Embed Size (px)
Citation preview

Biotechnology Advances 24 (2006) 452–481www.elsevier.com/locate/biotechadv
Research review paper
Outlook for cellulase improvement: Screening and selection strategies
Y.-H. Percival Zhang a,⁎, Michael E. Himmel b, Jonathan R. Mielenz c
a Biological Systems Engineering Department, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061, USAb National Bioenergy Center, National Renewable Energy Laboratory, Golden, CO 80401, USA
c Life Science Division, Oak Ridge National Laboratory, Oak Ridge, TN 37831, USA
Received 31 January 2006; received in revised form 6 March 2006; accepted 11 March 2006Available online 27 March 2006
Abstract
Cellulose is the most abundant renewable natural biological resource, and the production of biobased products and bioenergyfrom less costly renewable lignocellulosic materials is important for the sustainable development of human beings. A reduction incellulase production cost, an improvement in cellulase performance, and an increase in sugar yields are all vital to reduce theprocessing costs of biorefineries. Improvements in specific cellulase activities for non-complexed cellulase mixtures can beimplemented through cellulase engineering based on rational design or directed evolution for each cellulase component enzyme,as well as on the reconstitution of cellulase components. Here, we review quantitative cellulase activity assays using soluble andinsoluble substrates, and focus on their advantages and limitations. Because there are no clear relationships between cellulaseactivities on soluble substrates and those on insoluble substrates, soluble substrates should not be used to screen or selectimproved cellulases for processing relevant solid substrates, such as plant cell walls. Cellulase improvement strategies based ondirected evolution using screening on soluble substrates have been only moderately successful, and have primarily targetedimprovement in thermal tolerance. Heterogeneity of insoluble cellulose, unclear dynamic interactions between insoluble substrateand cellulase components, and the complex competitive and/or synergic relationship among cellulase components limit rationaldesign and/or strategies, depending on activity screening approaches. Herein, we hypothesize that continuous culture usinginsoluble cellulosic substrates could be a powerful selection tool for enriching beneficial cellulase mutants from the large librarydisplayed on the cell surface.© 2006 Elsevier Inc. All rights reserved.
Keywords: Cellulase activity assay; Cellulose; Cellulosome; Continuous culture; Enzymatic cellulose hydrolysis; High throughput screening;Selection; Sugar assay
Abbreviations:AFEX, ammonia fiber explosion; BC, bacterial cellulose; BCA, 2,2′-bicinchroninate; BMCC, bacterial microcrystalline cellulose;CMC, carboxymethyl cellulose; CBM, cellulose-binding module; CBP, consolidated bioprocessing; CrI, crystallinity index; DMAc, N,N-dimethylacetamide; DNS, dinitrosalicyclic acid; DP, degree of polymerization of cellulose; DS, degree of substitution; DTT, dithiothreitol; Fa,fraction of β-glucosidic bond accessible to cellulase; FPA, filter paper activity; FRE, fraction of the reducing end to all anhydroglucose units ofcellulose, 1/DP; HEC, hydroxyethyl cellulose; PASC, phosphoric acid swollen cellulose; RAC, regenerated amorphous cellulose; PAHBAH, 4-hydroxybenzoylhydrazine; RS, selection ratio; TNP-CMC, trinitrophenyl-carboxymethyl cellulose.⁎ Corresponding author. Tel.: +1 540 231 7414; fax: +1 540 231 3199.E-mail address: [email protected] (Y.-H. Percival Zhang).
0734-9750/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.biotechadv.2006.03.003

453Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4532. Cellulose hydrolysis mechanisms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4553. Substrates for cellulase activity assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 457
3.1. Soluble substrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4583.2. Insoluble substrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 458
4. Quantitative assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4604.1. Hydrolysis products . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4614.2. Cellulase activity assays. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 462
4.2.1. Endoglucanases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4624.2.2. Exoglucanases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4634.2.3. β-D-glucosidases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4644.2.4. Total cellulase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 464
5. Cellulase improvement and screening/selection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4655.1. Rational design . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4655.2. Directed evolution. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4675.3. Screening . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4695.4. Selection . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 470
6. Summary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 472Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 472References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 472
1. Introduction
Cellulose is the primary product of photosynthesis interrestrial environments, and the most abundant renew-able bioresource produced in the biosphere (∼100billion dry tons/year) (Holtzapple, 1993; Jarvis, 2003;Zhang and Lynd, 2004b). Cellulose biodegradation bycellulases and cellulosomes, produced by numerousmicroorganisms, represents a major carbon flow fromfixed carbon sinks to atmospheric CO2 (Berner, 2003;Falkowski et al., 2000; Melillo et al., 2002), is veryimportant in several agricultural and waste treatmentprocesses (Angenent et al., 2004; Das and Singh, 2004;Haight, 2005; Hamer, 2003; Humphrey et al., 1977;Russell and Rychlik, 2001; Schloss et al., 2005; vanWyk, 2001), and could be widely used to producesustainable biobased products and bioenergy to replacedepleting fossil fuels (Angenent et al., 2004; Demain etal., 2005; Galbe and Zacchi, 2002; Hall et al., 1993;Hoffert et al., 2002; Kamm and Kamm, 2004; Lynd,1996; Lynd et al., 1991, 2002, 1999; Mielenz, 2001;Mohanty et al., 2000; Moreira, 2005; Reddy and Yang,2005; Wyman, 1994, 1999, 2003). Additionally, studieshave shown that the use of biobased products andbioenergy can achieve zero net carbon dioxide emission(Demain, 2004; Demain et al., 2005; Hoffert et al.,2002; Lynd et al., 1991, 1999). Development oftechnologies for effectively converting less costlyagricultural and forestry residues to fermentable sugars
offers outstanding potential to benefit the nationalinterest through: (1) improved strategic security, (2)decreased trade deficits, (3) healthier rural economies,(4) improved environmental quality, (5) technologyexports, and (6) a sustainable energy resource supply(Angenent et al., 2004; Caldeira et al., 2003; Demain etal., 2005; Hoffert et al., 1998, 2002; Kamm and Kamm,2004; Lynd, 1996; Lynd et al., 1991, 1999, 2002;Moreira, 2005; Wirth et al., 2003; Wyman, 1999).
Effective conversion of recalcitrant lignocelluloseto fermentable sugars requires three sequential steps:(1) size reduction, (2) pretreatment/fractionation, and(3) enzymatic hydrolysis (Wyman, 1999; Zhang andLynd, 2004b). One of the most important anddifficult technological challenges is to overcome therecalcitrance of natural lignocellulosic materials,which must be enzymatically hydrolyzed to producefermentable sugars (Chang et al., 1981; Demain etal., 2005; Fan et al., 1982; Grethlein, 1984; Hsu,1996; Lin et al., 1981; McMillian, 1994; Millett etal., 1976; Moreira, 2005; Mosier et al., 2005; Saddleret al., 1993; Weil et al., 1994; Wyman, 1999; Wymanet al., 2005a).
Cellulases are relatively costly enzymes, and asignificant reduction in cost will be important for theircommercial use in biorefineries. Cellulase-based strat-egies that will make the biorefinery processing moreeconomical include: increasing commercial enzymevolumetric productivity, producing enzymes using

454 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
cheaper substrates, producing enzyme preparations withgreater stability for specific processes, and producingcellulases with higher specific activity on solidsubstrates. Recently, the biotechnology companiesGenencor International and Novozymes Biotech havereported the development of technology that hasreduced the cellulase cost for the cellulose-to-ethanolprocess from US$5.40 per gallon of ethanol toapproximately 20 cents per gallon of ethanol (Moreira,2005), in which the two main strategies were (1) aneconomical improvement in production of cellulase toreduce US$ per gram of enzyme by process and strainenhancement, e.g., cheaper medium from lactose toglucose and alternative inducer system and (2) animprovement in the cellulase enzyme performance toreduce grams of enzyme for achieving equivalenthydrolysis by cocktails and component improvement(Knauf and Moniruzzaman, 2004). But this claim hasnot yet been widely accepted because the cellulasemixture was tested only for the specific pretreatedlignocellulosic substrate and cannot be applied to otherpretreated lignocelluloses.
Currently, most commercial cellulases (including β-glucosidase) are produced by Trichoderma species andAspergillus species (Cherry and Fidantsef, 2003;Esterbauer et al., 1991; Kirk et al., 2002). Cellulasesare used in the textile industry for cotton softening anddenim finishing; in the detergent market for color care,cleaning, and anti-deposition; in the food industry formashing; and in the pulp and paper industries for de-
Rational Design
Scre
so
Improved cellulase
components
Reconstitute
cellulase
cocktail
W
Cellulase
Fig. 1. Scheme of cellulase engineering for non-complexed cellulases. EndexosNR, exoglucanases acting on non-reducing ends; β-Gase, β-glucosidase
inking, drainage improvement, and fiber modification(Cherry and Fidantsef, 2003; Kirk et al., 2002). Thecellulase market is expected to expand dramaticallywhen cellulases are used to hydrolyze pretreatedcellulosic materials to sugars, which can be fermentedto commodities such as bioethanol and biobasedproducts on a large scale (Cherry and Fidantsef, 2003;Himmel et al., 1999; van Beilen and Li, 2002). Forexample, the potential cellulase market has beenestimated to be as high as US$400 million per year ifcellulases are used for hydrolyzing the available cornstover in the midwestern United States (van Beilen andLi, 2002). This market scenario represents an increase of∼33% in the total US industrial enzyme market(Wolfson, 2005). The large market potential and theimportant role that cellulases play in the emergingbioenergy and bio-based products industries provide agreat motivation to develop better cellulase preparationsfor plant cell wall cellulose hydrolysis. These improvedcellulases must also have characteristics necessary forbiorefineries, such as higher catalytic efficiency oninsoluble cellulosic substrates, increased stability atelevated temperature and at a certain pH, and highertolerance to end-product inhibition.
Fig. 1 shows that cellulase engineering for non-complexed cellulase systems contains three majorresearch directions: (1) rational design for eachcellulase, based on knowledge of the cellulase structureand the catalytic mechanism (Schulein, 2000; Wilson,2004; Wither, 2001); (2) directed evolution for each
Directed Evolution
endos
exosR
expsNR
β-Gase
en or select on
lid substrate
ild type
Components
os, endoglucanases; exosR, exoglucanases acting on reducing ends;.

455Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
cellulase, in which the improved enzymes or ones withnew properties were selected or screened after randommutagenesis and/or molecular recombination (Arnold,2001; Cherry and Fidantsef, 2003; Hibbert et al., 2005;Schmidt-Dannert and Arnold, 1999; Shoemaker et al.,2003; Tao and Cornish, 2002); and (3) the reconstitutionof cellulase mixtures (cocktails) active on insolublecellulosic substrates, yielding an improved hydrolysisrate or higher cellulose digestibility (Baker et al., 1998;Boisset et al., 2001; Himmel et al., 1999; Irwin et al.,1993; Kim et al., 1998; Sheehan and Himmel, 1999;Walker et al., 1993; Wilson and Walker, 1991; Zhangand Lynd, 2004b). With respect to engineering com-plexed cellulase systems (cellulosomes), the idea ofchimeric constructs of cellulosomal domains/compo-nents was proposed by Bayer et al. (1994), and thereconstruction of cellulosome components is becominganother hot research area (Fierobe et al., 2001, 2002,2005; Mingardon et al., 2005; Sabathe and Soucaille,2003), which we do not review here.
The cornerstone of enzyme engineering is to achievea direct correlation between the enzyme assays orscreening approaches and the changes in enzymefunctions in the desired application. Development of auseful, predictive cellulase assay or screening isparticularly difficult because of the nature of solidheterogeneous substrates, such as plant cell walls.Available quantitative cellulase assays and screeningshave been analyzed and compared herein, includingtheir advantages and limitations. Also, successfulcellulase examples using directed evolution are exam-ined, and a possible strategy of combinatorial molecularbreeding and continuous culture with solid cellulosicmaterials to select a cellulase with higher activity isdiscussed.
2. Cellulose hydrolysis mechanisms
Cellulose is a linear condensation polymer consistingof D-anhydroglucopyranose joined together by β-1,4-glycosidic bonds with a degree of polymerization (DP)from 100 to 20,000 (Krassig, 1993; O'Sullivan, 1997;Tomme et al., 1995; Zhang and Lynd, 2004b).Anhydrocellobiose is the repeating unit of cellulose.Coupling of adjacent cellulose chains and sheets ofcellulose by hydrogen bonds and van der Waal's forcesresults in a parallel alignment and a crystalline structurewith straight, stable supra-molecular fibers of greattensile strength and low accessibility (Demain et al.,2005; Krassig, 1993; Nishiyama et al., 2003; Notley etal., 2004; Zhang and Lynd, 2004b; Zhbankov, 1992).The cellulose molecule is very stable, with a half life of
5–8 million years for β-glucosidic bond cleavage at 25°C (Wolfenden and Snider, 2001), while the much fasterenzyme-driven cellulose biodegradation process is vitalto return the carbon in sediments to the atmosphere(Berner, 2003; Cox et al., 2000; Falkowski et al., 2000;Schlamadinger and Marland, 1996).
The widely accepted mechanism for enzymaticcellulose hydrolysis involves synergistic actions byendoglucanase (EC 3.2.1.4), exoglucanase or cellobio-hydrolase (EC 3.2.1.91), and β-glucosidase (EC3.2.1.21) (Henrissat, 1994; Knowles et al., 1987;Lynd et al., 2002; Teeri, 1997; Wood and Garica-Campayo, 1990; Zhang and Lynd, 2004b). Endoglu-canases hydrolyze accessible intramolecular β-1,4-glucosidic bonds of cellulose chains randomly toproduce new chain ends; exoglucanases processivelycleave cellulose chains at the ends to release solublecellobiose or glucose; and β-glucosidases hydrolyzecellobiose to glucose in order to eliminate cellobioseinhibition. These three hydrolysis processes occursimultaneously as shown in Fig. 2. Primary hydrolysisthat occurs on the surface of solid substrates releasessoluble sugars with a degree of polymerization (DP) upto 6 into the liquid phase upon hydrolysis byendoglucanases and exoglucanases. The enzymaticdepolymerization step performed by endoglucanasesand exoglucanases is the rate-limiting step for thewhole cellulose hydrolysis process. Secondary hydro-lysis that occurs in the liquid phase involves primarilythe hydrolysis of cellobiose to glucose by β-glucosi-dases, although some β-glucosidases also hydrolyzelonger cellodextrins (Zhang and Lynd, 2004b). Duringcellulose hydrolysis, the solid substrate characteristicsvary, including (1) changes in the cellulose chain endnumber resulting from generation by endoglucanasesand consumption by exoglucanases (Kleman-Leyer etal., 1992, 1994, 1996; Kongruang et al., 2004;Srisodsuk et al., 1998; Zhang and Lynd, 2005b) and(2) changes in cellulose accessibility resulting fromsubstrate consumption and cellulose fragmentation(Banka et al., 1998; Boisset et al., 2000; Chanzy etal., 1983; Din et al., 1991, 1994; Halliwell and Riaz,1970; Lee et al., 1996, 2000; Saloheimo et al., 2002;Walker et al., 1990, 1992; Wang et al., 2003;Woodward et al., 1992). The combined actions ofendoglucanases and exoglucanases modify the cellu-lose surface characteristics (topography) over time,resulting in rapid changes in hydrolysis rates.
The complicated interactions among endogluca-nases, exoglucanases, and the changing substratecharacteristics during hydrolysis have been simulatedby a new functionally based mathematical model

Liquid
Phase
(primary
hydrolysis)
(secondary
hydrolysis)
β-Gase
Solid
Phase
endos
exosR
exosNR
Fig. 2. Mechanistic scheme of enzymatic cellulose hydrolysis by Trichoderma non-complexed cellulase system.
456 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
(Zhang and Lynd, in press), applying a set of enzymaticparameters for endoglucanase I, cellobiohydrolases Iand II to a variety of substrates with two importantsubstrate properties: the fraction of β-glucosidic bondaccessible to cellulase (Fa) (Zhang and Lynd, 2004b)and the degree of polymerization (DP) (Okazaki andMoo-Young, 1978; Zhang and Lynd, 2004b) (seeTable 1). In this way, disparate information from the
Table 1Summary of typical values of model celluloses for crystallinity index (CrI),estimated by maximum cellulase adsorption capacity (Zhang and Lynd, 2004of reducing ends (FRE), and relative ratio of FRE/Fa
Substrates CrI Fa(%)
DP
SolubleCellodextrins and their derivatives N.A. 100CMC N.A. 100 1
InsolubleCotton 0.8–0.95 0.2 10Whatman No. 1 filter paper ∼0.45 1.8 7Bacterial cellulose 0.8–0.95 6 6Microcrytalline cellulose (Avicel) 0.5–0.6 0.6 1PASC 0 12 1Pulp (Solka Floc) 0.4–0.7 1.8 7Pretreated cellulosic substrates 0.4–0.7 0.6 4
literature was framed in a coherent way to facilitate anunderstanding of enzymatic cellulose hydrolysis. Forexample, the reaction rates simulated by the modelwere consistent with a substantial number of observa-tions reported in the literature, including the effects ofsubstrate characteristics on exoglucanase and endo-glucanase activities; the effects of substrate character-istics and experimental conditions on the degree of
the fraction of β-glucosidic bond accessible to cellulase (Fa), which isb), the number average of degree of polymerization (DPN), the fraction
N FRE (%) FRE/Fa
Low High Low High
2–6 16.67 50 0.167 0.500–2000 0.05 1 0.0005 0.01
00–3000 0.033 0.1 0.167 0.550–2800 0.036 0.133 0.0198 0.074100–2000 0.05 0.167 0.00833 0.027850–500 0.2 0.667 0.333 1.1100–1000 0.1 1 0.00833 0.083350–1500 0.067 0.133 0.0370 0.074100–1000 0.1 0.25 0.167 0.417

457Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
endo-exo synergy; the effects of endoglucanasepartition coefficient on the hydrolysis rates; and theeffects of enzyme loading on relative reaction rates fordifferent substrates. The model also suggests that it isnearly impossible to predict hydrolysis performance ofcellulase mixtures from one solid substrate to anothersolid substrate, because of large variations in totalcellulase concentration, ratio of endo/exocellulases,reaction time, and substrate characteristics. Therefore,enzyme reconstitution may have be conducted so as toachieve better performance for a specific substrate(Knauf and Moniruzzaman, 2004).
Unlike non-complexed fungal cellulase, anaerobicmicroorganisms possess complexed cellulase systems,called cellulosomes (Bayer et al., 1994, 1998, 2004;Beguin and Alzari, 1998; Demain et al., 2005; Doiand Kosugi, 2004; Doi et al., 1998; Doi and Tamaru,2001; Leschine, 1995; Schwarz, 2001). Leschine(1995) estimated that anaerobic cellulose degradationcould account for only 5–10% of total cellulosebiodegradation, but it could be underestimated becauseanaerobic cellulose hydrolysis is responsible forconsiderable carbon recycling in the anoxic zones ofponds, lakes, oceans, and intestines of ruminants andguts of termites (P.J. Weimer, personal communica-tion). Furthermore, an understanding anaerobic cellu-lase systems are of significant importance to basic
Table 2Substrates containing β-1,4-glucosidic bonds hydrolyzed by cellulases and t
Substrate
SolubleShort chain (low DP)CellodextrinsRadio-labeled cellodextrinsCellodextrin derivativesβ-methylumbelliferyl-oligosaccharidesp-nitrophenol-oligosaccharidesLong chain cellulose derivativesCarboxymethyl cellulose (CMC), hydroxyethyl cellulose (HEC)Dyed CMC
InsolubleCrystalline cellulose-Cotton, microcrystalline cellulose (Avicel),Valonia cellulose, bacterial cellulose
Amorphous cellulose - PASC, alkali-swollen cellulose RACDyed celluloseFluorescent celluloseChromogenic and fluorephoric derivativesTrinitrophenyl-carboxymethylcellulose (TNP-CMC)Fluram-cellulose
Practical cellulose-containing substratesα-cellulose, pretreated lignocellulosic biomass
a RS, reducing sugars; TSS, total soluble sugars.
sciences, such as the evolution of cellulase genes, thestructures of cellulases, and the formation andhydrolysis of reacting biofilms on cellulose surfaces(Lynd et al., in press). Anaerobic cellulose fermenta-tion has both current and future applications, such asagricultural processes anaerobic waste treatment, andconsolidated bioprocessing (CBP), respectively (Lynd,1996; Lynd et al., 1999, 2002, 2005). Recently, amicrobial cellulose hydrolysis mechanism has beenreported for the anaerobic cellulolytic bacteriumClostridium thermocellum that assimilates longersoluble hydrolysis products with an average degreeof polymerization of 4 rather than glucose andcellobiose. The improved bioenergetics resultingfrom longer chain sugar assimilation supports thebiological feasibility of anaerobic fermentation withoutadded saccharolytic enzymes (Zhang and Lynd,2005c). More information about the cellulosome-based microbial cellulose hydrolysis research isavailable elsewhere (Lynd, 1996; Lynd et al., 2002,1999, 2005; Zhang and Lynd, 2003a, 2004a, 2005a).
3. Substrates for cellulase activity assays
Substrates for cellulase activity assays can be dividedinto two categories, based on their solubility in water(Table 2).
heir detections
Detection a Enzymes
RS, HPLC; TLC Endo, Exo, BGTLC plus liquid scintillation Endo, Exo, BG
Fluorophore liberation, TLC Endo, Exo, BGChromophore liberation, TLC Endo, Exo, BG
RS; viscosity EndoDye liberation Endo
RS, TSS, HPLC Total, Endo, ExoRS, TSS, HPLCRS, TSS, HPLC, TLC Total, Endo, ExoDye liberation Total, EndoFluorophore liberation Total
Chromophore liberation EndoFluorophore liberation Endo, Total
HPLC, RS Total

458 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
3.1. Soluble substrates
Soluble substrates include low DP cellodextrins from2 to 6 sugar units and their derivatives, as well as longDP cellulose derivatives (ca. several hundreds of sugarunits). They are often used for measuring individualcellulase component activity (Table 2).
Cellodextrins are soluble for DP≤6, and veryslightly soluble for 6<DP<12 (Miller, 1960; Miller,1963; Pereira et al., 1988; Zhang and Lynd, 2003b,2005b). Their solubility decreases drastically withincreasing DP because of strong intermolecular hydro-gen bonds and system entropic effects. Cellodextrins areoften prepared through cellulose hydrolysis by fumingHCl (Miller, 1960, 1963), sulfuric acid (Voloch et al.,1984), acetylation (Schmid et al., 1988; Wolfram andDacons, 1952), or mixed acids (HCl and H2SO4) (Zhangand Lynd, 2003b). Cellodextrins are also preparedthrough biosynthesis using C. thermocellum cellobioseand cellodextrin phosphorylases (Ng and Zeikus, 1986;Strobel et al., 1995; Zhang and Lynd, 2005c, 2006), or T.reesei β-glucosidase (Chirico and Brown, 1987).Cellodextrin mixtures can be separated into singlecomponents using chromatographic methods such ascharcoal-celite (Miller, 1963), thin layer (Chirico andBrown, 1985; Zhang and Lynd, 2006), cation exchange(Huebner et al., 1978; Voloch et al., 1984), or size-exclusion (Schmid et al., 1988; Shintate et al., 2003;Zhang and Lynd, 2003b, 2006).
Chromogenic p-nitrophenyl glycosides and fluoro-genic methylumbelliferyl-D-glycosides derived fromsoluble cellodextrins are very useful for the study ofinitial cellulase kinetics (Tuohy et al., 2002; Wolfgangand Wilson, 1999), reaction specificity (Bhat et al.,1990; Claeyssens and Aerts, 1992; Tomme et al., 1996;van Tilbeurgh and Claeyssens, 1985; van Tilbeurgh etal., 1982, 1985; Zverlov et al., 2002b), and binding sitethermodynamics (Barr and Holewinski, 2002). They arealso used to determine the inhibition constants ofcellulase in the presence of added cellobiose andglucose (Tuohy et al., 2002), because chromophoresreleased from substituted glycosides can be easilymeasured independently of sugars.
Long DP cellulose derivatives can be dissolved inwater because of their chemical substitutions. Ionic-substituted carboxymethyl cellulose (CMC) is oftenused for determining endoglucanase activity, calledCMCase, because endoglucanases cleave intramolecu-lar β-1,4-glucosidic bonds randomly, resulting in adramatic reduction in the DP (i.e., specific viscosity) ofCMC. CMC has two very important physical parameters—the degree of substitution (DS) and DP. The solubility
of CMC is closely associated with the DS that has amaximum stoichiometric value of 3. CMC is soluble inwater when DS>0.3–0.7 (Karlsson et al., 2001; Klemmet al., 1998a; Wood and Bhat, 1988). CommercialCMCs usually have a DS<1.5. It is strongly recom-mended that a reducing sugar assay or viscosity assayusing CMC as a substrate should be limited to the first2% hydrolysis of substrate when DS=0.7 (Wood andBhat, 1988). This is important because only non-substituted glucose units are accessible to cellulase,and hydrolysis action requires at least two or threecontiguous non-substituted residues. The DP of CMC isnot important for the reducing sugar assay, but it is veryimportant for determining viscosity reduction. CMCdissolution in water should be done by gentle swirling toavoid DP reduction (Sharrock, 1988). Also, theviscosity of ionic CMC is influenced by pH, ionicstrength, and polyvalent cation concentration. There-fore, it is recommended to use nonionic substitutedcelluloses, such as hydroxyethyl cellulose (HEC), fordetermining endoglucanase activity (Wood and Bhat,1988).
Dyed soluble CMC is made by mixing CMC withdyes—Remazol Brilliant Blue R (Fülöp and Ponyi,1997; McCleary, 1980; Wirth and Wolf, 1992) orRuthenium Red (Rescigno et al., 1994). The colorsreleased from soluble cellulose derivatives in thesupernatant can be measured after precipitation of thesoluble substrates. A dyed CMC is also used as asubstrate on solid agar plates, sometimes called“zymograms” (Bera-Maillet et al., 2000; Eriksson andPetterson, 1973; Holt and Hartman, 1994). Afterhydrolysis, staining, and washing, halo zones can beobserved in the colored background.
3.2. Insoluble substrates
Insoluble cellulose-containing substrates for cellu-lase activity assays include nearly pure celluloses(cotton linter, Whatman No. 1 filter paper, bacterialcellulose, microcrystalline cellulose, and amorphouscellulose) and impure cellulose-containing substrates(dyed cellulose, α-cellulose, and pretreated lignocellu-lose). Native cellulose, referred to as cellulose I, has twodistinct crystallite forms, Iα which is dominant inbacterial and algal cellulose, and Iβ, which is dominantin higher plants (Atalla and Vanderhart, 1984). Nativecellulose (cellulose I) can be converted to othercrystalline forms (II–IV) by various treatments (Kleinand Snodgrass, 1993; Krassig, 1993; O'Sullivan, 1997).Table 1 shows several key physical values—crystallin-ity index, degree of polymerization, and cellulose

459Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
accessibility to cellulase that can be estimated based onmaximum cellulase adsorption (Zhang and Lynd,2004b).
The crystallinity index (CrI) of cellulose, quantita-tively measured from its wide range X-ray diffractionpattern (Krassig, 1993; Ramos et al., 2005; Zhang andLynd, 2004b), is not strongly associated with hydrolysisrates (Converse, 1993; Mansfield et al., 1999; Zhangand Lynd, 2004b). Nevertheless, it is still a convenientindicator representing the change in cellulose character-istics for one material before and after treatment. Cotton,bacterial cellulose, and the Valonia ventricosa algalcellulose are examples of highly crystalline cellulose(Boisset et al., 1999; Fierobe et al., 2002), whereasamorphous cellulose is at the other extreme. Microcrys-talline cellulose, filter paper, α-cellulose, and pretreatedcellulosic substrates have modest CrI values, and can beregarded as a combination of crystalline fraction andamorphous fraction, but there is no clear borderlinebetween two fractions.
Cotton fiber is made from natural cotton afterimpurities, such as wax, pectin, and colored matter,have been removed (Wood, 1988). Whatman No.1filter paper is made from long fiber cotton pulp with alow CrI=∼45% (Dong et al., 1998; Henrissat et al.,1985). Microcrystalline cellulose, called hydrocelluloseor avicel (the commercial name), can be purchasedfrom several companies, such as FMC, Merck, andSigma. It is made through the following steps:hydrolysis of wood pulp by dilute hydrochloric acidto remove the amorphous cellulose fraction, formationof colloidal dispersions by high shear fields, followedby spray drying of the washed pulp slurry (Fleming etal., 2001; Zhang and Lynd, 2004b). However,microcrystalline cellulose still contains a significantfraction of amorphous cellulose. Avicel is a goodsubstrate for exoglucanase activity assay, because ithas a low DP and relatively low accessibility (i.e., thehighest ratio of FRE/Fa) (Table 1). Therefore, someresearchers feel that “avicelase” activity is equivalentto exoglucanase activity (Wood and Bhat, 1988).However, some endoglucanases can release consider-able reducing sugars from avicel (Zhang and Lynd,2004b).
Bacterial cellulose (BC) is prepared from the pellicleproduced by Acetobacter xylinum (ATCC 23769)(Hestrin, 1963) or from Nata de Coco (Daiwa FineProduces, Singapore) (Boisset et al., 2000). Bacterialmicrocrystalline cellulose (BMCC) can be preparedfrom BC by partial acid hydrolysis to remove theamorphous cellulose fraction, resulting in a reduction inDP (Valjamae et al., 1999).
Amorphous cellulose is prepared by converting thecrystalline fraction of cellulose to the amorphous formby mechanical or chemical methods. These cellulosesinclude mechanically made amorphous cellulose, alkali-swollen cellulose, and phosphorous acid swollencellulose (PASC, Walseth cellulose). Mechanicallymade amorphous cellulose is often prepared by ballmilling or severe blending (Fan et al., 1980; Ghose,1969; Henrissat et al., 1985; Wood, 1988). Alkali-swollen amorphous cellulose is made by swellingcellulose power in a high concentration of NaOH(e.g., 16% wt/wt) producing the cellulose type II fromtype I (O'Sullivan, 1997; Wood, 1988). Phosphoric acidswollen cellulose (PASC) is most commonly made byswelling dry cellulose powder by adding 85% o-phosphoric acid (Walseth, 1952; Wood, 1988). Highconcentration phosphoric acid treatment could result insome degree of conversion of type II cellulose from typeI (Weimer et al., 1990). The properties of amorphouscellulose made by ball milling, NaOH and H3PO4, varygreatly, depending on cellulose origins, reaction tem-perature and time, as well as reagent types andconcentrations. Therefore, it is nearly impossible tocompare hydrolysis rates on various types of amorphouscellulose from different laboratories or even differentbatches of amorphous cellulose preparations from thesame laboratory. Amorphous cellulose should be kept inhydrated condition; simple air-drying dehydrationresults in a loss of substrate reactivity (Zhang andLynd, 2004b). The loss of substrate reactivity duringdehydration can be minimized through freeze drying ordrying after solvent exchange (Fan et al., 1981; Lee etal., 1980).
Regenerated cellulose is often made by convertinginsoluble cellulose to soluble form using cellulosesolvents, such as nitric acid, sulfuric acid, ammoniacalcupric hydroxide (Cu(NH3)4(OH)2), N,N-dimethylace-tamide (DMAc)/LiCl (Striegel, 1997), and 1-butyl-3-methylimidazolium Cl (Swatloski et al., 2002), followedby restoration to physically insoluble form. The majorcommercial regenerated cellulose is viscose rayon,which is not pure amorphous cellulose due to some re-crystallization. Regenerated amorphous cellulose (RAC)can be made by using cold ∼85% H3PO4 to dissolvecellulose slurry, followed by precipitation with coldwater. RAC is a very good homogeneous substrate forcellulase activity assays (Zhang et al., 2006), and isdifferent from Walseth cellulose, prepared from hetero-geneous swollen cellulose (Walseth, 1952). RAC has aconsistent quality from batch to batch, and is an idealinsoluble nonsubstitutation cellulose substrate for mea-suring extremely low cellulase activity.

460 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
α-Cellulose contains major cellulose and a smallamount of hemicellulose. The commercial Sigma α-cellulose is often used as a reference cellulosic materialto evaluate the hydrolysis ability of total cellulase (Kimet al., 2003). Holocellulose is a solid residue of wood(lignocellulose) after removal of lignin; α-cellulose is asolid residue of holocellulose after removal of majorhemicellulose by alkali extraction (Green, 1963); afterthe neutralization of soluble alkali extract materials fromholocellulose, the insoluble fraction and the solublefraction are β-cellulose and γ-cellulose, respectively(Corbett, 1963a; Corbett, 1963b).
Lignocellulose pretreatment breaks up the recalci-trant structure of lignocellulose so that cellulase canhydrolyze pretreated lignocellulose faster and moreefficiently. Current leading lignocellulose pretreatmenttechnologies, including dilute acid, hot water, flowthrough, ammonia fiber explosion (AFEX), ammoniarecycle percolation, and lime, have been recentlyreviewed elsewhere (Mosier et al., 2005; Wyman etal., 2005a,b). In addition, two other pretreatments–steam explosion and organosolv–have been intensivelyinvestigated (Arato et al., 2005; Bura et al., 2002, 2003;Galbe and Zacchi, 2002; Ohgren et al., 2005; Pan et al.,2005a,b; Pye and Lora, 1991; Sassner et al., 2005 ;Soderstrom et al., 2003; Wingren et al., 2003).
The substrate characteristics (e.g., cellulose acces-sibility, DP, hemicellulose content, and lignin content)of pretreated lignocelluloses vary greatly, stronglydepending on pretreatment methods and severity, andon lignocellulose origins. For example, the goal ofAFEX is to break up the linkages among lignin,hemicellulose, and cellulose, but not to remove anymain component. Therefore, the addition of hemi-cellulase into the cellulase mixture would be importantfor improving overall hydrolysis performance forAFEX-treated feedstock (Teymouri et al., 2005).Dilute acid pretreatment not only to breaks the linkageamong lignin, hemicellulose, and cellulose, but alsoremoves major hemicellulose. Therefore, the additionof hemicellulase is not necessary for an improvementin cellulase mixture performance; while the addition ofnon-hydrolysis proteins (e.g. bovine serum albumin)into the cellulase mixture could reduce the use ofcellulase because of minimization of non-hydrolysisadsorption of cellulase to lignin (Pan et al., 2005b;Wyman CE, personal communication). Organosolvpretreatment significantly removes both hemicelluloseand lignin (Arato et al., 2005; Pan et al., 2005a; Pyeand Lora, 1991). Therefore, neither hemicellulase norother protein blockers need to be added. A novelcellulose-solvent-based lignocellulose fractionation is
under development by our laboratory; the hydrolysisrates of residual cellulose samples containing littlehemicellulose and lignin cannot be improved by theaddition of either hemicellulase or non-hydrolysisprotein (Zhang et al., unpublished). In a word,improvements in the overall performance of cellulasemixture by cocktailing are strongly dependent onresidual lignocellulose properties, and remains in thetrial-and-test stage.
Dyed cellulose is prepared by mixing cellulosewith a variety of dyes, such as Remazol Brilliant Blue(Holtzapple et al., 1984; Wood, 1988), ReactiveOrange (Gusakov et al., 1985), Reactive Blue 19(Yamada et al., 2005), and fluorescent dye 5-(4,6-dichlorotriazinyl) aminofluresceinsm (Helbert et al.,2003). Because of large variations in the surface areasof cellulose and the binding conditions, the quantita-tive relationship between released dye and reducingsugars must be established for each batch of dyedcellulose.
Insoluble cellulose derivatives, such as slightlysubstituted CMC, can be mixed with a variety of dyes,including Cibacron Blue 3GA and Reactive Orange 14to produce insoluble dyed-CMC (Ten et al., 2004).Insoluble cellulose derivatives can also be chemicallysubstituted with trinitrophenyl groups to producechromogenic trinitrophenyl-carboxymethyl cellulose(TNP-CMC) and fluorophoric Fluram cellulose(Huang and Tang, 1976). The TNP-CMC has a 25-fold greater sensitivity for endoglucanase activity thandoes the reducing sugar dinitrosalicyclic acid method,and Fluram cellulose gives another 10-fold increase insensitivity over TNP-CMC (Huang and Tang, 1976).However, an increased substitution of TNP-CMCreduces substrate solubility and impairs cellulase actionalong β-linked chains (Wood and Bhat, 1988). Some-times, TNP-CMC is a useful substrate for enzymesolutions containing reducing agents when the reducingsugar assay cannot be conducted (Shinmyo et al., 1979).For example, the cellulosome from the anaerobicbacterium C. thermocellum requires the presence ofreducing agents such as DTT or cysteine for activity(Johnson et al., 1982a; Morag et al., 1992; Zhang andLynd, 2003a).
4. Quantitative assays
All existing cellulase activity assays can be dividedinto three types: (1) the accumulation of productsafter hydrolysis, (2) the reduction in substratequantity, and (3) the change in the physical propertiesof substrates.
461Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
4.1. Hydrolysis products
The majority of assays involve the accumulation ofhydrolysis products, including reducing sugars, totalsugars, and chromophores. The most common reducingsugar assays include the dinitrosalicyclic acid (DNS)method (Ghose, 1987; Miller, 1959), the Nelson-Somogyi method (Nelson, 1944; Somogyi, 1952), the2,2′-bicinchroninate (BCA) method (Waffenschmidtand Janeicke, 1987; Zhang and Lynd, 2005b), the 4-hydroxybenzoylhydrazine (PAHBAH) method (Lever,1972; Lever et al., 1973), and the ferricyanide methods(Kidby and Davidson, 1973; Park and Johnson, 1949) inTable 3. Total soluble sugars, regardless of their chainlengths, can be measured directly by the phenol-H2SO4
method (Dubois et al., 1956; Zhang and Lynd, 2005b) orthe anthrone-H2SO4 method (Roe, 1955; Viles andSilverman, 1949). Glucose can be measured by anenzymatic glucose kit using coupled hexokinase andglucose-6-phosphate dehydrogenase (Zhang and Lynd,2004a), or HPLC after post-hydrolysis conversion toglucose.
Detection ranges of many sugar assays can bemodified using two strategies: (1) a further dilutionafter the color reaction and (2) varying sugar volume persample prior to the reaction. For example, the DNSmethod was originally designed for 20–600 μg reducingsugar per sample (Miller, 1959), but its detection range
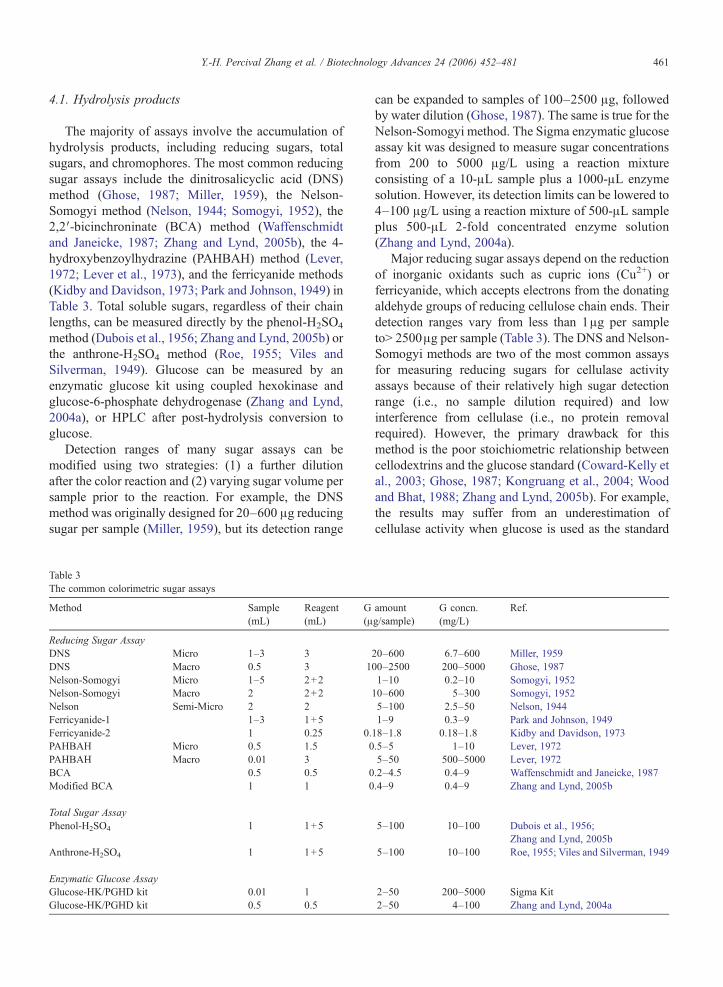
Table 3The common colorimetric sugar assays
Method Sample(mL)
Reagent(mL)
G(μ
Reducing Sugar AssayDNS Micro 1–3 3DNS Macro 0.5 3 1Nelson-Somogyi Micro 1–5 2+2Nelson-Somogyi Macro 2 2+2Nelson Semi-Micro 2 2Ferricyanide-1 1–3 1+5Ferricyanide-2 1 0.25 0.PAHBAH Micro 0.5 1.5 0PAHBAH Macro 0.01 3BCA 0.5 0.5 0Modified BCA 1 1 0
Total Sugar AssayPhenol-H2SO4 1 1+5
Anthrone-H2SO4 1 1+5
Enzymatic Glucose AssayGlucose-HK/PGHD kit 0.01 1Glucose-HK/PGHD kit 0.5 0.5
can be expanded to samples of 100–2500 μg, followedby water dilution (Ghose, 1987). The same is true for theNelson-Somogyi method. The Sigma enzymatic glucoseassay kit was designed to measure sugar concentrationsfrom 200 to 5000 μg/L using a reaction mixtureconsisting of a 10-μL sample plus a 1000-μL enzymesolution. However, its detection limits can be lowered to4–100 μg/L using a reaction mixture of 500-μL sampleplus 500-μL 2-fold concentrated enzyme solution(Zhang and Lynd, 2004a).
Major reducing sugar assays depend on the reductionof inorganic oxidants such as cupric ions (Cu2+) orferricyanide, which accepts electrons from the donatingaldehyde groups of reducing cellulose chain ends. Theirdetection ranges vary from less than 1μg per sampleto> 2500μg per sample (Table 3). The DNS and Nelson-Somogyi methods are two of the most common assaysfor measuring reducing sugars for cellulase activityassays because of their relatively high sugar detectionrange (i.e., no sample dilution required) and lowinterference from cellulase (i.e., no protein removalrequired). However, the primary drawback for thismethod is the poor stoichiometric relationship betweencellodextrins and the glucose standard (Coward-Kelly etal., 2003; Ghose, 1987; Kongruang et al., 2004; Woodand Bhat, 1988; Zhang and Lynd, 2005b). For example,the results may suffer from an underestimation ofcellulase activity when glucose is used as the standard
amountg/sample)
G concn.(mg/L)
Ref.
20–600 6.7–600 Miller, 195900–2500 200–5000 Ghose, 19871–10 0.2–10 Somogyi, 195210–600 5–300 Somogyi, 19525–100 2.5–50 Nelson, 19441–9 0.3–9 Park and Johnson, 194918–1.8 0.18–1.8 Kidby and Davidson, 1973.5–5 1–10 Lever, 19725–50 500–5000 Lever, 1972.2–4.5 0.4–9 Waffenschmidt and Janeicke, 1987.4–9 0.4–9 Zhang and Lynd, 2005b
5–100 10–100 Dubois et al., 1956;Zhang and Lynd, 2005b
5–100 10–100 Roe, 1955; Viles and Silverman, 1949
2–50 200–5000 Sigma Kit2–50 4–100 Zhang and Lynd, 2004a

462 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
and β-glucosidase is not in excess (Breuil and Saddler,1985a,b; Schwarz et al., 1988). The ferricyanide,PAHBAH, and BCA methods, having higher sensitivityto reducing sugar, can detect as little as severalmicrograms per sample, but suffer from non-specificinterference from protein.
Total carbohydrate assays, including the phenol-H2SO4 method and the anthrone-H2SO4 method, offertwo obvious advantages as compared with reducingsugar assays: a strict stoichemetic relationship betweencellodextrins (glucose equivalent) and the glucosestandard, and little or no interference from protein. Butthey are limited for application to pure celluloses,because any carbohydrates and their derivatives canhave strong interference readings. Using an enzymaticglucose assay kit or HPLC can overcome nonspecificreadings from other sugars, but this requires an extrastep—conversion of longer cellodextrins to glucose.
Total loss of substrate can be measured by severalmeans, such as gravimetry and chemical methods. Thesemethods are not as popular as those involving productaccumulation because they involve tedious procedures,such as sample centrifugation or filtration followed bydrying. Gravimetry should be employed with care,because the standard deviation of this method is stronglyassociated with sample weight. For example, twosamples of 1mg and 100mg weighed by an analyticalbalance with accuracy of 0.1mg have 10% and 0.1%standard deviation, respectively. Chemical methods fordetermining substrate loss include the phenol-H2SO4
(Dubois et al., 1956), the anthrone-H2SO4 (Viles andSilverman, 1949), and the K2Cr2O7–H2SO4 methods(Wood, 1988) for residual cellulose, and quantitativesaccharification for different carbohydrate components(Ruiz and Ehrman, 1996).
Measurable physical cellulose properties represent-ing cellulase activity include swollen factor, fiberstrength, structure collapse, turbidity, and viscosity.Earlier assays, involving measurement of the physicalchanges of the residual solid cellulose, are reviewedhere for historical interest. Examples of these assaysinclude the “swelling factor” (measured by alkaliuptake) and the reduction in tensile strength of threadand pulp (Oksanen et al., 2000; Wood, 1975). Typically,the lack of sensitivity limits the use of these assays,except on special occasions (Oksanen et al., 2000; Pereet al., 2001; Wong et al., 2000). For example, Toyama etal. measured total cellulase activity based on the timeneeded to disintegrate a 1×1 cm filter paper square(Wood, 1988). The turbidometric assay measures areduction in the absorbance of particle suspensionduring the hydrolysis process (Enari and Niku-Paavola,
1988; Johnson et al., 1982a,b; Nummi et al., 1981),which monitors the overall hydrolysis rate over a longtime but does not measure well the initial hydrolysis ratefor individual enzymes. Amorphous cellulose is recom-mended for turbidometric assays (Enari and Niku-Paavola, 1988) because crystalline cellulose hydrolysiscould lead to an initial absorbance increase (Zhang,unpublished).
Viscosimetric determinations have been used as anassay for the initial hydrolysis rate for endoglucanasesusing soluble cellulose derivatives (Demeester et al.,1976; Hulme, 1988; Manning, 1981; Miller et al., 1960).Application of this method relies on the assumption thatthe ratio of viscosity-average molecular weight tonumber-average molecular weight should remain con-stant during the period of the assay, which may be trueonly for a short time (Hulme, 1988). This method is alsoexperimentally cumbersome and difficult to automate.
4.2. Cellulase activity assays
The two basic approaches to measuring cellulaseactivity are (1) measuring the individual cellulase(endoglucanases, exoglucanases, and β-glucosidases)activities, and (2) measuring the total cellulase activity.In general, hydrolase enzyme activities are expressed inthe form of the initial hydrolysis rate for the individualenzyme component within a short time, or the end-pointhydrolysis for the total enzyme mixture to achieve afixed hydrolysis degree within a given time. Forcellulase activity assays, there is always a gap betweeninitial cellulase activity assays and final hydrolysismeasurement (Sheehan and Himmel, 1999). To be mostmeaningful, individual cellulase component assaysmust also be based on a reliable estimation of theamount of individual enzyme component present in theassay. This information permits the calculation ofspecific activity, i.e., bonds broken per milligramenzyme per unit time.
4.2.1. EndoglucanasesEndoglucanases cleave intramolecular β-1,4-gluco-
sidic linkages randomly, and their activities are oftenmeasured on a soluble high DP cellulose derivative,such as CMC with the lowest ratio of FRN/Fa (Table 1).The modes of actions of endoglucanases and exoglu-canases differ in that endoglucanases decrease thespecific viscosity of CMC significantly with littlehydrolysis due to intramolecular cleavages, whereasexoglucanases hydrolyze long chains from the ends in aprocessive process (Irwin et al., 1993; Teeri, 1997;Zhang and Lynd, 2004b). Endoglucanase activities can

463Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
be measured based on a reduction in substrate viscosityand/or an increase in reducing ends determined by areducing sugar assay. Because exoglucanases alsoincrease the number of reducing ends, it is stronglyrecommended that endoglucanase activities be mea-sured by both methods (viscosity and reducing ends).Because the carboxymethyl substitutions on CMC makesome glucosidic bonds less susceptible to enzymeaction, a linear relationship between initial hydrolysisrates and serially diluted enzyme solutions requires (1)dilute enzyme preparation, (2) a short incubation period(e.g., 2–4min) or a very low enzyme loading, (3) a lowDS CMC, and (4) a sensitive reducing sugar assay.Many workers agree that the BCA method for reducingsugar assay is superior to the DNS method (Carcia et al.,1993). For example, the modified BCA method, whichis conducted at 75 °C to avoid β-glucosidic bondcleavage during the assay, delivers a strict stoichiometryfor the reducing ends of cellodextrins regardless of sugarchain lengths (Zhang and Lynd, 2005b) and offers amuch higher sensitivity as shown in Table 3 (Zhang andLynd, 2005b).
Soluble oligosaccharides and their chromophore-substituted substrates, such as p-nitrophenyl glucosidesand methylumbelliferyl-β-D-glucosides, are also used tomeasure endoglucanase activities based on the release ofchromophores or the formation of shorter oligosaccha-ride fragments, which are measured by HPLC or TLC(Bhat et al., 1990; Claeyssens and Aerts, 1992; vanTilbeurgh and Claeyssens, 1985; Zverlov et al., 2002a,2002b, 2003, 2005).
Endoglucanase activities can also be easily detectedon agar plates by staining residual polysaccharides(CMC, cellulose) with various dyes because these dyesare adsorbed only by long chains of polysaccharides(Fülöp and Ponyi, 1997; Hagerman et al., 1985; Jang etal., 2003; Jung et al., 1998; Kim et al., 2000; Murashimaet al., 2002a; Piontek et al., 1998; Rescigno et al., 1994;Ten et al., 2004). These methods are semi-quantitative,and are well suited to monitoring large numbers ofsamples. Precision is limited because of the relationshipbetween the cleared zone diameters and the logarithm ofenzyme activities. For example, differences in enzymeactivity levels less than 2-fold are difficult to detect byeye (Sharrock, 1988). Unfortunately, most exoglucanaseactivities are not detected by these methods, since theprocessive action of exoglucanases is blocked bycarboxymethyl substitutions, which prohibits cellulosechain from shortening. The lack of efficient exogluca-nase plate screening method explains some of thedifficulty in detecting exoglucanase genes cloned fromC. thermocellum (Demain et al., 2005).
4.2.2. ExoglucanasesExoglucanases cleave the accessible ends of cellu-
lose molecules to liberate glucose and cellobiose. T.reesei cellobiohydrolase (CBH) I and II act on thereducing and non-reducing cellulose chain ends,respectively (Teeri, 1997; Teeri et al., 1998; Zhangand Lynd, 2004b). Avicel has been used for measuringexoglucanase activity because it has the highest ratio ofFNR/Fa among insoluble cellulosic substrates (Table 1).During chromatographic fractionation of cellulasemixtures, enzymes with little activity on soluble CMC,but showing relatively high activity on avicel, areusually identified as exoglucanases. Unfortunately,amorphous cellulose and soluble cellodextrins aresubstrates for both purified exoglucanases and endoglu-canases. Therefore, unlike endoglucanases and β-glucosidases, there are no substrates specific forexoglucanases within the cellulase mixtures (Sharrock,1988; Wood and Bhat, 1988).
Claeyssens and his coworkers (van Tilbeurgh et al.,1982) found that 4-methylumbelliferyl-β-D-lactosidewas an effective substrate for T. reesei CBH I, yieldinglactose and phenol as reaction products, but it was not asubstrate for T. reesei CBH II (van Tilbeurgh andClaeyssens, 1985) and some endoglucanases (vanTilbeurgh et al., 1982). T. reesei EG I, structurallyhomologous to CBH I, also cleaves 4-methylumbelli-feryl-β-D-lactoside, yet these enzymes can be differen-tiated by adding cellobiose, an inhibitor that stronglysuppresses cellobiohydrolase activity (Claeyssens andAerts, 1992). T. reesei CBH II does not hydrolyze 4-methylumbelliferyl-β-D-aglycones of either glucose orcellobiose units, but does cleave 4-methylumbelliferyl-β-D-glycosides with longer glucose chains (van Til-beurgh et al., 1985).
Deshpande et al. (1984) reported a selective assay forexoglucanases in the presence of endoglucanases and β-glucosidases. This assay is based on the following: (1)exoglucanase specifically hydrolyzes the aglyconicbond of p-nitrophenyl-β-D-cellobioside to yield cello-biose and p-nitrophenol, (2) β-glucosidase activity isinhibited by D-glucono-1,5-δ-lactone (Holtzapple et al.,1990), and (3) the influence of exoglucanase hydrolysisactivities must be quantified in the assay procedure inthe presence of added purified endoglucanases. How-ever, this technique has its own limitations: (1) CBH IIactivity cannot be measured using p-nitrophenyl-β-D-cellobioside, (2) the specific activity of the availablepurified endoglucanases may not be representative of allexisting endoglucanases in the mixture, and (3) theproduct ratio from endoglucanase actions may beinfluenced by the presence of exoglucanases.

464 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
4.2.3. β-D-glucosidasesβ-D-glucosidases hydrolyze soluble cellobiose and
other cellodextrins with a DP up to 6 to produce glucosein the aqueous phase. The hydrolysis rates decreasemarkedly as the substrate DPs increase (Zhang andLynd, 2004b). The term “cellobiase” is often misleadingdue to this key enzyme's broad substrate specificitybeyond a DP of 2. β-D-glucosidases are very amenableto a wide range of simple sensitive assay methods, basedon colored or fluorescent products released from p-nitrophenyl β-D-1,4-glucopyranoside (Deshpande et al.,1984; Strobel and Russell, 1987), β-naphthyl-β-D-glucopyranoside, 6-bromo-2-naphthyl-β-D-glucopyra-noside (Polacheck et al., 1987), and 4-methylumbelli-feryl-β-D-glucopyranoside (Setlow et al., 2004). Also,β-D-glucosidase activities can be measured usingcellobiose, which is not hydrolyzed by endoglucanasesand exoglucanases, and using longer cellodextrins,which are hydrolyzed by endoglucanases and exoglu-canases (Ghose, 1987; Gong et al., 1977; McCarthy etal., 2004; Zhang and Lynd, 2004b).
4.2.4. Total cellulaseThe total cellulase system consists of endoglucanases,
exoglucanases, and β-D-glucosidases, all of whichhydrolyze crystalline cellulose synergically. Total cellu-lase activity assays are always measured using insolublesubstrates, including pure cellulosic substrates such asWhatman No. 1 filter paper, cotton linter, microcrystal-line cellulose, bacterial cellulose, algal cellulose; andcellulose-containing substrates such as dyed cellulose,α-cellulose, and pretreated lignocellulose.
The heterogeneity of insoluble cellulose and thecomplexity of the cellulase system cause formidableproblems in measuring total cellulase activity. Experi-mental results show that the heterogeneous structure ofcellulose (filter paper and bacterial cellulose) gives riseto a rapid decrease in the hydrolysis rate within a shorttime (less than an hour), even when the effects ofcellulase deactivation and product inhibition are takeninto account (Valjamae et al., 1998; Zhang et al., 1999).In an attempt to clarify this situation, a functionallybased model has been developed to demonstrate that thedegree of synergism between endoglucanase andexoglucanase is influenced by substrate characteristics,experimental conditions, and enzyme loading/composi-tion ratio (Zhang and Lynd, in press). This model clearlysuggests the complexity of total cellulase activity assaysand infers that it is nearly impossible to apply the resultsof the total cellulase activity assay measured on onesolid substrate to a different solid substrate. This is oneof the reasons that the U.S. DOE-sponsored cellulase
development projects, conducted by Genencor Interna-tional and Novozymes Biotech, tailored cellulasemixture performance based only on an identicalsample—dilute acid pretreated corn stover substratethat was prepared in the pilot plant of the NationalRenewable Energy Laboratory (Golden, CO) (Knaufand Moniruzzaman, 2004).
The most common total cellulase activity assay is thefilter paper assay (FPA) using Whatman No. 1 filterpaper as the substrate, which was established andpublished by the International Union of Pure andApplied Chemistry (IUPAC) (Ghose, 1987). Thisassay requires a fixed amount (2mg) of glucose releasedfrom a 50-mg sample of filter paper (i.e., 3.6%hydrolysis of the substrate), which ensures that bothamorphous and crystalline fractions of the substrate arehydrolyzed. A series of enzyme dilution solutions isrequired to achieve the fixed degree of hydrolysis. Thestrong points of this assay are (1) it is based on a widelyavailable substrate, (2) it uses a substrate that ismoderately susceptible to cellulases, and (3) it is basedon a simple procedure (the removal of residual substrateis not necessary prior to the addition of the DNSreagent). However, the FPA is reproduced in mostlaboratories with some considerable effort and it haslong been recognized for its complexity and suscepti-bility to operators' errors (Coward-Kelly et al., 2003;Decker et al., 2003). Reliability of results could beinfluenced by (1) the β-D-glucosidase level present inthe cellulase mixture (Breuil and Saddler, 1985a,b;Schwarz et al., 1988; Sharrock, 1988), because the DNSreadings are strongly influenced by the reducing endratio of glucose, cellobiose, and longer cellodextrins(Ghose, 1987; Kongruang et al., 2004; Wood and Bhat,1988; Zhang and Lynd, 2005b); (2) the freshness of theDNS reagent, which is often ignored (Miller, 1959); (3)the DNS reaction conditions, such as boiling severity,heat transfer, and reaction time (Coward-Kelly et al.,2003); (4) the variations in substrate weight based on thearea size (1×6 cm a strip), because this method does notrequire substrate excess (i.e., substrate amounts stronglyinfluence enzyme activity) (Griffin, 1973); and (5) filterpaper cutting methods, because the different paper-cutting methods such as paper punching, razoring, orscissoring could lead to different accessible reducingends of the substrate (Zhang and Lynd, 2005b).
Dyed celluloses are widely used for determiningsugar inhibition for total cellulase because they avoidthe high background interference from added sugars(Gusakov et al., 1985c; Holtzapple et al., 1984; Wood,1988). Fluorescent-dyed cellulose is also used for thesame purpose, and the higher signal per molecule of

465Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
fluorescent dye permits detection of lower cellulaseactivities. Researchers should consider the following:(1) the calibration curve between dye release andreducing sugar accumulation should be established foreach batch of substrate, because dye adsorption dependson cellulosic substrate properties and preparationconditions; (2) the calibration curve works only for asmall hydrolysis conversion range, because dye mole-cules cannot enter into the internal cellulose structure;and (3) the different hydrolysis modes of endogluca-nases and exoglucanases have different dye releasepreferences (Helbert et al., 2003). Using dyed cellulose,Holtzapple et al. (1984) showed that glucose andcellobiose were noncompetitive inhibitors to the T.reesei cellulase. On the contrary, the T. longibrachiatumcellulase was competitively inhibited by cellobiose andglucose (Gusakov et al., 1985c). Some feel that thedifferent inhibition patterns may be attributed to largevariations in characteristics of dyed celluloses (Gruno etal., 2004).
Cotton fiber, microcrystalline cellulose, bacterialcellulose, and algal cellulose are several other commonpure cellulosic substrates. Powder microcrystallinecellulose could become a preferred substrate to replacefilter paper because (1) it can be rapidly dispensedvolumetrically as a slurry and thus permits roboticsmethods; (2) it can be easily pelleted by centrifugation,and the total sugars released are measured more exactlyby the phenol-H2SO4 method than by the DNS assay;(3) it is a more recalcitrant substrate, yielding a morestringent substrate for total cellulase activity than doesfilter paper; and (4) activities measured on microcrys-talline cellulose could more accurately representhydrolysis ability on pretreated lignocellulose, becauseits characteristics are closer to those of pretreatedlignocelluloses, based on cellulose accessibility tocellulase and the degree of polymerization (Zhang andLynd, 2004b). Sigmacell-20, a readily available micro-crystalline cellulose powder, could also be a goodalternative substrate for a total cellulase activity assay,replacing Whatman No. 1 filter paper. Keep that inmind, some of the pretreated lignocellulose still containssignificant amounts of hemicellulose and lignin, whilemicrocrystalline cellulose does not contain hemicellu-lose and lignin.
α-Cellulose and pretreated lignocellulose are oftenused to evaluate the digestibility of commercial cellulaseor of a reconstituted cellulase mixture for a prolongedreaction. The primary difference, as compared tocellulase activity assays using model cellulosic sub-strates, is the time required for assays, which rangesfrom several minutes to hours for model substrates
(initial hydrolysis rate) to several days for pretreatedlignocellulose to obtain the final digestibility (celluloseconversion). Clearly, the presence of hemicellulose andeven lignin results in more complexity. Again, thedesired outcome of the experiment must indicate thesubstrate chosen, especially in the case of total cellulaseperformance.
In conclusion, the measurement of isolated individ-ual cellulase activity is relatively easy, but it is stillchallenging to measure T. reesei CBH I and CBH IIactivities specifically in the presence of endoglucanases.There is no clear relationship between the hydrolysisrates obtained on soluble substrates and those oninsoluble substrates, mainly because of huge differencesin substrate accessibility and DP. For insoluble cellu-lose, it is highly unlikely that any substantial solubili-zation of crystalline or semicrystalline cellulose willproceed linearly with time, due to varying β-glucosidic-bond accessibilities and chain end availability fordifferent regions of fibers. Researchers must stateclearly all parameters of their assay conditions, andresist temptation to compare their results to those ofother researchers using different substrates, assaymethods, etc. For example, the specific activity ofThermobifida fusca YX endoglucanase is reported to beat least ten-fold higher than that of T. reesei endoglu-canase on soluble CMC (Himmel et al., 1993); however,this activity ratio is not maintained if the assays areperformed with insoluble cellulose (Himmel et al.,1999).
5. Cellulase improvement and screening/selection
Two strategies are available for improving theproperties of individual cellulase components: (1)rational design and (2) directed evolution.
5.1. Rational design
Rational design is the earliest approach to proteinengineering, was introduced after the development ofrecombinant DNA methods and site-directed mutagen-esis more than 20years ago, and is still widely used.This strategy requires detailed knowledge of the proteinstructure, of the structural causes of biological catalysisor structure-based molecular modeling, and of theideally structure–function relationship. As shown inFig. 3, the process of rational design involves (1) choiceof a suitable enzyme, (2) identification of the amino acidsites to be changed, based usually on a high resolutioncrystallographic structure, and (3) characterization ofthe mutants. The availability of data on the protein
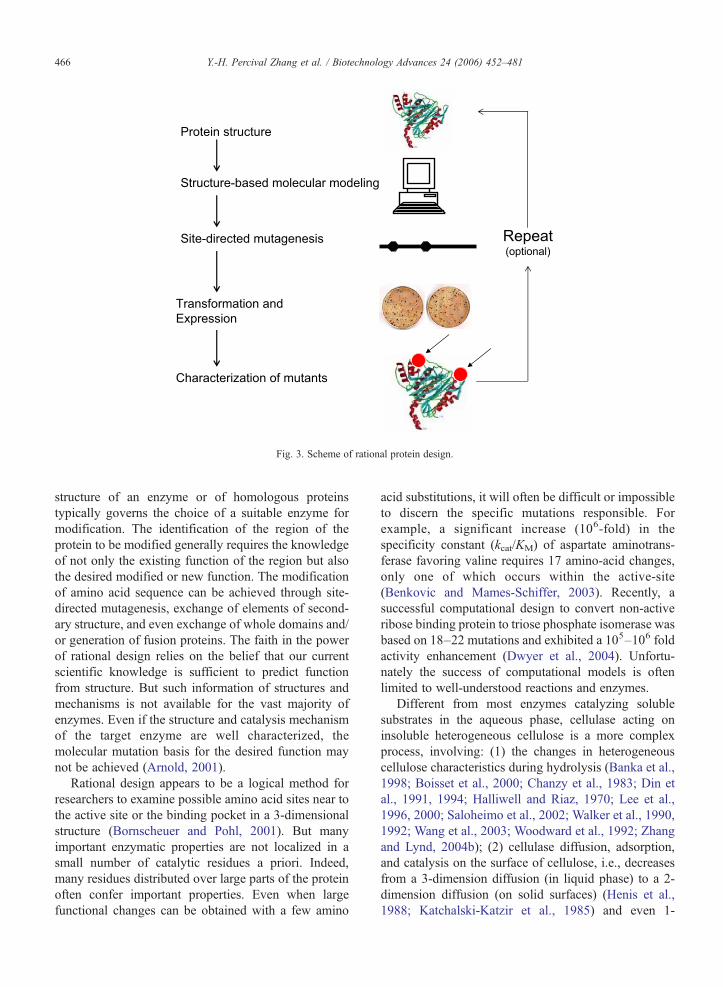
Protein structure
Structure-based molecular modeling
Site-directed mutagenesis
Characterization of mutants
Transformation and
Expression
Repeat(optional)
Fig. 3. Scheme of rational protein design.
466 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
structure of an enzyme or of homologous proteinstypically governs the choice of a suitable enzyme formodification. The identification of the region of theprotein to be modified generally requires the knowledgeof not only the existing function of the region but alsothe desired modified or new function. The modificationof amino acid sequence can be achieved through site-directed mutagenesis, exchange of elements of second-ary structure, and even exchange of whole domains and/or generation of fusion proteins. The faith in the powerof rational design relies on the belief that our currentscientific knowledge is sufficient to predict functionfrom structure. But such information of structures andmechanisms is not available for the vast majority ofenzymes. Even if the structure and catalysis mechanismof the target enzyme are well characterized, themolecular mutation basis for the desired function maynot be achieved (Arnold, 2001).
Rational design appears to be a logical method forresearchers to examine possible amino acid sites near tothe active site or the binding pocket in a 3-dimensionalstructure (Bornscheuer and Pohl, 2001). But manyimportant enzymatic properties are not localized in asmall number of catalytic residues a priori. Indeed,many residues distributed over large parts of the proteinoften confer important properties. Even when largefunctional changes can be obtained with a few amino
acid substitutions, it will often be difficult or impossibleto discern the specific mutations responsible. Forexample, a significant increase (106-fold) in thespecificity constant (kcat/KM) of aspartate aminotrans-ferase favoring valine requires 17 amino-acid changes,only one of which occurs within the active-site(Benkovic and Mames-Schiffer, 2003). Recently, asuccessful computational design to convert non-activeribose binding protein to triose phosphate isomerase wasbased on 18–22 mutations and exhibited a 105–106 foldactivity enhancement (Dwyer et al., 2004). Unfortu-nately the success of computational models is oftenlimited to well-understood reactions and enzymes.
Different from most enzymes catalyzing solublesubstrates in the aqueous phase, cellulase acting oninsoluble heterogeneous cellulose is a more complexprocess, involving: (1) the changes in heterogeneouscellulose characteristics during hydrolysis (Banka et al.,1998; Boisset et al., 2000; Chanzy et al., 1983; Din etal., 1991, 1994; Halliwell and Riaz, 1970; Lee et al.,1996, 2000; Saloheimo et al., 2002; Walker et al., 1990,1992; Wang et al., 2003; Woodward et al., 1992; Zhangand Lynd, 2004b); (2) cellulase diffusion, adsorption,and catalysis on the surface of cellulose, i.e., decreasesfrom a 3-dimension diffusion (in liquid phase) to a 2-dimension diffusion (on solid surfaces) (Henis et al.,1988; Katchalski-Katzir et al., 1985) and even 1-

467Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
dimension processivity along cellulose chains forcellobiohydrolases (Teeri, 1997); (3) the non-productivecellulase binding on the cellulose surface (Beldman etal., 1987; Sheehan and Himmel, 1999); and (4) the yetunexplained dynamic interactions among the cellulose-binding module (CBM), the catalytic domain, and asingle glucan chain end lifted from the cellulose surface(Skopec et al., 2003).
Several excellent reviews summarize numerousstudies using site-directed mutagenesis for investigatingcellulase mechanisms and improving enzyme properties(Schulein, 2000; Wilson, 2004; Wither, 2001). Notsurprisingly, few researchers using site-directed muta-genesis have reported successful examples of signifi-cantly higher activity cellulase mutants on insolublesubstrates (Escovar-Kousen et al., 2004; Sakon et al.,1996; Zhang et al., 2000a,b; Zhang and Wilson, 1997).One clear example, however, is the report by Baker andcoworkers of a 20% improvement in the activity onmicrocrystalline cellulose of a modified endoglucanaseCel5A from Acidothermus cellulolyticus (Baker et al.,2005). The Cel5A endoglucanase, whose high-resolu-tion crystallographic structure has been available(Sakon et al., 1996), was subjected to a series ofmutations designed to alter the chemistry of theproduct-leaving side of the active site cleft. Usingstructural information and following a thesis that endproduct inhibition could be relieved by a substitution ofnon-aromatic residue at site 245, a mutant (Y245G) wasshown to increase KI of cellobiose by 15-fold.However, today there are no general rules for site-directed mutagenesis strategies for improving cellulaseactivity on solid cellulase substrates and it remains in atrial-and-test process.
Fig. 4. Scheme of directed
5.2. Directed evolution
Our still limited knowledge about the characteristicsof insoluble cellulose substrates, the dynamic interac-tions between cellulases and insoluble substrates, andthe complex synergetic and/or competitive relationshipsamong cellulase components, significantly limitsrational design for improving cellulase properties,despite increasing understanding of cellulase structuresand hydrolysis mechanisms, characterization of cellu-lose properties, and cellulase adsorption (Bothwell et al.,1997; Bothwell and Walker, 1995; Bourne and Henris-sat, 2001; Lynd et al., 2002; Wither, 2001; Zhang andLynd, 2004b). In 1999, Michael Himmel (Sheehan andHimmel, 1999) wrote: “…non-informational approachesto protein engineering should be used to complementexisting efforts based on informational or rational designstrategies in order to ensure success of the DOEcellulase improvement program”. One approach tonon-informational mutant identification is irrationaldesign using directed evolution.
The greatest advantage of directed evolution is that itis independent of knowledge of enzyme structure and ofthe interactions between enzyme and substrate. Thegreatest challenge of this method is developing tools tocorrectly evaluate the performance of mutants generatedby recombinant DNA techniques. The success of adirected evolution experiment depends greatly on themethod chosen for finding the best mutant enzyme,often stated as “you get what you screen for” (Hibbert etal., 2005; Schmidt-Dannert, 2001; Schmidt-Dannert andArnold, 1999) (see Fig. 4).
Table 4 lists the published examples of the cellulaseswith properties altered using directed evolution. Four
protein evolution.

Table 4List of cellulases and relevant enzymes whose properties have been changed using directed evolution techniques
Enzyme Alteredproperty
DNA technique Screening/Selection Ref.
Endoglucanase Thermalstability
Family shuffling Facilitated screening-Congo red+CMC agar Murashima et al., 2002b
Endoglucanase Activity DNA shuffling Facilitated screening-Congo red+CMC agar Kim et al., 2000Endoglucanase Alkali pH epPCR Facilitated screening-Congo red+CMC agar Wang et al., 2005Endoglucanase – Family shuffling Facilitated screening-Congo red+CMC agar Catcheside et al., 2003β-D-glucosidase Cold adoption DNA shuffling Random Screening-chromogenic substrate Lebbink et al., 2000β-D -glucosidase Thermal
stabilityepPCR Random Screening-chromogenic substrate Gonzalez-Blasco et al., 2000
β-D -glucosidase Thermalstability
epPCR+Familyshuffling
Random Screening-chromogenic substrate Arrizubieta and Polaina, 2000
β-D -glucosidase Activity epPCR Random Screening-coupled to color reaction McCarthy et al., 2004β-glycosidase Activity Family shuffling Random Screening-chromogenic substrate Kaper et al., 2002Mutated β-glucosidase(glycosynthase)
Activity epPCR Facilitated Screening-fluorogenic substrate Kim et al., 2004
Mutated endoglucanase(glycosynthase)
Activity cassette mutegenesis Chemical complementation Lin et al., 2004
468 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
directed evolution examples have been reported forendoglucanases, all of which are identified by facilitatedscreening on solid plates containing CMC, followed byCongo Red staining (Catcheside et al., 2003; Kim et al.,2000; Murashima et al., 2002a; Wang et al., 2005). Kimet al. (2000) reported that a 5-fold higher specificactivity Bacillus subtilis endoglucanase mutant wasfound by screening cellulase mutants, generated byDNA shuffling and displayed on the surface of E. coliby fusion of the Pseudomonas syringae ice-nucleationprotein. Doi et al. (Murashima et al., 2002b) enhancedthe thermostability of an endoglucanase by seven-foldusing the family shuffling technique based on theparental Clostridium cellulosomal endoglucanases-EngB and EngD. Gao et al. (Wang et al., 2005) foundthat a T. reesei EG III mutant generated using the error-prone PCR technique and expressed in Saccharyomycescerevisiae was found to have an optimal pH of 5.4,corresponding to a basic pH shift of 0.6. Anotherexample identified hybrid mutants using the familyshuffling technique for T. reesei cel12A and Hypocreaschweinitzii cel12A genes (Catcheside et al., 2003).
β-D-glucosidase mutants have been reported to bescreened blindly using 96-microplate wells because oflack of facilitated screening tools (Arrizubieta andPolaina, 2000; Gonzalez-Blasco et al., 2000; Lebbinket al., 2000; McCarthy et al., 2004). Improvements inthe low temperature catalysis (3-fold) for the hyperther-mostable Pyrococcus furiosus β-D-glucosidase CelB(Lebbink et al., 2000) and the thermostabilities andcatalytic efficiencies for the Paenibacillus polymyxaBgblA and BglA were obtained using the chromogenicsubstrate, p-nitrophenyl-β-D-glucopyranoside (Arrizu-
bieta and Polaina, 2000; Gonzalez-Blasco et al., 2000).The hydrolysis rate of the Thermotoga neapolitana 1,4-β-D-glucan β-glucohydrolase (GghA) (EC 3.2.1.74)mutant is increased by 31% after error-prone PCRmutagenesis, in which blind screening was based onglucose released from a non-chromogenic substrate(cellobiose) and measured by the coupled reactions ofthermostable glucokinase and glucose-6-phosphatedehydrogenase (McCarthy et al., 2004). In anotherrecent example, after DNA family shuffling, a β-glycosidase mutant was found to display lactosehydrolysis rates 3.5-fold and 8.6-fold higher than theparental P. furiosus CelB and Sulfolobus solfataricusLacS, respectively, where glucose released from lactosewas measured using a coupled glucose oxidase andphenol 4-aminophenazone peroxidase reaction (Kaperet al., 2002).
In some cases, glycosyl hydrolases, e.g., Agrobacter-ium sp. β-D-glucosidase, can be converted to glyco-synthases by site-directed mutagenesis (Mackenzie etal., 1998). There is no intrinsic way to screen or select forglycosynthase activities today. The specific activity ofglycosynthase from Agrobacterium sp. β-D-glucosidasewas improved (Kim et al., 2004) using a novel coupled-enzyme assay and screening on solid plates becauseanother endoglucanase releases fluorophores from thefluorogenic product synthesized by glycosynthase(Mayer et al., 2001). Another selection method for aglycosynthase mutant library is the chemical comple-mentation method (Lin et al., 2004), based on theprinciple that the glycosynthase activity is linked to thetranscription of a LEU2 reporter gene, resulting in cellgrowth dependant on glycosynthase activity. A 5-fold

469Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
higher activity of glycosynthase is obtained using thisapproach (Lin et al., 2004).
Although a number of successful examples usingdirected evolution for desired cellulases have beenpublished, the largest limitation of all current selectionand screening methods is based on soluble substrates.It is still very challenging to design a method toscreen or select cellulase mutants using solid cellulosicsubstrates.
5.3. Screening
The screening strategy is a critical step for finding thedesired mutants from a large mutant library. Screeningcan be divided into two categories: (1) facilitatedscreening, which distinguishes mutants on the basis ofdistinct phenotypes, such as chromospheres released orhalos formed, and (2) random screening, which picksmutants blindly (Taylor et al., 2001).
A typical facilitated screening, carried out on solidagar, relies on product solubilization followed by anenzymatic reaction that gives rise to a zone of identity,such as chromophores released from chromogenicsubstrates. The assays may be coupled to a secondenzyme whose product can in turn be easily monitored,as demonstrated by a successful coupling for cyto-chrome P450 to horseradish peroxidase (Joo et al.,1999a,b). With the help of microscopic plate images, itis feasible to screen a much larger number of clones onsolid plates (e.g., several hundreds per cm2) (Delagraveet al., 2001; Joo et al., 1999b; Youvan et al., 1995).Recently, an ultra-high throughput facilitated screeningmethod, based on solid microbeads, has been developedin which single cells containing mutant genes areimmobilized on solid beads. After a chromogenicsubstrate is applied, stronger colored beads containingdesired mutants are identified under the microscope(Freeman et al., 2004). Another facilitated screeningmethod, conducted in the liquid phase, applies a flowcytometer for detecting chromospheres released fromchromogenic substrates, which are catalyzed by the cell-displayed enzyme. Numerous reviews pertaining to cellsurface displayed enzyme library screening by flowcytometers are available elsewhere (Aharoni et al.,2005; Becker et al., 2004; Cohen et al., 2001; Goddardand Reymond, 2004; Lin and Cornish, 2002; Wahlerand Reymond, 2001; Wittrup, 2001).
Endoglucanase activities are detected easily byexamination of “halos” on solid agar plates usingCMC as the substrate, followed by Congo Red stainingand washing. Higher hydrolysis rates of mutants usuallyresult in larger halos (in Section 4.2.1). It is not
surprising that all reported endoglucanase examplesusing directed evolution have been screened using theCMC/Congro Red method (in Section 5.2). It may beoperative to screen exoglucanase mutants on solid platesusing soluble chromogenic substrates, such as nitrophe-nol-cellobioside. However, it is worth noting that thebest screening methods for endoglucanases and exoglu-canases, capable of hydrolyzing insoluble cellulose,must be implemented on insoluble cellulose rather thanon soluble cellulose derivatives.
Random screening is another choice, if facilitatedscreening is not available. It is often implemented using96-well microtiter plates, although some researchers aremoving towards 384-well and higher density plateformats with the help of accurate, low-volume dispens-ing instruments (Sundberg, 2000). For example, Diversahas developed an ultra-throughput screening platform,the Gigamatrix, having 400,000 wells containing only50 nL of liquid substrate per well (Wolfson, 2005). Butthe reformatting of 96-well plates into higher densityrequires high assay sensitivity and high evaporationcontrol. Additional product measurement can beachieved using HPLC, mass spectrometry, capillaryelectrophoresis, or IR-thermography (Wahler and Rey-mond, 2001).
A number of improved β-glycosidase mutants afterrandom mutagenesis are found using 96-well micro-plates, as reported in Section 5.2. Recently, in order tomeasure total cellulase activity, the FPA has beenminiaturized from a 1.5-ml enzyme solution to 60 μL,which is implemented in a 96-microplate well (Xiao etal., 2004). Water evaporation from samples is preventedusing a PCR thermocycler having a built-in 105 °C hotlid. Also, Decker and coworkers (2003) have developeda high throughput cellulase assay system using 96-microplates equipped in a Cyberlabs C400 robotics deckwith the substrates such as Whatman No. 1 filter paperdisks (0.25in. diameter), Solka-Floc, SigmaCell-20,Avicel PH101 (FMC, Philadelphia, PA), and cottonlinters (Fluka/Sigma Aldrich). This custom system has amaximum output of 84 samples per day and producesvalues that correlate to the traditional FPA. However, noapplication of these systems to the screening of higheractivity cellulases has been reported. Considering theinherent limitations of the FPA (see Section 4.2.4), thisautomated approach could be benefited by replacing theDNS method with the phenol-H2SO4 method becausethe latter (1) has a higher sugar sensitivity (Table 3), (2)is independent of oxygen presence (unlike the DNSreagent) (Miller, 1959), especially for miniaturizationthat has a very high surface/volume ratio, (3) yields astrict stoichiometric relationship between color

470 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
formation and total soluble sugars released, and (4) isthus independent of β-D-glucosidase levels. Differentfrom FPA, the recommended method requires centrifu-gation for soluble sugars and solid cellulose residueprior to the phenol-sulfuric acid assay.
5.4. Selection
Selection is always preferred over screening becauseit has several-order-of-magnitude higher efficiency thanscreening (Griffithsa et al., 2004; Olsen et al., 2000;Otten and Quax, 2005). However, selection requires aphenotypic functional link between the target gene andits encoding product that confers selective advantage toits producer. This method is often implemented based onthe principles of resistance to cytotoxic agents (e.g.,antibiotics) (Stemmer, 1994a,b) or of complementationof auxotroph (Griffithsa et al., 2004; Jurgens et al.,2000; Smiley and Benkovic, 1994). Today, selection onsolid media in petri dishes (200–300 colonies per dish)is commonly used because a large number of mutantscan be identified conveniently by visual inspection ofgrowth or zones around the colonies as a consequence ofa diffusing product.
Recently, more and more attention has been paid tothe traditional liquid selection (or enrichment) technique(Sauer et al., 2000). An exceedingly higher cellconcentration of ∼1012 individual cells per liter and alonger cultivation time (generations) allow continuousculture to become a powerful selection system for theultra-large size of the mutant library even when selectiveadvantages are very small.
Experimental selection strategy is associated with (1)the selection ratio, (2) the location of the targetedenzyme, and (3) the solubility of the substrate for the
Enzyme
Intracellular Extr
Sub
stra
te S
olub
ility
Sol
uble
Inso
lubl
e
High RS: Yes. Solid Media
Low RS: Yes. Chemostat
e.g. kan gene in the presence of kanamycin
e.g. β-galactosidase gene on lactose
High RS: No.
Low RS: No.
Hig
e.gam
L
H
e.gin t
Lo
e.
No
Fig. 5. Selection strategies depending on selection ratio
targeted enzyme. Fig. 5 presents possible selectionstrategies and real examples.
The selection ratio (RS) is defined as
RS ¼ lmutant
lparent
in which μmutant and μparent are the specific growth ratesof the beneficial mutant strain and the parental control,respectively, in terms of time (h-1). RS values could varyfrom 1 to infinity. A value of “one” implies no selectiveadvantage for the mutants in growth rates, and a value of“infinity” implies an essentially infinite selectiveadvantage, e.g., no growth for wild strains in thepresence of antibiotic. When RS>1.2, a selectionexperiment can be implemented preferentially on solidmedia because of easy operation. When RS<1.2, liquidcontinuous culture could be the only choice, because alimited number of generations (e.g., ∼25) cannotgenerate the significant difference detected, but thislimitation can be overcome by employing the continu-ous culture chemostat.
The location of the targeted enzyme is anotherimportant concern for selection. Selection for intracel-lular enzymes is relatively easy, because such benefit isconfined to the same organism that produces the protein.A typical example is kanamycin selection, in which onlybacteria containing the aminoglycoside phosphototrans-ferase (kan) gene expressed intracellularly can grow inthe presence of kanamycin. Other examples of carbo-hydrate-metabolizing enzymes are intracellular β-ga-lactosidase (Horiuchi et al., 1962) and cell-membrane-associated lactose permease (Tsen et al., 1996), whenlactose is the controlling growth carbon source. ASaccharomyces cerevisiae mutant containing severalrecombinant key intracellular genes for xylose
Location
acellular - tethered Extracellular - secreted
h RS: Yes. Solid Media.
. β-lactamase gene in the presence of picillin
ow RS: Yes. Microcolonization
igh RS: Yes. Solid Media
. Isolation of cellulolytic microorganisms he presence of cellulose
w RS: Possible. Low RS: No.
g. subiltis gene on BSA
example
(RS), enzyme location, and substrate solubility.

471Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
catabolism has been selected to utilize xylose efficientlyunder anaerobic conditions after 460 generations(Sonderegger and Sauer, 2003). When a key enzymeis secreted extracellularly, the benefit from this enzymeis shared with others, resulting in cross feeding. Atypical example is the growth of plasmid-deficient stainsin liquid fermentations when fermentations are pro-longed or the formation of micro-colonies on petri platesbecause the secreted β-lactamase degrades ampicillinwith time. A novel method, called microcolonization,has been invented to solve cross feeding challenges for asecreted enzyme (Naki et al., 1998). To employ thismethod, cells are compartmented into hollow fibers andeach single colony grows in its own segment. Whensoluble substrate is fed to each single cell, diffusion ofthe secreted enzymes and hydrolytic products (crossfeeding) is limited spatially, resulting in effectiveselective pressure. A 5-fold greater protease-producingmutant was obtained using bovine serum albumin as asole nitrogen source and by this selection technique(Naki et al., 1998).
The examples pertaining to the influence of enzymelocation on evolution of specific cellulase activity couldexist in nature. The genes encoding the secretory T.reesei cellobiohydrolases are thought to have noselection pressure to increase their catalytic efficiency(Divne et al., 1994; Konstantinidis et al., 1993; Sinnott,1998). The fact that the cell surface associated cellulasefrom C. thermocellum (∼2.4 IU/mg, measured atsubstrate-excess conditions) (Zhang and Lynd, 2003a)has several-fold higher specific activity than thesecretory T. reesei cellulase (∼0.6 IU/mg, measured atsubstrate-excess) (Lynd et al., 2002) on crystallinecellulose could be attributed to the enzyme location.(Note, here we do not use the old data that the specifichydrolysis rate of C. thermocellum cellulase on crystal-line cellulose (substrate-excess) was 50-fold higher thanthe T. reesei cellulase hydrolysis (substrate-limited)(Johnson et al., 1982a,b).
Another comparative example (cellulase vs. amylase)also suggests the influence of different enzymes'locations on their evolution rates. The different mainbiological roles (cellulose as a major component of theplant cell wall and starch as an energy reserve)determine that the majority of cellulases and amylasesare located extracellularly and intracellularly, respec-tively. Therefore, it is not surprising that amylases couldhave experienced higher evolution rates for achievinghigher catalytic efficiency. The difference in catalyticefficiencies for two enzymes on soluble substrates isverified by the results from the Sinot laboratory(Sinnott, 1998). Furthermore, if amylases already have
a little higher specific activity than cellulases, lessexpression of amylase for utilizing substrates couldresult in a higher evolution rate than cellulases becauseof different expression levels (Drummond et al., 2005).Therefore, the different enzyme locations and theresulting expression levels could result in differentobserved catalysis efficiencies for cellulases andamylases over a long evolution.
Substrate solubility of the targeted enzyme is alsoimportant for selection, but often is ignored. Becauseinsoluble substrates cannot diffuse across cell mem-branes, selection requires that a microorganism produceextracellular enzymes that hydrolyze insoluble sub-strates to produce utilized nutrients. On solid plates, anexample of this selection is the isolation of cellulolyticmicroorganisms that produce extracellular cellulasesfrom non-cellulolytic microorganisms using cellulose asthe sole carbon source. In liquid medium, Lelieveld(1982) proposed the use of continuous culture forselection of beneficial hydrolase mutants on solidsubstrates because there is a diffusion layer of hydro-lyzed products between solid substrates and the bulkphase, causing the difference in growth rates. Anothertheoretical model, proposed by Lynd and coworkers (Fanet al., 2005), further predicted that the tethered hydrolaseon the cell surface was a prerequisite for selection on aninsoluble substrate. If a hydrolase is secreted into theliquid phase, the cross feeding effect leads to little or noselective pressure (Fan et al., 2005). It must be noted thatthe above selection analysis is based on the positive orneutral gene control assumption that be right for majorcarbohydrate hydrolases. In nature, cellulase regulationis more complicated, including negative control andglobal catabolite regulation (Dror et al., 2003a,b, 2005;Han et al., 2004, 2003; Ilmen et al., 1997; Lin andWilson, 1988; Mach and Zeilinger, 2003; Spiridonovand Wilson, 1998, 2000; Stevenson and Weimer, 2005;Zhang and Lynd, 2005a).
Herein we hypothesize that several key requirementsfor effective cellulase selection experiments are (1) thecellulase gene is expressed extracellularly in a non-cellulolysis host, (2) the cellulase is tethered on the cellsurface, and (3) the insoluble cellulosic substrate is usedas the limiting carbon source. Difficulty may be posedby the inability of such cell-tethered enzymes toproperly access the essential cellulose surface, especial-ly in real plant cell wall (microfibril) fractions.However, aggressive particle size reduction and pre-treatment may permit adequate selection. Other chal-lenges lie in heterologous expression of cellulases inhosts suitable for selection. For example, considerableefforts have been made to express heterologous

472 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
cellulase genes in non-hydrolytic microorganisms,including yeasts, Saccharomyces cereisiae (Fujita etal., 2004, 2002; Hong et al., 2003a,b; Ito et al., 2004;Murai et al., 1997, 1998; Penttila et al., 1988; vanRensburg et al., 1994, 1996; 1998), and Pichia pastoris(Boer et al., 2000; Godbole et al., 1999); and bacteria, E.coli (Francisco et al., 1993; Zhou et al., 1999), andKlebsiella oxytoca (Wood and Ingram, 1992; Zhou etal., 2001; Zhou and Ingram, 1999, 2001). Only the non-cellulolytic microorganism, K. oxytoca SZ21, can growon insoluble amorphous cellulose after transformed withheterologous cellulase genes (Zhou and Ingram, 2001).Insufficient cellulase expression or reduced activity ofrecombinant cellulases are the two largest obstacles tothe selection approach for obtaining higher activitycellulase mutants (Boer et al., 2000; Godbole et al.,1999; Lynd et al., 2005).
6. Summary
Effective cellulose hydrolysis requires several differ-ent kinds of cellulases working synergically. Althoughnumerous soluble substrates have been developed formeasuring endoglucanase and β-D-glucosidase activi-ties, specific soluble substrates for exoglucanase activityassays lack. It is emphasized that hydrolysis data fromsoluble substrates cannot yield useful information onhydrolysis of insoluble substrates. The heterogeneity ofinsoluble celluloses, the complex dynamic interactionsbetween insoluble substrates and cellulase components,and the complexity of cellulase components result informidable problems in extrapolating the activitymeasured on one solid substrate to other solid substrates,especially those with biorefinery impact. This point iskey to the eventual improvement of cellulases for theconversion of pretreated plant cell walls in energy cropsand agricultural residues. Actual screening and selectioneffort must be based on physically and chemicallyrelevant industrial substrates. This objective is clearlychallenging at this time.
The past success of rational design campaigns forimproving properties of enzymes have been notable, butthe results are costly and come far too slowly (Arnold,2001). Some feel it is still more of an art than a scienceand is at a heuristic stage (Arnold, 2001; Arnold et al.,2001; Gustafsson et al., 2003). In contrast, directedevolution is sufficiently robust that improved biocata-lysts can be obtained independently of knowledge of theprotein structure and the interaction between enzymesand substrate. Therefore, we propose that the employ-ment of continuous culture with insoluble cellulosicsubstrate as the sole carbon source could be a powerful
tool to select higher activity cellulase mutants that aredisplayed on the cell surface.
Acknowledgments
This work was made possible in part by support toYHPZ from the Biological Systems EngineeringDepartment of Virginia Polytechnic Institute and StateUniversity. Funding from the U.S. Department ofEnergy Office of the Biomass Program (to MEH), andUT-Battelle (to JRM) also supported this work.
References
Aharoni A, Griffiths AD, Tawfik DS. High-throughput screens andselections of enzyme-encoding genes. Curr Opin Chem Biol2005;9:210–6.
Angenent LT, Karim K, Al-Dahhan MH, Wrenn BA, Domiguez-Espinosa R. Production of bioenergy and biochemicals fromindustrial and agricultural wastewater. Trends Biotechnol 2004;22:477–85.
Arato C, Pye EK, Gjennestad G. The lignol approach to biorefining ofwoody biomass to produce ethanol and chemicals. Appl BiochemBiotechnol 2005;121/124:871–82.
Arnold FH. Combinatorial and computational challenges for biocata-lyst design. Nature 2001;409:253–7.
Arnold FH, Wintrode PL, Miyazaki K, Gershenson A. How enzymesadapt: lessons from directed evolution. Trends Biochem Sci2001;26:100–6.
Arrizubieta MJ, Polaina J. Increased thermal resistance and modifi-cation of the catalytic properties of a beta-glucosidase by randommutagenesis and in vitro recombination. J Biol Chem 2000;275:28843–8.
Atalla RH, Vanderhart DL. Native cellulose: a composite of twodistinct crystalline forms. Science 1984;223:283–5.
Baker JO, Ehrman CI, Adney WS, Thomas SR, Himmel ME.Hydrolysis of cellulose using ternary mixtures of purifiedcellulases. Appl Biochem Biotechnol 1998;70/72:395–403.
Baker JO, McCarley JR, Lovett R, Yu CH, Adney WS, Rignall TR,et al. Catalytically enhanced endocellulase Cel5A from Acid-othermus cellulolyticus. Appl Biochem Biotechnol 2005;121–124:129–48.
Banka RR, Mishra S, Ghose TK. Fibril formation from cellulose by anovel protein from Trichoderma reesei: a non-hydrolytic cellulo-lytic component? World J Microbiol Biotechnol 1998;14:551–8.
Barr BK, Holewinski RJ. 4-Methyl-7-thioumbelliferyl-beta-D-cello-bioside: a fluorescent, nonhydrolyzable substrate analogue forcellulases. Biochemistry 2002;41:4447–52.
Bayer EA, Morag E, Lamed R. The cellulosome—a treasure-trove forbiotechnology. Trends Biotechnol 1994;12:379–86.
Bayer EA, Shimon LJ, Shoham Y, Lamed R. Cellulosomes—structureand ultrastructure. J Struct Biol 1998;124:221–34.
Bayer EA, Belaich JP, Shoham Y, Lamed R. The cellulosomes:multienzyme machines for degradation of plant cell wallpolysaccharides. Annu Rev Microbiol 2004;58:521–54.
Becker S, Schmoldt HU, Adams TM, Wilhelm S, Kolmar H. Ultra-high-throughput screening based on cell-surface display andfluorescence-activated cell sorting for the identification of novelbiocatalysts. Curr Opin Biotechnol 2004;15:323–9.

473Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Beguin P, Alzari PM. The cellulosome of Clostridium thermocellum.Biochem Soc Trans 1998;26:178–85.
Beldman G, Voragen AGJ, Rombouts FM, Searle-van Leeuwen MF,Pilnik W. Adsorption and kinetic behavior of purified endogluca-nases and exoglucanases from Trichoderma viride. BiotechnolBioeng 1987;30:251–7.
Benkovic SJ, Mames-Schiffer S. A perspective on enzyme catalysis.Science 2003;301:1196–202.
Bera-Maillet C, Arthaud L, Abad P, Rosso M-N. Biochemicalcharacterization of MI-ENG1, a family 5 endoglucanase secretedby the root-knot nematode Meloidogyne incognita. Eur J Biochem2000;267(11):3255–63.
Berner RA. The long-term carbon cycle, fossil fuels and atmosphericcomposition. Nature 2003;426:323–6.
Bhat KM, Hay AJ, Claeyssens M, Wood TM. Study of the mode ofaction and site-specificity of the endo-(1-4)-beta-D-glucanases ofthe fungus Penicillium pinophilum with normal, 1-3H-labelled,reduced and chromogenic cello-oligosaccharides. Biochem J1990;266:371–8.
Boer H, Teeri TT, Koivula A. Characterization of Trichoderma reeseicellobiohydrolase Cel7A secreted from Pichia pastoris using twodifferent promoters. Biotechnol Bioeng 2000;69:486–94.
Boisset C, Chanzy H, Henrissat B, Lamed RL, Shoham Y, Bayer EA.Digestion of crystalline cellulose substrates by the Clostridiumthermocellum cellulosome: structural and morphological aspects.Biochem J 1999;340:829–35.
Boisset C, Fraschini C, Schulein M, Henrissat B, Chanzy H. Imagingthe enzymatic digestion of bacterial cellulose ribbons reveals theendo character of the cellobiohydrolase Cel6A from Humicolainsolens and its mode of synergy with cellobiohydrolase Cel7A.Appl Environ Microbiol 2000;66:1444–52.
Boisset C, Petrequin C, Chanzy H, Henrissat B, Schulein M.Optimized mixtures of recombinant Humicola insolens cellulasesfor biodegradation of crystalline cellulose. Biotechnol Bioeng2001;72:339–45.
Bornscheuer UT, Pohl M. Improved biocatalysts by directedevolution and rational protein design. Curr Opin Chem Biol2001;5:137–43.
Bothwell MK, Walker LP. Evalution of parameter estimation methodsfor estimating cellulase binding constants. Bioresour Technol1995;53:21–9.
Bothwell MK, Daughhetee SD, Chaua GY, Wilson DB, Walker LP.Binding capacities for thermomonospora fusca E3, E4 and E5, theE3 binding domain, and Trichoderm reesei CBHI on Avicel andBacterial Microcrystalline cellulose. Bioresour Technol 1997;60:69–178.
Bourne Y, Henrissat B. Glycoside hydrolases and glycosyltransferases:families and functional modules. Curr Opin Struct Biol 2001;11:593–600.
Breuil C, Saddler JN. Comparison of the 3,5-dinitrosalicylic acid andNelson-Somogyi methods of assaying for reducing sugars anddetermining cellulase activity. Enzyme Microb Technol 1985a;7:327–32.
Breuil C, Saddler JN. A comparison of various cellulase assayprocedures for measuring the cellulolytic activity of Trichodermaharzianum E58. Biochem Soc Trans 1985b;13:449–50.
Bura R, Mansfield SD, Saddler JN, Bothast RJ. SO2-catalyzed steamexplosion of corn fiber for ethanol production. Appl BiochemBiotechnol 2002;98/100:59–72.
Bura R, Bothast RJ, Mansfield SD, Saddler JN. Optimization of SO2-catalyzed steam pretreatment of corn fiber for ethanol production.Appl Biochem Biotechnol 2003;105/108:319–35.
Caldeira K, Jain AK, Hoffert MI. Climate sensitivity uncertainty andthe need for energy without CO2 emission. Science 2003;299:2052–4.
Carcia E, Johnston D, Whitaker JR, Shoemaker SP. Assessment ofendo-1,4-beta-D-glucanase activity by a rapid colorimetric assayusing disodium 2,2'-bicinchoninate. J Food Biochem 1993;17:135–45.
Catcheside DE, Rasmussen JP, Yeadon PJ, Bowring FJ, Cambareri EB,et al. Diversification of exogenous genes in vivo in Neurospora.Appl Microbiol Biotechnol 2003;62:544–9.
Chang MM, Chou TYC, Tsao GT. Structure, pretreatment andhydrolysis of cellulose. Adv Biochem Eng 1981;20:15–42.
Chanzy H, Henrissat B, Vuong R, Schulein M. The action of 1,4-b-D-glucan cellobio-hydrolase on Valonia cellulose microcrystals. Anelectron microscopic study. FEBS Lett 1983;153:113–7.
Cherry JR, Fidantsef AL. Directed evolution of industrial enzymes: anupdate. Curr Opin Biotechnol 2003;14:438–43.
ChiricoWJ, Brown RDJ. Separation of [1-3H]cellooligosaccharides bythin-layer chromatography: assay for cellulolytic enzymes. AnalBiochem 1985;150:264–72.
Chirico WJ, Brown RDJ. Beta-glucosidase from Trichoderma reesei:substrate-binding region and mode of action on [3H]cello-oligosaccharides. Eur J Biochem 1987;165:343–51.
Claeyssens M, Aerts G. Characterisation of cellulolytic activities incommercial Trichoderma reesei preparations: an approachusing small, chromogenic substrates. Bioresour Technol 1992;39:143–6.
Cohen N, Abramov S, Dror Y, Freeman A. In vitro enzyme evolution:the screening challenge of isolating the one in a million. TrendsBiotechnol 2001;19:507–10.
Converse AO. Substrate factors limiting enzymatic hydrolysis. In:Saddler JN, editor. Bioconversion of forest and agricultural plantresidues. CAB International; 1993. p. 93–106.
Corbett WM. Determination of the alpha-cellulose content of cottonand wood cellulose. Methods Carbohydr Chem 1963a;3:27–8.
Corbett WM. Purification of cotton cellulose. Methods CarbohydrChem 1963b;3:3–4.
Coward-Kelly G, Aiello-Mazzari C, Kim S, Granda C, Holtzapple M.Suggested improvements to the standard filter paper assay used tomeasure cellulase activity. Biotechnol Bioeng 2003;82:745–9.
Cox PM, Betts RA, Jones CD, Spall SA, Totterdell IJ. Acceleration ofglobal warming due to carbon-cycle feedbacks in a coupledclimate model. Nature 2000;408:184–7.
Das H, Singh SK. Useful byproducts from cellulosic wastes ofagriculture and food industry-a critical appraisal. Crit Rev FoodSci Nutr 2004;44:77–89.
Decker SR, Adney WS, Jennings E, Vinzant TB, Himmel ME.Automated filter paper assay for determination of cellulase activity.Appl Biochem Biotechnol 2003;105-108:689–703.
Delagrave S, Murphy DJ, Pruss JL, Maffia AMr, Bylina EJ, et al.Application of a very high-throughput digital imaging screen toevolve the enzyme galactose oxidase. Protein Eng 2001;4:261–7.
Demain AL. Pickles, pectin, and penicillin. Annu Rev Microbiol2004;58:1–42.
Demain AL, NewcombM, Wu JHD. Cellulase, clostridia, and ethanol.Microbiol Mol Biol Rev 2005;69:124–54.
Demeester J, Cooreman W, Bracke M, Lauwers A. Determination ofendoglucanase activity by viscosimetry. Biochem Soc Trans1976;5:1115–7.
Deshpande MV, Eriksson KE, Pettersson LG. An assay for selectivedetermination of exo-1,4,-beta-glucanases in a mixture ofcellulolytic enzymes. Anal Biochem 1984;138:481–7.

474 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Din N, Gilkes NR, Tekant B, Miller RC, Anthony JR, Warren RAJ,et al. Non-hydrolytic disruption of cellulose fibres by thebinding domain of a bacterial cellulase. Bio/Technology 1991;9:1096–9.
Din N, Damude HG, Gilkes NR, Miller RC, Warren RAJ, Kilburn DG.C1-Cx revisited: intramolecular synergism in a cellulase. Proc NatlAcad Sci U S A 1994;91:11383–7.
Divne C, Stahlberg J, Reinikainen T, Ruohonen L, Pettersson G,Knowles JKC, et al. The three dimensional structure ofcellobiohydrolase I from Trichoderma reesei. Science 1994;265:524–8.
Doi RH, Kosugi A. Cellulosomes: plant-cell-wall-degrading enzymecomplexes. Nat Rev Microbiol 2004;2:541–51.
Doi RH, Tamaru Y. The Clostridium cellulovorans cellulosome: anenzyme complex with plant cell wall degrading activity. Chem Rec2001;1:24–32.
Doi RH, Park JS, Liu CC, Malburg LM, Tamaru Y, Ichiishi A, et al.Cellulosome and noncellulosomal cellulases of Clostridiumcellulovorans. Extremophiles 1998;2:53–60.
Dong XM, Revol J-F, Gray DG. Effect of microcrystallite preparationconditions on the formation of colloid crystals of cellulose.Cellulose 1998;5:19–32.
Dror TW, Morag E, Rolider A, Bayer EA, Lamed R, Shoham Y.Regulation of the cellulosomal CelS (cel48A) gene of Clostridiumthermocellum is growth rate dependent. J Bacteriol 2003a;185:3042–8.
Dror TW, Rolider A, Bayer EA, Lamed RL, Shoham Y. Regulation ofexpression of scaffoldin-related genes in Clostridium thermocel-lum. J Bacteriol 2003b;185:5109–16.
Dror TW, Rolider A, Bayer EA, Lamed R, Shoham Y. Regulation ofmajor cellulosomal endoglucanases of Clostridium thermocellumdiffers from that of a prominent cellulosomal xylanase. J Bacteriol2005;187:2261–6.
Drummond DA, Bloom JD, Adami C, Wilke CO, Arnold FH. Whyhighly expressed proteins evolve slowly. Proc Natl Acad Sci U S A2005;102:14338–43.
Dubois B, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colormetricmethod for determination of sugars and relative substances. AnalChem 1956;28:350–6.
Dwyer MA, Looger LL, Hellinga HW. Computational design of abiologically active enzyme. Science 2004;304:1967–71.
Enari TM, Niku-Paavola ML. Nephelometric and turbidometric assayfor cellulase. Methods Enzymol 1988;160:117–26.
Eriksson KE, Pettersson B. A zymogram technique for the detection ofcarbohydrases. Anal Biochem 1973;56:618–20.
Escovar-Kousen JM, Wilson DB, Irwin D. Integration of computermodeling and initial studies of site-directed mutagenesis toimprove cellulase activity on Cel9A from Thermobifida fusca.Appl Biochem Biotechnol 2004;113-116:287–97.
Esterbauer H, Steiner W, Labudova I, Hermann A, Hayn M.Production of Trichoderma cellulase in laboratory and pilotscale. Bioresour Technol 1991;36:51–65.
Falkowski P, Scholes RJ, Boyle E, Canadell J, Canfield D, Elser J,et al. The global carbon cycle: a test of our knowledge of earth as asystem. Science 2000;290:291–6.
Fan LT, Lee Y-H, Beardmore DH. Mechanism of the enzymatichydrolysis of cellulose: effects of major structural features ofcellulose on enzymatic hydrolysis. Biotechnol Bioeng 1980;22:177–99.
Fan LT, Lee Y-H, Beardmore DR. The influence of major structuralfeatures of cellulose on rate of enzymatic hydrolysis. BiotechnolBioeng 1981;23:419–24.
Fan LT, Lee Y-H, Gharpuray MM. The nature of lignocellulosics andtheir pretreatments for enzymatic hydrolysis. Adv Biochem Eng1982;23:157–87.
Fan Z, McBride JE, van Zyl WH, Lynd LR. Theoretical analysis ofselection-based strain improvement for microorganisms withgrowth dependent upon extracytoplasmic enzymes. BiotechnolBioeng 2005;92:35–44.
Fierobe HP, Mechaly A, Tardif C, Belaich A, Lamed R, Shoham Y,et al. Design and production of active cellulosome chimeras.Selective incorporation of dockerin-containing enzymes intodefined functional complexes. J Biol Chem 2001;276:21257–61.
Fierobe HP, Bayer EA, Tardif C, Czjzek M, Mechaly A, Belaich A,et al. Degradation of cellulose substrates by cellulosome chimeras.Substrate targeting versus proximity of enzyme components. J BiolChem 2002;277:49621–30.
Fierobe HP, Mingardon F, Mechaly A, Belaich A, Rincon M, Pages S,et al. Action of designer cellulosomes on homogeneous versuscomplex substrates: controlled incorporation of three distinctenzymes into a defined tri-functional scaffoldin. J Biol Chem2005;280:16325–34.
Fleming K, Gray DG, Matthews S. Cellulose crystallites. Chemistry2001;7:1831–5.
Francisco JA, Stathopoulos C, Warren RAJ, Kilburn DG, Georgiou G.Specific adhesion and hydrolysis of cellulose by intact Escherichiacoli expressing surface anchored cellulase or cellulose bindingdomains. Bio/Technology 1993;11:491–5.
Freeman A, Cohen-Hadar N, Abramov S, Modai-Hod R, Dror Y,Georgiou G. Screening of large protein libraries by the cellimmobilized on adsorbed bead approach. Biotechnol Bioeng2004;86:196–200.
Fujita Y, Takahashi S, UedaM, Tanaka A, Okada H, Morikawa Y, et al.Direct and efficient production of ethanol from cellulosic materialwith a yeast strain displaying cellulolytic enzymes. Appl EnvironMicrobiol 2002;68:5136–41.
Fujita Y, Ito J, Ueda M, Fukuda H, Kondo A. Synergisticsaccharification, and direct fermentation to ethanol, of amorphouscellulose by use of an engineered yeast strain codisplaying threetypes of cellulolytic enzyme. Appl Environ Microbiol 2004;70:1207–12.
Fülöp L, Ponyi T. Rapid screening for endo-b-1,4-glucanase and endo-b-1,4-mannanase activities and specific measurement usingsoluble dye-labelled substrates. J Microbiol Methods 1997;29:15–21.
Galbe M, Zacchi G. A review of the production of ethanol fromsoftwood. Appl Microbiol Biotechnol 2002;59:618–28.
Ghose TK. Continuous enzymatic saccharification of cellulose withculture filtrates of Trichoderma viride QM 6a. Biotechnol Bioeng1969;11:239–61.
Ghose TK. Measurement of cellulase activities. Pure Appl Chem1987;59:257–68.
Godbole S, Decker SR, Nieves RA, Adney WS, Vinzant TB, et al.Cloning and expression of Trichoderma reesei cellobiohydrolase Iin Pichia pastoris. Biotechnol Prog 1999;15:828–33.
Goddard JP, Reymond JL. Enzyme assays for high-throughputscreening. Curr Opin Biotechnol 2004;15:314–22.
Gong C-S, Ladisch MR, Tsao G. Cellobiase from Trichoderma viride:purification, properties, kinetics and mechanism. BiotechnolBioeng 1977;19:959–81.
Gonzalez-Blasco G, Sanz-Aparicio J, Gonzalez B, Hermoso JA,Polaina J. Directed evolution of beta-glucosidase A fromPaenibacillus polymyxa to thermal resistance. J Biol Chem2000;275:13012–708.

475Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Green JW. Wood cellulose. Methods Carbohydr Chem 1963;3:9–21.Grethlein HE. Pretreatment for enhanced hydrolysis of cellulosic
biomass. Biotechnol Adv 1984;2:43–62.Griffin HL. Filter paper assay-effect of time and substrate concentra-
tion on cellulase activity. Anal Biochem 1973;56:621–5.Griffithsa JS, Cheriyanb M, Corbella JB, Pocivavseka L, Fierke CA,
Toone EJ. A bacterial selection for the directed evolution ofpyruvate aldolases. Bioorg Med Chem 2004;12:4067–74.
Gruno M, Valjamae P, Pettersson G, Johansson G. Inhibition of theTrichoderma reesei cellulases by cellobiose is strongly dependenton the nature of the substrate. Biotechnol Bioeng 2004;86:503–11.
Gusakov AV, Sinitsyn AP, Gerasimas VB, Savitskene RY, Stepona-vichus YY. A product inhibition study of cellulases fromTrichoderma longibrachiatum using dyed cellulose. J Biotechnol1985;3:167–74.
Gustafsson C, Govindarajan S, Minshull J. Putting engineering backinto protein engineering: bioinformatic approaches to catalystdesign. Curr Opin Biotechnol 2003;14:366–70.
Hagerman AE, Blau DH, McClure AL. Plate assay for determining thetime of production of protease, cellulase and pectinases bygerminating fungal spores. Anal Biochem 1985;151:334–42.
Haight M. Assessing the environmental burdens of anaerobic digestionin comparison to alternative options for managing the biodegrad-able fraction of municipal solid wastes. Water Sci Technolnolol2005;52:553–9.
Hall DO, Rosillo-Calle F, Williams RH, Woods J. Biomass for energy:supply prospects. In: Johansson TB, Kelly H, Reddy AKN,WillianRH, editors. Renewable energy: sources for fuels and electricity.Washington (DC): Island; 1993.
Halliwell G, Riaz M. The formation of short fibres from nativecellulose by the components of Trichoderma koningii cellulase.Biochem J 1970;116:35–42.
Hamer G. Solid waste treatment and disposal: effects on public healthand environmental safety. Biotechnol Adv 2003;22:71–9.
Han SO, Yukawa H, Inui M, Doi RH. Regulation of expression ofcellulosomal cellulase and hemicellulase genes in Clostridiumcellulovorans. J Bacteriol 2003;185:6067–75.
Han SO, Cho HY, Yukawa H, Inui M, Doi RH. Regulation ofexpression of cellulosomes and noncellulosomal (hemi)cellulolyticenzymes in Clostridium cellulovorans during growth on differentcarbon sources. J Bacteriol 2004;186:4218–27.
Helbert W, Chanzy H, Husum TL, Schulein M, Ernst S. Fluorescentcellulose microfibrils as substrate for the detection of cellulaseactivity. Biomacromolecules 2003;4:481–7.
Henis YI, Yaron T, Lamed RL, Rishpon J, Sahar E, Katchalski-KatzirE. Mobility of enzymes on insoluble substrates: the beta-amylase-starch gel system. Biopolymers 1988;27:123–38.
Henrissat B. Cellulases and their interaction with cellulose. Cellulose1994;1:169–96.
Henrissat B, Driguez H, Viet C, Shulein M. Synergism of cellulasesfrom Trichoderma reesei in the degradation of cellulose. Bio/Technology 1985;3:722–6.
Hestrin S. Bacterial cellulose. Methods Carbohydr. Chem. AcademicPress; 1963. p. 4–9.
Hibbert EG, Baganz F, Hailes HC, Ward JM, Lye GJ, Woodley JM,Dalby PA. Directed evolution of biocatalytic processes. BiomolEng 2005;22:11–9.
Himmel ME, Adney WS, Baker JO, Nieves RA, Thomas SR.Cellulases: structure, function and applications. In: Wyman CE,editor. Handbook on Bioethanol. 1993. p. 144–61.
Himmel ME, Ruth MF, Wyman CE. Cellulase for commodity productsfrom cellosic biomass. Curr Opin Biotechnol 1999;10:358–64.
Hoffert MI, Caldeira K, Jain AK, Haites EF, Harvey LDD, Potter SD,et al. Energy implications of future stabilization of atmosphericCO2 content. Nature 1998;395:881–4.
Hoffert MI, Caldeira K, Benford G, Criswell DR, Green C,Herzog H, et al. Advanced technology paths to global climatestability: energy for a greenhouse planet. Science 2002;298:981–7.
Holt SM, Hartman PA. A zymogram method to detect endoglucanasesfrom Bacillus subtilis, Myrothecium verrucaria and Trichodermareesei. J Ind Microbiol Biotechnol 1994;13(1):2–4.
Holtzapple MT. Cellulose. In: Macrae R, Robinson RK, Saddler MJ,editors. Encyclopedia of food science food technology andnutrition. London: Academic Press; 1993. p. 758–67.
Holtzapple MT, Caram H, Humphrey AE. Determining the inhibitionconstants in the HCH-1 model of cellulose hydrolysis. BiotechnolBioeng 1984;26:735–57.
Holtzapple MT, Cognata M, Shu Y, Hendrickson C. Inhibition ofTrichoderma reesei cellulase by sugars and solvents. BiotechnolBioeng 1990;36:275–87.
Hong J, Tamaki H, Yamamoto K, Kumagai H. Cloning of a geneencoding a thermo-stable endo-beta-1,4-glucanase from Thermo-ascus aurantiacus and its expression in yeast. Biotechnol Lett2003a;25:657–61.
Hong J, Tamaki H, Yamamoto K, Kumagai H. Cloning of a geneencoding thermostable cellobiohydrolase from Thermoascusaurantiacus and its expression in yeast. Appl Microbiol Biotechnol2003b;63:42–50.
Horiuchi T, Tomizawa J-I, Novick A. Isolation and properties ofbacteria capable of high rates of b-galactosidase synthesis.Biochim Biophys Acta 1962;55:152–63.
Hsu T-A. Pretreatment of biomass. In: Wyman CE, editor. Handbookon bioethanol. Taylor & Francis; 1996.
Huang JS, Tang J. Sensitive assay for cellulase and dextranase. AnalBiochem 1976;75:369–77.
Huebner A, Ladisch MR, Tsao GT. Preparation of cellodextrin: anengineering approach. Biotechnol Bioeng 1978;20:1669–77.
Hulme MA. Viscosimetric determination of carboxymethylcellulaseactivity. Methods Enzymol 1988;160:130–5.
Humphrey AE, Moreira AA, Arminger W, Zabriskie D. Production ofsingle cell protein from cellulose wastes. Biotechnol Bioeng Symp1977;7:45–64.
Ilmen M, Saloheimo A, Onnela M-L, Penttila ME. Regulation ofcellulase gene expression in the filamentous fungus Trichodermareesei. Appl Environ Microbiol 1997;63:1298–306.
Irwin D, Spezio M, Walker LP, Wilson DB. Activity studies of eightpurifiied cellulases: specificity, synergism and binding domaineffects. Biotechnol Bioeng 1993;42:1002–13.
Ito J, Fujita Y, Ueda M, Fukuda H, Kondo A. Improvement ofcellulose-degrading ability of a yeast strain displaying Tricho-derma reesei endoglucanase II by recombination of cellulose-binding domains. Biotechnol Prog 2004;20:688–91.
Jang SJ, Ham MS, Lee JM, Chung SK, Lee HJ, Kim JH, et al. Newintegration vector using a cellulase gene as a screening marker forLactobacillus. FEMS Microbiol Lett 2003;224:191–5.
Jarvis M. Cellulose stacks up. Science 2003;426:611–2.Johnson EA, Sakajoh M, Halliwell G, Madia A, Demain AL.
Saccharification of complex cellulosic substrates by the cellulasesystem from Clostridium thermocellum. Appl Environ Microbiol1982a;43:1125–32.
Johnson EA, Reese ET, Demain AL. Inhibition of Clostridiumthermocellum cellulase by end products of cellulolysis. J ApplBiochem 1982b;4:64–71.

476 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Joo H, Lin Z, Arnold FH. Laboratory evolution of peroxide-mediatedcytochrome P450 hydroxylation. Nature 1999a;399:670–3.
Joo H, Lin Z, Arnold FH. Laboratory evolution of peroxide-mediatedcytochrome P450 hydroxylation. Nature 1999b;399:670–3.
Jung HC, Park JH, Park SH, Lebeault JM, Pan JG. Expression ofcarboxymethylcellulase on the surface of Escherichia coli usingPseudomonas syringae ice nucleation protein. Enzyme MicrobTechnol 1998;22:348–54.
Jurgens C, Strom A, Wegener D, Hettwer S, Wilmanns M, Sterner R.Directed evolution of a (beta alpha)8-barrel enzyme to catalyzerelated reactions in two different metabolic pathways. Proc NatlAcad Sci U S A 2000;97:9925–30.
Kamm B, Kamm M. Principles of biorefineries. Appl MicrobiolBiotechnol 2004;64:137–45.
Kaper T, Brouns SJ, Geerling AC, De Vos WM, Van der Oost J. DNAfamily shuffling of hyperthermostable beta-glycosidases. BiochemJ 2002;368:461–70.
Karlsson J, Momcilovic D, Wittgren B, Schulein M, Tjerneld F,Brinkmalm G. Enzymatic degradation of carboxymethyl cellulosehydrolyzed by the endoglucanases Cel5A, Cel7B, and Cel45AfromHumicola insolens and Cel7B, Cel12A and Cel45Acore fromTrichoderma reesei. Biopolymers 2001;63:32–40.
Katchalski-Katzir E, Rishpon J, Sahar E, Lamed RL, Henis YI.Enzyme diffusion and action on soluble and insoluble substratebiopolymers. Biopolymers 1985;24:257–77.
Kidby DK, Davidson DJ. A convenient ferricyanide estimation ofreducing sugars in the nanomole range. Anal Biochem1973;55:321–5.
Kim E, Irwin DC, Walker LP, Wilson DB. Factorial optimization of asix-cellulase mixture. Biotechnol Bioeng 1998;58:494–501.
Kim YS, Jung HC, Pan JG. Bacterial cell surface display of an enzymelibrary for selective screening of improved cellulase variants. ApplEnviron Microbiol 2000;66:788–93.
Kim TH, Kim JS, Sunwoo C, Lee YY. Pretreatment of corn stover byaqueous ammonia. Bioresour Technol 2003;90:39–47.
Kim YW, Lee SS, Warren RA, Withers SG. Directed evolution of aglycosynthase from Agrobacterium sp. increases its catalyticactivity dramatically and expands its substrate repertoire. J BiolChem 2004;279:42787–93.
Kirk O, Borchert TV, Fuglsang CC. Industrial enzyme applications.Curr Opin Biotechnol 2002;13:345–51.
Klein GL, Snodgrass WR. Cellulose. In: Macrae R, Robinson RK,Saddler MJ, editors. Encyclopedia of Food Science, Food Technol-ogy and Nutrition. London: Academic Press; 1993. p. 758–67.
Kleman-Leyer K, Agosin E, Conner AH, Kirk TK. Changes inmolecular size distribution of cellulose during attack by white rotand brown rot fungi. Appl Environ Microbiol 1992;58:1266–70.
Kleman-Leyer KM, Gilkes NR, Miller JRC, Kirk TK. Changes in themolecular-size distribution of insoluble celluloses by the action ofrecombinant Cellulomonas fimi cellulases. Biochem J1994;302:463–9.
Kleman-Leyer KM, Siika-Aho M, Teeri TT, Kirk TK. The cellulaseendoglucanase I and cellobiohydrolase II of Trichoderma reesei actsyngeristically to solubilize native cotton cellulose but not to dec-rease its molecular size. Appl Environ Microbiol 1996;62:2883–7.
Klemm D, Philipp B, Heinze T, Heinze U, Wagenknecht W.Comprehensive cellulose chemistry: I. Fundementals and analyt-ical methods. Weinheim, Germany: Wiley-VCH; 1998.
Knauf M, Moniruzzaman M. Lignocellulosic biomass processing: aperspective. Int Sugar J 2004;106:147–50.
Knowles J, Lethtovaara P, Reeri TT. Cellulase families and their genes.Trends Biotechnol 1987;5:255–61.
Kongruang S, Han MJ, Breton CIG, Penner MH. Quantitative analysisof cellulose-reducing ends. Appl Biochem Biotechnol 2004;113-116:213–31.
Konstantinidis AK, Marsden I, Sinnott ML. Hydrolyses of alpha-andbeta-cellobiosyl fluorides by cellobiohydrolases of Trichodermareesei. Biochem J 1993;291:883–8.
Krassig HA. Cellulose: structure, accessibility, and reactivity.Yverdon, Switzerland: Gordon and Breach Sci. Publishers;1993.
Lebbink JH, Kaper T, Bron P, van der Oost J, de Vos WM. Improvinglow-temperature catalysis in the hyperthermostable Pyrococcusfuriosus beta-glucosidase CelB by directed evolution. Biochemis-try 2000;39:3656–65.
Lee Y-H, Fan LT, Fan L-S. Kinetics of hydrolysis of insoluble celluloseby cellulose. Adv Biochem Eng 1980;17:131–68.
Lee I, Evans BR, Lane LM, Woodward J. Substrate-enzymeinteractions in cellulase systems. Bioresour Technol 1996;58:163–9.
Lee I, Evans BR, Woodward J. The mechanism of cellulase action oncotton fibers: evidence from atomic force microscopy. Ultramicro-scopy 2000;82:213–21.
Lelieveld HLM. The use of continuous cultures for selection andisolation of microorganisms producing extracellular enzymesadapted to extreme environments. Biotechnol Bioeng 1982;24:1419–25.
Leschine SB. Cellulose degradation in anaerobic environments. AnnuRev Microbiol 1995;49:399–426.
Lever M. A new reaction for colorimetric determination ofcarbohydratesnext term. Anal Biochem 1972;47:273–9.
Lever M, Powell JC, Killip M, Small CW. A comparison of 4-hydroxybenzoic acid hydrazide (PAHBAH) with other reagents forthe determination of glucose. J Lab Clin Med 1973;82:649–55.
Lin H, Cornish VW. Screening and selection methods for large-scaleanalysis of protein function. Angew Chem Int Ed Engl 2002;41:4402–25.
Lin E, Wilson DB. Transpription of the celE gene in Thermomonos-pora fusca. J Bacteriol 1988;170:3838–42.
Lin KW, Ladisch MR, Schaeffer DM, Noller CH, Lechtenberg V, TsaoGT. Review on effect of pretreatment on digestibility of cellulosicmaterials. AIChE Symp Ser 1981;203:102–6.
Lin H, Tao H, Cornish VW. Directed evolution of a glycosynthasevia chemical complementation. J Am Chem Soc 2004;126:15051–15059.
Lynd LR. Overview and evaluation of fuel ethanol from cellulosicbiomass: technology, economics, the environment, and policy.Annu Rev Energy Environ 1996;21:403–65.
Lynd LR, Cushman JH, Nichols RJ, Wyman CE. Fuel ethanol fromcellulosic biomass. Science 1991;251:1318–23.
Lynd LR, Wyman CE, Gerngross TU. Biocommodity engineering.Biotechnol Prog 1999;15:777–93.
Lynd LR, Weimer PJ, van Zyl WH, Pretorius IS. Microbial celluloseutilization: fundamentals and biotechnology. Microbiol Mol BiolRev 2002;66:506–77.
Lynd LR, van Zyl WH, McBride JE, Laser M. Consolidatedbioprocessing of cellulosic biomass: an update. Curr OpinBiotechnol 2005;16:577–83.
Lynd LR, Weimer PJ, Wolfaardt G, Zhang Y-HP. Cellulose hydrolysisby Clostridium thermocellum: a microbial perspective. In: KataevaIA, editor. Cellulosome. Hauppauge (NY) USA: Nova SciencePublishers, Inc. in press. [ISBN 1594549508]
Mach RL, Zeilinger S. Regulation of gene expression in industrialfungi: Trichoderma. Appl Microbiol Biotechnol 2003;60:515–22.

477Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Mackenzie LF, Wang Q, Warren RAJ, Withers SG. Glycosynthases:mutant glycosidases for oligosaccharide synthesis. J Am ChemSoc 1998;120:5583–4.
Manning K. Improved viscometric assay for cellulase. J BiochemBiophys Methods 1981;5:189–202.
Mansfield SD, Mooney C, Saddler JN. Substrates and enzymecharacteristics that limit cellulose hydrolysis. Biotechnol Prog1999;15:804–16.
Mayer C, Jakeman DL, Mah M, Karjala G, Gal L, Warren RA, et al.Directed evolution of new glycosynthases from Agrobacteriumbeta-glucosidase: a general screen to detect enzymes foroligosaccharide synthesis. Chem Biol 2001;8:437–43.
McCarthy JK, Uzelac A, Davis DF, Eveleigh DE. Improved catalyticefficiency and active site modification of 1,4-beta-D-glucanglucohydrolase A from Thermotoga neapolitana by directedevolution. J Biol Chem 2004;279:11495–502.
McCleary BV. New chromogenic substrates for the assay of alpha-amylase and (1-4)-beta-glucanase. Carbohydr Res 1980;86: 97–104.
McMillian JD. Pretreatment of lignocellulosic biomass. ACS Ser1994;566:234–92.
Melillo JM, Steudler PA, Aber JD, Newkirk K, Lux H, Bowles FP,et al. Soil warming and carbon-cycle feedbacks to the climatesystem. Science 2002;298:2173–6.
Mielenz JR. Ethanol production from biomass: technology andcommercialization status. Curr Opin Microbiol 2001;4:324–9.
Miller GL. Use of dinitrosalicylic acid reagent for determination ofreducing sugar. Anal Chem 1959;31:426–8.
Miller GL. Micro column chromatographic method for analysis ofoligosaccharides. Anal Biochem 1960;2:133–40.
Miller GL. Cellodextrins. Methods Carbohydr Chem 1963;3:134–9.Miller GL, Blum R, Glennon WE, Burton AL. Measurement of
carboxymethylcellulase activity. Anal Biochem 1960;1:127–32.Millett MA, Baker AJ, Satter LD. Physical and chemical pretreatments
for enhancing cellulose saccharification. Biotechnol Bioeng Symp1976;6:125–53.
Mingardon F, Perret S, Belaich A, Tardif C, Belaich JP, Fierobe HP.Heterologous production, assembly, and secretion of a minicellu-losome by Clostridium acetobutylicum ATCC 824. Appl EnvironMicrobiol 2005;71:1215–22.
Mohanty AK, Misra M, Hinrichsen G. Biofibres, biodegradablepolymers and biocomposites: an overview. Macromol Mater Eng2000;276-277:1–24.
Morag E, Bayer EA, Lamed RL. Affinity digestion for the near-totalrecovery of purified cellulosome from Clostridium thermocellum.Enzyme Microb Technol 1992;14:289–92.
Moreira N. Growing expectations: new technology could turn fuel intoa bump crop. Sci News Online 2005;168(14):209–24.
Mosier N, Wyman CE, Dale BE, Elander RT, Lee YY, Holtzapple M,et al. Features of promising technologies for pretreatment oflignocellulosic biomass. Bioresour Technol 2005;96:673–86.
Murai T, Ueda M, Atomi H, Shibasaki Y, Kamasawa N, Osumi M,et al. Genetic immobilization of cellulase on the cell surface ofSaccharomyces cerevisiae. Appl Microbiol Biotechnol 1997;48:499–503.
Murai T, Ueda M, Kawaguchi T, Arai M, Tanaka A. Assimilation ofcellooligosaccharides by a cell surface-engineered yeast expres-sing beta-glucosidase and carboxymethylcellulase from Aspergil-lus aculeatus. Appl Environ Microbiol 1998;64:4857–61.
Murashima K, Chen CL, Kosugi A, Tamaru Y, Doi RH, Wong SL.Heterologous production ofClostridium cellulovorans engB, usingprotease-deficient Bacillus subtilis, and preparation of activerecombinant cellulosomes. J Bacteriol 2002a;184:76–81.
Murashima K, Kosugi A, Doi RH. Thermostabilization of celluloso-mal endoglucanase EngB from Clostridium cellulovorans by invitro DNA recombination with non-cellulosomal endoglucanaseEngD. Mol Microbiol 2002b;45:617–26.
Naki D, Paech C, Ganshaw G, Schellenberger V. Selection of asubtilisin-hyperproducing Bacillus in a highly strcutured environ-ment. Appl Microbiol Biotechnol 1998;49:290–4.
Nelson N. A photometric adaptation of the Somogyi method for thedetermination of glucose. J Biol Chem 1944;153:375–80.
Ng TK, Zeikus JG. Synthesis of [14C] cellobiose with Clostridiumthermocellum: cellobiose phosphorylase. Appl Environ Microbiol1986;52:902–4.
Nishiyama Y, Sugiyama J, Chanzy H, Langan P. Crystal structure andhydrogen bonding system in cellulose Ia from aynchrotron X-rayand neutron fiber diffraction. J Am Chem Soc 2003;125(47):14300–6.
Notley SM, Pettersson B,Wagberg L. Direct measurement of attractivevan der Waals' forces between regenerated cellulose surfaces in anaqueous environment. J Am Chem Soc 2004;126(43):13930–1.
Nummi M, Fox PC, Niku-Passvolva M-L, Enari T-M. Nephelometricand turbidometric assays of cellulase activity. Anal Biochem1981;116:133–6.
Ohgren K, Galbe M, Zacchi G. Optimization of steam pretreatment ofSO2-impregnated corn stover for fuel ethanol production. ApplBiochem Biotechnol 2005;121/124:1055–67.
Okazaki M, Moo-Young M. Kinetics of enzymatic hydrolysis ofcellulose: analytical description of a mechanistic model. Biotech-nol Bioeng 1978;20:637–63.
Oksanen T, Pere J, Paavilainen L, Buchert J, Viikari L. Treatment ofrecycled kraft pulps with Trichoderma reesei hemicellulases andcellulases. J Biotechnol 2000;78:39–48.
Olsen M, Iverson B, Georgiou G. High-throughput screening ofenzyme libraries. Curr Opin Biotechnol 2000;11:331–7.
O'Sullivan AC. Cellulose: the structure slowly unravels. Cellulose1997;4:173–207.
Otten LG, Quax WJ. Directed evolution: selecting today's biocata-lysts. Biomol Eng 2005;22:1–9.
Pan X, Arato C, Gilkes NR, Gregg D, Mabee W, Pye K, et al.Biorefining of softwood using ethanol organosolv pulping:preliminary evaluation of process streams for manufacture offuel-grade ethanol and co-products. Biotechnol Bioeng2005a;90:473–81.
Pan X, Xie D, Gilkes N, Gregg DJ, Saddler JN. Strategies to enhancethe enzymatic hydrolysis of pretreated softwood with high residuallignin content. Appl Biochem Biotechnol 2005b;124:1069–80.
Park JT, Johnson MJ. A submicrodetermination of glucose. J BiolChem 1949;181:149–51.
Penttila M, Andre L, Lehtovaara P, Bailey M, Teeri TT, Knowles J.Efficient secretion of two fungal cellobiohydrolases by Saccharo-myces cerevisiae. Gene 1988;63:103–12.
Pere J, Puolakka A, Nousiainan P, Buchert J. Action of purifiedTrichoderma reesei cellulases on cotton fibers and yarn. JBiotechnol 2001;89:247–55.
Pereira AN,Mobedshahi M, LadischMR. Preparation of cellodextrins.Methods Enzymol 1988;160:26–43.
PiontekM, Hagedorn J, Hollenberg CP, Gellissen G, Strasser AW. Twonovel gene expression systems based on the yeasts Schwannio-myces occidentalis and Pichia stipitis. Appl Microbiol Biotechnol1998;50:331–8.
Polacheck I, Melamed M, Bercovier H, Salkin IF. beta-Glucosidase inCandida albicans and its application in yeast identification. J ClinMicrobiol 1987;25:907–10.

478 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Pye EK, Lora JH. The Alcell process: a proven alternative to krfatpulping. Tappi J 1991;74(3):113–8.
Ramos LA, Assaf JM, ElSeoud OA, Frollini E. Influence of thesupramolecular structure and physicochemical properties ofcellulose on its dissolution in a lithium chloride/N,N-dimethyla-cetamide solvent system. Biomacromolecules 2005;6:2638–47.
Reddy N, Yang Y. Biofibers from agricultural byproducts for industrialapplications. Trends Biotechnol 2005;23:22–7.
Rescigno A, Rinaldi AC, Curreli N, Olianas A, Sanjust E. A dyedsubstrate for the assay of endo-1,4-beta-glucanases. J BiochemBiophys Methods 1994;28:123–9.
Roe JH. The determination of sugar in blood and spinal fluid withanthrone reagent. J Biol Chem 1955;212:335–43.
Ruiz R, Ehrman T. Determination of carbohydrates in biomass by highperformance liquid chromatography Laboratory Analytic Proce-dure LAP-002, 1996.
Russell JB, Rychlik JL. Factors that alter rumen microbial ecology.Science 2001;292:1119–22.
Sabathe F, Soucaille P. Characterization of the CipA scaffoldingprotein and in vivo production of a minicellulosome in Clostridiumacetobutylicum. J Bacteriol 2003;185:1092–6.
Saddler JN, Ramos LP, Breuil C. Steam pretreatment of lignocellulosicresidues. In: Saddler JN, editor. Bioconversion of forest andagricultural plant residues. CAB International; 1993. p. 73–91.
Sakon J, Adney W, Himmel M, Thomas S, Karplus P. Crystallinestructure of thermostable family 5 endoglucanase EG1 fromAcidothermus cellulolyticum in complex with cellotetraose.Biochemistry 1996;35:10648–60.
SaloheimoM, PaloheimoM, Hakola S, Pere J, Swanson B, NyyssonenE, et al. Trichoderma reesei protein with sequence similarity to theplant expansins, exhibits disruption activity on cellulosic materials.Eur J Biochem 2002;269:4202–11.
Sassner P, Galbe M, Zacchi G. Steam pretreatment of Salix with andwithout SO2 impregnation for production of bioethanol. ApplBiochem Biotechnol 2005;121/124:1101–17.
Sauer J, Sigurskjold BW, Christensen U, Frandsen TP, MirgorodskayaE, Harrison M, et al. Glucoamylase: structure/function relation-ships, and protein engineering. Biochim Biophys Acta 2000;1543:275–93.
Schlamadinger B, Marland G. The role of forest and bioenergystrategies in the global carbon cycle. Biomass Bioenergy1996;10:275–300.
Schloss PD, Hay AG,Wilson DB, Gossett JM,Walker LP. Quantifyingbacterial population dynamics in compost using 16S rRNA geneprobes. Appl Microbiol Biotechnol 2005;66:457–63.
Schmid G, Bisell M, Wandrey C. Preparation of cellodextrins andisolation of oligomeric side components and their characterization.Anal Biochem 1988;175:573–83.
Schmidt-Dannert C. Directed evolution of single proteins, metabolicpathways, and viruses. Biochemistry 2001;40:13125–36.
Schmidt-Dannert C, Arnold FH. Directed evolution of industrialenzymes. Trends Biotechnol 1999;17:135–6.
Schulein M. Protein engineering of cellulases. Biochim Biophys Acta2000;1543:239–52.
Schwarz WH. The cellulosome and cellulose degradation by anaerobicbacteria. Appl Microbiol Biotechnol 2001;56(5–6):634–49.
Schwarz W, Chan M, Breuil C, Saddler JN. Comparison of HPLC andcolorometric methods for measuring cellulolytic activity. ApplMicrobiol Biotechnol 1988;28:398–403.
Setlow B, Cabrera-Hernandez A, Cabrera-Martinez RM, Setlow P.Identification of aryl-phospho-beta-D-glucosidases in Bacillussubtilis. Arch Microbiol 2004;181:60–7.
Sharrock KR. Cellulase assay methods: a review. J Biochem BiophysMethods 1988;17:81–105.
Sheehan J, Himmel M. Enzymes, energy, and the environment: astrategic perspective on the U.S. department of energy's researchand development activities for bioethanol. Biotechnol Prog1999;15:817–27.
Shinmyo A, Garcia-Martinez DV, Demain AL. Studies on theextracellular cellulolytic enzyme complex produced by Clostrid-ium thermocellum. J Appl Biochem 1979;1:202–9.
Shintate K, Kitaoka M, Kim YK, Hayashi K. Enzymatic synthesis of alibrary of beta-(1->4) hetero-D-glucose and D-xylose-basedoligosaccharides employing cellodextrin phosphorylase. Carbo-hydr Res 2003;338:1981–90.
Shoemaker HE, Mink D, Wubbolts MG. Dispelling the myths-biocatalysis in industrial synthesis. Science 2003;299:1694–7.
Sinnott ML. The cellobiohydrolases of Trichoderma reesei: areview of indirect and direct evidence that their function is notjust glycosidic bond hydrolysis. Biochem Soc Trans1998;26:160–4.
Skopec CE, Himmel ME, Matthews JF, Brady JW. Energetics fordisplacing a single chain from the surface of microcrystallinecellulose into the active site of Acidothermus cellulolyticus Cel5A.Protein Eng 2003;16:1005–15.
Smiley JA, Benkovic SJ. Selection of catalytic antibodies for abiosynthetic reaction from a combinatorial cDNA library bycomplementation of an auxotrophic Escherichia coli: antibodiesfor orotate decarboxylation. Proc Natl Acad Sci U S A 1994;91:8319–23.
Soderstrom J, Pilcher L, Galbe M, Zacchi G. Combined use of H2SO4
and SO2 impregnation for steam pretreatment of spruce in ethanolproduction. Appl Biochem Biotechnol 2003;105/108:127–40.
Somogyi M. Notes on sugar determination. J Biol Chem1952;195:19–23.
Sonderegger M, Sauer U. Evolutionary engineering of Saccharomycescerevisiae. Appl Environ MicrobiolVol nr 69 2003:1990–8.
Spiridonov NA, Wilson DB. Regulation of biosynthesis of individualcellulases in Thermomonospora fusca. J Bacteriol 1998;180:3549–52.
Spiridonov NA,Wilson DB. A celR mutation affecting transcription ofcellulase genes in Thermobifida fusca. J Bacteriol 2000;182:252–5.
Srisodsuk M, Kleman-Leyer K, Keranen S, Kirk TK, Teeri TT. Modesof action on cotton and bacterial cellulose of a homologousendoglucanase-exoglucanase pair from Trichoderma reesei. Eur JChem 1998;251:885–92.
Stemmer WPC. Rapid evolution of a protein in vitro by DNAshuffling. Nature 1994a;370:389–91.
Stemmer WPC. DNA shuffling by random fragmentation andreassembly: in vitro recombination for molecular evolution. ProcNatl Acad Sci U S A 1994b;91:10747–51.
Stevenson DM, Weimer PJ. Expression of 17 genes in Clostridiumthermocellum ATCC 27405 during fermentation of cellulose orCellobiose in continuous culture. Appl Environ Microbiol2005;71:4672–8.
Striegel AM. Theory and applications of DMAc/LiCl in the analysis ofpolysacharrides. Carbohydr Polym 1997;34:267–74.
Strobel HJ, Russell JB. Regulation of beta-glucosidase in Bacteroidesruminicola by a different mechanism: growth rate-dependentderepression. Appl Environ Microbiol 1987;53:2505–10.
Strobel HJ, Caldwell FC, Dawson KA. Carbohydrate transport by theanaerobic thermophilic Clostridium thermocellum LQRI. ApplEnviron Microbiol 1995;61:4012–5.

479Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Sundberg SA. High-throughput and ultra-high-throughput screening:solution-and cell-based approaches. Curr Opin Biotechnol2000;11:47–53.
Swatloski RP, Spear SK, Holbrey JD, Rogers RD. Dissolution ofcellulose with ionic liquids. J Am Chem Soc 2002;124:4974–5.
Tao H, Cornish VW. Milestones in directed enzyme evolution. CurrOpin Chem Biol 2002;6:858–64.
Taylor SV, Kast P, Hilvert D. Investigating and engineering enzymesby genet ic select ion. Angew Chem Int Ed Engl2001;40:3310–35.
Teeri TT. Crystalline cellulose degradation: new insights into thefunction of cellobiohydrolases. Trends Biotechnol 1997;15:160–7.
Teeri TT, Koivula A, Linder M, Wohlfahrt G, Divne C, Jones TA.Trichoderma reesei cellobiohydrolases: why so efficient oncrystalline cellulose? Biochem Soc Trans 1998;26:173–8.
Ten LN, Im W-T, Kim M-K, Kang MS, Lee S-T. Development of aplate technique for screening of polysaccharide-degrading micro-organisms by using a mixture of insoluble chromogenic substrates.J Microbiol Methods 2004;56:375–82.
Teymouri F, Laureano-Perez L, Alizadeh H, Dale BE. Optimization ofthe ammonia fiber explosion (AFEX) treatment parameters forenzymatic hydrolysis of corn stover. Bioresour Technol2005;96:2014–8.
Tomme P, Driver DP, Amandoron EA, Miller JRC, Warren RAJ,Kilburn DG. Comparsion of a fungal (family I) and bacterial(family II) cellulose-binding domain. J Bacteriol 1995;177:4356–63.
Tomme P, Kwan E, Gilkes NR, Kilburn DG, Warren RA.Characterization of CenC, an enzyme from Cellulomonas fimiwith both endo-and exoglucanase activities. J Bacteriol1996;178:4216–23.
Tsen S-D, Lai S-C, Pang C-P, Lee J-I, Wilson TH. Chemostat selectionof an Escherichia coli mutant containing permease withenhanced lactose affinity. Biochem Biophys Res Commun1996;224:351–7.
Tuohy MG, Walsh DJ, Murray PG, Claeyssens M, Cuffe MM, SavageAV, et al. Kinetic parameters and mode of action of thecellobiohydrolases produced by Talaromyces emersonii. BiochimBiophys Acta 2002;1596:366–80.
Valjamae P, Sild V, Pettersson G, Johansson G. The initial kinetics ofhydrolysis by cellobiohydrolases I and II is consistent with acellulose surface- erosion model. Eur J Biochem 1998;253:469–75.
Valjamae P, Sild V, Pettersson G, Johansson G. Acid hydrolysis ofbacterial cellulose reveals different modes of synergistic actionbetween cellobiohydrolase I and endoglucanase I. Eur J Biochem1999;266:327–34.
van Beilen JB, Li Z. Enzyme technology: an overview. Curr OpinBiotechnol 2002;13:338–42.
van Rensburg P, van Zyl WH, Pretorius IS. Expression of theButyrivibrio fibrisolvens endo-beta-1,4-glucanase gene togetherwith the Erwinia pectate lyase and polygalacturonase genes inSaccharomyces cerevisiae. Curr Genet 1994;27:17–22.
van Rensburg P, van Zyl WH, Pretorius IS. Co-expression of aPhanerochaete chrysosporium cellobiohydrolase gene and aButyrivibrio fibrisolvens endo-beta-1,4-glucanase gene in Saccha-romyces cerevisiae. Curr Genet 1996;30:246–50.
van Rensburg P, Van Zyl WH, Pretorius IS. Engineering yeast forefficient cellulose degradation. Yeast 1998;14:67–76.
van Tilbeurgh H, Claeyssens M. Detection and differentiation ofcellulase components using low molecular mass fluorogenicsubstrates. FEBS Lett 1985;187:283–8.
van Tilbeurgh H, Claeyssens M, de Bruyne CK. The use of 4-methylumbelliferyl and other chromophoric glycosides in thestudy of cellulolytic enzymes. FEBS Lett 1982;149:152–6.
van Tilbeurgh H, Pettersson G, Bhikabhai R, De Boeck H, ClaeyssensM. Studies of the cellulolytic system of Trichoderma reesei QM9414. Reaction specificity and thermodynamics of interactions ofsmall substrates and ligands with the 1,4-beta-glucan cellobiohy-drolase II. Eur J Biochem 1985;148:329–34.
van Wyk JP. Biotechnology and the utilization of biowaste as aresource for bioproduct development. Trends Biotechnol2001;19:172–7.
Viles FJ, Silverman L. Determination of starch and cellulose withanthrone. Anal Chem 1949;21:950–3.
Voloch M, Ladisch MR, Cantarlla M, Tsao GT. Preparation ofcellodextrins using sulfuric acid. Biotechnol Bioeng 1984;26:557–9.
Waffenschmidt S, Janeicke L. Assay of reducing sugars in thenanomole range with 2,2-bicinchoninate. Anal Biochem 1987;165:337–40.
Wahler D, Reymond J-L. Novel methods for biocatalyst screening.Curr Opin Chem Biol 2001;5:152–8.
Walker LP, Wilson DB, Irwin DC. Measuring fragmentation ofcellulose by Thermomonospora fusca cellulase. Enzyme MicrobTechnol 1990;12:378–86.
Walker LP, Wilson DB, Irwin DC. Fragmentation of cellulose by themajor Thermomonospora fusca cellulase cellulases, Trichodermareesei CBH1 and their mixtures. Biotechnol Bioeng 1992;40:1019–26.
Walker LP, Belair CD, Wilson DB, Irwin DC. Engineering cellulasemixtures by varying the mole fraction of Thermomonospora fuscaE5 and E3, Trichoderma reesei CBH1 and Caldocellumsaccharolyticum b-glucosidase. Biotechnol Bioeng 1993;42:1019–28.
Walseth CS. Occurrence of cellulases in enzyme preparations frommicroorganisms. TAPPI 1952;35:228–33.
Wang W, Liu J, Chen G, Zhang Y, Gao P. Function of a low molecularweight peptide from Trichoderma pseudokoningii S38 duringcellulose biodegradation. Curr Microbiol 2003;46:371–9.
Wang T, Liu X, Yu Q, Zhang X, Qu Y, Gao P, et al. Directed evolutionfor engineering pH profile of endoglucanase III from Trichodermareesei. Biomol Eng 2005;22:89–94.
Weil JWP, Kohlmann K, Ladisch MR. Cellulose pretreatments oflignocellulosic substrates. Enzyme Microb Technol 1994;16:1002–14.
Weimer PJ, Lopez-Guisa JM, French AD. Effect of cellulosefine structure on kinetics of its digestion by mixed ruminalmicroorganisms in vitro. Appl Environ Microbiol 1990;56:2419–21.
Wilson DB. Studies of Thermobifida fusca plant cell wall degradingenzymes. Chem Rec 2004;4:72–82.
Wilson DB, Walker LP. Engineering cellulases. Bioresour Technol1991;36:97–9.
Wingren A, Galbe M, Zacchi G. Techno-economic evaluation ofproducing ethanol from softwood: comparison of SSF and SHFand identification of bottlenecks. Biotechnol Prog 2003;19:1109–17.
Wirth SJ, Wolf GA. Micro-plate colourimetric assaynext term forEndo-acting previous termcellulase, next term xylanase, chitinase,1,3-b-glucanase and amylase extracted from forest soil horizons.Soil Biol Biochem 1992;24:511–9.
Wirth TE, Gray CB, Podesta JD. The future of energy policy. ForeignAff 2003;82:125–44.

480 Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Wither SG. Mechanism of glycosyl transferase and hydrolases.Carbohydr Polym 2001;44:325–37.
Wittrup KD. Protein engineering by cell-surface display. Curr OpinBiotechnol 2001;12:395–9.
Wolfenden R, Snider MJ. The depth of chemical time and the powderof enzyme as catalysts. Acc Chem Res 2001;34:938–45.
Wolfgang DE, Wilson DB. Mechanistic studies of active site mutantsof Thermomonospora fusca endocellulase E2. Biochemistry1999;38:9746–51.
Wolfram ML, Dacons JC. The polymer-homogous series of oligo-sacchardies from cellulose. J Am Chem Soc 1952;74:5331–3.
Wolfson W. Diversa builds a business with designer bacteria. ChemBiol 2005;12:503–5.
Wong KK, Richardson JD, Mansfield SD. Enzymatic treatment ofmechanical pulp fibers for improving papermaking properties.Biotechnol Prog 2000;16:1025–9.
Wood TM. Properties and modes of action of cellulases. BiotechnolBioeng Symp 1975;5:111–37.
Wood TM. Preparation of crystalline, amorphous and dyed cellulasesubstrate. Methods Enzymol 1988;166:19–45.
Wood TM, Bhat KM. Methods for measuring cellulase activities.Methods Enzymol 1988;160:87–117.
Wood TM, Garica-Campayo V. Enzymology of cellulose degradation.Biodegradation 1990;1:147–61.
Wood BE, Ingram LO. Ethanol production from cellobiose,amorphous cellulose, and crystalline cellulose by recombi-nant Klebsiella oxytoca containing chromosomally integratedZymomonas mobilis genes for ethanol production andplasmids expressing thermostable cellulase genes fromClostridium thermocellum. Appl Environ Microbiol1992;58:2103–10.
Woodward J, Affholter KA, Noles KK, Troy NT, Gaslightwala SF.Does cellobiohydrolase II core protein from Trichoderma reeseidisperse cellulose macrofibrils? Enzyme Microb Technol1992;14:625–30.
Wyman CE. Ethanol from lignocellulosic biomass: technology,economics, and opportunities. Bioresour Technol 1994;50:3–15.
Wyman CE. Biomass ethanol: technical progress, opportunities, andcommercial challenges. Annu Rev Energy Environ 1999;24:189–226.
Wyman CE. Potential synergies and challenges in refining cellulosicbiomass to fuels, chemicals, and power. Biotechnol Prog 2003;19:254–262.
Wyman CE, Dale BE, Elander RT, Holtzapple M, Ladisch MR, LeeYY. Coordinated development of leading biomass pretreatmenttechnologies. Bioresour Technol 2005a;96:1959–66.
Wyman CE, Dale BE, Elander RT, Holtzapple M, Ladisch MR, LeeYY. Comparative sugar recovery data from laboratory scaleapplication of leading pretreatment technologies to corn stover.Bioresour Technol 2005b;96:2026–32.
Xiao Z, Storms R, Tsang A. Microplate-based filter paper assay tomeasure total cellulase activity. Biotechnol Bioeng 2004;88:832–7.
Yamada M, Amano Y, Horikawa E, Nozaki K, Kanda T. Mode ofaction of cellulases on dyed cotton with a reactive dye. BiosciBiotechnol Biochem 2005;69:45–50.
Youvan DC, Goldman E, Delagrave S, Yang MM. Digital imagingspectroscopy for massively parallel screening of mutants. MethodsEnzymol 1995;246:732–48.
Zhang Y-HP, Lynd LR. Quantification of cell and cellulase massconcentrations during anaerobic cellulose fermentation: develop-ment of an ELISA-based method with application to
Clostridium thermocellum batch cultures. Anal Chem 2003a;75:219–27.
Zhang Y-HP, Lynd LR. Cellodextrin preparation by mixed-acidhydrolysis and chromatographic separation. Anal Biochem2003b;322:225–32.
Zhang Y-HP, Lynd LR. Kinetics and relative importance ofphosphorolytic and hydrolytic cleavage of cellodextrins andcellobiose in cell extracts of Clostridium thermocellum. ApplEnviron Microbiol 2004a;70:1563–9.
Zhang Y-HP, Lynd LR. Toward an aggregated understanding ofenzymatic hydrolysis of cellulose: noncomplexed cellulasesystems. Biotechnol Bioeng 2004b;88:797–824.
Zhang Y-HP, Lynd LR. Regulation of cellulase synthesis in batch andcontinuous cultures of Clostridium thermocellum. J Bacteriol2005a;187:99–106.
Zhang Y-HP, Lynd LR. Determination of the number-average degreeof polymerization of cellodextrins and cellulose with application toenzymatic hydrolysis. Biomacromolecules 2005b;6:1510–5.
Zhang Y-HP, Lynd LR. Cellulose utilization by Clostridium thermo-cellum: bioenergetics and hydrolysis product assimilation. ProcNatl Acad Sci U S A 2005c;102:7321–5.
Zhang Y-HP, Lynd LR. Biosynthesis of radio-labeled cellodextrins bythe Clostridium thermocellum cellobiose and cellodextrin phos-phorylases for measurement of intracellular sugars. Appl Micro-biol Biotechnol 2006;70(1):123–9.
Zhang Y-HP, Lynd LR. A functionally-based model for hydrolysis ofcellulose by fungal cellulase. Biotechnol Bioeng [in press].doi:10.1002/bit.20906.
Zhang S, Wilson DB. Surface residue mutations which change thesubstrate specificity of Thermomonospora fusca endoglucanaseE2. J Biotechnol 1997;57:101–13.
Zhang S, Wolfgang DE, Wilson DB. Substrate heterogeneity causesthe nonlinear kinetics of insoluble cellulose hydrolysis. BiotechnolBioeng 1999;66:35–41.
Zhang S, Barr BK, Wilson DB. Effects of noncatalytic residuemutations on substrate specificity and ligand binding of Thermo-bifida fusca endocellulase cel6A. Eur J Biochem 2000a;267:244–52.
Zhang S, Irwin DC, Wilson DB. Site-directed mutation of noncatalyticresidues of Thermobifida fusca exocellulase Cel6B. Eur J Biochem2000b;267:3101–15.
Zhang Y-HP, Cui J-B, Lynd LR, Kuang LR. A transition from celluloseswelling to cellulose dissolution by o-phosphoric acid: evidencesfrom enzymatic hydrolysis and supramolecular structure. Bioma-cromolecules 2006;7(2):644–8.
Zhbankov RG. Hydrogen bonds and structure of carbohydrates. J MolStruct 1992;270:523–39.
Zhou S, Ingram LO. Engineering endoglucanase-secreting strains ofethanologenic Klebsiella oxytoca P2. J Ind Microbiol Biotechnol1999;22:600–7.
Zhou S, Ingram LO. Simultaneous saccharification and fermentationof amorphous cellulose to ethanol by recombinant Klebsiellaoxytoca SZ21 without supplemental cellulase. Biotechnol Lett2001;23:1455–62.
Zhou S, Yomano LP, Saleh AZ, Davis FC, Aldrich HC, Ingram LO.Enhancement of expression and apparent secretion of Erwiniachrysanthemi endoglucanase (encoded by celZ) in Escherichia coliB. Appl Environ Microbiol 1999;65:2439–45.
Zhou S, Davis FC, Ingram LO. Gene integration and expression andextracellular secretion of Erwinia chrysanthemi endoglucanaseCelY (celY) and CelZ (celZ) in ethanologenic Klebsiella oxytocaP2. Appl Environ Microbiol 2001;67:6–14.

481Y.-H. Percival Zhang et al. / Biotechnology Advances 24 (2006) 452–481
Zverlov VV, Fuchs KP, Schwarz WH. Chi18A, the endochitinase inthe cellulosome of the thermophilic, cellulolytic bacteriumClostridium thermocellum. Appl Environ Microbiol 2002a;68:3176–9.
Zverlov VV, Velikodvorskaya GA, Schwarz WH. A newly describedcellulosomal cellobiohydrolase, CelO, from Clostridium thermo-cellum: investigation of the exo-mode of hydrolysis, and bindingcapacity to crystalline cellulose. Microbiology 2002b;148:247–55.
Zverlov VV, Velikodvorskaya GA, Schwarz WH. Two new cellulo-some components encoded downstream of celI in the genome of
Clostridium thermocellum: the non-processive endoglucanaseCelN and the possibly structural protein CseP. Microbiology2003;149:515–24.
Zverlov VV, Schantz N, Schwarz WH. A major new component in thecellulosome of Clostridium thermocellum is a processive endo-beta-1,4-glucanase producing cellotetraose. FEMS Microbiol Lett2005;249:353–8.





























