Embed Size (px)
Citation preview

Chapter 30:
DNA Replication, Repair,and Recombination

1. DNA Replication: An overview2. Enzymes of Replication3. Prokaryotic Replication4. Eukaryotic Replication5. Repair of DNA6. Recombination and Mobile Genetic
Elements7. DNA Methylation and Trinucleotide
Repeat Expansion

DNA Replication
• DNA double strand -> template for duplication, Replication
• Chemically similar to transcription• As complex as translation but enzymes in only few copies/cell
• Extremely accurate: 10-10 mistakes/base• Extremely regulated: only once per cell division
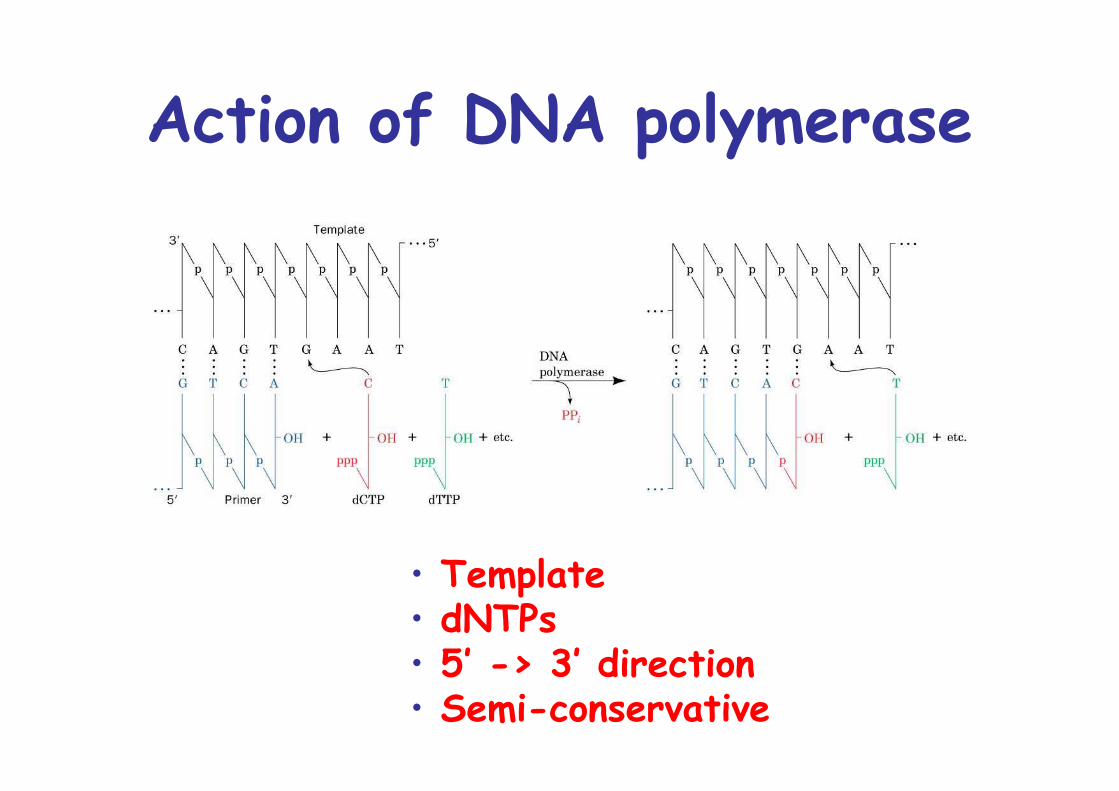
Action of DNA polymerase
• Template• dNTPs• 5’ -> 3’ direction• Semi-conservative
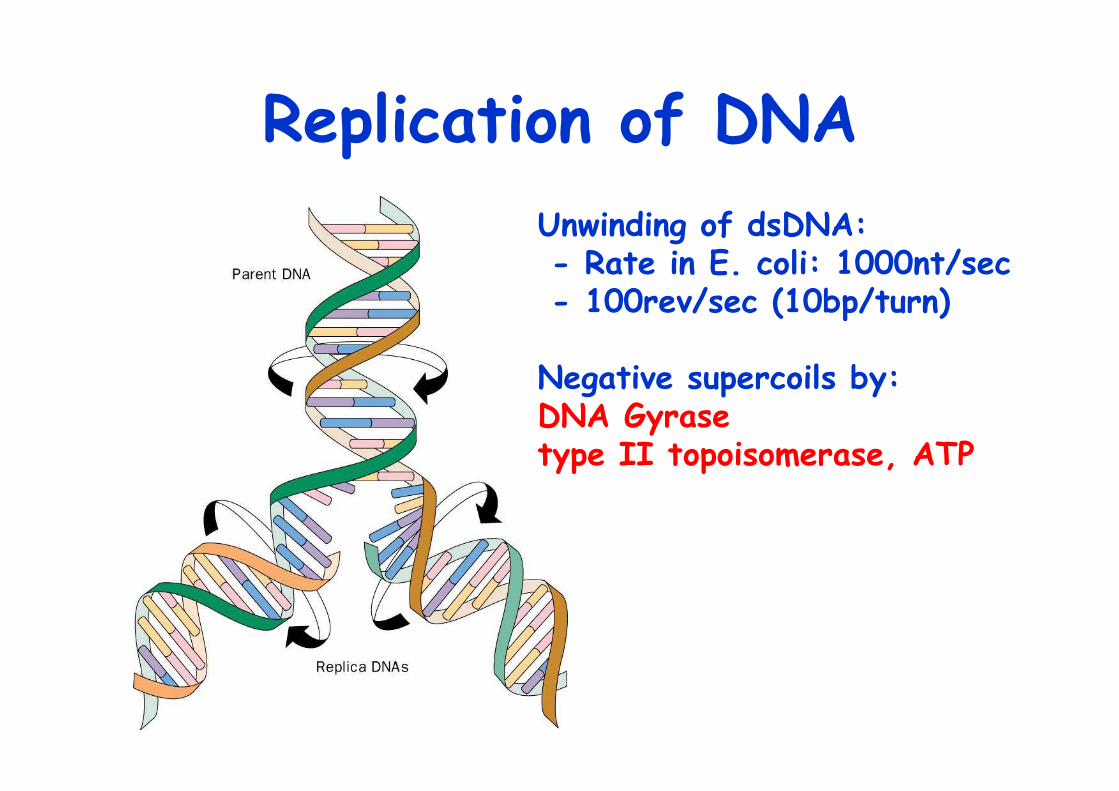
Replication of DNAUnwinding of dsDNA: - Rate in E. coli: 1000nt/sec - 100rev/sec (10bp/turn)
Negative supercoils by: DNA Gyrase type II topoisomerase, ATP
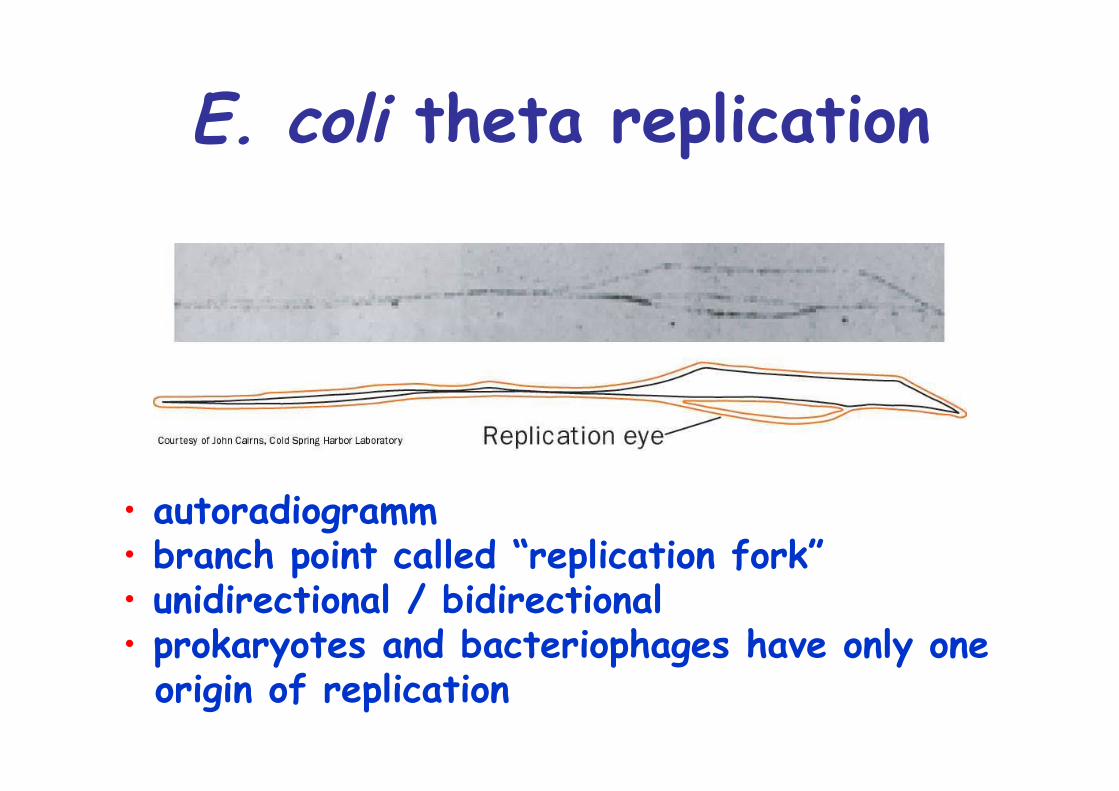
E. coli theta replication
• autoradiogramm• branch point called “replication fork”• unidirectional / bidirectional• prokaryotes and bacteriophages have only one origin of replication
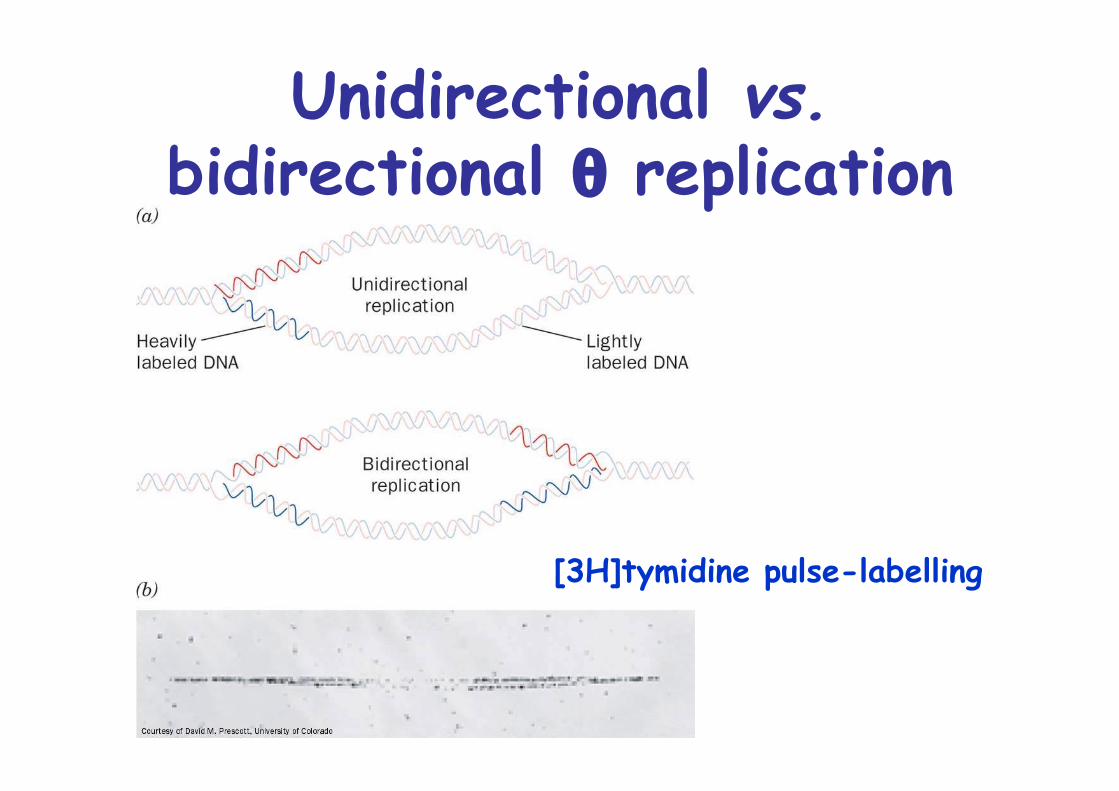
Unidirectional vs.bidirectional θ replication
[3H]tymidine pulse-labelling
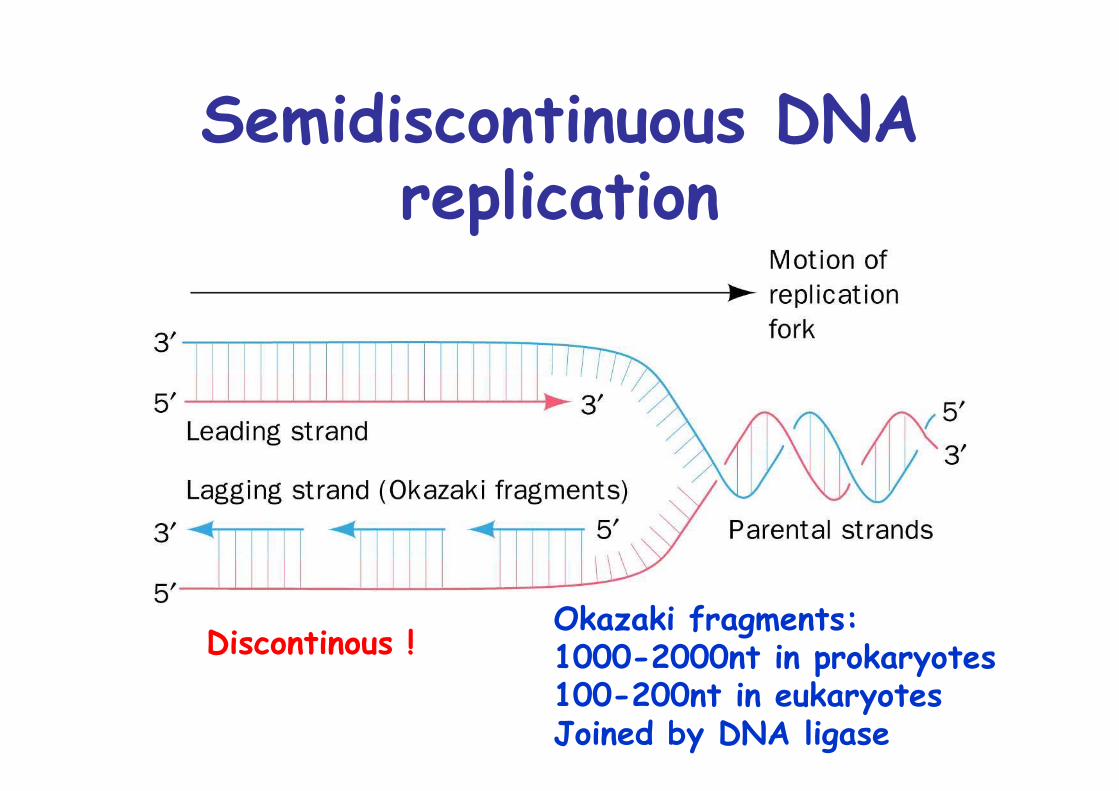
Semidiscontinuous DNAreplication
Okazaki fragments: 1000-2000nt in prokaryotes100-200nt in eukaryotesJoined by DNA ligase
Discontinous !
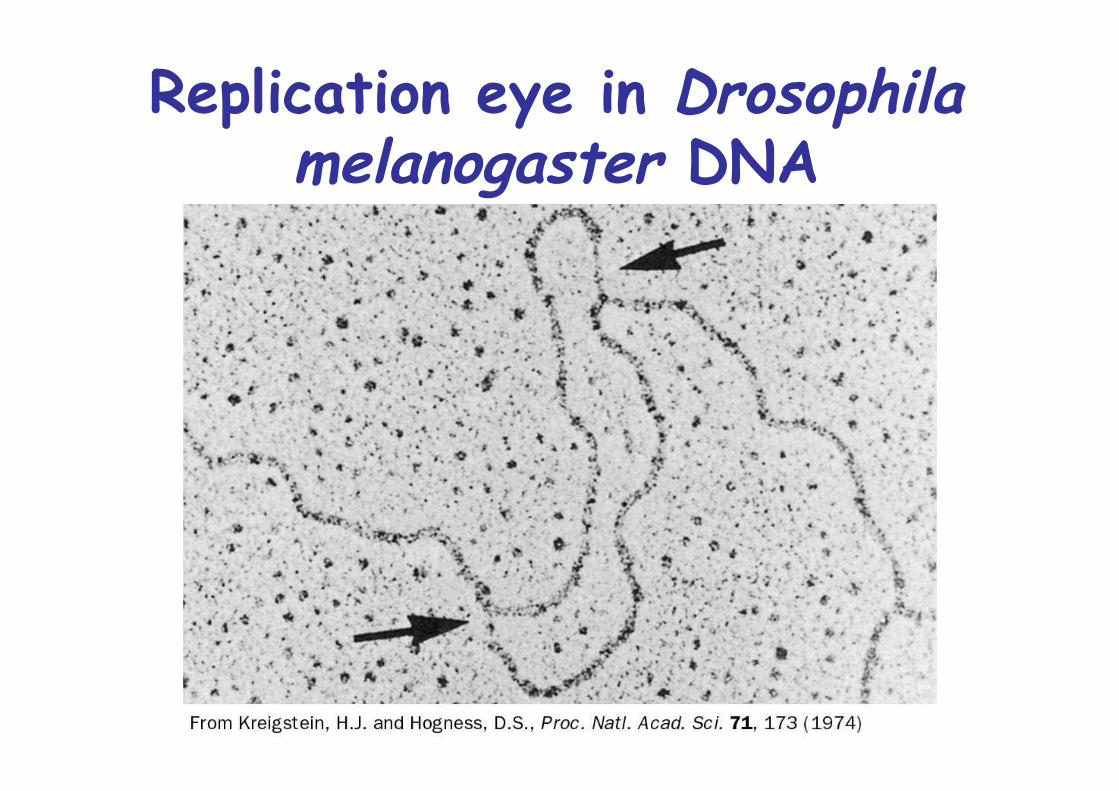
Replication eye in Drosophilamelanogaster DNA
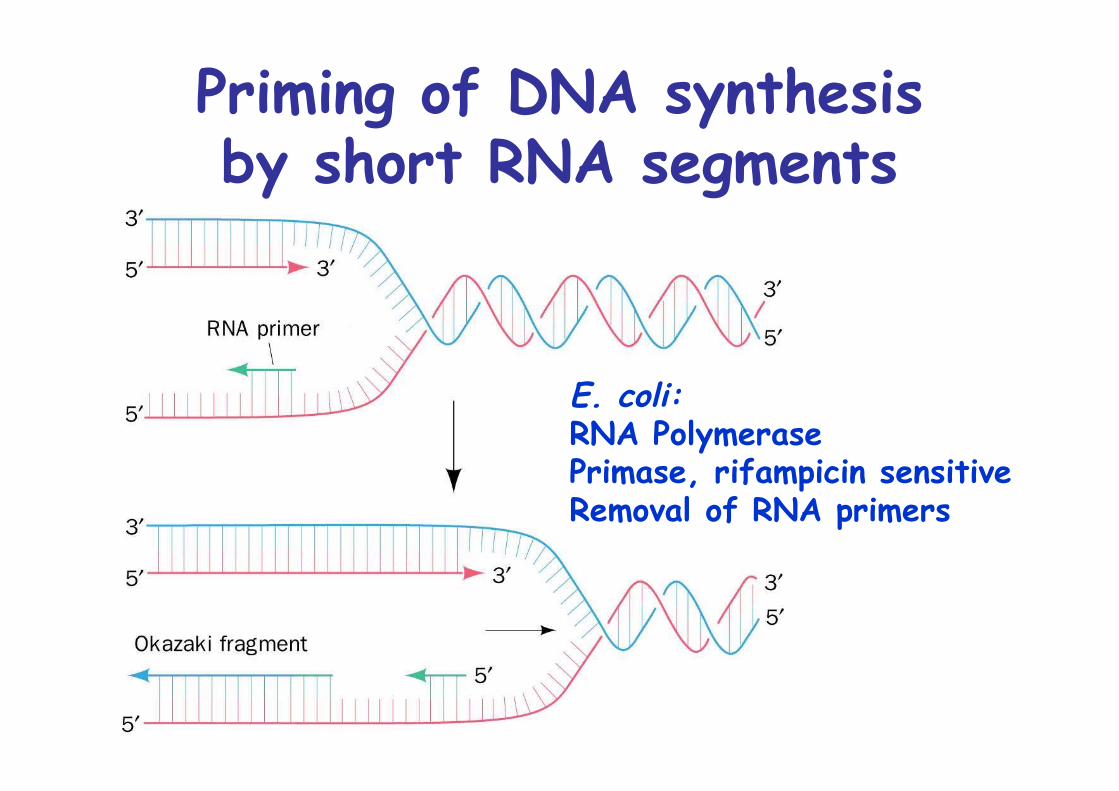
Priming of DNA synthesisby short RNA segments
E. coli: RNA PolymerasePrimase, rifampicin sensitiveRemoval of RNA primers

2. Enzymes of replication
DNA Replication requires (in order of appearance):1. DNA Topoisomerase2. Helicases 3. ssDNA binding proteins4. RNA primer synthesis5. DNA polymerase6. Enzyme to remove RNA primers7. Link Okazaki fragments
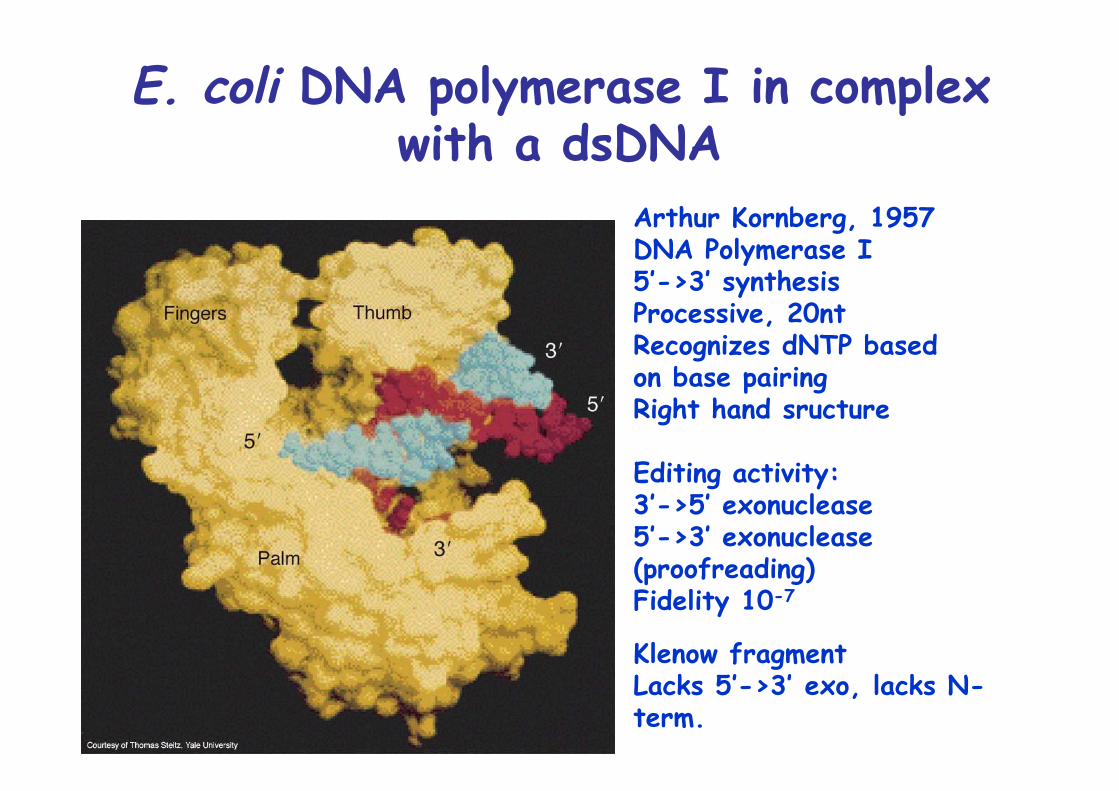
E. coli DNA polymerase I in complexwith a dsDNA
Arthur Kornberg, 1957DNA Polymerase I5’->3’ synthesisProcessive, 20ntRecognizes dNTP basedon base pairingRight hand sructure
Editing activity:3’->5’ exonuclease5’->3’ exonuclease(proofreading)Fidelity 10-7
Klenow fragmentLacks 5’->3’ exo, lacks N-term.

Nick translation as catalyzedby Pol I
Used to radiolabel DNA probes for Southern/NorthernDNaseI, αP32dNTP

Pol I functions to repairDNA
E. coli, Pol I mutant are viable but sensitive to UVand chemical mutagens
Essentisl physiological function of Pol I 5’->3’ exonuclease is to excise RNA primers, role in replication

DNA Polymerase III
Pol III is replicase of E. coliHoloenzyme consists of more than 10 subunitsβ subunit confers processivity >5000ntβ subunit form a ring like sliding clampwith 80Å diameter hole, sliding clamp/ β clamp

Properties of E. coli DNAPolymerases

Components of E. coli DNAPolymerase III Holoenzyme
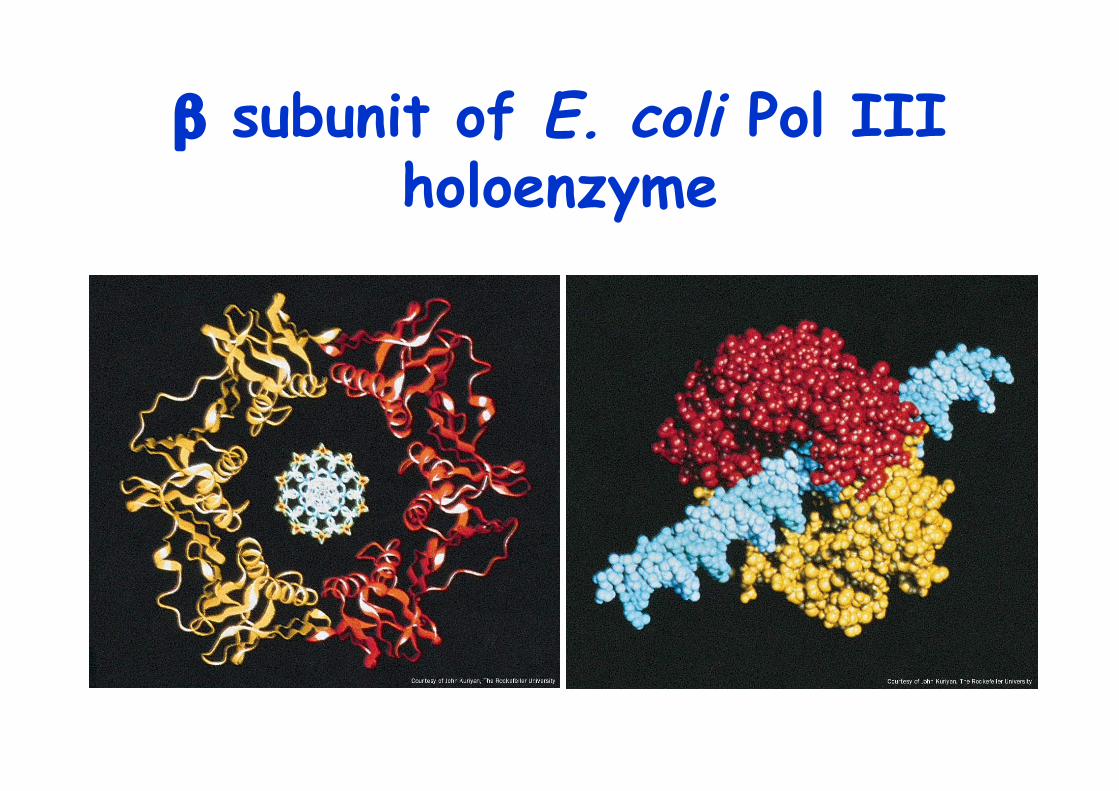
β subunit of E. coli Pol IIIholoenzyme
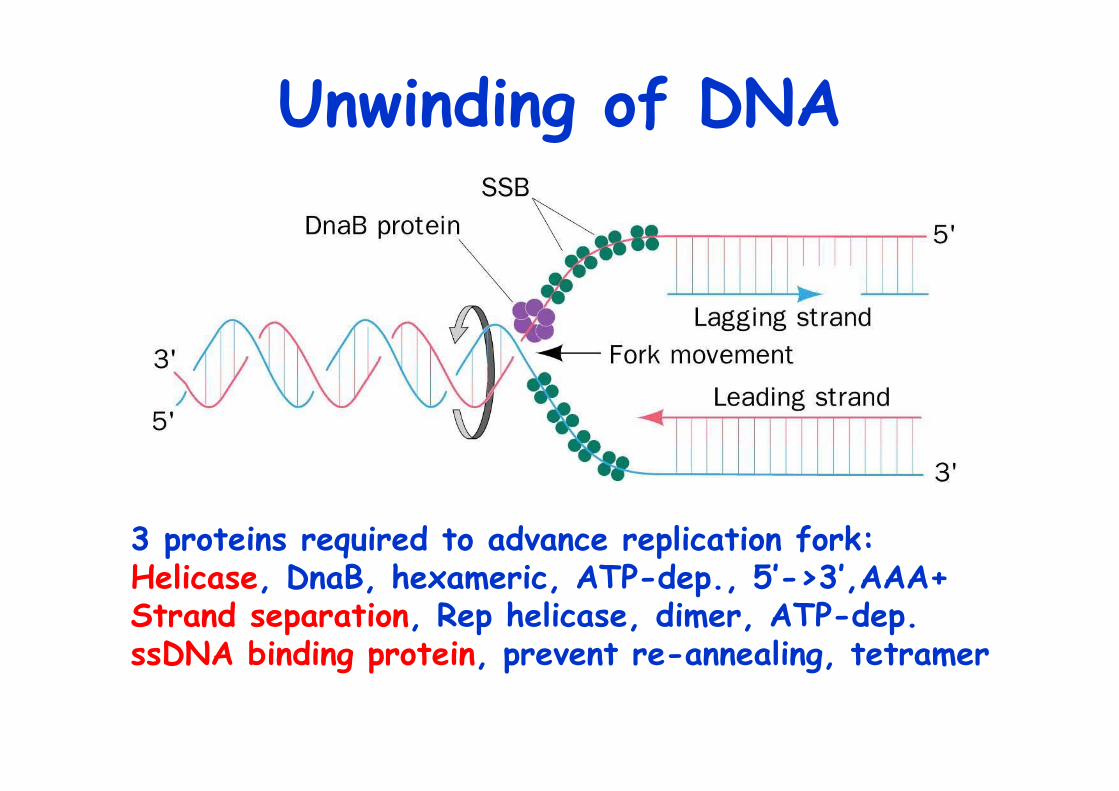
Unwinding of DNA
3 proteins required to advance replication fork:Helicase, DnaB, hexameric, ATP-dep., 5’->3’,AAA+ Strand separation, Rep helicase, dimer, ATP-dep.ssDNA binding protein, prevent re-annealing, tetramer
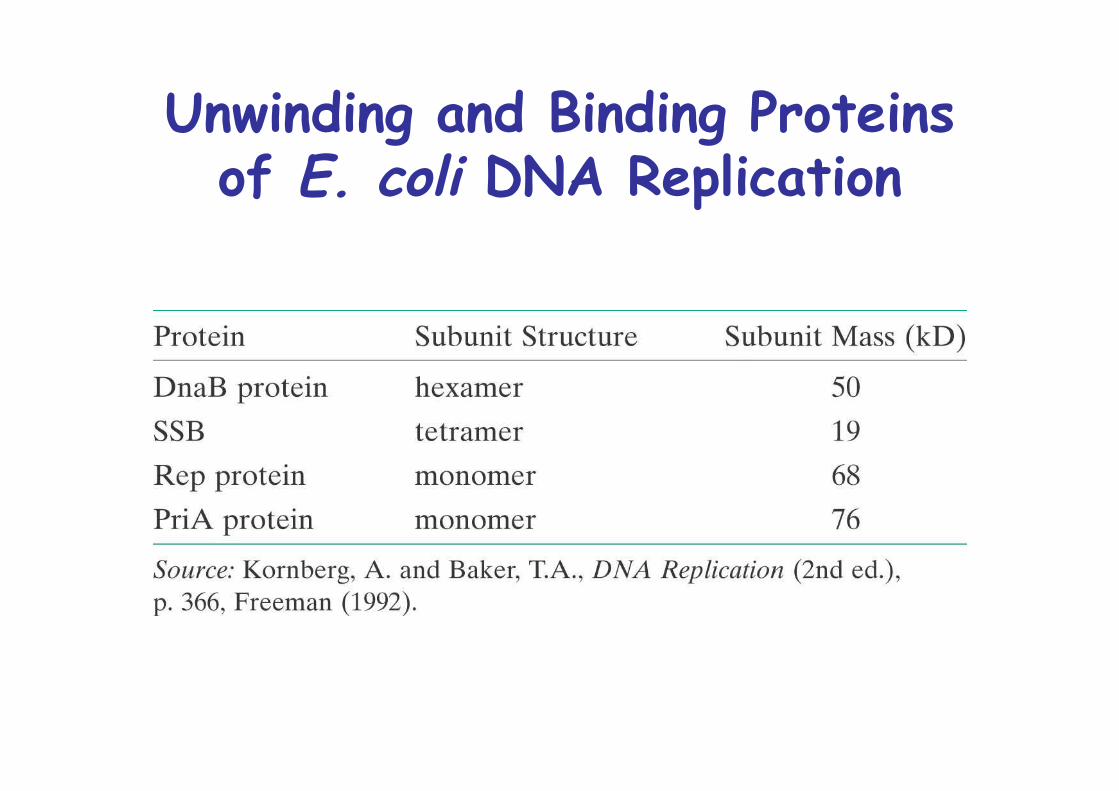
Unwinding and Binding Proteinsof E. coli DNA Replication
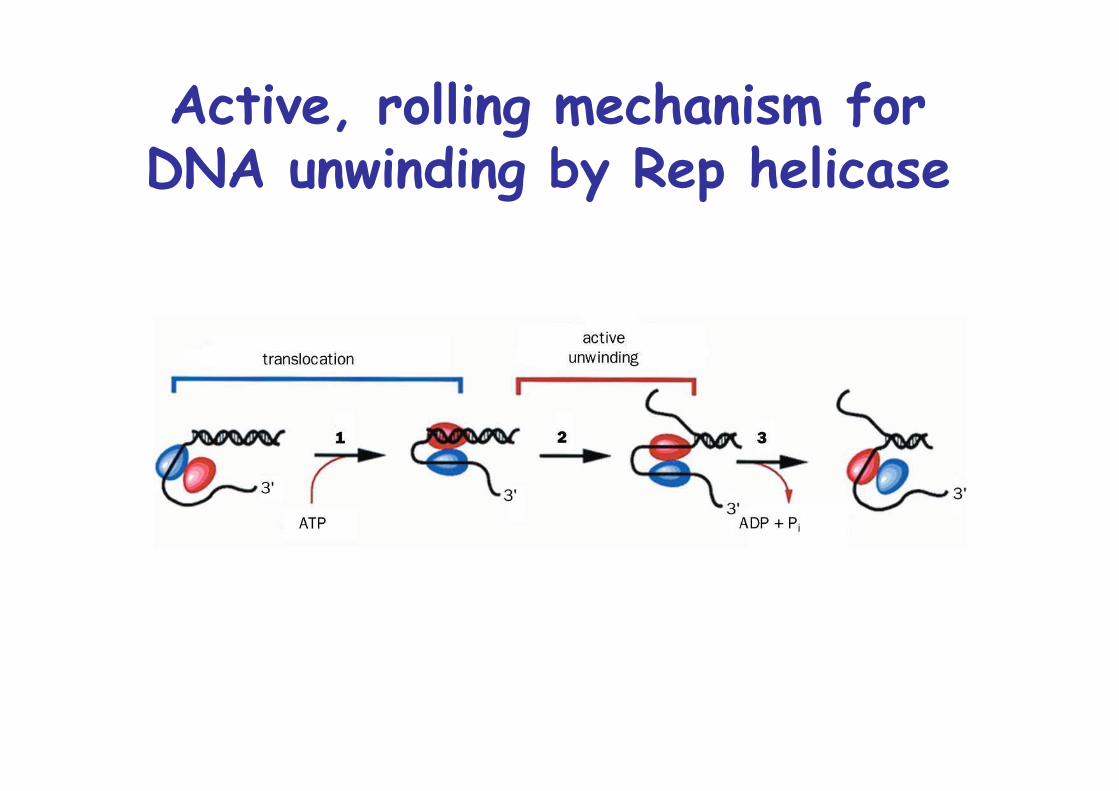
Active, rolling mechanism forDNA unwinding by Rep helicase
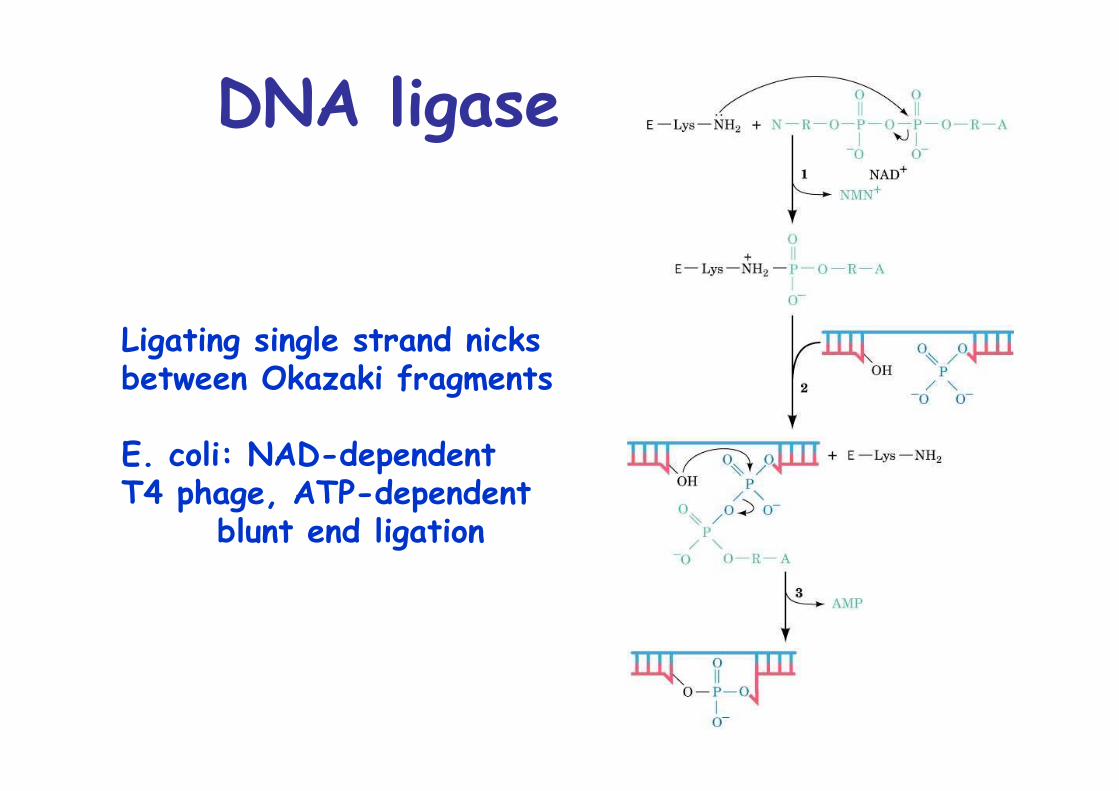
DNA ligase
Ligating single strand nicks between Okazaki fragments
E. coli: NAD-dependentT4 phage, ATP-dependent
blunt end ligation

PrimaseSynthesis of RNA primers fro Okazaki fragments:5’->3’In vitro 11nt ±1

Prokaryotic Replication
BacteriophagesColiphages: M13, φX174
M13: 6408nt ssDNA(+), circularReplication->RFLeading strand synthesis
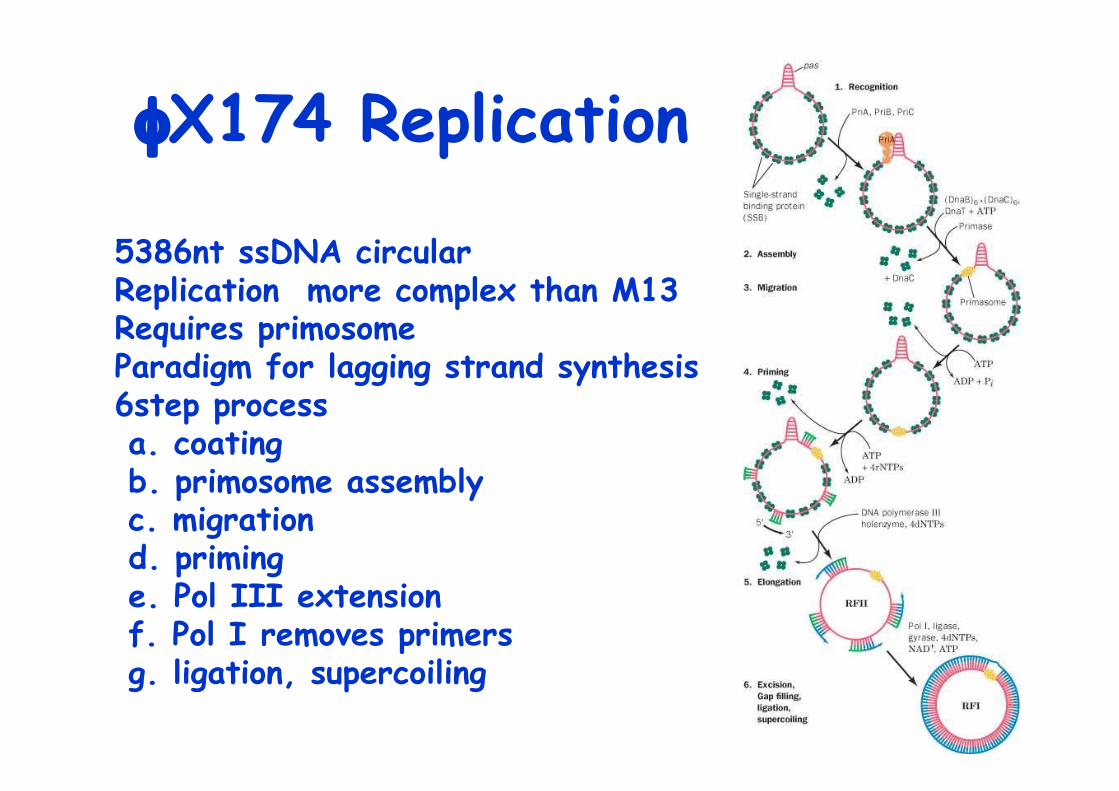
φX174 Replication
5386nt ssDNA circularReplication more complex than M13Requires primosomeParadigm for lagging strand synthesis6step process a. coating b. primosome assembly c. migration d. priming e. Pol III extension f. Pol I removes primers g. ligation, supercoiling
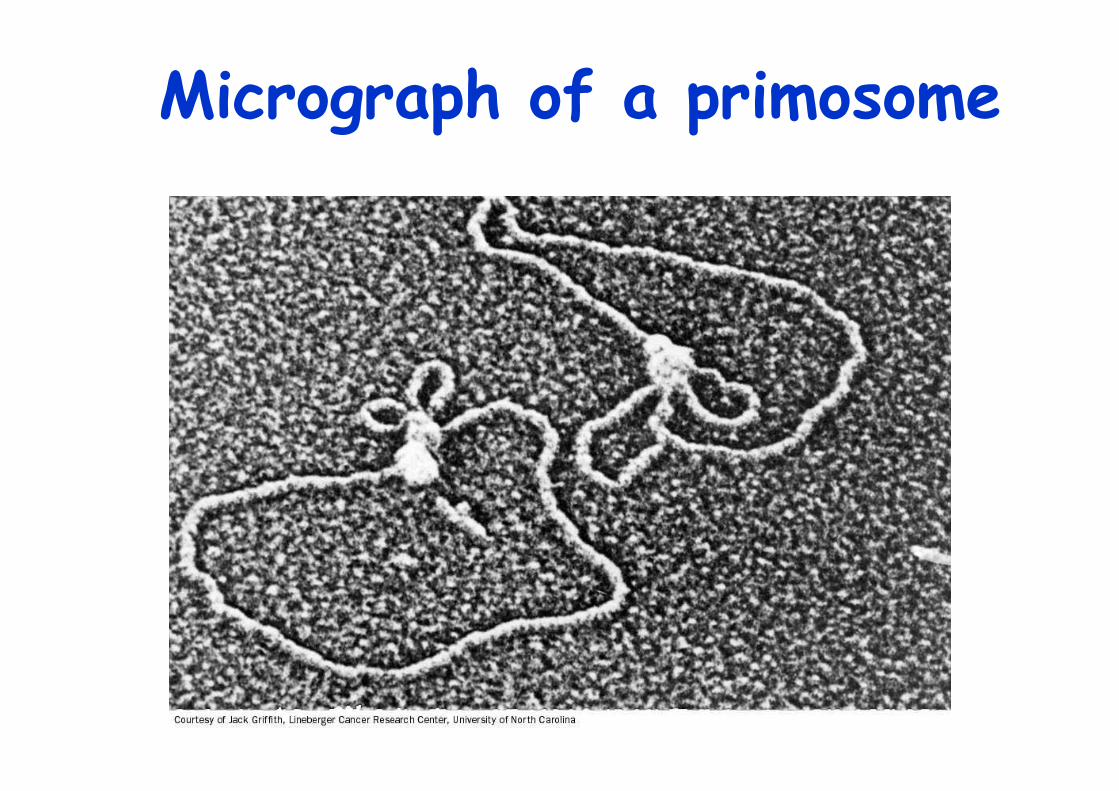
Micrograph of a primosome

Proteins of the Primosomea
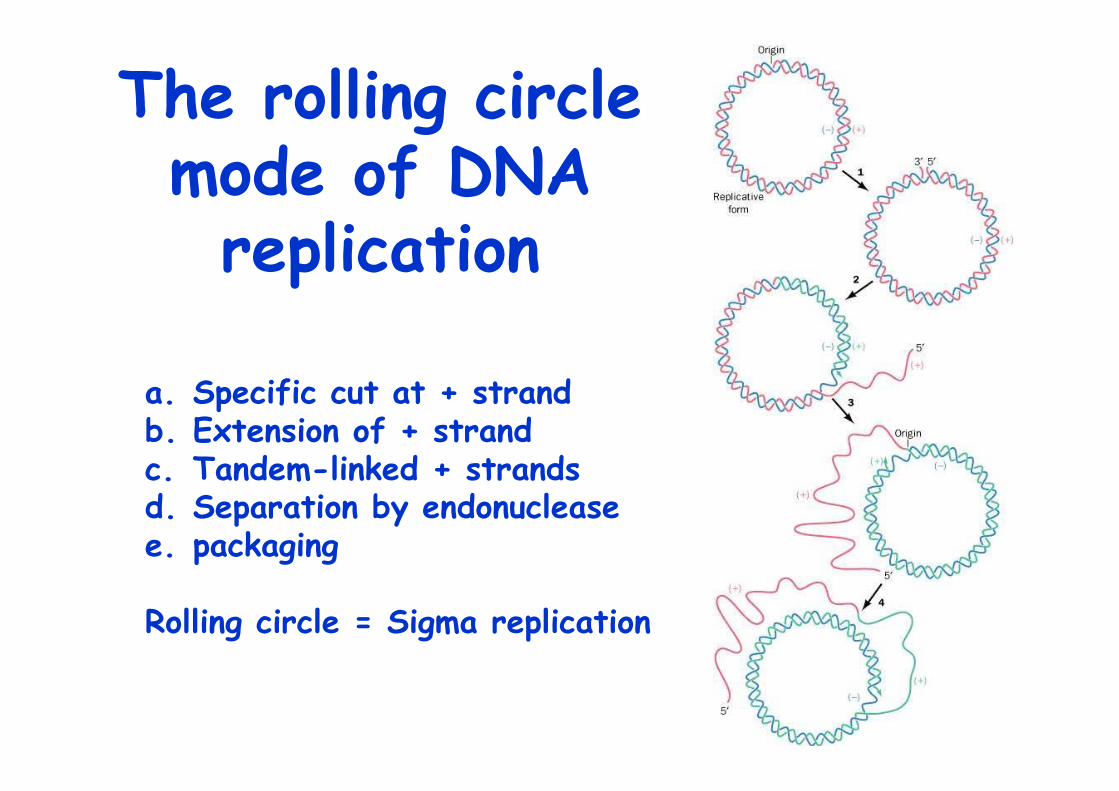
The rolling circlemode of DNAreplication
a. Specific cut at + strandb. Extension of + strandc. Tandem-linked + strandsd. Separation by endonucleasee. packaging
Rolling circle = Sigma replication
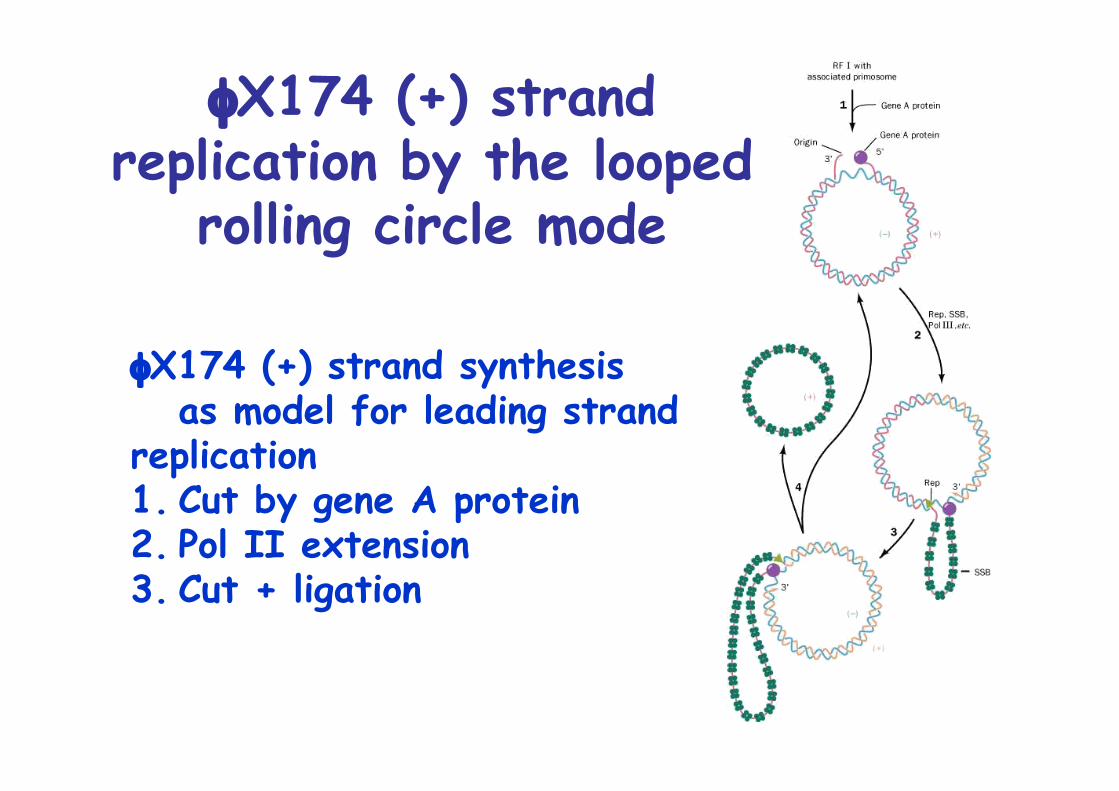
φX174 (+) strandreplication by the looped
rolling circle mode
φX174 (+) strand synthesisas model for leading strand
replication1. Cut by gene A protein2. Pol II extension3. Cut + ligation

The replication of E. coli DNA
Bidirectional, theta replicationleading and lagging strand synthesis occurson a common 900kD multisubunit particle:the replisome -> loop of lagging strand
Initiation: at oriC, 245bp segment
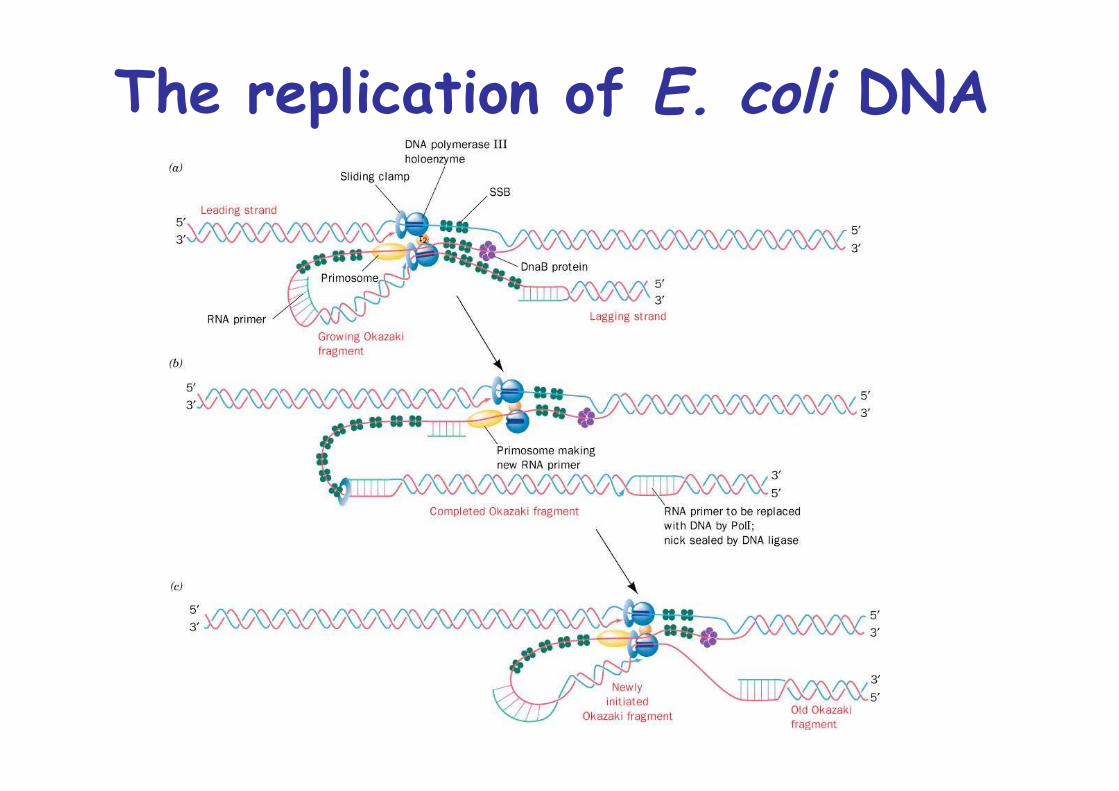
The replication of E. coli DNA
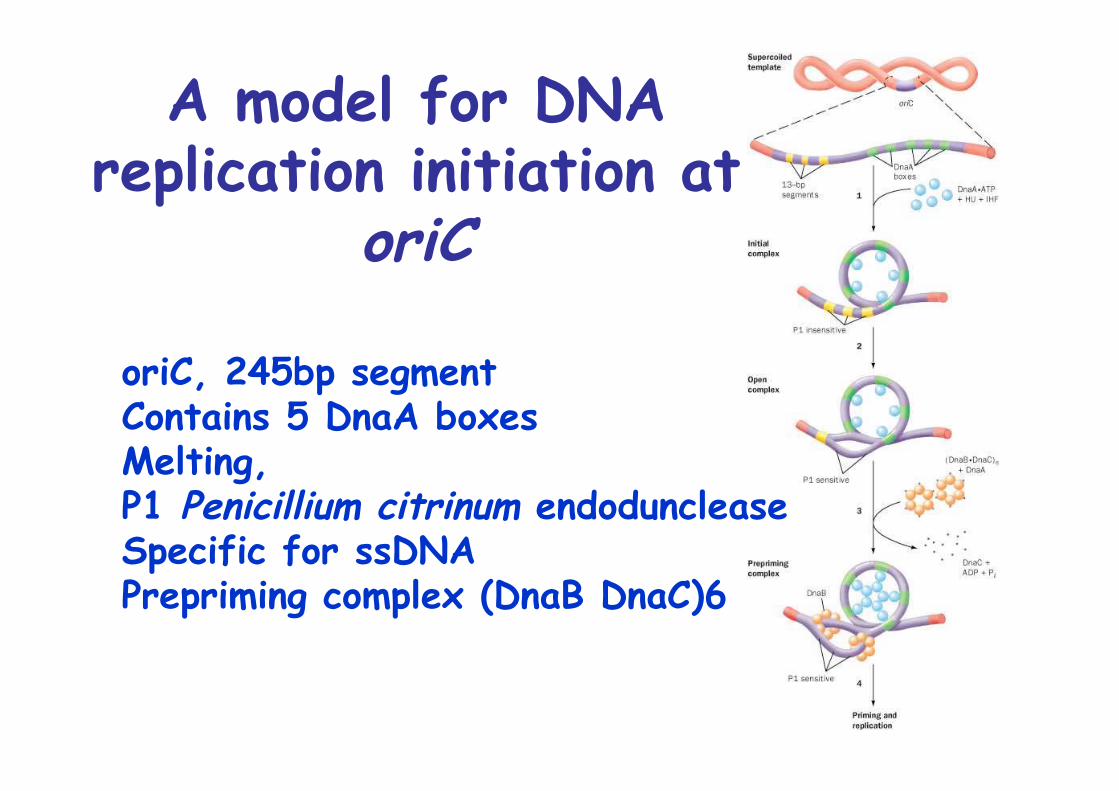
A model for DNAreplication initiation at
oriC
oriC, 245bp segmentContains 5 DnaA boxesMelting, P1 Penicillium citrinum endoduncleaseSpecific for ssDNAPrepriming complex (DnaB DnaC)6
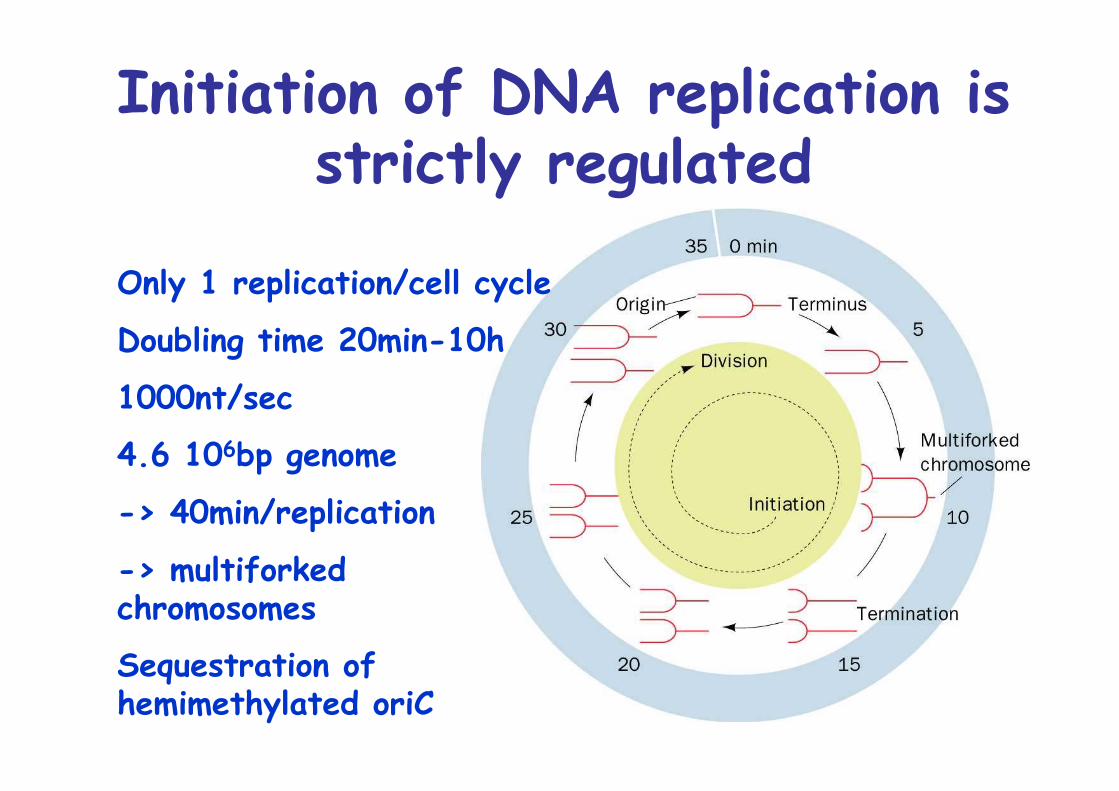
Initiation of DNA replication isstrictly regulated
Only 1 replication/cell cycle
Doubling time 20min-10h
1000nt/sec
4.6 106bp genome
-> 40min/replication
-> multiforkedchromosomes
Sequestration ofhemimethylated oriC
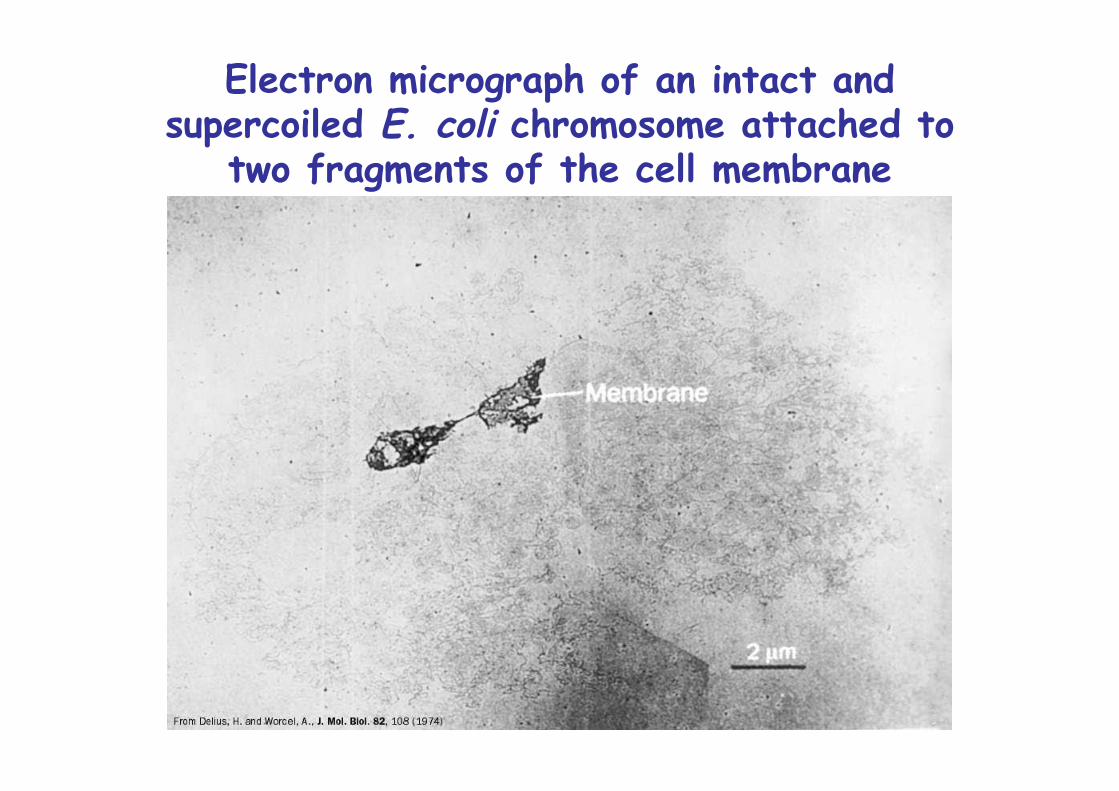
Electron micrograph of an intact andsupercoiled E. coli chromosome attached to
two fragments of the cell membrane
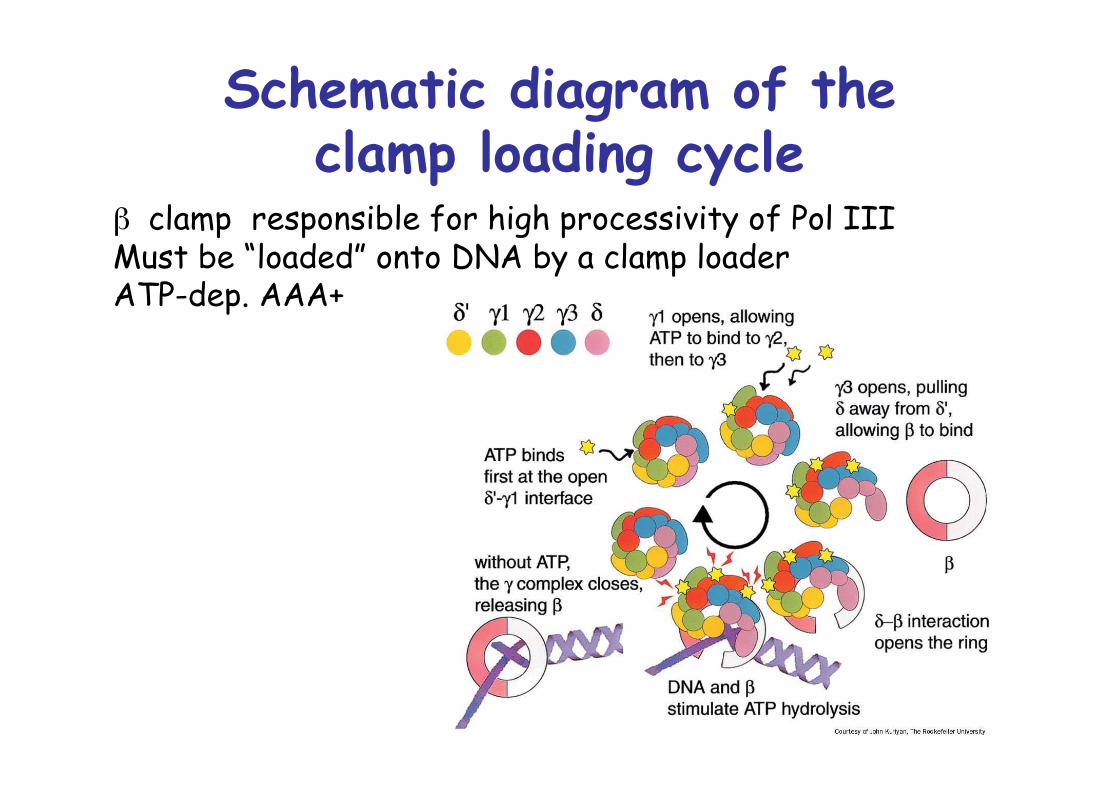
Schematic diagram of theclamp loading cycle
β clamp responsible for high processivity of Pol IIIMust be “loaded” onto DNA by a clamp loaderATP-dep. AAA+
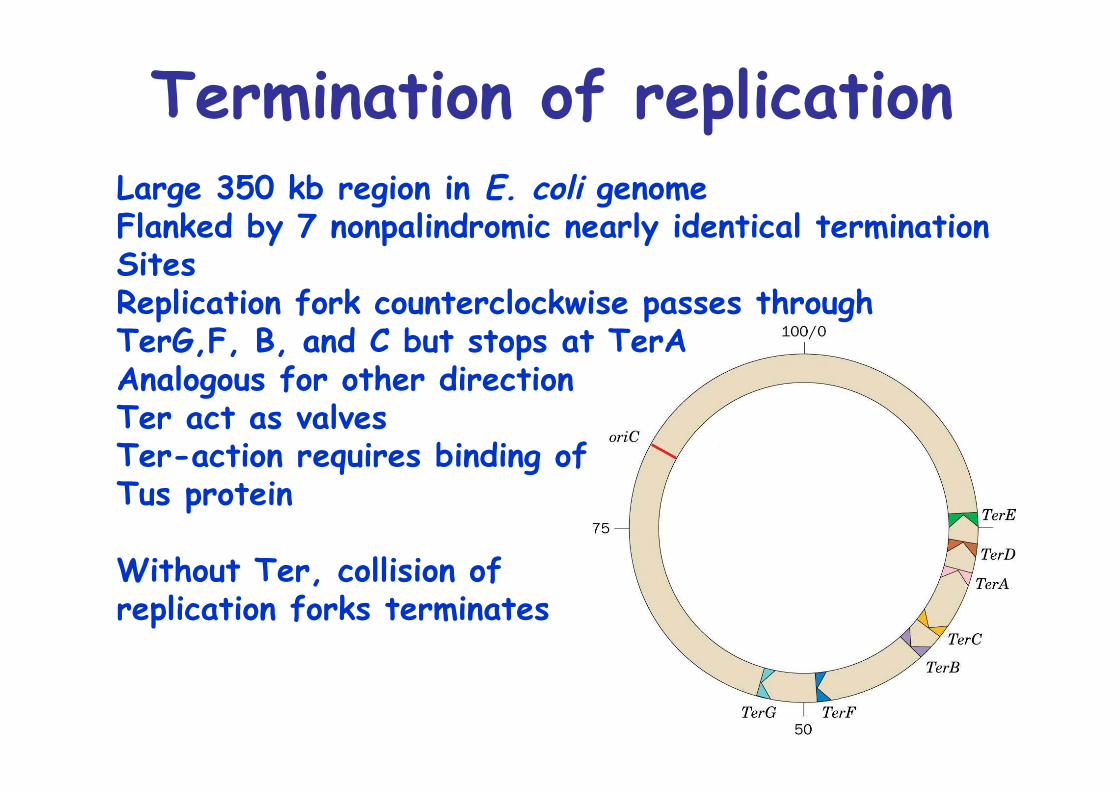
Termination of replicationLarge 350 kb region in E. coli genomeFlanked by 7 nonpalindromic nearly identical termination SitesReplication fork counterclockwise passes through TerG,F, B, and C but stops at TerAAnalogous for other directionTer act as valvesTer-action requires binding ofTus protein
Without Ter, collision of replication forks terminates

Fidelity of Replication
Complexity of replication (>20 proteins) important for high fidelity:T4 phage reversion 10-8 - 10-10
High accuracy due to: 1. Balanced dNTP levels 2. Polymerase reaction itself, pairing 3. 3’->5’ exonuclease of Pol I and Pol III 4. Repair systems -> see later

Why only 5’->3’ synthesis ?
3’->5’ extension would require retention of 5’ triphosphateThis would be lost upon editing !

Eukaryotic Replication
Remarkable degree of similarity to prok. replicationBut linear chromosomes -> ends ?
Cell cycle regulation, can last 8h to > 100 daysMost variation in G1 phase/Go phaseIrreversible decision to proliferate is made in G1CheckpointControlled by cyclins and cyclin-dep. kinases
Best understood from yeast (budding, fission)
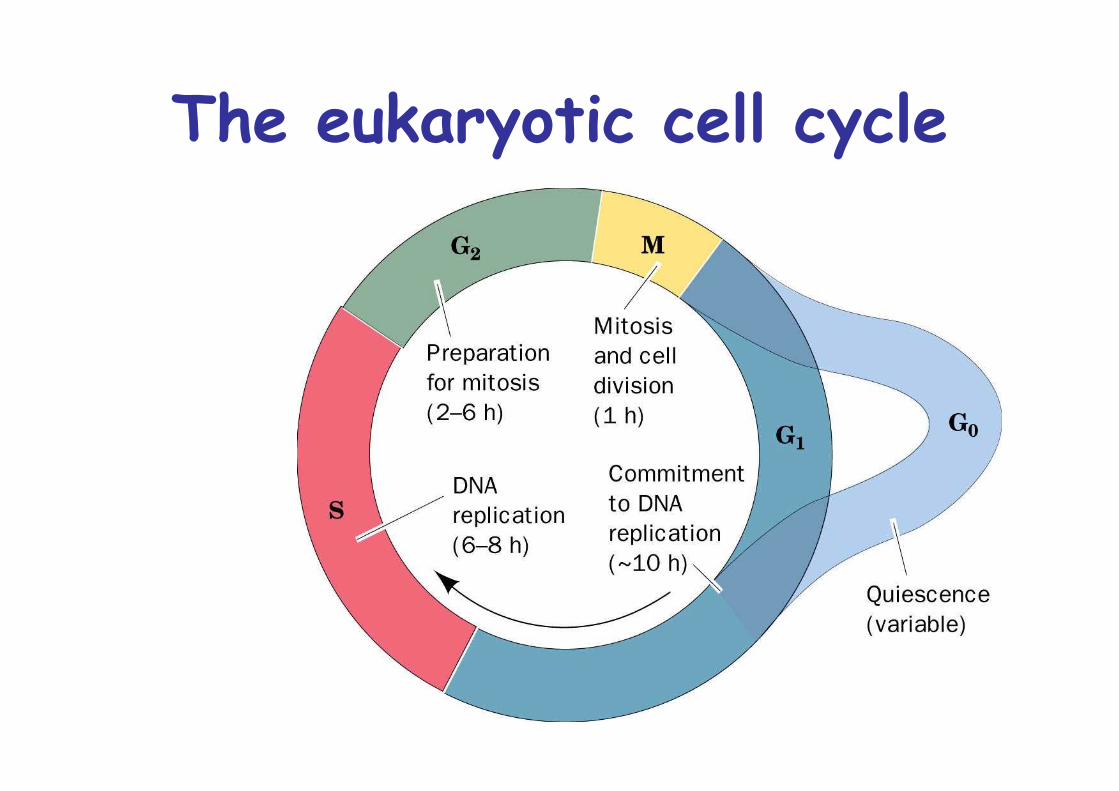
The eukaryotic cell cycle

Eukaryotic cells containmany polymerases
6 families: A, E. coli Pol I, Pol γ (mitochondrial) B, E. coli Pol II, Pol α, Pol δ C, E. coli Pol III D, X, Y
Pol δ, unlimited processivity when incomplex with PCNA, proliferating cellnuclear antigen (systemic lupuserythematosus), β clamp function

Properties of Some AnimalDNA Polymerases

Structure of PCNA

Multiple replication origins, every 3-300kbReplication rate 50nt/sec, 20x slower than E. coliBut 60x more DNAReplication would require 1 monthClusters of 20-80 adjacent repliconsNot simultaneously, but ensure they initiate only once
Eukaryotic chromosomes consistof numerous replicons
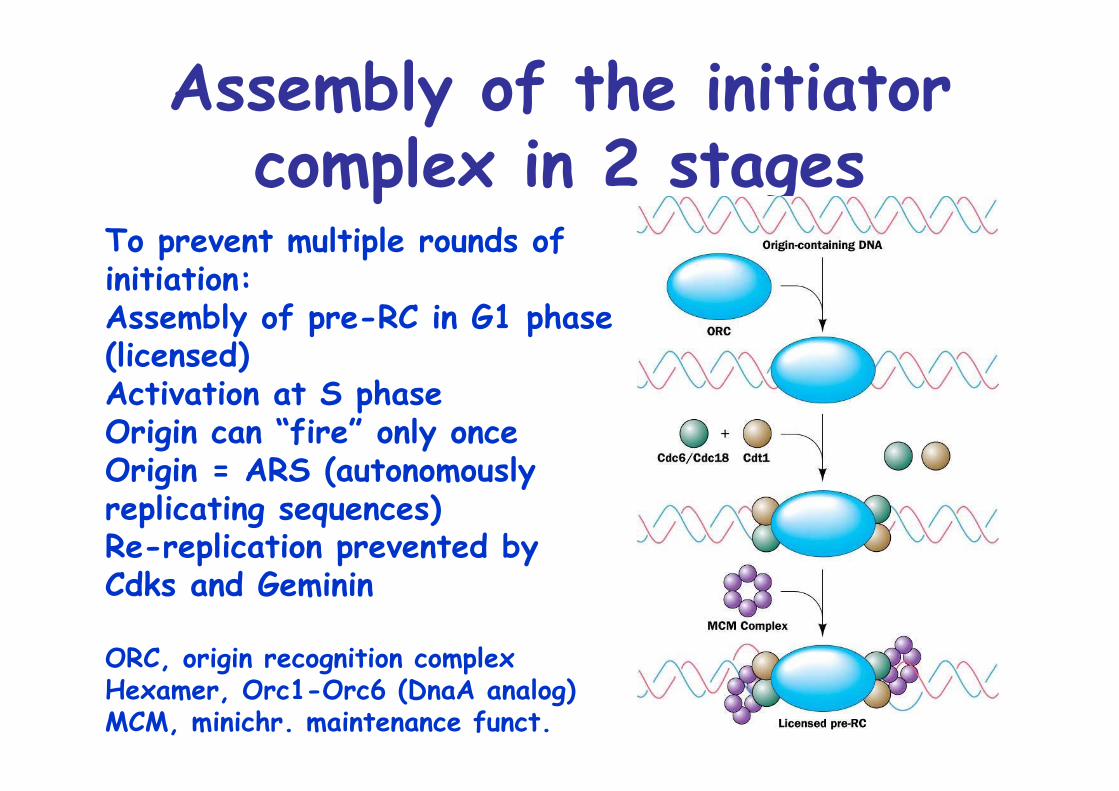
Assembly of the initiatorcomplex in 2 stages
To prevent multiple rounds of initiation:Assembly of pre-RC in G1 phase(licensed)Activation at S phaseOrigin can “fire” only onceOrigin = ARS (autonomously replicating sequences)Re-replication prevented byCdks and Geminin
ORC, origin recognition complexHexamer, Orc1-Orc6 (DnaA analog)MCM, minichr. maintenance funct.

Removal of RNA primers
2 enzymes: RNase H1, removes most of the RNA leaving a single 5’ ribonucleotide (H, hybrid)
Flap endonuclease-1 (FEN1) removes single single 5’ ribonucleotide
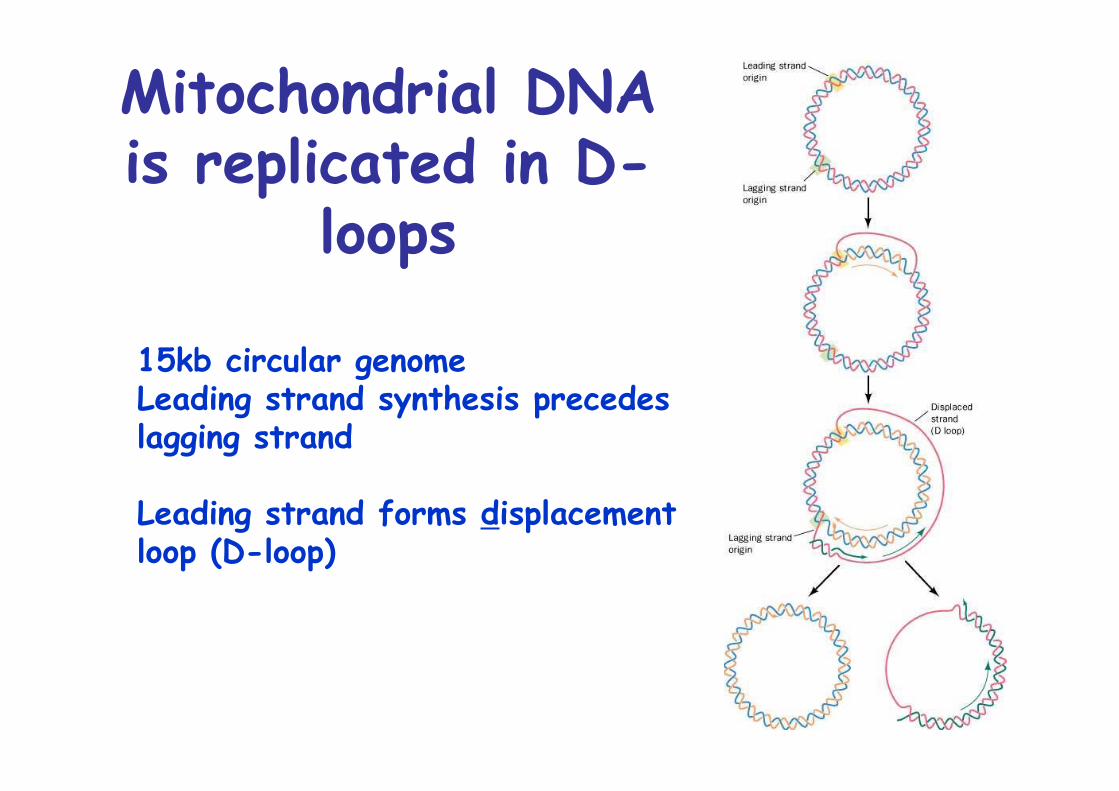
Mitochondrial DNAis replicated in D-
loops
15kb circular genomeLeading strand synthesis precedes lagging strand
Leading strand forms displacement loop (D-loop)

Reverse transcriptaseRetroviruses:
RNA containing eukaryotic viruses, e.g. HIVReplicate from RNA genomeCopy RNA into DNA by Reverse Transcripase (RT)
Similar to Pol I, 5’->3’ synthesis of DNA from RNA template, primed by host tRNARNA is degraded by RNase HssDNA directs dsDNA synthesisdsDNA integration into host genome
RT: important tool for cDNA synthesis, oligo-dT primed
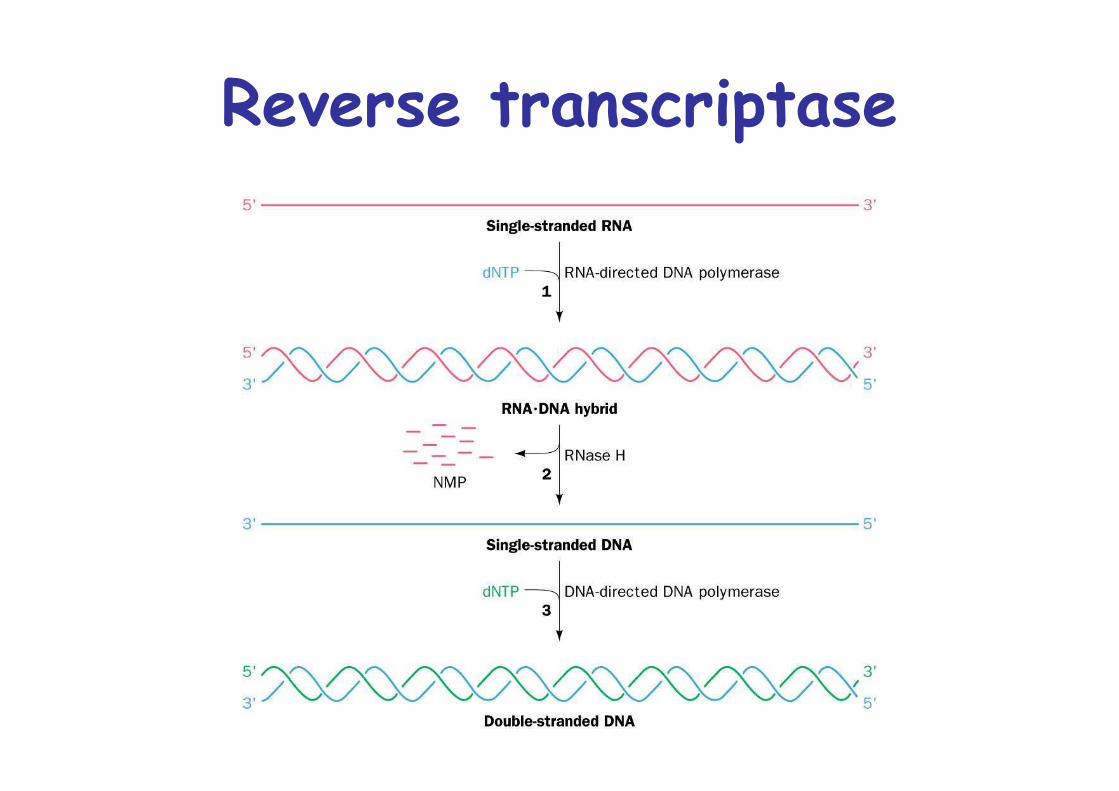
Reverse transcriptase

Structure of HIV-1reverse transcriptase
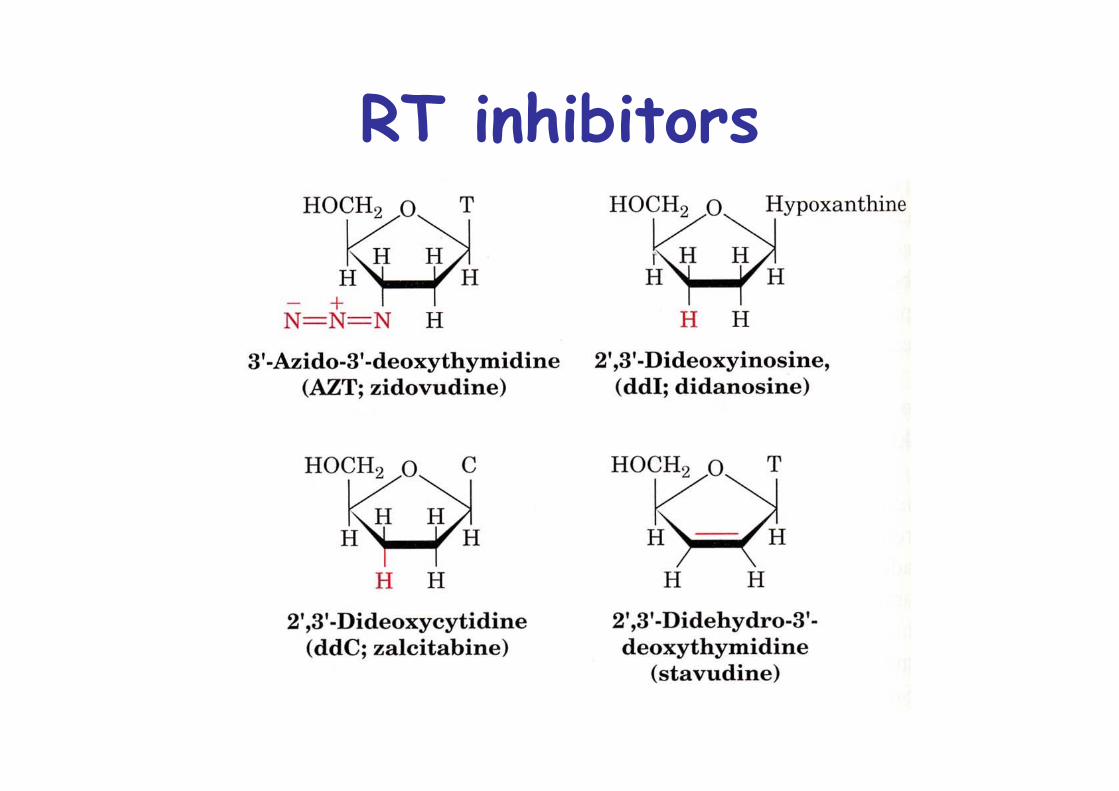
RT inhibitors

Telomers and Telomerase
How are the ends of linear chromosomes replicated ?
Problem: no priming at 5’ of lagging strand possible without shortening of the chromosome upon every replication
Telomer sequence: unusual, G-rich, 3’ overhang (20-200bp)
Specialized enzyme: telomerase adds G-rich repeats without teplate, is ribonucleoprotein, RNA acts as template
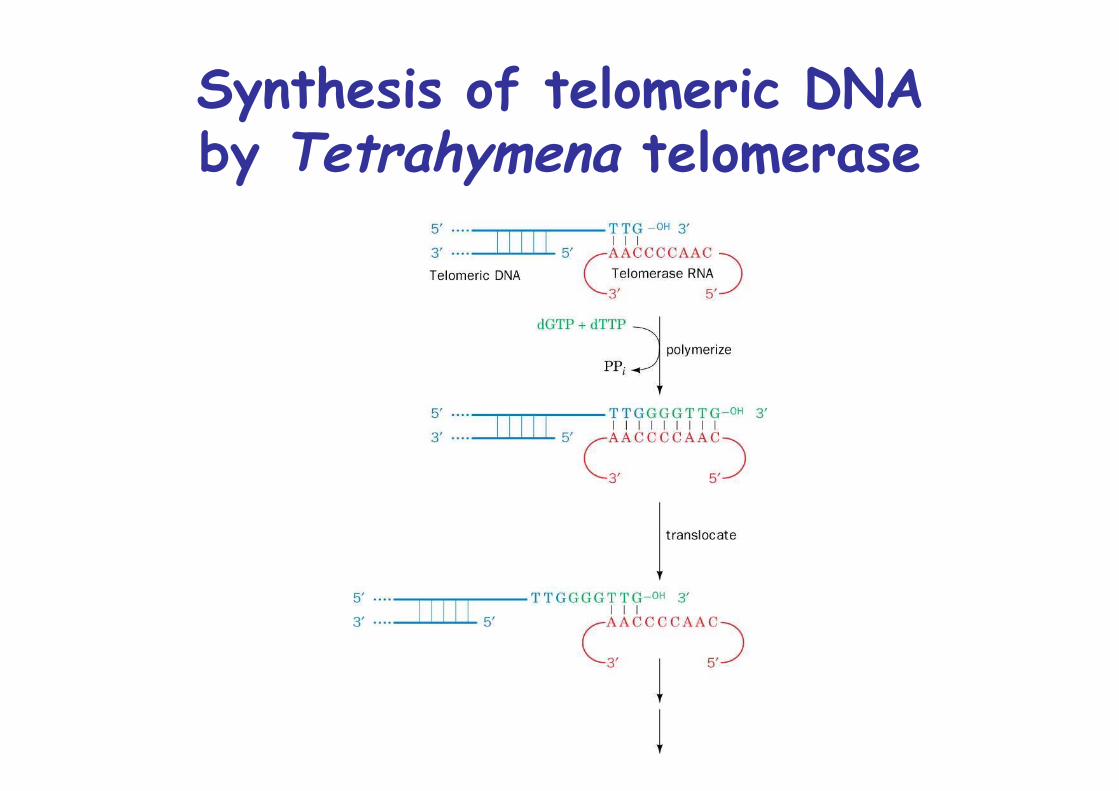
Synthesis of telomeric DNAby Tetrahymena telomerase

Telomers must be cappedWithout telomerase, chromosome would shorten 50-100ntwith every cell division
Exposed telomeric ssDNA must be protected by cappingwith proteins, Pot1

Telomere length correlateswith aging
Primary cells in culture die after 20-60 divisions
Such somatic cells have no telomerase activity ->Telomers shorten with every divisionTelomerase is active only in germ cells
Analysis of fibroblast from donors of different age:No correlation with numbers of doublings in cultureBut correlation of telomer length with numbers of doublings
Progeria: premature aging diseasepatients have short telomers

Cancer cells have activetelomerase
Why do somatic cells down regulate telomerase ?Senescence may be a mechanism to protect from cancer
All immortal cells express telomerase
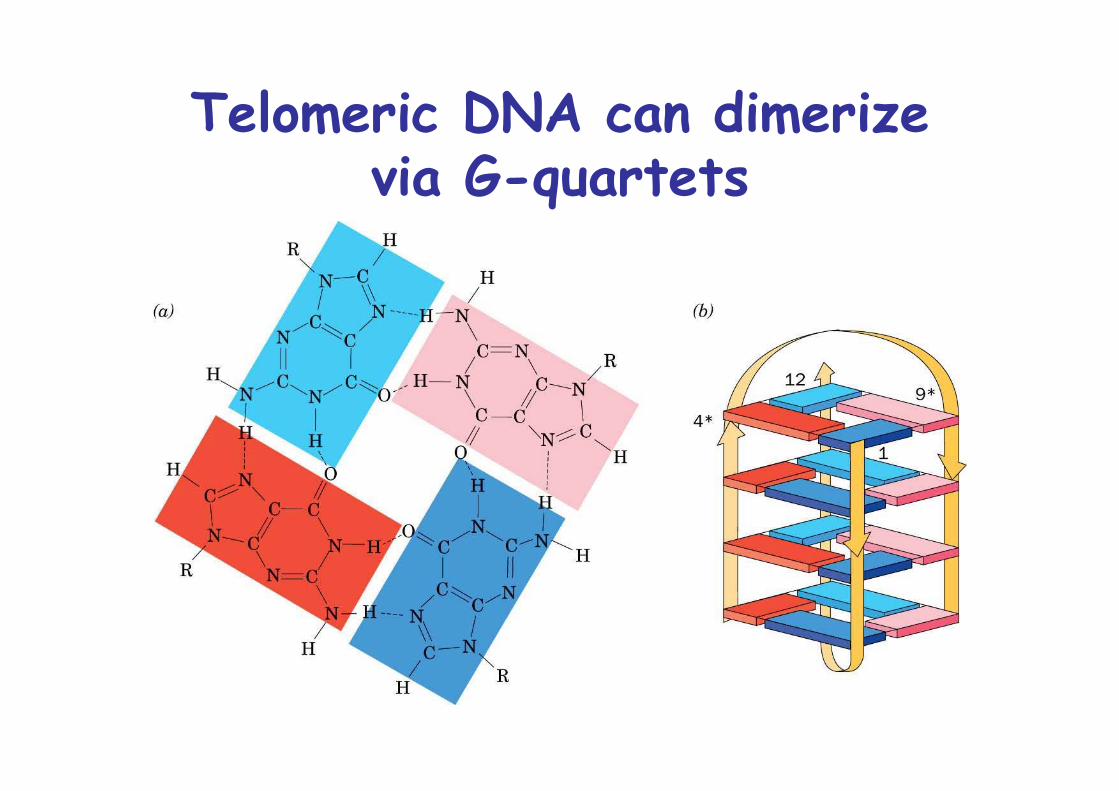
Telomeric DNA can dimerizevia G-quartets
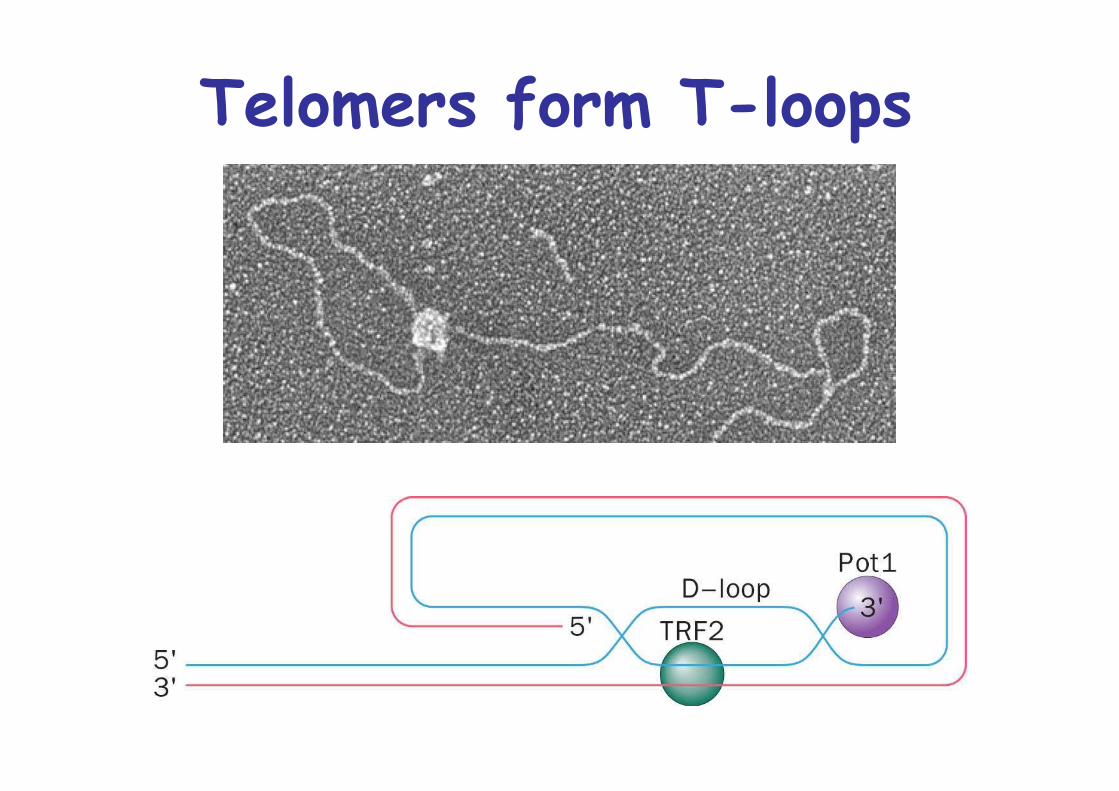
Telomers form T-loops

Repair of DNA
DNA is not inert
UV radiation, ionizing radiation, toxic chemicals, oxidative metabolism can harm DNA
Spontaneous hydrolysis of 10’000 glycosidic bonds in every cell every day....
Human genome 130 genes dedicated to DNA repairChemically similar in E. coli
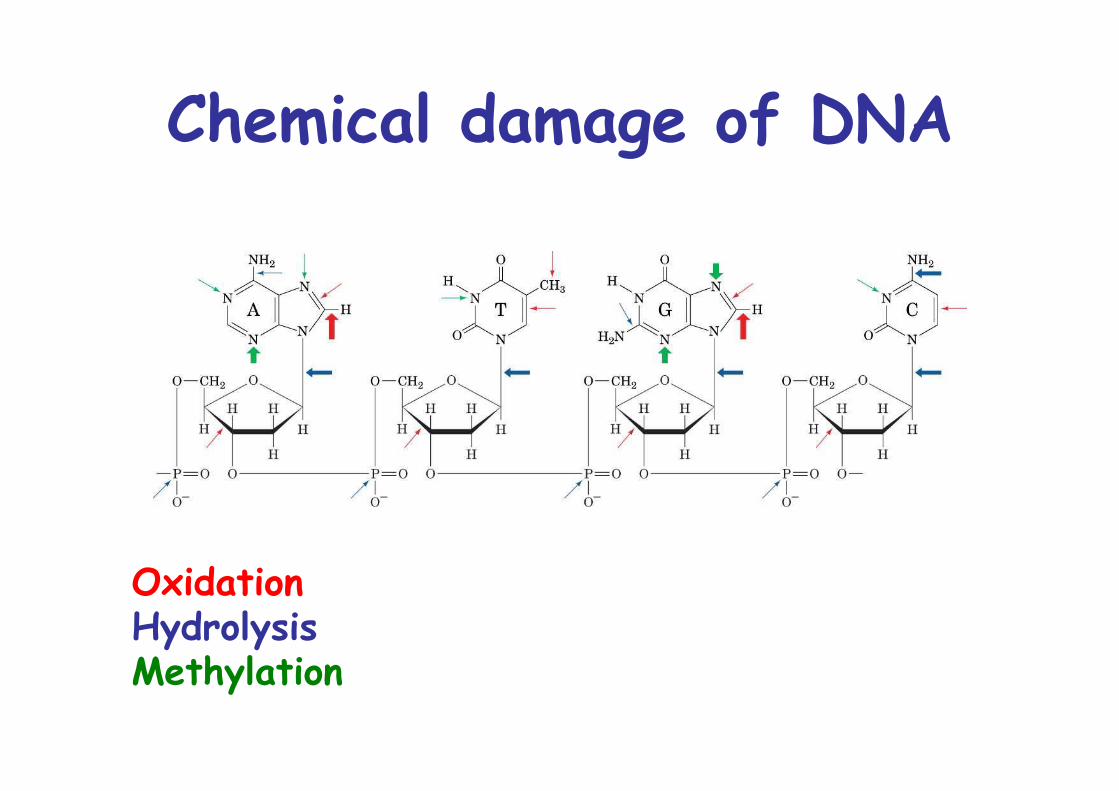
Chemical damage of DNA
OxidationHydrolysisMethylation
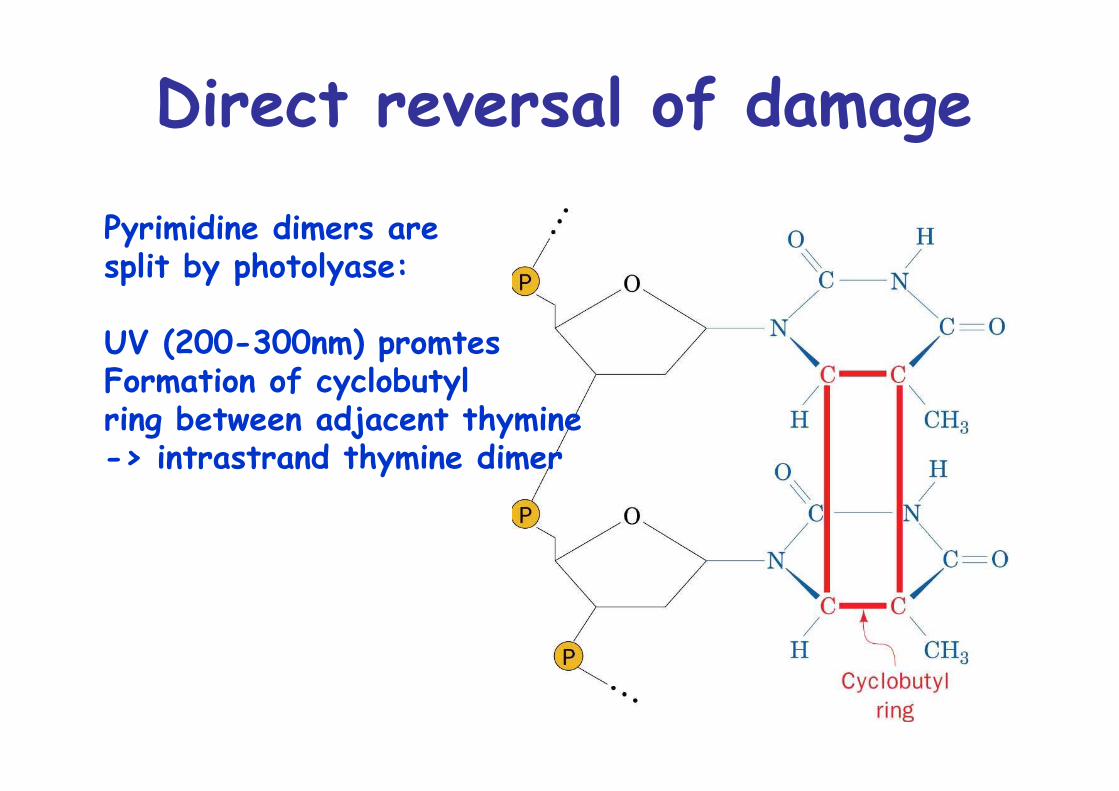
Direct reversal of damage
Pyrimidine dimers are split by photolyase:
UV (200-300nm) promtesFormation of cyclobutyl ring between adjacent thymine-> intrastrand thymine dimer

DNA photolyase
Photoreactive enzyme:Absorbed light is transferred to FADH-
Electron used to split thymine dimer
Base flipping:Often used to repair damaged DNA

Excision repair
Cells have two types of excision repair:1. Nucleotide excision repair, NER
repairs bulky lesions2. Base excision repair, BER
repairs nonbulky lesions involving a single base
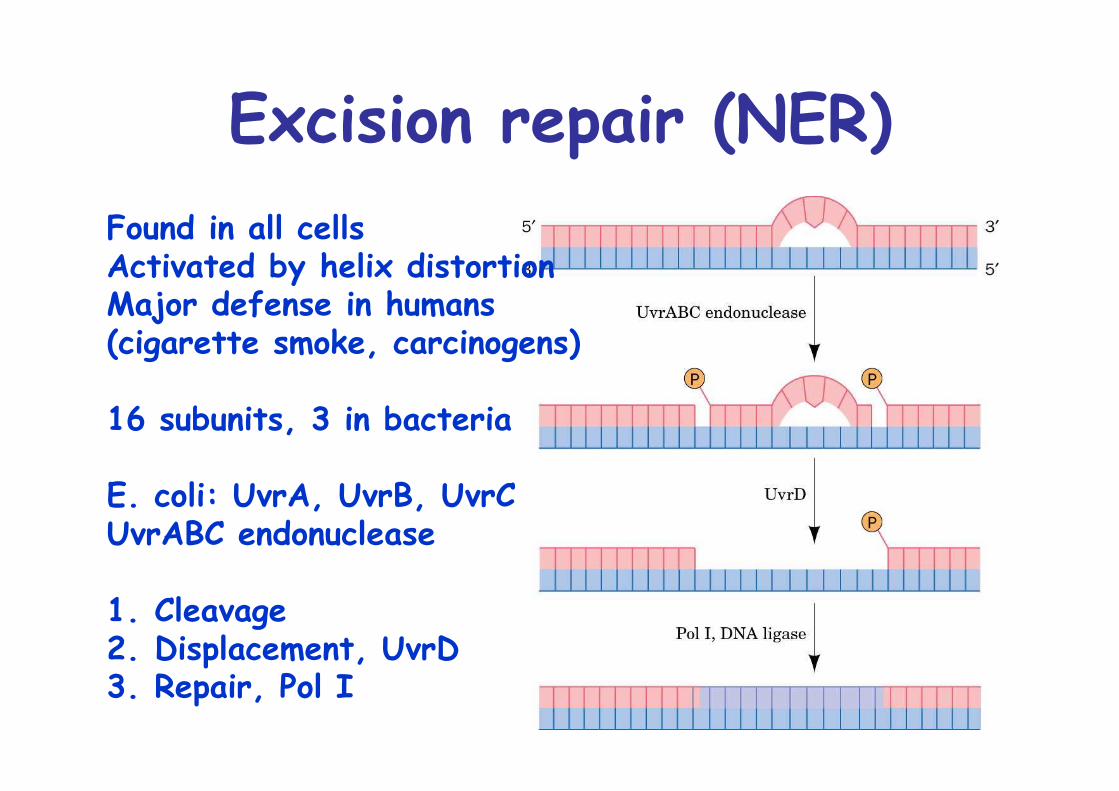
Excision repair (NER)Found in all cellsActivated by helix distortionMajor defense in humans(cigarette smoke, carcinogens)
16 subunits, 3 in bacteria
E. coli: UvrA, UvrB, UvrCUvrABC endonuclease
1. Cleavage2. Displacement, UvrD3. Repair, Pol I

NER diseasesXenoderma pigmentosum
skin cells cannot repair UV damageIndividuals extremely sensitive to sun lightskin tumors risk 2000-fold elevatedcultured skin cells are defective in repairingtymidine dimersCell fusion experiments: 8 complementation groups
Cockayne syndromelight sensitive and neurological defectsdemyelination-> oxidative damage in neurons
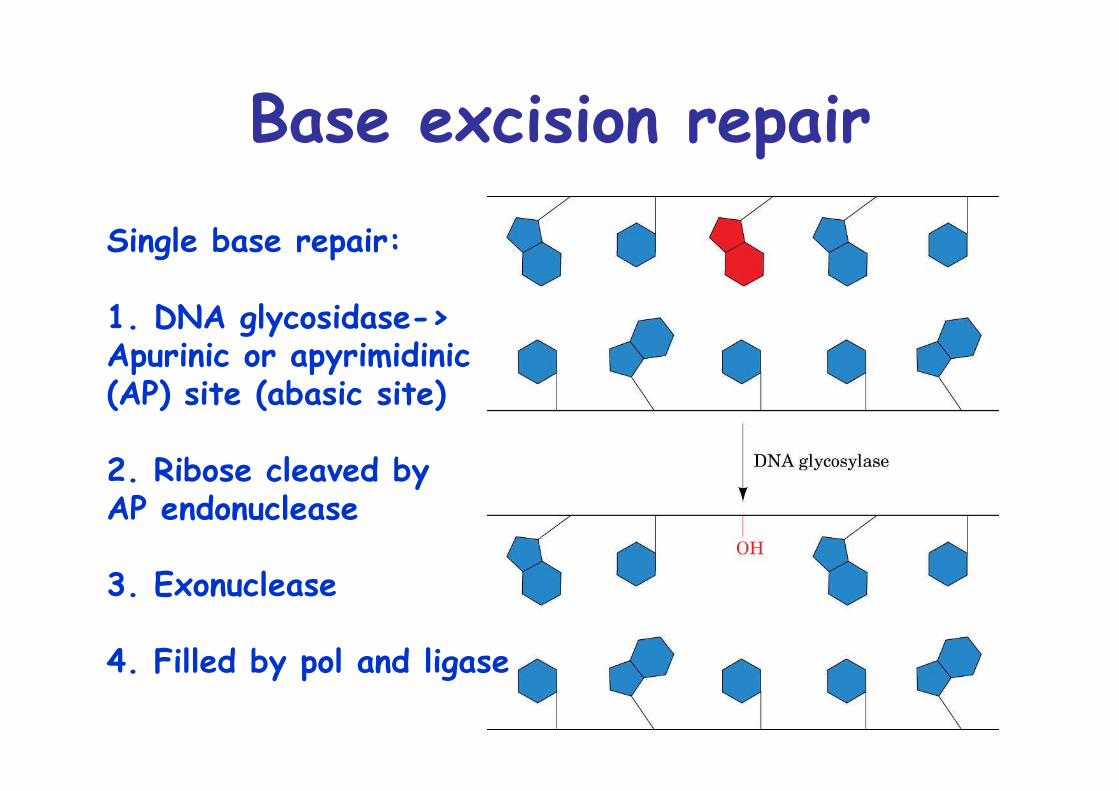
Base excision repairSingle base repair:
1. DNA glycosidase->Apurinic or apyrimidinic(AP) site (abasic site)
2. Ribose cleaved byAP endonuclease
3. Exonuclease
4. Filled by pol and ligase

Uracil in DNA would be highlymutagenic
Why use thymine in DNA and uracil in RNA ?
Cytosine deaminates to uracil
If U in DNA: no way to discriminate whetherG-U mismatch is due to:
G-C -> deaminated to UA-U
Since T is normal in DNA, every U is due to deaminated C

Mismatch repair
Replicational mispairing is repaired by mismatch repair (MMR)
Defects result in hereditary nonpolyposis colorectal cancer (HNPCC)
Must distinguish between correct and wrong baseIn E. coli, possible due to hemimethylation3 proteins, MutS, MutL, MutH
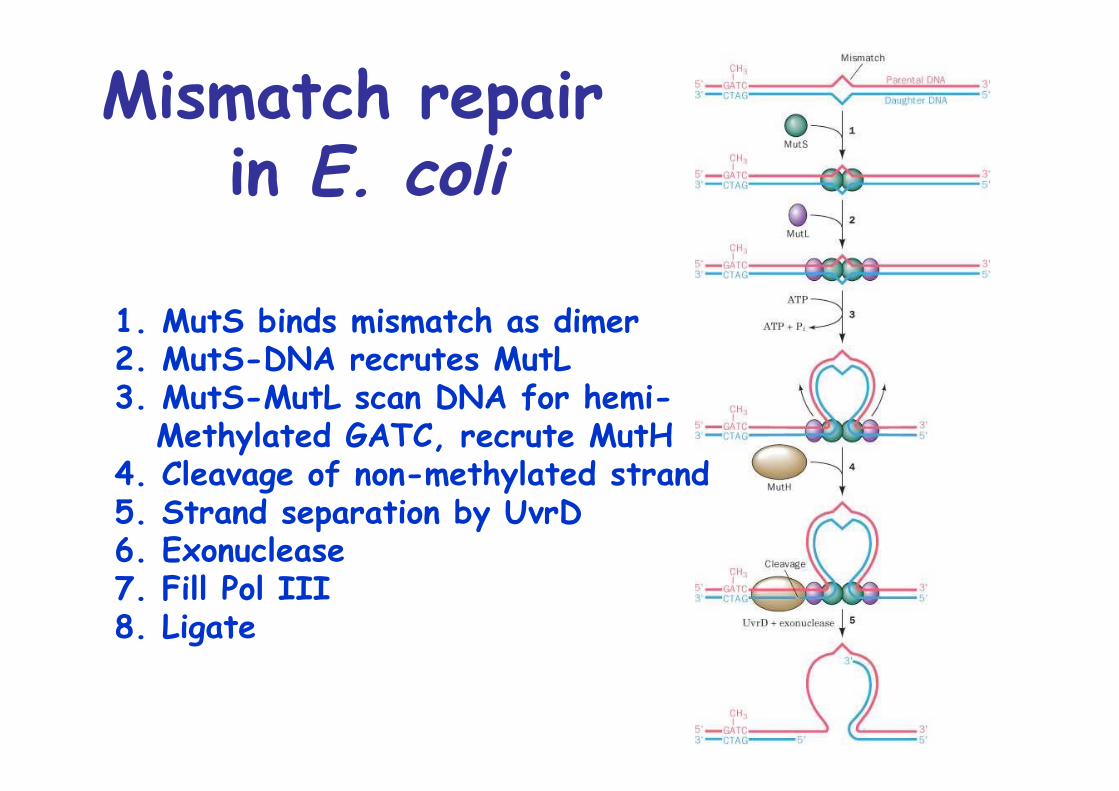
Mismatch repair in E. coli
1. MutS binds mismatch as dimer2. MutS-DNA recrutes MutL3. MutS-MutL scan DNA for hemi- Methylated GATC, recrute MutH4. Cleavage of non-methylated strand5. Strand separation by UvrD6. Exonuclease7. Fill Pol III8. Ligate

The SOS response
On heavy DNA damage, E. coli stops to grow and inducesDNA repair system, SOS system
SOS operon, recA, uvrA, uvrB repressed by LexA
RecA is ssDNA binding protein, induces cleavage of LexA upon ssDNA binding -> release repression of SOS operon
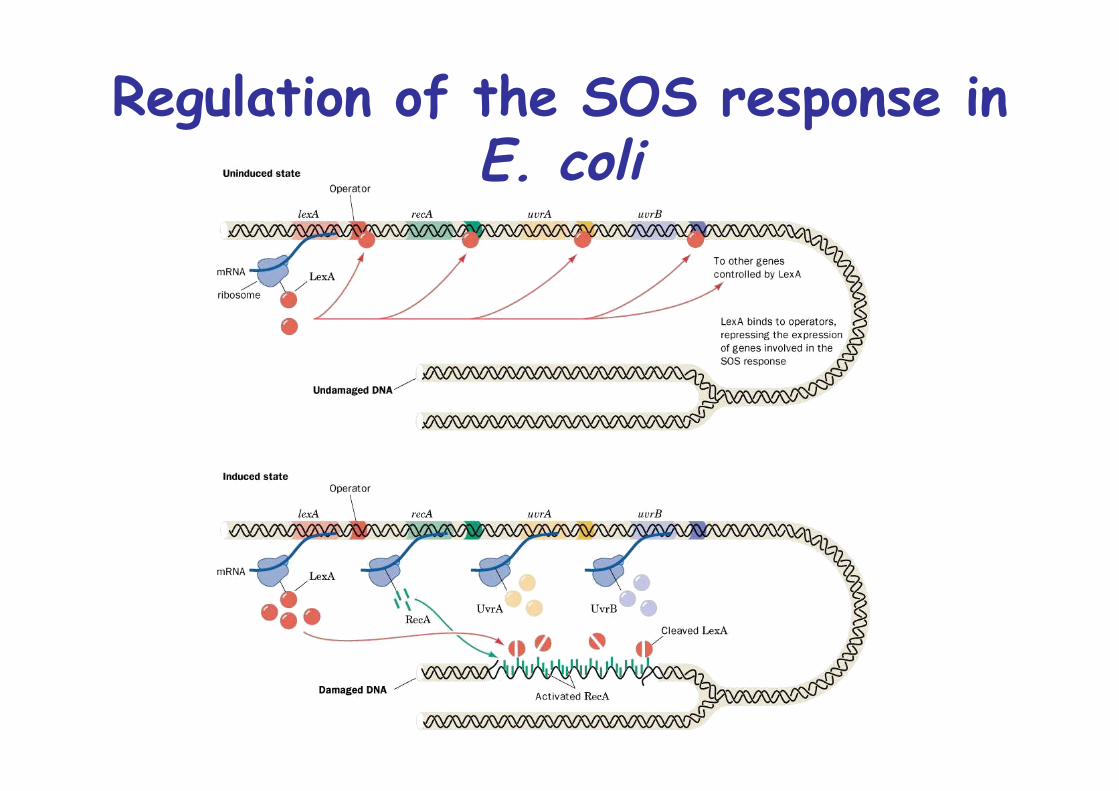
Regulation of the SOS response inE. coli

SOS repair is error prone
If replisome encounters DNA lesion:Stallment, relase Pol III core, collapse of replication fork
To resume: either SOS repair or recombination repair
Recombination repair: circumvents lesion and uses homologous recombination to restore damaged site (->later)
In SOS repair, Pol III is replaced by bypass DNA polymerase, Pol IV or Pol VError prone polymerases -> SOS response is mutagenic ->Adaptation to difficult situation by generating diversity

Double-strand break repair
Ionizing radiation and free radicals can induce double strand breaks in DNA (DSB)Also induced by some cellular processes, e.g. VDJ recomb.
2 ways to repair DSBs:1. Recombination repair-> later2. Nonhomologous end-joining (NHEJ)
involves DNA end binding protein Ku
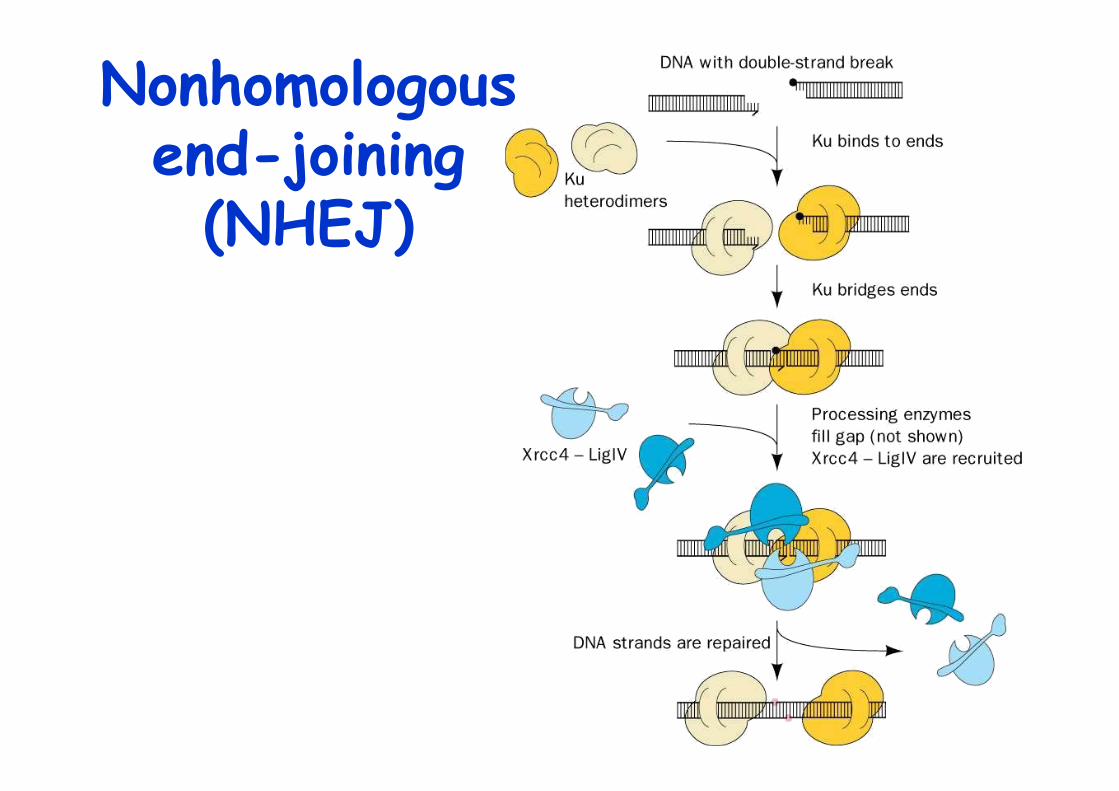
Nonhomologousend-joining
(NHEJ)

Identification ofcarcinogens
Many forms of cancers are caused by exposure to certain chemical agents, carcinogens (man-made or natural)
Ames test assay for carcinogenicity
Salmonelle typhimuriumhis- incubate with chemical -> rate of reversionto his+ correlates with mutagenecity of tested chemical

The Ames test formutagenesis
Filter disc containing Substance:
1. Zone lethal2. Zone mutagenic3. Zone spontaneous
reversion

Recombination and mobilegenetic elements
Pairs of allelic genes may exchange chromosomal locationby genetic recombination via homologous recombination
Homologous recombination:Exchange of homologous segments between two DNA molecules
Bacteria, haploid, exchange via conjugation (mating) orTransduction (viral)

The Holliday model ofhomologous recombination
1. ssDNA nick2. Strand invasion3. Branch migration4. Holliday interm.
Chi structure5. Resolution
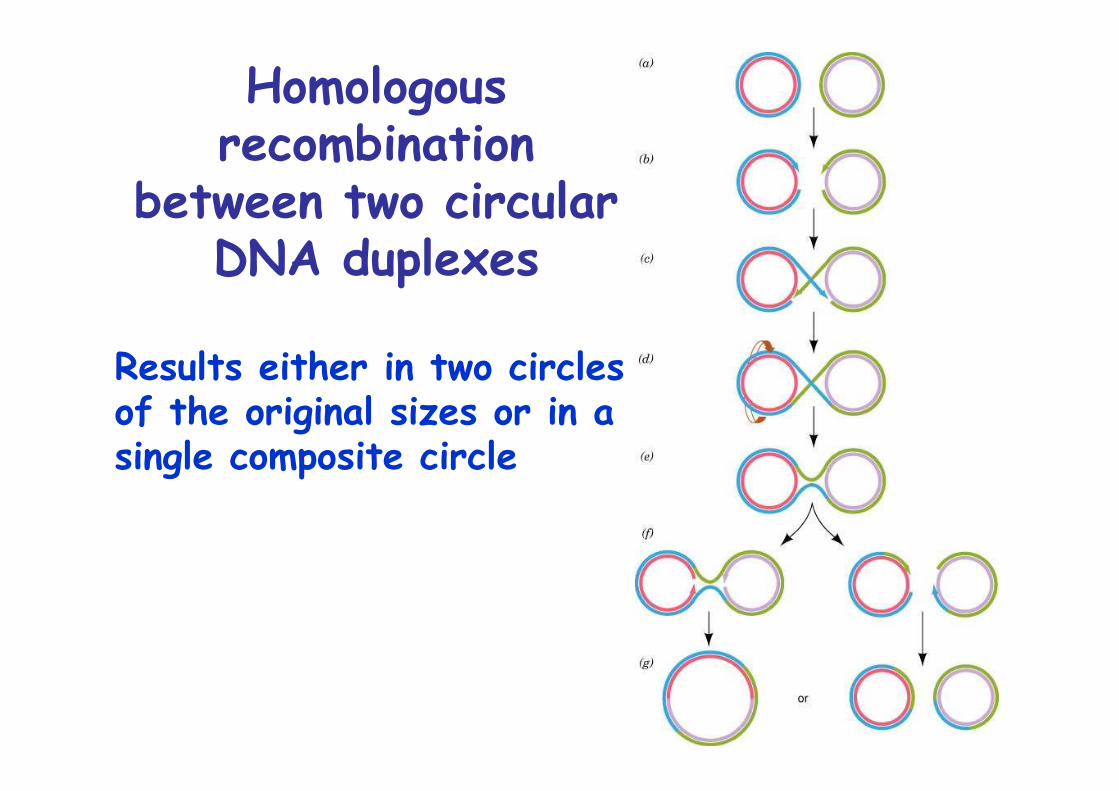
Homologousrecombination
between two circularDNA duplexes
Results either in two circles of the original sizes or in asingle composite circle
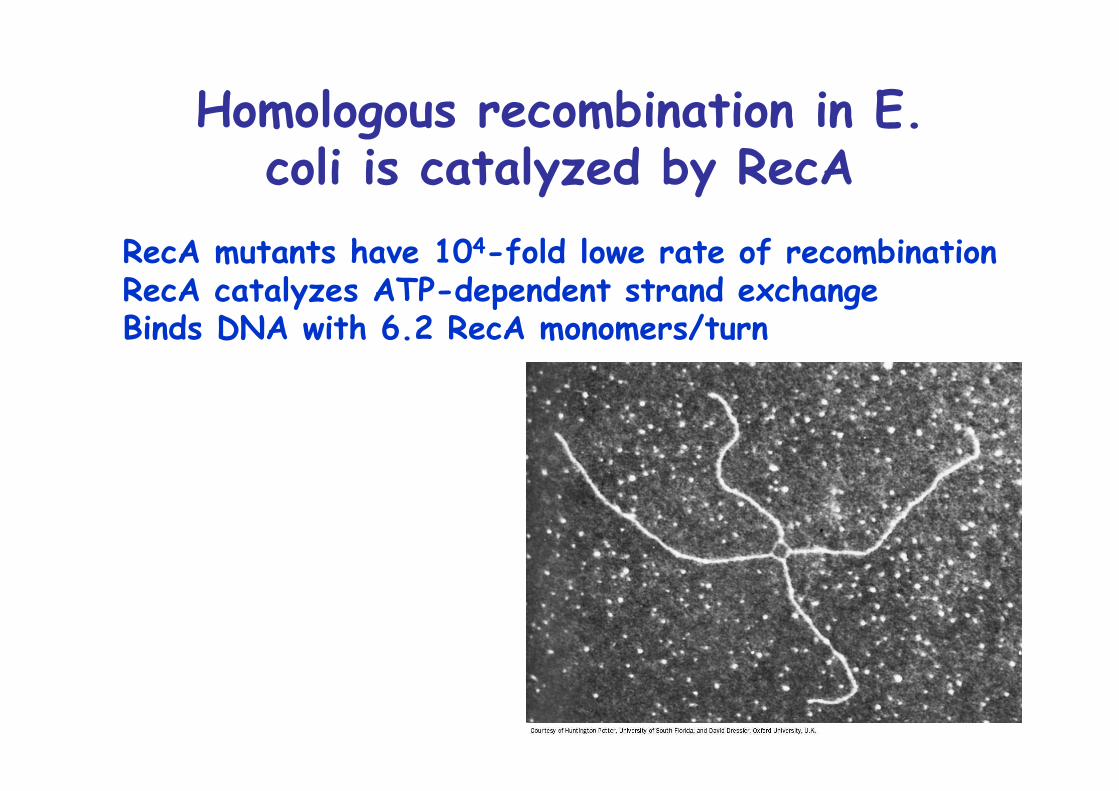
Homologous recombination in E.coli is catalyzed by RecA
RecA mutants have 104-fold lowe rate of recombinationRecA catalyzes ATP-dependent strand exchangeBinds DNA with 6.2 RecA monomers/turn
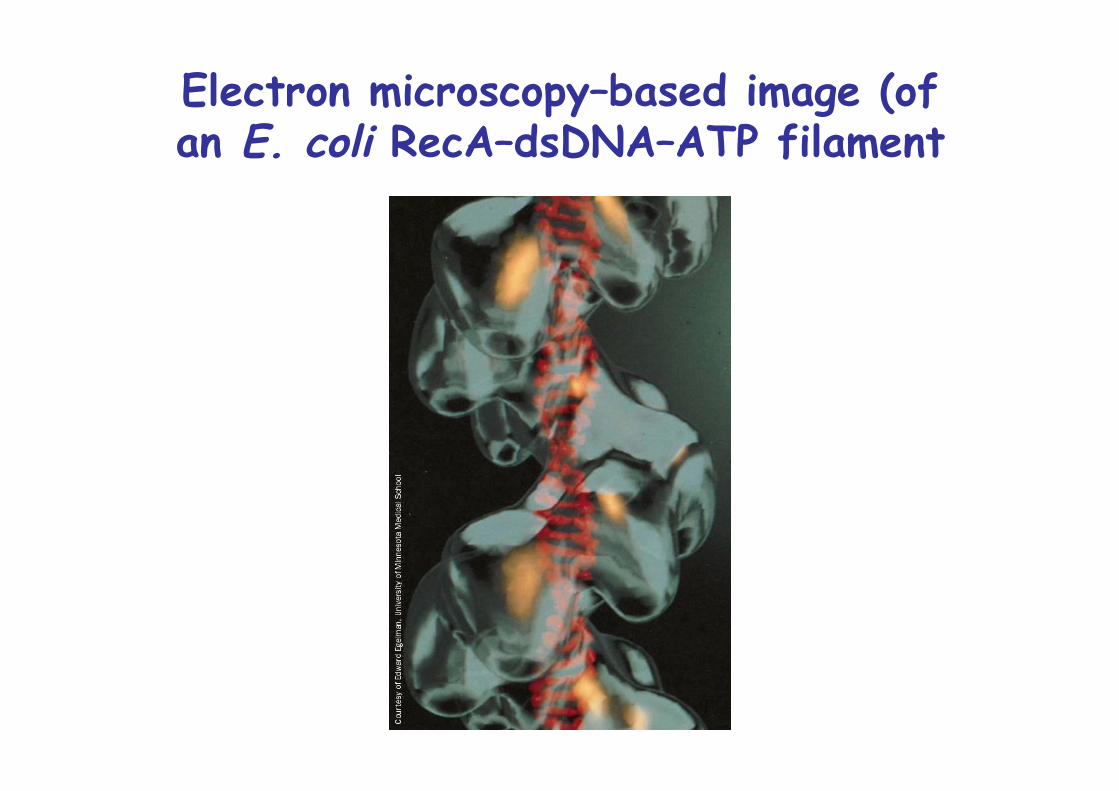
Electron microscopy–based image (ofan E. coli RecA–dsDNA–ATP filament
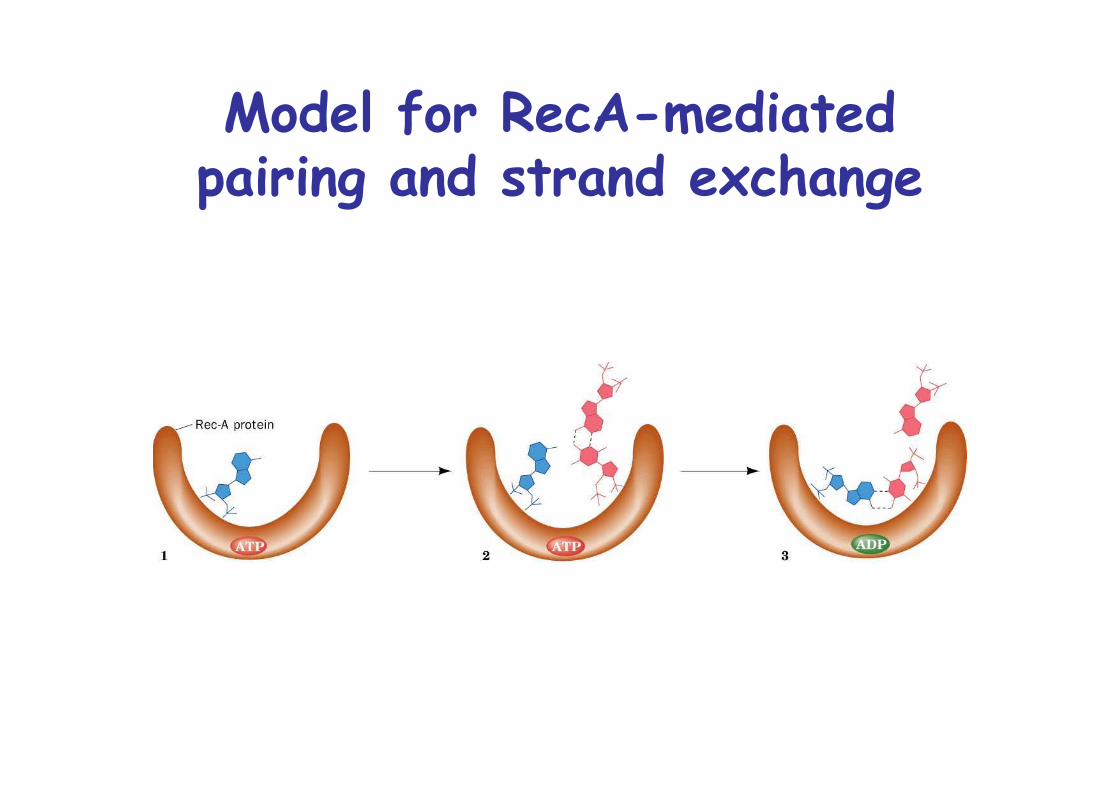
Model for RecA-mediatedpairing and strand exchange
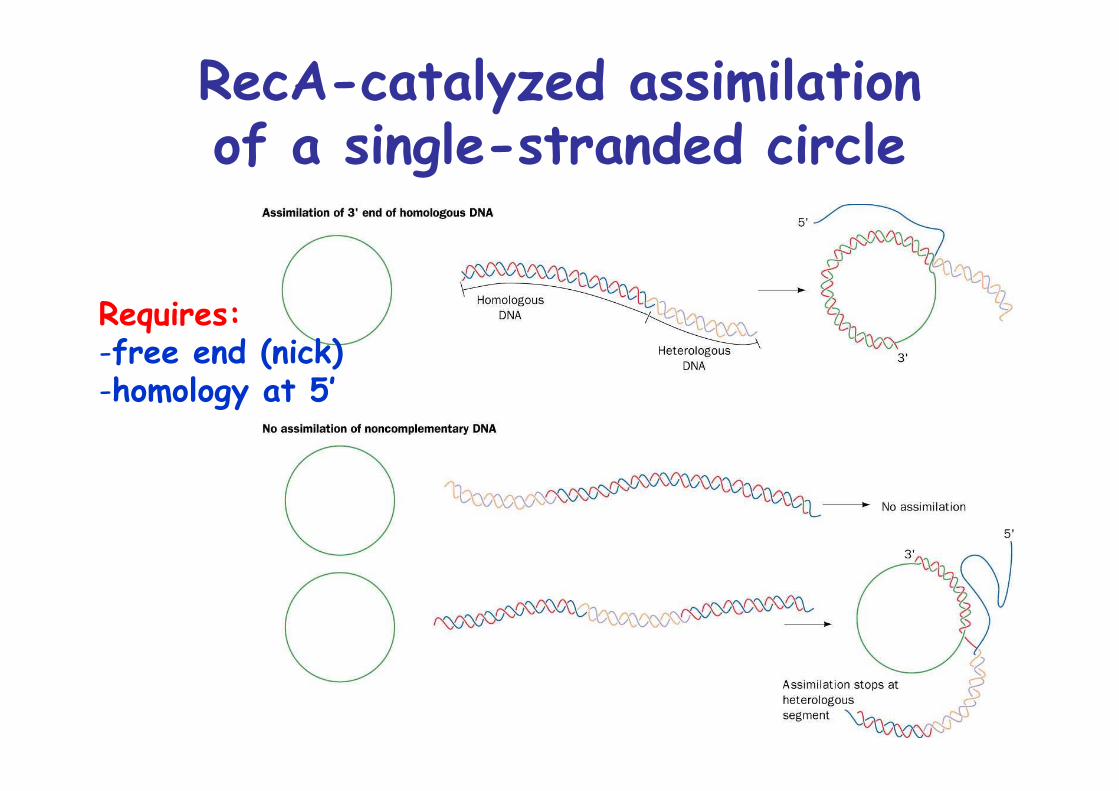
RecA-catalyzed assimilationof a single-stranded circle
Requires:-free end (nick)-homology at 5’
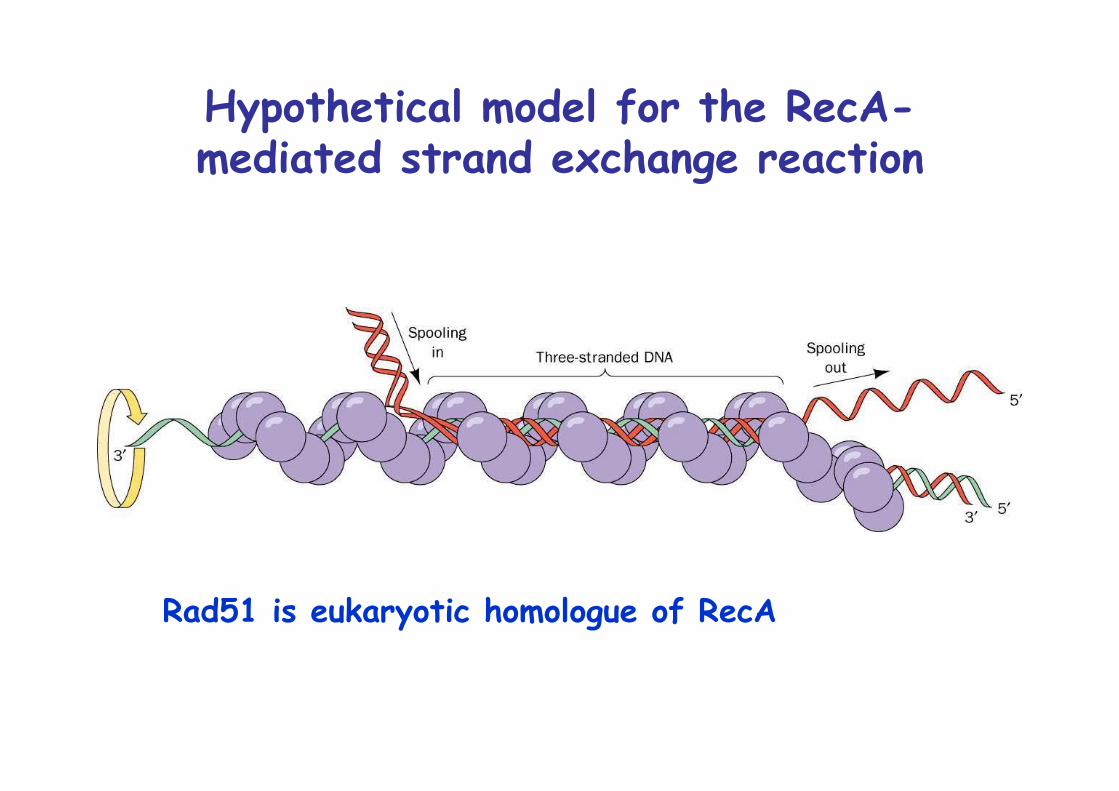
Hypothetical model for the RecA-mediated strand exchange reaction
Rad51 is eukaryotic homologue of RecA
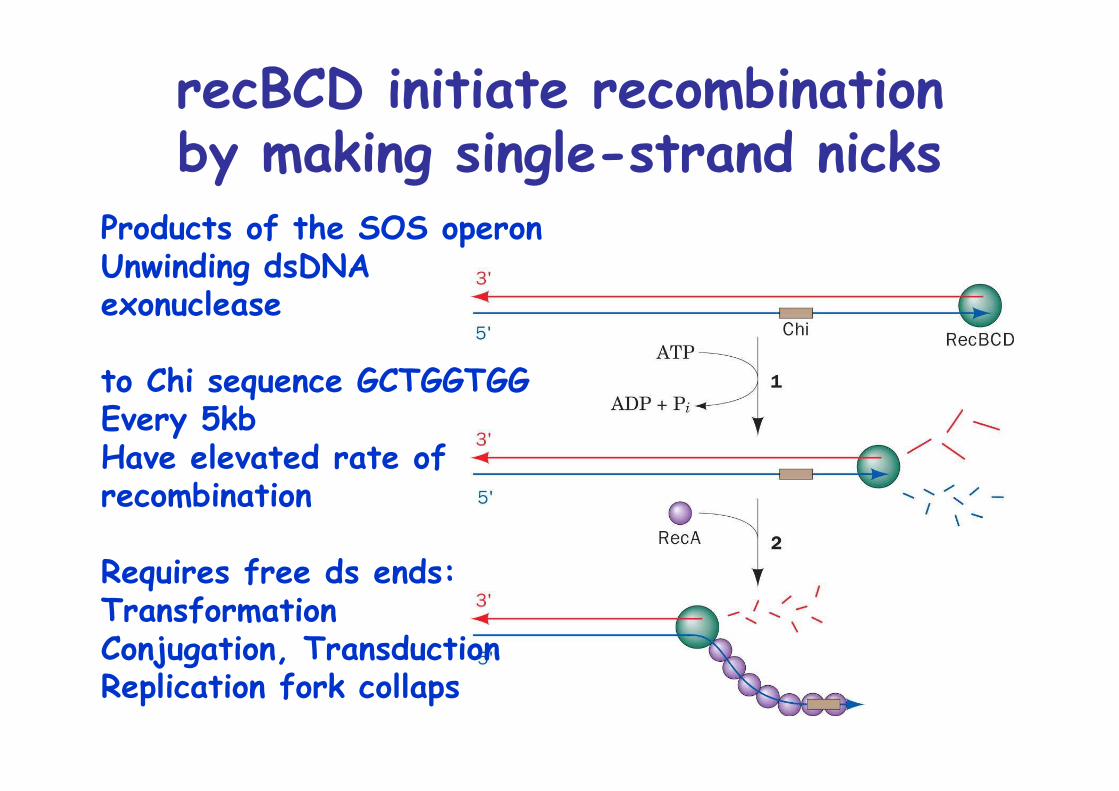
recBCD initiate recombinationby making single-strand nicks
Products of the SOS operonUnwinding dsDNAexonuclease
to Chi sequence GCTGGTGGEvery 5kbHave elevated rate of recombination
Requires free ds ends:TransformationConjugation, TransductionReplication fork collaps

RuvABC mediates branch migration andthe resolution of the Holliday junction
Branch migration is ATP-dependent, unidirectionalMediated by SOS-induced proteins: RuvB, ATP-dep. Pump, hexamer, AAA+ RuvA, binds Holliday junction, homotetramer RuvC, exonuclease
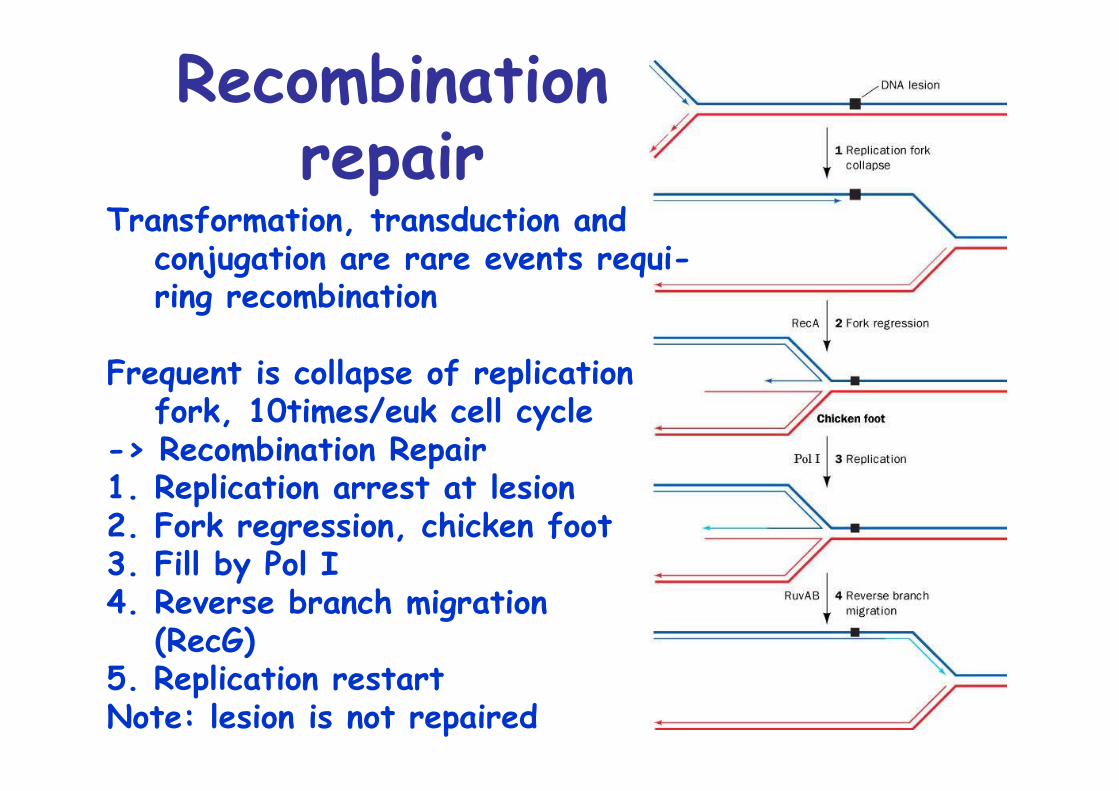
Recombinationrepair
Transformation, transduction and conjugation are rare events requi-ring recombination
Frequent is collapse of replicationfork, 10times/euk cell cycle
-> Recombination Repair1. Replication arrest at lesion2. Fork regression, chicken foot3. Fill by Pol I4. Reverse branch migration
(RecG)5. Replication restartNote: lesion is not repaired
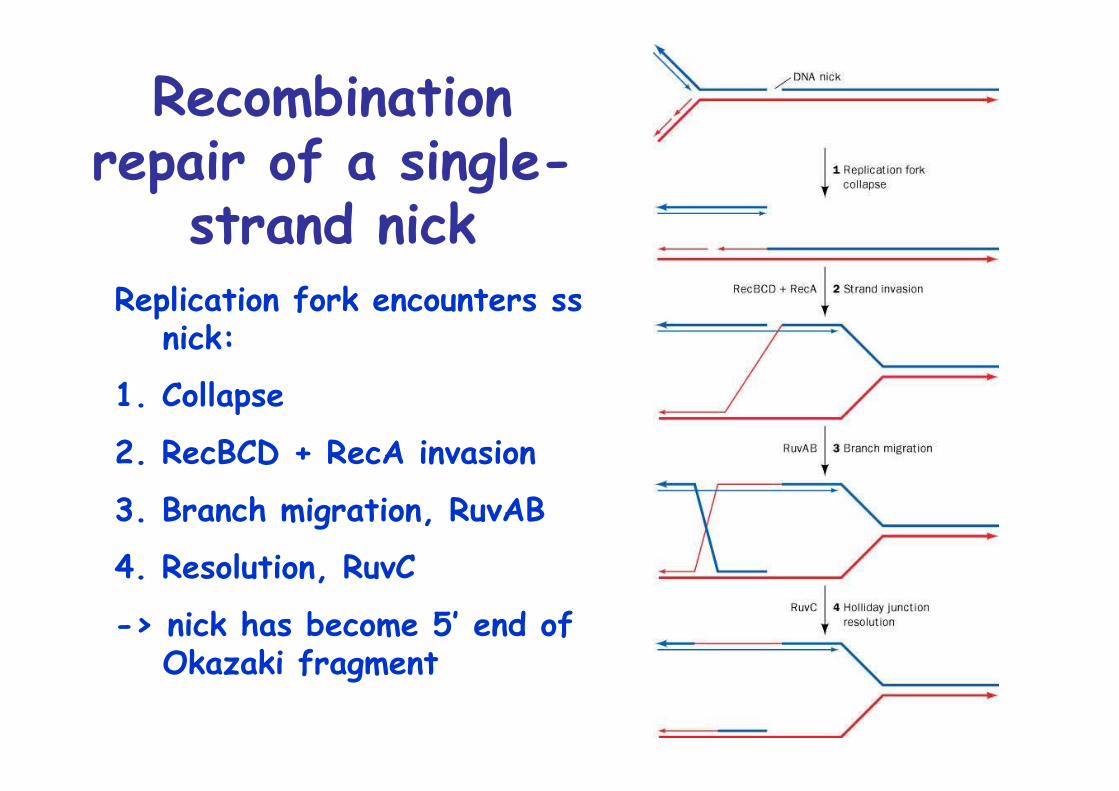
Recombinationrepair of a single-
strand nickReplication fork encounters ss
nick:
1. Collapse
2. RecBCD + RecA invasion
3. Branch migration, RuvAB
4. Resolution, RuvC
-> nick has become 5’ end ofOkazaki fragment
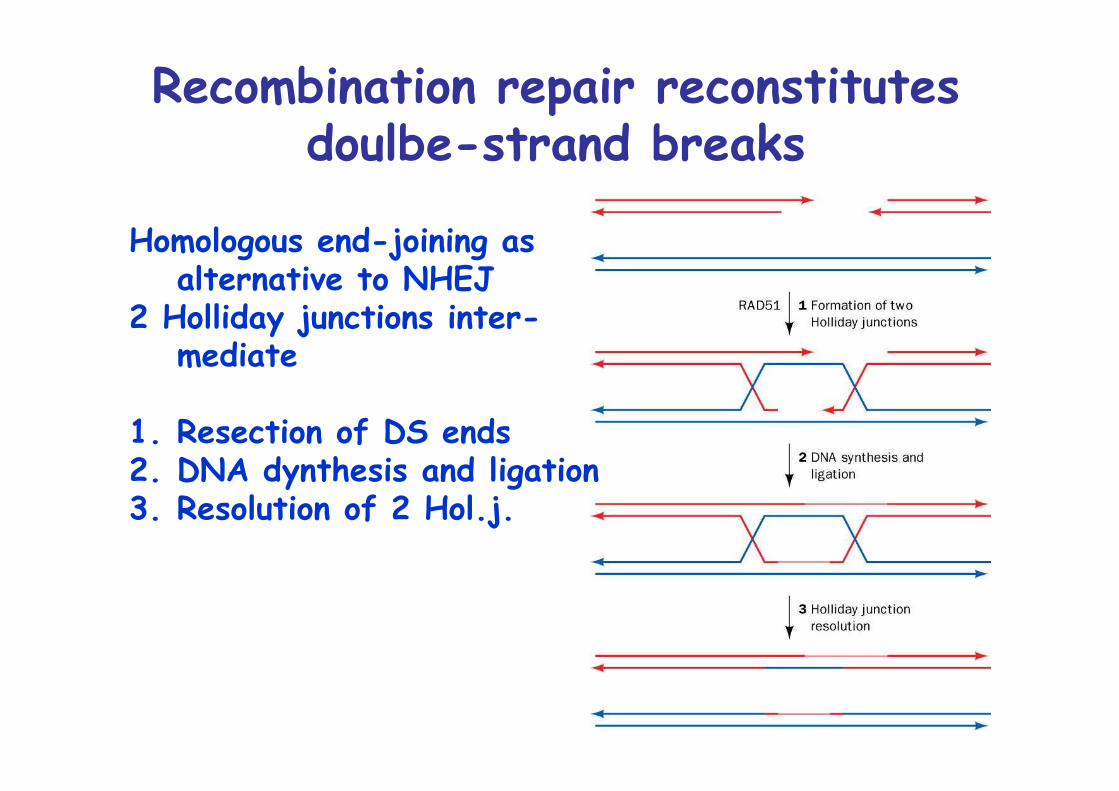
Recombination repair reconstitutesdoulbe-strand breaks
Homologous end-joining asalternative to NHEJ
2 Holliday junctions inter-mediate
1. Resection of DS ends2. DNA dynthesis and ligation3. Resolution of 2 Hol.j.

Transposition and site-specific recombination
1950 Barbara McClintock, varied pigmentation on maize
Due to the action of variable genetic elements, i.e.non-Mendelian inheritance
20 years later, evidence for mobile genetic elements inE. coli
Transposable elements, transposons in prokaryotes andeuk.
Each transposon encodes for a transposase thatcatalyzes illegitimate recombination, because it requiresno homology between donor and acceptor
Transposition is mutagenic and dangerous, tightlyregulated: 10-5 to 10-7 events per cell division

Prokaryotic transposons3 Types:
1. Simplest, insertion sequences, IS Elements<2000bp, transposase, flanked by shortinverted repeats, flanked by direct repeatat insertion site, E. coli: 8 copies of IS1,5 copies of IS2
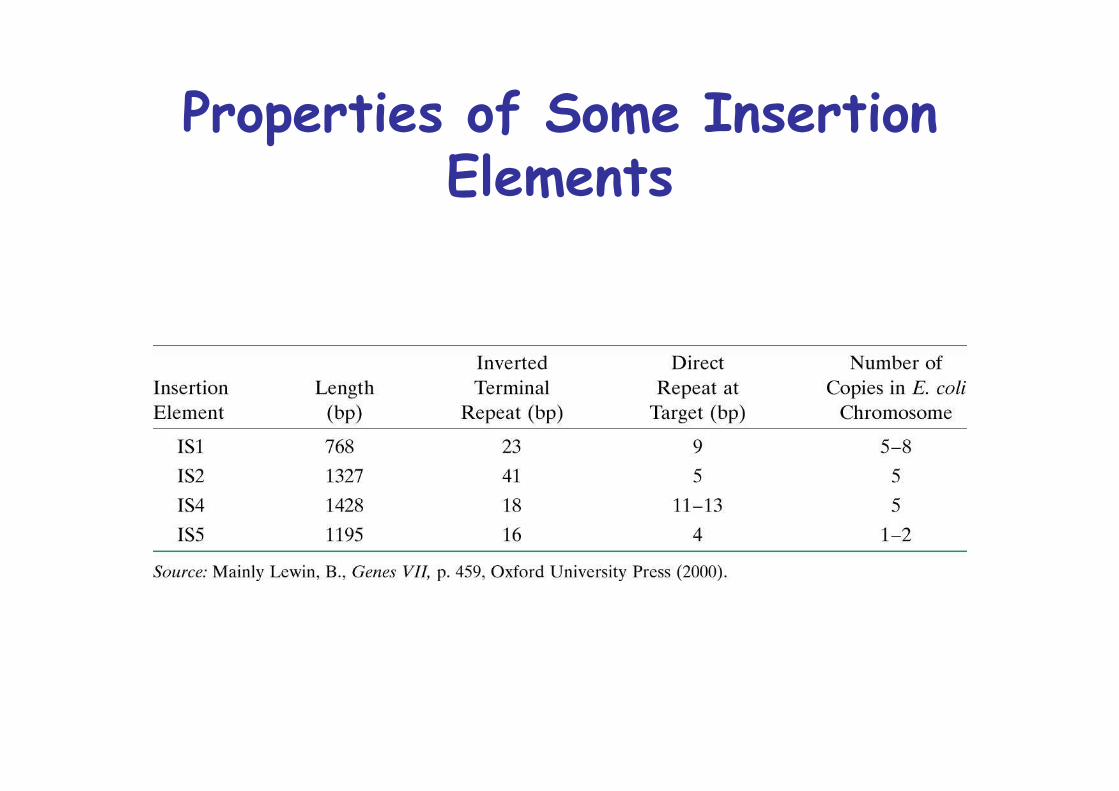
Properties of Some InsertionElements
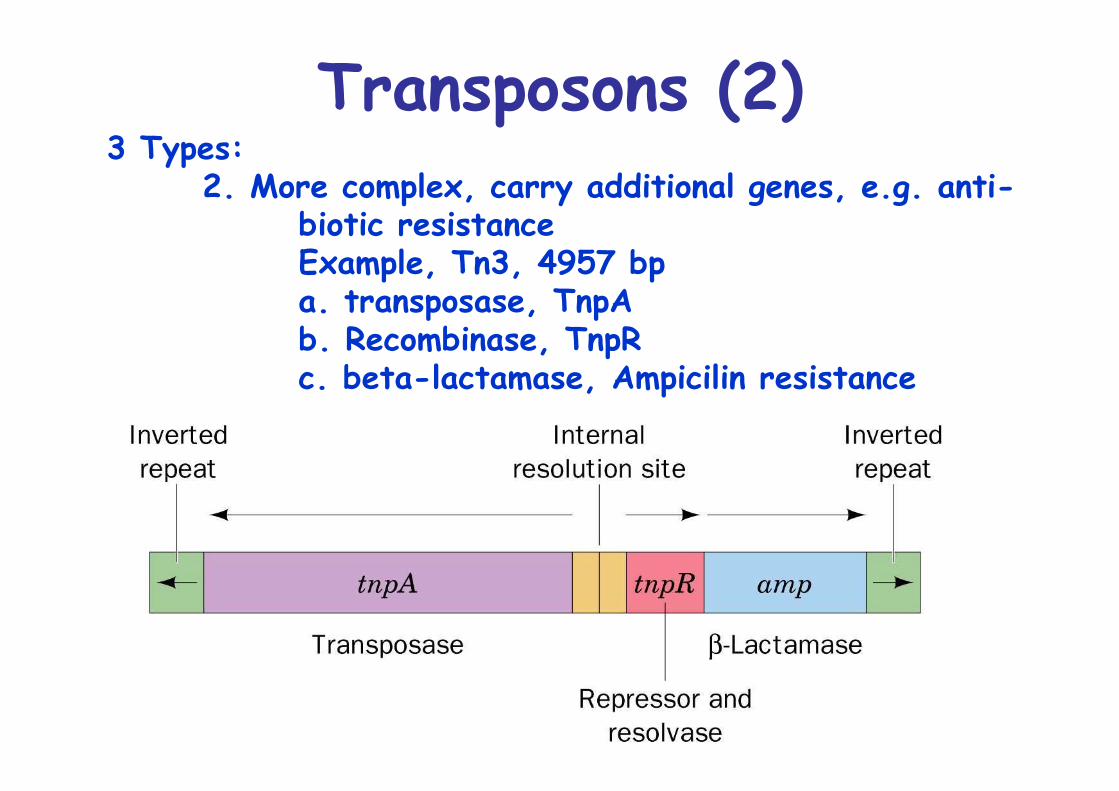
Transposons (2)3 Types:
2. More complex, carry additional genes, e.g. anti-biotic resistanceExample, Tn3, 4957 bpa. transposase, TnpAb. Recombinase, TnpRc. beta-lactamase, Ampicilin resistance
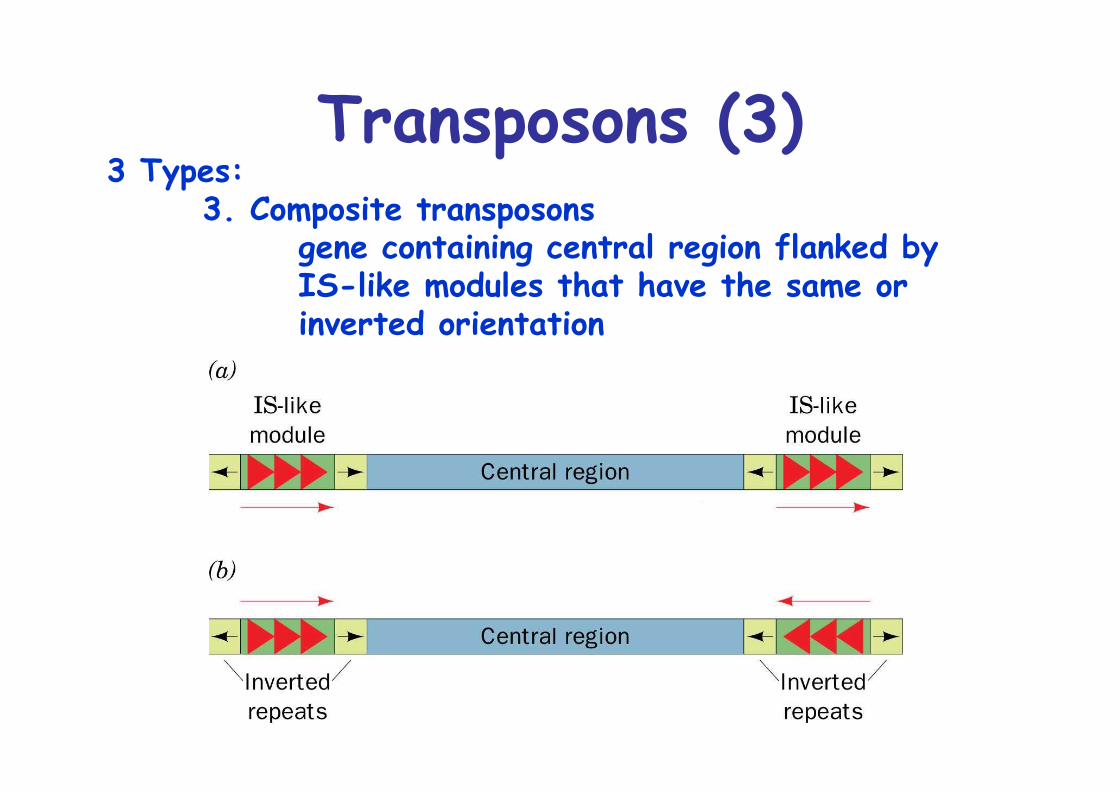
Transposons (3)3 Types:
3. Composite transposonsgene containing central region flanked byIS-like modules that have the same orinverted orientation
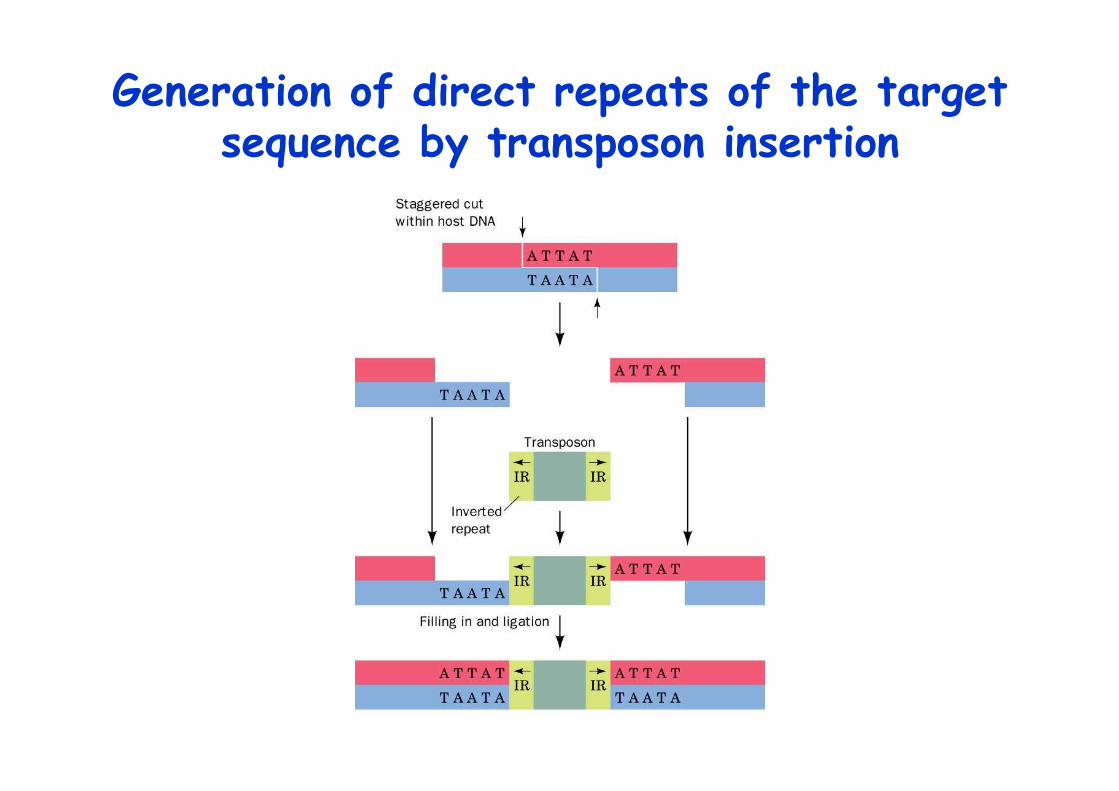
Generation of direct repeats of the targetsequence by transposon insertion

Two modes for transposition
1. Direct or simple transposition -> transposon moves fromposition A to position B
2. Replicative transposition -> transposon remains + newcopy at position B
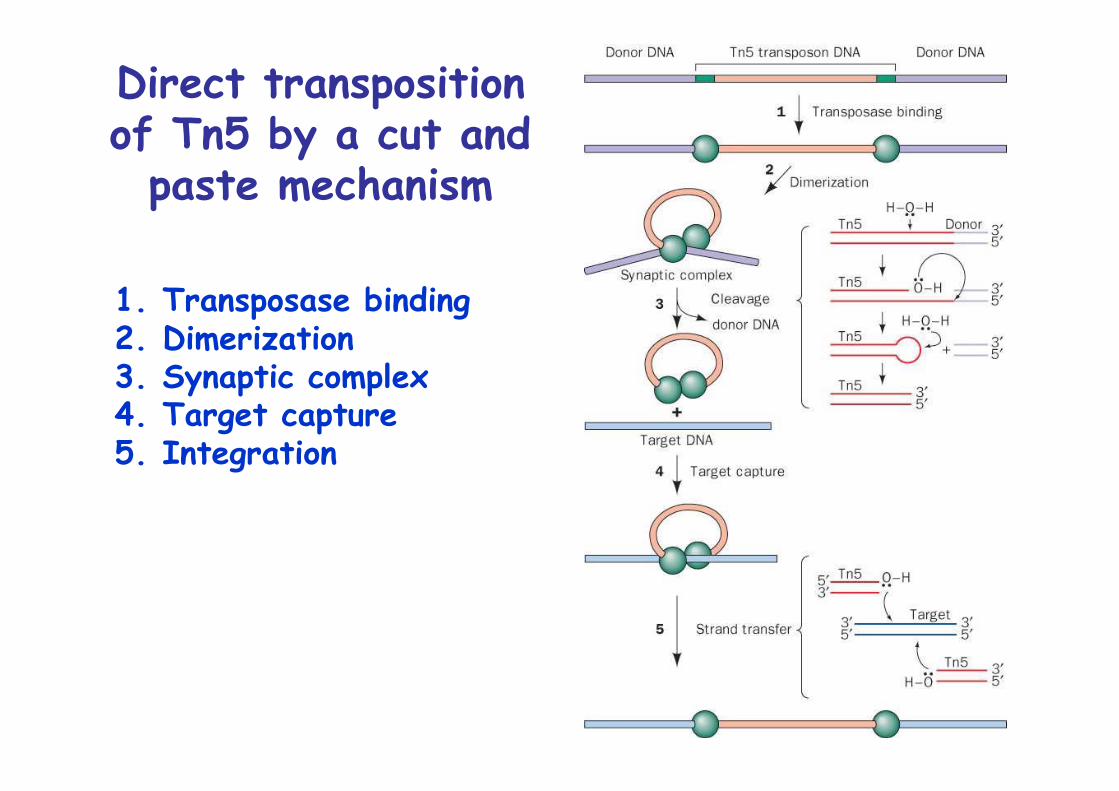
Direct transpositionof Tn5 by a cut and
paste mechanism
1. Transposase binding2. Dimerization3. Synaptic complex4. Target capture5. Integration
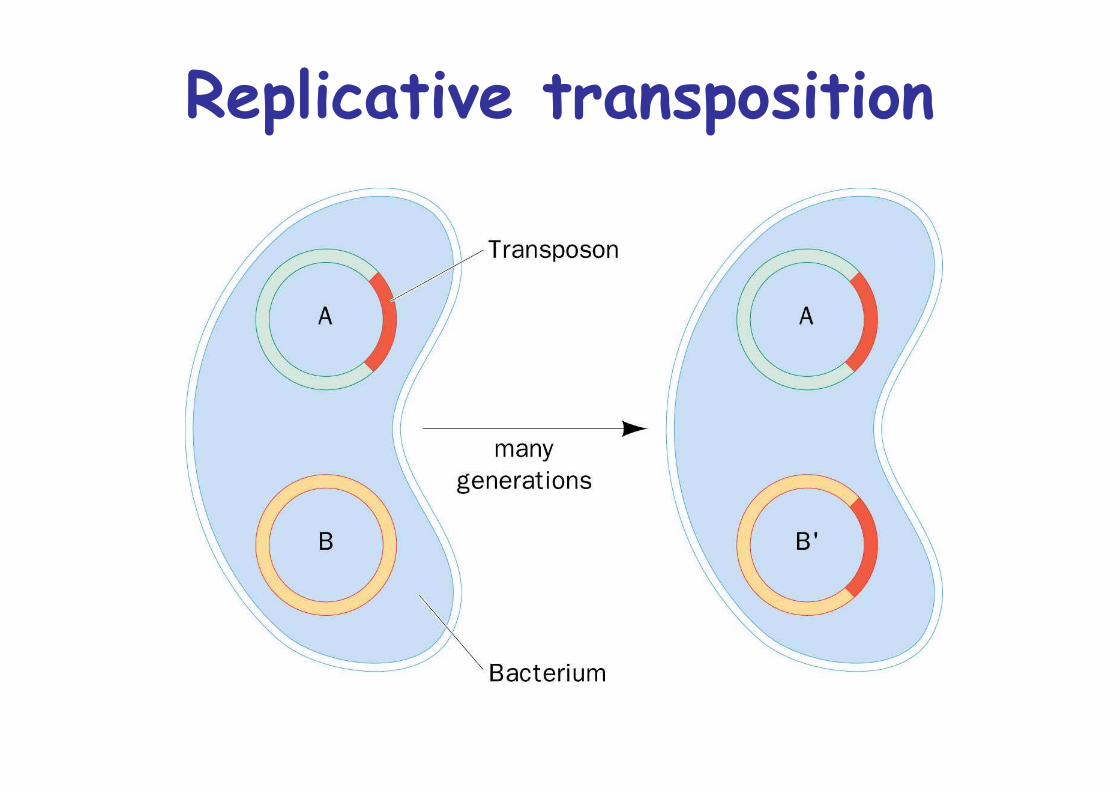
Replicative transposition
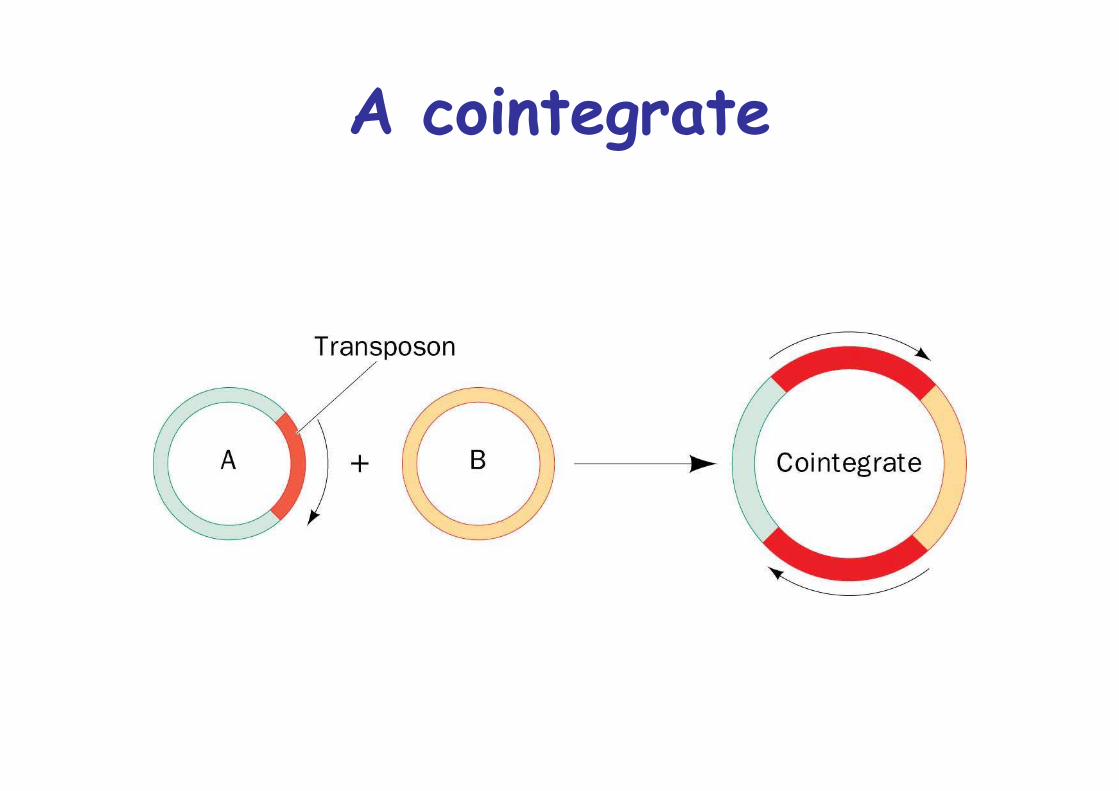
A cointegrate
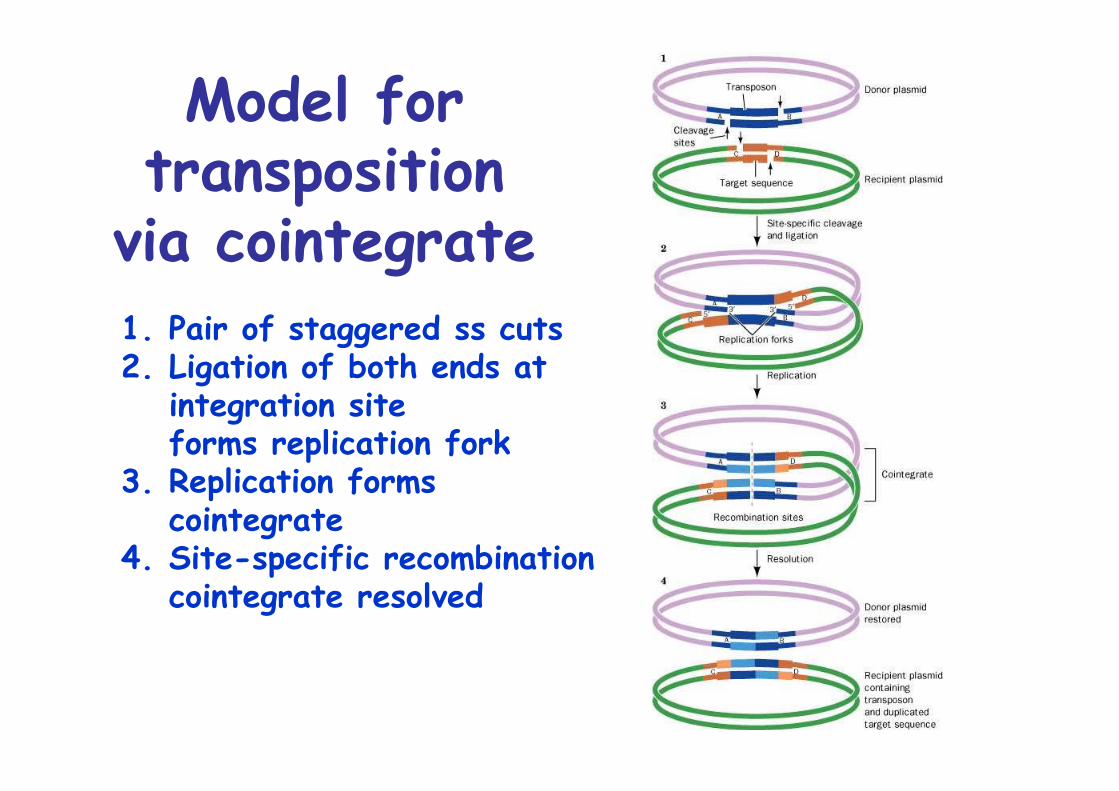
Model fortransposition
via cointegrate1. Pair of staggered ss cuts2. Ligation of both ends at
integration siteforms replication fork
3. Replication formscointegrate
4. Site-specific recombinationcointegrate resolved
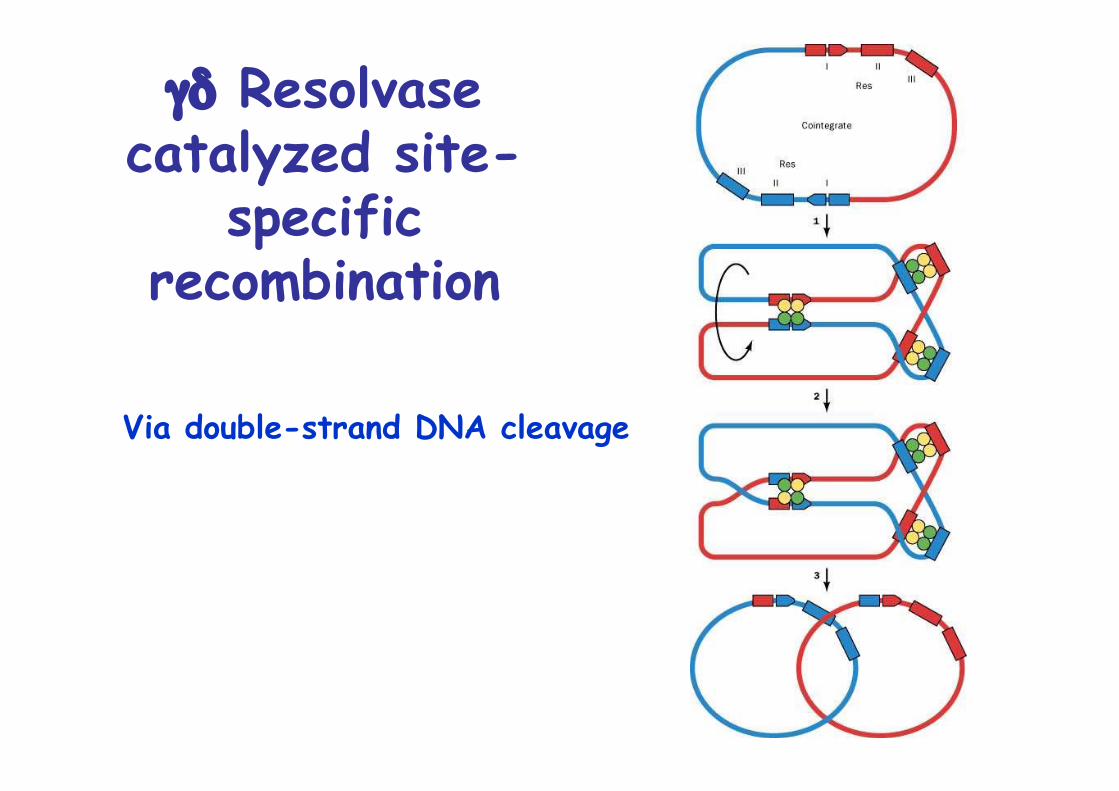
γδ Resolvasecatalyzed site-
specificrecombination
Via double-strand DNA cleavage
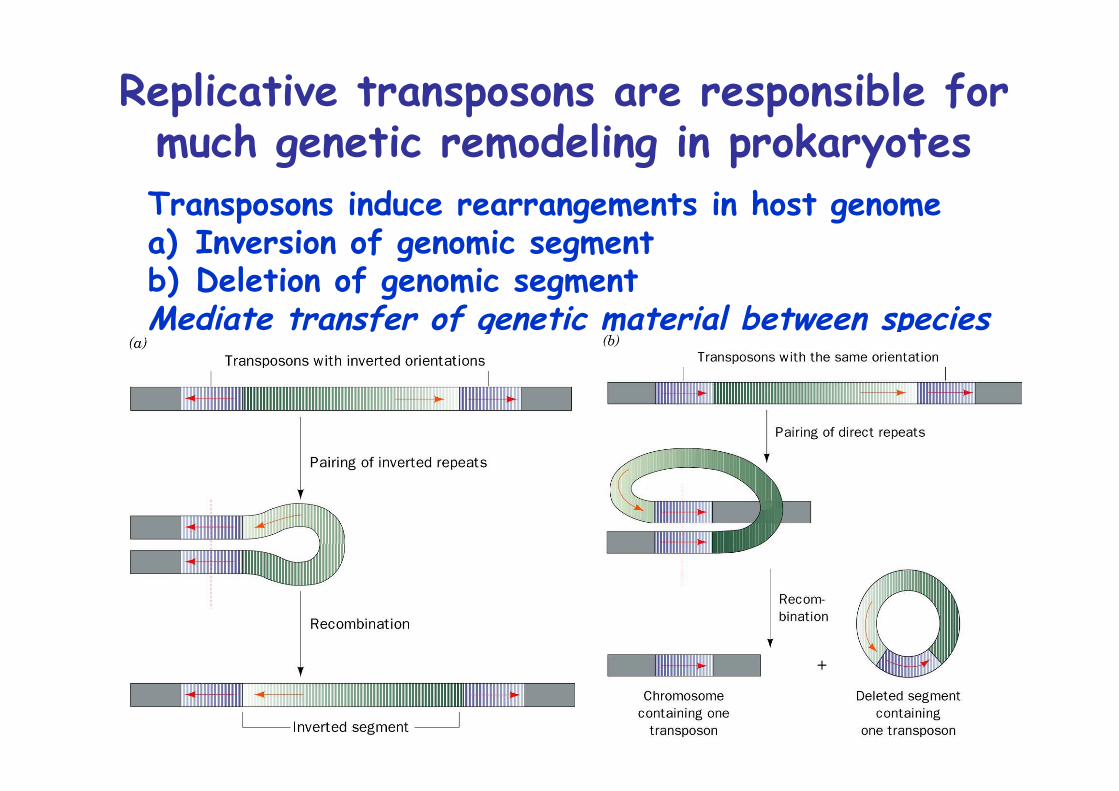
Replicative transposons are responsible formuch genetic remodeling in prokaryotesTransposons induce rearrangements in host genomea) Inversion of genomic segmentb) Deletion of genomic segmentMediate transfer of genetic material between species

Phase variation is mediated bysite-specific Recombination
Salmonella typhimurium make 2 antigenetically distinctversions of flagellin, H1 and H2
only one of the two is expressedswitch every 1000 cell generations, phase variationmay help evade host immune response
H2 is linked to rh1, that encodes a repressor for H1Expression of H2-rh1 unit is controlled by a 995bp segment that contains
1. Promoter for H2-rh12. Hin gene coding for Hin DNA invertase3. Two closely related 26bp sites, hixL and hixR
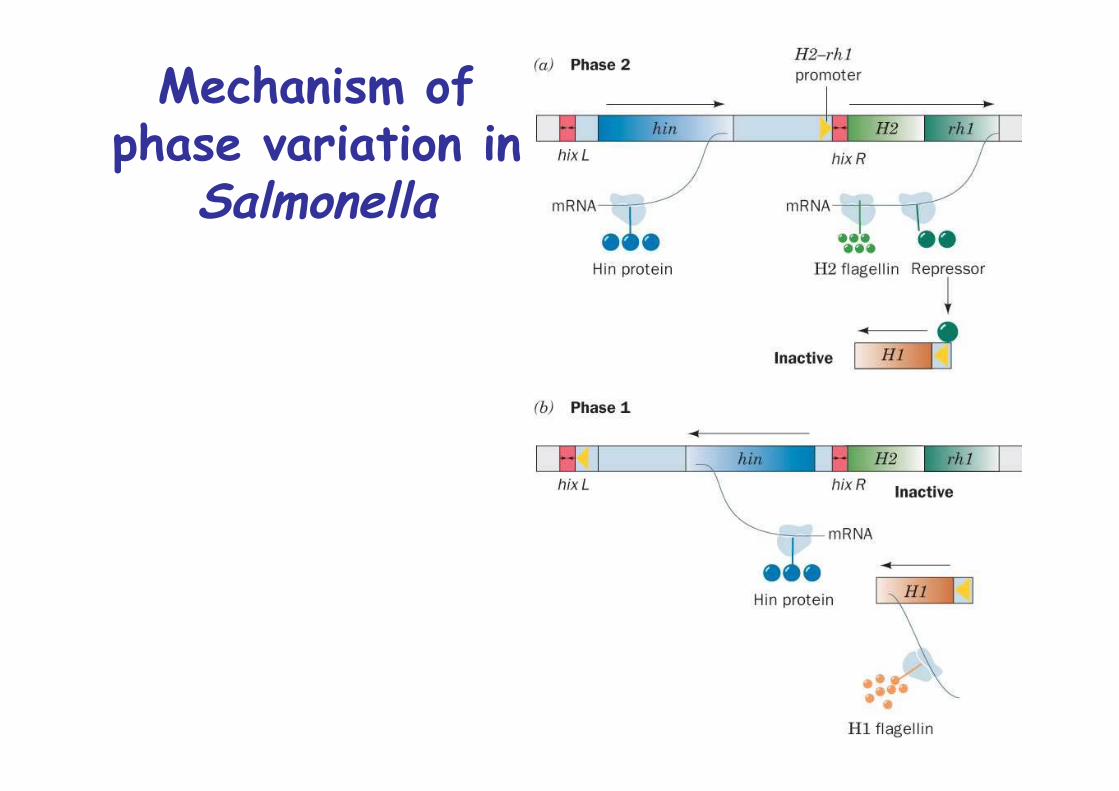
Mechanism ofphase variation in
Salmonella

Cre-mediated site-specificrecombination
Many bacteriophages have two modes to propagate: 1.lytic, lysis of cells 2. Lysogeic, integration into host genome
Examples: Bacteriophage lambda, λ integraseP1 bacteriophage, Cre recombinase
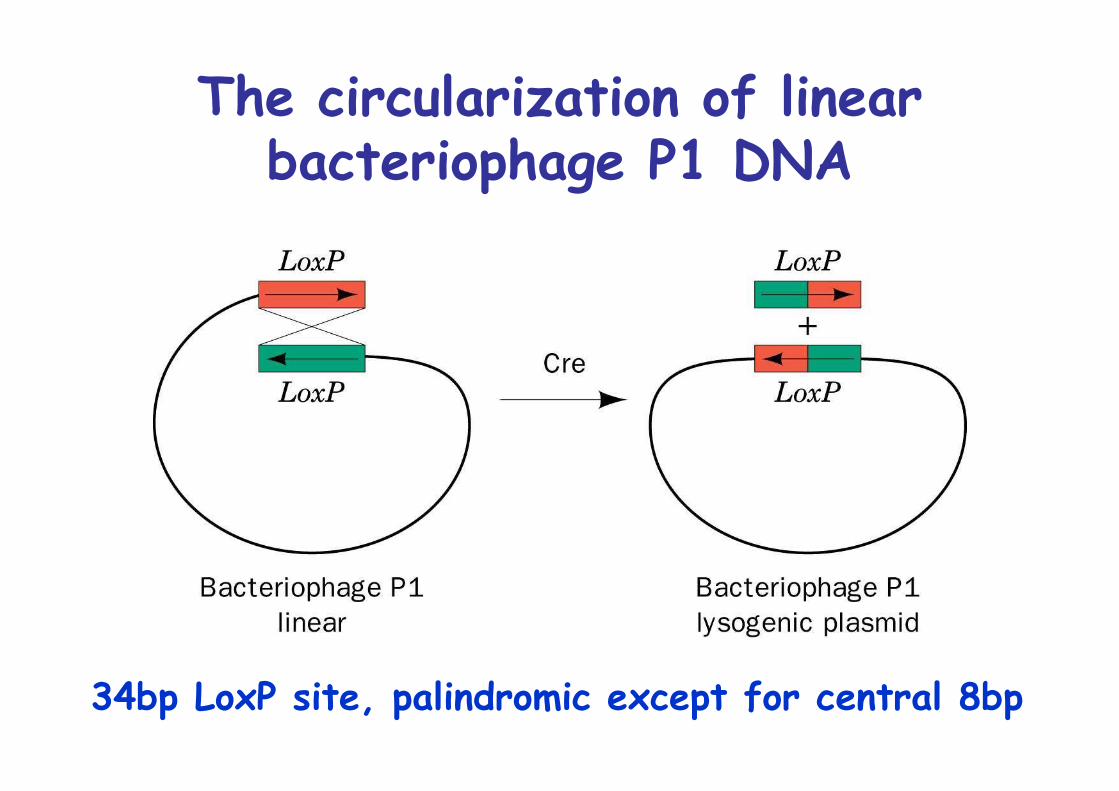
The circularization of linearbacteriophage P1 DNA
34bp LoxP site, palindromic except for central 8bp
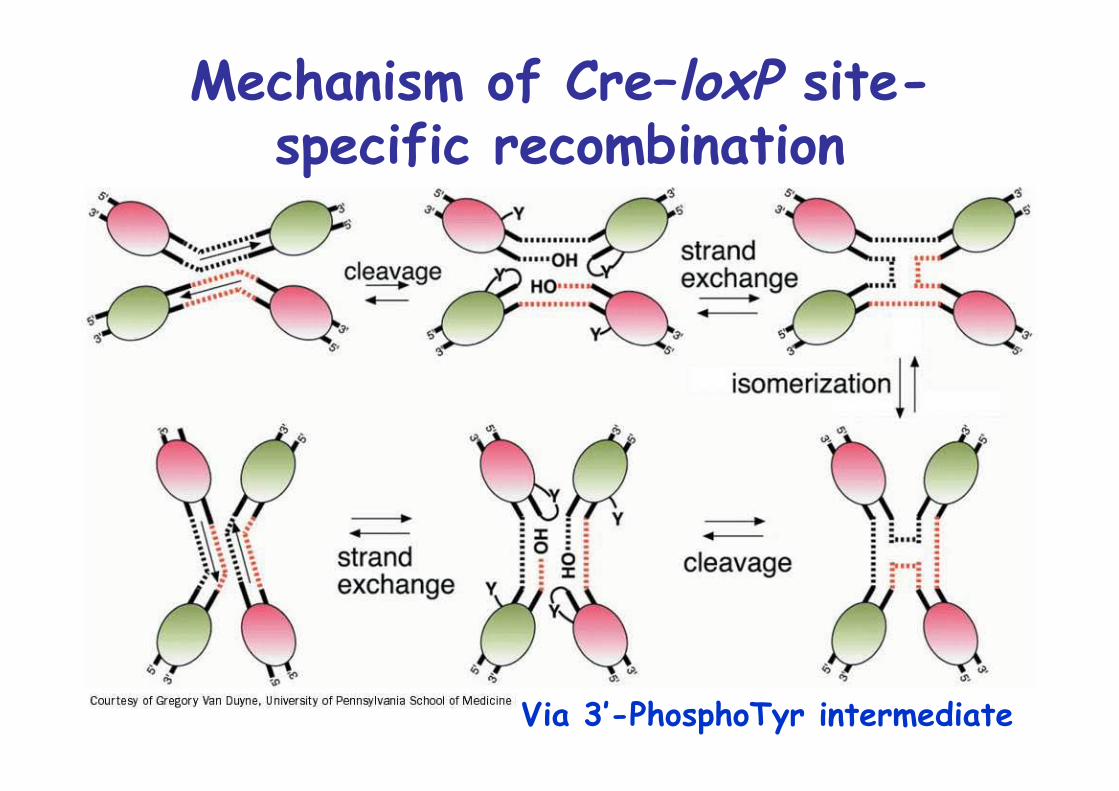
Mechanism of Cre–loxP site-specific recombination
Via 3’-PhosphoTyr intermediate
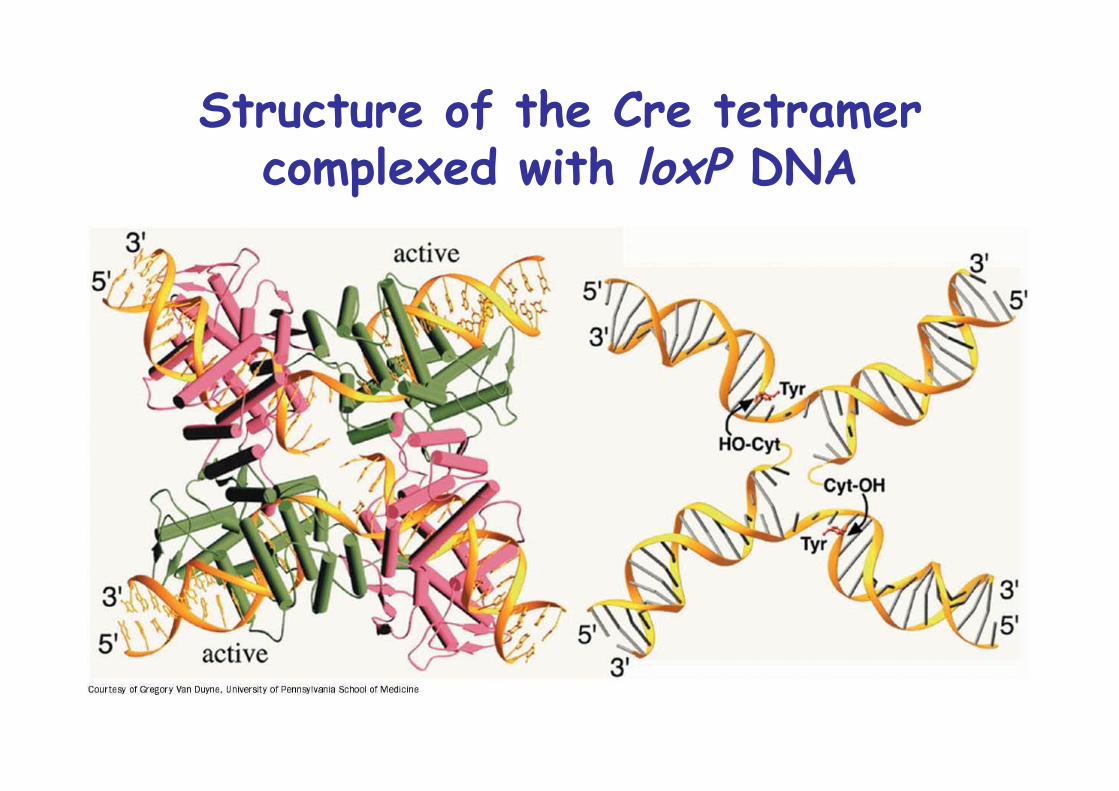
Structure of the Cre tetramercomplexed with loxP DNA

Most transposition in eukaryotesinvolve RNA intermediates
3% of the human genome consists of transposonsMany are fosils, i.e. sequence mutated to be inactiveMany ressemble retroviruses in sequence
RetroposonsTransposition via RNA intermediate, tanscriptiondsDNA via reverse transcriptase, cDNARandom integration by integrase
Retroviral genome flanked by LTR, long terminal repeats(250-600bp)3 polyproteins: gag (viral core)
pol (reverse transcriptase)env (viral envelope)

Organization of retrovirusesand the Ty1 retrotransposon

Non-viralretroposons
Vertebrate genomes containRetroposons that lack LTRsNon-viral retroposons,e.g.LINEs, long intersoersed nuclearelements, 1-7kn longContain 2 ORFsORF1, similar to gagORF2, similar to pol
In humans, LINEs account for20% of genome !
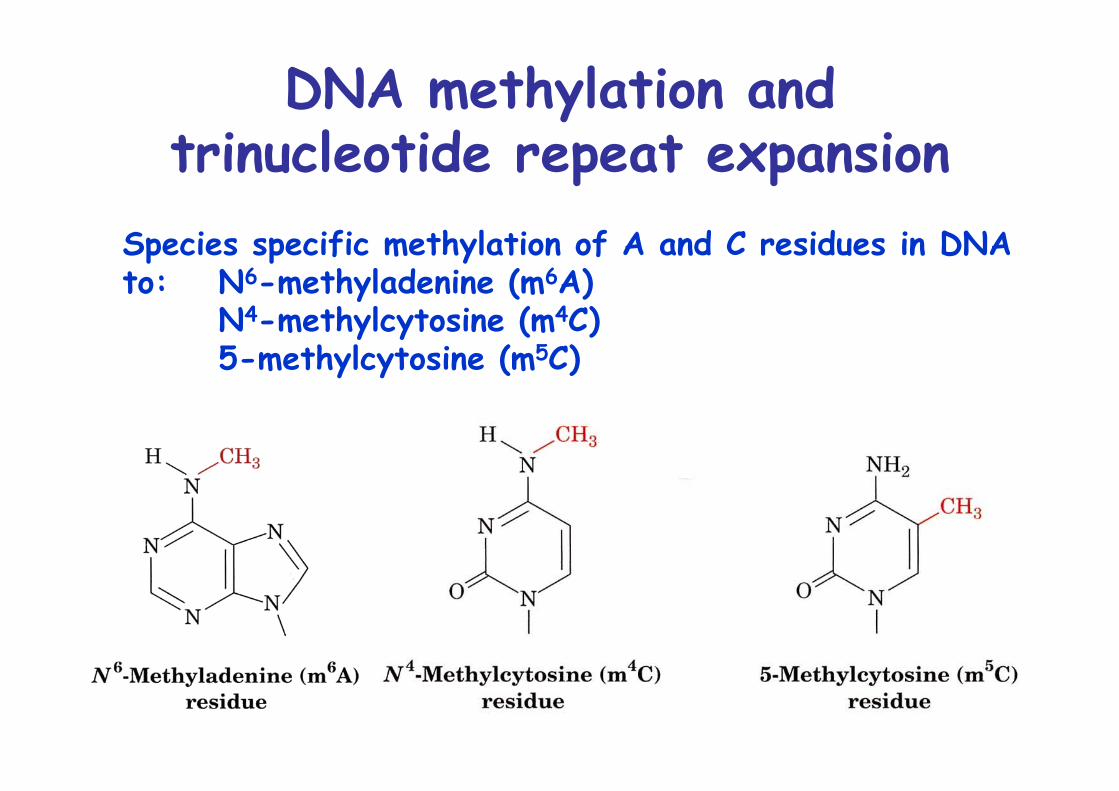
DNA methylation andtrinucleotide repeat expansion
Species specific methylation of A and C residues in DNAto: N6-methyladenine (m6A)
N4-methylcytosine (m4C)5-methylcytosine (m5C)

DNA methylation
Bacterial DNA is methylated at own restriction site
E.coli, Dam methyltransferase (dam MTase), A in GATC Dcm MTase bith C in CCA/TGG at pos 5 both palindromic, mismatch repair and oriC
Methyl groups project into major groove of B-DNA, interact with DNA-binding proteins
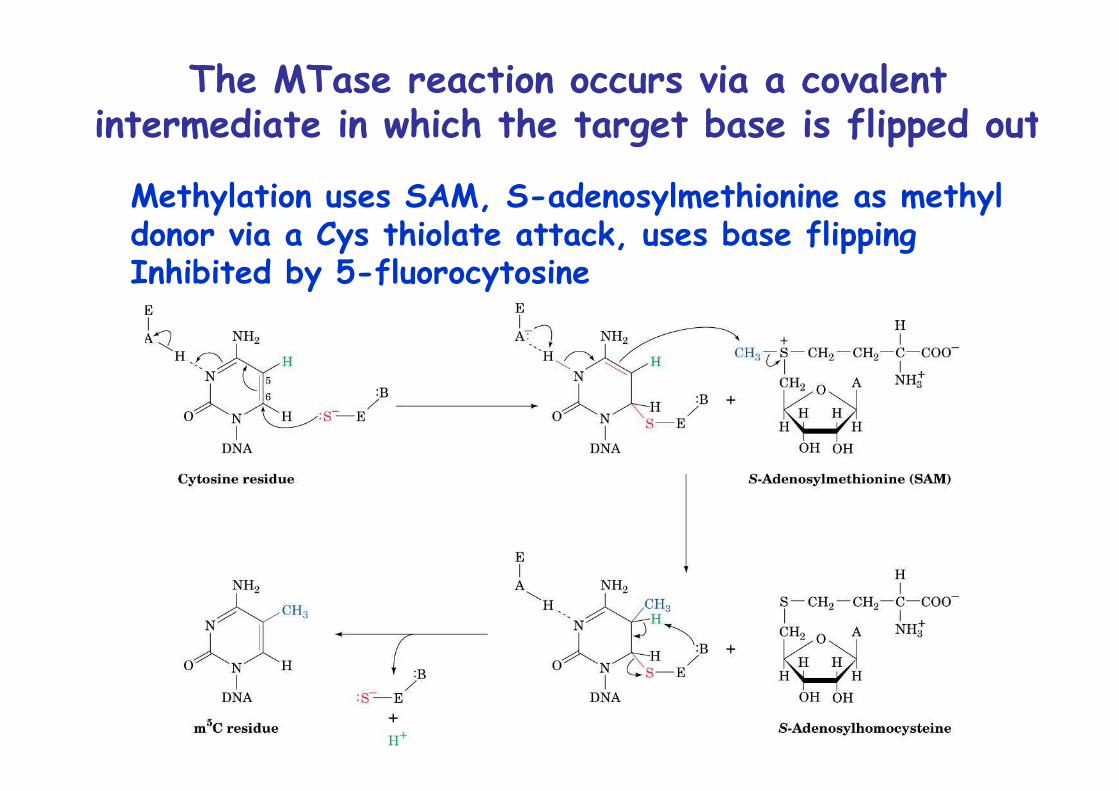
The MTase reaction occurs via a covalentintermediate in which the target base is flipped out
Methylation uses SAM, S-adenosylmethionine as methyl donor via a Cys thiolate attack, uses base flippingInhibited by 5-fluorocytosine
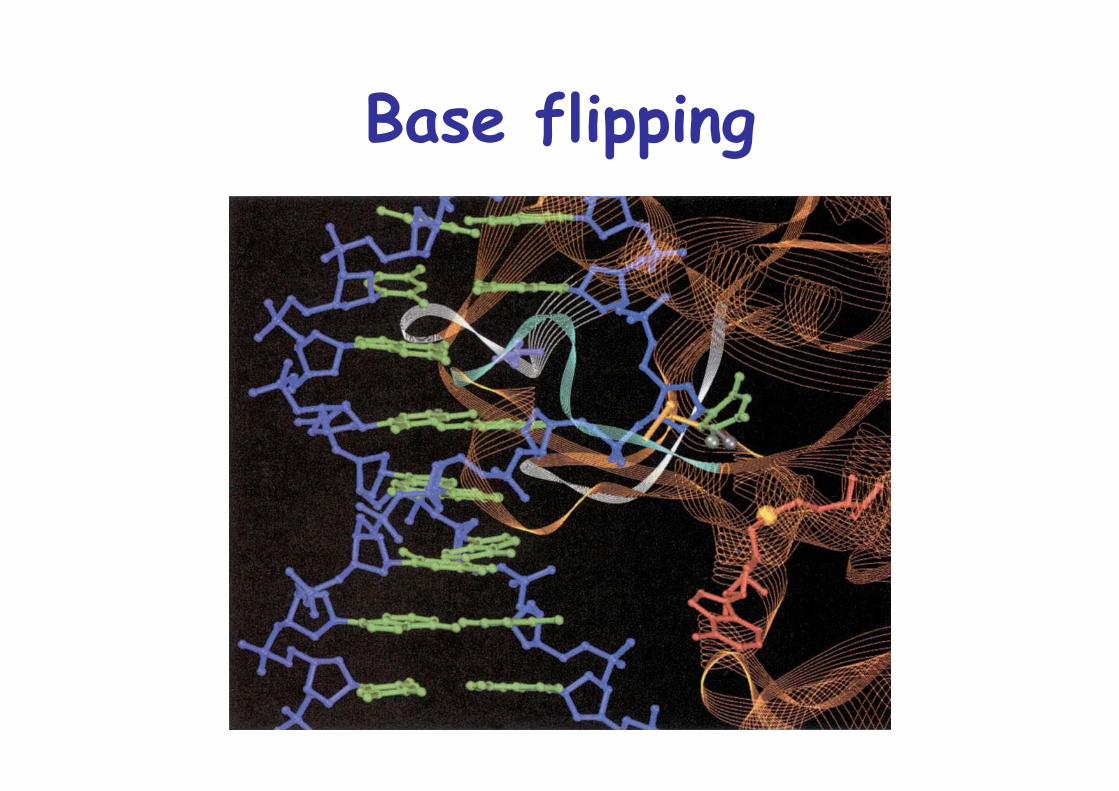
Base flipping

DNA methylation in eukaryotesfunctions in gene regulation
5-methylcytosine is the only methylated base in mosteukaryotesModification in largely in GC dinucleotideCG is present at 1/5 of statistical expectationUpstream regions of many genes have CpG island

DNA methylation in eukaryotes
Experimental assessment:Comparative southern blot of DANN cut withHpaII, cleaves CCGG, but not C-m5C-GG andMspI, cleaves both
Identification of m5C residues through bisulfitesequencingDNA is reacted with bisulfite (HSO3
-) whichdeaminates C to U, but not m5C, followed by PCR amplification:copies U to T and m5C to CSequence and compare to untreated

DNA methylation in eukaryotes (2)Methylation switches off eukaryotic gene expression,particularly when methylation occurs in promoter regionFor example, globin genes are less methylated inerythroid cells
Recognized by methyl-CpG binding domain (MBD)May also affect chromatin packaging
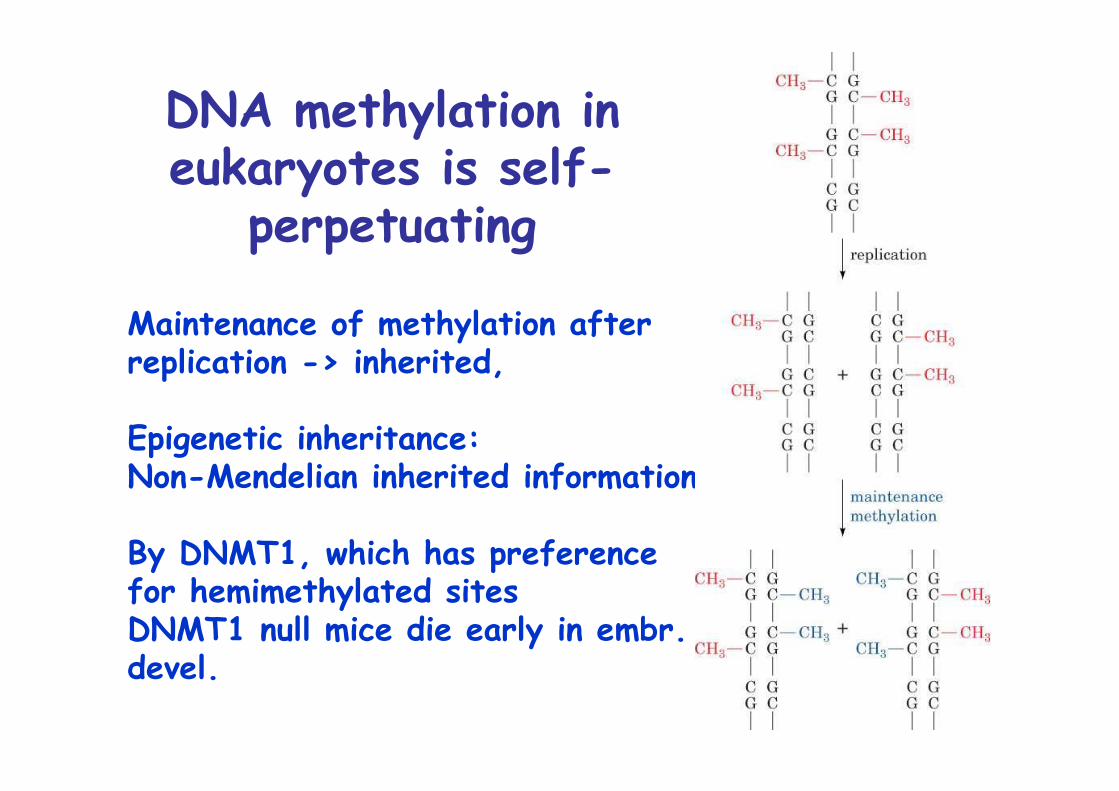
DNA methylation ineukaryotes is self-
perpetuating
Maintenance of methylation afterreplication -> inherited,
Epigenetic inheritance:Non-Mendelian inherited information
By DNMT1, which has preference for hemimethylated sitesDNMT1 null mice die early in embr.devel.

Methylation is dynamicPattern of DNA methylation varies in early embryologicaldevelopment:
Methylation levels high in gamets (sperm, ova) butnearly eliminated in blastocyst stageMethylation then rises again till gastrula stagewhen it reaches that found in adults, remain constantExcept germ line cells, remain unmethylated
Pattern of expression differs in embryonic and somaticcells=> Explains high failure of cloning experiments, few survivers, early death, abnormalities, large size

Genomic imprinting results fromdifferential DNA methylation
Difference in maternal and paternal inheritance:Mare x Male donkey -> muleFemale donkey x stallion -> hinnyBoth are sterile
Maternal and paternal genes are differentially expressed= genomic imprinting, only in mammalsNo embry from transplant of two male or female pronuclei
mule hinny

DNA methylation is associatedwith cancer
Most prevalent mutation is is m5C to T, covert proto-oncogens to oncogens or inactivate tumor suppressors

Several neurological diseases areassociated with trinucleotide repeat
expansionFragile X syndrome: mental retardation, long narrow face1 in 4500 males, 1 in 9000 femalesActivated by passage through femaleAffects FMR1 gene, which contains (CGG)n, n=6-60 in5’ region, n can increase from 60 to 200 = premutationCan the expand upon transmission to a daughter to >200= full mutation
Expansion arises through slippage during replication
FMR1 is unmethylated in normal individualsBut is methylated when premutation is maternally transmitted

Other important trinucleotiderepeat diseases
Huntington’s disease (HD), 1 in 10’000, onset at age of approx. 40, 18-year course, fatalProtein huntingtin contains (CAG)n repeats (Gln)Normal 11-34, sick 37-86Repeat length is unstable, changes in >80% meiotic transmissionsNumber of repeats inversely correlates with age of onsetpolyGln aggregates as β sheetsNeurons contain inclusions
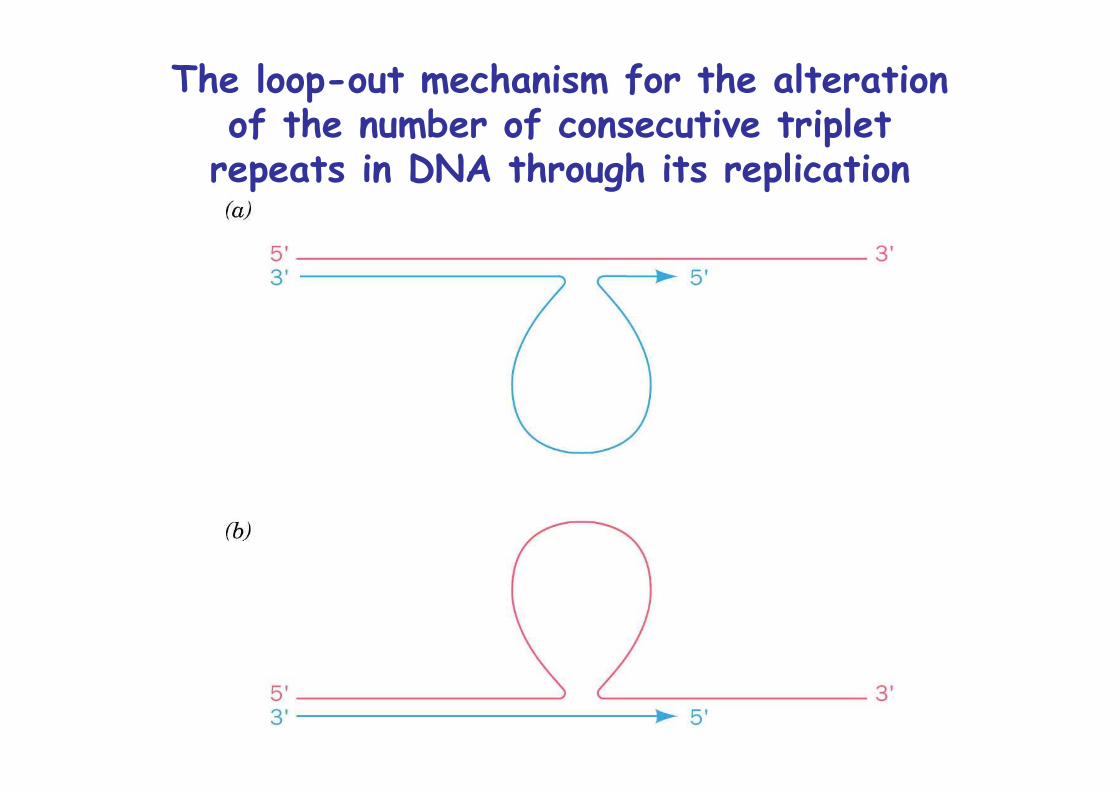
The loop-out mechanism for the alterationof the number of consecutive triplet
repeats in DNA through its replication
























