Embed Size (px)
Citation preview

Review ArticleCalcium Influx and Male Fertility in the Context ofthe Sperm Proteome An Update
Md Saidur Rahman Woo-Sung Kwon and Myung-Geol Pang
Department of Animal Science and Technology Chung-Ang University 4726 Seodong-daero AnseongGyeonggi-Do 456-756 Republic of Korea
Correspondence should be addressed to Myung-Geol Pang mgpangcauackr
Received 17 December 2013 Accepted 14 January 2014 Published 27 April 2014
Academic Editor Jeroen Krijgsveld
Copyright copy 2014 Md Saidur Rahman et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Freshly ejaculated spermatozoa are incapable or poorly capable of fertilizing an oocyte The fertilization aptness of spermatozoadepends on the appropriate and time-dependent acquisition of hyperactivation chemotaxis capacitation and the acrosomereaction where calcium (Ca2+) is extensively involved in almost every step A literature review showed that several ion channelproteins are likely responsible for regulation of the Ca2+ uptake in spermatozoaTherefore manipulation of the functions of channelproteins is closely related to Ca2+ influx ultimately affecting male fertility Recently it has been shown that together with differentphysiological stimuli protein-protein interaction also modifies the Ca2+ influx mechanism in spermatozoa Modern proteomicanalyses have identified several sperm proteins and therefore these findings might provide further insight into understanding theCa2+ influx protein functions and regulation of fertility The objective of this review was to synthesize the published findings onthe Ca2+ influx mechanism in mammalian spermatozoa and its implications for the regulation of male fertility in the context ofsperm proteins Finally Pathway Studio (90) was used to catalog the sperm proteins that regulate the Ca2+ influx signaling by usingthe information available from the PubMed database following a MedScan Reader (50) search
1 Introduction
Spermatozoa are atypical cells with peculiar functionalitythey are produced in one organism and released and thenthey invade another organism and deliver their geneticmaterial into a host cell to produce offspring by sexualreproduction It is a well-known fact that only about 1 in25000 spermatozoa finally reaches the fallopian tube andgets the opportunity to fertilize an oocyte In the mid-20thcentury it had been claimed that mammalian spermatozoaare unable to fertilize an oocyte before achieving functionalmaturation which occurs during their journey through thefemale reproductive tract for a finite period of time [1 2]This fundamental maturational process is chiefly regulatedby numerous signaling cascades and calcium (Ca2+) playsa dynamic role in this process as an intracellular secondmessenger Several studies have hypothesized that elevationof sperm intracellular Ca2+ ([Ca2+]i)Ca
2+ influx regulatesmotility hyperactivation chemotaxis capacitation and the
acrosome reaction and facilitates the spermatozoa reachingand fertilizing of an oocyte [3ndash8] Therefore understandingthe mechanism that regulates the Ca2+ influx in spermatozoais a matter of utmost importance
Previous studies have shown that the Ca2+ entry mecha-nisms are regulated via numerous Ca2+ permeable channelproteins in spermatozoa [6 9 10] Therefore the factorsthat regulate the functions of those channels will ulti-mately help us understand how male fertility is regulatedRecent applications of proteomic approaches such as two-dimensional polyacrylamide gel electrophoresis mass spec-trometry and differential in-gel electrophoresis have yieldedthe identification of several sperm-specific proteins [11 12]These discoveries have provided new insight into proteinfunctions and enabled us to recognize diverse sperm-specificprocesses in order to differentiate normal from abnormalspermatozoa [11] Mature spermatozoa are widely known tobe silent in both transcription and translation [11 13 14]or poorly capable of translation [15] therefore studies on
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 841615 13 pageshttpdxdoiorg1011552014841615
2 BioMed Research International
individual sperm proteomes have described the importanceof spermatozoal posttranslational modifications and theirability to induce physiological changes as a prerequisite forsuccessful fertilization
Torres-Flores et al [16] have shown that human sperma-tozoa exposed to the phosphodiesterase inhibitor papaverinecause activation of protein kinase A (PKA) and stimulate theprogesterone-induced Ca2+ influx via the cyclic adenosinemonophosphate- (cAMP-) dependent pathway Althoughthese authors did not evaluate the relationship between invitro fertility and Ca2+ influx changes in intracellular pHand increased tyrosine phosphorylation ultimately providea potential clue regarding sperm fertility competence Inanother study to evaluate hamster spermatozoa capacitationcapability comparative association was observed betweenpyruvate dehydrogenase A Ca2+ influx cAMP and reactiveoxygen species [17] Additionally Breitbart et al [18] reportedthat polymerization of globular- (G-) actin to filamentous-(F-) actin occurs during capacitation As capacitation andthe acrosome reaction are Ca2+-mediated events [4 5] onecan without considering further signaling cascade assumethat remodeling the actin structure might be linked with theregulation of Ca2+ influx in spermatozoa
Recently in our laboratory we found that the manip-ulation of sperm proteins such as ubiquinol-cytochrome-creductase core protein 2 (UQCRC2) [39] voltage-dependentanion channels proteins (VDACs) [4] and arginine vaso-pressin [5] could control the Ca2+ influx in spermatozoa andregulate capacitation the acrosome reaction and fertilityTherefore design and construction of a similar study withmost of the identified sperm proteins available from severalprotein databases might provide a more realistic insight intothe Ca2+ influx protein functions and fertility The presentwork reviews the latest information published by other labo-ratories as well as our research team on the aforementionedaspects of spermatozoa and their potential implications fordiagnosis and prognosis of male fertility
2 Mechanism of Ca2+ Influx in MammalianSpermatozoa
The ultimate goal of fertilization of mammalian sperm is tofuse with and deliver their geneticmaterials into an oocyte [240 41] For fertilization to occur completely the spermatozoamust experience various obstacles both in vitro and in vivo[40 41] Ca2+ ions act as central signaling molecules oncethey enter the spermatozoa they exert allosteric regulatoryeffects on enzymes and many proteins [10 21 42] Indeednumerous elegant research findings have contributed signif-icantly to our understanding of the molecular signaling ofCa2+ influx especially through monitoring the activity ofindividual cells However most of the studies are discreteand often do not represent a cumulative idea This sectionpresents a compilation of some basic information regardingthe Ca2+ entry mechanism into mammalian spermatozoa byrecapitulating scientific evidence
The literature reviewed shows that the primary source ofCa2+ for spermatozoa is the external environment the fallop-ian tube in the female reproductive tract (in vivo) and culturemedia (in vitro) [8] and simultaneously increasing [Ca2+]iregulates the release of Ca2+ into the cellTherefore howCa2+crosses into cells through the sperm plasma membrane isa matter of paramount importance In eukaryotic cells theCa2+ influx occurs through specificCa2+ permeable ion chan-nel proteins located on the plasmamembrane [43 44] such asclassical voltage-gated (high and low) Ca2+ (Cavs) transientreceptor potential (TRP) and cyclic nucleotide-gated (CNG)channels [9] Recently Ren and Xia have proposed fourcriteria to identify sperm ion channel proteins detectabilityin sperm preferably with knockout sperm as a negativecontrol ability to produce ion channel current detectable bypatch-clamp recording blocking of the channels that impairsnormal sperm function and mutations of gene encoding theion channel proteins leading to sperm malfunctions [10]
The CatSper family of channels is the newest and onlyfamily of voltage-gated Ca2+ channels that meets most of theaforementioned criteria and essentially regulates Ca2+ entryinto cells and is therefore crucial for sperm fertility [9 45]Four pore-forming CatSper channel proteins CatSper 1ndash4and at least two auxiliary subunits CatSper120573 and CatSper120574have been identified in a wide range of animals includinghumans andmice [46 47] Physiologically CatSpermembersare permeable to Ca2+whereas the CatSper knockdownsperm does not have the channel current that is detectedin the principal piece of wild-type sperm [20 48] Mostof the channel proteins including CatSper members havebeen identified in the principal piece of spermatozoa [2046 47 49] (Figure 1) Although the explanation of suchsubcellular localization is still debated it might be becauseof interactions among the channel proteins and with theauxiliary subunits although a further study is needed toresolve this issue Collectively these proteins play a key rolein various cellular processes via regulation of the membranepotential and intracellular ionic balance Carlson et al [50]and Quill et al [51] have conclusively proved that CatSper1and CatSper2 null mice are sterile owing to their inabilityto generate the sperm-hyperactivated motility prerequisitefor penetration of an oocyte extracellular matrix In effectthe complete or partial absence of single or multiple Ca2+channels is responsible for infertility or subfertility althoughtheir underlying signaling cascade has not been properlystudied
Previously it has been reported that CatSper-dependentincreases of [Ca2+]i in spermatozoa are induced byseveral psychological stimuli such as cyclic nucleotides(eg cAMP and cGMP) [29 30 52] soluble adenylylcyclase [29 52] zona pellucida glycoprotein [34 35 38]serum albumin [37 38] secretion of cumulus oophorus[38] intracellular alkalization [3 53] and pH [6 21] Arecent study showed that endocrine disruptors such as pp1015840-dichlorodiphenyldichloroethylene (pp1015840-DDE) promotedCa2+ entry into spermatozoa by activating CatSper channelseven at a physiological concentration [36] In additionseveral other components are also known to play an
BioMed Research International 3
GPC
RC
Alkalinization
CatS
per
120574120573
Ca2+
[Ca 2+]i
sACY
ATP cAMPcGMP
ZP receptors
HCN
Stimulate
Opening
CN
G
K+
Ca2+
Progesterone
Spermatozoa
Principalpiece
H+
Extracellular space
Intracellular space
Na+
NBC
Na+
Follicle
nHCO3minus
HCO3minus
Hv1
pH
Figure 1 Possible signal transduction mechanisms of mammalian sperm Ca2+ influx through the Ca2+ permeable channel proteinsPreviously published studies were used as references to summarize the list of channel proteins in spermatozoa The channel proteins arelocalized mainly in the principle piece of spermatozoa The follicular fluid and several factors in the fallopian tube (in vitromedia) stimulatethe receptors for spermatozoa Ca2+ influx Ca2+ influx in spermatozoa is principally regulated by CatSper channels however the possibleinteraction between other channels that are responsible for controlling the opening of CatSper and allowing the Ca2+ into cells is indicatedby arrow signs (red circle) The different channel proteins that are depicted in the diagram include the Na+-coupled HCO
3
minus transporter(NBC) family soluble adenylyl cyclase (sACY) adenosine triphosphate (ATP) cyclic adenosine monophosphate (cAMP) cyclic guanosinemonophosphate (cGMP) cyclic nucleotide-gated ion channel (CNG) hyperpolarization-activated cyclic nucleotide-gated channel (HCN)zona pellucida (ZP) the voltage-gated proton channel (Hv1) glutamate receptor family class-C (GPCRC) and an unknown mechanism ()
important role in Ca2+ influx mechanisms in mammalianspermatozoa by regulating the opening of CatSper membersincluding the flagellar voltage-gated proton channel (Hv1)[21] Ca2+-ATPase pump [33] several cyclic nucleotide-gatedion channels (CNG) [27 54] hyperpolarization-activatedcyclic nucleotide-gated (HCN) channels [27] and G-proteincoupled receptors (GPCRs)
A hypothetical signaling cascade of Ca2+ influx pathwaysand interaction of several channel proteins is depicted inFigure 1 Although the functions of several ion channelproteins together with their concurrent relationship withnumerous stimuli have been well studied [21 27 38] severalfundamental questions remain unanswered for examplehowdo these channelsstimuli regulate theCa2+ influx duringspermatozoa processes such as capacitation the acrosomereaction and fertilization Do they work alone or togetherwith other channel proteins to regulate Ca2+ influx More-over which other parameters that remain undetected couldhave an effect on Ca2+ influx Therefore future researchshould focus on resolving these issues Table 1 summarizesthe proposed effect of Ca2+ ion channels and their physio-logical role that ultimately helps Ca2+ influx into mammalianspermatozoa
3 Effect of Ca2+ Influx on Male Fertility
Ca2+ triggers multiple physiological events in sperma-tozoa such as hyperactivation chemotaxis capacitationand the acrosomal reaction all of which are essential for
successful fertilization In mammalian spermatozoa numer-ous Ca2+ permeable channel proteins control intracellularpH and the pH-dependent Ca2+ influx is measured by thewhole-cell patch clamp technique [9 20] A review of theliterature showed that a potential functional interaction existsbetween the sperm proteins and Ca2+ permeable chan-nel proteins thus modulating the Ca2+ influx mechanism[4 5 39] and playing a vital role in adjusting male fertilityHowever themechanism bywhich Ca2+ triggers intracellularsignaling to regulate physiological events in spermatozoaand the role of sperm proteins in adjustment of Ca2+influx into cells remains unclear This topic is emphasizedbelow
31 Ca2+ Influx Sperm Hyperactivation Chemotaxis andProtein Functions In general mature spermatozoa are heldimmotile within the epididymis However they quickly beginto swim following release This is known as activation ofmotility and is characterized by symmetrical flagellar beats[55 56]The terms sperm activation and hyperactivation havequite different meanings The spermatozoa become hyperac-tivatedwhen the amplitude of the flagellar bend increases andproduces a highly asymmetrical beat In vivo hyperactivationof spermatozoa facilitates the release of sperm from oviductalstorage and boosts them through mucus in the oviductallumen and matrix of the cumulus oophorus during fertiliza-tion [7] In contrast chemotaxis is a formof spermmovementin which spermatozoa move toward a concentration gradientof a chemoattractant released from the oocyte [57 58]
4 BioMed Research International
Table 1 Summary of published works on ion channels and physiological stimuli of mammalian spermatozoa that regulate the Ca2+ influxmechanism
Name ofchannelstimuli
Localization onspermatozoaavailability Role in Ca2+ influx Role in sperm
physiologyEffect of knockingdownabsence Reference
CatSperCatSper 1CatSper 2CatSper 3CatSper 4CatSper120573CatSper120574
Principal piece Regulates Ca2+ influxCa2+ uptakehyperactivated
motilitySterile Barratt and Publicover
[19] Qi et al [20]
Hv1 Principal pieceIntracellular pHalkalization thus
stimulate Ca2+ influx
Extrudesprotons from
flagellaalkalization
Fertile Lishko et al [21]Lishko et al [22]
119868ATP Midpiece Selectively transports theCa2+
Ca2+ influxalkalization Fertile Navarro et al [23]
TRPC Principal piecemidpiece
Stimulates opening ofCatSper
Ca2+ influx celldepolarization Fertile Gees et al [24]
Castellano et al [25]
CNG Sperm flagellum headStimulates opening of
CatSper viacAMPcGMP
Ca2+ influx Fertile Biel and Michalakis [26]
HCN Flagellum Depolarization andopening of CatSper Ca2+ influx Fertile Wiesner et al [27]
SOC Plasma membrane ZP-induced Ca2+ influx Spermchemotactic Subfertile Yoshida et al [28]
sACYcAMPcGMP
Intracellular space andcell membrane
Activates CatSper CNGand HCN to regulate
Ca2+ influx
Ca2+ influxalkalization Sterile Esposito et al [29]
Hess et al [30]
GPCR(s) Principal piecemidpiece
ZP-induced Ca2+ influxincreases in [Ca2+]i
Maintainsfertilization Subfertile Fukami et al [31]
Fukami et al [32]
PLC120575 Acrosome ZP induced increases in[Ca2+]i
Ca2+ influx Subfertile Fukami et al [32]
Ca2+-ATPasepump Principal piece Intracellular pH and
alkalizationCa2+ influxcapacitation
Motility loss resultsin infertility Wennemuth et al [33]
ZPglycoproteins Follicle Induced Ca2+-dependent
increase in [Ca2+]iHyperactivationcapacitation
Delayedcapacitation
Florman [34]Florman et al [35]
Endocrinedisruptor(pp1015840-DDE)
Female reproductivetract Activates CatSper Ca2+ influx
Motility lossdelayed
capacitationTavares et al [36]
BSA Extracellular space Similar to ZPglycoprotein
In vitrocapacitation
Motility losssubfertility
Xia and Ren [37]Bailey and Storey [38]
Oviductal andfollicular fluid
Extracellular space(in vivo)
Ca2+-dependent increasein [Ca2+]i in sperm Ca2+ influx
Motility lossdelayed
capacitationXia and Ren [37]
Hv1 voltage-gated proton channel 119868ATP ATP-gated channel TRPC transient receptor potential channels CNG cyclic nucleotide-gated ion channel HCNhyperpolarization-activated cyclic nucleotide-gated channel SOC store-operated Ca2+ channel cAMP cyclic adenosine monophosphate cGMP cyclicguanosine monophosphate sACY soluble adenylyl cyclase GPCR glutamate receptor family class-C PLC120575 phospholipase C zeta ZP zona pellucida pp1015840-DDE pp1015840-dichlorodiphenyldichloroethylene BSA bovine serum albumin
However themolecular event that characterizes spermatozoachemotaxis is only partially known [57]
There is strong evidence to support that sperm hyper-activation and chemotaxis are required for penetrating thezona pellucida [48 57 59 60] Incubation of spermatozoawith an extracellular Ca2+ source induces hyperactivationin mammalian spermatozoa [61 62] and chemotaxis instarfish [57] In addition measuring cytoplasmic Ca2+ levels
by using the fluorescent Ca2+ indicator indo-1 proved thatspermatozoa hyperactivation is potentially regulated by Ca2+influx However it is unknown whether Ca2+ influx indepen-dently induces hyperactivationchemotaxis in mammalianspermatozoaHo and Suarez [56] proposed that spermhyper-activation induced by Ca2+ influx is mainly pH-dependentbecause sperm require a pH of 79ndash85 for hyperactivationwhereas activation can occur at a pH lt 70 The proposed
BioMed Research International 5
model of Ca2+-induced hyperactivation is represented inFigure 2
It has recently been found by our laboratory thattreatment of mouse spermatozoa with nutlin-3a a smallmolecule antagonist of the mouse double minute 2 repressorpotentially downregulates the functions of the ubiquinol-cytochrome-c reductase complex component UQCRC2 andcorrelated with significantly reduced [Ca2+]i and spermhyperactivation This study provided insight that the Ca2+influx in spermatozoa is partially regulated by UQCRC2protein Kwon et al [4] reported that blocking VDACwith 441015840-diisothiocyanostilbene-221015840-disulfonic acid (DIDS)significantly decreased sperm hyperactivation A signifi-cant decrease in [Ca2+]i was observed in (minus) DIDS con-ditions while [pH]i significantly increased in (minus) DIDSregardless of Ca2+ Simultaneously a significantly elevated[pH]i was observed in (+) Ca2+ This study providesstrong evidence that the modulation of Ca2+ influx byVDACs is pH-dependent which is consistent with the resultof a previous study by Ho and Suarez [56] Moreoveranother study proposed that deamino [Cys 1 d-ArgS] vaso-pressin (dDAVP) an AVPR2 agonist significantly decreasedsperm motility and intracellular pH but interestingly itincreased [Ca2+]i by regulating the function of argininevasopressin in mice spermatozoa However it remains tobe clarified as to why spermatozoa motility is decreased evenin increased [Ca2+]i conditions
On the basis of the findings of the aforementionedstudies it is tempting to hypothesize that spermatozoahyperactivation is mostly controlled by Ca2+ influx How-ever potential interactions exist between protein func-tions Therefore Ca2+ influx protein interaction andhyperactivation might give numerous different annotationsof upcoming research in this field We have illustrateda schematic representation of different signaling path-ways involving sperm proteins by using Pathway StudioThese proteins exhibit significant modifications to inducesperm hyperactivation and chemotaxis in spermatozoa byregulating Ca2+ influx (Figure 3)
32 Ca2+ Influx versus Capacitation the Acrosomal ReactionFertilization and Sperm Proteome Mammalian fertilizationis a species-specific episode that is accomplished by a complexset of molecular events To fertilize an oocyte multipleextreme changes occur in spermatozoa that begin from itsformation in the testes of the male reproductive tract to itspenetration and fusionwith an egg in the female reproductivetract Although spermatozoa are motile as well asmorphologically normal after ejaculation they are unable tofertilize an oocyte [59]They gain the fertilization ability onlyafter educating in the female reproductive tract [40] and themodifications that spermatozoa experience during this timeare collectively known as ldquocapacitationrdquo Only capacitatedspermatozoa can undergo the acrosome reaction throughbinding to the egg zona pellucida and they finally becomecapable of penetrating and fertilizing the egg [4 18 39]
The term ldquocapacitationrdquo was proposed by Austin in 1952[1] although this concept was initially described by bothChang and Austin in 1951 [2 41] In fact in vivo capacitationtakes place in the female reproductive tract however it is alsopossible to capacitate spermatozoa in vitro by using particularmedia containing appropriate electrolytes and pH [2] Inan elegant review Visconti summarized that the early stageof capacitation mainly comprises the bicarbonate-mediatedactivation of sperm motility whereas the late stages includeintracellular alkalinization increase in protein tyrosine phos-phorylation and preparation for the acrosomal reaction [63]These temporal differences in capacitation and the acrosomereaction require numerous mechanisms and Ca2+ influxplays a significant role in the process [63 64] Fraser [65]reported that capacitation is a comparatively slow event thatrequires several hours to complete and is mainly regulated bya modest rise in [Ca2+]i whereas the acrosome reaction is anexocytosis process that occurs very rapidly (within a minute)and is triggered by a large influx of [Ca2+]i [65 66]
Although the biochemical phenomenon of Ca2+regulated capacitation and the acrosome reaction havebeen known for the last two decades the molecular basisof this process is still poorly understood For capacitationthe cholesterol influx initially stimulates the elevationof [Ca2+]i and bicarbonate into the spermatozoa andfinally activates PKA and tyrosine phosphorylationrespectively via the production of the cAMP [66ndash68]In addition binding to the zona pellucida causes additionalactivation of cAMPPKA and protein kinase C (PKC)[68ndash70] Spermatozoa need [Ca2+]i influx to proceed furtherand they are believed to be activated by PKC through theopening of the calcium channels Interestingly PKA togetherwith a secondary messenger inositol trisphosphate activatescalcium channels localized in the outer acrosomalmembraneand increases the calcium concentration in the cytosolFurther increase of cytosolic Ca2+ influx occurs through astore-operated calciumentrymechanism in the plasmamem-brane resulting in further depletion of Ca2+ in the acrosome[68 69]
In support of the aforesaid studies several recent stud-ies on the same topic have also hypothesized that afterthe morphological maturation of spermatozoa for sperm-oocyte fusion [Ca2+]i decreases because acrosome-reactedspermatozoa release a substantial amount of Ca2+ from theirinner cell layers [71 72] Ca2+-mediated capacitation and theacrosome reaction have been illustrated in Figure 2 for betterunderstandingHowever for amore in-depth understandingwe recommend reading some excellent reviews on this topic[63 67 73ndash77]
A review of the literature showed that several spermproteins potentially regulate the Ca2+-dependent capacita-tion and the acrosome reaction in mammalian spermato-zoa [4 5 39] However how these proteins regulate theCa2+ influx in spermatozoa is a matter that remains tobe elucidated Breitbart et al [18] reported that forma-tion of F-actin mostly depends on PKA protein tyrosinephosphorylation and phospholipase D activation duringcapacitation Ca2+ is one of the principle regulators of
6 BioMed Research International
Acrosome reaction
Proteinphosphorylation
Flagellar beatinghyperactivation
CatS
per
sAC
ATP
cAMP
CNG
Calm
MPL
PLD
PLD
Acrosomalenzymes
Acrosome
IP3R
ZRK
G-proteins
Flagellarbeating
PLC
Aniontransport
sAC
mAC
T-type calcium channel
PKC
PTK PKG
mGC
GTPcGMP
sGC
CO NO
NO
CO
STK
PDE3
Nucleus
IP3
Fallopian tube (follicular fluid)
Sperm headPrinciple piece
Transporter
[pH]i
ZP3Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+ Ca2+
Ca2+G-
proteins
H+
H+
Ca2+HCO minus3
ZP3
ATP
cAMPPKA
Figure 2 Schematic diagram showing the mechanism of Ca2+ regulated hyperactivation capacitation and the acrosome reaction ofspermatozoa which are three principal events of fertilization Ca2+ together with ZP3 (zona pellucida glycoprotein-3) exhibits the mostimportant role in sperm binding and acrosomal reaction Ca2+ triggers the zona pellucida (ZP) receptors of cell membrane that activateG-proteins in the sperm head Activated G-proteins stimulate the H+ transporter to increase intracellular pH ultimately inducing theacrosomal reaction and hyperactivation by catalyzing the acrosomal enzymes [91] Cyclic adenosine monophosphate (cAMP) and cyclicguanosine monophosphate (cGMP) are produced from adenosine triphosphate (ATP) owing to enzymatic catalysis by soluble adenylatecyclase (sAC) and guanylate cyclase (sGC) respectively in mature spermatozoa The bicarbonate ions activate the sAC however follicularfluid also stimulates the sAC through release of Ca2+ ions via the CatSper channel (principal piece) However G-protein mediated signaltransduction activates sAC and phospholipase-C (PLC) that ultimately causes tyrosine phosphorylation [51 92] which is responsible forevents such as capacitation and the acrosomal reaction Likewise extracellular signals such as nitric oxide (NO) and carbon monoxide (CO)stimulate membrane-bound GC (mGC) and sGC respectively to synthesize cGMP Increases in cGMP level evoke a concomitant increasein cAMP by inhibiting its PDE3 However the increased Ca2+ level can also directly catalyze cAMP [93 94] Activated sAC sGC and PLCstimulate the generation of the second messengersrsquo inositol trisphosphate (IP3) like cAMP cGMP The IP3 binds to the IP3 receptor (IP3R)to increase [Ca2+]i via the release of the [Ca2+]i storage ions Concurrently the second messengers activate protein kinases (PKA PKCand PKG) in turn gating ions through the T-type calcium channels cyclic-nucleotide gated ion channel (CNG) and so on that togetherwith the activation of protein tyrosine kinases (PTK) and serinethreonine protein kinase (STK) cause increased protein phosphorylation[93 94] Additionally the CatSper Ca2+ activates calmodulin (Calm) phospholipase-A (PLA) and phospholipase-D (PLD) with increasedgeneration of other second messengers during the acrosome reaction Ca2+ influx together with increased protein phosphorylation bringsabout the capacitation response that is responsible for the waveform asymmetry of motility termed hyperactivation during fertilization Bothhyperactivation and the acrosomal reaction boost flagellar beating ultimately resulting in the penetration of the outer egg coat and subsequentfertilization of the mature ovum [91ndash95]
capacitation and it is therefore tempting to hypothesizethat organizational modification of F-actin in spermatozoatogether with interacting with other sperm proteins haspotential influence on Ca2+ influx A similar finding hasbeen established more precisely by another study [78] whereboar sperm capacitation was studied by combined applica-tion of computational and experimental approaches Theseauthors reported that the boar spermatozoa capacitationnetwork contains several connecting cascades whereas onlythree nodes bound to all the subcellular compartments are
involved in spermatozoa postejaculatory signaling such as[Ca2+]i ATP and actin polymerization Removal of the actinpolymerization node from this aforesaid network causesdisorganization of the network topography and affects capac-itation and this has been confirmed by zona pellucida-induced capacitation and the acrosomal reaction in an in vitrodemonstration [78]
In another study Patrat et al [79] showed that pro-gesterone (P
4) that is secreted by cumulus cells directly
acts on the sperm plasma membrane and triggers the
BioMed Research International 7
AVP
AVPR2 CNR1CATSPE
RB
Motilityhyperactivation CATSPER1
CATSPER2
CATSPERG
Calmoduli-n MARCKS
BASP1
MAPK1
PRKCSH
CALRITPR1
ADCY
SRCDLD
HSPA2
AKAP3
RON1
CABYR
SPINLW1
SEMG1
RAPGEF4
TNF
PLCZ1
Ca +2
PLC
PKC
PIP2NSF
PTPN1
SNAPreceptor
SYT6
RAB3A
RAPGEF3 GEF
SLC8A1
NPR1
NPPA
Chemotaxis
Progest-erone
Proteintyrosinekinase
Protein
Cell processFunctional class
Small molecule
RegulationMol synthesisMol transportBindingProt modifcation
Figure 3 Schematic representation of interactions among sim35 proteins related to Ca2+ regulated spermatozoa hyperactivation andchemotaxisThefigurewas produced by use of Pathway Studio (90) following theMedScanReader (50) protein search fromPubMeddatabase[12]
intracellular signals and enzymatic pathways involved inthe acrosome reaction P
4regulates the acrosome reaction
and is mediated by a compulsory Ca2+ increase This studyfound that P
4induced the activation of GiGo protein-
coupled and protein tyrosine kinase receptors and it affectedcapacitation and the acrosome reaction In contrast Ca2+ reg-ulated exocytosis of spermatozoa requires active acrosomalproteins such as N-ethylmaleimide-sensitive factor (NSF)[66] Additionally the same research team showed that the
ras-related protein Rab-3A (RAB3A) is also necessary forCa2+-dependent exocytosis Interestingly Rab3A activationof acrosomal exocytosis requires active NSF Thereforeprotein-protein interaction might also play a potential rolein regulating Ca2+ influx All of these observations seemto be consistent with the idea that Ca2+ functions areregulated by sperm proteins during fertilization Howeverthe key question is how do these proteins modify Ca2+ influxin spermatozoa
8 BioMed Research International
SEMG1AVP
PGR
CATSPER1
CATSPER2
ZP3
CDC42
DNM1
CHRNA7
PLCB1
PLCD4
GNA13
F-actin
Spinkl
GSN
IP3
PIP2
Protein tyrosinekinase
MAPK1
cAMPSperm
capacitation
PKAPRKAR2A
ADCY10
PLC
PKC
Actinflament
PLCZ1
L-tyrosine
CAMP
AKAP3ADCY
RON1LY6G6C
SLC8A1
OR10J5
CAMK2B
SRC
PLCG1
CS
ABCC1
NPPA
ABCB1
CAPN1
VDAC
Phospholipa-
IntracellularpH
Acrosome reaction
se A2
Ca2+
Fertilization
Protein
Cell processFunctional class Small molecule
Regulation
Complex
Axone-mal
dynein
Figure 4 Schematic representation of interactions among sim40 proteins related to Ca2+ regulated spermatozoa capacitation the acrosomereaction and fertilization The figure was produced by use of Pathway Studio (90) following the MedScan Reader (50) protein search fromPubMed database [12]
Recently in our laboratory we used mice spermatozoato evaluate the interrelationship of proteins related to Ca2+influx includingUQCRC2 [39] arginine vasopressin [5] andVDACs [4] and evaluate their effects on capacitation andthe acrosome reaction It is likely that a sustained phase ofCa2+ is required for fertilization andmight be regulated by thecomplex interaction of numerous sperm proteins Thereforestudies to identify proteins that might have the ability toinduce such a change are worth undertaking Application
of Pathway Studio helped us represent over 40 proteinsthat are potentially implicated in Ca2+ mediated regulationof capacitation the acrosome reaction and male fertility(Figure 4)
33 Ca2+ Influx and Postfertilization Egg Activation in Con-text of Sperm Proteome Ca2+ influx in spermatozoa is notonly important for sperm maturation but it is also equally
BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

2 BioMed Research International
individual sperm proteomes have described the importanceof spermatozoal posttranslational modifications and theirability to induce physiological changes as a prerequisite forsuccessful fertilization
Torres-Flores et al [16] have shown that human sperma-tozoa exposed to the phosphodiesterase inhibitor papaverinecause activation of protein kinase A (PKA) and stimulate theprogesterone-induced Ca2+ influx via the cyclic adenosinemonophosphate- (cAMP-) dependent pathway Althoughthese authors did not evaluate the relationship between invitro fertility and Ca2+ influx changes in intracellular pHand increased tyrosine phosphorylation ultimately providea potential clue regarding sperm fertility competence Inanother study to evaluate hamster spermatozoa capacitationcapability comparative association was observed betweenpyruvate dehydrogenase A Ca2+ influx cAMP and reactiveoxygen species [17] Additionally Breitbart et al [18] reportedthat polymerization of globular- (G-) actin to filamentous-(F-) actin occurs during capacitation As capacitation andthe acrosome reaction are Ca2+-mediated events [4 5] onecan without considering further signaling cascade assumethat remodeling the actin structure might be linked with theregulation of Ca2+ influx in spermatozoa
Recently in our laboratory we found that the manip-ulation of sperm proteins such as ubiquinol-cytochrome-creductase core protein 2 (UQCRC2) [39] voltage-dependentanion channels proteins (VDACs) [4] and arginine vaso-pressin [5] could control the Ca2+ influx in spermatozoa andregulate capacitation the acrosome reaction and fertilityTherefore design and construction of a similar study withmost of the identified sperm proteins available from severalprotein databases might provide a more realistic insight intothe Ca2+ influx protein functions and fertility The presentwork reviews the latest information published by other labo-ratories as well as our research team on the aforementionedaspects of spermatozoa and their potential implications fordiagnosis and prognosis of male fertility
2 Mechanism of Ca2+ Influx in MammalianSpermatozoa
The ultimate goal of fertilization of mammalian sperm is tofuse with and deliver their geneticmaterials into an oocyte [240 41] For fertilization to occur completely the spermatozoamust experience various obstacles both in vitro and in vivo[40 41] Ca2+ ions act as central signaling molecules oncethey enter the spermatozoa they exert allosteric regulatoryeffects on enzymes and many proteins [10 21 42] Indeednumerous elegant research findings have contributed signif-icantly to our understanding of the molecular signaling ofCa2+ influx especially through monitoring the activity ofindividual cells However most of the studies are discreteand often do not represent a cumulative idea This sectionpresents a compilation of some basic information regardingthe Ca2+ entry mechanism into mammalian spermatozoa byrecapitulating scientific evidence
The literature reviewed shows that the primary source ofCa2+ for spermatozoa is the external environment the fallop-ian tube in the female reproductive tract (in vivo) and culturemedia (in vitro) [8] and simultaneously increasing [Ca2+]iregulates the release of Ca2+ into the cellTherefore howCa2+crosses into cells through the sperm plasma membrane isa matter of paramount importance In eukaryotic cells theCa2+ influx occurs through specificCa2+ permeable ion chan-nel proteins located on the plasmamembrane [43 44] such asclassical voltage-gated (high and low) Ca2+ (Cavs) transientreceptor potential (TRP) and cyclic nucleotide-gated (CNG)channels [9] Recently Ren and Xia have proposed fourcriteria to identify sperm ion channel proteins detectabilityin sperm preferably with knockout sperm as a negativecontrol ability to produce ion channel current detectable bypatch-clamp recording blocking of the channels that impairsnormal sperm function and mutations of gene encoding theion channel proteins leading to sperm malfunctions [10]
The CatSper family of channels is the newest and onlyfamily of voltage-gated Ca2+ channels that meets most of theaforementioned criteria and essentially regulates Ca2+ entryinto cells and is therefore crucial for sperm fertility [9 45]Four pore-forming CatSper channel proteins CatSper 1ndash4and at least two auxiliary subunits CatSper120573 and CatSper120574have been identified in a wide range of animals includinghumans andmice [46 47] Physiologically CatSpermembersare permeable to Ca2+whereas the CatSper knockdownsperm does not have the channel current that is detectedin the principal piece of wild-type sperm [20 48] Mostof the channel proteins including CatSper members havebeen identified in the principal piece of spermatozoa [2046 47 49] (Figure 1) Although the explanation of suchsubcellular localization is still debated it might be becauseof interactions among the channel proteins and with theauxiliary subunits although a further study is needed toresolve this issue Collectively these proteins play a key rolein various cellular processes via regulation of the membranepotential and intracellular ionic balance Carlson et al [50]and Quill et al [51] have conclusively proved that CatSper1and CatSper2 null mice are sterile owing to their inabilityto generate the sperm-hyperactivated motility prerequisitefor penetration of an oocyte extracellular matrix In effectthe complete or partial absence of single or multiple Ca2+channels is responsible for infertility or subfertility althoughtheir underlying signaling cascade has not been properlystudied
Previously it has been reported that CatSper-dependentincreases of [Ca2+]i in spermatozoa are induced byseveral psychological stimuli such as cyclic nucleotides(eg cAMP and cGMP) [29 30 52] soluble adenylylcyclase [29 52] zona pellucida glycoprotein [34 35 38]serum albumin [37 38] secretion of cumulus oophorus[38] intracellular alkalization [3 53] and pH [6 21] Arecent study showed that endocrine disruptors such as pp1015840-dichlorodiphenyldichloroethylene (pp1015840-DDE) promotedCa2+ entry into spermatozoa by activating CatSper channelseven at a physiological concentration [36] In additionseveral other components are also known to play an
BioMed Research International 3
GPC
RC
Alkalinization
CatS
per
120574120573
Ca2+
[Ca 2+]i
sACY
ATP cAMPcGMP
ZP receptors
HCN
Stimulate
Opening
CN
G
K+
Ca2+
Progesterone
Spermatozoa
Principalpiece
H+
Extracellular space
Intracellular space
Na+
NBC
Na+
Follicle
nHCO3minus
HCO3minus
Hv1
pH
Figure 1 Possible signal transduction mechanisms of mammalian sperm Ca2+ influx through the Ca2+ permeable channel proteinsPreviously published studies were used as references to summarize the list of channel proteins in spermatozoa The channel proteins arelocalized mainly in the principle piece of spermatozoa The follicular fluid and several factors in the fallopian tube (in vitromedia) stimulatethe receptors for spermatozoa Ca2+ influx Ca2+ influx in spermatozoa is principally regulated by CatSper channels however the possibleinteraction between other channels that are responsible for controlling the opening of CatSper and allowing the Ca2+ into cells is indicatedby arrow signs (red circle) The different channel proteins that are depicted in the diagram include the Na+-coupled HCO
3
minus transporter(NBC) family soluble adenylyl cyclase (sACY) adenosine triphosphate (ATP) cyclic adenosine monophosphate (cAMP) cyclic guanosinemonophosphate (cGMP) cyclic nucleotide-gated ion channel (CNG) hyperpolarization-activated cyclic nucleotide-gated channel (HCN)zona pellucida (ZP) the voltage-gated proton channel (Hv1) glutamate receptor family class-C (GPCRC) and an unknown mechanism ()
important role in Ca2+ influx mechanisms in mammalianspermatozoa by regulating the opening of CatSper membersincluding the flagellar voltage-gated proton channel (Hv1)[21] Ca2+-ATPase pump [33] several cyclic nucleotide-gatedion channels (CNG) [27 54] hyperpolarization-activatedcyclic nucleotide-gated (HCN) channels [27] and G-proteincoupled receptors (GPCRs)
A hypothetical signaling cascade of Ca2+ influx pathwaysand interaction of several channel proteins is depicted inFigure 1 Although the functions of several ion channelproteins together with their concurrent relationship withnumerous stimuli have been well studied [21 27 38] severalfundamental questions remain unanswered for examplehowdo these channelsstimuli regulate theCa2+ influx duringspermatozoa processes such as capacitation the acrosomereaction and fertilization Do they work alone or togetherwith other channel proteins to regulate Ca2+ influx More-over which other parameters that remain undetected couldhave an effect on Ca2+ influx Therefore future researchshould focus on resolving these issues Table 1 summarizesthe proposed effect of Ca2+ ion channels and their physio-logical role that ultimately helps Ca2+ influx into mammalianspermatozoa
3 Effect of Ca2+ Influx on Male Fertility
Ca2+ triggers multiple physiological events in sperma-tozoa such as hyperactivation chemotaxis capacitationand the acrosomal reaction all of which are essential for
successful fertilization In mammalian spermatozoa numer-ous Ca2+ permeable channel proteins control intracellularpH and the pH-dependent Ca2+ influx is measured by thewhole-cell patch clamp technique [9 20] A review of theliterature showed that a potential functional interaction existsbetween the sperm proteins and Ca2+ permeable chan-nel proteins thus modulating the Ca2+ influx mechanism[4 5 39] and playing a vital role in adjusting male fertilityHowever themechanism bywhich Ca2+ triggers intracellularsignaling to regulate physiological events in spermatozoaand the role of sperm proteins in adjustment of Ca2+influx into cells remains unclear This topic is emphasizedbelow
31 Ca2+ Influx Sperm Hyperactivation Chemotaxis andProtein Functions In general mature spermatozoa are heldimmotile within the epididymis However they quickly beginto swim following release This is known as activation ofmotility and is characterized by symmetrical flagellar beats[55 56]The terms sperm activation and hyperactivation havequite different meanings The spermatozoa become hyperac-tivatedwhen the amplitude of the flagellar bend increases andproduces a highly asymmetrical beat In vivo hyperactivationof spermatozoa facilitates the release of sperm from oviductalstorage and boosts them through mucus in the oviductallumen and matrix of the cumulus oophorus during fertiliza-tion [7] In contrast chemotaxis is a formof spermmovementin which spermatozoa move toward a concentration gradientof a chemoattractant released from the oocyte [57 58]
4 BioMed Research International
Table 1 Summary of published works on ion channels and physiological stimuli of mammalian spermatozoa that regulate the Ca2+ influxmechanism
Name ofchannelstimuli
Localization onspermatozoaavailability Role in Ca2+ influx Role in sperm
physiologyEffect of knockingdownabsence Reference
CatSperCatSper 1CatSper 2CatSper 3CatSper 4CatSper120573CatSper120574
Principal piece Regulates Ca2+ influxCa2+ uptakehyperactivated
motilitySterile Barratt and Publicover
[19] Qi et al [20]
Hv1 Principal pieceIntracellular pHalkalization thus
stimulate Ca2+ influx
Extrudesprotons from
flagellaalkalization
Fertile Lishko et al [21]Lishko et al [22]
119868ATP Midpiece Selectively transports theCa2+
Ca2+ influxalkalization Fertile Navarro et al [23]
TRPC Principal piecemidpiece
Stimulates opening ofCatSper
Ca2+ influx celldepolarization Fertile Gees et al [24]
Castellano et al [25]
CNG Sperm flagellum headStimulates opening of
CatSper viacAMPcGMP
Ca2+ influx Fertile Biel and Michalakis [26]
HCN Flagellum Depolarization andopening of CatSper Ca2+ influx Fertile Wiesner et al [27]
SOC Plasma membrane ZP-induced Ca2+ influx Spermchemotactic Subfertile Yoshida et al [28]
sACYcAMPcGMP
Intracellular space andcell membrane
Activates CatSper CNGand HCN to regulate
Ca2+ influx
Ca2+ influxalkalization Sterile Esposito et al [29]
Hess et al [30]
GPCR(s) Principal piecemidpiece
ZP-induced Ca2+ influxincreases in [Ca2+]i
Maintainsfertilization Subfertile Fukami et al [31]
Fukami et al [32]
PLC120575 Acrosome ZP induced increases in[Ca2+]i
Ca2+ influx Subfertile Fukami et al [32]
Ca2+-ATPasepump Principal piece Intracellular pH and
alkalizationCa2+ influxcapacitation
Motility loss resultsin infertility Wennemuth et al [33]
ZPglycoproteins Follicle Induced Ca2+-dependent
increase in [Ca2+]iHyperactivationcapacitation
Delayedcapacitation
Florman [34]Florman et al [35]
Endocrinedisruptor(pp1015840-DDE)
Female reproductivetract Activates CatSper Ca2+ influx
Motility lossdelayed
capacitationTavares et al [36]
BSA Extracellular space Similar to ZPglycoprotein
In vitrocapacitation
Motility losssubfertility
Xia and Ren [37]Bailey and Storey [38]
Oviductal andfollicular fluid
Extracellular space(in vivo)
Ca2+-dependent increasein [Ca2+]i in sperm Ca2+ influx
Motility lossdelayed
capacitationXia and Ren [37]
Hv1 voltage-gated proton channel 119868ATP ATP-gated channel TRPC transient receptor potential channels CNG cyclic nucleotide-gated ion channel HCNhyperpolarization-activated cyclic nucleotide-gated channel SOC store-operated Ca2+ channel cAMP cyclic adenosine monophosphate cGMP cyclicguanosine monophosphate sACY soluble adenylyl cyclase GPCR glutamate receptor family class-C PLC120575 phospholipase C zeta ZP zona pellucida pp1015840-DDE pp1015840-dichlorodiphenyldichloroethylene BSA bovine serum albumin
However themolecular event that characterizes spermatozoachemotaxis is only partially known [57]
There is strong evidence to support that sperm hyper-activation and chemotaxis are required for penetrating thezona pellucida [48 57 59 60] Incubation of spermatozoawith an extracellular Ca2+ source induces hyperactivationin mammalian spermatozoa [61 62] and chemotaxis instarfish [57] In addition measuring cytoplasmic Ca2+ levels
by using the fluorescent Ca2+ indicator indo-1 proved thatspermatozoa hyperactivation is potentially regulated by Ca2+influx However it is unknown whether Ca2+ influx indepen-dently induces hyperactivationchemotaxis in mammalianspermatozoaHo and Suarez [56] proposed that spermhyper-activation induced by Ca2+ influx is mainly pH-dependentbecause sperm require a pH of 79ndash85 for hyperactivationwhereas activation can occur at a pH lt 70 The proposed
BioMed Research International 5
model of Ca2+-induced hyperactivation is represented inFigure 2
It has recently been found by our laboratory thattreatment of mouse spermatozoa with nutlin-3a a smallmolecule antagonist of the mouse double minute 2 repressorpotentially downregulates the functions of the ubiquinol-cytochrome-c reductase complex component UQCRC2 andcorrelated with significantly reduced [Ca2+]i and spermhyperactivation This study provided insight that the Ca2+influx in spermatozoa is partially regulated by UQCRC2protein Kwon et al [4] reported that blocking VDACwith 441015840-diisothiocyanostilbene-221015840-disulfonic acid (DIDS)significantly decreased sperm hyperactivation A signifi-cant decrease in [Ca2+]i was observed in (minus) DIDS con-ditions while [pH]i significantly increased in (minus) DIDSregardless of Ca2+ Simultaneously a significantly elevated[pH]i was observed in (+) Ca2+ This study providesstrong evidence that the modulation of Ca2+ influx byVDACs is pH-dependent which is consistent with the resultof a previous study by Ho and Suarez [56] Moreoveranother study proposed that deamino [Cys 1 d-ArgS] vaso-pressin (dDAVP) an AVPR2 agonist significantly decreasedsperm motility and intracellular pH but interestingly itincreased [Ca2+]i by regulating the function of argininevasopressin in mice spermatozoa However it remains tobe clarified as to why spermatozoa motility is decreased evenin increased [Ca2+]i conditions
On the basis of the findings of the aforementionedstudies it is tempting to hypothesize that spermatozoahyperactivation is mostly controlled by Ca2+ influx How-ever potential interactions exist between protein func-tions Therefore Ca2+ influx protein interaction andhyperactivation might give numerous different annotationsof upcoming research in this field We have illustrateda schematic representation of different signaling path-ways involving sperm proteins by using Pathway StudioThese proteins exhibit significant modifications to inducesperm hyperactivation and chemotaxis in spermatozoa byregulating Ca2+ influx (Figure 3)
32 Ca2+ Influx versus Capacitation the Acrosomal ReactionFertilization and Sperm Proteome Mammalian fertilizationis a species-specific episode that is accomplished by a complexset of molecular events To fertilize an oocyte multipleextreme changes occur in spermatozoa that begin from itsformation in the testes of the male reproductive tract to itspenetration and fusionwith an egg in the female reproductivetract Although spermatozoa are motile as well asmorphologically normal after ejaculation they are unable tofertilize an oocyte [59]They gain the fertilization ability onlyafter educating in the female reproductive tract [40] and themodifications that spermatozoa experience during this timeare collectively known as ldquocapacitationrdquo Only capacitatedspermatozoa can undergo the acrosome reaction throughbinding to the egg zona pellucida and they finally becomecapable of penetrating and fertilizing the egg [4 18 39]
The term ldquocapacitationrdquo was proposed by Austin in 1952[1] although this concept was initially described by bothChang and Austin in 1951 [2 41] In fact in vivo capacitationtakes place in the female reproductive tract however it is alsopossible to capacitate spermatozoa in vitro by using particularmedia containing appropriate electrolytes and pH [2] Inan elegant review Visconti summarized that the early stageof capacitation mainly comprises the bicarbonate-mediatedactivation of sperm motility whereas the late stages includeintracellular alkalinization increase in protein tyrosine phos-phorylation and preparation for the acrosomal reaction [63]These temporal differences in capacitation and the acrosomereaction require numerous mechanisms and Ca2+ influxplays a significant role in the process [63 64] Fraser [65]reported that capacitation is a comparatively slow event thatrequires several hours to complete and is mainly regulated bya modest rise in [Ca2+]i whereas the acrosome reaction is anexocytosis process that occurs very rapidly (within a minute)and is triggered by a large influx of [Ca2+]i [65 66]
Although the biochemical phenomenon of Ca2+regulated capacitation and the acrosome reaction havebeen known for the last two decades the molecular basisof this process is still poorly understood For capacitationthe cholesterol influx initially stimulates the elevationof [Ca2+]i and bicarbonate into the spermatozoa andfinally activates PKA and tyrosine phosphorylationrespectively via the production of the cAMP [66ndash68]In addition binding to the zona pellucida causes additionalactivation of cAMPPKA and protein kinase C (PKC)[68ndash70] Spermatozoa need [Ca2+]i influx to proceed furtherand they are believed to be activated by PKC through theopening of the calcium channels Interestingly PKA togetherwith a secondary messenger inositol trisphosphate activatescalcium channels localized in the outer acrosomalmembraneand increases the calcium concentration in the cytosolFurther increase of cytosolic Ca2+ influx occurs through astore-operated calciumentrymechanism in the plasmamem-brane resulting in further depletion of Ca2+ in the acrosome[68 69]
In support of the aforesaid studies several recent stud-ies on the same topic have also hypothesized that afterthe morphological maturation of spermatozoa for sperm-oocyte fusion [Ca2+]i decreases because acrosome-reactedspermatozoa release a substantial amount of Ca2+ from theirinner cell layers [71 72] Ca2+-mediated capacitation and theacrosome reaction have been illustrated in Figure 2 for betterunderstandingHowever for amore in-depth understandingwe recommend reading some excellent reviews on this topic[63 67 73ndash77]
A review of the literature showed that several spermproteins potentially regulate the Ca2+-dependent capacita-tion and the acrosome reaction in mammalian spermato-zoa [4 5 39] However how these proteins regulate theCa2+ influx in spermatozoa is a matter that remains tobe elucidated Breitbart et al [18] reported that forma-tion of F-actin mostly depends on PKA protein tyrosinephosphorylation and phospholipase D activation duringcapacitation Ca2+ is one of the principle regulators of
6 BioMed Research International
Acrosome reaction
Proteinphosphorylation
Flagellar beatinghyperactivation
CatS
per
sAC
ATP
cAMP
CNG
Calm
MPL
PLD
PLD
Acrosomalenzymes
Acrosome
IP3R
ZRK
G-proteins
Flagellarbeating
PLC
Aniontransport
sAC
mAC
T-type calcium channel
PKC
PTK PKG
mGC
GTPcGMP
sGC
CO NO
NO
CO
STK
PDE3
Nucleus
IP3
Fallopian tube (follicular fluid)
Sperm headPrinciple piece
Transporter
[pH]i
ZP3Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+ Ca2+
Ca2+G-
proteins
H+
H+
Ca2+HCO minus3
ZP3
ATP
cAMPPKA
Figure 2 Schematic diagram showing the mechanism of Ca2+ regulated hyperactivation capacitation and the acrosome reaction ofspermatozoa which are three principal events of fertilization Ca2+ together with ZP3 (zona pellucida glycoprotein-3) exhibits the mostimportant role in sperm binding and acrosomal reaction Ca2+ triggers the zona pellucida (ZP) receptors of cell membrane that activateG-proteins in the sperm head Activated G-proteins stimulate the H+ transporter to increase intracellular pH ultimately inducing theacrosomal reaction and hyperactivation by catalyzing the acrosomal enzymes [91] Cyclic adenosine monophosphate (cAMP) and cyclicguanosine monophosphate (cGMP) are produced from adenosine triphosphate (ATP) owing to enzymatic catalysis by soluble adenylatecyclase (sAC) and guanylate cyclase (sGC) respectively in mature spermatozoa The bicarbonate ions activate the sAC however follicularfluid also stimulates the sAC through release of Ca2+ ions via the CatSper channel (principal piece) However G-protein mediated signaltransduction activates sAC and phospholipase-C (PLC) that ultimately causes tyrosine phosphorylation [51 92] which is responsible forevents such as capacitation and the acrosomal reaction Likewise extracellular signals such as nitric oxide (NO) and carbon monoxide (CO)stimulate membrane-bound GC (mGC) and sGC respectively to synthesize cGMP Increases in cGMP level evoke a concomitant increasein cAMP by inhibiting its PDE3 However the increased Ca2+ level can also directly catalyze cAMP [93 94] Activated sAC sGC and PLCstimulate the generation of the second messengersrsquo inositol trisphosphate (IP3) like cAMP cGMP The IP3 binds to the IP3 receptor (IP3R)to increase [Ca2+]i via the release of the [Ca2+]i storage ions Concurrently the second messengers activate protein kinases (PKA PKCand PKG) in turn gating ions through the T-type calcium channels cyclic-nucleotide gated ion channel (CNG) and so on that togetherwith the activation of protein tyrosine kinases (PTK) and serinethreonine protein kinase (STK) cause increased protein phosphorylation[93 94] Additionally the CatSper Ca2+ activates calmodulin (Calm) phospholipase-A (PLA) and phospholipase-D (PLD) with increasedgeneration of other second messengers during the acrosome reaction Ca2+ influx together with increased protein phosphorylation bringsabout the capacitation response that is responsible for the waveform asymmetry of motility termed hyperactivation during fertilization Bothhyperactivation and the acrosomal reaction boost flagellar beating ultimately resulting in the penetration of the outer egg coat and subsequentfertilization of the mature ovum [91ndash95]
capacitation and it is therefore tempting to hypothesizethat organizational modification of F-actin in spermatozoatogether with interacting with other sperm proteins haspotential influence on Ca2+ influx A similar finding hasbeen established more precisely by another study [78] whereboar sperm capacitation was studied by combined applica-tion of computational and experimental approaches Theseauthors reported that the boar spermatozoa capacitationnetwork contains several connecting cascades whereas onlythree nodes bound to all the subcellular compartments are
involved in spermatozoa postejaculatory signaling such as[Ca2+]i ATP and actin polymerization Removal of the actinpolymerization node from this aforesaid network causesdisorganization of the network topography and affects capac-itation and this has been confirmed by zona pellucida-induced capacitation and the acrosomal reaction in an in vitrodemonstration [78]
In another study Patrat et al [79] showed that pro-gesterone (P
4) that is secreted by cumulus cells directly
acts on the sperm plasma membrane and triggers the
BioMed Research International 7
AVP
AVPR2 CNR1CATSPE
RB
Motilityhyperactivation CATSPER1
CATSPER2
CATSPERG
Calmoduli-n MARCKS
BASP1
MAPK1
PRKCSH
CALRITPR1
ADCY
SRCDLD
HSPA2
AKAP3
RON1
CABYR
SPINLW1
SEMG1
RAPGEF4
TNF
PLCZ1
Ca +2
PLC
PKC
PIP2NSF
PTPN1
SNAPreceptor
SYT6
RAB3A
RAPGEF3 GEF
SLC8A1
NPR1
NPPA
Chemotaxis
Progest-erone
Proteintyrosinekinase
Protein
Cell processFunctional class
Small molecule
RegulationMol synthesisMol transportBindingProt modifcation
Figure 3 Schematic representation of interactions among sim35 proteins related to Ca2+ regulated spermatozoa hyperactivation andchemotaxisThefigurewas produced by use of Pathway Studio (90) following theMedScanReader (50) protein search fromPubMeddatabase[12]
intracellular signals and enzymatic pathways involved inthe acrosome reaction P
4regulates the acrosome reaction
and is mediated by a compulsory Ca2+ increase This studyfound that P
4induced the activation of GiGo protein-
coupled and protein tyrosine kinase receptors and it affectedcapacitation and the acrosome reaction In contrast Ca2+ reg-ulated exocytosis of spermatozoa requires active acrosomalproteins such as N-ethylmaleimide-sensitive factor (NSF)[66] Additionally the same research team showed that the
ras-related protein Rab-3A (RAB3A) is also necessary forCa2+-dependent exocytosis Interestingly Rab3A activationof acrosomal exocytosis requires active NSF Thereforeprotein-protein interaction might also play a potential rolein regulating Ca2+ influx All of these observations seemto be consistent with the idea that Ca2+ functions areregulated by sperm proteins during fertilization Howeverthe key question is how do these proteins modify Ca2+ influxin spermatozoa
8 BioMed Research International
SEMG1AVP
PGR
CATSPER1
CATSPER2
ZP3
CDC42
DNM1
CHRNA7
PLCB1
PLCD4
GNA13
F-actin
Spinkl
GSN
IP3
PIP2
Protein tyrosinekinase
MAPK1
cAMPSperm
capacitation
PKAPRKAR2A
ADCY10
PLC
PKC
Actinflament
PLCZ1
L-tyrosine
CAMP
AKAP3ADCY
RON1LY6G6C
SLC8A1
OR10J5
CAMK2B
SRC
PLCG1
CS
ABCC1
NPPA
ABCB1
CAPN1
VDAC
Phospholipa-
IntracellularpH
Acrosome reaction
se A2
Ca2+
Fertilization
Protein
Cell processFunctional class Small molecule
Regulation
Complex
Axone-mal
dynein
Figure 4 Schematic representation of interactions among sim40 proteins related to Ca2+ regulated spermatozoa capacitation the acrosomereaction and fertilization The figure was produced by use of Pathway Studio (90) following the MedScan Reader (50) protein search fromPubMed database [12]
Recently in our laboratory we used mice spermatozoato evaluate the interrelationship of proteins related to Ca2+influx includingUQCRC2 [39] arginine vasopressin [5] andVDACs [4] and evaluate their effects on capacitation andthe acrosome reaction It is likely that a sustained phase ofCa2+ is required for fertilization andmight be regulated by thecomplex interaction of numerous sperm proteins Thereforestudies to identify proteins that might have the ability toinduce such a change are worth undertaking Application
of Pathway Studio helped us represent over 40 proteinsthat are potentially implicated in Ca2+ mediated regulationof capacitation the acrosome reaction and male fertility(Figure 4)
33 Ca2+ Influx and Postfertilization Egg Activation in Con-text of Sperm Proteome Ca2+ influx in spermatozoa is notonly important for sperm maturation but it is also equally
BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

BioMed Research International 3
GPC
RC
Alkalinization
CatS
per
120574120573
Ca2+
[Ca 2+]i
sACY
ATP cAMPcGMP
ZP receptors
HCN
Stimulate
Opening
CN
G
K+
Ca2+
Progesterone
Spermatozoa
Principalpiece
H+
Extracellular space
Intracellular space
Na+
NBC
Na+
Follicle
nHCO3minus
HCO3minus
Hv1
pH
Figure 1 Possible signal transduction mechanisms of mammalian sperm Ca2+ influx through the Ca2+ permeable channel proteinsPreviously published studies were used as references to summarize the list of channel proteins in spermatozoa The channel proteins arelocalized mainly in the principle piece of spermatozoa The follicular fluid and several factors in the fallopian tube (in vitromedia) stimulatethe receptors for spermatozoa Ca2+ influx Ca2+ influx in spermatozoa is principally regulated by CatSper channels however the possibleinteraction between other channels that are responsible for controlling the opening of CatSper and allowing the Ca2+ into cells is indicatedby arrow signs (red circle) The different channel proteins that are depicted in the diagram include the Na+-coupled HCO
3
minus transporter(NBC) family soluble adenylyl cyclase (sACY) adenosine triphosphate (ATP) cyclic adenosine monophosphate (cAMP) cyclic guanosinemonophosphate (cGMP) cyclic nucleotide-gated ion channel (CNG) hyperpolarization-activated cyclic nucleotide-gated channel (HCN)zona pellucida (ZP) the voltage-gated proton channel (Hv1) glutamate receptor family class-C (GPCRC) and an unknown mechanism ()
important role in Ca2+ influx mechanisms in mammalianspermatozoa by regulating the opening of CatSper membersincluding the flagellar voltage-gated proton channel (Hv1)[21] Ca2+-ATPase pump [33] several cyclic nucleotide-gatedion channels (CNG) [27 54] hyperpolarization-activatedcyclic nucleotide-gated (HCN) channels [27] and G-proteincoupled receptors (GPCRs)
A hypothetical signaling cascade of Ca2+ influx pathwaysand interaction of several channel proteins is depicted inFigure 1 Although the functions of several ion channelproteins together with their concurrent relationship withnumerous stimuli have been well studied [21 27 38] severalfundamental questions remain unanswered for examplehowdo these channelsstimuli regulate theCa2+ influx duringspermatozoa processes such as capacitation the acrosomereaction and fertilization Do they work alone or togetherwith other channel proteins to regulate Ca2+ influx More-over which other parameters that remain undetected couldhave an effect on Ca2+ influx Therefore future researchshould focus on resolving these issues Table 1 summarizesthe proposed effect of Ca2+ ion channels and their physio-logical role that ultimately helps Ca2+ influx into mammalianspermatozoa
3 Effect of Ca2+ Influx on Male Fertility
Ca2+ triggers multiple physiological events in sperma-tozoa such as hyperactivation chemotaxis capacitationand the acrosomal reaction all of which are essential for
successful fertilization In mammalian spermatozoa numer-ous Ca2+ permeable channel proteins control intracellularpH and the pH-dependent Ca2+ influx is measured by thewhole-cell patch clamp technique [9 20] A review of theliterature showed that a potential functional interaction existsbetween the sperm proteins and Ca2+ permeable chan-nel proteins thus modulating the Ca2+ influx mechanism[4 5 39] and playing a vital role in adjusting male fertilityHowever themechanism bywhich Ca2+ triggers intracellularsignaling to regulate physiological events in spermatozoaand the role of sperm proteins in adjustment of Ca2+influx into cells remains unclear This topic is emphasizedbelow
31 Ca2+ Influx Sperm Hyperactivation Chemotaxis andProtein Functions In general mature spermatozoa are heldimmotile within the epididymis However they quickly beginto swim following release This is known as activation ofmotility and is characterized by symmetrical flagellar beats[55 56]The terms sperm activation and hyperactivation havequite different meanings The spermatozoa become hyperac-tivatedwhen the amplitude of the flagellar bend increases andproduces a highly asymmetrical beat In vivo hyperactivationof spermatozoa facilitates the release of sperm from oviductalstorage and boosts them through mucus in the oviductallumen and matrix of the cumulus oophorus during fertiliza-tion [7] In contrast chemotaxis is a formof spermmovementin which spermatozoa move toward a concentration gradientof a chemoattractant released from the oocyte [57 58]
4 BioMed Research International
Table 1 Summary of published works on ion channels and physiological stimuli of mammalian spermatozoa that regulate the Ca2+ influxmechanism
Name ofchannelstimuli
Localization onspermatozoaavailability Role in Ca2+ influx Role in sperm
physiologyEffect of knockingdownabsence Reference
CatSperCatSper 1CatSper 2CatSper 3CatSper 4CatSper120573CatSper120574
Principal piece Regulates Ca2+ influxCa2+ uptakehyperactivated
motilitySterile Barratt and Publicover
[19] Qi et al [20]
Hv1 Principal pieceIntracellular pHalkalization thus
stimulate Ca2+ influx
Extrudesprotons from
flagellaalkalization
Fertile Lishko et al [21]Lishko et al [22]
119868ATP Midpiece Selectively transports theCa2+
Ca2+ influxalkalization Fertile Navarro et al [23]
TRPC Principal piecemidpiece
Stimulates opening ofCatSper
Ca2+ influx celldepolarization Fertile Gees et al [24]
Castellano et al [25]
CNG Sperm flagellum headStimulates opening of
CatSper viacAMPcGMP
Ca2+ influx Fertile Biel and Michalakis [26]
HCN Flagellum Depolarization andopening of CatSper Ca2+ influx Fertile Wiesner et al [27]
SOC Plasma membrane ZP-induced Ca2+ influx Spermchemotactic Subfertile Yoshida et al [28]
sACYcAMPcGMP
Intracellular space andcell membrane
Activates CatSper CNGand HCN to regulate
Ca2+ influx
Ca2+ influxalkalization Sterile Esposito et al [29]
Hess et al [30]
GPCR(s) Principal piecemidpiece
ZP-induced Ca2+ influxincreases in [Ca2+]i
Maintainsfertilization Subfertile Fukami et al [31]
Fukami et al [32]
PLC120575 Acrosome ZP induced increases in[Ca2+]i
Ca2+ influx Subfertile Fukami et al [32]
Ca2+-ATPasepump Principal piece Intracellular pH and
alkalizationCa2+ influxcapacitation
Motility loss resultsin infertility Wennemuth et al [33]
ZPglycoproteins Follicle Induced Ca2+-dependent
increase in [Ca2+]iHyperactivationcapacitation
Delayedcapacitation
Florman [34]Florman et al [35]
Endocrinedisruptor(pp1015840-DDE)
Female reproductivetract Activates CatSper Ca2+ influx
Motility lossdelayed
capacitationTavares et al [36]
BSA Extracellular space Similar to ZPglycoprotein
In vitrocapacitation
Motility losssubfertility
Xia and Ren [37]Bailey and Storey [38]
Oviductal andfollicular fluid
Extracellular space(in vivo)
Ca2+-dependent increasein [Ca2+]i in sperm Ca2+ influx
Motility lossdelayed
capacitationXia and Ren [37]
Hv1 voltage-gated proton channel 119868ATP ATP-gated channel TRPC transient receptor potential channels CNG cyclic nucleotide-gated ion channel HCNhyperpolarization-activated cyclic nucleotide-gated channel SOC store-operated Ca2+ channel cAMP cyclic adenosine monophosphate cGMP cyclicguanosine monophosphate sACY soluble adenylyl cyclase GPCR glutamate receptor family class-C PLC120575 phospholipase C zeta ZP zona pellucida pp1015840-DDE pp1015840-dichlorodiphenyldichloroethylene BSA bovine serum albumin
However themolecular event that characterizes spermatozoachemotaxis is only partially known [57]
There is strong evidence to support that sperm hyper-activation and chemotaxis are required for penetrating thezona pellucida [48 57 59 60] Incubation of spermatozoawith an extracellular Ca2+ source induces hyperactivationin mammalian spermatozoa [61 62] and chemotaxis instarfish [57] In addition measuring cytoplasmic Ca2+ levels
by using the fluorescent Ca2+ indicator indo-1 proved thatspermatozoa hyperactivation is potentially regulated by Ca2+influx However it is unknown whether Ca2+ influx indepen-dently induces hyperactivationchemotaxis in mammalianspermatozoaHo and Suarez [56] proposed that spermhyper-activation induced by Ca2+ influx is mainly pH-dependentbecause sperm require a pH of 79ndash85 for hyperactivationwhereas activation can occur at a pH lt 70 The proposed
BioMed Research International 5
model of Ca2+-induced hyperactivation is represented inFigure 2
It has recently been found by our laboratory thattreatment of mouse spermatozoa with nutlin-3a a smallmolecule antagonist of the mouse double minute 2 repressorpotentially downregulates the functions of the ubiquinol-cytochrome-c reductase complex component UQCRC2 andcorrelated with significantly reduced [Ca2+]i and spermhyperactivation This study provided insight that the Ca2+influx in spermatozoa is partially regulated by UQCRC2protein Kwon et al [4] reported that blocking VDACwith 441015840-diisothiocyanostilbene-221015840-disulfonic acid (DIDS)significantly decreased sperm hyperactivation A signifi-cant decrease in [Ca2+]i was observed in (minus) DIDS con-ditions while [pH]i significantly increased in (minus) DIDSregardless of Ca2+ Simultaneously a significantly elevated[pH]i was observed in (+) Ca2+ This study providesstrong evidence that the modulation of Ca2+ influx byVDACs is pH-dependent which is consistent with the resultof a previous study by Ho and Suarez [56] Moreoveranother study proposed that deamino [Cys 1 d-ArgS] vaso-pressin (dDAVP) an AVPR2 agonist significantly decreasedsperm motility and intracellular pH but interestingly itincreased [Ca2+]i by regulating the function of argininevasopressin in mice spermatozoa However it remains tobe clarified as to why spermatozoa motility is decreased evenin increased [Ca2+]i conditions
On the basis of the findings of the aforementionedstudies it is tempting to hypothesize that spermatozoahyperactivation is mostly controlled by Ca2+ influx How-ever potential interactions exist between protein func-tions Therefore Ca2+ influx protein interaction andhyperactivation might give numerous different annotationsof upcoming research in this field We have illustrateda schematic representation of different signaling path-ways involving sperm proteins by using Pathway StudioThese proteins exhibit significant modifications to inducesperm hyperactivation and chemotaxis in spermatozoa byregulating Ca2+ influx (Figure 3)
32 Ca2+ Influx versus Capacitation the Acrosomal ReactionFertilization and Sperm Proteome Mammalian fertilizationis a species-specific episode that is accomplished by a complexset of molecular events To fertilize an oocyte multipleextreme changes occur in spermatozoa that begin from itsformation in the testes of the male reproductive tract to itspenetration and fusionwith an egg in the female reproductivetract Although spermatozoa are motile as well asmorphologically normal after ejaculation they are unable tofertilize an oocyte [59]They gain the fertilization ability onlyafter educating in the female reproductive tract [40] and themodifications that spermatozoa experience during this timeare collectively known as ldquocapacitationrdquo Only capacitatedspermatozoa can undergo the acrosome reaction throughbinding to the egg zona pellucida and they finally becomecapable of penetrating and fertilizing the egg [4 18 39]
The term ldquocapacitationrdquo was proposed by Austin in 1952[1] although this concept was initially described by bothChang and Austin in 1951 [2 41] In fact in vivo capacitationtakes place in the female reproductive tract however it is alsopossible to capacitate spermatozoa in vitro by using particularmedia containing appropriate electrolytes and pH [2] Inan elegant review Visconti summarized that the early stageof capacitation mainly comprises the bicarbonate-mediatedactivation of sperm motility whereas the late stages includeintracellular alkalinization increase in protein tyrosine phos-phorylation and preparation for the acrosomal reaction [63]These temporal differences in capacitation and the acrosomereaction require numerous mechanisms and Ca2+ influxplays a significant role in the process [63 64] Fraser [65]reported that capacitation is a comparatively slow event thatrequires several hours to complete and is mainly regulated bya modest rise in [Ca2+]i whereas the acrosome reaction is anexocytosis process that occurs very rapidly (within a minute)and is triggered by a large influx of [Ca2+]i [65 66]
Although the biochemical phenomenon of Ca2+regulated capacitation and the acrosome reaction havebeen known for the last two decades the molecular basisof this process is still poorly understood For capacitationthe cholesterol influx initially stimulates the elevationof [Ca2+]i and bicarbonate into the spermatozoa andfinally activates PKA and tyrosine phosphorylationrespectively via the production of the cAMP [66ndash68]In addition binding to the zona pellucida causes additionalactivation of cAMPPKA and protein kinase C (PKC)[68ndash70] Spermatozoa need [Ca2+]i influx to proceed furtherand they are believed to be activated by PKC through theopening of the calcium channels Interestingly PKA togetherwith a secondary messenger inositol trisphosphate activatescalcium channels localized in the outer acrosomalmembraneand increases the calcium concentration in the cytosolFurther increase of cytosolic Ca2+ influx occurs through astore-operated calciumentrymechanism in the plasmamem-brane resulting in further depletion of Ca2+ in the acrosome[68 69]
In support of the aforesaid studies several recent stud-ies on the same topic have also hypothesized that afterthe morphological maturation of spermatozoa for sperm-oocyte fusion [Ca2+]i decreases because acrosome-reactedspermatozoa release a substantial amount of Ca2+ from theirinner cell layers [71 72] Ca2+-mediated capacitation and theacrosome reaction have been illustrated in Figure 2 for betterunderstandingHowever for amore in-depth understandingwe recommend reading some excellent reviews on this topic[63 67 73ndash77]
A review of the literature showed that several spermproteins potentially regulate the Ca2+-dependent capacita-tion and the acrosome reaction in mammalian spermato-zoa [4 5 39] However how these proteins regulate theCa2+ influx in spermatozoa is a matter that remains tobe elucidated Breitbart et al [18] reported that forma-tion of F-actin mostly depends on PKA protein tyrosinephosphorylation and phospholipase D activation duringcapacitation Ca2+ is one of the principle regulators of
6 BioMed Research International
Acrosome reaction
Proteinphosphorylation
Flagellar beatinghyperactivation
CatS
per
sAC
ATP
cAMP
CNG
Calm
MPL
PLD
PLD
Acrosomalenzymes
Acrosome
IP3R
ZRK
G-proteins
Flagellarbeating
PLC
Aniontransport
sAC
mAC
T-type calcium channel
PKC
PTK PKG
mGC
GTPcGMP
sGC
CO NO
NO
CO
STK
PDE3
Nucleus
IP3
Fallopian tube (follicular fluid)
Sperm headPrinciple piece
Transporter
[pH]i
ZP3Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+ Ca2+
Ca2+G-
proteins
H+
H+
Ca2+HCO minus3
ZP3
ATP
cAMPPKA
Figure 2 Schematic diagram showing the mechanism of Ca2+ regulated hyperactivation capacitation and the acrosome reaction ofspermatozoa which are three principal events of fertilization Ca2+ together with ZP3 (zona pellucida glycoprotein-3) exhibits the mostimportant role in sperm binding and acrosomal reaction Ca2+ triggers the zona pellucida (ZP) receptors of cell membrane that activateG-proteins in the sperm head Activated G-proteins stimulate the H+ transporter to increase intracellular pH ultimately inducing theacrosomal reaction and hyperactivation by catalyzing the acrosomal enzymes [91] Cyclic adenosine monophosphate (cAMP) and cyclicguanosine monophosphate (cGMP) are produced from adenosine triphosphate (ATP) owing to enzymatic catalysis by soluble adenylatecyclase (sAC) and guanylate cyclase (sGC) respectively in mature spermatozoa The bicarbonate ions activate the sAC however follicularfluid also stimulates the sAC through release of Ca2+ ions via the CatSper channel (principal piece) However G-protein mediated signaltransduction activates sAC and phospholipase-C (PLC) that ultimately causes tyrosine phosphorylation [51 92] which is responsible forevents such as capacitation and the acrosomal reaction Likewise extracellular signals such as nitric oxide (NO) and carbon monoxide (CO)stimulate membrane-bound GC (mGC) and sGC respectively to synthesize cGMP Increases in cGMP level evoke a concomitant increasein cAMP by inhibiting its PDE3 However the increased Ca2+ level can also directly catalyze cAMP [93 94] Activated sAC sGC and PLCstimulate the generation of the second messengersrsquo inositol trisphosphate (IP3) like cAMP cGMP The IP3 binds to the IP3 receptor (IP3R)to increase [Ca2+]i via the release of the [Ca2+]i storage ions Concurrently the second messengers activate protein kinases (PKA PKCand PKG) in turn gating ions through the T-type calcium channels cyclic-nucleotide gated ion channel (CNG) and so on that togetherwith the activation of protein tyrosine kinases (PTK) and serinethreonine protein kinase (STK) cause increased protein phosphorylation[93 94] Additionally the CatSper Ca2+ activates calmodulin (Calm) phospholipase-A (PLA) and phospholipase-D (PLD) with increasedgeneration of other second messengers during the acrosome reaction Ca2+ influx together with increased protein phosphorylation bringsabout the capacitation response that is responsible for the waveform asymmetry of motility termed hyperactivation during fertilization Bothhyperactivation and the acrosomal reaction boost flagellar beating ultimately resulting in the penetration of the outer egg coat and subsequentfertilization of the mature ovum [91ndash95]
capacitation and it is therefore tempting to hypothesizethat organizational modification of F-actin in spermatozoatogether with interacting with other sperm proteins haspotential influence on Ca2+ influx A similar finding hasbeen established more precisely by another study [78] whereboar sperm capacitation was studied by combined applica-tion of computational and experimental approaches Theseauthors reported that the boar spermatozoa capacitationnetwork contains several connecting cascades whereas onlythree nodes bound to all the subcellular compartments are
involved in spermatozoa postejaculatory signaling such as[Ca2+]i ATP and actin polymerization Removal of the actinpolymerization node from this aforesaid network causesdisorganization of the network topography and affects capac-itation and this has been confirmed by zona pellucida-induced capacitation and the acrosomal reaction in an in vitrodemonstration [78]
In another study Patrat et al [79] showed that pro-gesterone (P
4) that is secreted by cumulus cells directly
acts on the sperm plasma membrane and triggers the
BioMed Research International 7
AVP
AVPR2 CNR1CATSPE
RB
Motilityhyperactivation CATSPER1
CATSPER2
CATSPERG
Calmoduli-n MARCKS
BASP1
MAPK1
PRKCSH
CALRITPR1
ADCY
SRCDLD
HSPA2
AKAP3
RON1
CABYR
SPINLW1
SEMG1
RAPGEF4
TNF
PLCZ1
Ca +2
PLC
PKC
PIP2NSF
PTPN1
SNAPreceptor
SYT6
RAB3A
RAPGEF3 GEF
SLC8A1
NPR1
NPPA
Chemotaxis
Progest-erone
Proteintyrosinekinase
Protein
Cell processFunctional class
Small molecule
RegulationMol synthesisMol transportBindingProt modifcation
Figure 3 Schematic representation of interactions among sim35 proteins related to Ca2+ regulated spermatozoa hyperactivation andchemotaxisThefigurewas produced by use of Pathway Studio (90) following theMedScanReader (50) protein search fromPubMeddatabase[12]
intracellular signals and enzymatic pathways involved inthe acrosome reaction P
4regulates the acrosome reaction
and is mediated by a compulsory Ca2+ increase This studyfound that P
4induced the activation of GiGo protein-
coupled and protein tyrosine kinase receptors and it affectedcapacitation and the acrosome reaction In contrast Ca2+ reg-ulated exocytosis of spermatozoa requires active acrosomalproteins such as N-ethylmaleimide-sensitive factor (NSF)[66] Additionally the same research team showed that the
ras-related protein Rab-3A (RAB3A) is also necessary forCa2+-dependent exocytosis Interestingly Rab3A activationof acrosomal exocytosis requires active NSF Thereforeprotein-protein interaction might also play a potential rolein regulating Ca2+ influx All of these observations seemto be consistent with the idea that Ca2+ functions areregulated by sperm proteins during fertilization Howeverthe key question is how do these proteins modify Ca2+ influxin spermatozoa
8 BioMed Research International
SEMG1AVP
PGR
CATSPER1
CATSPER2
ZP3
CDC42
DNM1
CHRNA7
PLCB1
PLCD4
GNA13
F-actin
Spinkl
GSN
IP3
PIP2
Protein tyrosinekinase
MAPK1
cAMPSperm
capacitation
PKAPRKAR2A
ADCY10
PLC
PKC
Actinflament
PLCZ1
L-tyrosine
CAMP
AKAP3ADCY
RON1LY6G6C
SLC8A1
OR10J5
CAMK2B
SRC
PLCG1
CS
ABCC1
NPPA
ABCB1
CAPN1
VDAC
Phospholipa-
IntracellularpH
Acrosome reaction
se A2
Ca2+
Fertilization
Protein
Cell processFunctional class Small molecule
Regulation
Complex
Axone-mal
dynein
Figure 4 Schematic representation of interactions among sim40 proteins related to Ca2+ regulated spermatozoa capacitation the acrosomereaction and fertilization The figure was produced by use of Pathway Studio (90) following the MedScan Reader (50) protein search fromPubMed database [12]
Recently in our laboratory we used mice spermatozoato evaluate the interrelationship of proteins related to Ca2+influx includingUQCRC2 [39] arginine vasopressin [5] andVDACs [4] and evaluate their effects on capacitation andthe acrosome reaction It is likely that a sustained phase ofCa2+ is required for fertilization andmight be regulated by thecomplex interaction of numerous sperm proteins Thereforestudies to identify proteins that might have the ability toinduce such a change are worth undertaking Application
of Pathway Studio helped us represent over 40 proteinsthat are potentially implicated in Ca2+ mediated regulationof capacitation the acrosome reaction and male fertility(Figure 4)
33 Ca2+ Influx and Postfertilization Egg Activation in Con-text of Sperm Proteome Ca2+ influx in spermatozoa is notonly important for sperm maturation but it is also equally
BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

4 BioMed Research International
Table 1 Summary of published works on ion channels and physiological stimuli of mammalian spermatozoa that regulate the Ca2+ influxmechanism
Name ofchannelstimuli
Localization onspermatozoaavailability Role in Ca2+ influx Role in sperm
physiologyEffect of knockingdownabsence Reference
CatSperCatSper 1CatSper 2CatSper 3CatSper 4CatSper120573CatSper120574
Principal piece Regulates Ca2+ influxCa2+ uptakehyperactivated
motilitySterile Barratt and Publicover
[19] Qi et al [20]
Hv1 Principal pieceIntracellular pHalkalization thus
stimulate Ca2+ influx
Extrudesprotons from
flagellaalkalization
Fertile Lishko et al [21]Lishko et al [22]
119868ATP Midpiece Selectively transports theCa2+
Ca2+ influxalkalization Fertile Navarro et al [23]
TRPC Principal piecemidpiece
Stimulates opening ofCatSper
Ca2+ influx celldepolarization Fertile Gees et al [24]
Castellano et al [25]
CNG Sperm flagellum headStimulates opening of
CatSper viacAMPcGMP
Ca2+ influx Fertile Biel and Michalakis [26]
HCN Flagellum Depolarization andopening of CatSper Ca2+ influx Fertile Wiesner et al [27]
SOC Plasma membrane ZP-induced Ca2+ influx Spermchemotactic Subfertile Yoshida et al [28]
sACYcAMPcGMP
Intracellular space andcell membrane
Activates CatSper CNGand HCN to regulate
Ca2+ influx
Ca2+ influxalkalization Sterile Esposito et al [29]
Hess et al [30]
GPCR(s) Principal piecemidpiece
ZP-induced Ca2+ influxincreases in [Ca2+]i
Maintainsfertilization Subfertile Fukami et al [31]
Fukami et al [32]
PLC120575 Acrosome ZP induced increases in[Ca2+]i
Ca2+ influx Subfertile Fukami et al [32]
Ca2+-ATPasepump Principal piece Intracellular pH and
alkalizationCa2+ influxcapacitation
Motility loss resultsin infertility Wennemuth et al [33]
ZPglycoproteins Follicle Induced Ca2+-dependent
increase in [Ca2+]iHyperactivationcapacitation
Delayedcapacitation
Florman [34]Florman et al [35]
Endocrinedisruptor(pp1015840-DDE)
Female reproductivetract Activates CatSper Ca2+ influx
Motility lossdelayed
capacitationTavares et al [36]
BSA Extracellular space Similar to ZPglycoprotein
In vitrocapacitation
Motility losssubfertility
Xia and Ren [37]Bailey and Storey [38]
Oviductal andfollicular fluid
Extracellular space(in vivo)
Ca2+-dependent increasein [Ca2+]i in sperm Ca2+ influx
Motility lossdelayed
capacitationXia and Ren [37]
Hv1 voltage-gated proton channel 119868ATP ATP-gated channel TRPC transient receptor potential channels CNG cyclic nucleotide-gated ion channel HCNhyperpolarization-activated cyclic nucleotide-gated channel SOC store-operated Ca2+ channel cAMP cyclic adenosine monophosphate cGMP cyclicguanosine monophosphate sACY soluble adenylyl cyclase GPCR glutamate receptor family class-C PLC120575 phospholipase C zeta ZP zona pellucida pp1015840-DDE pp1015840-dichlorodiphenyldichloroethylene BSA bovine serum albumin
However themolecular event that characterizes spermatozoachemotaxis is only partially known [57]
There is strong evidence to support that sperm hyper-activation and chemotaxis are required for penetrating thezona pellucida [48 57 59 60] Incubation of spermatozoawith an extracellular Ca2+ source induces hyperactivationin mammalian spermatozoa [61 62] and chemotaxis instarfish [57] In addition measuring cytoplasmic Ca2+ levels
by using the fluorescent Ca2+ indicator indo-1 proved thatspermatozoa hyperactivation is potentially regulated by Ca2+influx However it is unknown whether Ca2+ influx indepen-dently induces hyperactivationchemotaxis in mammalianspermatozoaHo and Suarez [56] proposed that spermhyper-activation induced by Ca2+ influx is mainly pH-dependentbecause sperm require a pH of 79ndash85 for hyperactivationwhereas activation can occur at a pH lt 70 The proposed
BioMed Research International 5
model of Ca2+-induced hyperactivation is represented inFigure 2
It has recently been found by our laboratory thattreatment of mouse spermatozoa with nutlin-3a a smallmolecule antagonist of the mouse double minute 2 repressorpotentially downregulates the functions of the ubiquinol-cytochrome-c reductase complex component UQCRC2 andcorrelated with significantly reduced [Ca2+]i and spermhyperactivation This study provided insight that the Ca2+influx in spermatozoa is partially regulated by UQCRC2protein Kwon et al [4] reported that blocking VDACwith 441015840-diisothiocyanostilbene-221015840-disulfonic acid (DIDS)significantly decreased sperm hyperactivation A signifi-cant decrease in [Ca2+]i was observed in (minus) DIDS con-ditions while [pH]i significantly increased in (minus) DIDSregardless of Ca2+ Simultaneously a significantly elevated[pH]i was observed in (+) Ca2+ This study providesstrong evidence that the modulation of Ca2+ influx byVDACs is pH-dependent which is consistent with the resultof a previous study by Ho and Suarez [56] Moreoveranother study proposed that deamino [Cys 1 d-ArgS] vaso-pressin (dDAVP) an AVPR2 agonist significantly decreasedsperm motility and intracellular pH but interestingly itincreased [Ca2+]i by regulating the function of argininevasopressin in mice spermatozoa However it remains tobe clarified as to why spermatozoa motility is decreased evenin increased [Ca2+]i conditions
On the basis of the findings of the aforementionedstudies it is tempting to hypothesize that spermatozoahyperactivation is mostly controlled by Ca2+ influx How-ever potential interactions exist between protein func-tions Therefore Ca2+ influx protein interaction andhyperactivation might give numerous different annotationsof upcoming research in this field We have illustrateda schematic representation of different signaling path-ways involving sperm proteins by using Pathway StudioThese proteins exhibit significant modifications to inducesperm hyperactivation and chemotaxis in spermatozoa byregulating Ca2+ influx (Figure 3)
32 Ca2+ Influx versus Capacitation the Acrosomal ReactionFertilization and Sperm Proteome Mammalian fertilizationis a species-specific episode that is accomplished by a complexset of molecular events To fertilize an oocyte multipleextreme changes occur in spermatozoa that begin from itsformation in the testes of the male reproductive tract to itspenetration and fusionwith an egg in the female reproductivetract Although spermatozoa are motile as well asmorphologically normal after ejaculation they are unable tofertilize an oocyte [59]They gain the fertilization ability onlyafter educating in the female reproductive tract [40] and themodifications that spermatozoa experience during this timeare collectively known as ldquocapacitationrdquo Only capacitatedspermatozoa can undergo the acrosome reaction throughbinding to the egg zona pellucida and they finally becomecapable of penetrating and fertilizing the egg [4 18 39]
The term ldquocapacitationrdquo was proposed by Austin in 1952[1] although this concept was initially described by bothChang and Austin in 1951 [2 41] In fact in vivo capacitationtakes place in the female reproductive tract however it is alsopossible to capacitate spermatozoa in vitro by using particularmedia containing appropriate electrolytes and pH [2] Inan elegant review Visconti summarized that the early stageof capacitation mainly comprises the bicarbonate-mediatedactivation of sperm motility whereas the late stages includeintracellular alkalinization increase in protein tyrosine phos-phorylation and preparation for the acrosomal reaction [63]These temporal differences in capacitation and the acrosomereaction require numerous mechanisms and Ca2+ influxplays a significant role in the process [63 64] Fraser [65]reported that capacitation is a comparatively slow event thatrequires several hours to complete and is mainly regulated bya modest rise in [Ca2+]i whereas the acrosome reaction is anexocytosis process that occurs very rapidly (within a minute)and is triggered by a large influx of [Ca2+]i [65 66]
Although the biochemical phenomenon of Ca2+regulated capacitation and the acrosome reaction havebeen known for the last two decades the molecular basisof this process is still poorly understood For capacitationthe cholesterol influx initially stimulates the elevationof [Ca2+]i and bicarbonate into the spermatozoa andfinally activates PKA and tyrosine phosphorylationrespectively via the production of the cAMP [66ndash68]In addition binding to the zona pellucida causes additionalactivation of cAMPPKA and protein kinase C (PKC)[68ndash70] Spermatozoa need [Ca2+]i influx to proceed furtherand they are believed to be activated by PKC through theopening of the calcium channels Interestingly PKA togetherwith a secondary messenger inositol trisphosphate activatescalcium channels localized in the outer acrosomalmembraneand increases the calcium concentration in the cytosolFurther increase of cytosolic Ca2+ influx occurs through astore-operated calciumentrymechanism in the plasmamem-brane resulting in further depletion of Ca2+ in the acrosome[68 69]
In support of the aforesaid studies several recent stud-ies on the same topic have also hypothesized that afterthe morphological maturation of spermatozoa for sperm-oocyte fusion [Ca2+]i decreases because acrosome-reactedspermatozoa release a substantial amount of Ca2+ from theirinner cell layers [71 72] Ca2+-mediated capacitation and theacrosome reaction have been illustrated in Figure 2 for betterunderstandingHowever for amore in-depth understandingwe recommend reading some excellent reviews on this topic[63 67 73ndash77]
A review of the literature showed that several spermproteins potentially regulate the Ca2+-dependent capacita-tion and the acrosome reaction in mammalian spermato-zoa [4 5 39] However how these proteins regulate theCa2+ influx in spermatozoa is a matter that remains tobe elucidated Breitbart et al [18] reported that forma-tion of F-actin mostly depends on PKA protein tyrosinephosphorylation and phospholipase D activation duringcapacitation Ca2+ is one of the principle regulators of
6 BioMed Research International
Acrosome reaction
Proteinphosphorylation
Flagellar beatinghyperactivation
CatS
per
sAC
ATP
cAMP
CNG
Calm
MPL
PLD
PLD
Acrosomalenzymes
Acrosome
IP3R
ZRK
G-proteins
Flagellarbeating
PLC
Aniontransport
sAC
mAC
T-type calcium channel
PKC
PTK PKG
mGC
GTPcGMP
sGC
CO NO
NO
CO
STK
PDE3
Nucleus
IP3
Fallopian tube (follicular fluid)
Sperm headPrinciple piece
Transporter
[pH]i
ZP3Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+ Ca2+
Ca2+G-
proteins
H+
H+
Ca2+HCO minus3
ZP3
ATP
cAMPPKA
Figure 2 Schematic diagram showing the mechanism of Ca2+ regulated hyperactivation capacitation and the acrosome reaction ofspermatozoa which are three principal events of fertilization Ca2+ together with ZP3 (zona pellucida glycoprotein-3) exhibits the mostimportant role in sperm binding and acrosomal reaction Ca2+ triggers the zona pellucida (ZP) receptors of cell membrane that activateG-proteins in the sperm head Activated G-proteins stimulate the H+ transporter to increase intracellular pH ultimately inducing theacrosomal reaction and hyperactivation by catalyzing the acrosomal enzymes [91] Cyclic adenosine monophosphate (cAMP) and cyclicguanosine monophosphate (cGMP) are produced from adenosine triphosphate (ATP) owing to enzymatic catalysis by soluble adenylatecyclase (sAC) and guanylate cyclase (sGC) respectively in mature spermatozoa The bicarbonate ions activate the sAC however follicularfluid also stimulates the sAC through release of Ca2+ ions via the CatSper channel (principal piece) However G-protein mediated signaltransduction activates sAC and phospholipase-C (PLC) that ultimately causes tyrosine phosphorylation [51 92] which is responsible forevents such as capacitation and the acrosomal reaction Likewise extracellular signals such as nitric oxide (NO) and carbon monoxide (CO)stimulate membrane-bound GC (mGC) and sGC respectively to synthesize cGMP Increases in cGMP level evoke a concomitant increasein cAMP by inhibiting its PDE3 However the increased Ca2+ level can also directly catalyze cAMP [93 94] Activated sAC sGC and PLCstimulate the generation of the second messengersrsquo inositol trisphosphate (IP3) like cAMP cGMP The IP3 binds to the IP3 receptor (IP3R)to increase [Ca2+]i via the release of the [Ca2+]i storage ions Concurrently the second messengers activate protein kinases (PKA PKCand PKG) in turn gating ions through the T-type calcium channels cyclic-nucleotide gated ion channel (CNG) and so on that togetherwith the activation of protein tyrosine kinases (PTK) and serinethreonine protein kinase (STK) cause increased protein phosphorylation[93 94] Additionally the CatSper Ca2+ activates calmodulin (Calm) phospholipase-A (PLA) and phospholipase-D (PLD) with increasedgeneration of other second messengers during the acrosome reaction Ca2+ influx together with increased protein phosphorylation bringsabout the capacitation response that is responsible for the waveform asymmetry of motility termed hyperactivation during fertilization Bothhyperactivation and the acrosomal reaction boost flagellar beating ultimately resulting in the penetration of the outer egg coat and subsequentfertilization of the mature ovum [91ndash95]
capacitation and it is therefore tempting to hypothesizethat organizational modification of F-actin in spermatozoatogether with interacting with other sperm proteins haspotential influence on Ca2+ influx A similar finding hasbeen established more precisely by another study [78] whereboar sperm capacitation was studied by combined applica-tion of computational and experimental approaches Theseauthors reported that the boar spermatozoa capacitationnetwork contains several connecting cascades whereas onlythree nodes bound to all the subcellular compartments are
involved in spermatozoa postejaculatory signaling such as[Ca2+]i ATP and actin polymerization Removal of the actinpolymerization node from this aforesaid network causesdisorganization of the network topography and affects capac-itation and this has been confirmed by zona pellucida-induced capacitation and the acrosomal reaction in an in vitrodemonstration [78]
In another study Patrat et al [79] showed that pro-gesterone (P
4) that is secreted by cumulus cells directly
acts on the sperm plasma membrane and triggers the
BioMed Research International 7
AVP
AVPR2 CNR1CATSPE
RB
Motilityhyperactivation CATSPER1
CATSPER2
CATSPERG
Calmoduli-n MARCKS
BASP1
MAPK1
PRKCSH
CALRITPR1
ADCY
SRCDLD
HSPA2
AKAP3
RON1
CABYR
SPINLW1
SEMG1
RAPGEF4
TNF
PLCZ1
Ca +2
PLC
PKC
PIP2NSF
PTPN1
SNAPreceptor
SYT6
RAB3A
RAPGEF3 GEF
SLC8A1
NPR1
NPPA
Chemotaxis
Progest-erone
Proteintyrosinekinase
Protein
Cell processFunctional class
Small molecule
RegulationMol synthesisMol transportBindingProt modifcation
Figure 3 Schematic representation of interactions among sim35 proteins related to Ca2+ regulated spermatozoa hyperactivation andchemotaxisThefigurewas produced by use of Pathway Studio (90) following theMedScanReader (50) protein search fromPubMeddatabase[12]
intracellular signals and enzymatic pathways involved inthe acrosome reaction P
4regulates the acrosome reaction
and is mediated by a compulsory Ca2+ increase This studyfound that P
4induced the activation of GiGo protein-
coupled and protein tyrosine kinase receptors and it affectedcapacitation and the acrosome reaction In contrast Ca2+ reg-ulated exocytosis of spermatozoa requires active acrosomalproteins such as N-ethylmaleimide-sensitive factor (NSF)[66] Additionally the same research team showed that the
ras-related protein Rab-3A (RAB3A) is also necessary forCa2+-dependent exocytosis Interestingly Rab3A activationof acrosomal exocytosis requires active NSF Thereforeprotein-protein interaction might also play a potential rolein regulating Ca2+ influx All of these observations seemto be consistent with the idea that Ca2+ functions areregulated by sperm proteins during fertilization Howeverthe key question is how do these proteins modify Ca2+ influxin spermatozoa
8 BioMed Research International
SEMG1AVP
PGR
CATSPER1
CATSPER2
ZP3
CDC42
DNM1
CHRNA7
PLCB1
PLCD4
GNA13
F-actin
Spinkl
GSN
IP3
PIP2
Protein tyrosinekinase
MAPK1
cAMPSperm
capacitation
PKAPRKAR2A
ADCY10
PLC
PKC
Actinflament
PLCZ1
L-tyrosine
CAMP
AKAP3ADCY
RON1LY6G6C
SLC8A1
OR10J5
CAMK2B
SRC
PLCG1
CS
ABCC1
NPPA
ABCB1
CAPN1
VDAC
Phospholipa-
IntracellularpH
Acrosome reaction
se A2
Ca2+
Fertilization
Protein
Cell processFunctional class Small molecule
Regulation
Complex
Axone-mal
dynein
Figure 4 Schematic representation of interactions among sim40 proteins related to Ca2+ regulated spermatozoa capacitation the acrosomereaction and fertilization The figure was produced by use of Pathway Studio (90) following the MedScan Reader (50) protein search fromPubMed database [12]
Recently in our laboratory we used mice spermatozoato evaluate the interrelationship of proteins related to Ca2+influx includingUQCRC2 [39] arginine vasopressin [5] andVDACs [4] and evaluate their effects on capacitation andthe acrosome reaction It is likely that a sustained phase ofCa2+ is required for fertilization andmight be regulated by thecomplex interaction of numerous sperm proteins Thereforestudies to identify proteins that might have the ability toinduce such a change are worth undertaking Application
of Pathway Studio helped us represent over 40 proteinsthat are potentially implicated in Ca2+ mediated regulationof capacitation the acrosome reaction and male fertility(Figure 4)
33 Ca2+ Influx and Postfertilization Egg Activation in Con-text of Sperm Proteome Ca2+ influx in spermatozoa is notonly important for sperm maturation but it is also equally
BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

BioMed Research International 5
model of Ca2+-induced hyperactivation is represented inFigure 2
It has recently been found by our laboratory thattreatment of mouse spermatozoa with nutlin-3a a smallmolecule antagonist of the mouse double minute 2 repressorpotentially downregulates the functions of the ubiquinol-cytochrome-c reductase complex component UQCRC2 andcorrelated with significantly reduced [Ca2+]i and spermhyperactivation This study provided insight that the Ca2+influx in spermatozoa is partially regulated by UQCRC2protein Kwon et al [4] reported that blocking VDACwith 441015840-diisothiocyanostilbene-221015840-disulfonic acid (DIDS)significantly decreased sperm hyperactivation A signifi-cant decrease in [Ca2+]i was observed in (minus) DIDS con-ditions while [pH]i significantly increased in (minus) DIDSregardless of Ca2+ Simultaneously a significantly elevated[pH]i was observed in (+) Ca2+ This study providesstrong evidence that the modulation of Ca2+ influx byVDACs is pH-dependent which is consistent with the resultof a previous study by Ho and Suarez [56] Moreoveranother study proposed that deamino [Cys 1 d-ArgS] vaso-pressin (dDAVP) an AVPR2 agonist significantly decreasedsperm motility and intracellular pH but interestingly itincreased [Ca2+]i by regulating the function of argininevasopressin in mice spermatozoa However it remains tobe clarified as to why spermatozoa motility is decreased evenin increased [Ca2+]i conditions
On the basis of the findings of the aforementionedstudies it is tempting to hypothesize that spermatozoahyperactivation is mostly controlled by Ca2+ influx How-ever potential interactions exist between protein func-tions Therefore Ca2+ influx protein interaction andhyperactivation might give numerous different annotationsof upcoming research in this field We have illustrateda schematic representation of different signaling path-ways involving sperm proteins by using Pathway StudioThese proteins exhibit significant modifications to inducesperm hyperactivation and chemotaxis in spermatozoa byregulating Ca2+ influx (Figure 3)
32 Ca2+ Influx versus Capacitation the Acrosomal ReactionFertilization and Sperm Proteome Mammalian fertilizationis a species-specific episode that is accomplished by a complexset of molecular events To fertilize an oocyte multipleextreme changes occur in spermatozoa that begin from itsformation in the testes of the male reproductive tract to itspenetration and fusionwith an egg in the female reproductivetract Although spermatozoa are motile as well asmorphologically normal after ejaculation they are unable tofertilize an oocyte [59]They gain the fertilization ability onlyafter educating in the female reproductive tract [40] and themodifications that spermatozoa experience during this timeare collectively known as ldquocapacitationrdquo Only capacitatedspermatozoa can undergo the acrosome reaction throughbinding to the egg zona pellucida and they finally becomecapable of penetrating and fertilizing the egg [4 18 39]
The term ldquocapacitationrdquo was proposed by Austin in 1952[1] although this concept was initially described by bothChang and Austin in 1951 [2 41] In fact in vivo capacitationtakes place in the female reproductive tract however it is alsopossible to capacitate spermatozoa in vitro by using particularmedia containing appropriate electrolytes and pH [2] Inan elegant review Visconti summarized that the early stageof capacitation mainly comprises the bicarbonate-mediatedactivation of sperm motility whereas the late stages includeintracellular alkalinization increase in protein tyrosine phos-phorylation and preparation for the acrosomal reaction [63]These temporal differences in capacitation and the acrosomereaction require numerous mechanisms and Ca2+ influxplays a significant role in the process [63 64] Fraser [65]reported that capacitation is a comparatively slow event thatrequires several hours to complete and is mainly regulated bya modest rise in [Ca2+]i whereas the acrosome reaction is anexocytosis process that occurs very rapidly (within a minute)and is triggered by a large influx of [Ca2+]i [65 66]
Although the biochemical phenomenon of Ca2+regulated capacitation and the acrosome reaction havebeen known for the last two decades the molecular basisof this process is still poorly understood For capacitationthe cholesterol influx initially stimulates the elevationof [Ca2+]i and bicarbonate into the spermatozoa andfinally activates PKA and tyrosine phosphorylationrespectively via the production of the cAMP [66ndash68]In addition binding to the zona pellucida causes additionalactivation of cAMPPKA and protein kinase C (PKC)[68ndash70] Spermatozoa need [Ca2+]i influx to proceed furtherand they are believed to be activated by PKC through theopening of the calcium channels Interestingly PKA togetherwith a secondary messenger inositol trisphosphate activatescalcium channels localized in the outer acrosomalmembraneand increases the calcium concentration in the cytosolFurther increase of cytosolic Ca2+ influx occurs through astore-operated calciumentrymechanism in the plasmamem-brane resulting in further depletion of Ca2+ in the acrosome[68 69]
In support of the aforesaid studies several recent stud-ies on the same topic have also hypothesized that afterthe morphological maturation of spermatozoa for sperm-oocyte fusion [Ca2+]i decreases because acrosome-reactedspermatozoa release a substantial amount of Ca2+ from theirinner cell layers [71 72] Ca2+-mediated capacitation and theacrosome reaction have been illustrated in Figure 2 for betterunderstandingHowever for amore in-depth understandingwe recommend reading some excellent reviews on this topic[63 67 73ndash77]
A review of the literature showed that several spermproteins potentially regulate the Ca2+-dependent capacita-tion and the acrosome reaction in mammalian spermato-zoa [4 5 39] However how these proteins regulate theCa2+ influx in spermatozoa is a matter that remains tobe elucidated Breitbart et al [18] reported that forma-tion of F-actin mostly depends on PKA protein tyrosinephosphorylation and phospholipase D activation duringcapacitation Ca2+ is one of the principle regulators of
6 BioMed Research International
Acrosome reaction
Proteinphosphorylation
Flagellar beatinghyperactivation
CatS
per
sAC
ATP
cAMP
CNG
Calm
MPL
PLD
PLD
Acrosomalenzymes
Acrosome
IP3R
ZRK
G-proteins
Flagellarbeating
PLC
Aniontransport
sAC
mAC
T-type calcium channel
PKC
PTK PKG
mGC
GTPcGMP
sGC
CO NO
NO
CO
STK
PDE3
Nucleus
IP3
Fallopian tube (follicular fluid)
Sperm headPrinciple piece
Transporter
[pH]i
ZP3Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+ Ca2+
Ca2+G-
proteins
H+
H+
Ca2+HCO minus3
ZP3
ATP
cAMPPKA
Figure 2 Schematic diagram showing the mechanism of Ca2+ regulated hyperactivation capacitation and the acrosome reaction ofspermatozoa which are three principal events of fertilization Ca2+ together with ZP3 (zona pellucida glycoprotein-3) exhibits the mostimportant role in sperm binding and acrosomal reaction Ca2+ triggers the zona pellucida (ZP) receptors of cell membrane that activateG-proteins in the sperm head Activated G-proteins stimulate the H+ transporter to increase intracellular pH ultimately inducing theacrosomal reaction and hyperactivation by catalyzing the acrosomal enzymes [91] Cyclic adenosine monophosphate (cAMP) and cyclicguanosine monophosphate (cGMP) are produced from adenosine triphosphate (ATP) owing to enzymatic catalysis by soluble adenylatecyclase (sAC) and guanylate cyclase (sGC) respectively in mature spermatozoa The bicarbonate ions activate the sAC however follicularfluid also stimulates the sAC through release of Ca2+ ions via the CatSper channel (principal piece) However G-protein mediated signaltransduction activates sAC and phospholipase-C (PLC) that ultimately causes tyrosine phosphorylation [51 92] which is responsible forevents such as capacitation and the acrosomal reaction Likewise extracellular signals such as nitric oxide (NO) and carbon monoxide (CO)stimulate membrane-bound GC (mGC) and sGC respectively to synthesize cGMP Increases in cGMP level evoke a concomitant increasein cAMP by inhibiting its PDE3 However the increased Ca2+ level can also directly catalyze cAMP [93 94] Activated sAC sGC and PLCstimulate the generation of the second messengersrsquo inositol trisphosphate (IP3) like cAMP cGMP The IP3 binds to the IP3 receptor (IP3R)to increase [Ca2+]i via the release of the [Ca2+]i storage ions Concurrently the second messengers activate protein kinases (PKA PKCand PKG) in turn gating ions through the T-type calcium channels cyclic-nucleotide gated ion channel (CNG) and so on that togetherwith the activation of protein tyrosine kinases (PTK) and serinethreonine protein kinase (STK) cause increased protein phosphorylation[93 94] Additionally the CatSper Ca2+ activates calmodulin (Calm) phospholipase-A (PLA) and phospholipase-D (PLD) with increasedgeneration of other second messengers during the acrosome reaction Ca2+ influx together with increased protein phosphorylation bringsabout the capacitation response that is responsible for the waveform asymmetry of motility termed hyperactivation during fertilization Bothhyperactivation and the acrosomal reaction boost flagellar beating ultimately resulting in the penetration of the outer egg coat and subsequentfertilization of the mature ovum [91ndash95]
capacitation and it is therefore tempting to hypothesizethat organizational modification of F-actin in spermatozoatogether with interacting with other sperm proteins haspotential influence on Ca2+ influx A similar finding hasbeen established more precisely by another study [78] whereboar sperm capacitation was studied by combined applica-tion of computational and experimental approaches Theseauthors reported that the boar spermatozoa capacitationnetwork contains several connecting cascades whereas onlythree nodes bound to all the subcellular compartments are
involved in spermatozoa postejaculatory signaling such as[Ca2+]i ATP and actin polymerization Removal of the actinpolymerization node from this aforesaid network causesdisorganization of the network topography and affects capac-itation and this has been confirmed by zona pellucida-induced capacitation and the acrosomal reaction in an in vitrodemonstration [78]
In another study Patrat et al [79] showed that pro-gesterone (P
4) that is secreted by cumulus cells directly
acts on the sperm plasma membrane and triggers the
BioMed Research International 7
AVP
AVPR2 CNR1CATSPE
RB
Motilityhyperactivation CATSPER1
CATSPER2
CATSPERG
Calmoduli-n MARCKS
BASP1
MAPK1
PRKCSH
CALRITPR1
ADCY
SRCDLD
HSPA2
AKAP3
RON1
CABYR
SPINLW1
SEMG1
RAPGEF4
TNF
PLCZ1
Ca +2
PLC
PKC
PIP2NSF
PTPN1
SNAPreceptor
SYT6
RAB3A
RAPGEF3 GEF
SLC8A1
NPR1
NPPA
Chemotaxis
Progest-erone
Proteintyrosinekinase
Protein
Cell processFunctional class
Small molecule
RegulationMol synthesisMol transportBindingProt modifcation
Figure 3 Schematic representation of interactions among sim35 proteins related to Ca2+ regulated spermatozoa hyperactivation andchemotaxisThefigurewas produced by use of Pathway Studio (90) following theMedScanReader (50) protein search fromPubMeddatabase[12]
intracellular signals and enzymatic pathways involved inthe acrosome reaction P
4regulates the acrosome reaction
and is mediated by a compulsory Ca2+ increase This studyfound that P
4induced the activation of GiGo protein-
coupled and protein tyrosine kinase receptors and it affectedcapacitation and the acrosome reaction In contrast Ca2+ reg-ulated exocytosis of spermatozoa requires active acrosomalproteins such as N-ethylmaleimide-sensitive factor (NSF)[66] Additionally the same research team showed that the
ras-related protein Rab-3A (RAB3A) is also necessary forCa2+-dependent exocytosis Interestingly Rab3A activationof acrosomal exocytosis requires active NSF Thereforeprotein-protein interaction might also play a potential rolein regulating Ca2+ influx All of these observations seemto be consistent with the idea that Ca2+ functions areregulated by sperm proteins during fertilization Howeverthe key question is how do these proteins modify Ca2+ influxin spermatozoa
8 BioMed Research International
SEMG1AVP
PGR
CATSPER1
CATSPER2
ZP3
CDC42
DNM1
CHRNA7
PLCB1
PLCD4
GNA13
F-actin
Spinkl
GSN
IP3
PIP2
Protein tyrosinekinase
MAPK1
cAMPSperm
capacitation
PKAPRKAR2A
ADCY10
PLC
PKC
Actinflament
PLCZ1
L-tyrosine
CAMP
AKAP3ADCY
RON1LY6G6C
SLC8A1
OR10J5
CAMK2B
SRC
PLCG1
CS
ABCC1
NPPA
ABCB1
CAPN1
VDAC
Phospholipa-
IntracellularpH
Acrosome reaction
se A2
Ca2+
Fertilization
Protein
Cell processFunctional class Small molecule
Regulation
Complex
Axone-mal
dynein
Figure 4 Schematic representation of interactions among sim40 proteins related to Ca2+ regulated spermatozoa capacitation the acrosomereaction and fertilization The figure was produced by use of Pathway Studio (90) following the MedScan Reader (50) protein search fromPubMed database [12]
Recently in our laboratory we used mice spermatozoato evaluate the interrelationship of proteins related to Ca2+influx includingUQCRC2 [39] arginine vasopressin [5] andVDACs [4] and evaluate their effects on capacitation andthe acrosome reaction It is likely that a sustained phase ofCa2+ is required for fertilization andmight be regulated by thecomplex interaction of numerous sperm proteins Thereforestudies to identify proteins that might have the ability toinduce such a change are worth undertaking Application
of Pathway Studio helped us represent over 40 proteinsthat are potentially implicated in Ca2+ mediated regulationof capacitation the acrosome reaction and male fertility(Figure 4)
33 Ca2+ Influx and Postfertilization Egg Activation in Con-text of Sperm Proteome Ca2+ influx in spermatozoa is notonly important for sperm maturation but it is also equally
BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004
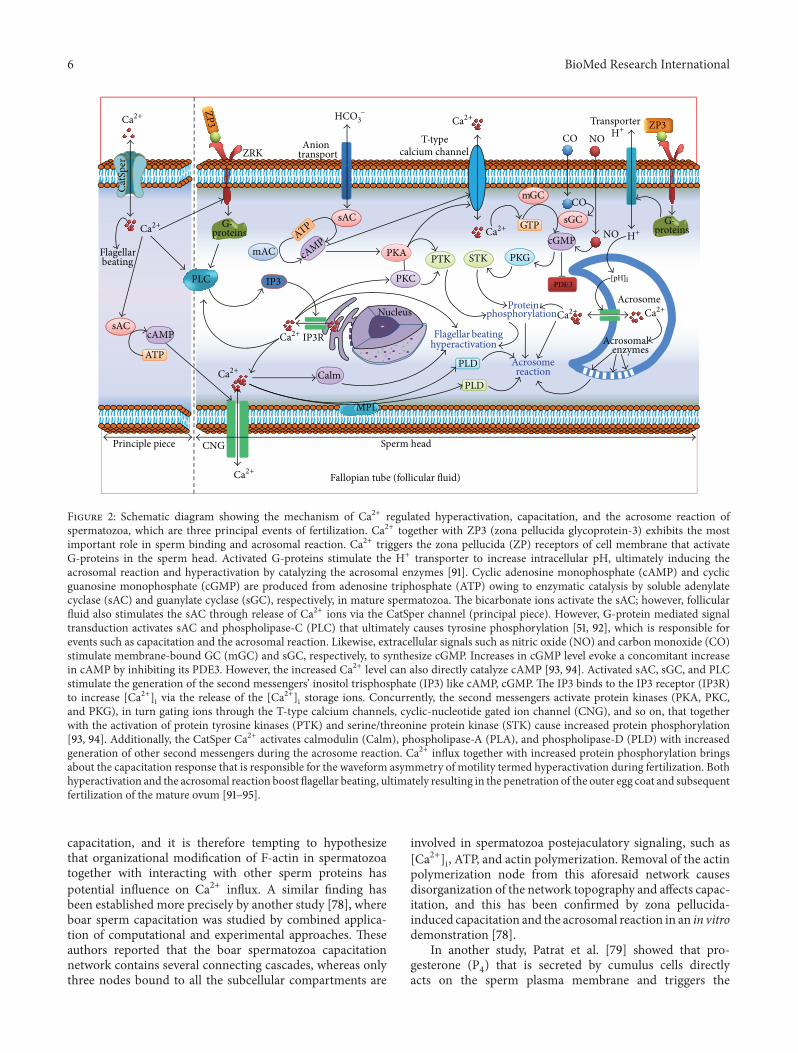
6 BioMed Research International
Acrosome reaction
Proteinphosphorylation
Flagellar beatinghyperactivation
CatS
per
sAC
ATP
cAMP
CNG
Calm
MPL
PLD
PLD
Acrosomalenzymes
Acrosome
IP3R
ZRK
G-proteins
Flagellarbeating
PLC
Aniontransport
sAC
mAC
T-type calcium channel
PKC
PTK PKG
mGC
GTPcGMP
sGC
CO NO
NO
CO
STK
PDE3
Nucleus
IP3
Fallopian tube (follicular fluid)
Sperm headPrinciple piece
Transporter
[pH]i
ZP3Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+ Ca2+
Ca2+G-
proteins
H+
H+
Ca2+HCO minus3
ZP3
ATP
cAMPPKA
Figure 2 Schematic diagram showing the mechanism of Ca2+ regulated hyperactivation capacitation and the acrosome reaction ofspermatozoa which are three principal events of fertilization Ca2+ together with ZP3 (zona pellucida glycoprotein-3) exhibits the mostimportant role in sperm binding and acrosomal reaction Ca2+ triggers the zona pellucida (ZP) receptors of cell membrane that activateG-proteins in the sperm head Activated G-proteins stimulate the H+ transporter to increase intracellular pH ultimately inducing theacrosomal reaction and hyperactivation by catalyzing the acrosomal enzymes [91] Cyclic adenosine monophosphate (cAMP) and cyclicguanosine monophosphate (cGMP) are produced from adenosine triphosphate (ATP) owing to enzymatic catalysis by soluble adenylatecyclase (sAC) and guanylate cyclase (sGC) respectively in mature spermatozoa The bicarbonate ions activate the sAC however follicularfluid also stimulates the sAC through release of Ca2+ ions via the CatSper channel (principal piece) However G-protein mediated signaltransduction activates sAC and phospholipase-C (PLC) that ultimately causes tyrosine phosphorylation [51 92] which is responsible forevents such as capacitation and the acrosomal reaction Likewise extracellular signals such as nitric oxide (NO) and carbon monoxide (CO)stimulate membrane-bound GC (mGC) and sGC respectively to synthesize cGMP Increases in cGMP level evoke a concomitant increasein cAMP by inhibiting its PDE3 However the increased Ca2+ level can also directly catalyze cAMP [93 94] Activated sAC sGC and PLCstimulate the generation of the second messengersrsquo inositol trisphosphate (IP3) like cAMP cGMP The IP3 binds to the IP3 receptor (IP3R)to increase [Ca2+]i via the release of the [Ca2+]i storage ions Concurrently the second messengers activate protein kinases (PKA PKCand PKG) in turn gating ions through the T-type calcium channels cyclic-nucleotide gated ion channel (CNG) and so on that togetherwith the activation of protein tyrosine kinases (PTK) and serinethreonine protein kinase (STK) cause increased protein phosphorylation[93 94] Additionally the CatSper Ca2+ activates calmodulin (Calm) phospholipase-A (PLA) and phospholipase-D (PLD) with increasedgeneration of other second messengers during the acrosome reaction Ca2+ influx together with increased protein phosphorylation bringsabout the capacitation response that is responsible for the waveform asymmetry of motility termed hyperactivation during fertilization Bothhyperactivation and the acrosomal reaction boost flagellar beating ultimately resulting in the penetration of the outer egg coat and subsequentfertilization of the mature ovum [91ndash95]
capacitation and it is therefore tempting to hypothesizethat organizational modification of F-actin in spermatozoatogether with interacting with other sperm proteins haspotential influence on Ca2+ influx A similar finding hasbeen established more precisely by another study [78] whereboar sperm capacitation was studied by combined applica-tion of computational and experimental approaches Theseauthors reported that the boar spermatozoa capacitationnetwork contains several connecting cascades whereas onlythree nodes bound to all the subcellular compartments are
involved in spermatozoa postejaculatory signaling such as[Ca2+]i ATP and actin polymerization Removal of the actinpolymerization node from this aforesaid network causesdisorganization of the network topography and affects capac-itation and this has been confirmed by zona pellucida-induced capacitation and the acrosomal reaction in an in vitrodemonstration [78]
In another study Patrat et al [79] showed that pro-gesterone (P
4) that is secreted by cumulus cells directly
acts on the sperm plasma membrane and triggers the
BioMed Research International 7
AVP
AVPR2 CNR1CATSPE
RB
Motilityhyperactivation CATSPER1
CATSPER2
CATSPERG
Calmoduli-n MARCKS
BASP1
MAPK1
PRKCSH
CALRITPR1
ADCY
SRCDLD
HSPA2
AKAP3
RON1
CABYR
SPINLW1
SEMG1
RAPGEF4
TNF
PLCZ1
Ca +2
PLC
PKC
PIP2NSF
PTPN1
SNAPreceptor
SYT6
RAB3A
RAPGEF3 GEF
SLC8A1
NPR1
NPPA
Chemotaxis
Progest-erone
Proteintyrosinekinase
Protein
Cell processFunctional class
Small molecule
RegulationMol synthesisMol transportBindingProt modifcation
Figure 3 Schematic representation of interactions among sim35 proteins related to Ca2+ regulated spermatozoa hyperactivation andchemotaxisThefigurewas produced by use of Pathway Studio (90) following theMedScanReader (50) protein search fromPubMeddatabase[12]
intracellular signals and enzymatic pathways involved inthe acrosome reaction P
4regulates the acrosome reaction
and is mediated by a compulsory Ca2+ increase This studyfound that P
4induced the activation of GiGo protein-
coupled and protein tyrosine kinase receptors and it affectedcapacitation and the acrosome reaction In contrast Ca2+ reg-ulated exocytosis of spermatozoa requires active acrosomalproteins such as N-ethylmaleimide-sensitive factor (NSF)[66] Additionally the same research team showed that the
ras-related protein Rab-3A (RAB3A) is also necessary forCa2+-dependent exocytosis Interestingly Rab3A activationof acrosomal exocytosis requires active NSF Thereforeprotein-protein interaction might also play a potential rolein regulating Ca2+ influx All of these observations seemto be consistent with the idea that Ca2+ functions areregulated by sperm proteins during fertilization Howeverthe key question is how do these proteins modify Ca2+ influxin spermatozoa
8 BioMed Research International
SEMG1AVP
PGR
CATSPER1
CATSPER2
ZP3
CDC42
DNM1
CHRNA7
PLCB1
PLCD4
GNA13
F-actin
Spinkl
GSN
IP3
PIP2
Protein tyrosinekinase
MAPK1
cAMPSperm
capacitation
PKAPRKAR2A
ADCY10
PLC
PKC
Actinflament
PLCZ1
L-tyrosine
CAMP
AKAP3ADCY
RON1LY6G6C
SLC8A1
OR10J5
CAMK2B
SRC
PLCG1
CS
ABCC1
NPPA
ABCB1
CAPN1
VDAC
Phospholipa-
IntracellularpH
Acrosome reaction
se A2
Ca2+
Fertilization
Protein
Cell processFunctional class Small molecule
Regulation
Complex
Axone-mal
dynein
Figure 4 Schematic representation of interactions among sim40 proteins related to Ca2+ regulated spermatozoa capacitation the acrosomereaction and fertilization The figure was produced by use of Pathway Studio (90) following the MedScan Reader (50) protein search fromPubMed database [12]
Recently in our laboratory we used mice spermatozoato evaluate the interrelationship of proteins related to Ca2+influx includingUQCRC2 [39] arginine vasopressin [5] andVDACs [4] and evaluate their effects on capacitation andthe acrosome reaction It is likely that a sustained phase ofCa2+ is required for fertilization andmight be regulated by thecomplex interaction of numerous sperm proteins Thereforestudies to identify proteins that might have the ability toinduce such a change are worth undertaking Application
of Pathway Studio helped us represent over 40 proteinsthat are potentially implicated in Ca2+ mediated regulationof capacitation the acrosome reaction and male fertility(Figure 4)
33 Ca2+ Influx and Postfertilization Egg Activation in Con-text of Sperm Proteome Ca2+ influx in spermatozoa is notonly important for sperm maturation but it is also equally
BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

BioMed Research International 7
AVP
AVPR2 CNR1CATSPE
RB
Motilityhyperactivation CATSPER1
CATSPER2
CATSPERG
Calmoduli-n MARCKS
BASP1
MAPK1
PRKCSH
CALRITPR1
ADCY
SRCDLD
HSPA2
AKAP3
RON1
CABYR
SPINLW1
SEMG1
RAPGEF4
TNF
PLCZ1
Ca +2
PLC
PKC
PIP2NSF
PTPN1
SNAPreceptor
SYT6
RAB3A
RAPGEF3 GEF
SLC8A1
NPR1
NPPA
Chemotaxis
Progest-erone
Proteintyrosinekinase
Protein
Cell processFunctional class
Small molecule
RegulationMol synthesisMol transportBindingProt modifcation
Figure 3 Schematic representation of interactions among sim35 proteins related to Ca2+ regulated spermatozoa hyperactivation andchemotaxisThefigurewas produced by use of Pathway Studio (90) following theMedScanReader (50) protein search fromPubMeddatabase[12]
intracellular signals and enzymatic pathways involved inthe acrosome reaction P
4regulates the acrosome reaction
and is mediated by a compulsory Ca2+ increase This studyfound that P
4induced the activation of GiGo protein-
coupled and protein tyrosine kinase receptors and it affectedcapacitation and the acrosome reaction In contrast Ca2+ reg-ulated exocytosis of spermatozoa requires active acrosomalproteins such as N-ethylmaleimide-sensitive factor (NSF)[66] Additionally the same research team showed that the
ras-related protein Rab-3A (RAB3A) is also necessary forCa2+-dependent exocytosis Interestingly Rab3A activationof acrosomal exocytosis requires active NSF Thereforeprotein-protein interaction might also play a potential rolein regulating Ca2+ influx All of these observations seemto be consistent with the idea that Ca2+ functions areregulated by sperm proteins during fertilization Howeverthe key question is how do these proteins modify Ca2+ influxin spermatozoa
8 BioMed Research International
SEMG1AVP
PGR
CATSPER1
CATSPER2
ZP3
CDC42
DNM1
CHRNA7
PLCB1
PLCD4
GNA13
F-actin
Spinkl
GSN
IP3
PIP2
Protein tyrosinekinase
MAPK1
cAMPSperm
capacitation
PKAPRKAR2A
ADCY10
PLC
PKC
Actinflament
PLCZ1
L-tyrosine
CAMP
AKAP3ADCY
RON1LY6G6C
SLC8A1
OR10J5
CAMK2B
SRC
PLCG1
CS
ABCC1
NPPA
ABCB1
CAPN1
VDAC
Phospholipa-
IntracellularpH
Acrosome reaction
se A2
Ca2+
Fertilization
Protein
Cell processFunctional class Small molecule
Regulation
Complex
Axone-mal
dynein
Figure 4 Schematic representation of interactions among sim40 proteins related to Ca2+ regulated spermatozoa capacitation the acrosomereaction and fertilization The figure was produced by use of Pathway Studio (90) following the MedScan Reader (50) protein search fromPubMed database [12]
Recently in our laboratory we used mice spermatozoato evaluate the interrelationship of proteins related to Ca2+influx includingUQCRC2 [39] arginine vasopressin [5] andVDACs [4] and evaluate their effects on capacitation andthe acrosome reaction It is likely that a sustained phase ofCa2+ is required for fertilization andmight be regulated by thecomplex interaction of numerous sperm proteins Thereforestudies to identify proteins that might have the ability toinduce such a change are worth undertaking Application
of Pathway Studio helped us represent over 40 proteinsthat are potentially implicated in Ca2+ mediated regulationof capacitation the acrosome reaction and male fertility(Figure 4)
33 Ca2+ Influx and Postfertilization Egg Activation in Con-text of Sperm Proteome Ca2+ influx in spermatozoa is notonly important for sperm maturation but it is also equally
BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004
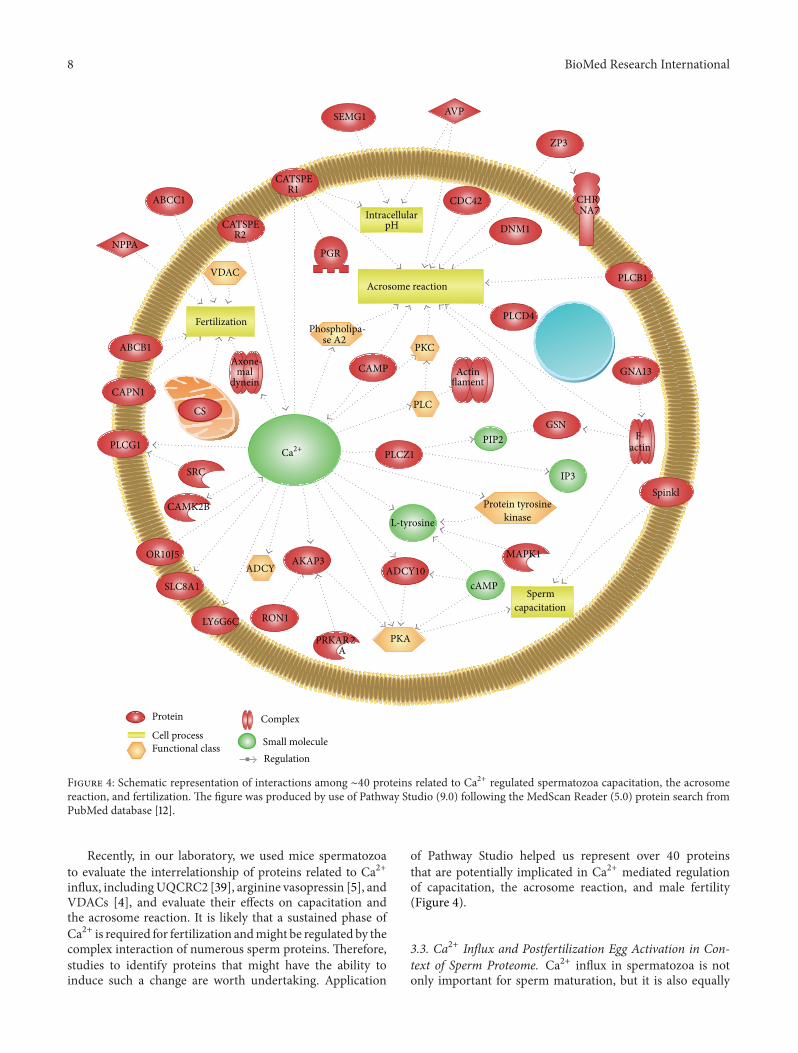
8 BioMed Research International
SEMG1AVP
PGR
CATSPER1
CATSPER2
ZP3
CDC42
DNM1
CHRNA7
PLCB1
PLCD4
GNA13
F-actin
Spinkl
GSN
IP3
PIP2
Protein tyrosinekinase
MAPK1
cAMPSperm
capacitation
PKAPRKAR2A
ADCY10
PLC
PKC
Actinflament
PLCZ1
L-tyrosine
CAMP
AKAP3ADCY
RON1LY6G6C
SLC8A1
OR10J5
CAMK2B
SRC
PLCG1
CS
ABCC1
NPPA
ABCB1
CAPN1
VDAC
Phospholipa-
IntracellularpH
Acrosome reaction
se A2
Ca2+
Fertilization
Protein
Cell processFunctional class Small molecule
Regulation
Complex
Axone-mal
dynein
Figure 4 Schematic representation of interactions among sim40 proteins related to Ca2+ regulated spermatozoa capacitation the acrosomereaction and fertilization The figure was produced by use of Pathway Studio (90) following the MedScan Reader (50) protein search fromPubMed database [12]
Recently in our laboratory we used mice spermatozoato evaluate the interrelationship of proteins related to Ca2+influx includingUQCRC2 [39] arginine vasopressin [5] andVDACs [4] and evaluate their effects on capacitation andthe acrosome reaction It is likely that a sustained phase ofCa2+ is required for fertilization andmight be regulated by thecomplex interaction of numerous sperm proteins Thereforestudies to identify proteins that might have the ability toinduce such a change are worth undertaking Application
of Pathway Studio helped us represent over 40 proteinsthat are potentially implicated in Ca2+ mediated regulationof capacitation the acrosome reaction and male fertility(Figure 4)
33 Ca2+ Influx and Postfertilization Egg Activation in Con-text of Sperm Proteome Ca2+ influx in spermatozoa is notonly important for sperm maturation but it is also equally
BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

BioMed Research International 9
Ca2+
Ca2+PLC120577
InsP3
+
+
Endoplasmicreticulum
Perivitelline space
Zona pellucida
Oocyte
Sperm
Figure 5 Schematic representation showing theCa2+ influxmechanism inmammalian eggs stimulated bymature spermatozoa Spermatozoadonate the phospholipase C isoform zeta (PLC120577) protein within a fewminutes of sperm-egg fusion (represented by green color circle) Inositol145-trisphosphate (InsP
3) is produced due to the hydrolysis of PLC120577 which subsequently triggers the nsP
3receptor-mediated Ca2+ release
(indicated by red color circle) from the endoplasmic reticulum of the oocyte Simultaneously the increased cytoplasmic Ca2+ leads to furtherPLC120577 stimulation leading to the positive feedback loop of Ca2+ and InsP
3riseThe hypothesis has beenmodified from Swann [96] and Swann
et al [97]
important for activation and development of the oocyte Astudy of egg activation by Ca2+ was conducted by Steinhardtand colleagues in 1974 and showed remarkable findings[80] Steinhardt et al [80] reported that administration ofCa2+ ionophores induced the early events of hamster eggactivationThus far it has been shown that the eggs of almostall species are activated by an increase in Ca2+ oscillationby spermatozoa during fertilization [81 82] However howthe spermatozoa trigger the oocyte Ca2+ oscillation remainsto be elucidated Several hypotheses have been proposed todescribe these mechanisms [83ndash86]
It has been reported previously [83] that the spermatozoaintroduce Ca2+ influx into oocytes by a specific proteincalled oscillogen in hamsters Recent studies have shownthat phospholipase C zeta (PLC120577) a novel sperm-specificagent is responsible for induction of Ca2+ oscillation ineggs after sperm-egg membrane fusion [87ndash89] Accordingto this mechanism the sperm protein PLC120577 causes therelease of [Ca2+]i in eggs and is mediated via inositol 145-trisphosphate (InsP
3) receptors (hypothetical depiction in
Figure 5) Even when the InsP3or its derivatives are injected
into unfertilized mature eggs oscillation occurs due to theunique feedback properties of the InsP
3receptors in mouse
eggs [90] However it is still unknown whether there areany other factorsproteins available in spermatozoa that also
have similar effects We illustrated the relevant signaling andmetabolic pathways by using sperm proteins to facilitate theunderstanding of the mechanisms behind Ca2+ mediatedactivation of oocytes (Figure 6)
4 Future Prospects
The maturational events of mammalian spermatozoa arestrictly regulated through the well-coordinated Ca2+ influx Itis the central regulator of many key activities in spermatozoaall of which are necessary for fertilization However ourcurrent understanding at the molecular level concerningCa2+ signaling in the spermatozoa is insufficient Thereforea better understanding of such an event can provide a morecomplete comprehension of Ca2+ regulated sperm functionsand fertility optimization
A large number of Ca2+ permeable ion channel proteinshave been identified [10 43] that collectively regulate theCa2+ influx mechanism in spermatozoa Although the recentapplication of patch-clamp recordings of channel currentsignificantly improves our understanding of the functions ofthese channel proteins several basic aspects remain unsolvedsuch as identifying the functions of individual channels inspermatozoa and how these channels coordinate Ca2+ influx
10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

10 BioMed Research International
Disease
Chemical reaction
Direct regulationExpression
EGF
KITPLCZ1
KCNMA1PLCG1
PLCB1
PLCG1
MAPK1
PAEP
GiCR
PKA
Calmod-ulin
CATSPE
OR10J5
R1
ADCYPIP2
Glutamyl
Ovarianneoplasms
PDGF
PDGFPLCG2
VPS54
ZP3
CS
TPA
Phorbolesters
Microtu-bule
PKC
PLC
RYR
YES1
CCNB1
CDK1Guanine
nucleotide-s
SRC
receptor
Embryonaldevelopment
Egg activation
endopepti-dase
G-protein
H2O2
HCO minus3
L-tyrosin-
e Cationchannel
Calciumchannel
Ca2+
Na+H+
antiporter
Voltage-gated Ca2+
channel
Progester-one
Intracell-ular pH
Spermcapacitation
G-Actin
Phosphol-ipase D
Phospho-diesteras-
e
Protein
Cell processFunctional class
Small molecule
Regulation
Mol synthesisMol transport
BindingProt modifcation
Complex
Figure 6 Schematic representation of interactions among sim30 proteins related to Ca2+ regulated egg activation and embryonic developmentThe figure was produced by the use of Pathway Studio (90) following the MedScan Reader (50) protein search from PubMed database [12]
Therefore production of knockdown animals and using themas negative controls compared with their wild counterpartsmight provide more specific ideas about channel functionsCatSper is the one of the well-studied channel proteins [10]and the functions of different pore-forming CatSper channels(1ndash4) and auxiliary subunits (CatSper120573 andCatSper120574) remaina matter to be elucidated
A literature review found that the Ca2+ influxmechanismin spermatozoa is regulated by several physical stimuli
although the underlying mechanism is less clearly definedProtein-protein interactions also potentially regulate theCa2+ uptake mechanism in spermatozoa Although recentlyapplied proteomic approaches have identified several sperm-specific proteins their functions in Ca2+ regulation andinteraction with channel proteins are unclear Thereforefuture research should target this topic to provide a robustunderstanding of Ca2+ andmale fertility in both humans andother animal species
BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

BioMed Research International 11
Conflict of Interests
The authors have declared there is no conflict of intereststhat could be perceived as prejudicing the impartiality of thispaper
Acknowledgments
This study was financially supported by the CooperativeResearch Program for Agriculture Science and TechnologyDevelopment (Project no PJ008415) Rural DevelopmentAdministration Republic of Korea Md Saidur Rahmanwas supported through the ldquoChung-Ang University YoungScientist Scholarship (CAYSS)rdquo Chung-Ang UniversityRepublic of Korea
References
[1] C R Austin ldquoThe capacitation of the mammalian spermrdquoNature vol 170 no 4321 p 326 1952
[2] M C Chang ldquoFertilizing capacity of spermatozoa depositedinto the fallopian tubesrdquoNature vol 168 no 4277 pp 697ndash6981951
[3] N Mannowetz N M Naidoo S-A Choo J F Smith and PV Lishko ldquoSlo1 is the principal potassium channel of humanspermatozoardquo eLife vol 2 Article ID e01009 2013
[4] W-S Kwon Y-J Park A M El-Sayed and M-G PangldquoVoltage-dependent anion channels are a key factor of malefertilityrdquo Fertility and Sterility vol 99 no 2 pp 354ndash361 2013
[5] W-S Kwon Y-J Park Y-H Kim Y-A You I C Kim andM-G Pang ldquoVasopressin effectively suppressesmale fertilityrdquoPLoSONE vol 8 no 1 Article ID e54192 2013
[6] P V Lishko and Y Kirichok ldquoThe role of Hv1 and CatSperchannels in sperm activationrdquo The Journal of Physiology vol588 no 23 pp 4667ndash4672 2010
[7] S S Suarez ldquoControl of hyperactivation in spermrdquo HumanReproduction Update vol 14 no 6 pp 647ndash657 2008
[8] C Foresta andM Rossato ldquoCalcium influx pathways in humanspermatozoardquoMolecular Human Reproduction vol 3 no 1 pp1ndash4 1997
[9] Y Kirichok B Navarro and D E Clapham ldquoWhole-cellpatch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channelrdquo Nature vol 439 no 7077 pp 737ndash7402006
[10] D Ren and J Xia ldquoCalcium signaling throughCatSper channelsin mammalian fertilizationrdquo Physiology vol 25 no 3 pp 165ndash175 2010
[11] M S Rahman J-S Lee W-S Kwon and M-G Pang ldquoSpermproteomics road to male fertility and contraceptionrdquo Interna-tional Journal of Endocrinology vol 2013 Article ID 360986 11pages 2013
[12] Y-J Park W-S Kwon S-A Oh and M-G Pang ldquoFertility-related proteomic profiling bull spermatozoa separated bypercollrdquo Journal of Proteome Research vol 11 no 8 pp 4162ndash4168 2012
[13] M A Baker B Nixon N Naumovski and R J AitkenldquoProteomic insights into the maturation and capacitation ofmammalian spermatozoardquo Systems Biology in ReproductiveMedicine vol 58 no 4 pp 211ndash217 2012
[14] R Oliva S de Mateo and J M Estanyol ldquoSperm cell pro-teomicsrdquo Proteomics vol 9 no 4 pp 1004ndash1017 2009
[15] Y Gur and H Breitbart ldquoMammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomesrdquo Genes ampDevelopment vol 20 no 4 pp 411ndash416 2006
[16] V Torres-Flores Y L Hernandez-Rueda P D Neri-Vidaurri etal ldquoActivation of protein kinase A stimulates the progesterone-induced calcium influx in human sperm exposed to the phos-phodiesterase inhibitor papaverinerdquo Journal of Andrology vol29 no 5 pp 549ndash557 2008
[17] V Kumar V Kota and S Shivaji ldquoHamster sperm capacitationrole of pyruvate dehydrogenase A and dihydrolipoamide dehy-drogenaserdquo Biology of Reproduction vol 79 no 2 pp 190ndash1992008
[18] H Breitbart G Cohen and S Rubinstein ldquoRole of actincytoskeleton in mammalian sperm capacitation and the acro-some reactionrdquo Reproduction vol 129 no 3 pp 263ndash268 2005
[19] C L R Barratt and S J Publicover ldquoSperm are promiscuousand CatSper is to blamerdquoThe EMBO Journal vol 31 no 7 pp1624ndash1626 2012
[20] H Qi M M Moran B Navarro et al ldquoAll four CatSper ionchannel proteins are required for male fertility and sperm cellhyperactivated motilityrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 4 pp1219ndash1223 2007
[21] P V Lishko I L Botchkina and Y Kirichok ldquoProgesteroneactivates the principal Ca2+ channel of human spermrdquo Naturevol 471 no 7338 pp 387ndash391 2011
[22] P V Lishko I L Botchkina A Fedorenko and Y KirichokldquoAcid extrusion from human spermatozoa is mediated byflagellar voltage-gated proton channelrdquo Cell vol 140 no 3 pp327ndash337 2010
[23] B Navarro K Miki and D E Clapham ldquoATP-activated P2X2current in mouse spermatozoardquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 108 no34 pp 14342ndash14347 2011
[24] M Gees B Colsoul and B Nilius ldquoThe role of transientreceptor potential cation channels in Ca2+ signalingrdquo ColdSpring Harbor Perspectives in Biology vol 2 no 10 Article IDa003962 2010
[25] L E Castellano C L Trevino D Rodrıguez et al ldquoTransientreceptor potential (TRPC) channels in human sperm expres-sion cellular localization and involvement in the regulation offlagellarmotilityrdquo FEBSLetters vol 541 no 1ndash3 pp 69ndash74 2003
[26] M Biel and S Michalakis ldquoCyclic nucleotide-gated channelsrdquoHandbook of Experimental Pharmacology vol 191 pp 111ndash1362009
[27] B Wiesner J Weiner R Middendorff V Hagen U B Kauppand I Weyand ldquoCyclic nucleotide-gated channels on the flagel-lum control Ca2+ entry into spermrdquoThe Journal of Cell Biologyvol 142 no 2 pp 473ndash484 1998
[28] M Yoshida M Ishikawa H Izumi R de Santis and M Mori-sawa ldquoStore-operated calcium channel regulates the chemo-tactic behavior of ascidian spermrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 100 no1 pp 149ndash154 2003
[29] G Esposito B S Jaiswal F Xie et al ldquoMice deficient for solubleadenylyl cyclase are infertile because of a severe sperm-motilitydefectrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 9 pp 2993ndash2998 2004
12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

12 BioMed Research International
[30] K CHess BH Jones BMarquez et al ldquoThe ldquosolublerdquo adenylylcyclase in sperm mediates multiple signaling events requiredfor fertilizationrdquo Developmental Cell vol 9 no 2 pp 249ndash2592005
[31] K Fukami K Nakao T Inoue et al ldquoRequirement of phospho-lipase C1205754 for the zona pellucida-induced acrosome reactionrdquoScience vol 292 no 5518 pp 920ndash923 2001
[32] K Fukami M Yoshida T Inoue et al ldquoPhospholipase C1205754 isrequired for Ca2+ mobilization essential for acrosome reactionin spermrdquoThe Journal of Cell Biology vol 161 no 1 pp 79ndash882003
[33] GWennemuth D F Babcock and B Hille ldquoCalcium clearancemechanisms of mouse spermrdquo The Journal of General Physiol-ogy vol 122 no 1 pp 115ndash128 2003
[34] HM Florman ldquoSequential focal and global elevations of spermintracellular Ca2+ are initiated by the zona pellucida duringacrosomal exocytosisrdquoDevelopmental Biology vol 165 no 1 pp152ndash164 1994
[35] H M Florman R M Tombes N L First and D F Bab-cock ldquoAn adhesion-associated agonist from the zona pellucidaactivates G protein-promoted elevations of internal Ca2+ andpH that mediate mammalian sperm acrosomal exocytosisrdquoDevelopmental Biology vol 135 no 1 pp 133ndash146 1989
[36] R S Tavares SMansell C L R Barratt S MWilson S J Pub-licover and J Ramalho-Santos ldquo119901 1199011015840-DDE activates CatSperand compromises human sperm function at environmentallyrelevant concentrationsrdquo Human Reproduction vol 28 no 12pp 3167ndash3177 2013
[37] J Xia and D Ren ldquoThe BSA-induced Ca2+ influx during spermcapacitation is CATSPER channel-dependentrdquo ReproductiveBiology and Endocrinology vol 7 article 119 2009
[38] J L Bailey and B T Storey ldquoCalcium influx into mousespermatozoa activated by solubilized mouse zona pellucidamonitored with the calcium fluorescent indicator fluo-3 Inhi-bition of the influx by three inhibitors of the zona pellucidainduced acrosome reaction tyrphostin A48 pertussis toxinand 3-quinuclidinyl benzilaterdquo Molecular Reproduction andDevelopment vol 39 no 3 pp 297ndash308 1994
[39] K K Shukla W-S Kwon M S Rahman Y-J Park Y-AYou and M-G Pang ldquoNutlin-3a decreases male fertility viaUQCRC2rdquo PLoS ONE vol 9 no 8 Article ID e76959 2013
[40] R Yanagimachi ldquoMammalian fertilizationrdquo inThePhysiology ofReproduction E Knobil and J D Neill Eds pp 189ndash317 RavenPress New York NY USA 1994
[41] C R Austin ldquoObservations on the penetration of the sperm inthemammalian eggrdquoAustralian Journal of Scientific Research BBiological Sciences vol 4 no 4 pp 581ndash596 1951
[42] T Strunker N Goodwin C Brenker et al ldquoThe CatSperchannel mediates progesterone-induced Ca2+ influx in humanspermrdquo Nature vol 471 no 7338 pp 382ndash386 2011
[43] A Darszon J J Acevedo B E Galindo et al ldquoSperm channeldiversity and functional multiplicityrdquo Reproduction vol 131 no6 pp 977ndash988 2006
[44] D Pietrobon F di Virgilio and T Pozzan ldquoStructural andfunctional aspects of calcium homeostasis in eukaryotic cellsrdquoEuropean Journal of Biochemistry vol 193 no 3 pp 599ndash6221990
[45] Y Kirichok G Krapivinsky and D E Clapham ldquoThe mito-chondrial calcium uniporter is a highly selective ion channelrdquoNature vol 427 no 6972 pp 360ndash364 2004
[46] J Liu J Xia K-HChoD E Clapham andDRen ldquoCatSper120573 anovel transmembrane protein in the CatSper channel complexrdquoThe Journal of Biological Chemistry vol 282 no 26 pp 18945ndash18952 2007
[47] H Wang J Liu K-H Cho and D Ren ldquoA novel single trans-membrane protein CATSPERG is associated with CATSPER1channel proteinrdquo Biology of Reproduction vol 81 no 3 pp 539ndash544 2009
[48] D Ren B Navarro G Perez et al ldquoA sperm ion channelrequired for sperm motility and male fertilityrdquo Nature vol 413no 6856 pp 603ndash609 2001
[49] D Wang S M King T A Quill L K Doolittle and D LGarbers ldquoA new sperm-specific Na+H+ exchanger required forsperm motility and fertilityrdquo Nature Cell Biology vol 5 no 12pp 1117ndash1122 2003
[50] A E Carlson R E Westenbroek T Quill et al ldquoCatSper1required for evoked Ca2+ entry and control of flagellar functionin spermrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 100 no 25 pp 14864ndash14868 2003
[51] T A Quill S A Sugden K L Rossi L K Doolittle R EHammer and D L Garbers ldquoHyperactivated sperm motilitydriven by CatSper2 is required for fertilizationrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 100 no 25 pp 14869ndash14874 2003
[52] Y Chen M J Cann T N Litvin et al ldquoSoluble adenylyl cyclaseas an evolutionarily conserved bicarbonate sensorrdquo Science vol289 no 5479 pp 625ndash628 2000
[53] D E Clapham ldquoSperm BerserKersrdquo eLife vol 2 Article IDe01469 2013
[54] I Weyand M Godde S Frings et al ldquoCloning and functionalexpression of acyclic-nucleotide-gated channel from mam-malian spermrdquo Nature vol 368 no 6474 pp 859ndash863 1994
[55] S S Suarez and X Dai ldquoHyperactivation enhances mousesperm capacity for penetrating viscoelastic mediardquo Biology ofReproduction vol 46 no 4 pp 686ndash691 1992
[56] H-C Ho and S S Suarez ldquoHyperactivation of mammalianspermatozoa function and regulationrdquo Reproduction vol 122no 4 pp 519ndash526 2001
[57] M Bohmer Q Van I Weyand et al ldquoCa2+ spikes in theflagellum control chemotactic behavior of spermrdquo The EMBOJournal vol 24 no 15 pp 2741ndash2752 2005
[58] B Al-Anzi and D E Chandler ldquoA sperm chemoattractant isreleased from xenopus egg jelly during spawningrdquoDevelopmen-tal Biology vol 198 no 2 pp 366ndash375 1998
[59] S S Suarez and A A Pacey ldquoSperm transport in the femalereproductive tractrdquo Human Reproduction Update vol 12 no 1pp 23ndash37 2006
[60] S S Suarez S M Varosi and X Dai ldquoIntracellular calciumincreases with hyperactivation in intact moving hamster spermand oscillates with the flagellar beat cyclerdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 90 no 10 pp 4660ndash4664 1993
[61] B Marquez and S S Suarez ldquoSoluble adenylyl cyclase isrequired for activation of sperm but does not have a direct effecton hyperactivationrdquo Reproduction Fertility and Developmentvol 20 no 2 pp 247ndash252 2008
[62] J Xia D Reigada C H Mitchell and D Ren ldquoCATSPERchannel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagationrdquo Biology of Reproduction vol 77 no 3 pp551ndash559 2007
BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004

BioMed Research International 13
[63] P E Visconti ldquoUnderstanding the molecular basis of spermcapacitation through kinase designrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 106 no3 pp 667ndash668 2009
[64] P E Visconti V A Westbrook O Chertihin I DemarcoS Sleight and A B Diekman ldquoNovel signaling pathwaysinvolved in sperm acquisition of fertilizing capacityrdquo Journal ofReproductive Immunology vol 53 no 1-2 pp 133ndash150 2002
[65] L R Fraser ldquoSperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 13 no 1 pp 9ndash19 1998
[66] M Michaut C N Tomes G de Blas R Yunes and L SMayorga ldquoCalcium-triggered acrosomal exocytosis in humanspermatozoa requires the coordinated activation of Rab3A and119873-ethylmaleimide-sensitive factorrdquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 97 no18 pp 9996ndash10001 2000
[67] H Breitbart ldquoIntracellular calcium regulation in spermcapacitation and acrosomal reactionrdquo Molecular and CellularEndocrinology vol 187 no 1-2 pp 139ndash144 2002
[68] H Breitbart ldquoSignaling pathways in sperm capacitation andacrosome reactionrdquo Cellular and Molecular Biology vol 49 no3 pp 321ndash327 2003
[69] H Breitbart and Z Naor ldquoProtein kinases in mammaliansperm capacitation and the acrosome reactionrdquo Reviews ofReproduction vol 4 no 3 pp 151ndash159 1999
[70] H Breitbart and B Spungin ldquoThe biochemistry of the acrosomereactionrdquoMolecular Human Reproduction vol 3 no 3 pp 195ndash202 1997
[71] H M Florman M K Jungnickel and K A Sutton ldquoSheddingLight on SpermpHertilityrdquoCell vol 140 no 3 pp 310ndash312 2010
[72] N Etkovitz S Rubinstein L Daniel and H Breitbart ldquoRoleof PI3-kinase and PI4-kinase in actin polymerization duringbovine sperm capacitationrdquo Biology of Reproduction vol 77 no2 pp 263ndash273 2007
[73] P E Visconti and H M Florman ldquoMechanisms of sperm-egg interactions between sugars and broken bondsrdquo ScienceSignaling vol 3 no 142 article pe35 2010
[74] A Abou-haila and D R P Tulsiani ldquoSignal transductionpathways that regulate sperm capacitation and the acrosomereactionrdquo Archives of Biochemistry and Biophysics vol 485 no1 pp 72ndash81 2009
[75] R K Naz and P B Rajesh ldquoRole of tyrosine phosphorylationin sperm capacitationacrosome reactionrdquo Reproductive Biologyand Endocrinology vol 2 article 75 2004
[76] D R P Tulsiani A Abou-Haila C R Loeser and B M JPereira ldquoThebiological and functional significance of the spermacrosome and acrosomal enzymes in mammalian fertilizationrdquoExperimental Cell Research vol 240 no 2 pp 151ndash164 1998
[77] L J D Zaneveld C J de Jonge R A Anderson and S RMack ldquoHuman sperm capacitation and the acrosome reactionrdquoHuman Reproduction vol 6 no 9 pp 1265ndash1274 1991
[78] N Bernabo P Berardinelli A Mauro et al ldquoThe role of actinin capacitation-related signaling an in silico and in vitro studyrdquoBMC Systems Biology vol 5 article 47 2011
[79] C Patrat J Auer P Fauque R L Leandri P Jouannet and CSerres ldquoZona pellucida from fertilised human oocytes inducesa voltage-dependent calcium influx and the acrosome reactionin spermatozoa but cannot be penetrated by spermrdquo BMCDevelopmental Biology vol 6 article 59 2006
[80] R A Steinhardt D Epel E J Carroli and R Yanagimachi ldquoIscalcium ionophore a universal activator for unfertilised eggsrdquoNature vol 252 no 5478 pp 41ndash43 1974
[81] M Theodoridou M Nomikos D Parthimos et al ldquoChimerasof sperm PLC120577 reveal disparate protein domain functions in thegeneration of intracellular Ca2+ oscillations inmammalian eggsat fertilizationrdquoMolecular Human Reproduction vol 19 no 12pp 852ndash864 2013
[82] J R Gonzalez-Garcia ZMachaty F A Lai and K Swann ldquoThedynamics of PKC-induced phosphorylation triggered by Ca2+oscillations in mouse eggsrdquo Journal of Cellular Physiology vol228 no 1 pp 110ndash119 2013
[83] M J Whitaker and R A Steinhardt ldquoIonic regulation of eggactivationrdquo Quarterly Reviews of Biophysics vol 15 no 4 pp593ndash666 1982
[84] L F Jaffe ldquoSources of calcium in egg activation a review andhypothesisrdquo Developmental Biology vol 99 no 2 pp 265ndash2761983
[85] S Miyazaki H Shirakawa K Nakada and Y Honda ldquoEssentialrole of the inositol 145-trisphosphate receptorCa2+ releasechannel in Ca2+ waves and Ca2+ oscillations at fertilization ofmammalian eggsrdquoDevelopmental Biology vol 158 no 1 pp 62ndash78 1993
[86] K Swann and J-P Ozil ldquoDynamics of the calcium signal thattriggers mammalian egg activationrdquo International Review ofCytology vol 152 pp 183ndash222 1994
[87] K Swann and F A Lai ldquoPLC120577 and the initiation of Ca2+oscillations in fertilizing mammalian eggsrdquo Cell Calcium vol53 no 1 pp 55ndash62 2013
[88] A Yu M Nomikos M Theodoridou G Nounesis F A Laiand K Swann ldquoPLC120577 causes Ca2+ oscillations in mouse eggsby targeting intracellular and not plasma membrane PI(45)P
2rdquo
Molecular Biology of the Cell vol 23 no 2 pp 371ndash380 2012[89] S V Phillips Y Yu A Rossbach et al ldquoDivergent effect of
mammalian PLC120577 in generating Ca2+ oscillations in somaticcells compared with eggsrdquo Biochemical Journal vol 438 no 3pp 545ndash553 2011
[90] K Swann and Y Yu ldquoThe dynamics of calcium oscillationsthat activate mammalian eggsrdquo The International Journal ofDevelopmental Biology vol 52 no 5-6 pp 585ndash594 2008
[91] B-G Jeon J-S Moon K-C Kim H-J Lee S-Y Choe andG-J Rho ldquoFollicular fluid enhances sperm attraction andits motility in humanrdquo Journal of Assisted Reproduction andGenetics vol 18 no 8 pp 407ndash412 2001
[92] H-C Ho K A Granish and S S Suarez ldquoHyperactivatedmotility of bull sperm is triggered at the axoneme by Ca2+ andnot cAMPrdquo Developmental Biology vol 250 no 1 pp 208ndash2172002
[93] M Luconi G Forti and E Baldi ldquoPathophysiology of spermmotilityrdquo Frontiers in Bioscience vol 11 no 2 pp 1433ndash14472006
[94] A Revelli D Ghigo F Moffa M Massobrio and I Tur-KaspaldquoGuanylate cyclase activity and sperm functionrdquo EndocrineReviews vol 23 no 4 pp 484ndash494 2002
[95] K Tasken and E M Aandahl ldquoLocalized effects of cAMPmediated by distinct routes of protein kinase Ardquo PhysiologicalReviews vol 84 no 1 pp 137ndash167 2004
[96] K Swann ldquoSoluble sperm factors and Ca2+ release in eggs atfertilizationrdquo Reviews of Reproduction vol 1 no 1 pp 33ndash391996
[97] K Swann M G Larman C M Saunders and F A Lai ldquoThecytosolic sperm factor that triggers Ca2+ oscillations and eggactivation in mammals is a novel phospholipase C PLC120577rdquoReproduction vol 127 no 4 pp 431ndash439 2004





























