Embed Size (px)
Citation preview

Chemical and Immunochemical Detection of 8-HalogenatedDeoxyguanosines at Early Stage Inflammation*
Received for publication, August 10, 2009, and in revised form, December 28, 2009 Published, JBC Papers in Press, January 15, 2010, DOI 10.1074/jbc.M109.054213
Takashi Asahi‡, Hajime Kondo‡, Mitsuharu Masuda§, Hoyoku Nishino¶, Yasuaki Aratani�, Yuji Naito**,Toshikazu Yoshikawa**, Shinsuke Hisaka‡, Yoji Kato‡‡1, and Toshihiko Osawa‡
From the ‡Laboratory of Food and Biodynamics, Graduate School of Bioagricultural Sciences, Nagoya University,Nagoya 464-8601, Japan, the §Department of Health Sciences and Preventive Medicine, Kyoto Prefectural University of Medicine,Kyoto 602-8566, Japan, the ¶Cancer Center, Kyoto Prefectural University of Medicine, Kyoto 602-8566, Japan, the �Department ofGenome System Science, Graduate School of Nanobioscience, Yokohama City University, Kanazawa-ku, Yokohama 236-0027, Japan,**Molecular Gastroenterology and Hepatology, Graduate School of Medical Science, Kyoto Prefectural University of Medicine,Kyoto 602-8566, Japan, and the ‡‡School of Human Science and Environment, University of Hyogo, Himeji 670-0092, Japan
Myeloperoxidase (MPO) generates reactive halogenating spe-cies that can modify DNA. The aim of this study was to investi-gate the formation of 8-halogenated 2�-deoxyguanosines (8-halo-dGs) during inflammatory events. 8-Bromo-2�-dG (8-BrdG)and 8-chloro-2�-dG (8-CldG) were generated by treatment ofMPO with hydrogen peroxide at physiological concentrationsof Cl� and Br�. The formation of 8-halo-dGs with other oxida-tive stress biomarkers in lipopolysaccharide-treated rats wasassessed by liquid chromatography tandem mass spectrometryand immunohistochemistry using a novel monoclonal antibody(mAb8B3) to 8-BrdG-conjugated keyhole limpet hemocyanin.The antibody recognized both 8-BrdG and 8-CldG. In the liverof lipopolysaccharide-treated rats, immunostaining for 8-halo-dGs, halogenated tyrosines, and MPO were increased at 8 h,whereas those of 8-oxo-2�-dG (8-OxodG) and 3-nitrotyrosinewere increased at 24 h. Urinary excretion of both 8-CldG and8-BrdG was also observed earlier than those of 8-OxodG andmodified tyrosines (3-nitrotyrosine, 3-chlorotyrosine, and 3-bromotyrosine). Moreover, the levels of the 8-halo-dGs in urinefromhumandiabetic patientswere 8-fold higher than in healthysubjects (n � 10, healthy and diabetic, p < 0.0001), whereasthere was a moderate difference in 8-OxodG between the twogroups (p < 0.001). Interestingly, positive mAb8B3 antibodystaining was observed in liver tissue from hepatocellular carci-noma patients but not in liver tissue from human cirrhosispatients. These data suggest that 8-halo-dGs may be potentialbiomarkers of early inflammation.
Oxidative damage is implicated in the pathogenesis of manydiseases including atherosclerosis and cancer (1–4). Involve-ment of myeloperoxidase (MPO),2 a heme protein secreted by
activated leukocytes such as neutrophils andmonocytic cells, isa potential cause of oxidative damage during inflammation. Atsites of inflammation, activated leukocytes play a major role inhost defense against microorganisms using oxidants such asHOCl and HOBr. However, excessive oxidant release can alsoinduce damage; DNA bases (5–8) and proteins and lipids(9–12), for instance, are putative targets of oxidants.At plasma halide concentrations (100 mM Cl�, 20–100 �M
Br�) (13), MPO converts HOCl to HOBr using Br� as a halideexchange (14). HOBr is also generated by eosinophil peroxidaseusing H2O2 and Br�, such as by the MPO-H2O2-Cl� system.Because MPO catalyzes halogenation, oxidation, and nitration(15–17),MPO and its productmay be important general mark-ers for evaluating oxidative damage to the body during inflam-mation. Halogenation is a key reaction for the formation of anMPO activity marker because the presence of halogen in themarker molecule directly indicates a contribution of the halo-genating species at the site of oxidative damage. Moreover, noother halogenating pathways have been reported except forMPO and eosinophil peroxidase-derived damage. Therefore,halogenated products have the potential to be specific markersof inflammation.With respect to halogenation of nucleic acid, bromination of
2�-deoxycytidine (dC) and uracil has been reported in detail(18–20). At physiological conditions, 5-bromo-dC was gener-ated by human eosinophiles, whereas 5-bromouracil wasdetected by gas chromatography/mass spectrometry in theinflammatory tissue of human subjects suffering from a varietyof bacterial infections (19, 20). In human atherosclerotic tissue,5-chlorouracil and 5-bromouracil were generated by MPO (3).An antibody against N4,5-dichloro-deoxycytidine (N4,5-di-CldC) was developed and used for immunohistochemistry inlipopolysaccharide (LPS)-treated mice. In that inflammationmodel, immunopositive N4,5-diCldC staining was observed inthe liver (21). Chlorination of deoxyguanosine (dG) by expo-* This work was supported by a research grant from the Ministry of Education,
Culture, Sports, Science, and Technology, by the COE Program in the 21stCentury in Japan, and by research fellowships of the Japan Society for thePromotion of Science.
1 To whom correspondence should be addressed: School of Human Scienceand Environment, University of Hyogo, Himeji 670-0092, Japan. Tel.: 81-79-292-9413; Fax: 81-79-293-5710; E-mail: [email protected].
2 The abbreviations used are: MPO, myeloperoxidase; dG, 2�-deoxy-guanosine; N4,5-diCldC, N4,5-dichloro-deoxycytidine; 8-CldG, 8-chloro-2�-deoxyguanosine; 8-BrdG, 8-bromo-2�-deoxyguanosine; HPLC, highpressure liquid chromatography; LC-MS/MS, LC tandem mass spec-
trometry; LPS, lipopolysaccharide; 8-OxodG, 8-hydroxy-2�-deox-yguanosine; BSA, bovine serum albumin; KLH, keyhole limpet hemocy-anin; suc-8-BrdG, 5�-succinyl-8-BrdG; PBS, phosphate-buffered saline;ELISA, enzyme-linked immunosorbent assay; Tg, thymidine glycol;3-NO2Tyr, 3-nitrotyrosine; TBARS, thiobarbituric acid-reactive sub-stances; 8-halo, 8-halogenated; dC, 2�-deoxycytidine; hOGG1, human8-oxoguanine DNA glycosylase.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 285, NO. 12, pp. 9282–9291, March 19, 2010© 2010 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
9282 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

sure to HOCl was also reported (22). By contrast, there is littleinformation on modification of dG by brominating species.In the present study, we examined the patterns of bromi-
nated dG generation in vitro. We prepared the monoclonalantibody (mAb8B3) specific to 8-halogenated dGs (8-chloro-2�-deoxyguanosine (8-CldG) and 8-bromo-2�-deoxyguanosine(8-BrdG)). Using that antibody and liquid chromatography tan-dem mass spectrometry (LC-MS/MS), our results demon-strated facile formation of 8-halo-dGs in the liver and urine ofrats administered LPS. Furthermore, with respect to the orderofmodification of dG, halogenation occurred prior to oxidationand nitration, which may be useful for evaluation of the pro-gress of inflammatory diseases.
EXPERIMENTAL PROCEDURES
Materials
2�-Deoxynucleosides, DNase I, nuclease P1, 8-oxo-2�-deoxy-guanosine (8-OxodG), 3,3�,5,5�-tetramethylbenzidine, bovineserum albumin (BSA), LPS from Escherichia coli, and 3,5-di-aminobenzoic acid were obtained from Sigma-Aldrich. Methi-onine, taurine, 4-aminoantipyrine, NaOCl, metaphosphoricacid, and alkaline phosphatase were obtained fromWako PureChemicals Industries. Horseradish peroxidase-labeled goatanti-mouse IgG was obtained from Cappel. Anti-MPO poly-clonal antibody was obtained fromDAKO.HOBrwas preparedfrom an equimolar solution of NaOCl and KBr as described(20). 15N5-dG was obtained from Spectra. Keyhole limpethemocyanin (KLH) was obtained from Pierce.
Reaction Conditions for dG Modification by Myeloperoxidase
As typical conditions, 1 mM dG, 20 nM MPO, 100 �M H2O2,100 mM NaCl, and 100 �M NaBr were reacted for 60 min at37 °C in 50 mM phosphate buffer (pH 7.4). The reaction wasterminated by adding catalase to scavenge H2O2. Each compo-nent was varied as described in the text. The formation of bro-minated dG was quantified by LC-MS/MS, as described below.
Preparation of 8-CldG, 8-BrdG, and 8-OxodG and TheirInternal Standards
Modified dGs and their stable isotopic internal standardswere prepared as follows: (a) For 8-CldG, dG (2 mM) was sup-plemented with nicotine (20 �M) in 50 mM phosphate buffer(pH 8.0). The reaction was initiated by addingNaOCl (1mM) tothe reaction mixture at 37 °C for 1 h and terminated by addingmethionine (10 mM). (b) For 8-BrdG, dG (2 mM) was supple-mented with taurine (1 mM) in 50 mM phosphate buffer (pH7.4). The reaction was initiated by adding HOBr (1 mM) to thereaction mixture at 37 °C for 1 h and terminated by addingmethionine (10 mM). (c) For 8-OxodG, dG (2 mM) was supple-mented with ascorbate (1 mM) in 50 mM phosphate buffer (pH7.4). The reaction was initiated by adding H2O2 (1 mM) to thereaction mixture at 37 °C for 4 h. 8-CldG, 8-BrdG, and 8-OxodG were isolated by reverse phase high pressure liquidchromatography (HPLC) (Develosil C30, 20 � 250 mm) using10% acetonitrile containing 0.1% acetic acid at a flow rate of 6.0ml/min with monitoring at 280 nm. The internal standardswere prepared using 15N5-dG as a parent molecule by the same
methods as described above. The obtained compounds werecharacterized by LC-MS scanning the molecular ion peaks(8-CldG, m/z 302.1; 15N5-8-CldG, m/z 307.0; 8-BrdG, m/z345.9; 15N5-8-BrdG, m/z 350.9; 8-OxodG, m/z 284.0; 15N5-8-OxodG,m/z 288.9).
Conditions of Detection of 8-Modified dGs by LC-MS/MS
LC-MS/MS analyses were performed on an API 2000 triplequadrupolemass spectrometer (Applied Biosystems) through aTurboIonSpray source. Chromatography was carried out on aDevelosil ODS-HG-3 column (2.0 � 50 mm) using an Agilent1100 HPLC system.8-Halo-dGs—The chromatographic separation was per-
formed by a gradient elution as follows: 0–5 min, water con-taining 0.1% formic acid; 5–18 min, linear gradient to 26% ace-tonitrile containing 0.1% formic acid; 18–18.1 min, lineargradient to 100% acetonitrile containing 0.1% formic acid;18.1–24min, acetonitrile containing 0.1% formic acid; 24–24.1min, linear gradient to water containing 0.1% formic acid;24.1–34min, water containing 0.1% formic acid; flow rate� 0.2ml/min. The instrument response was optimized by infusionexperiments of the standard compounds using a syringe pumpat a flow rate of 5 �l/min. 8-Halo-dGs were detected usingelectrospray ionization tandem mass spectrometry in the mul-tiple reaction monitoring mode. Specific transitions used todetect products in the positive ionization mode were thosebetween the molecular cation of the products and the charac-teristic daughter ion formed from the loss of the 2�-deoxyribosemoiety.8-OxodG—The chromatographic separation was performed
by a gradient elution as follows: 0–5min, water containing 0.1%formic acid; 5–30 min, linear gradient to 50% acetonitrile con-taining 0.1% formic acid; 30–30.1 min, linear gradient to watercontaining 0.1% formic acid; 30.1–40 min, water containing0.1% formic acid; flow rate � 0.2 ml/min. Optimization of theinstrument response was performed as for 8-halo-dGs.
Preparation of the Monoclonal Antibody to 8-BrdG
To couple 8-BrdG to protein, the 5�-succinyl-8-BrdG deriv-ative (suc-8-BrdG) was synthesized. Briefly, 8-BrdG (6.5 mg,18.9 �mol) and succinic anhydride (3.8 mg, 37.8 �mol) weredissolved in pyridine (1 ml), and the mixture was kept at roomtemperature with stirring. After 2 days, the same amount ofsuccinic anhydride was added, and the mixture was kept over-night at room temperature. The solution was then evaporated,and the residue was dissolved in methanol. Suc-8-BrdG wasisolated by reverse phase HPLC (Develosil C30, 8 � 250 mm)using 10% acetonitrile containing 0.1% acetic acid at a flow rateof 2.0 ml/min with monitoring at 280 nm. The obtained suc-8-BrdG (2.0 mg, yield 23.4%) was identified by 1H NMR andLC-MS measurements (m/z 330, 332). The carboxyl group ofthe obtained suc-8-BrdGwas conjugated to the amino group ofKLH or BSA by the carbodiimide procedure as described pre-viously (23). The conjugate of suc-8-BrdG and KLH (8-BrdG-KLH) (0.6mg/ml) was emulsified with an equal volume of com-plete Freund’s adjuvant. Six-week-old female Balb/c mice wereimmunizedwith 100�l of this emulsion intraperitoneally. After2 weeks, the mice were boosted with the 8-BrdG-KLH (0.2
8-Halogenated dG as Inflammatory Marker
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9283
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

mg/ml) emulsified with an equal volume of incompleteFreund’s adjuvant. In the final boost, 100 �l of the conjugate(0.5 mg/ml) in phosphate-buffered saline (PBS) was injectedintravenously. Three days after the final boost, the mouse waskilled, and the spleen was removed for fusion with P3/U1myeloma cells. The fusion was carried out by polyethylene gly-col, and the cells were cultured in hypoxanthine/aminopterin/thymidine selectionmedium. Five days after the fusion, the cul-ture supernatants of hybridomas obtained were screened byenzyme-linked immunosorbent assay (ELISA) using 8-BrdG-BSA and untreated BSA as the coating agents. The positivehybridomas against the 8-BrdG-BSA were cloned by the limit-ing dilutionmethod. After repeated screening and cloning, fourspecific clones were obtained. Among them, a clone (termedmAb8B3) was used in the following experiments because of itsspecificity and high ability for cell growth.
ELISA
The indirect noncompetitive ELISA procedure has beendescribed previously (24). Briefly, 100 �l of antigen in PBS wascoated in wells and kept at 4 °C overnight. After washing andblocking with 4% Block Ace (Dainihon Seiyaku), 100 �l of themAb8B3 (1/2500 dilution) in PBS containing 0.05% Tween 20(TPBS)was added, and thewells were incubated at 37 °C for 2 h.After washing, 100 �l of anti-mouse IgG goat peroxidase-la-beled antibody (1/5000 in TPBS) was added, and the wells incu-bated at 37 °C for 1 h. After washing, 100 �l of reaction buffer(1% (w/v) 3,3�,5,5�-tetramethylbenzidine with 0.04% H2O2 in40 mM citrate-phosphate buffer, pH 5.0) was added. The colordeveloping reactionwas stopped by the addition of 100�l of 1 N
H3PO4. The binding of the antibody to the antigen was evalu-ated by measuring the optical density at 450 nm.Competitive indirect ELISA was performed for estimating
the cross-reactivity of the low molecular weight compoundswith the antibody. The 8-BrdG-BSA conjugate (0.05 �g/well)was used as the coating antigen. For the competitive reaction,50�l of competitors in PBSweremixedwith an equal volumeofthe antibody (1/2500 dilution) in PBS containing 4% BSA. Thecompetitor solution was kept at 4 °C overnight, and 90�l of themixture was used as the primary antibody. The cross-reactivityof the antibody to the competitorswas expressed asB/B0, whereB is the amount of antibody bound in the presence andB0 in theabsence of the competitor.
Animal Experiments
Female Wistar rats (7 weeks old) were given an intraperi-toneal injection of LPS from E. coli (3 mg/kg body weight;Sigma) in PBS or PBS alone. The rats were sacrificed at 2, 6,12, 24, or 72 h after LPS/PBS, and the livers were immedi-ately frozen until analysis. Rat urine was collected every 24 hfrom the day before LPS treatment to 10 days after adminis-tration. MPO�/� mice were developed and maintained asdescribed previously (25). The mice were given an intraper-itoneal injection of LPS (1 mg/kg of body weight) asdescribed above. All of the sections were counterstainedwith hematoxylin.
Immunohistochemistry
Livers were collected, fixed with buffered formalin, andembedded in paraffin. In the case of livers from MPO�/� andcontrol C57BL/6 mice, the specimens were fixed with Bouin’sfluid. The 3-�m tissue sections were affixed to slides, the spec-imenswere deparaffinized, and antigen retrievalwas performedby microwave treatment in 0.01 M citrate buffer (pH 6.0) for 10min. After cooling the slides at room temperature for 20 min, 1N HCl was added for 30 min (for the primary antibody of mod-ified DNA). The sections were blocked with normal rabbitserum (DAKO) in 1% BSA/PBS for 30 min and washed threetimes with PBS; normal swine serum (DAKO) was used for theanti-MPOantibody. The primary antibody (1/100 dilution)wasreacted at 4 °C overnight. After three PBS washes, biotinylatedsecond anti-mouse IgG antibody (DAKO, 1/100 dilution) wasreacted at room temperature for 40 min. Color developmentwas achieved using the ABC-AP (alkaline phosphatase) kit(Vectastain) with a Vector Red commercial kit as the chromo-gen according to the manufacturer’s recommendation. Thespecimens were finally counterstained with hematoxylin. Anti-3-nitrotyrosine (3-NO2Tyr)antibodywaspreparedasdescribedpre-viously (26).Anti-thymineglycol (Tg) antibodyanddihalotyrosineantibody (27) were obtained from Nikken Zeil Co. Human sam-ples were obtained from surgical operations. The present studywas approved by the Ethics Committee of Kyoto Prefectural Uni-versity ofMedicine.
Extraction of 8-CldG, 8-BrdG, and 8-OxodG from Liver Tissue
Liver tissue resected at surgery was immediately placed inliquid nitrogen and frozen at�80 °C until analysis. The thawedtissue (100 mg of wet weight) was homogenized at 4 °C in 1 mlof extraction buffer (10 mM methionine, 4 M urea, 0.2 M NaCl,0.5% sodium N-lauroyl sarcosine, 10 mM EDTA, 10 mM Tris-HCl, pH 7.5). In brief, the homogenate was mixed with 20 �lof proteinase K (20 mg/ml) and kept at 55 °C for 8 h. RNase(Qiagen; 16 �l) was added and incubated for 2 min. After vor-texing for 15 s, 1 ml of phenol was mixed and gently shaken for30min. Themixture was centrifuged for 10min at 3,000 rpm at4 °C. DNA was collected with phenol and chloroform/isoamy-lalcohol mixture according to a conventional protocol. Prior tothe enzymatic digestion of DNA, the buffer was supplementedwith isotope-labeled internal standards (15N5-labeled dG, 15N5-8-CldG, 15N5-8-BrdG, and 15N5-8-OxodG).DNAwas extractedas described (22, 28). Enzymatic digestion of DNA with DNaseI, nuclease P1, and alkaline phosphatase was performed asreported previously (29). After digestion, the reactionmixtureswere filtered throughultrafreeMCmembrane (nominalmolec-ular weight limit, 5000; Millipore) by centrifugation (10,000rpm) to remove enzymes. 8-CldG, 8-BrdG, and 8-OxodG of thefiltrates were fractionated by reverse phase HPLC, and 10 �l ofthe samples was injected for LC-MS/MS as described above.The multiple reaction monitoring for each compound was:15N5-8-CldG (m/z 307.0 3 190.9), 8-CldG (m/z 302.1 3185.9), 15N5-8-BrdG (m/z 350.93 235.0), 8-BrdG (m/z 345.93229.9), 15N5-8-OxodG (m/z 288.93 173.1), and 8-OxodG (m/z284.03 168.1), respectively.
8-Halogenated dG as Inflammatory Marker
9284 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from
MPO Activity of Rat Liver Tissue
Tissues were homogenized in 0.5% hexadecyltrimethyl-am-monium bromide, 50 mM KH2PO4 (pH 6) and centrifuged at10,000 rpm at 4 °C for 20 min. After 5 min, supernatant wasadded to the mixture of 150 �l of aqueous 2 mM H2O2 and 130�l of aqueous 25 mM 4-aminoantipyrine/1% phenol. The solu-tion was then reacted for 5 min. The absorbance at 510 nmwasmeasured (A5). Instead of 2mMH2O2/H2O, water was used as astandard (A0). TheMPOactivity was assessed by subtractingA0from A5 and normalized to the liver weight.
Measurement of Thiobarbituric Acid-reactive Substances(TBARS) in Rat Liver Tissue
The lipid peroxidation levels were evaluated by measure-ment of the TBARS as described previously (30). In brief, eachliver was homogenized in aqueous 1.15%KClwith 1% butylatedhydroxytoluene. After centrifugation at 13,000 rpm at 4 °C for10 min, a portion of supernatant was diluted 50� with 1.15%KCl to measure the protein concentration. The supernatant(200 �l) was added to 225 �l of 20% acetic acid and 225 �l of0.8% thiobarbituric acid and further incubated at 100 °C for 1 h.After cooling with water, the supernatant was centrifuged at4,000 rpm at 4 °C for 10 min, and the absorption was measuredat 532 nm. TBARS data were normalized to the proteinconcentration.
GSH of Rat Liver Tissue
The liver was homogenized in 0.1 M phosphate buffer con-taining 5mMEDTA and 1% butylated hydroxytoluene (pH 7.4).Homogenate was diluted 50� with buffer. A 100-�l volume of25 mM metaphosphoric acid was added to the sample (400 �l)and was ultracentrifuged at 47,000 rpm at 4 °C for 30 min. A50-�l volume of supernatant was reacted with 50 �l of 1 mg/mlo-phthalaldehyde in 900 �l of buffer for 15 min. Fluorescencewas measured at an excitation of 355 nm and an emission of460 nm.
Creatinine Assay
Urinary creatinine was determined by the Creatinine Test(WAKO) according to the manufacturer’s recommendation.
Analysis of Urinary 8-Modified dG and Modified TyrosinesUsing LC-MS/MS
Humanurinewas collected as a spot sample after approval bythe Nakatsugawa Municipal Hospital committee (31). The agerange was 56.8 � 10.6 in healthy patients and 56.9 � 11.0 indiabetic patients. Therewere fivemales and five females in eachgroup. For estimation of modified dGs, urine was supple-mented with each internal standard (15N5-8-CldG, 15N5-8-BrdG, and 15N5-8-OxodG). After centrifugation at 12,000 rpmat 4 °C for 10 min, the supernatant was diluted 10� with waterand then fractionated. Partial fractionations of 8-modified dGwere performed by reverse phase HPLC as described above.The samples (10 �l) were analyzed using LC-MS/MS asdescribed above. Quantification of urinary modified tyrosines,3-bromotyrosine, 3-chlorotyrosine, and 3-NO2Tyr, was per-formed as described previously (31).
Statistics
Statistical significance of the intergroup differences ofmeansfor multiple groups were determined using the Student New-
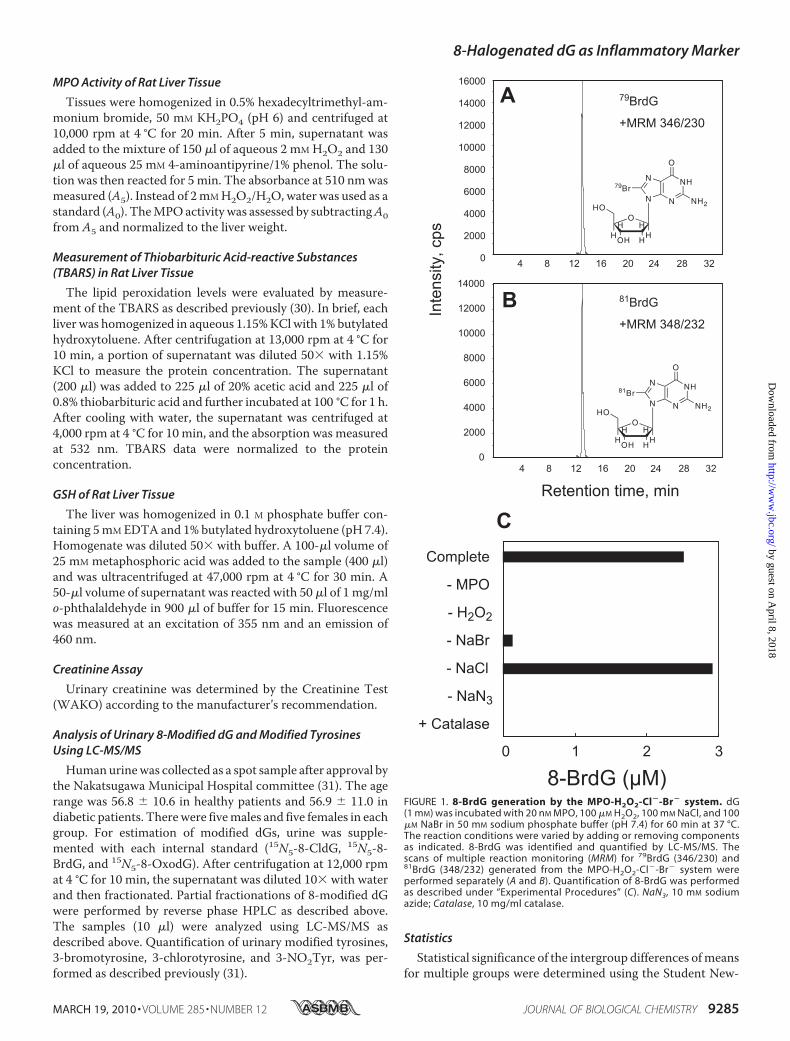
FIGURE 1. 8-BrdG generation by the MPO-H2O2-Cl�-Br� system. dG(1 mM) was incubated with 20 nM MPO, 100 �M H2O2, 100 mM NaCl, and 100�M NaBr in 50 mM sodium phosphate buffer (pH 7.4) for 60 min at 37 °C.The reaction conditions were varied by adding or removing componentsas indicated. 8-BrdG was identified and quantified by LC-MS/MS. Thescans of multiple reaction monitoring (MRM) for 79BrdG (346/230) and81BrdG (348/232) generated from the MPO-H2O2-Cl�-Br� system wereperformed separately (A and B). Quantification of 8-BrdG was performedas described under “Experimental Procedures” (C). NaN3, 10 mM sodiumazide; Catalase, 10 mg/ml catalase.
8-Halogenated dG as Inflammatory Marker
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9285
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from
man-Keuls multiple comparisons test after one-way analysisof variance to determine variations among the group means,followed by Bartlett’s test to determine the homogeneity ofvariance.
RESULTS
Reaction Requirement for 8-BrdG Production by the MPO-H2O2-Cl�-Br� System—We characterized the bromination ofdGbyMPOusing LC-MS/MS in vitro. During the incubation of
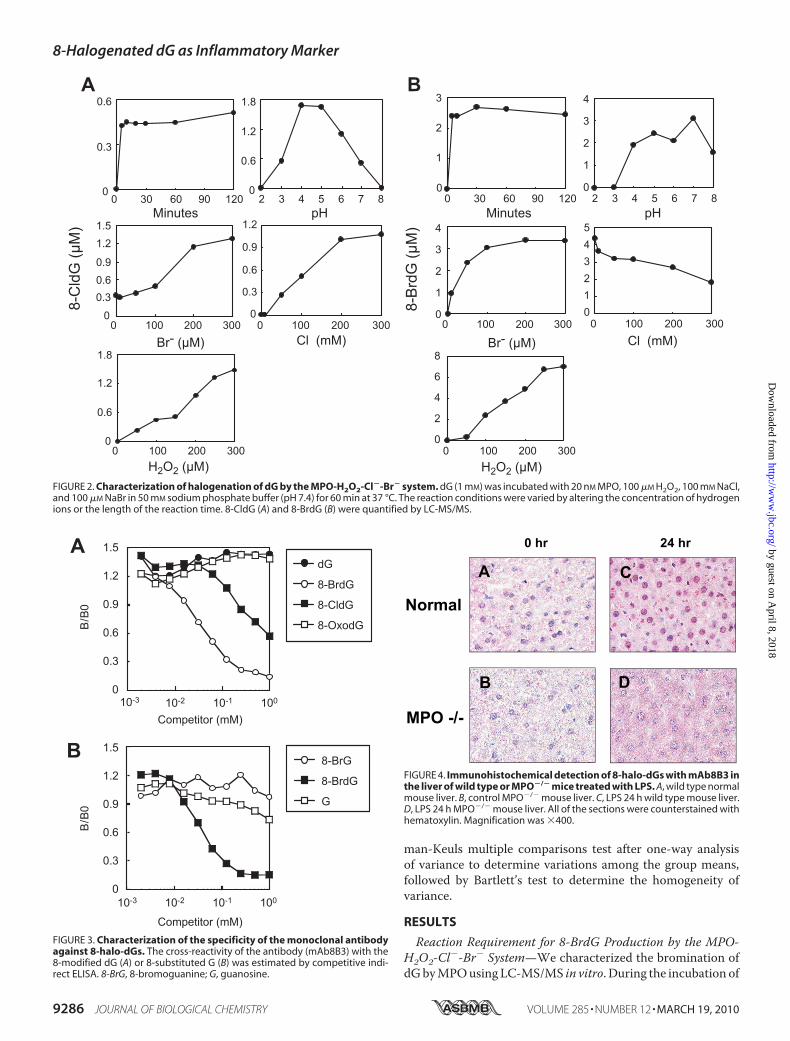
FIGURE 2. Characterization of halogenation of dG by the MPO-H2O2-Cl�-Br� system. dG (1 mM) was incubated with 20 nM MPO, 100 �M H2O2, 100 mM NaCl,and 100 �M NaBr in 50 mM sodium phosphate buffer (pH 7.4) for 60 min at 37 °C. The reaction conditions were varied by altering the concentration of hydrogenions or the length of the reaction time. 8-CldG (A) and 8-BrdG (B) were quantified by LC-MS/MS.
FIGURE 3. Characterization of the specificity of the monoclonal antibodyagainst 8-halo-dGs. The cross-reactivity of the antibody (mAb8B3) with the8-modified dG (A) or 8-substituted G (B) was estimated by competitive indi-rect ELISA. 8-BrG, 8-bromoguanine; G, guanosine.
FIGURE 4. Immunohistochemical detection of 8-halo-dGs with mAb8B3 inthe liver of wild type or MPO�/� mice treated with LPS. A, wild type normalmouse liver. B, control MPO�/� mouse liver. C, LPS 24 h wild type mouse liver.D, LPS 24 h MPO�/� mouse liver. All of the sections were counterstained withhematoxylin. Magnification was �400.
8-Halogenated dG as Inflammatory Marker
9286 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from
dG with MPO and H2O2 in a phosphate buffer containingNaBr, a novel product was observed on the mass chromato-gram, with a characteristic pattern of multiple reaction moni-toring of [M�H�]346/230 and [M�H�]348/232,with similarintensities (Fig. 1, A and B). The formation of the collision-induced fragment ion (m/z 230 and 232) indicated the libera-tion of the deoxyribosemoiety from themodified dG. TheM�2 isotopic pattern strongly suggested the incorporation of abromonium atom into dG (79Br and 81Br). Furthermore, thismass spectra pattern was consistent with 8-BrdG, the predom-inant reaction product from dG and HOBr. Therefore, theproduct was identified as 8-BrdG.The formation of 8-BrdG required MPO and H2O2 and
was blocked by adding catalase, a scavenger of H2O2 (Fig.1C). The reaction could not be completely prevented even inthe absence of NaBr, likely because the NaCl in the bufferwas contaminated with Br� (6). The heme enzyme inhibitor,NaN3, inhibited the reaction. These results demonstrate thatbromination of dG by MPO requires the active enzyme, Br�,and H2O2.The effects of varying the reaction conditions on the gener-
ation of 8-BrdG and 8-CldG can be seen in Fig. 2. As a basalcondition, dG (1 mM) was incubated with 20 nM MPO, 100 �M
H2O2, 100 mM NaCl, and 100 �M NaBr in 50 mM phosphatebuffer (pH 7.4). Enzymatic bromination increased with H2O2concentration up to 300 �M. At 100 �M of Br�, which is thephysiological concentration of Br�, the yield of 8-BrdG reacheda plateau. Both halogenations were completed within 30 min,and the optimal pH for 8-BrdG generation was �7. The pro-duction of 8-BrdG was decreased with increasing concentra-tion of Cl�, suggesting a competitive reaction between Cl� andBr� with MPO.Preparation andCharacterization ofMonoclonalAntibody to
8-BrdG—We prepared a monoclonal antibody to 8-BrdG(mAb8B3) using 8-BrdG-conjugated KLH as an immunogen(Fig. 3). Synthetic 8-BrdGwas chemically modified for succiny-lation, and the succinyl moiety was then coupled to a carrier
protein, KLH, as described previ-ously (24). Although the resultingantibody bound both 8-BrdG and8-CldG, the reactivity against8-BrdG was �10-fold higher thanfor 8-CldG. The antibody showedlittle cross-reactivity against8-OxodG and 8-bromoguanosineand could therefore distinguishthe 8-substituted moiety of dGand deoxyribose. Thus, the anti-body was considered to be an anti-body to 8-halo-dGs.Chlorination is considered a con-
sequence of MPO, whereas bromi-nation is related to both MPO andeosinophil peroxidase. To confirmthe availability of the antibodyagainst MPO-derived halogenationin animals, we examined its immu-noreactivity in the liver of LPS-
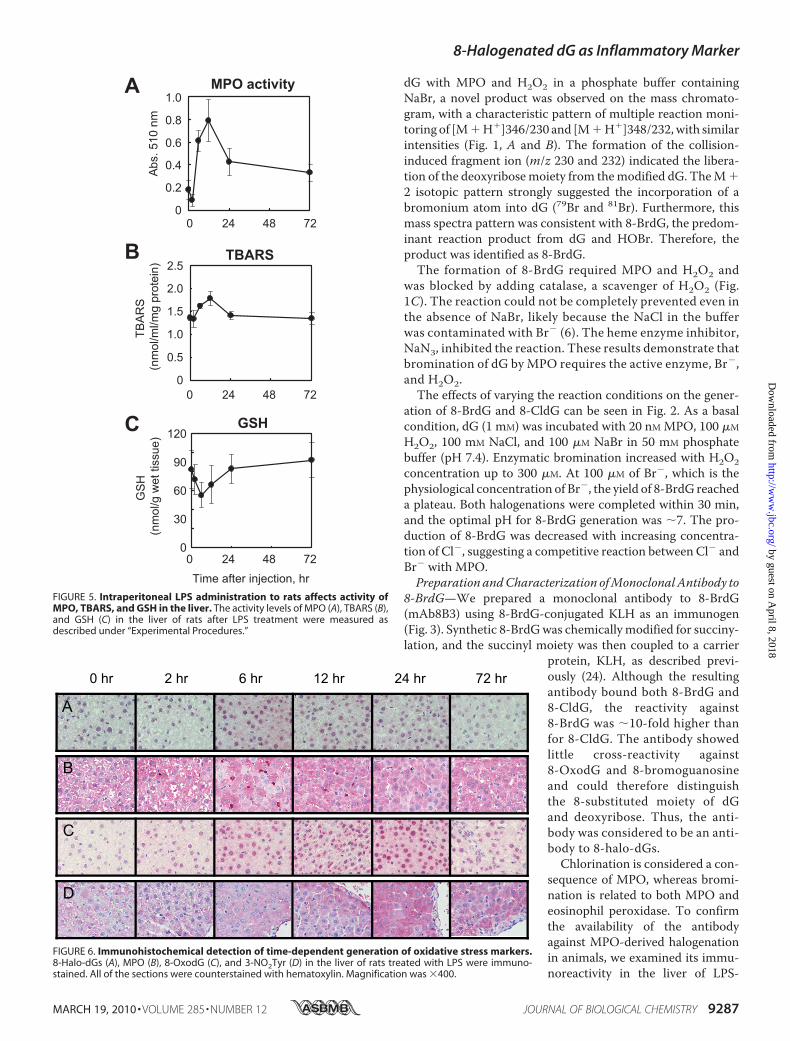
FIGURE 5. Intraperitoneal LPS administration to rats affects activity ofMPO, TBARS, and GSH in the liver. The activity levels of MPO (A), TBARS (B),and GSH (C) in the liver of rats after LPS treatment were measured asdescribed under “Experimental Procedures.”
FIGURE 6. Immunohistochemical detection of time-dependent generation of oxidative stress markers.8-Halo-dGs (A), MPO (B), 8-OxodG (C), and 3-NO2Tyr (D) in the liver of rats treated with LPS were immuno-stained. All of the sections were counterstained with hematoxylin. Magnification was �400.
8-Halogenated dG as Inflammatory Marker
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9287
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from
treated wild type andMPO�/� mice. Following LPS treatment,8-halo-dGs immunoreactivity was observed in the liver of wildtype mice but not MPO�/� mice (Fig. 4), suggesting that theantibody can be used for estimation ofMPO-derived modifica-tion. It is worthy to note that a weak signal in the cytoplasmicmatrix was also observed.Immunohistochemical Detection of Oxidative Stress Markers
in the Liver of LPS-treated Rats—Immunostaining for oxidativestressmarkers, including the novel antibody to 8-halo-dGs, wasexamined in the livers of rats treated with LPS. The levels ofMPO activity (i.e. a parameter of neutrophil infiltration) andTBARS (i.e. a parameter of lipid peroxidation) were signifi-cantly increased at 12 h after LPS treatment (Fig. 5, A and B).Moreover, a decrease of GSH (i.e. a parameter of reductantability) was observed at 6 h after LPS treatment (Fig. 5C).Immunopositive staining was observed in the livers of LPS-treated rats for all of the antibodies used, although the stainingpatterns varied between the antibodies (Fig. 6). An increase in8-halo-dGs and MPO immunoreactivity in the liver of LPS-treated animals was observed at 6 and 12 h, respectively (Fig. 6,A andB). By contrast, oxidized and nitrated products, 8-OxodGand 3-NO2Tyr, exhibited immunoreactivity at 12 and 24 h afterLPS. Although the antibody to 3-NO2Tyr was solely stained inthe cytosol, both halo-dGs and 8-OxodG were stained in boththe nuclei and a certain amount of the cytosol. A summaryof the various immunostainings can be seen in Table 1. N4,5-diCldC was previously detected by immunohistochemistry inLPS-treated mice (21). The time course of N4,5-diCldC immu-nostaining was consistent with that of 8-halo-dGs (Table 1). Tgis awell knownoxidized dT, and themonoclonal antibody toTgis commercially available. The time course of Tg immuno-staining was similar to that of 8-OxodG (Table 1). These datafurther suggest that halogenation of dG is an early and transientevent in tissue inflammation.Quantification of LPS-induced Time-dependent Generation
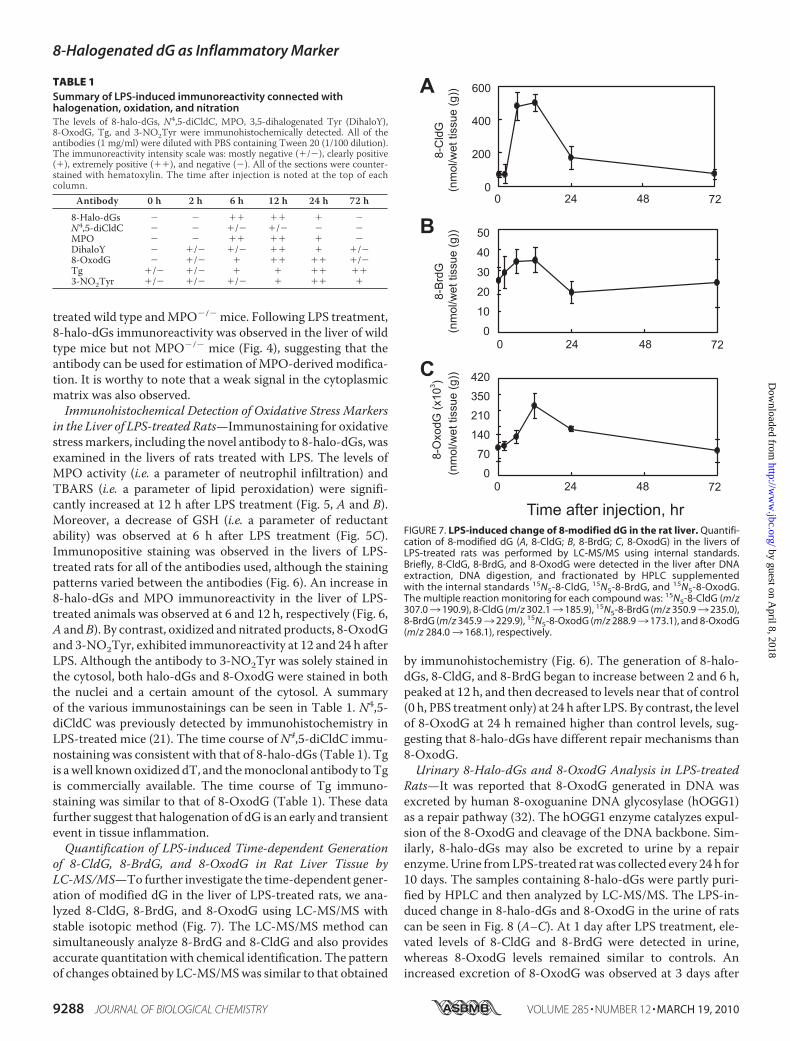
of 8-CldG, 8-BrdG, and 8-OxodG in Rat Liver Tissue byLC-MS/MS—To further investigate the time-dependent gener-ation of modified dG in the liver of LPS-treated rats, we ana-lyzed 8-CldG, 8-BrdG, and 8-OxodG using LC-MS/MS withstable isotopic method (Fig. 7). The LC-MS/MS method cansimultaneously analyze 8-BrdG and 8-CldG and also providesaccurate quantitationwith chemical identification. The patternof changes obtained by LC-MS/MSwas similar to that obtained
by immunohistochemistry (Fig. 6). The generation of 8-halo-dGs, 8-CldG, and 8-BrdG began to increase between 2 and 6 h,peaked at 12 h, and then decreased to levels near that of control(0 h, PBS treatment only) at 24 h after LPS. By contrast, the levelof 8-OxodG at 24 h remained higher than control levels, sug-gesting that 8-halo-dGs have different repair mechanisms than8-OxodG.Urinary 8-Halo-dGs and 8-OxodG Analysis in LPS-treated
Rats—It was reported that 8-OxodG generated in DNA wasexcreted by human 8-oxoguanine DNA glycosylase (hOGG1)as a repair pathway (32). The hOGG1 enzyme catalyzes expul-sion of the 8-OxodG and cleavage of the DNA backbone. Sim-ilarly, 8-halo-dGs may also be excreted to urine by a repairenzyme.Urine fromLPS-treated ratwas collected every 24 h for10 days. The samples containing 8-halo-dGs were partly puri-fied by HPLC and then analyzed by LC-MS/MS. The LPS-in-duced change in 8-halo-dGs and 8-OxodG in the urine of ratscan be seen in Fig. 8 (A–C). At 1 day after LPS treatment, ele-vated levels of 8-CldG and 8-BrdG were detected in urine,whereas 8-OxodG levels remained similar to controls. Anincreased excretion of 8-OxodG was observed at 3 days after
FIGURE 7. LPS-induced change of 8-modified dG in the rat liver. Quantifi-cation of 8-modified dG (A, 8-CldG; B, 8-BrdG; C, 8-OxodG) in the livers ofLPS-treated rats was performed by LC-MS/MS using internal standards.Briefly, 8-CldG, 8-BrdG, and 8-OxodG were detected in the liver after DNAextraction, DNA digestion, and fractionated by HPLC supplementedwith the internal standards 15N5-8-CldG, 15N5-8-BrdG, and 15N5-8-OxodG.The multiple reaction monitoring for each compound was: 15N5-8-CldG (m/z307.03 190.9), 8-CldG (m/z 302.13 185.9), 15N5-8-BrdG (m/z 350.93 235.0),8-BrdG (m/z 345.93 229.9), 15N5-8-OxodG (m/z 288.93 173.1), and 8-OxodG(m/z 284.03 168.1), respectively.
TABLE 1Summary of LPS-induced immunoreactivity connected withhalogenation, oxidation, and nitrationThe levels of 8-halo-dGs, N4,5-diCldC, MPO, 3,5-dihalogenated Tyr (DihaloY),8-OxodG, Tg, and 3-NO2Tyr were immunohistochemically detected. All of theantibodies (1 mg/ml) were diluted with PBS containing Tween 20 (1/100 dilution).The immunoreactivity intensity scale was: mostly negative (�/�), clearly positive(�), extremely positive (��), and negative (�). All of the sections were counter-stained with hematoxylin. The time after injection is noted at the top of eachcolumn.
Antibody 0 h 2 h 6 h 12 h 24 h 72 h
8-Halo-dGs � � �� �� � �N4,5-diCldC � � �/� �/� � �MPO � � �� �� � �DihaloY � �/� �/� �� � �/�8-OxodG � �/� � �� �� �/�Tg �/� �/� � � �� ��3-NO2Tyr �/� �/� �/� � �� �
8-Halogenated dG as Inflammatory Marker
9288 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from
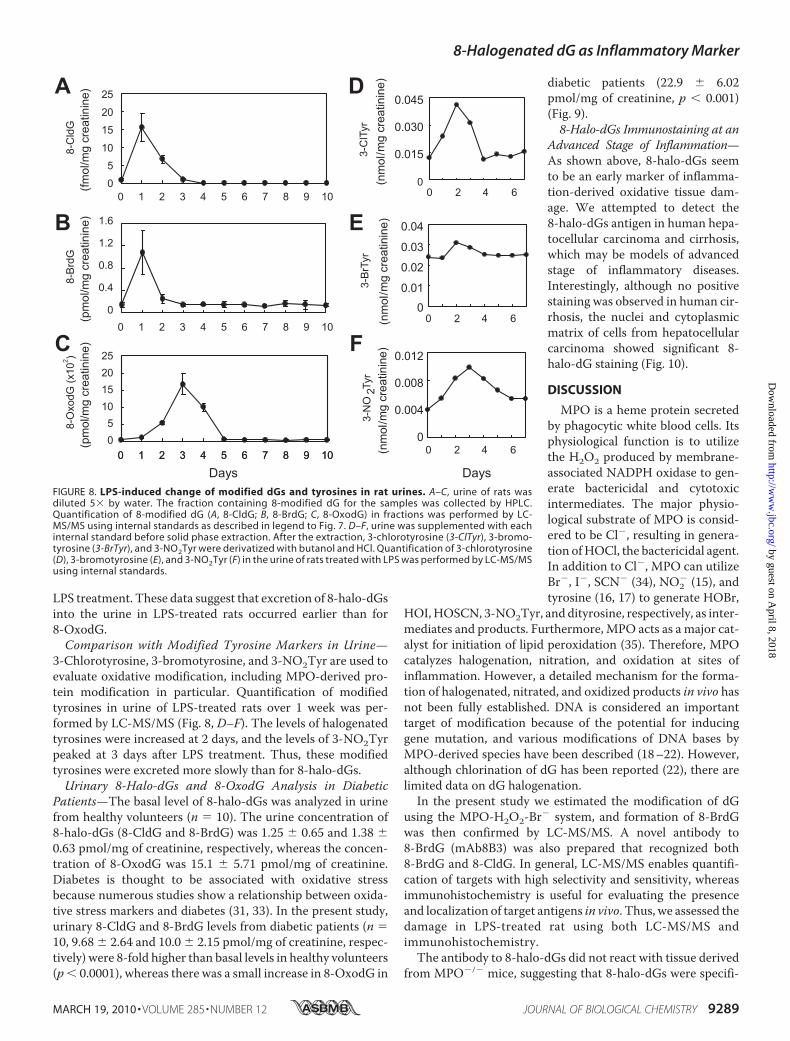
LPS treatment. These data suggest that excretion of 8-halo-dGsinto the urine in LPS-treated rats occurred earlier than for8-OxodG.Comparison with Modified Tyrosine Markers in Urine—
3-Chlorotyrosine, 3-bromotyrosine, and 3-NO2Tyr are used toevaluate oxidative modification, including MPO-derived pro-tein modification in particular. Quantification of modifiedtyrosines in urine of LPS-treated rats over 1 week was per-formed by LC-MS/MS (Fig. 8, D–F). The levels of halogenatedtyrosines were increased at 2 days, and the levels of 3-NO2Tyrpeaked at 3 days after LPS treatment. Thus, these modifiedtyrosines were excreted more slowly than for 8-halo-dGs.Urinary 8-Halo-dGs and 8-OxodG Analysis in Diabetic
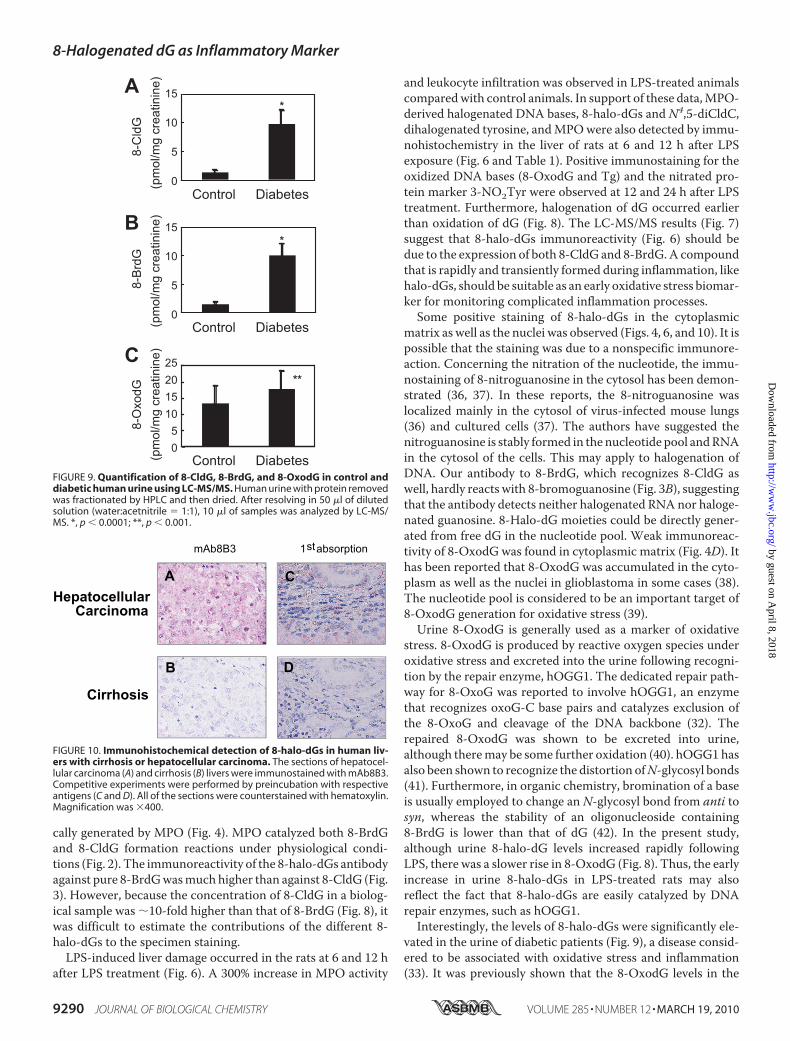
Patients—The basal level of 8-halo-dGs was analyzed in urinefrom healthy volunteers (n � 10). The urine concentration of8-halo-dGs (8-CldG and 8-BrdG) was 1.25 � 0.65 and 1.38 �0.63 pmol/mg of creatinine, respectively, whereas the concen-tration of 8-OxodG was 15.1 � 5.71 pmol/mg of creatinine.Diabetes is thought to be associated with oxidative stressbecause numerous studies show a relationship between oxida-tive stress markers and diabetes (31, 33). In the present study,urinary 8-CldG and 8-BrdG levels from diabetic patients (n �10, 9.68 � 2.64 and 10.0 � 2.15 pmol/mg of creatinine, respec-tively) were 8-fold higher than basal levels in healthy volunteers(p� 0.0001), whereas there was a small increase in 8-OxodG in
diabetic patients (22.9 � 6.02pmol/mg of creatinine, p � 0.001)(Fig. 9).8-Halo-dGs Immunostaining at an
Advanced Stage of Inflammation—As shown above, 8-halo-dGs seemto be an early marker of inflamma-tion-derived oxidative tissue dam-age. We attempted to detect the8-halo-dGs antigen in human hepa-tocellular carcinoma and cirrhosis,which may be models of advancedstage of inflammatory diseases.Interestingly, although no positivestaining was observed in human cir-rhosis, the nuclei and cytoplasmicmatrix of cells from hepatocellularcarcinoma showed significant 8-halo-dG staining (Fig. 10).
DISCUSSION
MPO is a heme protein secretedby phagocytic white blood cells. Itsphysiological function is to utilizethe H2O2 produced by membrane-associated NADPH oxidase to gen-erate bactericidal and cytotoxicintermediates. The major physio-logical substrate of MPO is consid-ered to be Cl�, resulting in genera-tion ofHOCl, the bactericidal agent.In addition to Cl�, MPO can utilizeBr�, I�, SCN� (34), NO2
� (15), andtyrosine (16, 17) to generate HOBr,
HOI,HOSCN, 3-NO2Tyr, and dityrosine, respectively, as inter-mediates and products. Furthermore, MPO acts as a major cat-alyst for initiation of lipid peroxidation (35). Therefore, MPOcatalyzes halogenation, nitration, and oxidation at sites ofinflammation. However, a detailed mechanism for the forma-tion of halogenated, nitrated, and oxidized products in vivo hasnot been fully established. DNA is considered an importanttarget of modification because of the potential for inducinggene mutation, and various modifications of DNA bases byMPO-derived species have been described (18–22). However,although chlorination of dG has been reported (22), there arelimited data on dG halogenation.In the present study we estimated the modification of dG
using the MPO-H2O2-Br� system, and formation of 8-BrdGwas then confirmed by LC-MS/MS. A novel antibody to8-BrdG (mAb8B3) was also prepared that recognized both8-BrdG and 8-CldG. In general, LC-MS/MS enables quantifi-cation of targets with high selectivity and sensitivity, whereasimmunohistochemistry is useful for evaluating the presenceand localization of target antigens in vivo. Thus, we assessed thedamage in LPS-treated rat using both LC-MS/MS andimmunohistochemistry.The antibody to 8-halo-dGs did not react with tissue derived
from MPO�/� mice, suggesting that 8-halo-dGs were specifi-
FIGURE 8. LPS-induced change of modified dGs and tyrosines in rat urines. A–C, urine of rats wasdiluted 5� by water. The fraction containing 8-modified dG for the samples was collected by HPLC.Quantification of 8-modified dG (A, 8-CldG; B, 8-BrdG; C, 8-OxodG) in fractions was performed by LC-MS/MS using internal standards as described in legend to Fig. 7. D–F, urine was supplemented with eachinternal standard before solid phase extraction. After the extraction, 3-chlorotyrosine (3-ClTyr), 3-bromo-tyrosine (3-BrTyr), and 3-NO2Tyr were derivatized with butanol and HCl. Quantification of 3-chlorotyrosine(D), 3-bromotyrosine (E), and 3-NO2Tyr (F) in the urine of rats treated with LPS was performed by LC-MS/MSusing internal standards.
8-Halogenated dG as Inflammatory Marker
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9289
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from
cally generated by MPO (Fig. 4). MPO catalyzed both 8-BrdGand 8-CldG formation reactions under physiological condi-tions (Fig. 2). The immunoreactivity of the 8-halo-dGs antibodyagainst pure 8-BrdGwasmuch higher than against 8-CldG (Fig.3). However, because the concentration of 8-CldG in a biolog-ical sample was �10-fold higher than that of 8-BrdG (Fig. 8), itwas difficult to estimate the contributions of the different 8-halo-dGs to the specimen staining.LPS-induced liver damage occurred in the rats at 6 and 12 h
after LPS treatment (Fig. 6). A 300% increase in MPO activity
and leukocyte infiltration was observed in LPS-treated animalscomparedwith control animals. In support of these data,MPO-derived halogenated DNA bases, 8-halo-dGs and N4,5-diCldC,dihalogenated tyrosine, andMPOwere also detected by immu-nohistochemistry in the liver of rats at 6 and 12 h after LPSexposure (Fig. 6 and Table 1). Positive immunostaining for theoxidized DNA bases (8-OxodG and Tg) and the nitrated pro-tein marker 3-NO2Tyr were observed at 12 and 24 h after LPStreatment. Furthermore, halogenation of dG occurred earlierthan oxidation of dG (Fig. 8). The LC-MS/MS results (Fig. 7)suggest that 8-halo-dGs immunoreactivity (Fig. 6) should bedue to the expression of both 8-CldG and 8-BrdG.A compoundthat is rapidly and transiently formed during inflammation, likehalo-dGs, should be suitable as an early oxidative stress biomar-ker for monitoring complicated inflammation processes.Some positive staining of 8-halo-dGs in the cytoplasmic
matrix as well as the nuclei was observed (Figs. 4, 6, and 10). It ispossible that the staining was due to a nonspecific immunore-action. Concerning the nitration of the nucleotide, the immu-nostaining of 8-nitroguanosine in the cytosol has been demon-strated (36, 37). In these reports, the 8-nitroguanosine waslocalized mainly in the cytosol of virus-infected mouse lungs(36) and cultured cells (37). The authors have suggested thenitroguanosine is stably formed in the nucleotide pool andRNAin the cytosol of the cells. This may apply to halogenation ofDNA. Our antibody to 8-BrdG, which recognizes 8-CldG aswell, hardly reacts with 8-bromoguanosine (Fig. 3B), suggestingthat the antibody detects neither halogenated RNA nor haloge-nated guanosine. 8-Halo-dG moieties could be directly gener-ated from free dG in the nucleotide pool. Weak immunoreac-tivity of 8-OxodG was found in cytoplasmic matrix (Fig. 4D). Ithas been reported that 8-OxodG was accumulated in the cyto-plasm as well as the nuclei in glioblastoma in some cases (38).The nucleotide pool is considered to be an important target of8-OxodG generation for oxidative stress (39).Urine 8-OxodG is generally used as a marker of oxidative
stress. 8-OxodG is produced by reactive oxygen species underoxidative stress and excreted into the urine following recogni-tion by the repair enzyme, hOGG1. The dedicated repair path-way for 8-OxoG was reported to involve hOGG1, an enzymethat recognizes oxoG-C base pairs and catalyzes exclusion ofthe 8-OxoG and cleavage of the DNA backbone (32). Therepaired 8-OxodG was shown to be excreted into urine,although theremay be some further oxidation (40). hOGG1hasalso been shown to recognize the distortion ofN-glycosyl bonds(41). Furthermore, in organic chemistry, bromination of a baseis usually employed to change an N-glycosyl bond from anti tosyn, whereas the stability of an oligonucleoside containing8-BrdG is lower than that of dG (42). In the present study,although urine 8-halo-dG levels increased rapidly followingLPS, there was a slower rise in 8-OxodG (Fig. 8). Thus, the earlyincrease in urine 8-halo-dGs in LPS-treated rats may alsoreflect the fact that 8-halo-dGs are easily catalyzed by DNArepair enzymes, such as hOGG1.Interestingly, the levels of 8-halo-dGs were significantly ele-
vated in the urine of diabetic patients (Fig. 9), a disease consid-ered to be associated with oxidative stress and inflammation(33). It was previously shown that the 8-OxodG levels in the
FIGURE 9. Quantification of 8-CldG, 8-BrdG, and 8-OxodG in control anddiabetic human urine using LC-MS/MS. Human urine with protein removedwas fractionated by HPLC and then dried. After resolving in 50 �l of dilutedsolution (water:acetnitrile � 1:1), 10 �l of samples was analyzed by LC-MS/MS. *, p � 0.0001; **, p � 0.001.
FIGURE 10. Immunohistochemical detection of 8-halo-dGs in human liv-ers with cirrhosis or hepatocellular carcinoma. The sections of hepatocel-lular carcinoma (A) and cirrhosis (B) livers were immunostained with mAb8B3.Competitive experiments were performed by preincubation with respectiveantigens (C and D). All of the sections were counterstained with hematoxylin.Magnification was �400.
8-Halogenated dG as Inflammatory Marker
9290 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 285 • NUMBER 12 • MARCH 19, 2010
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

urinewere affected by age, gender, and degree of disease (43). Inthe present study, urine 8-halo-dGswere not affected by genderin either healthy or diabetic patients (data not shown); however,as yet we have not assessed enough samples, and a moredetailed study is required. There was significant staining ofhepatocellular carcinoma cells with the anti-8-halo-dG anti-body (Fig. 10). This may relate to active gene transcription orreplication processes, which increase the chance ofmutation byreactive species or incorporation of a pooledmodified dG.Con-versely, there was no anti-8-halo-dGs immunostaining in livertissue from cirrhosis patients. The detection of MPO andHOCl-modified protein in the human liver, including thosewith cirrhosis, has been reported (44), although decreased pro-duction of reactive oxygen species by neutrophils in patientswith liver cirrhosis was also demonstrated (45). This reductionin reactive oxygen species may cause a reduced halogenatingability in the cirrhotic liver, because MPO requires hydrogenperoxide as a substrate to catalyze the production of halogenat-ing species. In addition, we have found that 8-BrdG was effec-tively generated by bromamines rather than HOBr itself invitro.3 Thus, intracellular amines including nucleotides couldact as halogenating species (as a carrier) in vivo.
In summary, we successively quantified the halogenated, oxi-dized, andnitrated products in the liver andurine of rats treatedwith LPS and suggest that halogenation might occur earlierthan nitration and oxidation in thismodel. The levels of urinary8-halo-dGs were relatively low compared with 8-OxodG butstill detectable even in control subjects. Furthermore, the levelsof urinary 8-halo-dGs in diabetic patients were higher than incontrol subjects. These data suggest that leukocyte-derivedhalogenated products such as 8-halo-dGs have potential asnovel and early biomarkers of oxidative stress for assessment ofinflammatory damage.
Acknowledgments—We thank Drs. Akihiro Yoshida (NakatsugawaMunicipal Hospital) and Michitaka Naito (Sugiyama UniversityGraduate School) for kindly supplying diabetic and control urine.
REFERENCES1. Hazen, S. L., and Heinecke, J. W. (1997) J. Clin. Invest. 99, 2075–20812. Ohshima, H., and Bartsch, H. (1994)Mutat. Res. 305, 253–2643. Takeshita, J., Byun, J., Nhan, T. Q., Pritchard, D. K., Pennathur, S.,
Schwartz, S. M., Chait, A., and Heinecke, J. W. (2006) J. Biol. Chem. 281,3096–3104
4. Weitzman, S. A., and Gordon, L. I. (1990) Blood 76, 655–6635. Henderson, J. P., Byun, J., and Heinecke, J. W. (1999) J. Biol. Chem. 274,
33440–334486. Henderson, J. P., Byun, J., Mueller, D. M., and Heinecke, J. W. (2001)
Biochemistry 40, 2052–20597. Shen, Z., Mitra, S. N., Wu,W., Chen, Y., Yang, Y., Qin, J., and Hazen, S. L.
(2001) Biochemistry 40, 2041–20518. Wiseman, H., and Halliwell, B. (1996) Biochem. J. 313, 17–299. Albert, C. J., Thukkani, A. K., Heuertz, R. M., Slungaard, A., Hazen, S. L.,
and Ford, D. A. (2003) J. Biol. Chem. 278, 8942–895010. Domigan, N.M., Charlton, T. S., Duncan,M.W.,Winterbourn, C. C., and
Kettle, A. J. (1995) J. Biol. Chem. 270, 16542–1654811. Hazen, S. L., Hsu, F. F., Mueller, D. M., Crowley, J. R., and Heinecke, J. W.
(1996) J. Clin. Invest. 98, 1283–1289
12. Masuda, M., Suzuki, T., Friesen, M. D., Ravanat, J. L., Cadet, J., Pignatelli,B., Nishino, H., and Ohshima, H. (2001) J. Biol. Chem. 276, 40486–40496
13. Thukkani, A. K., Albert, C. J.,Wildsmith, K. R.,Messner,M.C.,Martinson,B. D., Hsu, F. F., and Ford, D. A. (2003) J. Biol. Chem. 278, 36365–36372
14. Gaut, J. P., Yeh, G. C., Tran,H.D., Byun, J., Henderson, J. P., Richter, G.M.,Brennan, M. L., Lusis, A. J., Belaaouaj, A., Hotchkiss, R. S., and Heinecke,J. W. (2001) Proc. Natl. Acad. Sci. U.S.A. 98, 11961–11966
15. Eiserich, J. P., Hristova, M., Cross, C. E., Jones, A. D., Freeman, B. A.,Halliwell, B., and van der Vliet, A. (1998) Nature 391, 393–397
16. Heinecke, J. W. (2002) Toxicology 177, 11–2217. Heinecke, J. W., Li, W., Daehnke, H. L., 3rd, and Goldstein, J. A. (1993)
J. Biol. Chem. 268, 4069–407718. Byun, J., Henderson, J. P., and Heinecke, J. W. (2003) Anal. Biochem. 317,
201–20919. Henderson, J. P., Byun, J., Takeshita, J., and Heinecke, J. W. (2003) J. Biol.
Chem. 278, 23522–2352820. Henderson, J. P., Byun, J., Williams, M. V., McCormick, M. L., Parks,
W. C., Ridnour, L. A., and Heinecke, J. W. (2001) Proc. Natl. Acad. Sci.U.S.A. 98, 1631–1636
21. Kawai, Y., Morinaga, H., Kondo, H., Miyoshi, N., Nakamura, Y., Uchida,K., and Osawa, T. (2004) J. Biol. Chem. 279, 51241–51249
22. Badouard, C., Masuda, M., Nishino, H., Cadet, J., Favier, A., and Ravanat,J. L. (2005) J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 827, 26–31
23. Kato, Y., Mori, Y., Makino, Y., Morimitsu, Y., Hiroi, S., Ishikawa, T., andOsawa, T. (1999) J. Biol. Chem. 274, 20406–20414
24. Kato, Y., Makino, Y., and Osawa, T. (1997) J. Lipid Res. 38, 1334–134625. Aratani, Y., Koyama, H., Nyui, S., Suzuki, K., Kura, F., and Maeda, N.
(1999) Infect. Immun. 67, 1828–183626. Beckmann, J. S., Ye, Y. Z., Anderson, P. G., Chen, J., Accavitti, M. A., Tarpey,
M.M., andWhite, C. R. (1994) Biol. Chem. Hoppe-Seyler 375, 81–8827. Kato, Y., Kawai, Y.,Morinaga,H., Kondo,H., Dozaki,N., Kitamoto,N., and
Osawa, T. (2005) Free Radic. Biol. Med. 38, 24–3128. Toyokuni, S., Tanaka, T., Hattori, Y., Nishiyama, Y., Yoshida, A., Uchida,
K., Hiai, H., Ochi, H., and Osawa, T. (1997) Lab. Invest. 76, 365–37429. Huang, X., Powell, J., Mooney, L. A., Li, C., and Frenkel, K. (2001) Free
Radic. Biol. Med. 31, 1341–135130. Kobayashi, Y., Nakae, D., Akai, H., Kishida, H., Okajima, E., Kitayama,W.,
Denda, A., Tsujiuchi, T., Murakami, A., Koshimizu, K., Ohigashi, H., andKonishi, Y. (1998) Carcinogenesis 19, 1809–1814
31. Kato, Y., Dozaki, N., Nakamura, T., Kitamoto, N., Yoshida, A., Naito, M.,Kitamura, M., and Osawa, T. (2009) J. Clin. Biochem. Nutr. 44, 67–78
32. Bruner, S. D., Norman, D. P., and Verdine, G. L. (2000) Nature 403,859–866
33. Baynes, J. W., and Thorpe, S. R. (1999) Diabetes 48, 1–934. Furtmuller, P. G., Burner, U., and Obinger, C. (1998) Biochemistry 37,
17923–1793035. Zhang, R., Brennan, M. L., Shen, Z., MacPherson, J. C., Schmitt, D., Mo-
lenda, C. E., and Hazen, S. L. (2002) J. Biol. Chem. 277, 46116–4612236. Akaike, T., Okamoto, S., Sawa, T., Yoshitake, J., Tamura, F., Ichimori, K.,
Miyazaki, K., Sasamoto, K., and Maeda, H. (2003) Proc. Natl. Acad. Sci.U.S.A. 100, 685–690
37. Yoshitake, J., Akaike, T., Akuta, T., Tamura, F., Ogura, T., Esumi, H., andMaeda, H. (2004) J. Virol. 78, 8709–8719
38. Iida, T., Furuta, A., Kawashima, M., Nishida, J., Nakabeppu, Y., and Iwaki,T. (2001) Neuro. Oncol. 3, 73–81
39. Haghdoost, S., Sjolander, L., Czene, S., and Harms-Ringdahl, M. (2006)Free Radic. Biol. Med. 41, 620–626
40. Suzuki, T., and Ohshima, H. (2002) FEBS Lett. 516, 67–7041. Bhatnagar, S. K., Bullions, L. C., and Bessman, M. J. (1991) J. Biol. Chem.
266, 9050–905442. Fabrega, C., Macías, M. J., and Eritja, R. (2001) Nucleosides Nucleotides
Nucleic Acids 20, 251–26043. Kimura, S., Yamauchi, H., Hibino, Y., Iwamoto,M., Sera, K., andOgino, K.
(2006) Basic Clin. Pharmacol. Toxicol. 98, 496–50244. Brown, K. E., Brunt, E. M., and Heinecke, J. W. (2001) Am. J. Pathol. 159,
2081–208845. Itoh, K., Nakao, A., Kishimoto, W., Itoh, T., Harada, A., Nonami, T., Na-
kano, M., and Takagi, H. (1993) Gastroenterol. Jpn. 28, 541–5463 T. Asahi and T. Osawa, unpublished results.
8-Halogenated dG as Inflammatory Marker
MARCH 19, 2010 • VOLUME 285 • NUMBER 12 JOURNAL OF BIOLOGICAL CHEMISTRY 9291
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Yuji Naito, Toshikazu Yoshikawa, Shinsuke Hisaka, Yoji Kato and Toshihiko OsawaTakashi Asahi, Hajime Kondo, Mitsuharu Masuda, Hoyoku Nishino, Yasuaki Aratani,
Early Stage InflammationChemical and Immunochemical Detection of 8-Halogenated Deoxyguanosines at
doi: 10.1074/jbc.M109.054213 originally published online January 15, 20102010, 285:9282-9291.J. Biol. Chem.
10.1074/jbc.M109.054213Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/285/12/9282.full.html#ref-list-1
This article cites 45 references, 21 of which can be accessed free at
by guest on April 8, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























