Embed Size (px)
Citation preview

THE JOURNAL oe BIOLOGKAL CHEMISTRY Vol. 247, No. 19, Issue of October 10, pp. 6167-6175. 1972
Printed in U.S.A.
Calcium-mediated Interactions between the Antidiuretic
Hormone and Renal Plasma Membranes*
(Received for publication, April 25, 1972)
BENEDICT J. CAMPBELL, GERALD WOODWARD,$. AND VICTORIA BORBERG
From the Department of Biochemistry, University of Missouri, Columbia, Missouri 65~01
SUMMARY
The antidiuretic hormone-stimulated adenylate cyclase system has been characterized in a purified preparation of porcine renal plasma membranes. Adenylate cyclase activ- ity was linear over a plasma membrane protein concentration from 30 to 300 pg, and maximal enzyme activity occurred at approximately 7 x 1OV M ATP. Activation of the system by sodium fluoride was about IO-fold and was produced by 10 to 12 InM sodium fluoride. The adenylate cyclase was stimulated approximately j-fold by the antidiuretic hormone. One-half maximal stimulation by hormone was measured at 2.2 x lo-* M antidiuretic hormone.
Calcium exhibited an inhibitory effect upon antidiuretic hormone-stimulated adenylate cyclase at concentrations of the metal ion above 10m6 M, but it produced little or no inhibi- tion in the sodium fluoride-stimulated system. The chelat- ing agent, ethylene glycol-his@-aminoethyl ether)-N,N’- tetraacetic acid, at a concentration of 1 m&r, completely inhibited antidiuretic hormone-stimulated adenyl cyclase. The activity of the chelator-inhibited enzyme was completely restored by increasing concentrations of calcium.
Binding of tritiated antidiuretic hormone to the renal plasma membranes was measured by centrifugation tech- niques and by filtration through Millipore filters. Both methods demonstrated that binding of the hormone to the membranes occurred at a level of approximately 0.5 pmole of antidiuretic hormone per mg of protein when the hormone was added at a concentration that produced half-maximal stimulation of adenylate cyclase. The rates of association and dissociation for the interaction between the antidiuretic hormone and renal plasma membranes were measured. Binding equilibrium was reached after approximately 10 min at 37”. Dissociation of tritiated bound hormone was shown to be 100% complete after approximately 30 min. Graphical analysis of the time course of dissociation demonstrated that the kinetics of the reaction follows a simple first order rate equation. Neither adrenocorticotropic hormone nor calcitonin exchanged with antidiuretic hormone at its binding
* This work was supported by Grant AM-04660 from National Institutes of Health. A preliminary account of part of this work was presented at the 62nd annual meeting of the American Society of Biological Chemists, San Francisco, June 1971.
$ Part of the studies reported in this paper were taken from a thesis by Gerald Woodward submitted to the University of Mis- souri at Columbia in partial fulfillment of the requirements for the degree of Doctor of Philosophy.
site, indicating that the receptor sites exhibit a specificity for the antidiuretic hormone.
Calcium ion concentrations in the millimolar range in- hibited the rate and level of binding of ADH to renal plasma membranes with complete inhibition reached at 10 mM calcium. However, ethylene glycol-bis(&aminoethyl ether)- N,N’-tetraacetic acid at the same concentration at which it completely inhibited adenylate cyclase activity had no effect upon the binding of antidiuretic hormone to the membranes. These results suggest that calcium plays multiple regulatory roles with milhmolar concentrations inhibiting binding to membrane receptor sites and lower calcium concentrations mediating at a locus within the membrane matrix between the receptor site and the site occupied by adenylate cyclase.
An interdependency between the effects of the antidiuretic hormone and calcium has been observed in a variety of biological systems. The response of the hormone in increasing the water permeability of cell membranes in the toad bladder (1, 2) and in the kidney of Necturus (3) has been reduced by increasing con centrations of calcium. Furthermore, it has been reported that the antidiuretic hormone causes an increased rate of excretion of calcium during its action on the intact mammalian kidney (4-6).
These effects of calcium and antidiuretic hormone have in certain cases been observed at the enzyme level in adenylate cyclase systems. Hynie and Sharp (7), using a toad bladder mucosal cell homogenate, showed that 1 mM calcium completely inhibited vasopressin-stimulated adenylate cyclase activity but only slightly depressed the sodium fluoride-stimulated adenylate cyclase. More recently, Marumo and Edelman (8) reported that calcium inhibits the vasopressin-stimulated adenylate cyclase in crude homogenates of golden hamster kidney. Sepa- ration of the effect of the antidiuretic effect from that produced by the parathyroid hormone in the rat kidney was achieved bJ Melson et al. (9), who demonstrated that the antidiuretic hor- mone activated adenylate cyclase in crude homogenates of the renal medulla but not in homogenates prepared from the renal cortex.
In the present investigation a plasma membrane preparation has been isolated from the porcine renal medulla region. The effects of calcium and the antidiuretic hormone upon the mem- brane-bound adenylate cyclase system have been measured.
6167
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from

6168
By using tritium-labeled antidiuretic hormone it was possible to determine the degree of binding of the hormone to membrane preparations, and to observe the effects of calcium upon the properties of the hormone-receptor interaction.
EXPERIMENTAL PROCEDURES
Materials-An acetone powder of whole hog pituitary glands from which corticotropin had been removed by treatment with oxycellulose was supplied by Dr. J. B. Lesh of Armour Pharma- ceutical Company. Synthetic salmon calcitonin (-98% pure, 5030 units’ per mg) and porcine adrenocorticotropin hormone purified by CM-cellulose chromatography (150 units per mg) were provided by Dr. Robert Schlueter of Armour Pharma- ceutical Company. Crystalline disodium ATP, cyclic AMP, creatine phosphate, creatine phosphokinase (salt-free powder, 25 to 30 units per mg), and chromatographic neutral alumina (type WN-3) were obtained from Sigma Chemical Company. Trisodium [LU-~~P]ATP (0.6 to 1.5 Ci per mmole) and 3H-labeled cyclic AMP (0.3 to 1.0 Ci per mmole) were purchased from International Chemical and Nuclear Corporation.
Methods
Puri$calion. of Tritiated Antidiuretic Hormolze-Lysine vaso - pressin (antidiuretic hormone) was purified from the acetone powder of whole hog pituitary glands by extraction with glacial acetic acid, removal of lipid components by extraction with diethyl ether, CM-cellulose chromatography, and Sephadex G-25 gel filtration as previously described by Campbell et al. (10). The purified hormone was then iodinated at the tyrosine ring position according to the method of Lee and Wiseman (11). Substitution of tritium for iodine was achieved by dissolving approximately 200 mg of the iodinated and lyophilized reaction mixture in 1.0 ml of dimethyl formamide containing 10 Ci of tritium gas, 25 mg of palladium black, and 0.025 mg of triethyl- amine. The reaction mixture was stirred for 4 hours at room temperature.
The resulting tritiated lysine vasopressin was diluted with carrier hormone and purified using CM-cellulose ion exchange chromatography and Sephadex G-25 gel filtration (10). The specific activity of the labeled product was monitored after each purification step. The amino acid composition of the final product was determined using the Spinco amino acid analyzer and the method of Moore and Stein (12). Paper electrophoresis was carried out using the E-C Apparatus Co. model 1331 equip- ment and pyridine-acetic acid buffer at pH 6.2. The distribu- tion of radioactivity along the path of migration was established by cutting the paper into sections (1 x 3 cm) and counting the sections in Bray’s solution (13) using a Tri-Carb scintillation counter (Packard Instruments, Inc.). The biological activity of the tritiated hormone was measured using the toad bladder assay of Bentley (1).
Preparation of Renal Plasma Membranes-Hog kidneys were excised from freshly slaughtered animals within 1 hour after death. All subsequent preparative steps were carried out at O-2”. The medulla was separated from cortex material follow- ing a midsagittal cut dividing the kidney into equal halves.
1 Unitage is based on the l-hour hypocalcemic activity in the rat of the test preparation administered subcutaneously, compared with the Medical Research Council (MRC) porcine calcitonin Standard B.
*The abbreviations used are: cyclic AMP, cyclic adenosine 3’,5’-monophosphate; ADH, antidiuretic hormone or lysine vaso- pressin; ACTH, porcine adrenocorticotropic hormone; EGTA, ethylene glycol his@-aminoethyl ether) N,N’-tetraacetic acid.
Between 100 and 200 g of medulla tissue were collected for each preparation, and care was taken not to include any of the cortex region. The medulla tissue was placed in a homogenizing mix- ture of 0.25 M sucrose and 1 mM EDTA in the ratio of 1 part tissue to 4 parts sucrose-EDTA (w/v). This suspension was placed in heavy glass test tubes and homogenized for 30 set using a Polytron homogenizer (Kinematica, Gmbh., Luzern- Schweiz, type PT-SO-OD). The highly minced material was then homogenized with six complete strokes using a Potter- Elvehjem Teflon-glass homogenizer (Arthur I-1. Thomas Co., 4288-B, C). The completion of this step yielded a crude ho- mogenate which was filtered through one layer of cheesecloth. The residue on the cheesecloth was rinsed one time with 1 volume of 0.25 M sucrose-l mM EDTA. The residue was discarded and differential centrifugation of the filtrate was performed according to a procedure similar to that reported by Fitzpatrick et al. (14).
All centrifugations were carried out in a Sorvall RC2-B refrigerated centrifuge using an SS-34 rotor. The filtrate was centrifuged at 1,475 x gmax for 10 min. The supernatant was discarded, and the pellet was suspended in 1.5 ml of 2.0 M sucrose per g of starting material using the Teflon-glass homogenizer. This suspension was centrifuged at 13,000 x gmrx for 10 min. The supernatant from this second centrifugation was saved and the pellet discarded. The supernatant was diluted to 0.25 M
sucrose with 7 volumes of cold deionized water and centrifuged a third time at 35,000 x gmax for 15 min. The resultant pellet from this step was composed of a dark brown bottom layer and a fluffy pink top layer. The top layer was collected by adding 3 ml of 0.05 M Tris-HCl buffer at pH 7.6 to the centrifuge tube and swirling in a Vortex-Genie mixer at low speed. The fluffy pink upper layer distributed in the buffer and the brown layer remained at the bottom of the tube. The swirled upper frac- tions were pooled and washed twice by centrifuging at 35,000 X
gmax and resuspending in the Tris-HCI buffer at a ratio of 1 ml of buffer to 10 g of starting medulla tissue. This suspension will be referred to as renal plasma membranes in the subsequent investigations.
When plasma membranes were to be stored for later use, the 2 M sucrose supernatants from the second centrifugation step were collected, adjusted to 25 InM sodium fluoride, and frozen at -80”. The procedure described above for processing the 2 M sucrose supernatant was employed when these membranes were needed.
Electron Microscopy-Renal plasma membrane suspensions were centrifuged at 35,000 x g,%, for 15 min. The pellet was resuspended in 2’% glutaraldehyde in 0.05 M sodium cacodylate buffer at pH 7.2 and washed twice with this buffer. The mem- branes were then postfixed for 2 hours in 1% osmium tetroxide in Millonig’s phosphate buffer at pH 7.3. Dehydration was carried out using graded concentrations of ethanol, and the pellets were embedded in Epon 812. Sections were taken using a MT-2 Porter-Blum ultramicrotome. The ultrathin sections (600 to 900 A) were stained for 10 min each in a solution of Reynold’s lead citrate and uranyl acetate, and then examined using a RCA EMU-4 electron microscope. An accelerating voltage of 50 kv was employed.
Sodium-Potassium ATPase Assay-The membrane ATPase activity was measured as the difference between magnesium- dependent ATPase and the total ATPase activity obtained upon addition of NaCl and KCI. In a typical assay for magnesium- dependent ATPase, an aliquot of the membrane preparation was incubated with shaking for 20 min at 37” in a solution of 4 rnM MgC12, 4 mM ATP, and 100 pmoles of Tris-HCl buffer at pH 7.4.
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from

6169
The total volume of the reaction mixture was 3 ml. To assay the total magnesium-sodium-potassium-dependent ATPase ac- tivity the reaction mixture was adjusted to a final concentration of 67 InM NaCl and 3.3 mM KCl. ATPase activity was stopped by the addition of 3 ml of 5% trichloroacetic acid. The mixtures were centrifuged (International clinical centrifuge for 3 min), and aliquots were removed for analysis of inorganic phosphate by the method of Lowry and Lopez (15) and for analysis of protein by the Lowry et al. (16) method for insoluble protein. The protein standard employed was three times crystallized bovine serum albumin.
Adenylate Cyclase Assay-The adenylate cyclase activity of the renal plasma membranes was determined essentially by the method of White and Zenser (17, 18). This technique measures the conversion of [cx-~~P]ATP to cyclic 3’) 5’-AMP in the presence of an ATP-regenerating system. The standard assay mixture of 75 ~1 contained: 0.03$& bovine serum albumin; 25 mM creatine phosphate; 0.4 c(g of creatine phosphokinase; 6.67 InM MgCl2; 16.0 mM caffeine; 1.07 ITIM cyclic AMP; 50 mM Tris-HCl buffer at pH 7.6; 100 to 300 pg of plasma membrane protein; and 1.2 mM ATP. Sodium fluoride was present at 10 mM where indi- cated. The reaction was started by the addition of substrate, [c@P]ATP, to give 200,000 to 800,000 cpm per assay mixture, and incubation was for 10 min at 30”. The reaction was termi- nated by adding 2O;l of 0.1 M EDTA which contained 3,000 to 15,000 cpm of 3H-labeled cyclic AMP, followed by heating for 2 min at 90”. Then 1 ml of 0.05 M Tris-HCl buffer at pH 7.6 was added to each tube, and the tubes were centrifuged in the International centrifuge for 5 min.
The 3H- and 32P-labeled cyclic AMP was separated from other radioactive materials by chromatography on neutral alumina columns as described by White and Zenser (17). The labeled AMP was obtained in the 2nd and 3rd ml fractions of eluate from l-g alumina columns. Counting was performed as previ- ously described. The recovery of cyclic AMP from the reaction mixtures was approximately 75 to 80% and was determined for each experiment. All points are averages of duplicate or tripli- cate determinations.
Binding of Antidiuretic Hormone to Plasma Membranes-To measure the binding of the antidiuretic hormone to plasma membrane fractions, two independent techniques were employed. The first method involved the separation of membrane-bound tritiated hormone from unbound hormone by centrifugation. In these experiments the assay mixtures consisted of 2 ml of 0.05 M Tris-HCl buffer at pH 7.6 to which were added plasma membranes (1.0 to 5.0 mg of plasma membrane protein) and tritiated ADH at a final concentration from IO-” to lo-* M.
The mixtures were incubated for 1 min at 37”, chilled in an ice bath, and then immediately centrifuged at 35,000 x gmax for 15 min in a Sorvall RC-2B centrifuge. The pellet was washed and recentrifuged with Tris-HCI buffer at pH 7.6 until radio- activity above background no longer appeared in the supernatant (usually four times). The washed membrane pellets were taken up in 4 ml of Tris-HCI buffer at pH 7.6. Aliquots (1 ml) of this mixture were added to counting vials containing 1 ml of Bio- Solv BBS-3 (Beckman), and the capped vials heated for 4 hours at 80”. To each of the resulting solutions were added 10 ml of Bray’s solution, and the vials were counted as described above. The protein content of aliquots from each plasma membrane mixture was determined by the Lowry method (16) for insoluble proteins.
The second t,echnique used to measure AD11 binding to mem- branes employed filtration through cellulose acetate EAWP
Millipore filters. A typical assay mixture consisted of 1.40 ml of 0.05 M Tris-HCl buffer at pH 7.6 which contained 1.4 mg of plasma membrane protein and 2.2 x 10e8 M ADH, final concen- tration. The mixture was incubated at 37”, and at various times (30 set to 30 mm), 0.1~ml aliquots were withdrawn and filtered through the Millipore filters. The filters were washed with three lo-ml aliquots of the Tris-HCI buffer, placed in counting vials containing 10 ml of Bray’s solution, and counted as described above. Appropriate controls were run to measure binding of the labeled hormone to the Millipore filter. When dissociation of labeled hormone was measured the reaction mixtures were allowed to equilibrate for 20 min, and then un- labeled hormone (ADH, ACTH, calcitonin) was added to give a final total concentration of the hormone of 2.2 x lo-” i\~. Ali- quots were then removed at various times (2 min to 2 hours) for filtration, washing, and counting.
Because of the relatively low specific activity of the tritiated ADH, care was taken to establish the reliability of the binding data. Each binding determination in the centrifuge method and in the filtration method was counted for a 60.min rather than the normal 10.min period. Five independent experiments of each type were performed, and the results were averaged for comparison of the two methods. When binding was measured as a function of time, each point was established as the average of duplicate or triplicate measurements against controls with no plasma membranes present. Finally, each rate of binding ex- periment contained 11 different counting measurements per- formed in duplicate or triplicate.
RESULTS
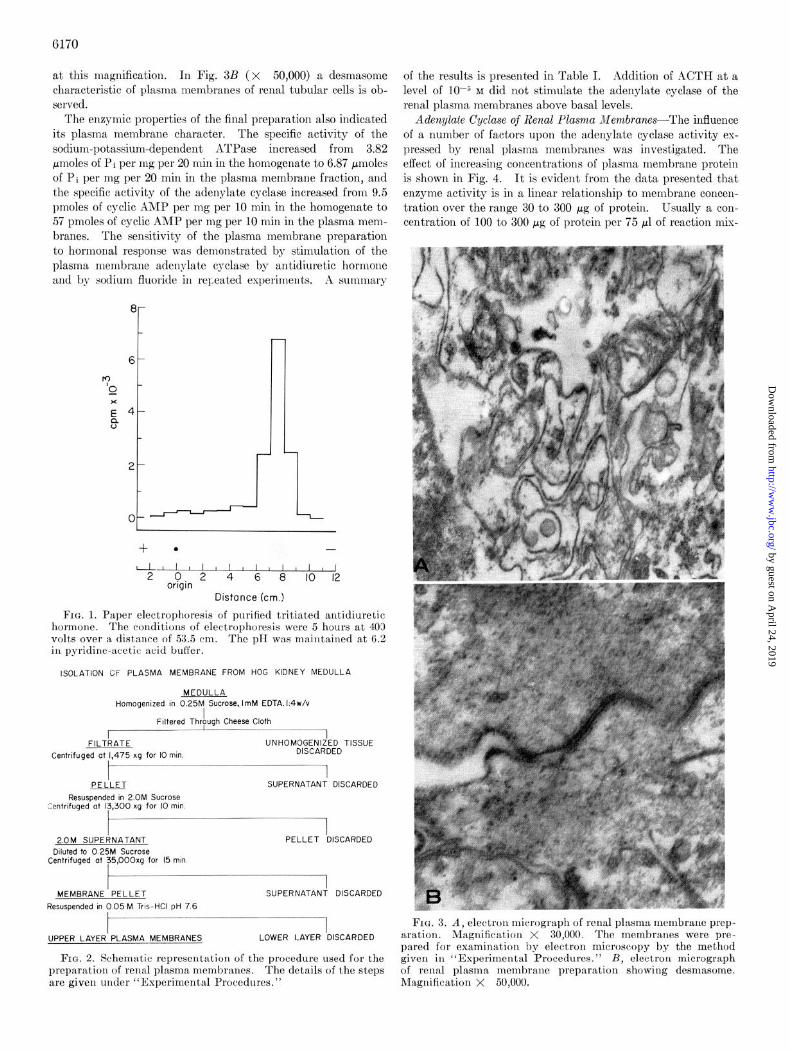
Purzlficafion of Tritiated Antidiuretic Hormone-In a typical iodination and tritiation experiment 68 mg of purified hormone were iodinated yielding 218 mg of lyophilized unresolved reaction mixture Following tritiation and isolation of the purified hor- mone, a final yield of 19 mg of labeled lysine vasopressin was ob- tained. The final product was taken up in 5 ml of deionized water and diluted with 20 mg of pure unlabeled ADH. The specific activity of various preparations was approximately 3.2 x lo6 cpm per pmole (3.61 PCi per pmole), and the specific activity remained constant through the final two purification steps. Amino acid analysis of the purified hormone indicated that the product was >99% pure according to its amino acid composition. The results of paper electrophoresis of the labeled hormone are shown in Fig. 1. Only one ninhydrin-positive spot was observed, and over 90y0 of the radioactivity was associated with that spot. Nitroprusside reacted with the radioactive spot only following treatment with cyanide, indicating that the disulfide bond of the hormone was intact. The tritiated hor- mone promoted water loss by the toad bladder in concentrations less than 1 pM.
Purifiealion and Characterization of Renal Plasma iiiembranes- The procedure for the isolation of renal plasma membranes is summarized in Fig. 2. In a typical experiment approximately 4 mg of purified renal plasma membranes were obtained from 1 g of medulla tissue. The first pellet examined by light microscopy was observed to contain cell debris, nuclei, and membranes. 111
the membrane pellet no cont,aminating nuclei were observed by light microscopy. Electron micrographs of the purified mem- brane preparation are presented in Fig. 3. In Fig. 3A (X 30,000) the membrane preparation appears to be composed of membrane components, usually in pairs in the presence of un- identified vesicle structure. Sonie contamination with intact and disrupted mit~ochondria was observed in micrographs taken
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from
6170
at this magnification. In Fig. 3B (X 50,000) a desmasome of the results is presented in Table I. Addition of ACTH at a characteristic of plasma membranes of renal tubular cells is ob- level of 10e5 M did not stimulate the adenylate cyclase of the served. renal plasma membranes above basal levels.
The enzymic properties of the final preparation also indicated its plasma membrane character. The specific activity of the sodium-potassium-dependent ATPase increased from 3.82 pmoles of P r per mg per 20 min in the homogenate to 6.87 hmoles of Pi per mg per 20 min in the plasma membrane fraction, and the specific activity of the adenylate cyclase increased from 9.5 pmoles of cyclic AMP per mg per 10 min in the homogenate to 57 pmoles of cyclic AMP per mg per 10 min in the plasma mem- branes. The sensitivity of the plasma membrane preparation to hormonal response was demonstrated by stimulation of the plasma membrane adenylate cyclase by antidiuretic hormone and by sodium fluoride in repeated experiments. A summary
Adenylate Cyclase of Renal Plasma Membranes-The influence of a number of fact,ors upon the adenylate cyclase activity ex- pressed by renal plasma membranes was investigated. The effect of increasing concentrations of plasma membrane protein is shown in Fig. 4. It is evident from the data presented that enzyme activity is in a linear relationship to membrane concen- tration over the range 30 to 300 pg of protein. Usually a con- centration of 100 to 300 pg of protein per 75 ~1 of reaction mix-
8 r
+ l -
I I I I I , , , , , , , ,
0 2 4 6 8 IO 12 origin
Distance km.!
FIG. 1. Paper electrophoresis of purified tritiated antidiuretic hormone. The conditions of electrophoresis were 5 hours at 403 volts over a distance of 53.5 cm. The pH was maintained at 6.2 in pyridine-acetic acid buffer.
ISOLATION CF PLASMA MEMBRANE FROM HOG KIDNEY MEDULLA
MEDULLA Homogenized in 0.25M Sucrose, ImM EDTA. I:4w/v
Filtered Thrbugh Cheese Cloth
I I FILTRATE UNHOMOGENIZED TISSUE
Centrifuged at 1,475 xg for IO min. DISCARDED
I PELLET
Resuspended I” 2 OM Sucrose
I SUPERNATANT DISCARDED
Centrifuged ot 17,300 xg for IO ml”
/---I 20M SUPEANATANT
Diluted to 0 25M Sucrose
PELLET DISCARDED
Centrifuged ot :S,OOOxg for 15 nvn
MEMBRANE ‘PELLET
Resuspended in 0.05 M Tris-HCI pH 7.6
SUPERNATANi DISCARDED
I UPPER LAYER PLASMA MEMBRANES
I LOWER LAYER DISCARDED
FIG. 2. Schematic representation of the procedure used for the preparation of renal plasma membranes. The details of the steps are given under “Experimental Procedures.”
Fro. 3. A, electron micrograph of renal plxsma membrane prep- aration. Magnification X 30,000. The membranes were pre- pared for examinat,ion by electron microscopy by the method given in “Experimental Procedures.” B, electron micrograph of renal plasma membrane preparation showing desmasome. Magnification X 50,000.
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from
6171
TABLE I
Activation of adenylate cyclase by antidiuretic hormone and by sodium jluoride
Renal plasma membranes were assayed for adenylate cyclase as
described in “Experimental Procedures.” The final concentra- tion of ADH was 10e5 M, and sodium fluoride was added to a final concentration of 10 mM. The results represent the mean f S.D.
for triplicate determinations.
Addition I
Specific activity
@mles cyclic AMP/m/l0 min
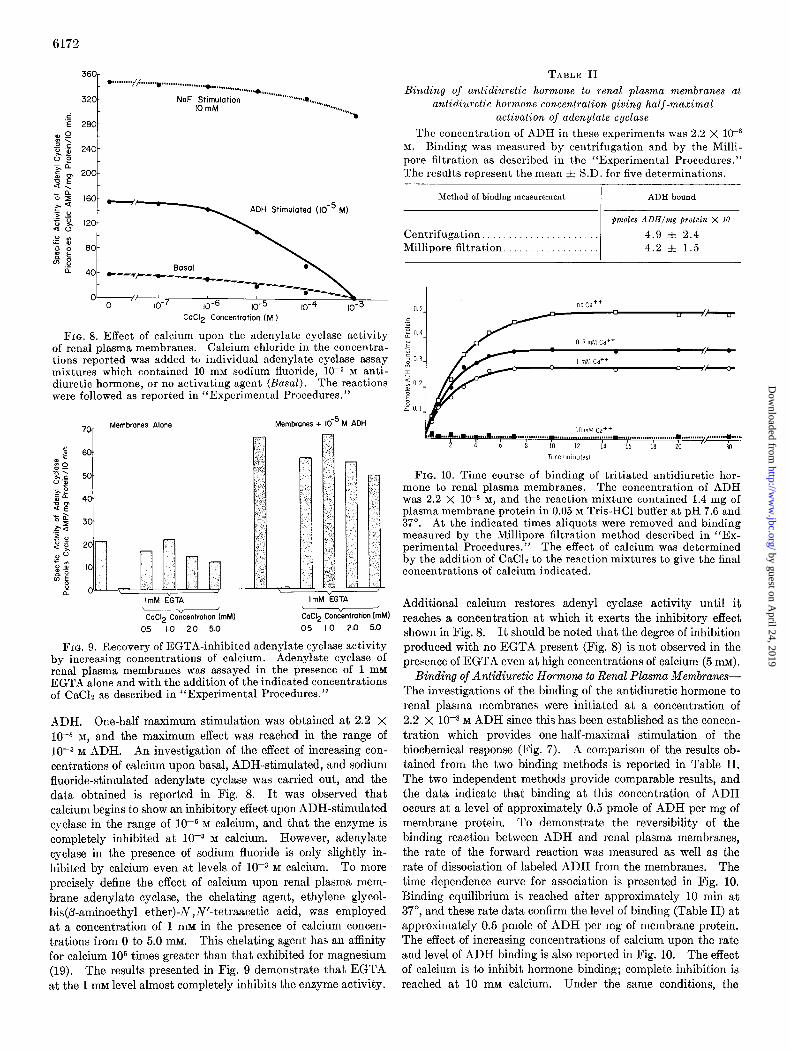
None ................................ 57 f 17
Antidiuretic hormone. ............... 169 f 24 Sodium fluoride ...................... 601 f 42
60
I / I /
Membrane Protein (pgm)
FIG. 4. Adenylate cyclase as a function of membrane protein concentration. Aliquots of a typical membrane preparation SUS- pended in 0.05 M Tris-HCl buffer at pH 7.6 were assayed as de- scribed under “Experimental Procedures.” The results are ex- pressed as picomoles of cyclic AMP formed per 7 min. Each point is the average of duplicate assays.
ATP Concentration (M x 104)
FIG. 5. The effect of different ATP concentrations on basal adenylate cyclase activity (- - -) and upon activity stimulated by antidiuretic hormone at 5 X 10T6 M (-). The assay con- ditions given under “Experimental Procedures” were employed.
I / 0 10 20 30 40 50
Sodium Fluorhde Cone hM)
FIG. 6. The effect of increasing concentrations (Cont.) of so- dium fluoride on basal adenylate cyclase activity. Plasma mem- branes at a concentration of 169 pg of plasma membrane protein were assayed in the presence of 1 to 50 mM NaF according to the procedure described in “Experimental Procedures.” Each point represents the average of four assays using the same membrane preparation.
10-10 10-g 10-e 10-l 10-G ‘5 IO-
Lysine Vasopressin Concentration (M)
FIG. 7. Dose response curve for the activation of adenylate cyclase by antidiuretic hormone. The antidiuretic hormone was added to reaction mixtures in the concentrations indicated and adenylate cyclase was assayed according to the method reported in the text. Graphical analysis of the above dose response curve demonstrated that half-maximal stimulation occurred at 2.2 X 1O-8 M ADH.
ture was employed in the adenylate cyclase assays reported in
this paper. A study of the effect of substrate concentration upon
reaction rate under basal and maximum ADH-stimulated con- ditions is presented in Fig. 5. The normal parabolic relation- ship is observed with maximal activity in both the basal and ADH-stimulated systems obtained at approximately 7 x 10e4 M
ATP. Levels approximately twice this concentration of sub- strate were usually employed to eliminate any effects of substrate depletion. Adenylate cyclase activity as a function of increasing sodium fluoride concentration up to 50 mu is reported in Fig. 6. 1Maximum stimulation by NaF is observed between 10 and 12 mu, and all subsequent NaF-stimulated assays were performed in this range. The stimulatory effect of ADH upon adenylate cyclase was measured over a hormone concentration of lo-r1 to 10e5 M ADH, and the results are shown in Fig. 7. The data indicate that the stimulatory effect of the hormone is expressed over the concentration range of approximately lo-lo to 10m6 M
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from
AOH Stimulated (10m5 M)
.- COCl2 Concentration (M )
FIG. 8. Effect of calcium upon the adenylate cyclase activity of renal plasma membranes. Calcium chloride in the concentra- tions reported was added to individual adenylate cyclase assay mixtures which contained 10 mM sodium fluoride, 1OF M anti- diuretic hormone, or no activating agent (Basal). The reactions were followed as reported in “Experimental Procedures.”
Membranes Alone Membranes + 10T5 M ADH
ImM EGTA ” <
C&l2 Concentration (mM1 CaCl2 Concentmtion (mM1
05 I.0 2.0 5.0 0.5 1.0 2.0 5.0
FIG. 9. Recovery of EGTA-inhibited adenylate cyclase activity by increasing concentrations of calcium. Adenylate cyclase of renal plasma membranes was assayed in the presence of 1 mM EGTA alone and with the addition of the indicated concentrations of CaCls as described in “Experimental Procedures.”
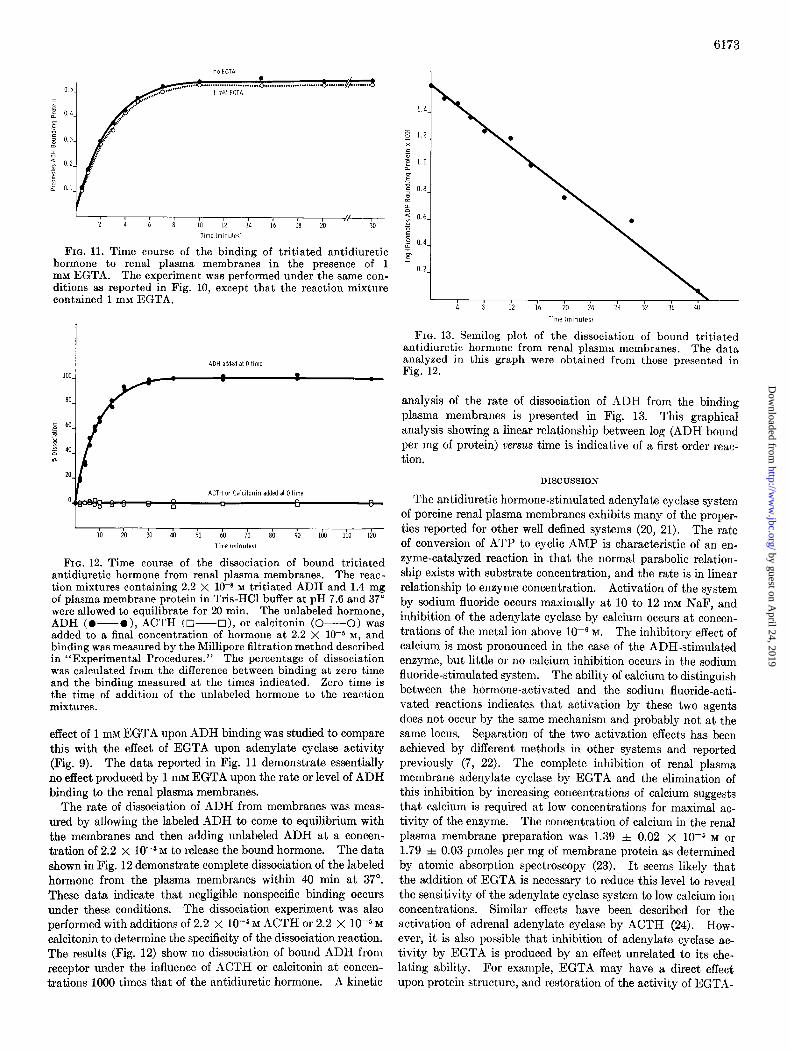
ADH. One-half maximum stimulation was obtained at 2.2 X 1O-8 M, and the maximum effect was reached in the range of 10-S M ADH. An investigation of the effect of increasing con- centrations of calcium upon basal, ADH-stimulated, and sodium fluoride-stimulated adenylate cyclase was carried out, and the data obtained is reported in Fig. 8. It was observed that calcium begins to show an inhibitory effect upon ADH-stimulated cyclase in the range of 1OW M calcium, and that the enzyme is completely inhibited at 10e3 M calcium. However, adenylate cyclase in the presence of sodium fluoride is only slightly in- hibited by calcium even at levels of 10W3 M calcium. To more precisely define the effect of calcium upon renal plasma mem- brane adenylate cyclase, the chelating agent, ethylene glycol- his@-aminoethyl ether)-N,N’-tetraacetic acid, was employed at a concentration of 1 mM in the presence of calcium concen- t.rations from 0 to 5.0 111~. This chelating agent has an affinity for calcium lo5 times greater than that exhibited for magnesium (19). The results presented in Fig. 9 demonstrate that EGTA a,t the 1 mM level almost completely inhibits the enzyme activity.
TABLE II
Binding of antidiuretic hormone to renal plasma membranes at antidiuretic hormone concentration giving half-maximal
activation of adenylate cyclase
The concentration of ADH in these experiments was 2.2 X lo-*
M. Binding was measured by centrifugation and by the Milli- pore filtration as described in the “Experimental Procedures.” The results represent the mean f S.D. for five determinations.
Method of binding measurement I ADH bound
~moles ADH/mg firotein X 10
Centrifugation. 4.9 f 2.4 Millipore filtration. 4.2 f 1.5
FIG. 10. Time course of binding of tritiated antidiuretic hor- mone to renal plasma membranes. The concentration of ADH was 2.2 X 1OW M, and the reaction mixture contained 1.4 mg of plasma membrane protein in 0.05 M Tris-HCl buffer at pH 7.6 and 37”. At the indicated times aliquots were removed and binding measured by the Millipore filtration method described in “Ex- perimental Procedures.” The effect of calcium was determined by the addition of CaCL to the reaction mixtures to give the final concentrations of calcium indicated.
Additional calcium restores adenyl cyclase activity until it reaches a concentration at which it exerts the inhibitory effect shown in Fig. 8. It should be noted that the degree of inhibition produced with no EGTA present (Fig. 8) is not observed in the presence of EGTA even at high concentrations of calcium (5 111~).
Binding of Antidiuretic Hormone to Renal Plasma Membranes- The investigations of the binding of the antidiuretic hormone to renal plasma membranes were initiated at a concentration of 2.2 x 10m8 M ADH since this has been established as the concen- tration which provides one-half-maximal stimulation of the biochemical response (Fig. 7). A comparison of the results ob- tained from the two binding methods is reported in Table II. The two independent methods provide comparable results, and the data indicate that binding at this concentration of ADH occurs at a level of approximately 0.5 pmole of ADH per mg of membrane protein. To demonstrate the reversibility of the binding reaction between ADH and renal plasma membranes, the rate of the forward reaction was measured as well as the rate of dissociation of labeled ADH from the membranes. The time dependence curve for association is presented in Fig. 10. Binding equilibrium is reached after approximately 10 min at 37”, and these rate data confirm the level of binding (Table II) at approximately 0.5 pmole of ADH per mg of membrane protein. The effect of increasing concentrations of calcium upon the rate and level of ADH binding is also reported in Fig. 10. The effect of calcium is to inhibit hormone binding; complete inhibition is reached at 10 mM calcium. Under the same conditions, the
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from
6173
FIG. 11. Time course of the binding of tritiated antidiuretic hormone to renal plasma membranes in the presence of 1 mM EGTA. The experiment was performed under the same con- ditions as reported in Fig. 10, except that the reaction mixture contained 1 mM EGTA.
io io ti 40 $I do io sb do iba Go Ii0 Time iminuterl
FIG. 12. Time course of the dissociation of bound tritiated antidiuretic hormone from renal plasma membranes. The reac- tion mixtures containing 2.2 X 10-n M tritiated ADH and 1.4 mg of plasma membrane protein in Tris-HCl buffer at pH 7.6 and 37” were allowed to equilibrate for 20 min. The unlabeled hormone, ADH (O-O), ACTH (O----O), or calcitonin (O--O) was added to a final concentration of hormone at 2.2 X 1CW M, and binding was measured by the Millipore filtration method described in “Experimental Procedures.” The percentage of dissociation was calculated from the difference between binding at zero time and the binding measured at the times indicated. Zero time is the time of addition of the unlabeled hormone to the reaction mixtures.
effect of 1 InM EGTA upon ADH binding was studied to compare this with the effect of EGTA upon adenylate cyclase activity (Fig. 9). The data reported in Fig. 11 demonstrate essentially no effect produced by 1 mu EGTA upon the rate or level of ADH binding to the renal plasma membranes.
The rate of dissociation of ADH from membranes was meas- ured by allowing the labeled ADH to come to equilibrium with the membranes and then adding unlabeled ADH at a concen- tration of 2.2 x lWs M to release the bound hormone. The data shown in Fig. 12 demonstrate complete dissociation of the labeled hormone from the plasma membranes within 40 min at 37”. These data indicate that negligible nonspecific binding occurs under these conditions. The dissociation experiment was also performed with additions of 2.2 x 10m5 M ACTH or 2.2 X 10F5 M
calcitonin to determine the specificity of the dissociation reaction. The results (Fig. 12) show no dissociat.ion of bound ADH from receptor under the influence of ACTH or calcitonin at concen- trations 1000 times that of the antidiuretic hormone. A kinetic
i i i2 i6 io i4 Zk j2 i6 4b
Time (minutes1
FIG. 13. Semilog plot of the dissociation of bound tritiated antidiuretic hormone from renal plasma membranes. The data analyzed in this graph were obtained from those presented in Fig. 12.
analysis of the rate of dissociation of ADH from the binding plasma membranes is presented in Fig. 13. This graphical analysis showing a linear relationship between log (ADH bound per mg of protein) versus time is indicative of a first order reac- tion.
DISCUSSION
The antidiuretic hormone-stimulated adenylate cyclase system of porcine renal plasma membranes exhibits many of the proper- ties reported for other well defined systems (20, 21). The rate of conversion of ATP to cyclic AMP is characteristic of an en- zyme-catalyzed reaction in that the normal parabolic relation- ship exists with substrate concentration, and the rate is in linear relationship to enzyme concentration. Activation of the system by sodium fluoride occurs maximally at 10 to 12 mM NaF, and inhibition of the adenylate cyclase by calcium occurs at concen- trations of the metal ion above 1OV M. The inhibitory effect of calcium is most pronounced in the case of the ADH-stimulated enzyme, but little or no calcium inhibition occurs in the sodium fluoride-stimulated system. The ability of calcium to distinguish between the hormone-activated and the sodium fluoride-acti- vated reactions indicates that activation by these two agents does not occur by the same mechanism and probably not at the same locus. Separation of the two activation effects has been achieved by different methods in other systems and reported previously (7, 22). The complete inhibition of renal plasma membrane adenylate cyclase by EGTA and the elimination of this inhibition by increasing concentrations of calcium suggests that calcium is required at low concentrations for maximal ac- tivity of the enzyme. The concentration of calcium in the renal plasma membrane preparation was 1.39 f 0.02 X 10m5 M or 1.79 + 0.03 pmoles per mg of membrane protein as determined by atomic absorption spectroscopy (23). It seems likely that the addition of EGTA is necessary to reduce this level to reveal the sensitivity of the adenylate cyclase system to low calcium ion concentrations. Similar effects have been described for the activation of adrenal adenylate cyclase by ACTH (24). How- ever, it is also possible that inhibition of adenylate cyclase ac- tivity by EGTA is produced by an effect unrelated to its che- lating ability. For example, EGTA may have a direct effect upon protein structure, and restoration of the activity of EGTA-
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from

6174
inhibited adenylate cyclase by additional calcium may be due to a saturation of EGTA with calcium which reverses the struc- tural effect produced by the free chelator. The same effect could also be produced by the prevention of chelation of another mem- brane-bound metal in the presence of excess calcium.
The dose response curve for activation of porcine renal adenyl- ate cyclase by the antidiuretic hormone is also similar to that reported in other activated adenylate cyclase systems (25). Half-maximal activation of adenylate cyclase in the ADH-sys- tern is approximately 2.2 X lo-* M. This concentration of hormone required for half-maximal stimulation is higher than reported circulating hormone levels (26). This difference be- tween the level of hormone required to produce the biochemical response of the in vitro system and the physiological concentra- tion of the hormone has been noted in other hormone-activated reactions (25, 27). It is probably produced by alterations of the hormone-receptor system as it is isolated from intact tissue.
Direct measurement of the binding of biologically active radio- active ADH to renal plasma membranes provides additional insight into the hormone-mediated reaction. Such measure- ments have been reported and correlated with the activation of adenylate cyclase by glucagon in the liver (28), by ACTH in the adrenal (24), and by angiotensin in the adrenal and kidney (29). It was not possible to establish a complete dose response curve for the binding of the antidiuretic hormone to plasma mem- branes, because, at the lower concentration level (lo+), the difference in radioactivity between controls and plasma mem- brane pellets or filters was too small to measure accurately the net amount of bound labeled hormone. Furthermore, at levels of hormone above 10m5 M binding continues to increase so that a maximum binding level to correspond with the maximal bio- chemical response is not reached. Binding at these high concen- trations of ADH probably represents nonspecific binding. Nonspecific binding at high concentrations of hormone, for example, has been reported for the binding of insulin to adipose tissue cells (30). For this reason a concentration of ADH was employed which produced half-maximal adenylate cyclase acti- vation. The binding of ADH observed at this concentration (0.5 pmole per mg of protein) corresponds closely to that re- ported for the binding of glucagon to liver plasma membranes (1 pmole per mg of protein). The rates of association and dis- sociation of the a&diuretic hormone from renal plasma mem- branes are close to those reported for the interaction of glucagon with liver plasma membranes (28). In both systems equilibrium is reached at a concentration of hormone which provides half- maximal stimulation within approximately 10 min. It is sig- nificant that the rate of association of ADH with renal plasma membranes is considerably faster than that reported for the disulfide interchange reaction between ADH and bovine serum albumin (31). However, when compared with the very rapid physiological response to ADH, the rate of association of hor- mone to membrane appears relatively slow. This may again be the result of alterations in the properties of the receptor complex that occur during the preparation of the plasma membranes. The complete release of labeled ADH from its binding site in the presence of excess purified unlabeled hormone demonstrates that the interaction is reversible and that nonspecific binding is not appreciable at 2.2 X lo-* M ADH. The failure of ACTH or calcitonin to exchange with ADH at its binding site provides evidence for the specificity of the reaction between the anti- diuretic hormone and its receptor on the renal plasma membrane. The observation t,hat the kinetic data of the dissociation reaction follow a simple first order rate equation may be an indication of
the homogeneity of the binding sites and of the lack of irreversible changes taking place during the time of binding. However, it has been shown that the antidiuretic hormone is degraded by this renal plasma membrane preparation (32), and for this reason the determination of the equilibrium constant from kinetic data was not attempted.
The inhibition of the rate and extent of ADH binding by calcium concentrations in the millimolar range may result from calcium ion competition with ADH at its binding site or from a calcium-induced effect upon the state of aggregation of the mem- branes which reduces binding. Quantitative measurements of calcium binding and investigations of membrane conformation will be required to resolve this question. Inhibition of binding may be related to calcium inhibition of ADH-activated adenylate cyclase at levels above 10e6 M calcium. But a comparison of the effect of EGTA upon binding and upon adenylate cyclase activity indicates a different role for calcium at low concentrations. EGTA completely abolished the adenylate cyclase activity of the plasma membranes and this activity was restored by in- creasing concentrations of calcium, but EGTA at the same concentration (1 mM) exerted absolutely no measurable effect upon the rate or extent of ADH binding to plasma membranes. These results indicate that the regulatory effect suggested for calcium at low concentrations is not a result of inhibition of binding of the hormone to the membrane. If these results are interpreted as a loss in activity because of a sequestering of cal- cium rather than a direct effect of the chelator upon membrane structure, it seems likely that this effect of calcium is produced at a locus between the binding site and the catalytic component which is directly involved in the conversion of ATP to cyclic AMP. The observation that closely similar effects are produced by calcium in the ACTH-activated adrenal adenylate cyclase system (24) suggests that a multiple regulatory role for calcium at the membrane surface and within the membrane matrix may operate in other polypeptide hormone-mediated processes (33).
Acknowledgments-We are indebted to the New England Nuclear Co. for tritiation of the iodinated antidiuretic hormone. We would like to thank Dr. James Esterly and Mr. Don Thomp- son for assistance with the electron microscopy. Finally, we are grateful to Dr. Robert Schlueter, Armour Pharmaceutical Co., for his gifts of purified adrenocorticotropic hormone and calci- tonin.
REFERENCES
1. BENTLEY, P. J. (1959) J. Endocrinol. 18, 327 2. PETIZRSON. M. J.. AND EDF:LMAN. I. S. (1964) J. Clin. Invest.
3. 43,583$94 ’
\ I
WHITTEMBURY, G., SUCISO, N., AND SOLOMON, A. K. (1960) Nature 187, 699
4. 5.
THORN, N. A. (1960) Dan. Med. Bull. ‘7, 110 DICKER, S. E., ‘ZND EGGLETON, M. G. (1961) J. Physiol. (Lon-
don)167, 351 6. 7.
8.
NIELSEN, B. (1964) Acta Endocrinol. 46, 151 HYNIE, S., AND SHARP, G. W. G. (1971) Biochim. Biophys.
Acta 230, 40 MARUMO, F., AND EDELM.\N, I. S. (1971) J. Clin. Invest. 60,
1613 9.
10.
11. 12. 13. 14.
15.
MELSON, G. L., CH.~SE, L. R., AND AURBACH, G. D. (1970) Endocrinology 86, 511-518
CAMPBELL, B. J., CHU, F. S., AND HUBBARD, S. (1963) Bio- chemistry 2, 764
LEE, N. D., ANC WISF,M.\N, R. (1959) J. Lab. Clin. Med. 64,326 MOORE, S., AND STEIN, W. (1963) Methods Enzymol. 6, 819 BRAY, G. A. (1960) Anal. Biochem. 1, 279-285 FITZPATRICK, D. F., DAVENPORT, G. R., FORTE, L., AND L-IN-
DON, E. J. (1969) J. Biol. Chem. 244, 3561-3569 LOWRY, 0. H., AND LOPEZ, J. A. (1946) J. Biol. Chem. 162, 421
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from

6175
16. LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL, R. J. (1951) J. Biol. Chem. 193, 265-275
17. WHITE, A. A., AND ZENSER, T. V. (1971) Anal. Biochem. 41, 372
18. WHITE, A. A., NORTHRUP, S. J., AND ZENSER, T. V. in Methods in Cyclic Nucleotide Research (CHASIN, M., ed) Vol. V, Marcel Dekker, Inc., New York, in press
19. CHABERCK, S., AND MARTELL, A. E. (1959) in Organic Seques- tering Agents, p. 557, Wiley, New York
20. TAUNTON, 0. D., ROTH, J., AND PASTAN, I. (1969) J. Biol. Chem. 244, 247-253
21. POHL, S. L., BIRNBAUMER, L., AND RODBELL, M. (1971) J. Biol. Chem. 246, 1849-1856
22. BIRNBAUMER, L., POHL, S. L., AND RODBELL, M. (1971) J. Biol. Chem. 246, 1857-1860
23. RENO, A. M., AND CAMPBELL, B. J. (1969) J. Biol. Chem. 244, 144551450
24. LEFKOWITZ, R. J., ROTH, J., AND PASTAN, I. (1970) Nature 228, 864
25. PASTAN, I., AND PERLMAN, R. A. (1971) Nature 229,5 26. BONJOUR, J. P., AND MALVIN, R. L. (1970) Amer. J. Physiol.
218, 1128 27. Bii~, H. P., HECHTER, O., SCHWARTZ, I. L., AND WALTER, R.
(1970) Proc. Nat. Acad. Sci. U. S. A. 67, 7 28. RODBELL, M., KRANS, H. M. J., POHL, S. L., AND BIRNBATJMER,
L. (1971) J. Biol. Chem. 246, 1861-1871 29. GOODFRIEND, T. L., AND LIN, S. Y. (1970) Circ. Res. Suppl.
26-27, I-163 30. CUATRECASAS, P. (1971) Proc. Nat. Acad. Sci. U. S. A. 68.1264 31. EDWARDS, F.. B., ROMBAUER, R. B., AND CAMPBELL, b. J.
(1969) Biochim. Biovhus. Acta 914. 234-245 32. N~RDA&I, N., BoRBE~G~V., AND CAMPBELL, B. J. (1972) Fed.
Proc. 31, 890 33. RASMUSSEN, H. (1970) Science, 170, 404
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Benedict J. Campbell, Gerald Woodward and Victoria BorbergPlasma Membranes
Calcium-mediated Interactions between the Antidiuretic Hormone and Renal
1972, 247:6167-6175.J. Biol. Chem.
http://www.jbc.org/content/247/19/6167Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/247/19/6167.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 24, 2019
http://ww
w.jbc.org/
Dow
nloaded from





























