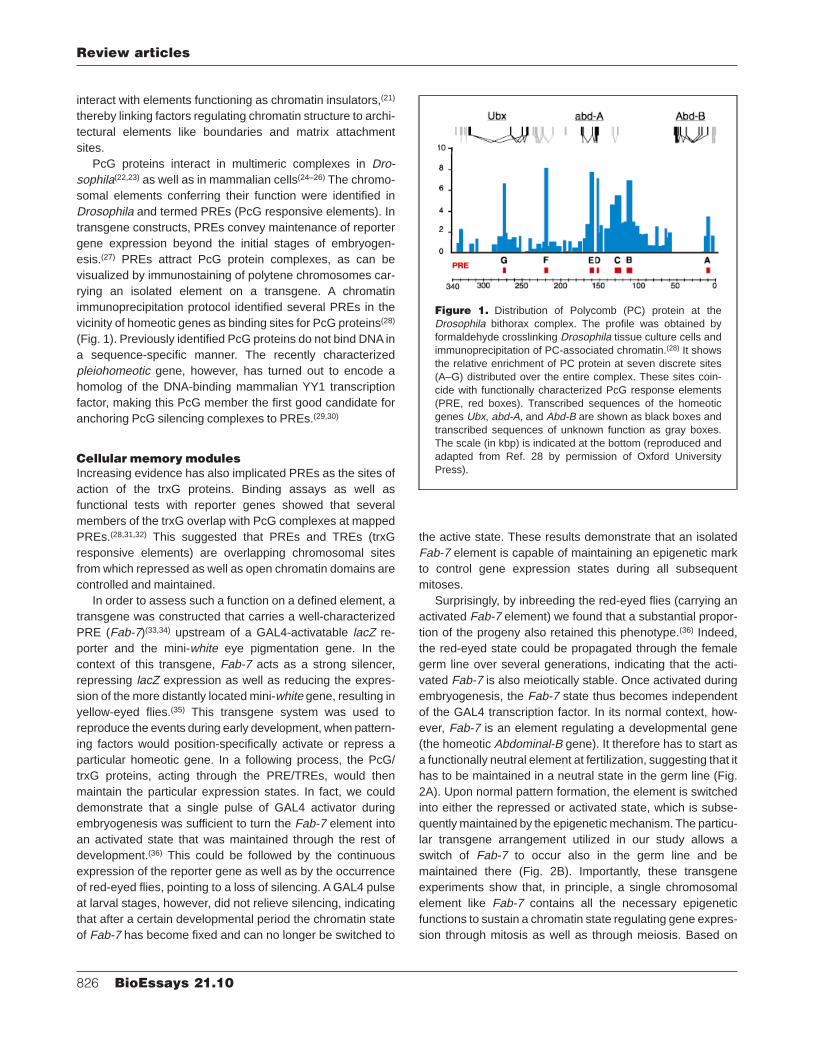
PcG proteins interact in multimeric complexes in Dro-sophila(22,23) as well as in mammalian cells(24–26) The chromo-somal elements conferring their function were identified inDrosophila and termed PREs (PcG responsive elements). Intransgene constructs, PREs convey maintenance of reportergene expression beyond the initial stages of embryogen-esis.(27) PREs attract PcG protein complexes, as can bevisualized by immunostaining of polytene chromosomes car-rying an isolated element on a transgene. A chromatinimmunoprecipitation protocol identified several PREs in thevicinity of homeotic genes as binding sites for PcG proteins(28)
(Fig. 1). Previously identified PcG proteins do not bind DNA ina sequence-specific manner. The recently characterizedpleiohomeotic gene, however, has turned out to encode ahomolog of the DNA-binding mammalian YY1 transcriptionfactor, making this PcG member the first good candidate foranchoring PcG silencing complexes to PREs.(29,30)
Cellular memory modulesIncreasing evidence has also implicated PREs as the sites ofaction of the trxG proteins. Binding assays as well asfunctional tests with reporter genes showed that severalmembers of the trxG overlap with PcG complexes at mappedPREs.(28,31,32) This suggested that PREs and TREs (trxGresponsive elements) are overlapping chromosomal sitesfrom which repressed as well as open chromatin domains arecontrolled and maintained.
In order to assess such a function on a defined element, atransgene was constructed that carries a well-characterizedPRE (Fab-7)(33,34) upstream of a GAL4-activatable lacZ re-porter and the mini-white eye pigmentation gene. In thecontext of this transgene, Fab-7 acts as a strong silencer,repressing lacZ expression as well as reducing the expres-sion of the more distantly located mini-white gene, resulting inyellow-eyed flies.(35) This transgene system was used toreproduce the events during early development, when pattern-ing factors would position-specifically activate or repress aparticular homeotic gene. In a following process, the PcG/trxG proteins, acting through the PRE/TREs, would thenmaintain the particular expression states. In fact, we coulddemonstrate that a single pulse of GAL4 activator duringembryogenesis was sufficient to turn the Fab-7 element intoan activated state that was maintained through the rest ofdevelopment.(36) This could be followed by the continuousexpression of the reporter gene as well as by the occurrenceof red-eyed flies, pointing to a loss of silencing. A GAL4 pulseat larval stages, however, did not relieve silencing, indicatingthat after a certain developmental period the chromatin stateof Fab-7 has become fixed and can no longer be switched to
the active state. These results demonstrate that an isolatedFab-7 element is capable of maintaining an epigenetic markto control gene expression states during all subsequentmitoses.
Surprisingly, by inbreeding the red-eyed flies (carrying anactivated Fab-7 element) we found that a substantial propor-tion of the progeny also retained this phenotype.(36) Indeed,the red-eyed state could be propagated through the femalegerm line over several generations, indicating that the acti-vated Fab-7 is also meiotically stable. Once activated duringembryogenesis, the Fab-7 state thus becomes independentof the GAL4 transcription factor. In its normal context, how-ever, Fab-7 is an element regulating a developmental gene(the homeotic Abdominal-B gene). It therefore has to start asa functionally neutral element at fertilization, suggesting that ithas to be maintained in a neutral state in the germ line (Fig.2A). Upon normal pattern formation, the element is switchedinto either the repressed or activated state, which is subse-quently maintained by the epigenetic mechanism. The particu-lar transgene arrangement utilized in our study allows aswitch of Fab-7 to occur also in the germ line and bemaintained there (Fig. 2B). Importantly, these transgeneexperiments show that, in principle, a single chromosomalelement like Fab-7 contains all the necessary epigeneticfunctions to sustain a chromatin state regulating gene expres-sion through mitosis as well as through meiosis. Based on
Figure 1. Distribution of Polycomb (PC) protein at theDrosophila bithorax complex. The profile was obtained byformaldehyde crosslinking Drosophila tissue culture cells andimmunoprecipitation of PC-associated chromatin.(28) It showsthe relative enrichment of PC protein at seven discrete sites(A–G) distributed over the entire complex. These sites coin-cide with functionally characterized PcG response elements(PRE, red boxes). Transcribed sequences of the homeoticgenes Ubx, abd-A, and Abd-B are shown as black boxes andtranscribed sequences of unknown function as gray boxes.The scale (in kbp) is indicated at the bottom (reproduced andadapted from Ref. 28 by permission of Oxford UniversityPress).
Review articles
826 BioEssays 21.10

these capabilities, Fab-7 was termed a ‘‘cellular memorymodule’’ (CMM). As the Drosophila genome has no recog-nized covalent modification like DNA methylation, the epige-netic mark is most likely formed by protein complexes such asthose encoded by the PcG and the trxG of genes.
Parent-of-origin-dependent silencing of allelesThe term ‘‘imprinting’’ was originally defined to describeparent-of-origin-dependent inheritance of a genetic trait ininsects.(37) The same term is now widely used to denoteparent-of-origin-dependent silencing of autosomal genes.Although related phenomena, affecting sex chromosome-
based markers, have been reported in Drosophila,(38,39) ge-nomic imprinting in the strict sense of its current definitionappears to be restricted to mammals. To date, more than 20imprinted genes have been identified in mice and humans.(40)
For some of these genes, silencing affects all cells of anorganism, whereas for others it appears to be highly tissue-specific. Tissue-specific imprinting is best illustrated by theUbe3A gene, which is silenced on the paternal allele only inhippocampal and Purkinje neurons.(41) In addition, imprintingcan be highly polymorphic. A prominent example is the IGF2Rgene, which is paternally silenced only in about 50% of thehuman population.(42) The tissue-specificity as well as the
Figure 2. Mitotic and meiotic maintenance of chromatin states at CMMs. A: During normal development in the germ line CMMs arenormally kept in a neutral state and marked by particular chromatin proteins sustaining the epigenetic tag. In the early embryo, the germline is soon separated from the somatic line, thus maintaining the neutral CMM state. In the somatic cells, the mechanism of patternformation switches a particular CMM either into the ON-state by gene activation or into the repressed OFF-state. At a particular timeduring embryogenesis, the specific chromatin state at a CMM becomes fixed by the respective PcG/trxG protein combinations and isclonally inherited in a mitotically stable manner. B: Isolating a CMM on a transgene, activatable by GAL4, allows the switch to occur in thegerm line. This particular CMM remains in the ON-state in the zygote and allows transcription of the nearby gene it is regulating. In thesomatic line, the ON-state is again mitotically stable. However, it can stochastically fall back into the OFF-state. Similarly, the ON-state ofthe CMM in the germ line can be maintained through multiple generations; however, with a strong tendency of the element to return to theOFF-state.
Review articles
BioEssays 21.10 827
polymorphic occurrence of silencing might be indicative of acomplex, metastable imprinting mechanism.
The imprinting machinery requires target sequences to actupon, and cis-regulatory imprinting control elements (ICEs)(43)
have been defined recently. One example is the imprintingcenter for human chromosome 15q11-q13, which is definedby microdeletions in a particular class of Prader-Willi syn-drome and Angelman syndrome patients.(44) Another ex-ample is the ICE of the mouse H19 gene, which is defined bytargeted deletions of the mouse H19 upstream region.(45)
These elements extend over several hundred base pairs ofDNA and are absolutely required for correct epigeneticinheritance. Up to now, sequence comparison of ICEs havefailed to reveal any similarities or other functionally character-ized motifs.
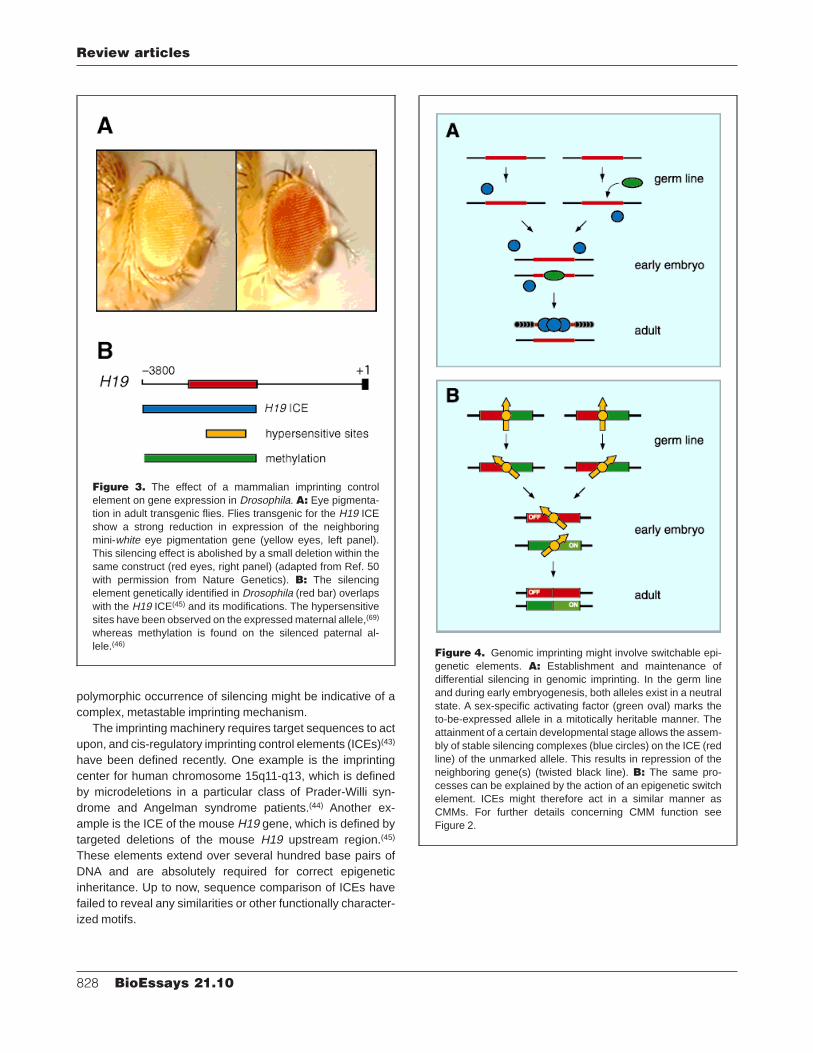
Figure 3. The effect of a mammalian imprinting controlelement on gene expression in Drosophila. A: Eye pigmenta-tion in adult transgenic flies. Flies transgenic for the H19 ICEshow a strong reduction in expression of the neighboringmini-white eye pigmentation gene (yellow eyes, left panel).This silencing effect is abolished by a small deletion within thesame construct (red eyes, right panel) (adapted from Ref. 50with permission from Nature Genetics). B: The silencingelement genetically identified in Drosophila (red bar) overlapswith the H19 ICE(45) and its modifications. The hypersensitivesites have been observed on the expressed maternal allele,(69)
whereas methylation is found on the silenced paternal al-lele.(46)
Figure 4. Genomic imprinting might involve switchable epi-genetic elements. A: Establishment and maintenance ofdifferential silencing in genomic imprinting. In the germ lineand during early embryogenesis, both alleles exist in a neutralstate. A sex-specific activating factor (green oval) marks theto-be-expressed allele in a mitotically heritable manner. Theattainment of a certain developmental stage allows the assem-bly of stable silencing complexes (blue circles) on the ICE (redline) of the unmarked allele. This results in repression of theneighboring gene(s) (twisted black line). B: The same pro-cesses can be explained by the action of an epigenetic switchelement. ICEs might therefore act in a similar manner asCMMs. For further details concerning CMM function seeFigure 2.
Review articles
828 BioEssays 21.10

Differential DNA methylation is widely held to be theprimary cause of genomic imprinting. Interallelic methylationdifferences appear to be exceptionally stable in the vicinity ofICEs,(46,47) suggesting that methylation confers imprinting byacting upon ICEs. Although very precise, these results arecorrelations and do not directly demonstrate a functional linkbetween DNA methylation and imprinting. To date, a func-tional role of methylation in imprinting can only be inferredfrom the observation that mice carrying a targeted disruptionof the DNA methyltransferase gene show loss of imprintingfor some, but not all, imprinted loci(48,49) This finding demon-strated that methylation is functionally involved in the mainte-nance of imprinting. Yet the precise level of action of DNAmethylation remains unclear, and other results clearly call foradditional mechanisms involved in the stable repression ofimprinted genes.
When the two aforementioned ICEs were introduced astransgenes into Drosophila, both elements were capable ofsignificantly reducing the expression of several neighboringreporter constructs(50,51) (see Fig. 3A). Silencing was ob-served irrespective of whether the transgenes were transmit-ted through the maternal or the paternal germ line. The H19silencing element could be genetically delineated and showsa substantial overlap with the mouse H19 ICE and itsepigenetic modifications (Fig. 3B). Similarly, the SNRPNsilencing element is entirely contained within the chromo-some 15q11-q13 ICE.(51) The striking overlap between silenc-ing elements defined in Drosophila and the respective mam-malian imprinting elements implies a mechanistic link betweenthe observed fly silencing and genomic imprinting.
The precise nature of the conserved silencing mechanismis presently unclear, but some conclusions can be drawn. AsDrosophila lacks cytosine methylation, this covalent modifica-tion of DNA cannot account for the observed effects. Instead,the results point to a protein-mediated mechanism that isconserved between flies and mammals participating in imprint-ing. The chromatin structure of imprinted genes has beendirectly investigated by nuclease digestion of isolated nuclei.The results revealed that at least two genes, H19 andU2af1-rs1, show differential nuclease accessibilities.(52–54) Inboth cases, the repressed allele was found to be significantlymore resistant to digestion, indicating a ‘‘closed’’ chromatinconformation. The involvement of chromatin in the repressionof the paternal H19 allele is further supported by the observa-tion that silencing can be partially relieved by trichostatin A, aknown inhibitor of histone deacetylation.(55) A special chroma-tin configuration of the repressed H19 allele might be ac-quired as early as in the germ line, as demonstrated bychromatin structural analysis in spermatozoa.(56)
Given these findings and the universal role of chromatinstructure in epigenetic regulation of gene expression, it is
conceivable that chromatin-mediated silencing participates inimprinting by using ICEs as target sequences. An imprintingmodel based on silencing can explain these findings. Silenc-ing elements dispersed over an imprinted domain could bringabout repression of imprinted genes. Every silencer shouldattract a certain number of silencing proteins. These proteinswould then interact by looping out intervening sequences toform a large silencing complex that leads to general repres-sion of the entire cluster. Similar structures have beenpreviously proposed to form the basis of PcG- and heterochro-matin-mediated silencing(57,58) (see Fig. 1). Finally, preventingthe build-up of silencing factors in a parent-of-origin-dependent manner (as described below) would result indifferential, imprinted gene expression.
Several findings concerning the higher order regulation ofimprinted genes can be explained by a cooperative silencingmodel. 1) For both known clusters of imprinted genes, it isalways one particular allele that is preferentially silenced: thepaternal allele for the cluster containing the H19 gene(59) andthe maternal allele for the 15q11-q13 cluster.(60) This can beinferred by the requirement for cooperative interactions be-tween silencing elements to achieve long-range repression.2) ICEs from two mammalian loci have been shown tofunction as methylation-independent silencers in Drosophila.These elements and the appropriate factors bound to themmight participate in a silencing mechanism, as outlinedabove. 3) Short transgenes derived from imprinted loci canshow appropriate imprinting, but consistent imprinting is onlyachieved by large YACs.(61,62) This presumably reflects acertain sensitivity of isolated transgenes towards genomicposition effects, since short constructs would fail to containthe required number of ICEs to form stable silencing com-plexes. Apart from its general simplicity, a cooperative silenc-ing model of genomic imprinting can thus explain severalobservations that are difficult to accommodate by othermodels.
As noted above, features distinct from chromatin-medi-ated silencing are also associated with imprinting. Theseinclude DNA methylation, replication timing,(63) and competi-tion for expression on opposite parental alleles.(64) While theoutlined model predicts a primary role for chromatin-mediated silencing, methylation could serve as a mainte-nance mechanism to stabilize repression. Accordingly, boththe silencing mechanism and the DNA methylation machineryappear to converge on the same DNA elements, the ICEs. Inthe case of imprinted genes, chromatin structure and DNAmethylation would thus be the primary determinants of therepressed epigenetic state. Once a particular epigenetic statehas been established, it could cause other properties ofimprinted loci such as asynchronous replication and expres-sion competition.
Review articles
BioEssays 21.10 829

Imprinting control elements as epigeneticswitchesThe differential expression of imprinted genes requires thatsilencing be overcome by activation of one allele betweengenerations. This could be established in the germ line bysex-specific activating factors through interaction with theICEs(65) (Fig. 4A). After resetting, in the germ line or duringearly embryogenesis, both alleles would exist in a neutralstate, not allowing for functional repression (Fig. 4A). Thiswould explain the observed biallelic expression of imprintedgenes in the germ line and during preimplantation develop-ment.(66,67) The mark acquired by the active allele could bepropagated in a mitotically stable manner, as for the previ-ously described CMMs. Only after a certain developmentalstage could functional silencing complexes be assembled onthe unprotected allele, resulting in differential repression (Fig.4A). In several ways, ICEs would thus be fundamentallysimilar to CMMs (Fig. 4B). Both classes of elements arecapable of conveying both the active and the inactive state ina stable and heritable manner.
Even though both ICEs and PREs might be functionallyrelated and considered CMMs, they do not necessarily haveto utilize the same set of trans-acting factors. Yet, theclassification of ICEs as CMMs predicts a dual role for theseelements in imprinting: on one allele they would be requiredfor repression, while on the other allele they would permitactivation. Interestingly, the two presently best-defined ICEsappear to meet this prediction: the inheritance pattern of thePrader-Willi syndrome requires a ‘‘switch element’’ in thechromosome 15q11-q13 ICE that is necessary for the estab-lishment of both the silenced and the active epigenotype.(68)
And a targeted deletion of the mouse H19 ICE revealed thatthe element is not only required for silencing of the normallyrepressed allele, but also for full activation of the normallyexpressed allele.(45)
ConclusionsThe stable and heritable silencing of chromosomal domainsforms an important part of epigenetic mechanisms. Analysesin genetic model organisms like yeast and Drosophila uncov-ered many chromatin components necessary to generateand sustain the repressive structures. In certain organisms,the additional recruitment of DNA methylation to furtherstabilize these structures seems to have occurred duringevolution. There is increasing evidence that features regulat-ing the epigenetically maintained OFF state of a domain andthe factors keeping the ON state act through common DNAelements, thus functioning as epigenetic switches. In yeastand Drosophila, such elements have been identified and theirfunctions have been tested in defined transgene assays. Inmammalian systems, analysis of mutant sequences as wellas transgene constructs points to the existence of equiva-lently acting chromosomal switches in regulating epigeneticphenomena like genomic imprinting. Part of the mechanism
appears to have been conserved during evolution. Futureanalyses will reveal whether the underlying chromosomalstructures and the trans-acting factors regulating the epige-netic features are also comparable between the systems.
AcknowledgmentsThe authors thank Justin Ainscough, Karin Buiting, BernhardHorsthemke, John Greally, and members of the group forcritically reading the manuscript and for valuable comments.
Note added in proofDetailed nuclease digestion of the mouse H19 ICE recentlydemonstrated strong differences between the maternal andthe paternal allele.(70) A similar analysis of the chromosome15q11-q13 imprinted region also revealed strong differentialhypersensitive sites within the chromosome 15q11-q13 ICE.(71)
Both findings provide further evidence that imprinting can beregulated by differential chromatin states.
References1. Reuter G, Spierer P. Position effect variegation and chromatin proteins.
BioEssays 1992;14:605–612.2. Pillus L, Grunstein M. Chromatin structure and epigenetic regulation in
yeast. In: Elgin SCR, editor. Chromatin structure and gene expression.Oxford: Oxford University Press; 1995. p 123–146.
3. Brown KE, Guest SS, Smale ST, Hahm K, Merkenschlager M, Fisher AG.Association of transcriptionally silent genes with Ikaros complexes atcentromeric heterochromatin. Cell 1997;91:845–854.
4. Festenstein R, Tolaini M, Corbella P, Mamalaki C, Parrington J, Fox M,Miliou A, Jones M, Kioussis D. Locus control region function andheterochromatin-induced position effect variegation. Science 1996;271:1123–1125.
5. Dobie KW, Lee M, Fantes JA, Graham E, Clark AJ, Springbett A, Lathe R,McClenaghan M. Variegated transgene expression in mouse mammarygland is determined by the transgene integration locus. Proc Natl AcadSci USA 1996;93:6659–6664.
6. Dorer DR, Henikoff S. Transgene repeat arrays interact with distantheterochromatin and cause silencing in cis and trans. Genetics 1997;147:1181–1190.
7. Garrick D, Fiering S, Martin DI, Whitelaw E. Repeat-induced genesilencing in mammals. Nat Genet 1998;18:56–59.
8. Grewal SI, Klar AJ. Chromosomal inheritance of epigenetic states infission yeast during mitosis and meiosis. Cell 1996;86:95–101.
9. Ekwall K, Olsson T, Turner BM, Cranston G, Allshire RC. Transient inhibitionof histone deacetylation alters the structural and functional imprint atfission yeast centromeres. Cell 1997;91:1021–1032.
10. Klar AJ. Propagating epigenetic states through meiosis: where Mendel’sgene is more than a DNA moiety. Trends Genet 1998;14:229–301.
11. Grunstein M. Molecular model for telomeric heterochromatin in yeast. CurrOpin Cell Biol 1997;9:383–387.
12. Palladino F, Laroche T, Gilson E, Axelrod A, Pillus L, Gasser SM. SIR3 andSIR4 proteins are required for the positioning and integrity of yeasttelomeres. Cell 1993;75:543–555.
13. Gotta M, Laroche T, Formenton A, Maillet L, Scherthan H, Gasser SM. Theclustering of telomeres and colocalization with Rap1, Sir3, and Sir4proteins in wild-type Saccharomyces cerevisiae. J Cell Biol 1996;134:1349–1363.
14. Henikoff S, Comai L. Trans-sensing effects: the ups and downs of beingtogether. Cell 1998;93:329–332.
15. Wakimoto BT. Beyond the nucleosome: epigenetic aspects of position-effect variegation in Drosophila. Cell 1998;93:321–324.
16. McGinnis W, Krumlauf R. Homeobox genes and axial patterning. Cell1992;68:283–302.
Review articles
830 BioEssays 21.10

17. Gould A. Functions of mammalian Polycomb group and trithorax grouprelated genes. Curr Opin Genet Dev 1997;4:488–494.
18. Jacobs JJL, Kieboom K, Marino S, DePinho RA, van Lohiuzen M. Theoncogene and Polycomb-group gene bmi-1 regulates cell proliferationand senescence through the ink4a locus. Nature 1999;397:164–168.
19. Buchenau P, Hodgson J, Strutt H, Arndt-Jovin DJ. The distribution ofpolycomb-group proteins during cell division and development in Dro-sophila embryos: impact on models for silencing. J Cell Biol 1998;141:469–481.
20. Dietzel S, Niemann H, Bruckner B, Maurange C, Paro R. The nucleardistribution of Polycomb during Drosophila melanogaster developmentshown with a GFP fusionprotein. Chromosoma 1999;108:83–94.
21. Gerasimova TI, Corces VG. Polycomb and trithorax group proteinsmediate the function of a chromatin insulator. Cell 1998;92:511–521.
22. Franke A, DeCamillis M, Zink D, Cheng N, Brock HW, Paro R. Polycomband polyhomeotic are constituents of a multimeric protein complex inchromatin of Drosophila melanogaster. EMBO J 1992;11:2941–2950.
23. Kyba M, Brock HW. The Drosophila polycomb group protein Psc contactsph and Pc through specific conserved domains. Mol Cell Biol 1998;18:2712–2720.
24. Alkema MJ, Bronk M, Verhoeven E, Otte A, van’t Veer LJ, Berns A, vanLohuizen M. Identification of Bmi1-interacting proteins as constituents of amultimeric mammalian polycomb complex. Genes Dev 1997;11:226–240.
25. Sewalt RG, van der Vlag J, Gunster MJ, Hamer KM, den Blaauwen JL,Satijn DP, Hendrix T, van Driel R, Otte AP. Characterization of interactionsbetween the mammalian polycomb-group proteins Enx1/EZH2 and EEDsuggests the existence of different mammalian polycomb-group proteincomplexes. Mol Cell Biol 1998;18:3586–3595.
26. Hashimoto N, Brock HW, Nomura M, Kyba M, Hodgson J, Fujita Y,Takihara Y, Shimada K, Higashinakagawa T. RAE28, BMI1, and M33 aremembers of heterogeneous multimeric mammalian Polycomb groupcomplexes. Biochem Biophys Res Commun 1998;245:356–365.
27. Simon J, Chiang A, Bender W, Shimell MJ, O’Connor M. Elements of theDrosophila bithorax complex that mediate repression by Polycomb groupproducts. Dev Biol 1993;158:131–144.
28. Strutt H, Cavalli G, Paro R. Co-localization of Polycomb protein and GAGAfactor on regulatory elements responsible for the maintenance of home-otic gene expression. EMBO J 1997;16:3621–3632.
29. Brown JL, Mucci D, Whiteley M, Dirksen ML, Kassis JA. The DrosophilaPolycomb group gene pleiohomeotic encodes a DNA binding protein withhomology to the transcription factor YY1. Mol Cell 1998;1:1057–1064.
30. Mihaly J, Mishra RK, Karch F. A conserved sequence motif in Polycomb-response elements. Mol Cell 1998;1:1065–1066.
31. Gindhart JGJ, Kaufman TC. Identification of Polycomb and trithorax groupresponsive elements in the regulatory region of the Drosophila homeoticgene Sex combs reduced . Genetics 1995;139:797–814.
32. Chang YL, King BO, O’Connor M, Mazo A, Huang DH. Functionalreconstruction of trans regulation of the Ultrabithorax promoter by theproducts of two antagonistic genes, trithorax and Polycomb. Mol Cell Biol1995;15:6601–6612.
33. Mihaly J, Hogga I, Gausz J, Gyurkovics H, Karch F. In situ dissection of theFab-7 region of the bithorax complex into a chromatin domain boundaryand a Polycomb-response element. Development 1997;124:1809–1820.
34. Hagstrom K, Muller M, Schedl P. A Polycomb and GAGA dependentsilencer adjoins the Fab-7 boundary in the Drosophila bit horax complex.Genetics 1997;146:1365–1380.
35. Zink D, Paro R. Drosophila Polycomb-group regulated chromatin inhibitsthe accessibility of a trans-activator to its target DNA. EMBO J 1995;14:5660–5671.
36. Cavalli G, Paro R. The Drosophila Fab-7 chromosomal element conveysepigenetic inheritance during mitosis and meiosis. Cell 1998;93:505–518.
37. Crouse HV. The controlling element in sex chromosome behaviour inSciara. Genetics 1960;45:1429–1443.
38. Dorn R, Krauss V, Reuter G, Saumweber H. The enhancer of position-effect variegation of Drosophila, E(var)3–93D, codes for a chromatinprotein containing a conserved domain common to several transcriptionalregulators. Proc Natl Acad Sci USA 1993;90:11376–11380.
39. Golic KG, Golic MM, Pimpinelli S. Imprinted control of gene activity inDrosophila. Curr Biol 1998;8:1273–1276.
40. Bartolomei MS, Tilghman SM. Genomic imprinting in mammals. Annu RevGenet 1997;1997:493–525.
41. Albrecht U, Sutcliffe JS, Cattanach BM, Beechey CV, Armstrong D,Eichele G, Beaudet AL. Imprinted expression of the murine Angelmansyndrome gene, Ube3A, in hippocampal and Purkinje neurons. Nat Genet1997;17:75–78.
42. Xu Y, Goodyer CG, Deal C, Polychronakos C. Functional polymorphism inthe parental imprinting of the human IGF2R gene. Biochem Biophys ResCommun 1993;197:747–754.
43. Ainscough JF-X, Surani MA. Organization and control of imprinted genes:the common features. In: Russo VEA, Martienssen RA, Riggs AD, editors.Epigenetic mechanisms of gene regulation. Plainview, NY: Cold SpringHarbor Laboratory Press; 1996. p 173–194.
44. Buiting K, Saitoh S, Gross S, Dittrich B, Schwartz S, Nicholls RD,Horsthemke B. Inherited microdeletions in the Angelman and Prader-Willisyndromes define an imprinting centre on human chromosome 15. NatGenet 1995;9:395–400.
45. Thorvaldsen JL, Duran KL, Bartolomei MS. Deletion of the H19 differen-tially methylated domain results in loss of imprinted expression of H19 andIgf2. Genes Dev 1998;12:3693–3702.
46. Tremblay KD, Duran KL, Bartolomei MS. A 58 2-kilobase-pair region of theimprinted mouse H19 gene exhibits exclusive paternal methylation through-out development. Mol Cell Biol 1997;17:4322–4329.
47. Zeschnigk M, Schmitz B, Dittrich B, Buiting K, Horsthemke B, Doerfler W.Imprinted segments in the human genome: different DNA methylationpatterns in the Prader-Willi/Angelman syndrome region as determined bythe genomic sequencing method. Hum Mol Genet 1997;6:387–395.
48. Li E, Beard C, Jaenisch R. Role for DNA methylation in genomicimprinting. Nature 1993;366:362–365.
49. Caspary T, Cleary MA, Baker CC, Guan X-J, Tilghman SM. Multiplemechanisms regulate imprinting of the mouse distal chromosome 7 genecluster. Mol Cell Biol 1998;18:3466–3474.
50. Lyko F, Brenton JD, Surani MA, Paro R. An imprinting element from themouse H19 locus functions as a silencer in Drosophila. Nat Genet1997;16:171–174.
51. Lyko F, Buiting K, Horsthemke B, Paro R. Identification of a silencingelement in the human 15q11-q13 imprinting center by using transgenicDrosophila. Proc Natl Acad Sci USA 1998;95:1698–1702.
52. Ferguson-Smith AC, Sasaki H, Cattanach BM, Surani MA. Parental-origin-specific epigenetic modification of the mouse H19 gene. Nature 1993;362:751–755.
53. Bartolomei MS, Webber AL, Brunkow ME, Tilghman SM. Epigeneticmechanisms underlying the imprinting of the mouse H19 gene. GenesDev 1993;7:1663–1673.
54. Feil R. Parental chromosome-specific chromatin conformation in theimprinted U2af1-rs1 gene in the mouse. J Biol Chem 1997;272:20893–20900.
55. Svensson K, Mattsson R, James TC, Wentzel P, Pilartz M, MacLaughlin J,Miller SJ, Olsson T, Eriksson UJ, Ohlsson R. The paternal allele of the H19gene is progressively silenced during early mouse development: theacetylation status of histones may be involved in the generation ofvariegated expression patterns. Development 1998;125:61–69.
56. Banerjee S, Smallwood A. Chromatin modification of imprinted H19 genein mammalian spermatozoa. Mol Reprod Dev 1998;50:474–484.
57. Pirrotta V, Rastelli L. white gene expression, repressive chromatin do-mains and homeotic gene regulation. BioEssays 1994;16:549–556.
58. Henikoff S. Dosage-dependent modification of position-effect variegationin Drosophila. BioEssays 1996;18:401–409.
59. Dao D, Frank D, Quian N, O’Keefe D, Vosatka RJ, Walsh CP, Tycko B.IMPT1, an imprinted gene similar to polyspecific transporter and multi-drug resistance genes. Hum Mol Genet 1998;7:597–608.
60. Saitoh S, Buiting K, Rogan PK, Buxton JL, Driscoll DJ, Arnemann J, KonigR, Malcolm S, Horsthemke B, Nicholls RD. Minimal definition of theimprinting center and fixation of a chromosome 15q11–13 epigenotype byimprinting mutations. Proc Natl Acad Sci USA 1996;93:7811–7815.
61. Ainscough JF, Koide T, Tada M, Barton S, Surani MA. Imprinting of Igf2and H19 from a 130 kb YAC transgene. Development 1997;124:3621–3632.
62. Wutz A, Smrzka OW, Schweifer N, Schellander K, Wagner EF, Barlow DP.Imprinted expression of the Igf2r gene depends on an intronic CpGisland. Nature 1997;389:745–749.
Review articles
BioEssays 21.10 831

63. Kitsberg D, Selig S, Brandeis M, Simon I, Keshet I, Driscoll DJ, NichollsRD, Cedar H. Allele-specific replication timing of imprinted gene regions.Nature 1993;364:459–463.
64. Barlow DP. Competition—a common motif for the imprinting mechanism?EMBO J 1997;16:6899–6905.
65. Brenton JD, Ainscough JF-X, Lyko F, Paro R, Surani MA. Imprinting andgene silencing in mice and Drosophila. In: Chadwick DJ, Cardew G,editors. Epigenetics. Chichester: Wiley; 1998. p 233–250.
66. Szabo P, Mann JR. Biallelic expression of imprinted genes in the mousegerm line: implications for erasure, establishment, and mechanisms ofgenomic imprinting. Genes Dev 1995;9:1857–1868.
67. Szabo PE, Mann JR. Allele-specific expression and total expression levelsof imprinted genes during early mouse development: implications forimprinting mechanisms. Genes Dev 1995;9:3097–3108.
68. Dittrich B, Buiting K, Korn B, Rickard S, Buxton J, Saitoh S, Nicholls RD,Poustka A, Winterpacht A, Zabel B, Horsthemke B. Imprint switching onhuman chromosome 15 may involve alternative transcripts of the SNRPNgene. Nat Genet 1996;14:163–170.
69. Hark AT, Tilghman SM. Chromatin conformation of the H19 epigeneticmark. Hum Mol Genet 1998;7:1979–1985.
70. Khosla S, Aitchison A, Gregory R, Allen ND, Feil R. Parental allele-specificchromatin configuration in a boundary-imprinting-control element up-stream of the mouse H19 gene. Mol Cell Biol 1999;19:2556–2566.
71. Schweizer J, Zynger D, Francke U. In vivo nuclease hypersensitivitystudies reveal multiple sites of parental origin-dependent differentialchromatin conformation in the 150 kb SNRPN transcription unit. Hum MolGenet 1999;8:555–566.
Review articles
832 BioEssays 21.10





























