Embed Size (px)
Citation preview

Cell Tiss. Res. 154, 265--274 (1974) �9 by Springer-Verlag 1974
Cell Autolysis and Deletion in the Planarian Polycelis tenuis Iijima
I. D. Bowen and T. A. Ryder *
Department of Zoology, University College, Cardiff, Wales, U.K.
Received August 14, 1974
Summary. Selective cell autolysis and deletion is described in the normal adult planarian. This appears to be a common feature and the sequence of events leading from cytoplasmic vesiculation and peripheral lysis to complete cellular breakdown is described. - In lysing cells acid phosphatase activity can be demonstrated in the cytoplasm as well as in lysosome- like vacuoles and occasionally in the Golgi apparatus. The role of cell autolysis and deletion in tissue regulation is discussed.
Key words: Cell autolysis - - Deletion - - Planaria - - Acid phosphatase - - Fine structure.
Introduction I n this s tudy, cellular lysis and deletion is described in normal, adult, uns tarved
planaria. The phenomenon of cell lysis and death was recognised long ago by Looss (1889) working on sarcolysis in amphibian metamorphosis and appears to be a common feature in the development of many animals. Glucksmann (1951) has reviewed seventy six examples of cell death occuring as par t of vertebrate ontogeny. Weber (1969) and Fox (1972) have dealt with cell deletion and tissue involution in the metamorphosing tadpole tail. Cell lysis and death, accompanying metamorphosis, has been described in invertebrates, especially insects (Lockshin and Williams, 1965a). To-date, no evidence is available of cell death and deletion in fully grown invertebrates which are not undergoing developmental change. The process of cell autolysis and deletion has been demonstra ted both cytochemic- ally and morphologically in the present study.
Sheldrake (1974) in a recent review has indicated tha t little is known about the chemistry of dying cells. I n this investigation acid phosphatase is employed as a marker of lytic activity. Precedence for this has been set by several authors (Brandes et al., 1965; Lockshin and Williams, 1965b). Fine structural evidence of lysis is also presented. Various studies on the nature of planarian parenchyma are available (Pedersen, 1959a, b, 1961, 1963; TSrSk and RShlich, 1960; Skaer, 1961 ; Klima, 1961). The fine structure of the gut and the pha rynx has been de- scribed by ishii (1962, 1964, 1965, 1966); Bowen and Ryder (1973) and Bowen et al. (1974). None of these reports, however, deal with the phenomenon of cell death.
Materials and Methods The materials and methods employed were identical to those presented by Bowen and
Ryder (1973). Briefly, animals 5 to 10 mm in length were pre-fixed for 3 hours at 0 to 4~
Send o/[print requests to: Dr. I. D. Rowen, Department of Zoology, University College, P. O. Box 78, Cardiff, Wales, U. K. * This research was supported by the Scientific Research Council, Grant No. B/RG/00869.
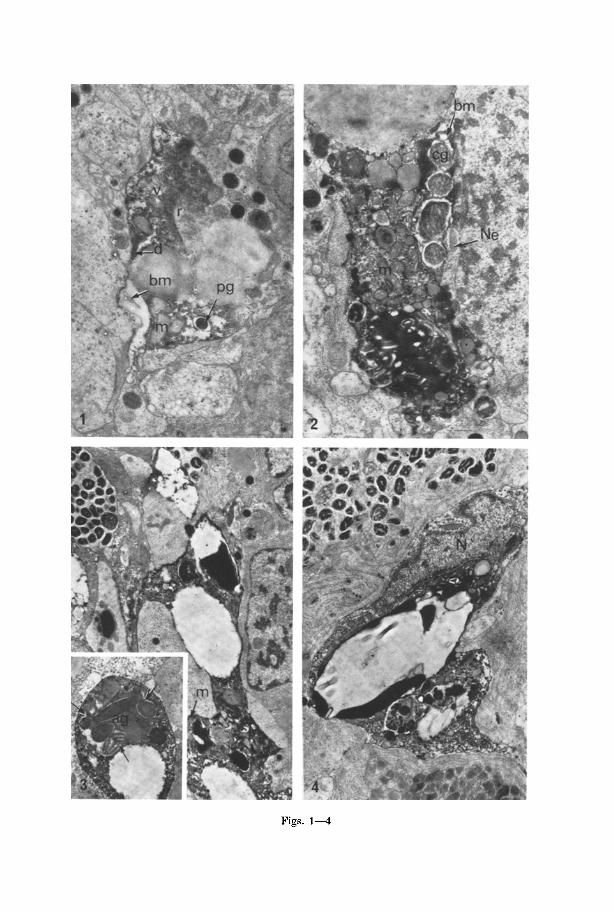
Figs. 1--4

Cell Deletion in Planarian 267
in 3% cacodylate buffered glutaraldehyde pH 7.2. They were then rinsed in the 0.2 M buffer overnight before post-fixing in buffered osmium tetroxide (Millonig, 1961) for 3 hours at 0 to 4~ The material was dehydrated in a series of alcohols and routinely embedded in Araldite. Ultrathin sections were cut with an LKB 111 ultramicrotome and these were mounted on carbon coated copper grids. Most of the sections were stained with uranyl acetate (Gibbons and Grimstone, 1960) and lead citrate (Reynolds, 1963). Some were stained in Potassium permanganate (Lawn, 1980) before treating with lead citrate. They were examined in an AEI EM 6 electron microscope operating at 80 KV.
Acid phosphatase activity was demonstrated as described by Bowen et al., (1974), tissues being incubated for 45-min at 37~ in a Gomori medium containing 0.01 M sodium-fl-glycero- phosphate (Sigma) as substrate and 2.5 mM lead nitrate made up in 0.1 M acetate buffer pH 4.0. p-Nitrophenyl phosphate was also employed as substrate as described by Ryder and Bowen (1974). In this case the incubating medium contained 3.8 mM substrate in 0.i M acetate buffer at pH 5.0 and 2.64 mM lead acetate as coupler. "No-substrate" and "sodium fluoride" controls were also performed. Tissues incubated in both media were washed in 0.2 M cacodylate buffer pH 7.2 before fixing in Millonigs osmium tetroxide for 1 hr at 0~]:~ C. After subsequent treatment for electron microscopy in a routine manner, sections were examined uncounterstained.
Results
Cell Autolysis and Deletion. Throughout the pa renchyma of the adul t planarian, whole cells can be seen exhibi t ing various degrees of lysis (Figs. 1 to 6). These can be observed next to perfectly normal well fixed tissue. Characteristic elements of specific cell types can often be recognised in these areas of lysis and a survey of the whole animal demonstrates tha t m a n y parenchymal cell types appear to be involved in this process. Numerous profiles indicate cells, and prolongations of them, in various degrees of breakdown, ranging from an apparent increase in cytoplasmic vesiculat ion and electron density, to a complete breakdown of the subcellular framework (compare Figs. 1 and 6). The peripheral areas of the cell appear to be degraded first and a high concentra t ion of ribosomes may often be seen at the boundary between the area of degradat ion and the more normal looking cytoplasm (Figs. 1, 5). The whole cell, however, including the nucleus (Fig. 4) is eventua l ly destroyed and deleted. In te rmedia te stages in cellular breakdown can be seen in Figs. 2-5. The final appearance of a lysed area generally takes the form of a space lined with highly electron dense lipid-like material and associated broken membranes and myelin-like whirls (Fig. 6).
Fig. 1. Part of a pigment cell. A dense concentration of ribosomes (r) can be seen bordering a lysed area of the cell. The area is characterised by vacuities (v), dense material (d) and broken membranes (bm). Mitochondria (m) and pigment granules (P9) can also be seen.
Note surrounding cells show "normal" well fixed profiles. • 14400 Fig. 2. Part of a cyanophil cell undergoing lysis. Characteristic reticular eyanophil granules (c9) can be seen among disintegrating mitochondria (m), broken membranes (bin) and dense
material. Part of a neoblast (Ne) can be seen next to the area of lysis. • 14500 Fig. 3. An extensive area of lysis in the parenchyma. Mitochondria (m) are still recognisable, otherwise the cell appears to be in an advanced state of decay and contains few identifiable components. Surrounding cells appear intact. • 6000. Inset: Part of a lysed acidophil cell in an advanced state of disintegration. Acidophil granules (ag) are still recognisable. Some appear
to be fused together (arrows) within a vacuole. • 4260 Fig. 4. The nucleus (N) of a lysed cell, surrounded by disintegrating cytoplasm, x8000
18b Cell Tiss. Res.. Vol. 154
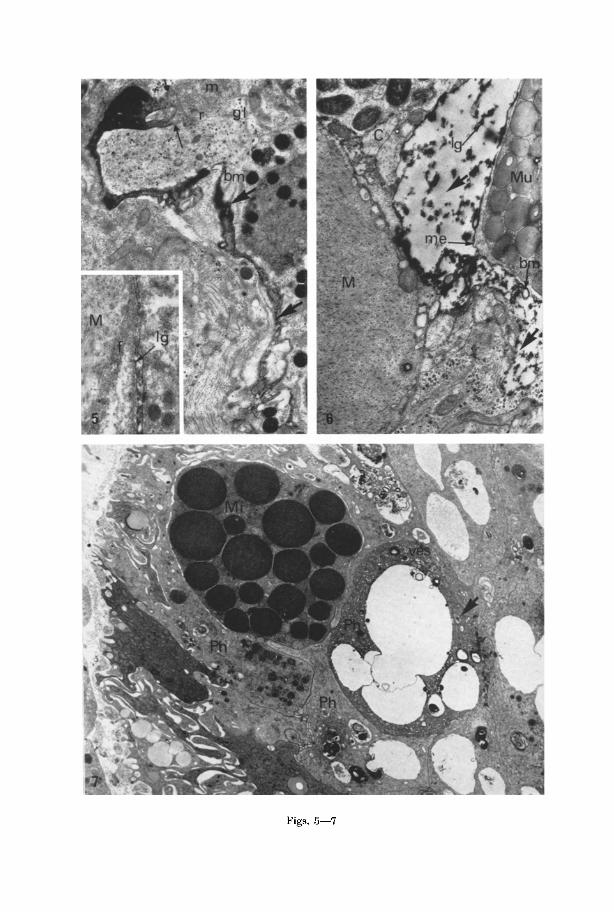
Figs. 5--7

Cell Deletion in Planarian 269
In the intermediate stages of breakdown elements of gland and pigment cells are easily identifiable, for example characteristic profiles of acidophil, cyanophil, mucous and pigment granules can be seen along with mitochondria and disinte- grating cndoplasmic ret iculum (Figs. 1-5). The areas of lysis are sharply deliniated and separated from surrounding intact cells by a distinct membrane which is coated on the lysing side with dense material (Fig. 6). Lysed areas are often ex- tended into thin prolongations (Fig. 5) again containing dense, frequently globular material (inset Fig. 5).
I n the gut, lysing cells appear to be more densely stained than normal cells (Fig. 7). Close examinat ion of such cells reveals tha t the endoplasmic reticulum is vesiculated. Broken membranes can also be seen as fur ther morphological evi- dence of lysis.
Taken together the morphological evidence indicates tha t a selective cell death involving a process of autolysis occurs in the normal animal.
Localization o /Ac id Phosphatase. The localization of acid phosphatase in non- lysing cells is described by Ryder and Bowen (1974). I n such cells the enzyme is generally membrane bound and associated with lysosome-like vacuoles. Although vacuolar act ivi ty often persists in lysing cells (Fig. 9), a more extensive cyto- plasmic distribution of the enzyme is usually apparent (Figs. 8-11). Interest ingly the cytoplasmic localization of the enzyme is extra-cisternal (Figs. 8, 9). A build up of act ivi ty in the Golgi apparatus of acidophil cells may be observed during the early stages of autolysis (Fig. 8) both the Golgi saccules and vacuoles showing a positive reaction. The Golgi appara tus of this part icular cell type is normal ly negative.
Cytoplasmic acid phosphatase act ivi ty is associated with the lysis of gastro- dermal phagocyt ic cells (Fig. 10). During cell lysis hydrolase act ivi ty initially appears peripherally near the plasma membrane. The distribution of the enzyme becomes more general as lysis proceeds.
All the lysing gland cells exhibit acid phosphatase activity. The distribution of enzyme act ivi ty in a lysing aeidophil gland cell is illustrated in Fig. 8. A more
Fig. 5. Prolongation of a cell (large arrows) in a lysed condition. Myelin-like whirls and broken membranes (bm) can be seen in the lysed area. The cytoplasm towards one end (small arrow) of the prolongation appears to be substantially normal containing ribosomes (r), mitochondria (m) and glycogen (gl). • 10500. Inset: The finer ending of a prolongation from a lysed cell showing the globular distribution of dense lipid-like material (lg) on the enclosing membrane. The prolongation ends in contact with the intercellular fibrous matrix (/) surrounding the
muscle cell (M). • 24000
Fig. 6. The final stage of cellular lysis. Cytoplasmic components have disappeared and the area previously occupied by the cell is largely a space or vacuity (large arrows). Dense globules of lipid-like material (lg) can be seen and this material also coats a surrounding membrane (me). Remains of broken membranes (bm) can also be seen within the lysed area. Surrounding cells, in this case, cyanophil (C), muscle (M) and mucous (Mu) are perfectly intact. • 14100
Fig. 7. Part of the gastrodermis showing a Minotian gland cell (Mi) and phagocytic cells (Ph). One of the phagocytic cells (arrow) appears darker. The cytoplasm of this cell contains a highly vesiculated endoplasmic reticulum (ges) and other signs of lysis in the form of
broken membranes. •
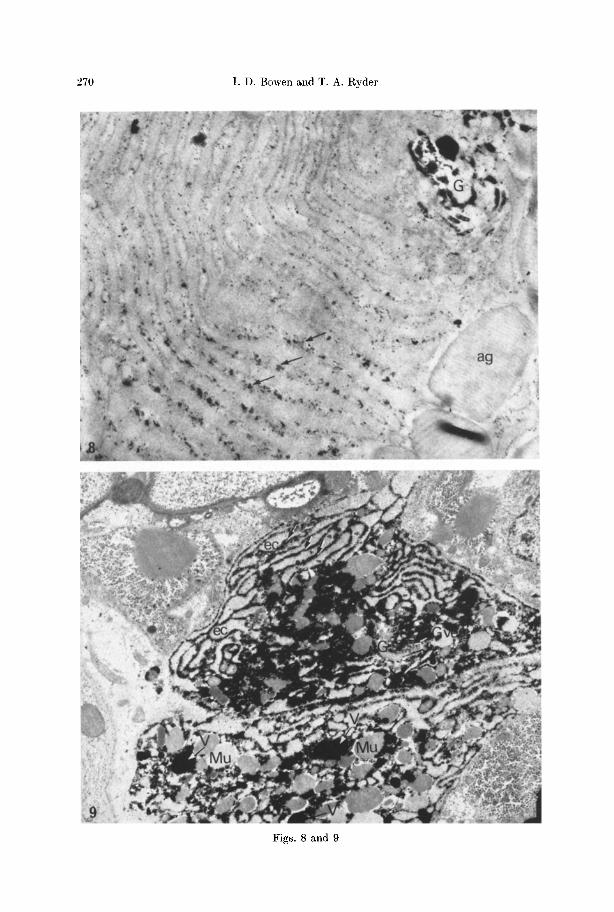
270 I . D . Bowen and T. A. Ryder
Figs. 8 and 9

Cell Deletion in Planarian 271
advanced state of lysis in the forming part of a mucous cell is shown in Fig. 9. A similar distribution of enzyme has been observed in lysing cyanophil and pig- ment cells. Fig. 11 depicts autolysis in the pharyngeal cavity lining. In all cases morphological evidence of lysis, in the form of endoplasmic vesiculation, broken organelles and membranes can be seen in the cytoplasm as well as cytochemical evidence of hydrolytic activity.
Both p-nitrophenyl phosphatase and fl-glycerophosphatase activities have been demonstrated, p-Nitrophenyl phosphatase activity appears to be a more useful marker for lysis in that it produced a greater deposition of reaction product at a larger number of sites. "No-substrate" and "sodium fluoride" controls were negative.
Discussion
Klima (1961) in his review of Triclad fine structure, does not mention the areas of lysis described here. Pedersen (1961), however, draws attention to the occurrence of dense intercellular material in the parenchyma and comments that these spaces are somewhat widened in certain areas. Some of these areas may represent the finer prolongations of lysing cells such as those illustrated in Fig. 5. Because they contain specifically recognizable organelles, lysing gut, pharyngeal cavity lining, gland and pigment cells have been conclusively identified. Whether the autolytic phenomenon applies to other cell types less easily identifiable in the disintegrated state, remains to be seen. Ramifications of lysed cells have been observed adjoining most cell types, including muscle; although no such areas have been observed in the brain and nerve cord.
If it is assumed that evidence of progressive morphological disintegration accompanied by increasing free acid hydrolase activity may be taken as an indi- cation of eventual cell death, then it may be concluded tha t a mechanism of selec- tive cell death operates in the normal adult planarian. A similar situation has been observed in the gut of the slug, Arion hortensis (Bowen and Davies, 1971). Changes in the ribosomal population also seem to occur. Such a phenomenon has been observed elsewhere, under conditions involving cellular damage (Young, 1970).
Autophagic lysosomes have been observed in most of the gland cells (Ryder and Bowen, 1974). There is no direct evidence, however, to indicate that autophagy is linked with the cellular lysis described here. Indeed, vacuolar acid phosphatase activity persists during autolysis. The earliest indication of autolysis appears to be a progressive vesiculation of the endoplasmic reticulum. This is generally pre- ceded by extra-cisternal acid phosphatase activity. The origin of the extra-
Fig. 8. p-Nitrophenyl phosph~tase activity in part of an acidophil gland cell. Reaction product in the form of lead crystals can be seen in the Golgi apparatus (G) and outside the cisternae (arrows) of the rough endoplasmic reticulum. The acidophil granules (ag) can also
be seen. • 25000
Fig. 9. A lysing mucous cell. Reaction product denoting intense p-Nitrophenyl phosphatase activity can be seen within vacuoles (V) in the cytoplasm and particularly outside the endo- plasmie cisternae (arrows). The endoplasmic cisternae (ec) themselves are negative as are the mucous droplets (Mu). The Golgi apparatus (G) in this case is also largely negative, though
there is slight vesicular staining (G ves).• 8000
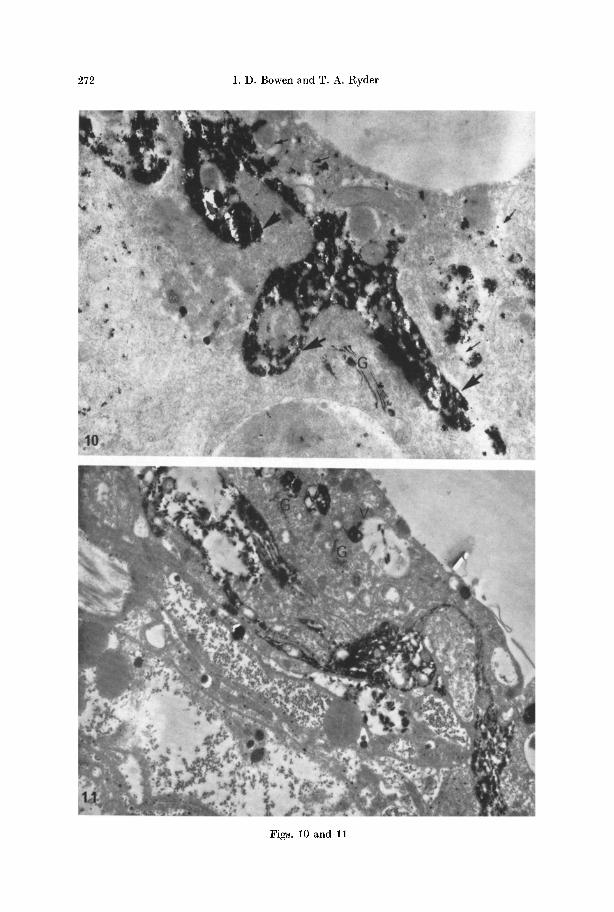
272 I . D . Bowen and T. A. Ryder
Figs. 10 and 11

Cell Deletion in Planarian 273
Fig. 10. Part of a lysing gastrodermal phagocyte. Note that reaction product indicating hydrolytic (p-nitrophenyl phosphatase) activity is localized peripherally in the cytoplasm (large arrows). Broken membranes and gaps c~n be seen in the cytoplasm (small arrows). The adjacent non-lysing cell is negative, except for some activity of the Golgi apparatus
(a). • 20000 Fig. 11. Reaction product released by p-nitrophenyl phosphatase activity delineating the tortous profile of a lysing cell in the cavity lining epithelium of the pharynx. An adjacent non-lysing epithelial cell shows evidence of vacuolar (V) activity and activity of the Golgi
apparatus (G). • 6000
cisternal activity remains obscure. According to the GERL hypothesis (Novikoff et al., 1964), enzyme might be expected within the cisternal space, not outside it, however, during autolysis changes in the permeability of cytomembranes or even their breakdown might account for the characteristic extra-cisternal localiza- tion obtained. Alternatively, the extra-cisternal acid phosphatase may represent enzyme, newly formed at the free ribosomes which occur outside the cisternae. In the case of the acidophil cell, Golgi apparatus associated acid phosphatase also appears as a prelude to autolysis. De Duve (1959), Brandes et al. (1965), Schin and Clever (1965) and Lockshin and Williams (1965b), all implicate lysosomes in the release of cytoplasmic acid phosphatase. Ballard and Holt (1968) and Weber (1964 on the other hand present evidence of cell lysis and death occurring without the initial involvement of lysosomal hydrolase.
Cell lysis and deletion in the normal adult planarian means that the majority of cells in the organism are undergoing a process of renewal, as defined by Leblond and Walker (1956). The neoblasts in the normal fully grown animal are undergoing a fairly high rate of mitosis (Bowen, Ryder and Dark, in preparation). This cell production must be balanced by cell loss (since the adult is not growing). Unlike the mammalian cell populations reviewed by Leblond and Walker, the major part of this loss would appear to take place by means of cell autolysis. Kerr et al. (1972, 1974) have shown that controlled cell lysis and deletion does occur in vertebrates. Indeed, this type of controlled cell death or apoptosis may prove to be a pheno- menon of general significance throughout the animal kingdom.
The factors which cause individual cells to lyse are unknown; perhaps each cell has a genetically programmed life span which is fairly short when compared with the animal as a whole. The lysis of entire cells followed by the possible re-cycling of cellular material may, like autophagy be a crucial factor in the ability of these animals to survive starvation.
References Ballard, J. K., Holt, S. J.: Cytological and eytochemical studies on cell death and digestion
in the foetal rat foot: the role of macrophages and hydrolytic enzymes. J. Cell Sci. 8, 245-262 (1968)
Bowen, I. D., Davies, P.: The fine structural distribution of acid phosphatase in the digestive gland of Arion hortensis (Fer.). Protoplasma 78 (1971)
Bowen, I. D., Ryder, T.A.: The fine structure of the planarian Polycelis tenuis Iijima. 1. The pharynx. Protoplasma 78, 223-241 (1973)
Bowen, I. D., Ryder, T. A., Thompson, J. A.: The fine structure of the planarian Polycelis tenuis Iijima. 11. The intestine and gastrodermal phagocytosis. Protoplasma 79, 1-17 (1974)

274 I . D . Bowen and T. A. Ryder
Brandes, D., Bertini, F., Smith, E. W.: Role of lysosomes in cellular lyric processes. 11. Cell death during holocrine secretion in sebaceous glands. Exp. molec. Path. 4, 245-265 (1965)
Duve, C. de: Lysosomes, a new group of cytoplasmic particles. In: Subcellular particles (ed. T. Hayashi), p. 128-159. New York: Ronald Press 1959
Fox, H.: Muscle degeneration in the tail of Rana temporaria at metamorphic climax; An electron microscopic study. Arch. Biol. 83, 407417 (1972)
Gibbons, I., Grimstone, A.V.: On flagellar structures in certain flagellates. J. biophys. biochem. Cytol. 7, 697-716, (1970)
Glucksmann, A.: Cell deaths in normal vertebrate ontogeny. Biol. Rev. 26, 59-86 (1951) Ishii, S.: Electron microscopic observations on the planarian tissues. 1. A survey of the
pharynx. Fukushima J. reed. Sci. 9-10, 51-73 (1962) Ishii, S.: The ultrastructure of the outer epithelium of the planarian pharnyx. Fukushima
J. med. Sci. l l , 109-125 (1964) Ishii, S.: Electron microscopic observations on the planarian tissues. 11. The intestine.
Fukushima J. med. Sci. 12, 67-87 (1965) Ishii, S.: The ultrastructure of the insunk epithelium lining the planarian pharyngeal cavity.
J. Ultrastruct. Res. 14, 345-355 (1966) Kerr, J .F.R. , Harmon, B., Searle, J . : Cell deletion in tadpole tail. J. Cell Sci. 14, 571-585 (1974) Kerr, J .F.R. , Wyllie, A. E., Currie, A. R.: Apoptosis: a basic biological phenomenon with
wide-ranging implications in tissue kinetics. Brit. J. Cancer 26, 239-257 (1973) Klima, J . : Elektronenmikroskopische Studien fiber die Feinstruktur der Tricladen (Turbel-
laria). Protoplasma 54, 101-162 (1961) Lawn, A. M.: The use of potassium permanganate as an electron dense stain for section of
tissue embedded in epoxy resin. J. biophys, biochem. Cytol. 7, 197 (1960) Leblond, C. P., Walker, B. E.: Renewal of cell populations. Physiol Rev. 36, 255-275 (1965) Lockshin, R. A., Williams, C. M.: Programmed cell death - 1. Cytology of degeneration in
the intersegumntal muscles of the pernyi silkmoth. J. Ins. Physiol. l l , 123-133 (1965a) Lockshin, R. A., Williams, C. M.: Programmed cell death - V. Cytolytic enzymes in relation
to the breakdown of the intersegmental muscles of silkmoths. J. Ins. Physiol. l l , 831-844 (19655)
Looss, A.: Ober Degenerationserscheinungen im Tierreich, besonders fiber die Reduktion des Froschlarvenschwanzes und die im Verlauf desselben auftretenden histolytischen Prozesse. Preisschr. jablonowsk. Ges. 10 (1889)
Novikoff, A. B., Essner, E., Quintana, N.: The Golgi apparatus and lysosomes. Fed. Proc. 23, 1010 1022 (1964)
Pedersen, K. J . : Some features of the fine structure and histochemistry of planarian sub- epidermal gland cells. Z. Zellforseh. 50, 121-142 (1959a)
Pedersen, K. J . : Cytological studies on the planarian neoblast. Z. Zellforsch. 50, 799 817 (1959b)
Pedersen, K. J . : Studies on the nature of the planarian connective tissue. Z. Zel]forsch. 53, 569-608 (1961)
Pedersen, K. J . : Slime secreting cells of planarians. Ann. N. Y. Acad. Sci. 106, 424-442 (1968) Reynolds, E. S.: The use of lead nitrate at high pH as an electron opaque stain in electron
microscopy. J. Cell Biol. 17, 208-212 (1963) Ryder, T.A. , Bowen, I. D.: The fine structural localization of acid phosphatase activity in
Polycelis tenuis (Iijima). Protoplasma, in press (1974) Schin, K. S., Clever, U.: Lysosomal and free acid phosphatase in salivary glands of Chiro-
riotous tetans. Science (N. Y.) 150, 1053 (1965) Sheldrake, A. R.: The ageing, growth and death of cells. Nature (Lond.) 250, 381-385 (1974) Skaer, R. J.: Some aspects of the cytology of Polycelis nigra. Quart. J. micr. Sci. 102, 295-318
(1961) TSrSk, L. J., RShlich, P.: Electronmikrozkopos vizsgalatok planariak eosinofil miriqysjtjein.
Biol. KSzl. 8, 117-124 (1960) Weber, R.: Ultrastructural changes in regressing tail muscles if Xenopus larvae at meta-
morphosis. J . Cell Biol. 22, 481487 (1964) Young, D., Ashhurst, D. E., Cohen, M. J . : The injury response of the neurones of Periplaneta
americana. Tiss. Cell. 2, 387-398 (1970)





























