Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, May 1993, p. 2826-28320021-9193/93/102826-07$02.00/0Copyright © 1993, American Society for Microbiology
Vol. 175, No. 10
Characterization and Symbiotic Importance of AcidicExtracellular Polysaccharides of Rhizobium sp. Strain GRH2
Isolated from Acacia NodulesISABEL M. LOPEZ-LARA,l GUY ORGAMBIDE,2 FRANK B. DAZZO,2 JOSE OLIVARES,1
AND NICOLAS TORO"*Department ofMicrobiology, Estacion Experimental del Zaidin, Consejo Superior de Investigaciones
Cientificas, CiProfesor Albareda 1, 18008 Granada, Spain, 1 and Department ofMicrobiology, Michigan State University, East Lansing, Michigan 488242
Received 26 October 1992/Accepted 14 March 1993
RIizobium sp. wild-type strain GRH2 was originally isolated from root nodules of the leguminous tree Acaciacyanophylla and has a broad host range which includes herbaceous legumes, e.g., Trifolium spp. We examinedthe extracellular exopolysaccharides (EPSs) produced by strain GRH2 and found three independent glycosidicstructures: a high-molecular-weight acidic heteropolysaccharide which is very similar to the acidic EPSproduced by Rhizobium leguminosarum biovar trifolii ANU843, a low-molecular-weight native heterooligosac-charide resembling a dimer of the repeat unit of the high-molecular-weight EPS, and low-molecular-weightneutral 13(1,2)-glucans. A TnS insertion mutant derivative of GRH2 (exo-57) that fails to form acidicheteropolysaccharides was obtained. This Exo- mutant formed nitrogen-fixing nodules on Acacia plants butinfected a smaller proportion of cells in the central zone of the nodules than did wild-type GRH2. In addition,the exo-57 mutant failed to nodulate several herbaceous legume hosts that are nodulated by wild-type strainGRH12.
Leguminous trees proliferate in arid and semiarid regionsof the world which are unsuited for crop production. Theseleguminous trees are of increasing interest since they pro-vide wood, fiber, gum, and food for animal and humannutrition, protect the soil from erosion, and form nitrogen-fixing nodules in symbiotic association with bacteria (2, 5,19, 24, 31, 41). Recent studies revealed that leguminous treesare infected by both fast-growing rhizobia and slowly grow-ing bradyrhizobia. Some rhizobial strains isolated from thenodules have a wide host range, even including herbaceouslegumes (15, 16, 24, 30, 45). This is the case for fast-growingRhizobium sp. strain GRH2, originally isolated from Acaciacyanophylla (24). Many reports suggest involvement ofRhizobium acidic heteropolysaccharides in infection of le-gume roots hairs, invasion of nodules, and nitrogen fixation(1, 6, 11, 21, 28, 32, 35, 37) and a possible contribution todetermination of the bacterial host range (13, 14, 20, 36).
Rhizobia with broad host ranges are very useful forinvestigations of the molecular mechanisms of symbioticinteraction. Rhizobium sp. strain NGR234 is one of the mostextensively studied strains with a broad host range, whichincludes several tropical legumes and the nonlegume Para-sponia spp. This strain shows functional and evolutionarygenetic equivalence to Rhizobium meliloti (44). In contrast,studies of restriction fragment length polymorphism suggestthat broad-host-range Rhizobium sp. strain GRH2 is moreclosely related to R. leguminosarum (30a). The aim of thepresent work was to evaluate the extracellular exopolysac-charide (EPS) of strain GRH2 and its role in the root nodulesymbiosis between the bacterial symbiont and its legumi-nous hosts. We show that the EPSs produced by Rhizobiumsp. strain GRH2 are very closely related to those producedby wild-type R. leguminosarum bv. tnfolii strain ANU843.
* Corresponding author.
In addition, a Tn5-induced Exo- mutant derivative of GRH2which does not produce acidic heteropolysaccharides wasobtained and found to be unable to induce nodules on severalherbaceous legumes hosts. However, it still nodulated Aca-cia sp. but infected fewer host cells in the nodule.
MATERIALS AND METHODS
Bacterial strains and genetic manipulations. Rhizobium sp.strain GRH2 was originally isolated from root nodules ofA.cyanophylla (24). Bacteria were routinely grown in minimalmedium (39) or TY medium (8). Transposon Tn5::mobrandom mutagenesis was done by mating Escherichia coliS17-1(pSUP5011) with Rhizobium sp. strain GRH2. Kana-mycin-resistant transconjugants were selected on minimalmedium-agar. Kanamycin sulfate and tetracycline were usedat 280 and 10 ptg/ml, respectively. Plasmid isolation, ran-domly primed DNA labeling, DNA hybridization, andSouthern blotting were performed by standard procedures(30).
Isolation and fractionation of EPS components. Bacteriawere grown at 280C on a rotary shaker in Erlenmeyer flaskscontaining 500 ml of minimal medium (39) for 4 days. Afterthe bacteria were pelleted by centrifugation, the supernatantwas lyophilized and 5 mg of this dry residue was redissolvedin 5 ml of 50 mM sodium phosphate buffer (pH 7.0) contain-ing 100 mM NaCl. The solution was fractionated on acolumn of Bio-Gel A-5m (2.6 by 84 cm; Bio-Rad Laborato-ries) equilibrated in the same phosphate buffer. The first 65ml of eluant was discarded, and then 7-ml fractions werecollected and assayed for carbohydrate content by thephenol-sulfuric acid method (22). The second, lower-molec-ular-weight (MW) peak of carbohydrate material was furtherfractionated by gel permeation chromatography on aSephacryl S-100-HR column (1 by 50 cm; Pharmacia LKBBiotechnology) with 1 mM ammonium formate (pH 6.8)
2826
on January 4, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

RHIZOBIUM SP. GRH2 EPS 2827
buffer. Gel permeation high-performance liquid chromatog-raphy (HPLC) of the oligosaccharides was performed with aSpherogel TSK 2000SW analytical column (Beckman Instru-ments, Inc.) with the same buffer.
Production of EPS-derived oligosaccharides. The high-MWacidic heteropolysaccharides were enzymatically depoly-merized by the action of a bacteriophage-borne glucuronicacid-specific lyase (PD-I) purified from a lysate of bacterio-phage 4S grown on R. trifoli 4S, and the oligosaccharideswere subsequently purified as described earlier (26).
Sugar and glycosidic linkage composition of glycosides.Oligo- and polysaccharides (1 mg) were treated with anhy-drous 1 M methanolic chloride for 7 h at 80'C. The neutral-ized methanolysate was then either peracetylated with aceticanhydride-pyridine (1:1; methylglycoside peracetate deriva-tives) or reduced with sodium borodeuteride (10 mg/ml inCH3OH-H20 [1:1]), hydrolyzed (2 M trifluoroacetic acid, 2h, 110'C), reduced with sodium borohydride, and peracety-lated (alditol acetate derivatives). Methylation analyses wereperformed on 1-mg samples as described by Waeghe et al.(43). Gas chromatography-mass spectrometry (MS) analysesof methylglycoside peracetates, alditol acetates, and par-tially methylated alditol acetate derivatives were done with aHewlett-Packard 5995C instrument equipped with a SupelcoSP-2330 column. The sugar derivatives were identified bycomparison of their mass spectra and gas-liquid chromatog-raphy (GLC) retention times with those of authentic stan-dards and quantitated by flame ionization detection-GLC.The relative ratios of hexose, uronic acid, and unsaturateduronic acid in the isolated polysaccharides or oligosaccha-rides from GRH2, ANU843, and 248 were compared bycolorimetric analysis. Samples (100 to 200 pg) were dis-solved in water, and aliquots were assayed colorimetricallyfor hexoses by the phenol-sulfuric acid method (22), foruronic acids by the carbazole reaction (12), and for theunsaturated uronic acid derivative by the thiobarbituric acidassay (26). For each glycoside, relative uronic acid contentwas estimated by the ratio of optical density at 525 nm(OD525) (carbazole assay) to OD490 (phenol-sulfuric acidassay), and unsaturated uronic acid derivative content wasestimated by the ratio of OD552 (thiobarbituric acid assay) toOD490.
Spectroscopic methods. 1H and 13C nuclear magnetic res-onance (NMR) analyses were performed on a VarianVXR500 instrument (500 MHz for H, 125 MHz for '3C) withdeuterated water (99.96%) as the solvent at 25°C for oli-gosaccharides and 80°C for polysaccharides. Chemical shiftswere referred to trimethylsilane as an external reference.Stoichiometry of noncarbohydrate substituents per oligosac-charidic unit was established by referring the relative inte-gration of proton resonances to H-4 of the unsaturatedglucuronic acid derivative at the nonreducing end of eacholigomer (8H-4, 5.75 ppm) (26, 27). Fast atom bombardment(FAB)-MS analysis of acidic oligosaccharides was con-ducted as described earlier (27).
Nodulation assays. Plants were grown on nitrogen-freemedium (38) and inoculated as described previously (35).Nodulation of tree legumes and herbaceous legumes wasmonitored for 6 months and 1 month, respectively.
Plant microscopy. Root nodules which developed on Aca-cia plants after 1 month of incubation with wild-type GRH2or mutant GRH2-57 were excised, fixed in glutaraldehyde,dehydrated through an ethanol series, and embedded inEpon resin. Two-micrometer sections were stained withtoluidine blue solution and examined by light microscopy.Differential counts of infected and uninfected nodule cells
within the central region bordered by the endodermis weremade from video prints. Six evaluations were done for eachstrain.
RESULTS
Structural analysis of GRH2 high-MW EPS. Structuralevaluation of GRH2 EPS was performed by comparativeanalysis with the different EPSs produced by R. meliloti 1021(3, 23), R. leguminosarum bv. viciae strain 248 (10), R.leguminosarum bv. trifolii strain ANU843 (27), and B.japonicum CC708 (17). Several lines of evidence indicated astrong similarity between GRH2 high-MW EPS and theacidic heteropolysaccharide produced by R. trifoliiANU843. The latter polysaccharide consists of a branchedoctasaccharide repeat unit containing glucose, glucuronicacid, and galactose in a 5:2:1 ratio and, in addition, nonsto-ichiometric amounts of pyruvyl, acetyl, and 3-hydroxybu-tyryl substitutions (27). (i) The GLC patterns of alditolacetates or methylglycoside peracetates derived from GRH2EPS were identical to those obtained with ANU843 EPS. Inparallel analyses, the Glc/GlcA/Gal ratios were 4.9:1.2:1.0for GRH2 EPS and 5.2:1.2:1.0 for ANU843 EPS. A colori-metric evaluation further supported the similarity in therelative uronic acid contents of GRH2 EPS and ANU843EPS since their OD52JOD490 ratios were 1.16 and 0.92,respectively. In contrast, the EPS from R. leguminosarumbv. viceae strain 248, which contains three GlcA residuesper repeat unit (10), yielded a ratio of 1.91 in the sameexperiment. (ii) The 1H NMR spectrum of the GRH2 poly-saccharide (Fig. 1A) displayed, in addition to the carbohy-drate signals, typical resonances of pyruvate (CH3, SH, 1.35to 1.45 ppm), acetate (CH3, bH, 2.0 to 2.2 ppm), and3-hydroxybutyrate (CH3, 8H, 1.20 ppm; CH2, 8H, 2.59ppm). These assignments were further assessed by alkalinetreatment and subsequent 1H NMR analysis of the GRH2EPS, which showed disappearance of the acetate and 3-hy-droxybutyrate signals as a result of the de-O-acylation,without alteration of the resonances assigned to the pyruvatemoieties. (iii) Like ANU843 EPS, GRH2 EPS was fullydepolymerized by the action of a bacteriophage-borne glu-curonic acid-specific lyase (PD-I) (26), yielding a singleoligosaccharidic unit (hereafter called EPSOLIGO). Its re-tention time on a TSK 2000SW gel permeation HPLCcolumn and sugar composition (Glc/GlcA/Gal ratios in oli-gomers, 5.4:0.8:1 for GRH2 and 4.9:0.7:1 for ANU843) weresimilar to those of the oligosaccharidic repeat unit obtainedby the same enzymatic treatment of ANU843 EPS. Under-estimation of the glucuronic acid contents of both acidicpolysaccharides and heterooligosaccharides occurred be-cause of incomplete reduction of the carboxylate groups,partial decomposition of this compound under acidic condi-tions (34), and resistance of the glycosidic bond engaged bythis sugar towards acidic solvolyses (7). Upon colorimetricevaluation, the relative uronic acid contents of the enzymat-ically produced oligosaccharides from GRH2 EPS andANU843 EPS were also found to be similar, as defined byOD525/OD490 ratios of 1.13 and 1.18, respectively. The 1HNMR spectra of the two EPS-derived oligosaccharides fromGRH2 and ANU843 (Fig. 1C and D) showed striking simi-larity in distribution and relative intensities of the reso-nances for glycosyl, pyruvyl, and 3-hydroxybutyryl signalsand only a slight difference in the distribution of their0-acetyl resonances (an additional minor signal was ob-served at bH, 2.02 ppm in the GRH2 oligomer). Quantitationof the noncarbohydrate substituents established the follow-
VOL. 175, 1993
on January 4, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
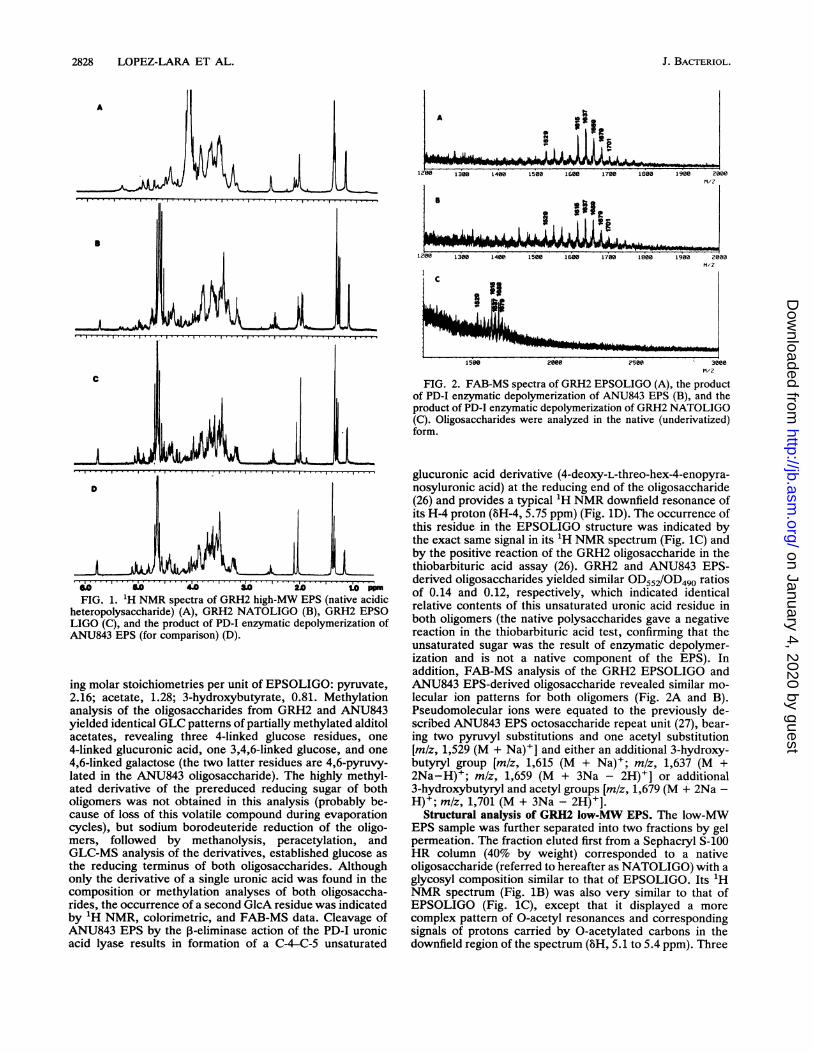
2828 LOPEZ-LARA ET AL.
A
II
A
P-'WW~WWW W WV-
w--V-Be 1300 1400 1509 1600 1700 1900 1900 2e
C
_A~~~~~~~d~2 1.OoFIG. 1. 'H NMR spectra of GRH2 high-MW EPS (native acidic
heteropolysaccharide) (A), GRH2 NATOLIGO (B), GRH2 EPSOLIGO (C), and the product of PD-I enzymatic depolymerization ofANU843 EPS (for comparison) (D).
ing molar stoichiometries per unit of EPSOLIGO: pyruvate,2.16; acetate, 1.28; 3-hydroxybutyrate, 0.81. Methylationanalysis of the oligosaccharides from GRH2 and ANU843yielded identical GLC patterns of partially methylated alditolacetates, revealing three 4-linked glucose residues, one4-linked glucuronic acid, one 3,4,6-linked glucose, and one4,6-linked galactose (the two latter residues are 4,6-pyruvy-lated in the ANU843 oligosaccharide). The highly methyl-ated derivative of the prereduced reducing sugar of botholigomers was not obtained in this analysis (probably be-cause of loss of this volatile compound during evaporationcycles), but sodium borodeuteride reduction of the oligo-mers, followed by methanolysis, peracetylation, andGLC-MS analysis of the derivatives, established glucose asthe reducing terminus of both oligosaccharides. Althoughonly the derivative of a single uronic acid was found in thecomposition or methylation analyses of both oligosaccha-rides, the occurrence of a second GlcA residue was indicatedby 1H NMR, calorimetric, and FAB-MS data. Cleavage ofANU843 EPS by the 13-eliminase action of the PD-I uronicacid lyase results in formation of a C-4-C-5 unsaturated
M/Z
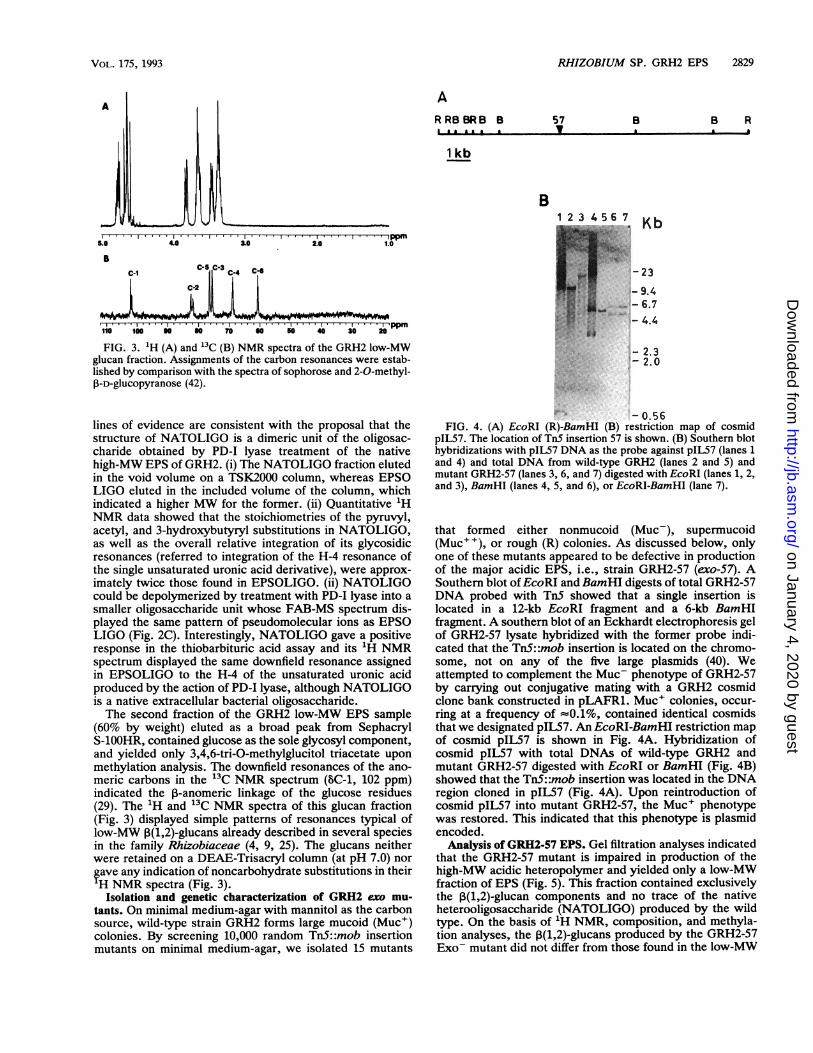
FIG. 2. FAB-MS spectra of GRH2 EPSOLIGO (A), the productof PD-I enzymatic depolymerization of ANU843 EPS (B), and theproduct of PD-I enzymatic depolymerization of GRH2 NATOLIGO(C). Oligosaccharides were analyzed in the native (underivatized)form.
glucuronic acid derivative (4-deoxy-L-threo-hex-4-enopyra-nosyluronic acid) at the reducing end of the oligosaccharide(26) and provides a typical 'H NMR downfield resonance ofits H-4 proton (BH-4, 5.75 ppm) (Fig. 1D). The occurrence ofthis residue in the EPSOLIGO structure was indicated bythe exact same signal in its 1H NMR spectrum (Fig. 1C) andby the positive reaction of the GRH2 oligosaccharide in thethiobarbituric acid assay (26). GRH2 and ANU843 EPS-derived oligosaccharides yielded similar OD552/OD490 ratiosof 0.14 and 0.12, respectively, which indicated identicalrelative contents of this unsaturated uronic acid residue inboth oligomers (the native polysaccharides gave a negativereaction in the thiobarbituric acid test, confirming that theunsaturated sugar was the result of enzymatic depolymer-ization and is not a native component of the EPS). Inaddition, FAB-MS analysis of the GRH2 EPSOLIGO andANU843 EPS-derived oligosaccharide revealed similar mo-lecular ion patterns for both oligomers (Fig. 2A and B).Pseudomolecular ions were equated to the previously de-scribed ANU843 EPS octosaccharide repeat unit (27), bear-ing two pyruvyl substitutions and one acetyl substitution[m/z, 1,529 (M + Na)+] and either an additional 3-hydroxy-butyryl group [mlz, 1,615 (M + Na)+; mlz, 1,637 (M +2Na-H)+; mlz, 1,659 (M + 3Na - 2H)+] or additional3-hydroxybutyryl and acetyl groups [m/z, 1,679 (M + 2Na -H)+; m/z, 1,701 (M + 3Na - 2H)+].
Structural analysis of GRH2 low-MW EPS. The low-MWEPS sample was further separated into two fractions by gelpermeation. The fraction eluted first from a Sephacryl S-100HR column (40% by weight) corresponded to a nativeoligosaccharide (referred to hereafter as NATOLIGO) with aglycosyl composition similar to that of EPSOLIGO. Its 1HNMR spectrum (Fig. 1B) was also very similar to that ofEPSOLIGO (Fig. 1C), except that it displayed a morecomplex pattern of O-acetyl resonances and correspondingsignals of protons carried by 0-acetylated carbons in thedownfield region of the spectrum (8H, 5.1 to 5.4 ppm). Three
II
,.
J. BACTERIOL.
on January 4, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
RHIZOBIUM SP. GRH2 EPS 2829
A
5.0 4.0 s0 2.0 1.0
Bc-1 C-s :-3 C-4.C-CC-2
11o 100 70 60 50 40 3s0..
20
FIG. 3. 'H (A) and 13C (B) NMR spectra of the GRH2 low-MWglucan fraction. Assignments of the carbon resonances were estab-lished by comparison with the spectra of sophorose and 2-0-methyl-P-D-glucopyranose (42).
lines of evidence are consistent with the proposal that thestructure of NATOLIGO is a dimeric unit of the oligosac-charide obtained by PD-I lyase treatment of the nativehigh-MW EPS of GRH2. (i) The NATOLIGO fraction elutedin the void volume on a TSK2000 column, whereas EPSOLIGO eluted in the included volume of the column, whichindicated a higher MW for the former. (ii) Quantitative 'HNMR data showed that the stoichiometries of the pyruvyl,acetyl, and 3-hydroxybutyryl substitutions in NATOLIGO,as well as the overall relative integration of its glycosidicresonances (referred to integration of the H-4 resonance ofthe single unsaturated uronic acid derivative), were approx-imately twice those found in EPSOLIGO. (ii) NATOLIGOcould be depolymerized by treatment with PD-I lyase into asmaller oligosaccharide unit whose FAB-MS spectrum dis-played the same pattern of pseudomolecular ions as EPSOLIGO (Fig. 2C). Interestingly, NATOLIGO gave a positiveresponse in the thiobarbituric acid assay and its 'H NMRspectrum displayed the same downfield resonance assignedin EPSOLIGO to the H-4 of the unsaturated uronic acidproduced by the action of PD-I lyase, although NATOLIGOis a native extracellular bacterial oligosaccharide.The second fraction of the GRH2 low-MW EPS sample
(60% by weight) eluted as a broad peak from SephacrylS-100HR, contained glucose as the sole glycosyl component,and yielded only 3,4,6-tri-O-methylglucitol triacetate uponmethylation analysis. The downfield resonances of the ano-meric carbons in the 13C NMR spectrum (BC-1, 102 ppm)indicated the P-anomeric linkage of the glucose residues(29). The 1H and '3C NMR spectra of this glucan fraction(Fig. 3) displayed simple patterns of resonances typical oflow-MW P(1,2)-glucans already described in several speciesin the family Rhizobiaceae (4, 9, 25). The glucans neitherwere retained on a DEAE-Trisacryl column (at pH 7.0) norgave any indication of noncarbohydrate substitutions in theirH NMR spectra (Fig. 3).Isolation and genetic characterization of GRH2 exo mu-
tants. On minimal medium-agar with mannitol as the carbonsource, wild-type strain GRH2 forms large mucoid (Muc')colonies. By screening 10,000 random Tn5::mob insertionmutants on minimal medium-agar, we isolated 15 mutants
ARRBBRB B
MEa^s a 2
57v
B B Ra a
1 kb
B1 2 3 L5 6 7 Kb
- 23- 9.4- 6.7- 4.4
X f -~~2.3||t, - 2.0
0.56FIG. 4. (A) EcoRI (R)-BamHI (B) restriction map of cosmid
pIL57. The location of Tn5 insertion 57 is shown. (B) Southern blothybridizations with pIL57 DNA as the probe against pIL57 (lanes 1and 4) and total DNA from wild-type GRH2 (lanes 2 and 5) andmutant GRH2-57 (lanes 3, 6, and 7) digested with EcoRI (lanes 1, 2,and 3), BamHI (lanes 4, 5, and 6), or EcoRI-BamHI (lane 7).
that formed either nonmucoid (Muc-), supermucoid(Muc++), or rough (R) colonies. As discussed below, onlyone of these mutants appeared to be defective in productionof the major acidic EPS, i.e., strain GRH2-57 (exo-57). ASouthern blot of EcoRI and BamHI digests of total GRH2-57DNA probed with TnS showed that a single insertion islocated in a 12-kb EcoRI fragment and a 6-kb BamHIfragment. A southern blot of an Eckhardt electrophoresis gelof GRH2-57 lysate hybridized with the former probe indi-cated that the Tn5::mob insertion is located on the chromo-some, not on any of the five large plasmids (40). Weattempted to complement the Muc- phenotype of GRH2-57by carrying out conjugative mating with a GRH2 cosmidclone bank constructed in pLAFR1. Muc+ colonies, occur-ring at a frequency of -0.1%, contained identical cosmidsthat we designated pIL57. An EcoRI-BamHI restriction mapof cosmid pIL57 is shown in Fig. 4A. Hybridization ofcosmid pIL57 with total DNAs of wild-type GRH2 andmutant GRH2-57 digested with EcoRI or BamHI (Fig. 4B)showed that the TnS::mob insertion was located in the DNAregion cloned in pIL57 (Fig. 4A). Upon reintroduction ofcosmid pIL57 into mutant GRH2-57, the Muc+ phenotypewas restored. This indicated that this phenotype is plasmidencoded.
Analysis of GRH2-57 EPS. Gel filtration analyses indicatedthat the GRH2-57 mutant is impaired in production of thehigh-MW acidic heteropolymer and yielded only a low-MWfraction of EPS (Fig. 5). This fraction contained exclusivelythe 3(1,2)-glucan components and no trace of the nativeheterooligosaccharide (NATOLIGO) produced by the wildtype. On the basis of 1H NMR, composition, and methyla-tion analyses, the 3(1,2)-glucans produced by the GRH2-57Exo- mutant did not differ from those found in the low-MW
VOL. 175, 1993
on January 4, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2830 LOPEZ-LARA ET AL.
60
'60
I
GRH2
100 200 300 400 ml
GRH2-57
0 100 200 300 400 ml
FIG. 5. Chromatographic profile on Bio-Gel A-5m of the EPSproduced by Rhizobium sp. strain GRH2 and mutant derivativeGRH2-57.
EPS fraction from the GRH2 parent. Production of thehigh-MW acidic heteropolymer and NATOLIGO was re-stored by genetic complementation of mutant GRH2-57 withcosmid pIL57.
A
Effects ofexo-57 mutation on infection of tree legumes. Likethe wild type, mutant GRH2-57 induced nitrogen-fixing rootnodules on A. cyanophylla, A. melanoxylon, and Prosopischilensis plants. However, plant microscopy of 1-month-oldroot nodules of A. cyanophylla revealed that the number ofinfected cells in the central zone of the nodules was consid-erably reduced compared with nodules elicited by the wild-type strain (Fig. 6). This cytological difference was con-firmed by quantitative evaluation indicating that percentagesof infected cells in this region were 51 + 2 versus 16 ± 3(mean ± the standard error of the mean; n = 6) for nodulesinduced by wild-type GRH2 and Exo- mutant GRH2-57,respectively.
Effect of exo-57 mutation on host range. Whereas wild-typeGRH2 had a broad host range which included the ability toform root nodules on Lotus corniculatus, Vicia hirsuta,Phaseolus vulganis, Trnfolium subterraneum, T. repens, andT. incamatum, Exo- mutant GRH2-57 was unable to nodu-late Vicia or Tifolium roots. The wild-type host range wasrestored by complementation with cosmid pIL57. Appar-ently, the exo gene mutated in strain GRH2-57 plays a role indetermining the leguminous host range of wild-type GRH2.
DISCUSSIONRhizobium sp. strain GRH2, originally isolated in Chile
from A. cyanophylla root nodules, is capable of infecting abroad range of legume species, including herbaceous le-gumes (24). In contrast to broad-host-range Rhizobium sp.strain NGR234, which shows functional and genetic equiv-alence to R. meliloti, Rhizobium sp. strain GRH2 is moreclosely related to R. leguminosarum (30a). Thus, genetic and
B
FIG. 6. Light microscopy of 2->m sections ofAcacia root nodules elicited by wild-type GRH2 (A) and mutant GRH2-57 (B). Bars, 50 ,um.
J. BACTERIOL.
on January 4, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

RHIZOBIUM SP. GRH2 EPS 2831
biochemical characterization of the EPS produced by Rhi-zobium sp. strain GRH2 can provide further evidence of itsgeneral role in leguminous root nodule symbiosis. In thisreport, three independent glycosidic structures were identi-fied as EPSs of wild-type strain GRH2. A high-MW acidicheteropolysaccharide was found to be very similar to theEPS produced by wild-type R. tnifolii ANU843 (27), on
the basis of the identity and stoichiometry of glycosyl(Glc, GlcA, and Gal) and noncarbohydrate (pyruvate, ace-
tate, and 3-hydroxybutyrate) substituents, glycosidic linkagecomposition, 1H NMR analysis of both the polymer and itsenzymatically produced oligosaccharide repeat unit, andFAB-MS analysis of the latter. A structurally related,low-MW native heterooligosaccharide was also isolatedfrom GRH2 EPS, and results of gel permeation HPLC, 1HNMR, and FAB-MS analyses indicated that its structure isconsistent with a dimer of an oligosaccharidic repeat unit ofthe high-MW EPS. In addition to its size, the only differencefound between the native acidic heterooligosaccharide andthe monomeric repeat unit obtained by PD-I lyase treatmentof the acidic EPS polymer was a more complex acetylationpattern in the former. A similar relationship has been re-
ported for R. tnifolii ANU843, in which the naturally occur-
ring extracellular hexadecasaccharide (14) contained theunsaturated uronic acid derivative and corresponded to thedimer of the enzymatically produced octasaccharide repeatunit of the heteropolysaccharide (27). A. cyanophylla treesare widespread in Australia, where R. trifolii ANU843 was
isolated (27). Thus, it is possible that both bacterial strainsshare a common ancestor. Whereas other symbiotic genes instrain GRH2, such as nod and fix, have evolved to provide a
broad host range, the striking identity between the EPSsproduced by Rhizobium sp. strain GRH2 and R. trifoliiANU843 suggests a very tightly conserved function. Thethird component of the GRH2 EPSs consisted of low-MW,neutral P(1,2)-glucans. As with the acidic EPS fractions, theimportance of this class of glycoconjugates in symbioticRhizobium-legume interactions has been emphasized (18,21).To study the symbiotic importance of the acidic het-
eropolysaccharide in nodulation of leguminous trees andherbaceous legumes, we obtained a TnS mutant of broad-host-range strain GRH2. Our results demonstrate a linkbetween the acidic EPS and the natural acidic heterooli-gosaccharide of GRH2, with its leguminous host range
depending on the type (determinate or indeterminate) ofnodule ontogeny, and that their production affects invasionof nodule cells in Acacia plants. Our results reinforce theimportance of the role of rhizobial glycoconjugates in devel-opment of the root nodule symbiosis with legumes.
ACKNOWLEDGMENTS
This work was supported by Comisi6n Asesora de Investigaci6nCientifica y Tecnica grants BT 87-0003 and B1090-0747 (J.O. andN.T.), National Institutes of Health grant GM-34331-06, and theMichigan State University Biotechnology Research Center(F.B.D.). I.L.-L. was supported by an M.E.C. fellowship. FAB-MSwas done at the NIH MS facility at Michigan State University andsupported by NIH-NCRR grant P-41-RR00480 to J. T. Watson.
REFERENCES1. Abe, M., J. E. Sherwood, R. I. Hollingsworth, and F. B. Dazzo.
1984. Stimulation of clover root hair infection by lectin-bindingoligosaccharides from the capsular and extracellular polysac-charides of Rhizobium trifolii. J. Bacteriol. 160:517-520.
2. Allen, 0. N., and E. K. Allen. 1981. The Leguminosae: a source
book of characteristics, uses and nodulation. The University of
Wisconsin Press, Madison.3. Aman, P., M. McNeil, L.-E. Franzen, A. G. Darvill, and P.
Albersheim. 1981. Structural elucidation, using H.P.L.C.-M.S.and G.L.C.-M.S., of the acidic polysaccharide secreted byRhizobium meliloti strain 1021. Carbohydr. Res. 95:263-282.
4. Amemura, A., M. Hisamatu, H. Mitani, and T. Harada. 1983.Cyclic (1-2)P-D-glucan and the octasaccharide repeating units ofextracellular acidic polysaccharides produced by Rhizobium.Carbohydr. Res. 114:277-285.
5. Basak, M. K., and S. K. Goyal. 1980. Studies on tree legumes.II. further additions to the list of nodulating tree legumes. PlantSoil 56:39-51.
6. Battisti, L., J. C. Lara, and J. A. Leigh. 1992. Specific oligosac-charide form of the Rhizobium meliloti exopolysaccharide pro-motes nodule invasion in alfalfa. Proc. Natl. Acad. Sci. USA89:5625-5629.
7. BeMiller, J. N. 1967. Acid-catalyzed hydrolysis of glycosides.Adv. Carbohydr. Chem. 22:25-108.
8. Beringer, J. E. 1974. R factor transfer in Rhizobium leguminosa-rum. J. Gen. Microbiol. 84:188-198.
9. Breedveld, M. W., L. P. T. M. Zevenhuizen, and A. J. B.Zehnder. 1990. Excessive excretion of cyclic f3(1,2)-glucan byRhizobium trifolii TA-1. Appl. Environ. Microbiol. 56:2080-2086.
10. Canter Cremers, H. C. J., K. Stevens, B. J. J. Lugtenberg, C. A.WUffelman, M. Batley, J. W. Redmond, M. W. Breedveld, andL. P. T. M. Zevenhuizen. 1991. Unusual structure of theexopolysaccharide of Rhizobium leguminosarum bv. viciaestrain 248. Carbohydr. Res. 218:185-200.
11. Carlson, R. W. 1982. Surface chemistry, p. 199-234. In W. J.Broughton (ed.), Nitrogen fixation. Rhizobium vol. 2. OxfordUniversity Press, Oxford.
12. Chaplin, M. F. 1986. Monosaccharides, p. 1-36. In M. F.Chaplin and J. F. Kennedy (ed.), Carbohydrate analysis, apractical approach. IRL Press, Oxford.
13. Dazzo, F. B., and R. I. Hollingsworth. 1984. Trifoliin A andcarbohydrate receptors as mediators of cellular recognition inthe Rhizobium tnfolii-clover symbiosis. Biol. Cell 51:267-274.
14. Djordjevic, S. P., H. Chen, M. Batley, J. W. Redmond, and B. G.Rolfe. 1987. Nitrogen fixation ability of exopolysaccharide syn-thesis mutants of Rhizobium sp. strain NGR234 and Rhizobiumtrifoldi is restored by the addition of homologous exopolysac-charides. J. Bacteriol. 169:53-60.
15. Dommergues, Y. R., H. G. Diem, D. L. Gauthier, B. L. Dreyfus,and F. Cornet. 1984. Nitrogen-fixing trees in the tropics: poten-tialities and limitations, p. 7-8. In C. Veeger and W. E. Newton(ed.), Martinus Nijhoff/Dr. W. Junk, Wagenigen, The Nether-lands.
16. Dreyfus, B. L., and Y. R. Dommergues. 1981. Nodulation ofacacia species by fast- and slow-growing tropical strains ofRhizobium. Appl. Environ. Microbiol. 41:97-99.
17. Dudman, W. F. 1978. Structural studies of the extracellularpolysaccharides of Rhizobium japonicum 71A, CC708, andCB1795. Carbohydr. Res. 66:9-23.
18. Dylan, T., L. Ielpi, S. Stanfield, L. Kashyap, C. Douglas, M.Yanofsky, E. Nester, D. R. Helinsky, and G. Ditta. 1986.Rhizobium meliloti genes required for nodule development arerelated to chromosomal virulence genes in Agrobacteriumtumefaciens. Proc. Natl. Acad. Sci. USA 83:4403-4407.
19. Felker, P., and P. R. Clark. 1980. Nitrogen fixation (acetylenereduction) and cross inoculation in 12 Prosopis (mesquite)species. Plant Soil 57:177-186.
20. Glazebrook, J., and G. C. Walker. 1989. A novel exopolysac-charide can function in place of the calcofluor-binding ex-opolysaccharide in nodulation of alfalfa by Rhizobium meliloti.Cell 56:661-672.
21. Gray, J. X., and B. G. Rolfe. 1990. Exopolysaccharide produc-tion in Rhizobium and its role in invasion. Mol. Microbiol.4:1425-1431.
22. Hanson, R. S., and J. A. Phillips. 1981. Chemical composition,p. 328-364. In P. Gerhardt, R. G. E. Murray, R. N. Costilow,E. W. Nester, W. A. Wood, N. R. Krieg, and G. B. Phillips(ed.), Manual of methods for general bacteriology. American
VOL. 175, 1993
on January 4, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2832 LOPEZ-LARA ET AL.
Society for Microbiology, Washington, D.C.23. Her, G. R., J. Glazebrook, G. C. Walker, and V. N. Reinhold.
1990. Structural studies of a novel exopolysaccharide producedby a mutant of Rhizobium meliloti strain RmlO21. Carbohydr.Res. 198:305-312.
24. Herrera, M. A., E. J. Bedmar, and J. Olivares. 1985. Hostspecificity of Rhizobium strains isolated from nitrogen-fixingtrees and nitrogenase activities of strain GRH2 in symbiosiswith Prosopis chilensis. Plant Sci. 42:177-182.
25. Hisamatsu, M., A. Amemura, K. Koizumi, T. Utamura, and Y.Okada. 1983. Structural studies on cyclic (1-2)-p-D-glucans(cyclosophoraoses) produced by Agrobacterium and Rhizo-bium. Carbohydr. Res. 121:31-40.
26. Hollingsworth, R. I., M. Abe, J. E. Sherwood, and F. B. Dazzo.1984. Bacteriophage-induced acidic heteropolysaccharidelyases that convert the acidic heteropolysaccharides of Rhizo-bium trifolii into oligosaccharide units. J. Bacteriol. 160:510-516.
27. Hollingsworth, R. I., F. B. Dazzo, K. Hallenga, and B. Mussel-man. 1988. The complete structure of the trifoliin A lectin-binding capsular polysaccharide of Rhizobium tnifolii 843. Car-bohydr. Res. 172:97-112.
28. Leigh, J. A., J. W. Reed, J. F. Hanks, A. M. Hirsch, and G. C.Walker. 1987. Rhizobium meliloti mutants that fail to succiny-late their calcofluor-binding exopolysaccharide are defective innodule invasion. Cell 51:579-587.
29. Lipkind, G. M., A. S. Shashkov, Y. A. Knirel, E. V. Vinogradov,and N. K. Kochetkov. 1988. A computer-assisted structuralanalysis of regular polysaccharides on the basis of 13C-NMRdata. Carbohydr. Res. 175:59-75.
30. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
30a.Martinez, E. 1992. Personal communication.31. Miettinen, P., 0. Luukkanen, S. Johansson, E. Eklund, and J.
Mulatya. 1988. Rhizobium nodulation in Prosopis jugifiora seed-ling at different irrigation levels in eastern Kenya. Plant Soil112:233-238.
32. Muller, P., M. Hynes, D. Kapp, K. Niehaus, and A. Puihler. 1988.Two classes of Rhizobium meliloti infection mutants differ inexopolysaccharide production and in coinoculation propertieswith nodulation mutants. Mol. Gen. Genet. 211:17-26.
33. Olivares, J., J. Casadesus, and E. J. Bedmar. 1980. Methods fortesting degree of infectivity of Rhizobium meliloti strains. Appl.Environ. Microbiol. 39:967-970.
34. Perkins, H. R. 1963. A polymer containing glucose and aminohexuronic acid isolated from cell walls ofMicrococcus lysodeik-ticus. Biochem. J. 86:475-483.
35. Philip-Hollingsworth, S., R. I. Hollingsworth, and F. B. Dazzo.1989. Host-range related structural features of the acidic extra-cellular polysaccharides of Rhizobium trifolii and Rhizobiumleguminosarum. J. Biol. Chem. 264:1461-1466.
36. Philip-Hollingsworth, S., R. I. Hollingsworth, F. B. Dazzo, M. A.Djordjevic, and B. G. Rolfe. 1989. The effects of interspeciestransfer of Rhizobium host-specific nodulation genes on acidicpolysaccharide structure and in situ binding by host lectin. J.Biol. Chem. 264:5710-5714.
37. Reuber, T. L., J. Reed, J. Glazebrook, M. A. Glucksmann, D.Ahman, A. Marra, and G. C. Walker. 1991. Rhizobium melilotiexopolysaccharides: genetic analysis and symbiotic importance.Biochem. Soc. Trans. 19:636-641.
38. Rigaud, J., and A. Puppo. 1975. Indol-3-acetic catabolism bysoybean bacteroids. J. Gen. Microbiol. 88:223-228.
39. Robertsen, B. K., P. Aman, A. G. Darvill, M. McNeil, and P.Albersheim. 1981. The structure of acidic extracellular polysac-charides secreted by Rhizobium leguminosarum and Rhizobiumtnifoldi. Plant Physiol. 67:339-340.
40. Toro, N., M. A. Herrera, and J. Olivares. 1984. Location of nifgenes on large plasmids in Rhizobium strains isolated fromlegume tree root nodules. FEMS Microbiol. Lett. 24:113-115.
41. Trinick, M. J. 1980. Relationships amongst the fast growingrhizobia ofLablabpurpureus, Leucaena leucocephala, Mimosaspp., Acacia famesiana and Sesbania grandiflora and theiraffinities with other rhizobia groups. J. Appl. Bacteriol. 49:39-53.
42. Usui, T., N. Yamaoka, K. Matsuda, and K. Tuzimura. 1973. Cnuclear magnetic resonance spectra of glucobioses, glucotri-oses, and glucans. J. Chem. Soc. Perkin Trans. 1:2425-2432.
43. Waeghe, T. J., A. G. Darvill, M. McNeil, and P. Albersheim.1983. Determination, by methylation analysis, of the glycosyl-linkage compositions of microgram quantities of complex car-bohydrates. Carbohydr. Res. 123:281-304.
44. Zhan, H., J. X. Gray, S. B. Levery, B. G. Rolfe, and J. A. Leigh.1990. Functional and evolutionary relatedness of genes forexopolysaccharide synthesis in Rhizobium meliloti and Rhizo-bium sp. strain NGR234. J. Bacteriol. 172:5245-5253.
45. Zhang, X., R. Harper, M. Karsisto, and K. Lindstrom. 1991.Diversity of Rhizobium bacteria isolated from root nodules ofleguminous trees. Int. J. Syst. Bacteriol. 41:104-113.
J. BAcrERIOL.
on January 4, 2020 by guesthttp://jb.asm
.org/D
ownloaded from




























