Embed Size (px)
Citation preview

Threat-Sensitive Nest Defense in Domino Damselfish, Dascyllus albisellaAuthor(s): Steven J. Oliver and Elise WatsonSource: Biological Bulletin, Vol. 197, No. 2, Centennial Issue: October, 1899-1999 (Oct., 1999),pp. 244-246Published by: Marine Biological LaboratoryStable URL: http://www.jstor.org/stable/1542628 .
Accessed: 28/06/2014 09:43
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Marine Biological Laboratory is collaborating with JSTOR to digitize, preserve and extend access toBiological Bulletin.
http://www.jstor.org
This content downloaded from 185.31.195.48 on Sat, 28 Jun 2014 09:43:53 AMAll use subject to JSTOR Terms and Conditions

REPORTS FROM THE MBL GENERAL SCIENTIFIC MEETINGS REPORTS FROM THE MBL GENERAL SCIENTIFIC MEETINGS
pattern found in other pomacentrids. The CAS were highly variable in pulse number and length, and the linear relationship between the two factors was very weak (r2 = 0.23; Fig. la). Furthermore, the CAS of A. sordidus were relatively low amplitude and could not be detected unless the hydrophone was within about 2 m. In contrast, D. albisella sounds were detected at distances up to about 9 m (8).
Male A. sordidus establish and maintain territories for nesting, often returning to the same nest site every year (9). Males compete aggressively for their own exclusive nest site. These nests are
arranged in a colonial arena layout. Courting males change col- oration from the drab barred pattern shown by non-courting males and by females to a bright contrasting pattern of white bars on a black background (Fig. lb, c). A male responds to a female in the
vicinity of the nest by rapidly swimming out toward her in a zigzag pattern, swimming first on one side, then flipping to the other. Often more than one male is seen swimming out to meet a female. The successful male leads the female back to his nest; when near the nest, he swims rapidly around her in a figure-eight pattern. During this courtship behavior, sounds are produced (Fig. ld, e). Clearly, the female will be aware of the male's activity in front of her, with or without sounds. The male continues this behavior, apparently trying to entice the female to enter the nest. Once she does, both male and female take turns swimming with pelvic fins
pressed against the nest substrate. This behavior may last up to 20 min before actual spawning begins. If the female tries to leave, or
pauses in her passing over the nest, the male resumes swimming in the rapid figure-eight pattern and producing sounds. (For an audio- video clip see web site http://www.mbl.edu/html/BB/VIDEO/ BB.video.html). Once spawning begins, the male no longer pro- duces sound. A male will often nest with several females over a few days.
A. sordidus possibly uses CAS acoustics only to attract atten- tion. In contrast, males in the genera Dascyllus and Stegastes repeatedly swim in vertical loops over their nest sites while mak-
ing a stereotypical, loud, clear, pulsed sound without detectable hydrodynamic noise. In these species, the female approaches the
pattern found in other pomacentrids. The CAS were highly variable in pulse number and length, and the linear relationship between the two factors was very weak (r2 = 0.23; Fig. la). Furthermore, the CAS of A. sordidus were relatively low amplitude and could not be detected unless the hydrophone was within about 2 m. In contrast, D. albisella sounds were detected at distances up to about 9 m (8).
Male A. sordidus establish and maintain territories for nesting, often returning to the same nest site every year (9). Males compete aggressively for their own exclusive nest site. These nests are
arranged in a colonial arena layout. Courting males change col- oration from the drab barred pattern shown by non-courting males and by females to a bright contrasting pattern of white bars on a black background (Fig. lb, c). A male responds to a female in the
vicinity of the nest by rapidly swimming out toward her in a zigzag pattern, swimming first on one side, then flipping to the other. Often more than one male is seen swimming out to meet a female. The successful male leads the female back to his nest; when near the nest, he swims rapidly around her in a figure-eight pattern. During this courtship behavior, sounds are produced (Fig. ld, e). Clearly, the female will be aware of the male's activity in front of her, with or without sounds. The male continues this behavior, apparently trying to entice the female to enter the nest. Once she does, both male and female take turns swimming with pelvic fins
pressed against the nest substrate. This behavior may last up to 20 min before actual spawning begins. If the female tries to leave, or
pauses in her passing over the nest, the male resumes swimming in the rapid figure-eight pattern and producing sounds. (For an audio- video clip see web site http://www.mbl.edu/html/BB/VIDEO/ BB.video.html). Once spawning begins, the male no longer pro- duces sound. A male will often nest with several females over a few days.
A. sordidus possibly uses CAS acoustics only to attract atten- tion. In contrast, males in the genera Dascyllus and Stegastes repeatedly swim in vertical loops over their nest sites while mak-
ing a stereotypical, loud, clear, pulsed sound without detectable hydrodynamic noise. In these species, the female approaches the
male in his nest before the male responds specifically to her. The notion is that females are attracted by the information content of the sounds or the pattern of production (1, 2, 4).
The CAS patterns produced by the four male A. sordidus ex- amined were irregular and highly variable. The sound appears to result from a combination of hydrodynamic noise produced by vigorous swimming movements with fins extended, and biological sounds produced by the movement of muscles and bones, possibly amplified by the swimbladder. The exact mechanism of sound
production is unknown. It seems unlikely that the pulsed sounds
produced by A. sordidus represent advanced coded signals; they are possibly, at least in part, adventitious. When compared to the
highly stylized and repetitious acoustic and swimming display exhibited by Dascyllus and Stegastes spp., the courtship-associated sounds and behavior of A. sordidus seem rudimentary. This dif- ference in acoustic displays probably reflects a species-specific reproductive tactic by which males attract females to the nest.
Supported by the Army Research Office grant DAAG 55-98-1- 0304 for the Johnston Atoll Reef Study.
Literature Cited
1. Myrberg, A. A. 1972. Anim. Behav. Monogr. 5: 197-283. 2. Myrberg, A. A., S. J. Ha, and M. J. Shamblott. 1993. J. Acoust.
Soc. Am. 94: 3067-3070. 3. Mann, D. A., and P. S. Lobel. 1998. Environ. Biol. Fishes 51:
421-428. 4. Lobel, P. S., and D. A. Mann. 1995. Bioacoustics 6: 187-198. 5. Mann, D. A., and P. S. Lobel. 1995. Bioacoustics 6: 199-213. 6. Kerr, L. M. 1996. Biol. Bull. 191: 306-307. 7. Kerr, L. M., K. L. Lang, and P. S. Lobel. 1997. Biol. Bull. 193:
279-281. 8. Mann, D. A., and P. S. Lobel. 1997. J. Acoust. Soc. Am. 101:
3783-3791. 9. Stanton, F. G. 1985. Proc. Fifth Int. Coral Reef Congr. Tahiti. 5:
361-366.
male in his nest before the male responds specifically to her. The notion is that females are attracted by the information content of the sounds or the pattern of production (1, 2, 4).
The CAS patterns produced by the four male A. sordidus ex- amined were irregular and highly variable. The sound appears to result from a combination of hydrodynamic noise produced by vigorous swimming movements with fins extended, and biological sounds produced by the movement of muscles and bones, possibly amplified by the swimbladder. The exact mechanism of sound
production is unknown. It seems unlikely that the pulsed sounds
produced by A. sordidus represent advanced coded signals; they are possibly, at least in part, adventitious. When compared to the
highly stylized and repetitious acoustic and swimming display exhibited by Dascyllus and Stegastes spp., the courtship-associated sounds and behavior of A. sordidus seem rudimentary. This dif- ference in acoustic displays probably reflects a species-specific reproductive tactic by which males attract females to the nest.
Supported by the Army Research Office grant DAAG 55-98-1- 0304 for the Johnston Atoll Reef Study.
Literature Cited
1. Myrberg, A. A. 1972. Anim. Behav. Monogr. 5: 197-283. 2. Myrberg, A. A., S. J. Ha, and M. J. Shamblott. 1993. J. Acoust.
Soc. Am. 94: 3067-3070. 3. Mann, D. A., and P. S. Lobel. 1998. Environ. Biol. Fishes 51:
421-428. 4. Lobel, P. S., and D. A. Mann. 1995. Bioacoustics 6: 187-198. 5. Mann, D. A., and P. S. Lobel. 1995. Bioacoustics 6: 199-213. 6. Kerr, L. M. 1996. Biol. Bull. 191: 306-307. 7. Kerr, L. M., K. L. Lang, and P. S. Lobel. 1997. Biol. Bull. 193:
279-281. 8. Mann, D. A., and P. S. Lobel. 1997. J. Acoust. Soc. Am. 101:
3783-3791. 9. Stanton, F. G. 1985. Proc. Fifth Int. Coral Reef Congr. Tahiti. 5:
361-366.
Reference: Biol. Bull. 197: 244-246. (October 1999)
Threat-Sensitive Nest Defense in Domino Damselfish, Dascyllus albisella Steven J. Oliver and Elise Watson (Boston University Marine Program, Woods Hole, Massachusetts)
Reference: Biol. Bull. 197: 244-246. (October 1999)
Threat-Sensitive Nest Defense in Domino Damselfish, Dascyllus albisella Steven J. Oliver and Elise Watson (Boston University Marine Program, Woods Hole, Massachusetts)
Among fishes with paternal care of eggs, damselfish (Pisces: Pomacentridae) are particularly well-studied (1). Species-specific aggression has been reported for algal-gardening damselfish (2, 3, 4, 5, 6). We initiated this study to determine whether the plank- tivorous Domino damselfish (Dascyllus albisella) exhibits species- specific nest-defense patterns. D. albisella is a common and con-
spicuous reef fish endemic to the Hawaiian archipelago. Males
guard nest-territories against conspecifics year round, and chase
potential egg predators when defending embryos. Courtship in- cludes an acoustic and visual display and is followed by the
deposition of demersal eggs, which the male fertilizes and guards until they hatch (4 days) as planktonic larvae (7).
Williams (8) suggested that paternal care in fishes adds little addi- tional cost to defending mating territories. In algal-gardening species,
Among fishes with paternal care of eggs, damselfish (Pisces: Pomacentridae) are particularly well-studied (1). Species-specific aggression has been reported for algal-gardening damselfish (2, 3, 4, 5, 6). We initiated this study to determine whether the plank- tivorous Domino damselfish (Dascyllus albisella) exhibits species- specific nest-defense patterns. D. albisella is a common and con-
spicuous reef fish endemic to the Hawaiian archipelago. Males
guard nest-territories against conspecifics year round, and chase
potential egg predators when defending embryos. Courtship in- cludes an acoustic and visual display and is followed by the
deposition of demersal eggs, which the male fertilizes and guards until they hatch (4 days) as planktonic larvae (7).
Williams (8) suggested that paternal care in fishes adds little addi- tional cost to defending mating territories. In algal-gardening species,
such as Stegastes spp., there is evidence of non-depreciable care: eggs do not appreciably increase defense costs because heterospecifics are
already excluded from the territories of these species. In D. albisella, however, the cost of nest defense in terms of energy expenditure (attacking of intruders) and energy uptake (time spent feeding) is largely additive because the fish undergo a behavioral shift in terri-
toriality upon reception of eggs. One strategy for ameliorating the cost of paternal care is selective defense proportional to the threat of egg predation posed by different species.
The authors observed male D. albisella (n = 23; standard length (SL) + SD = 89.9 + 11.2 mm) at Johnston Atoll, Central Pacific Ocean, in March-July 1998. Water depth of the lagoon study site ranged from 4 to 5 m. Using scuba, 15-min focal samples were conducted at times ranging from sunrise (0545 h) to sundown
such as Stegastes spp., there is evidence of non-depreciable care: eggs do not appreciably increase defense costs because heterospecifics are
already excluded from the territories of these species. In D. albisella, however, the cost of nest defense in terms of energy expenditure (attacking of intruders) and energy uptake (time spent feeding) is largely additive because the fish undergo a behavioral shift in terri-
toriality upon reception of eggs. One strategy for ameliorating the cost of paternal care is selective defense proportional to the threat of egg predation posed by different species.
The authors observed male D. albisella (n = 23; standard length (SL) + SD = 89.9 + 11.2 mm) at Johnston Atoll, Central Pacific Ocean, in March-July 1998. Water depth of the lagoon study site ranged from 4 to 5 m. Using scuba, 15-min focal samples were conducted at times ranging from sunrise (0545 h) to sundown
244 244
This content downloaded from 185.31.195.48 on Sat, 28 Jun 2014 09:43:53 AMAll use subject to JSTOR Terms and Conditions
PISCINE NEUROBIOLOGY AND BEHAVIOR
(2030 h), with more than 80% of the observations made between 0800 and 1100 h. Males were tagged to allow individuals to be
recognized. We measured attack distances by placing three 5-m
nylon lines (marked in 0.5-m intervals) on the substrate, radiating outward from the nest. The observer estimated attack distances to the nearest 0.25 m. Heterospecific intrusions within 1 m of the nest were recorded (noting species, response, and distance from nest), as were all attacks by the focal male. Encounters were defined as
any incursion within 1 m of the nest, or as an attack on an intruder
regardless of the distance from the nest. We defined attacks as those encounters in which males oriented towards, swam at, or
physically struck the intruder and the intruder fled. Males often
pursued intruders several meters from the nest. Encounters between D. albisella and other fishes occurred for 43
species. Twenty-three species (representing 10 families) were ex- cluded from analysis because they were encountered only in the
presence of eggs. A total of 346 encounters, resulting in 119 attacks (34.4% attack rate) were observed for 19 species (Table I). Wrasses (Labridae) were the only family in which individual
species differed in the response elicited from D. albisella, and are listed both by family and by species. The remaining 13 species are listed by family. One species, the piscivorous Oxychelinus unifas- ciatus (Labridae), was consistently attacked by D. albisella in the absence of embryos. Because 0. unifasciatus appears to be unique in this regard, we excluded it from further analysis.
To test the hypothesis that D. albisella selectively attacks active
egg predators, we divided the species into two groups. At Johnston atoll, the wrasses Thalassoma duperrey and T. lutescens hybridize (9), making identification of species difficult. This hybrid complex was the only group of fish that were observed to actively search for
nests, and to persistently return to a nest once it was discovered. We grouped the Thalassoma spp. together as the T. duperrey X T. lutescens species complex (hereafter referred to as "Thalassoma hybrids"). We grouped the other fishes (Acanthuridae, Monacan- thidae, Chaetodontidae, Mullidae, Scaridae, and the labrids Epi- belus insidiator, Gomphosis varius, and Coris gaimard) together because they were not observed to actively search for, and raid, nests, although they would consume embryos if they discovered a nest. With the exception of Aluterus scriptus (Monacanthidae), which can achieve a standard length of 760 mm, the fishes being attacked were roughly 2 to 3 times the size of the territorial male.
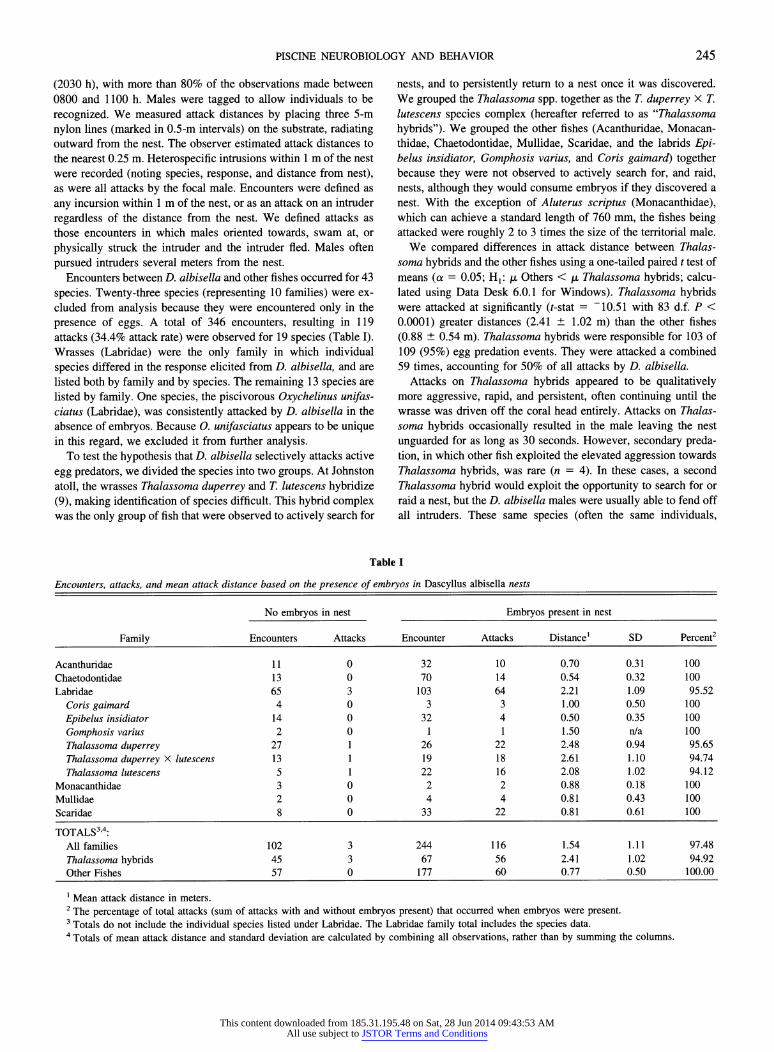
We compared differences in attack distance between Thalas- soma hybrids and the other fishes using a one-tailed paired t test of means (a = 0.05; HI: ,t Others < ,u Thalassoma hybrids; calcu- lated using Data Desk 6.0.1 for Windows). Thalassoma hybrids were attacked at significantly (t-stat = -10.51 with 83 d.f. P < 0.0001) greater distances (2.41 + 1.02 m) than the other fishes (0.88 + 0.54 m). Thalassoma hybrids were responsible for 103 of 109 (95%) egg predation events. They were attacked a combined 59 times, accounting for 50% of all attacks by D. albisella.
Attacks on Thalassoma hybrids appeared to be qualitatively more aggressive, rapid, and persistent, often continuing until the wrasse was driven off the coral head entirely. Attacks on Thalas- soma hybrids occasionally resulted in the male leaving the nest
unguarded for as long as 30 seconds. However, secondary preda- tion, in which other fish exploited the elevated aggression towards Thalassoma hybrids, was rare (n = 4). In these cases, a second Thalassoma hybrid would exploit the opportunity to search for or raid a nest, but the D. albisella males were usually able to fend off all intruders. These same species (often the same individuals,
Table I
Encounters, attacks, and mean attack distance based on the presence of embryos in Dascyllus albisella nests
No embryos in nest Embryos present in nest
Family Encounters Attacks Encounter Attacks Distance' SD Percent2
Acanthuridae 11 0 32 10 0.70 0.31 100 Chaetodontidae 13 0 70 14 0.54 0.32 100 Labridae 65 3 103 64 2.21 1.09 95.52
Coris gaimard 4 0 3 3 1.00 0.50 100
Epibelus insidiator 14 0 32 4 0.50 0.35 100
Gomphosis varius 2 0 1 1 1.50 n/a 100 Thalassoma duperrey 27 1 26 22 2.48 0.94 95.65 Thalassoma duperrey X lutescens 13 1 19 18 2.61 1.10 94.74 Thalassoma lutescens 5 1 22 16 2.08 1.02 94.12
Monacanthidae 3 0 2 2 0.88 0.18 100 Mullidae 2 0 4 4 0.81 0.43 100 Scaridae 8 0 33 22 0.81 0.61 100
TOTALS3'4: All families 102 3 244 116 1.54 1.11 97.48 Thalassoma hybrids 45 3 67 56 2.41 1.02 94.92 Other Fishes 57 0 177 60 0.77 0.50 100.00
1 Mean attack distance in meters. 2 The percentage of total attacks (sum of attacks with and without embryos present) that occurred when embryos were present. 3 Totals do not include the individual species listed under Labridae. The Labridae family total includes the species data. 4 Totals of mean attack distance and standard deviation are calculated by combining all observations, rather than by summing the columns.
245
This content downloaded from 185.31.195.48 on Sat, 28 Jun 2014 09:43:53 AMAll use subject to JSTOR Terms and Conditions
REPORTS FROM THE MBL GENERAL SCIENTIFIC MEETINGS REPORTS FROM THE MBL GENERAL SCIENTIFIC MEETINGS
which we identified by their unique markings) that were attacked when embryos were present were tolerated in the nest when there were no embryos. Since none of these species are cryptic, and many of them are highly conspicuous (see 9), it is unlikely that the ability of D. albisella to perceive the fishes explains this pattern.
These patterns suggest that D. albisella exhibits a threat-depen- dent level of aggression in which the size of the defended area around the nest varies as a function of relative threat of egg predation.
Funding provided by Army Research Office Grant DAAG 559810304.
which we identified by their unique markings) that were attacked when embryos were present were tolerated in the nest when there were no embryos. Since none of these species are cryptic, and many of them are highly conspicuous (see 9), it is unlikely that the ability of D. albisella to perceive the fishes explains this pattern.
These patterns suggest that D. albisella exhibits a threat-depen- dent level of aggression in which the size of the defended area around the nest varies as a function of relative threat of egg predation.
Funding provided by Army Research Office Grant DAAG 559810304.
Literature Cited
1. Petersen, C. W. 1995. Bull. Mar. Sci. 57: 690-704. 2. Draud, M. J., and M. Itzkowitz. 1995. Copeia 2: 431-435. 3. Ebersole, J. P. 1980. Am. Nat. 115(4): 492-509. 4. Myrberg, A. A., and R. E. Thresher. 1974. Am. Zool. 14: 81-96. 5. Thresher, R. E. 1976. Anim. Behav. 24: 562-569. 6. Thresher, R. E. 1979. Mar. Behav. Physiol. 6: 83-94. 7. Mann, D. A., and P. S. Lobel. 1995. Bioacoustics 6: 199-213. 8. Williams, G. C. 1975. Sex and Evolution. Princeton, New Jersey. 9. Randall, J. E. 1996. Shore Fishes of Hawaii. Natural World Press,
Vida, Oregon.
Literature Cited
1. Petersen, C. W. 1995. Bull. Mar. Sci. 57: 690-704. 2. Draud, M. J., and M. Itzkowitz. 1995. Copeia 2: 431-435. 3. Ebersole, J. P. 1980. Am. Nat. 115(4): 492-509. 4. Myrberg, A. A., and R. E. Thresher. 1974. Am. Zool. 14: 81-96. 5. Thresher, R. E. 1976. Anim. Behav. 24: 562-569. 6. Thresher, R. E. 1979. Mar. Behav. Physiol. 6: 83-94. 7. Mann, D. A., and P. S. Lobel. 1995. Bioacoustics 6: 199-213. 8. Williams, G. C. 1975. Sex and Evolution. Princeton, New Jersey. 9. Randall, J. E. 1996. Shore Fishes of Hawaii. Natural World Press,
Vida, Oregon.
Reference: Biol. Bull. 197: 246-247. (October 1999)
Predator-prey Interactions of Juvenile Toadfish, Opsanus tau Nichole N. Price1'2 and Allen F. Mensinger2
(Washington University School of Medicine, St. Louis, Missouri 63110)
Reference: Biol. Bull. 197: 246-247. (October 1999)
Predator-prey Interactions of Juvenile Toadfish, Opsanus tau Nichole N. Price1'2 and Allen F. Mensinger2
(Washington University School of Medicine, St. Louis, Missouri 63110)
The oyster toadfish, Opsanus tau, is a benthic teleost that inhabits estuaries and coastal waters along the eastern seaboard of the United States (1). The toadfish is an ambush predator that
commonly feeds on crabs, shrimps, and fishes (2). The toadfish has also been historically an important marine
model for research on auditory physiology, muscle physiology, vestibular system, and nerve regeneration (3, 4, 5). Although much is known about this animal in its adult stages, the behavior of
juvenile toadfish is poorly understood. A concurrent project at the Marine Biological Laboratory is attempting to culture toadfish (6). If this effort is to be successful, the feeding behaviors of these
young fish must be understood. The aim of this study was to
investigate the relationship between strike distance and strike success of first year toadfish.
One-year old juvenile Opsanus tau (n = 6; 5.9 ? 0.3 cm, standard length) were maintained separately in 80-1 Plexiglas tanks
provided with flow-through ambient seawater (20? + 2?C). The toadfish were transferred to feeding arenas (opaque 6.5-1 aquari- ums) containing seawater at 20? + 1?C and allowed to acclimate for 30 min, which was sufficient time for the fish to resume their normal respiration rates and initiate their feeding behaviors. For each feeding trial, five feeder guppies (about 2 cm sl) were placed in the arena, and the experiment was conducted for 30 min. The interval between trials was no less than 48 h to prevent satiation. Trials were recorded on 8-mm tape with a Sony video High 8
Handycam positioned 0.5 m above the tank. Grids (0.25 cm2) were
placed on the bottom of the tank so that measurements on the VHS
recordings could be calibrated. All strikes were re-recorded onto VHS to be viewed frame by frame on a ZenithInteq VCR (30 frames per second).
The typical toadfish strike sequence consists of an orientation towards the prey followed by a sudden contraction of the pectoral fins, which propels the toadfish at the prey. Just prior to striking,
The oyster toadfish, Opsanus tau, is a benthic teleost that inhabits estuaries and coastal waters along the eastern seaboard of the United States (1). The toadfish is an ambush predator that
commonly feeds on crabs, shrimps, and fishes (2). The toadfish has also been historically an important marine
model for research on auditory physiology, muscle physiology, vestibular system, and nerve regeneration (3, 4, 5). Although much is known about this animal in its adult stages, the behavior of
juvenile toadfish is poorly understood. A concurrent project at the Marine Biological Laboratory is attempting to culture toadfish (6). If this effort is to be successful, the feeding behaviors of these
young fish must be understood. The aim of this study was to
investigate the relationship between strike distance and strike success of first year toadfish.
One-year old juvenile Opsanus tau (n = 6; 5.9 ? 0.3 cm, standard length) were maintained separately in 80-1 Plexiglas tanks
provided with flow-through ambient seawater (20? + 2?C). The toadfish were transferred to feeding arenas (opaque 6.5-1 aquari- ums) containing seawater at 20? + 1?C and allowed to acclimate for 30 min, which was sufficient time for the fish to resume their normal respiration rates and initiate their feeding behaviors. For each feeding trial, five feeder guppies (about 2 cm sl) were placed in the arena, and the experiment was conducted for 30 min. The interval between trials was no less than 48 h to prevent satiation. Trials were recorded on 8-mm tape with a Sony video High 8
Handycam positioned 0.5 m above the tank. Grids (0.25 cm2) were
placed on the bottom of the tank so that measurements on the VHS
recordings could be calibrated. All strikes were re-recorded onto VHS to be viewed frame by frame on a ZenithInteq VCR (30 frames per second).
The typical toadfish strike sequence consists of an orientation towards the prey followed by a sudden contraction of the pectoral fins, which propels the toadfish at the prey. Just prior to striking,
the toadfish begins to raise its head and lower its mandible rapidly, generating an inward rush of water that may assist in the capture of the prey (7). Upon contacting the prey, the toadfish quickly snaps its jaws around it. Strike initiation was defined as the frame in which the pectoral fins begin to contract. Strike distance was defined as the distance between the head of the fish and the head of the guppy measured at the time of strike initiation (8).
The toadfish exhibited a 34.6% success rate at capturing the guppies (n = 78 attempts). Successful attempts were initiated at an
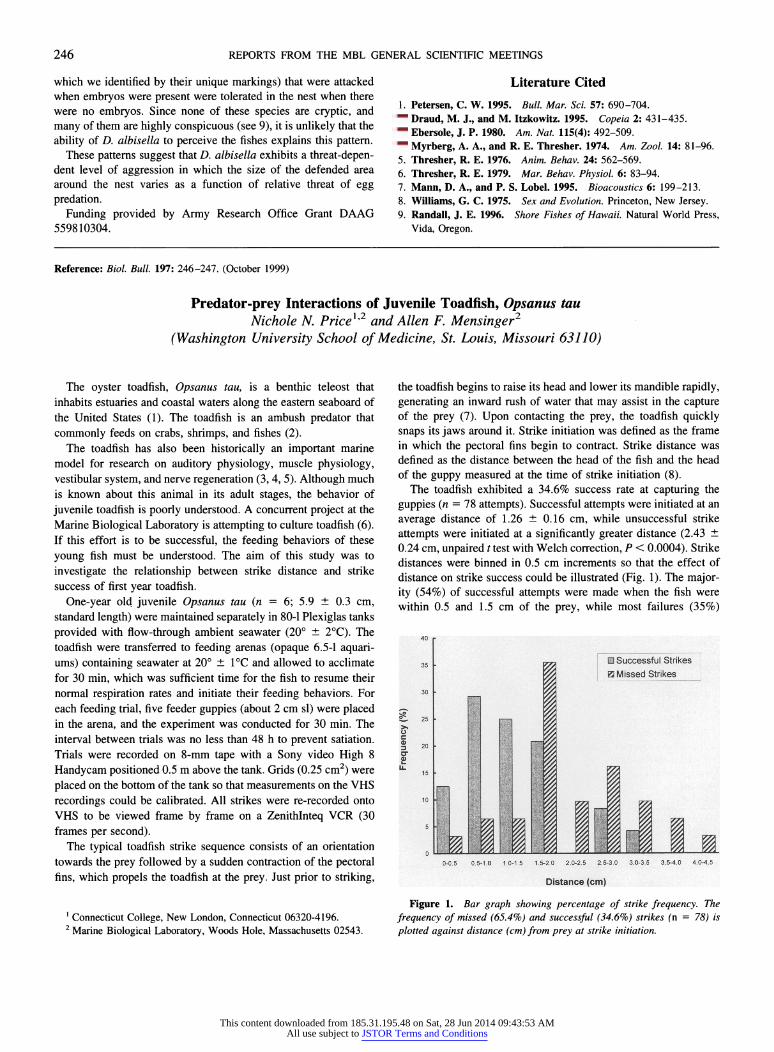
average distance of 1.26 ? 0.16 cm, while unsuccessful strike attempts were initiated at a significantly greater distance (2.43 + 0.24 cm, unpaired t test with Welch correction, P < 0.0004). Strike distances were binned in 0.5 cm increments so that the effect of distance on strike success could be illustrated (Fig. 1). The major- ity (54%) of successful attempts were made when the fish were within 0.5 and 1.5 cm of the prey, while most failures (35%)
the toadfish begins to raise its head and lower its mandible rapidly, generating an inward rush of water that may assist in the capture of the prey (7). Upon contacting the prey, the toadfish quickly snaps its jaws around it. Strike initiation was defined as the frame in which the pectoral fins begin to contract. Strike distance was defined as the distance between the head of the fish and the head of the guppy measured at the time of strike initiation (8).
The toadfish exhibited a 34.6% success rate at capturing the guppies (n = 78 attempts). Successful attempts were initiated at an
average distance of 1.26 ? 0.16 cm, while unsuccessful strike attempts were initiated at a significantly greater distance (2.43 + 0.24 cm, unpaired t test with Welch correction, P < 0.0004). Strike distances were binned in 0.5 cm increments so that the effect of distance on strike success could be illustrated (Fig. 1). The major- ity (54%) of successful attempts were made when the fish were within 0.5 and 1.5 cm of the prey, while most failures (35%)
40 r 40 r
35 F 35 F
30 o 30 o
I I
25
20
15
25
20
15
10
5
0
10
5
0
Sucessful Strikes 1 B Missed Strikes
Sucessful Strikes 1 B Missed Strikes
I I
0. 1.2.0 202. 2 3.0 3. .5 3.4.0 4. o. .1.0 1.5 .i e 2.02.6 S3.0 3.0 3.5. 4.cm.5
Distance (cm)
0. 1.2.0 202. 2 3.0 3. .5 3.4.0 4. o. .1.0 1.5 .i e 2.02.6 S3.0 3.0 3.5. 4.cm.5
Distance (cm)
'Connecticut College, New London, Connecticut 06320-4196. 2 Marine Biological Laboratory, Woods Hole, Massachusetts 02543. 'Connecticut College, New London, Connecticut 06320-4196. 2 Marine Biological Laboratory, Woods Hole, Massachusetts 02543.
Figure 1. Bar graph showing percentage of strike frequency. The
frequency of missed (65.4%) and successful (34.6%) strikes (n = 78) is
plotted against distance (cm) from prey at strike initiation.
Figure 1. Bar graph showing percentage of strike frequency. The
frequency of missed (65.4%) and successful (34.6%) strikes (n = 78) is
plotted against distance (cm) from prey at strike initiation.
246 246
This content downloaded from 185.31.195.48 on Sat, 28 Jun 2014 09:43:53 AMAll use subject to JSTOR Terms and Conditions





























