Embed Size (px)
DESCRIPTION
x
Citation preview

Cai si retele de transmitere a semnalelor celulare
Celulele organismului functioneaza ca un tot unitar. Ele emit
permanent ,,mesaje” sub forma de molecule specifice care dicteaza celulelor
receptoare ceea ce trebuie sa faca astfel incat organismul sa se adapteze permanent la
conditiile de mediu. Aceste mecanisme de semnalizare celulara sunt repartizate in
mai multe sisteme specializate.
Un sistem de specializare este un dispozitiv care permite transmiterea la
distanta a unei informatii numita semnal. Destinatarul trebuie sa capteze mesajul, s ail
inteleaga sis a il aplice adaptandusi propriul comportament la ,,ordinul” primit.
Semnalizarea biologica corespunde acestei descrieri. Clasic se disting 2 tipuri majore
de semnalizari biologice: semnalizarea nervoasa si semnalizarea hormonala.
In primul caz, mesajul este transmis la distanta dea lungul unui filet nervos prin
intermediul unui fenomen electricinfluxul nervos. In al IIlea caz, mesajul este
transmis sub forma unei substante chimice cu structura definitenumita mesager
primar, care circula pe cale sangvina catre celulele de destinatie. Cele 2 tipuri de
semnalizare, odata ajunse la destinatie, folosesc acelasi tip de molecule si reactii
chimice.
In afara acestor 2 cai majore de semnalizare, exista si sisteme de mesagerie
localizata, care actioneaza la nivelul anumitor organe si care moduleaza sistemul
neuroendocrin.
Mesagerul primar
Liganzii naturali, extracelulari, care se fixeaza la un receptor si pe care ulterior
il activeaza se numesc mesageri primari. Acestia se pot clasifica in hormoni,
neurotransmitatori, citokine, limfocite, factori de crestere (GF), chemoatractanti, etc.
Fiecare din acesti termeni defineste o clasa de agenti care actioneaza intrun mod
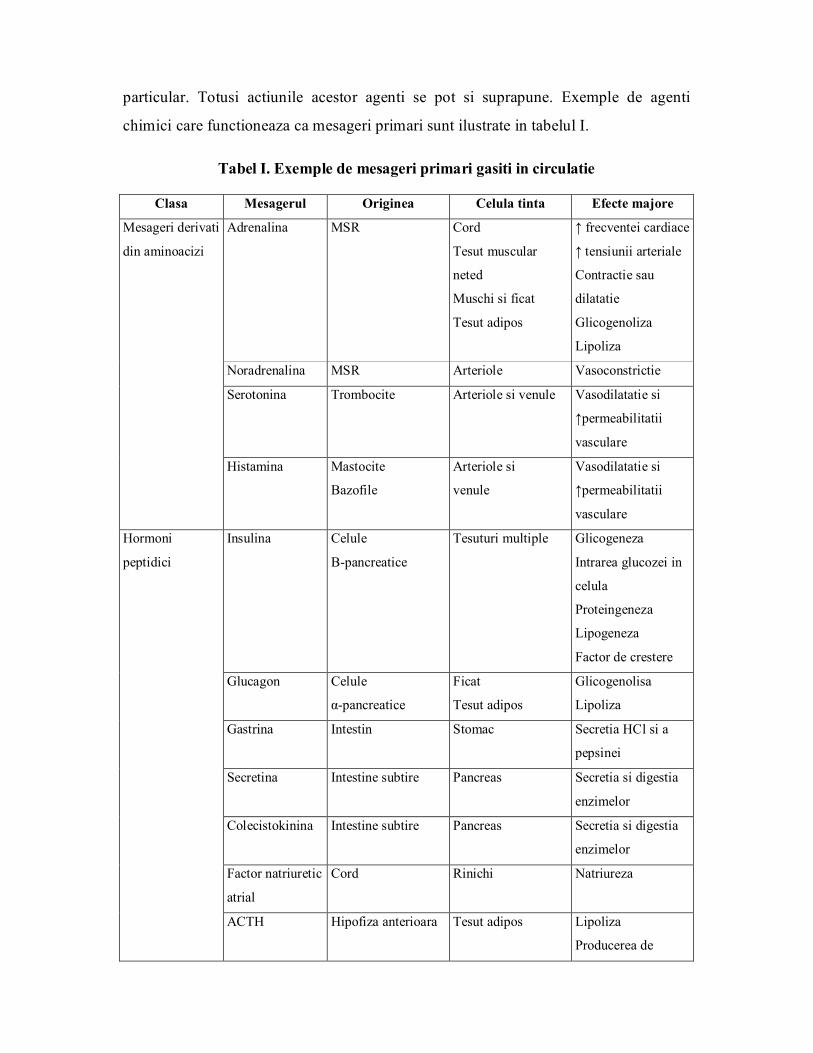
particular. Totusi actiunile acestor agenti se pot si suprapune. Exemple de agenti
chimici care functioneaza ca mesageri primari sunt ilustrate in tabelul I.
Tabel I. Exemple de mesageri primari gasiti in circulatie
Clasa Mesagerul Originea Celula tinta Efecte majore
Adrenalina MSR Cord
Tesut muscular
neted
Muschi si ficat
Tesut adipos
↑ frecventei cardiace
↑ tensiunii arteriale
Contractie sau
dilatatie
Glicogenoliza
Lipoliza
Noradrenalina MSR Arteriole Vasoconstrictie
Serotonina Trombocite Arteriole si venule Vasodilatatie si
↑permeabilitatii
vasculare
Mesageri derivati
din aminoacizi
Histamina Mastocite
Bazofile
Arteriole si
venule
Vasodilatatie si
↑permeabilitatii
vasculare
Insulina Celule
Βpancreatice
Tesuturi multiple Glicogeneza
Intrarea glucozei in
celula
Proteingeneza
Lipogeneza
Factor de crestere
Glucagon Celule
αpancreatice
Ficat
Tesut adipos
Glicogenolisa
Lipoliza
Gastrina Intestin Stomac Secretia HCl si a
pepsinei
Secretina Intestine subtire Pancreas Secretia si digestia
enzimelor
Colecistokinina Intestine subtire Pancreas Secretia si digestia
enzimelor
Factor natriuretic
atrial
Cord Rinichi Natriureza
Hormoni
peptidici
ACTH Hipofiza anterioara Tesut adipos Lipoliza
Producerea de
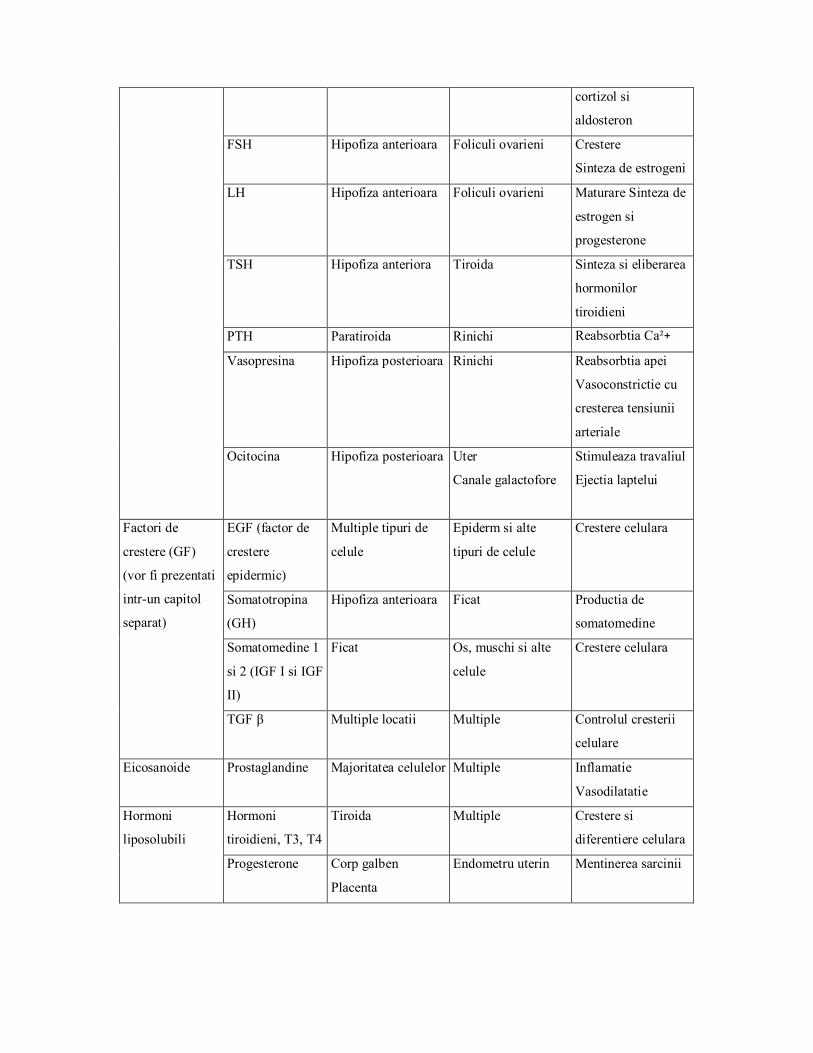
cortizol si
aldosteron
FSH Hipofiza anterioara Foliculi ovarieni Crestere
Sinteza de estrogeni
LH Hipofiza anterioara Foliculi ovarieni Maturare Sinteza de
estrogen si
progesterone
TSH Hipofiza anteriora Tiroida Sinteza si eliberarea
hormonilor
tiroidieni
PTH Paratiroida Rinichi Reabsorbtia Ca²+
Vasopresina Hipofiza posterioara Rinichi Reabsorbtia apei
Vasoconstrictie cu
cresterea tensiunii
arteriale
Ocitocina Hipofiza posterioara Uter
Canale galactofore
Stimuleaza travaliul
Ejectia laptelui
EGF (factor de
crestere
epidermic)
Multiple tipuri de
celule
Epiderm si alte
tipuri de celule
Crestere celulara
Somatotropina
(GH)
Hipofiza anterioara Ficat Productia de
somatomedine
Somatomedine 1
si 2 (IGF I si IGF
II)
Ficat Os, muschi si alte
celule
Crestere celulara
Factori de
crestere (GF)
(vor fi prezentati
intrun capitol
separat)
TGF β Multiple locatii Multiple Controlul cresterii
celulare
Eicosanoide Prostaglandine Majoritatea celulelor Multiple Inflamatie
Vasodilatatie
Hormoni
tiroidieni, T3, T4
Tiroida Multiple Crestere si
diferentiere celulara
Hormoni
liposolubili
Progesterone Corp galben
Placenta
Endometru uterin Mentinerea sarcinii

Hormonii
Hormonii sunt in general secretati in cantitati mici de cellule la distanta de
organele tinta. Odata intrati in circulatie ei sunt diluati in masa de sange, o parte
degradati de enzimele existente iar o parte se fixeaza de proteinele transportoare. Toate
aceste 3 mecanisme micsoreaza foarte mult cantitatea de hormone liber, disponibil
pentru celula tinta. De aceea, pe celula tinta se gasesc receptori de mare afinitate, care
ii fixeaza rapid si declanseaza un raspuns din partea celulei.
GH
Sunt agenti de natura peptidica absolute necesari celulei sa depaseasca punctele
de control “check points” din ciclul cellular. In lipsa lor nu are loc replicarea ADN si
respective proliferarea celulara.
Ca si hormonii, GH au receptori de mare afinitate care pot declansa multiple cai
de semnalizare. Pe langa efectul stimulator (inhibitor) asupra ↑ celulare, acesti agenti
chimici mai pot initia moartea celulara programata (apoptoza), diferentierea celulara si
expresia genica. Fata de hormoni, in general GH actioneaza local, de tip paracrin /
autocrin.
Citokinele
In paralel cu descoperirea GH, sau identificat si agenti chimici care
interactioneaza cu celulele sistemului imun. Deoarece acesti compusi activeaza /
moduleaza proliferarea acestei clase de celule, initial au fost denumite imunocitokine.
Dar pentru ca actioneaza si pe celule din afara sistemului imun, actual se accepta
termenul de citokine. Functia GH si a citokinelor e atat de diversa incat o distinctie
intre aceste 2 clase nu se poate face. Astfel citokinele includ atat GH de tipul PDGF,
EGF, insulina cat si molecule ca interferon, TNFα, numeroase interleukine, GMCSF
si altii. Chemokinele sunt citokine care au rol in inflamatie prin recrutarea celulelor
inflamatorii prin chemoxie la nivelul focarului inflamator si ulterior activarea
acestor celule.
Agenti vasoactivi

Orice distructie tisulara (cauzata fizic sau de un agent infectios) genereaza un
raspuns inflamator. Pentru recrutarea leucocitelor are loc o vasodilatatie si o crestere a
permeabilitatii vaselor locale. Aceste fenomene sunt declansate de histamine (eliberata
din bazofile si mastocite), serotonina (eliberata de trombocite) si de agenti
proinflamatori ca bradikinina.
Eicosanoizii sunt o alta clasa cu efecte vasoactive. Ei deriva din acidul
arahidonic si cuprind prostaglandine (PG), tromboxani (Tx), leucotriene (LT). Acesti
agenti actioneaza in maniera paracrina si autocrina, controland multiple functii
celulare atat in conditii fiziologice cat si patologice.
Neurotransmitatori si neuropeptide
Neurotransmitatorii sunt de asemenea, mesageri primari dar sunt eliberati si
detectati la nivelul sinapselor. In celula presinaptica neurotransmitatorul este stocat in
niste vezicule. In momentul depolarizarii membranei presinaptice, se produce fuziunea
acestor vezicule cu membrana si elibereaza neurotransmitatori in fanta sinaptica. Din
fanta, prin difuzie, mediatorul ajunge la nivelul membranei postsinaptice unde se
fixeaza pe receptorul de mare afinitate, initiind un influx nervos sau raspuns cellular.
In SNC, glutamatul are efect in principal excitator, in timp ce acidul gmaaminobutiric
(GABA) si glicina sunt inhibitori. Acetilcolina (Ach) isi exercita functia in special pe
jonctiunea neuromusculara si are efect excitator.
Hormoni lipofilici
Aceasta clasa de hormoni difera de hormonii hidrosolubili prin mai multe
caracteristici. Din punct de vedere structural sunt steroizi si hormoni tiroidieni. Fiind
liposolubili, in plasma circula fixati de proteine transportoare. Odata ajunsi la nivelul
celulelor tinta, penetreaza membrana celulei si isi gaseste receptorul fie in citoplasma,
fie in nucleu.
Complexul HR format va trece in nucleu, unde se va fixa pe o regiune
specifica din promotorul unei gene – numita HRE ( “hormone response element”) –
ceea ce va determina transcriptia acelei gene si secundar sinteza unei proteine. In afara
influentei asupra transcriptiei genice, acesti hormoni pot actiona si prin alte mecanisme.

De exemplu, progesteronul are 2 efecte principale: intracelular activeaza transcriptia
unor proteine specifice implicate in mentinerea sarcinii, iar la nivel membranei
celulare stimuleaza internalizarea receptorului pentru oxitocina, scazand efectul
acestui hormon asupra uterului gravid.
Astfel in tabelul II sunt prezentate diferentele generale intre hormonii
hidrosolubili si liposolubili.
Tabel .II. Principalele caracteristici ale hormonilor hidrosolubili si liposolubili
Hormoni hidrosolubili Hormoni liposolubili
Transport sangvin Liber Legat de proteina specifica
T½ ↓ ↑
Receptor Membranar Intracelular
Mesager da nu
Receptori hormonali
Sensibilitatea unei celule la actiunea unui hormone este determinate de prezenta
pe suprafata / intracelular a unui component proteic capabil sa recunoasca hormonal,
sal fixeze sis a initieze raspunsul specific. Aceasta proteina se numeste receptor.
Specificitatea receptorului este foarte mare.
Substantele care fixate pe receptori ii activeaza se numesc agonisti, iar
substantele care fixate pe receptori ii inhiba se numesc antagonisti.
Dintre receptorii celulari, cei mai bine studiati sunt receptorii pentru Ach si receptorii
pentru adrenalina (A), noradrenalina(NA).
Receptorul pentru Ach
Ach este un mesager primar care interactioneaza cu receptorul si care nu are
functie hormonala. Se gaseste la nivelul sinapselor sistemului nervos somatic si in
sinapsele sistemului nervos parasimpatic.
Clasic receptorii pentru Ach sunt nicotinici si muscarinici.
Receptorii muscarinici se gasesc la nivelul cordului si prin stimulare scad frecventa
cardiaca si forta de contractie. Apartin familiei de receptori care traverseaza de 7 ori
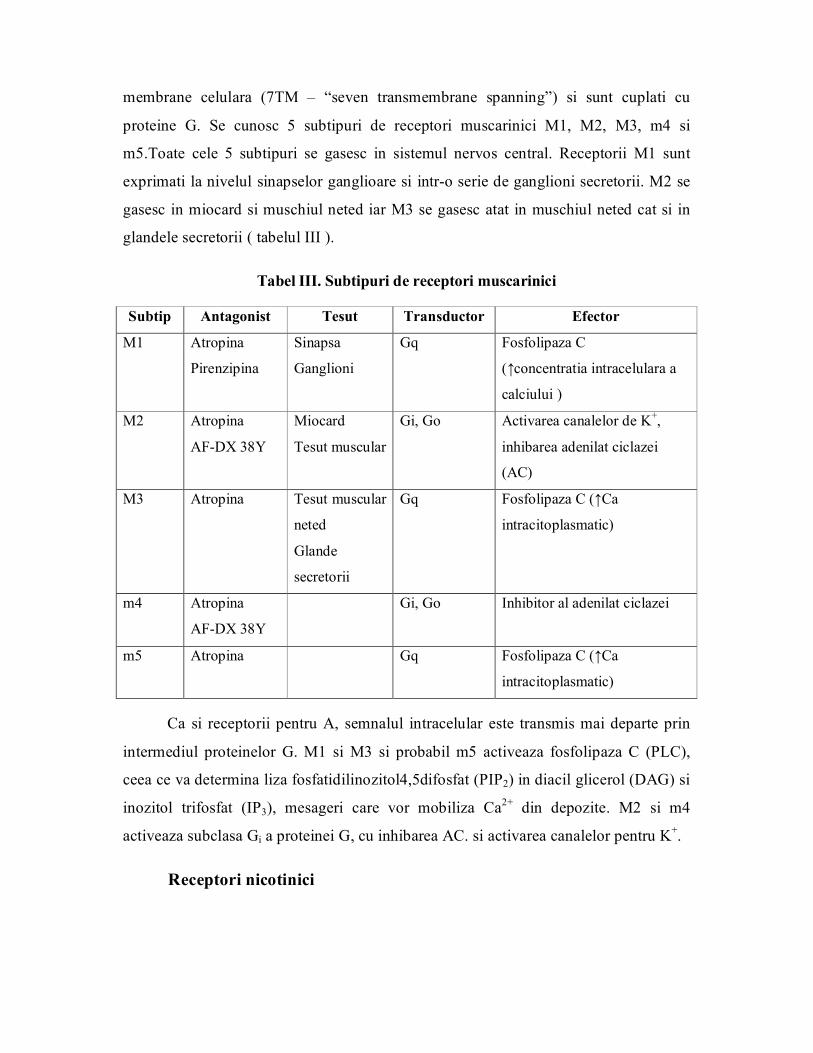
membrane celulara (7TM – “seven transmembrane spanning”) si sunt cuplati cu
proteine G. Se cunosc 5 subtipuri de receptori muscarinici M1, M2, M3, m4 si
m5.Toate cele 5 subtipuri se gasesc in sistemul nervos central. Receptorii M1 sunt
exprimati la nivelul sinapselor ganglioare si intro serie de ganglioni secretorii. M2 se
gasesc in miocard si muschiul neted iar M3 se gasesc atat in muschiul neted cat si in
glandele secretorii ( tabelul III ).
Tabel III. Subtipuri de receptori muscarinici
Subtip Antagonist Tesut Transductor Efector
M1 Atropina
Pirenzipina
Sinapsa
Ganglioni
Gq Fosfolipaza C
(↑concentratia intracelulara a
calciului )
M2 Atropina
AFDX 38Y
Miocard
Tesut muscular
Gi, Go Activarea canalelor de K + ,
inhibarea adenilat ciclazei
(AC)
M3 Atropina Tesut muscular
neted
Glande
secretorii
Gq Fosfolipaza C (↑Ca
intracitoplasmatic)
m4 Atropina
AFDX 38Y
Gi, Go Inhibitor al adenilat ciclazei
m5 Atropina Gq Fosfolipaza C (↑Ca
intracitoplasmatic)
Ca si receptorii pentru A, semnalul intracelular este transmis mai departe prin
intermediul proteinelor G. M1 si M3 si probabil m5 activeaza fosfolipaza C (PLC),
ceea ce va determina liza fosfatidilinozitol4,5difosfat (PIP2) in diacil glicerol (DAG) si
inozitol trifosfat (IP 3 ), mesageri care vor mobiliza Ca 2+ din depozite. M2 si m4
activeaza subclasa Gi a proteinei G, cu inhibarea AC. si activarea canalelor pentru K + .
Receptori nicotinici

Se gasesc la nivelul placii motorii neuromusculare, stimuland contractia
musculaturii scheletice. Acest tip de receptori functioneaza ca niste canale nespecifice
atat pentru Na+ cat si pentru K.
Acesti receptori apartin familiei de receptori ionotropi alaturi de receptorii
pentru serotonina, glicina, GABA, ultimii 2 fiind canale pentru anioni, raspunzand la
liganzi care determina inhibitie.
Receptorul nicotinic este un hexamer membranar, cu doua subunitati identice, si
complexul cuprinde α2, β, γ (fig. 1)(fig. 3.7). Toate subunitatile sunt necesare pentru
buna functionare a receptorilor. ACH se fixeaza in niste buzunare formate la
interfetele celor doua subunitati α cu δ si γ. Ambele situsuri trebuie sa fie ocupate
pentru ca receptorul sa se activeze sis a trimita mai departe semnalul celular. Structura
primara a subunitatilor arata un grad mediu de omologie (35 40%) intre subunitati
sugerand ca ar avea un stramos ancestral comun.
Fig. 1 Organizarea receptorului nicotinic
Fiecare subunitate este alcatuita dintrun peptid care traverseaza de 4 ori
membrane, aceste domenii transmembranare fiind notate M1, M2, M3, M4 si sunt
formate din aminoacizi hidrofobi. Atat capatul N terminal cat si C terminal al fiecarei
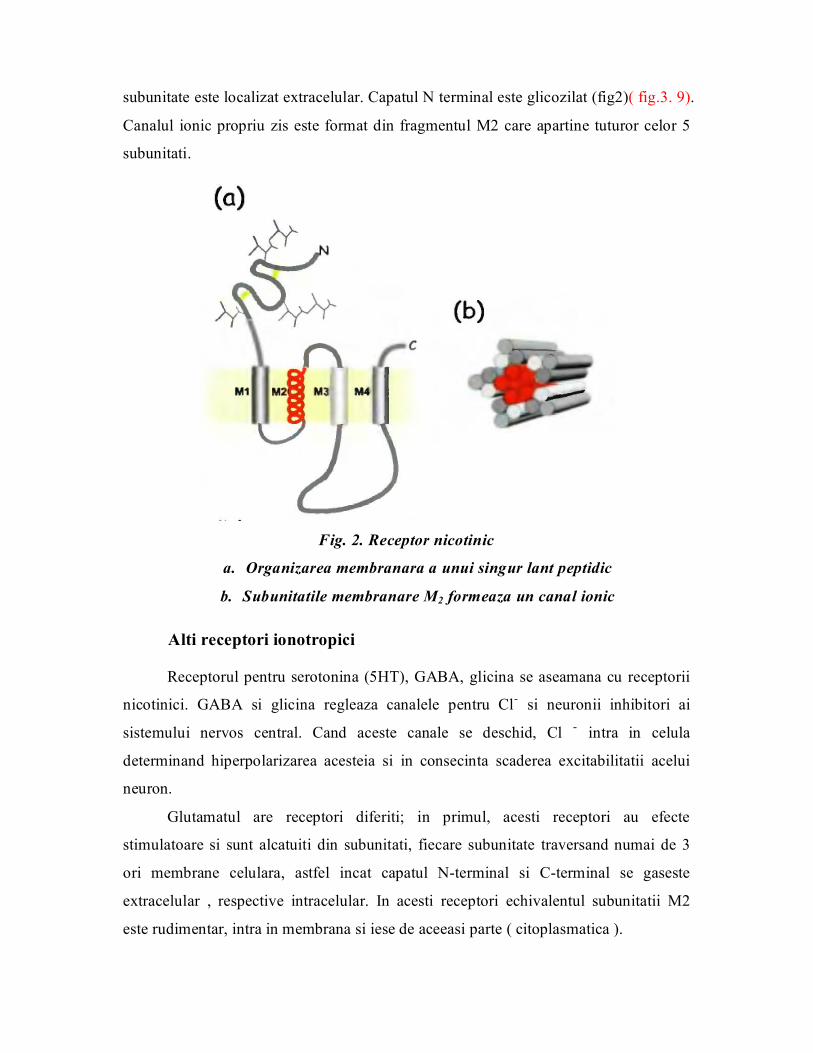
subunitate este localizat extracelular. Capatul N terminal este glicozilat (fig2)( fig.3. 9).
Canalul ionic propriu zis este format din fragmentul M2 care apartine tuturor celor 5
subunitati.
Fig. 2. Receptor nicotinic
a. Organizarea membranara a unui singur lant peptidic
b. Subunitatile membranare M2 formeaza un canal ionic
Alti receptori ionotropici
Receptorul pentru serotonina (5HT), GABA, glicina se aseamana cu receptorii
nicotinici. GABA si glicina regleaza canalele pentru Cl si neuronii inhibitori ai
sistemului nervos central. Cand aceste canale se deschid, Cl intra in celula
determinand hiperpolarizarea acesteia si in consecinta scaderea excitabilitatii acelui
neuron.
Glutamatul are receptori diferiti; in primul, acesti receptori au efecte
stimulatoare si sunt alcatuiti din subunitati, fiecare subunitate traversand numai de 3
ori membrane celulara, astfel incat capatul Nterminal si Cterminal se gaseste
extracelular , respective intracelular. In acesti receptori echivalentul subunitatii M2
este rudimentar, intra in membrana si iese de aceeasi parte ( citoplasmatica ).
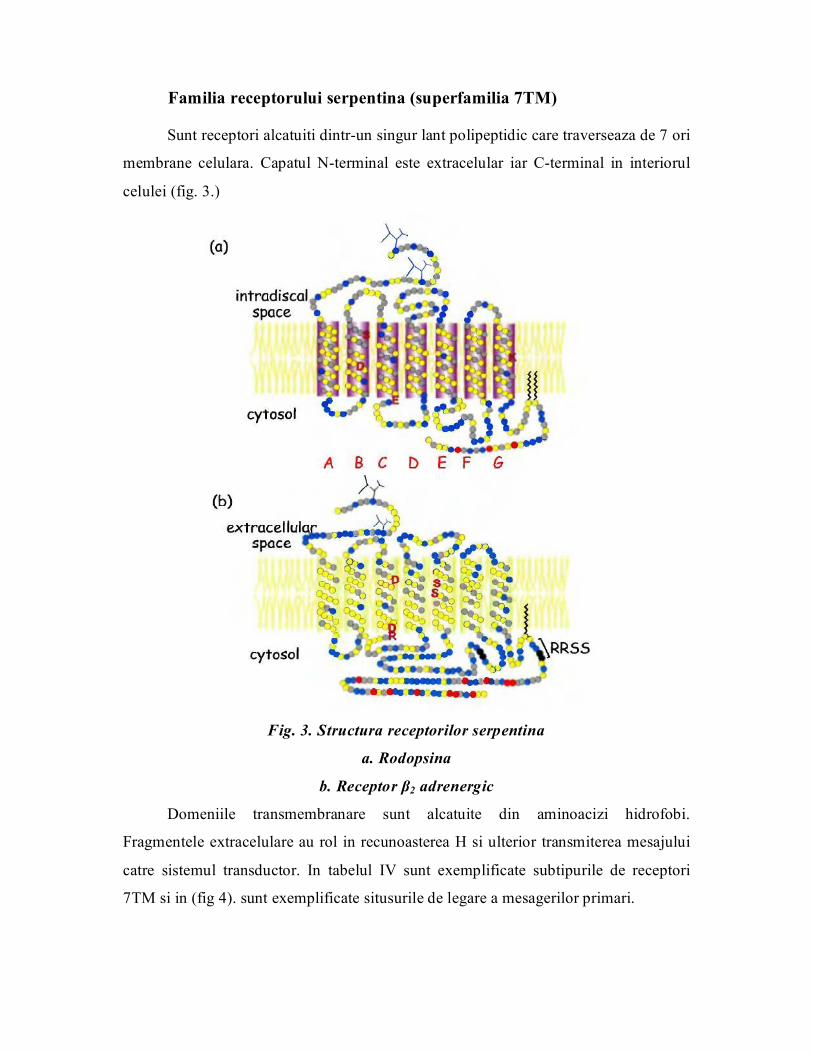
Familia receptorului serpentina (superfamilia 7TM)
Sunt receptori alcatuiti dintrun singur lant polipeptidic care traverseaza de 7 ori
membrane celulara. Capatul Nterminal este extracelular iar Cterminal in interiorul
celulei (fig. 3.)
Fig. 3. Structura receptorilor serpentina
a. Rodopsina
b. Receptor β2 adrenergic
Domeniile transmembranare sunt alcatuite din aminoacizi hidrofobi.
Fragmentele extracelulare au rol in recunoasterea H si ulterior transmiterea mesajului
catre sistemul transductor. In tabelul IV sunt exemplificate subtipurile de receptori
7TM si in (fig 4). sunt exemplificate situsurile de legare a mesagerilor primari.

Fig. 4. Situsurile de legare ale receptorilor serpentina
Tabel nr IV. Subtipuri de receptori serpentina
Proprietatile receptorului Ligand
Ligandul se fixeaza intrun miez
alcatuit din toate cele 7 elicii
transmembranare
11cis retinal (rodopsina)
ACH
Catecolamine
Amine biogene (histamina, serotonina)
Nucleozide si nucleotide
LT, PG, PGi, Tx
Ligandul (un peptid scurt) se fixeaza
partial in regiunea hidrofoba la nivelul
buclelor extracelulare
Hormoni peptidici: ACTH, glucagon,
GH, PTH, calcitonina
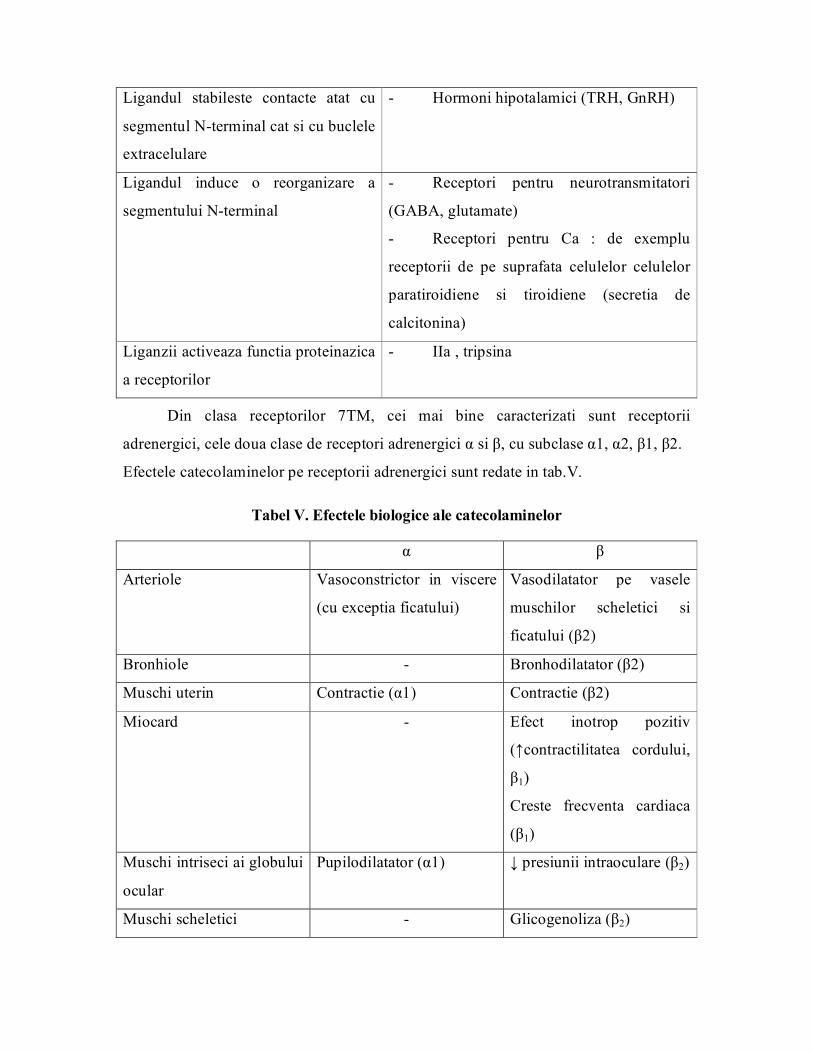
Ligandul stabileste contacte atat cu
segmentul Nterminal cat si cu buclele
extracelulare
Hormoni hipotalamici (TRH, GnRH)
Ligandul induce o reorganizare a
segmentului Nterminal
Receptori pentru neurotransmitatori
(GABA, glutamate)
Receptori pentru Ca : de exemplu
receptorii de pe suprafata celulelor celulelor
paratiroidiene si tiroidiene (secretia de
calcitonina)
Liganzii activeaza functia proteinazica
a receptorilor
IIa , tripsina
Din clasa receptorilor 7TM, cei mai bine caracterizati sunt receptorii
adrenergici, cele doua clase de receptori adrenergici α si β, cu subclase α1, α2, β1, β2.
Efectele catecolaminelor pe receptorii adrenergici sunt redate in tab.V.
Tabel V. Efectele biologice ale catecolaminelor
α β
Arteriole Vasoconstrictor in viscere
(cu exceptia ficatului)
Vasodilatator pe vasele
muschilor scheletici si
ficatului (β2)
Bronhiole Bronhodilatator (β2)
Muschi uterin Contractie (α1) Contractie (β2)
Miocard Efect inotrop pozitiv
(↑contractilitatea cordului,
β 1 )
Creste frecventa cardiaca
(β 1 )
Muschi intriseci ai globului
ocular
Pupilodilatator (α1) ↓ presiunii intraoculare (β 2 )
Muschi scheletici Glicogenoliza (β 2 )

Ficat Glicogenoliza (α 1 ) Glicogenoliza (β 2 )
Tesut adipos Inhibarea lipolizei (α2) Lipoliza (β1)
Pancreas Inhiba secretia de insulina
(α 2 )
Stimuleaza secretia de
insulina (β 2 )
Tr Inhiba agregarea Tr (α 2 )
Fixarea ligandului la receptor, determina modificarea informationala a acestuia,
cu deplasarea buclelor extracelulare (fig. 5) in jurul ligandului si a buclelor
intracelulare in jurul subunitatii α a proteinei G, ceea ce va determina activarea
acesteia.
Fig. 5. Deplasarea elicelor intramembranare determina modificarea conformatiei
proteinei G cu activarea sa

Fosforilarea receptorilor si inhibarea transmisiei celulare
Receptorii muscarinici si adrenergici, fosforilati la resturile de la serina/treonina
din capatul C terminal (fig. 3) devin tinte pentru legarea de arestina care astfel
ii ,,prepara” eliminarea lor prin endocitoza. Acest mechanism are importanta la nivelul
sinapselor, prevenind transmiterea in exces a influxului nervos in neuronal
postsinaptic/organul tinta.Mai mult, in timp ce fosforilarea previne comunicarea cu
proteinele G (desensibilizarea receptorilor), legarea (necovalenta) de arestina il
transforma in substrat pentru protein kinaza tirozin specifica solubila Src. Aceasta
noua fosforilare initiaza o noua cale de semnalizare pe seama MAPkinazelor.
Pe masura ce concentratia stimulului creste, fosforilarea receptorului (si in consecinta
inactivarea lui) se realizeaza cu doua enzime: PKA (protein kinaza cAMPdep) si
βARK (β adrenergic receptor kinase). Cele doua kinaze actioneaza situsuri diferite din
domeniul C terminal a receptorului (fig.3). Activarea PKA se face in conditiile unei
concentratii medii a stimulului, in timp ce activarea βARK se produce la concentratii
crescute ale ligandului. Prin fosforilare, receptorul devine inactive sin u mai poate
activa anticorpii, deci transmitere semnalului este blocata.
PKA fosforileaza atat receptorul care a activateo cat si receptorii situati in apropiere si
care prezinta aceeasi secventa de aminoacizi (desensibilizare heterologa). βARK,
activata de sbunitatea βγ, fosforileaza numai receptorul care are in apropiere aceste
subunitati (desensibilizarea homologa) (fig. 6).
Fig. 6. Desensibilizarea receptorilor β adrenergici
a. de tip heterolog
b. de tip homolog

Proteinele G
Din punct de vedere structural se disting doua clase de proteine G: trimere (50
KDa) si monomere (21 KDa).Primele sunt permanent fixate de fata interna a
membranei plasmatice, pe cand proteinele mici, monomere, circula liber in citoplasma.
Ambele tipuri de proteine G (trimere si monomere) au rolul de a traduce celulei
semnalul primit de la complexul HR, fenomen denumit transductie.
Proteinele G trimere au fost primele descries si sunt formate din 3 subunitati
α, β, γ. Subunitatea α contine situsul de fixare pentru GDP (in repaus). In repaus
proteinele G fixeaza GDP si este ancorata de fata interna a membranei prin legaturi
hidrofobe.
Activarea proteinelor G este redata in (fig7).
Fig. 7. (1) activarea proteinei G prin schimb cu GTP;
(2) activarea GTPazei intrinseci cu revenirea la forma de repaus a proteinei G
In repaus, proteina G este in contact cu receptorii si are fixat de subunitatea α
nucleotidul GDP.In momentul fixarii H de R, are loc o modificare conformationala a
receptorilor si rapid subunitatea α pierde GDP si fixeaza GTP. In acest moment se
produce si disocierea complexului cu eliberarea HR care poate fixa si active o noua
proteina G. Astfel, in acest punct are loc o prima amplificare a semnalului Indus de
ligand. Proteinele G disociaza: αGTP de βγ, iar αGTP va modula activitatea unei
enzime implicate in sinteza unei molecule mesager secund.

Ireversibilitatea ciclului este determinate de clivarea moleculei GTP in GDP si
Pa, de catre GTPaza intriseca subunitatea α. GDP ramane fixata de subunitatea α, care
revine la forma de repaus, recupleaza βγ si ciclul se poate relua.
Recent sau descoperit o clasa de proteine RGS (regulator of Gprotein
signaling), care contine cel putin 20 de membri, implicate in activarea functiei GTP
azice a subunitatii α a proteinei G; actionand ca reglatori de mare viteza a unor variate
procese fiziologice. Aceste proteine G heterotrimere sunt cuplate cu receptorul 7TM.
Proteinele G sunt diferite prin subunitatea α, care ii imprima functii diverse. Se cunosc
pana in present patru mari clase de proteine G: Gs, Gi, Gq111, G12 cu numeroase
subclase (fig. 8).
Fig. 8. Omologia structurala dintre subunitatile α ale proteinei G
Actual si dimerului βγ i se atribuie functii diverse:
• asigura localizarea , cuplarea efectiva si dezactivarea subunitatii α;
• regleaza afinitatea receptorului pentru liganzii sai;
• reduce tendinta GDP de a disocia pe subunitatea α (astfel stabilizand formele
inactive);
• toxina pertusis produce ADPribozilarea subunitatii α numai in prezenta
dimerului βγ;

• interactioneaza direct cu o serie de efectori situati mai jos in lantul de
transmitere intracelulara;
• regleaza fosforilarea unor receptori de catre kinazele specifice.
Proteinele G activate (forma αGTP) transmit semnalul unor enzyme
generatoare de mesager second: astfel subgrupa αsGTP activeaza AC, enzima
responsabila de transformarea ATP in cAMP, compus de numit mesager second
deoarece sinteza sa este dependenta de mesagerul primar hormonul. Mai mult, odata
sintetizat, mesagerul secund transmite intracelular semnalul preluat de la primul
mesager. cAMP este o molecula mica, hidrosolubila, care difuzeaza rapid in
citoplasma activand o kinaza specifica denumita protein kinaza cAMP dependenta
(PKA). Din punct de vedere structural PKA este un tetramer format din 2 subunitati
reglatorii R si 2 subunitati catalitice C (R2C2). Mediatorul secund cAMP se fixeaza de
subunitatile R ale PKa disociindule de subunitatile C, care fiind libere devin active.
PKa astfel activate, recunoaste numeroase substrate:
• fosforileaza proteine care devin “canale pentru apa”, asiguranduse astfel
la nivel renal reabsortia apei;
• fosforileaza proteine de pe diverse cai metabolice – de exemplu
fosforilazkinaza care devenind active prin fosforilare, va active glicogen
fosforilaza, enzima implicata in glicogenoliza;
• fosforileaza unele proteine care migrand catre nucleu influenteaza
transcriptia unor gene.
Controlul transmiterii intracelulare se realizeaza prin modularea in sens negativ
a acestei cai, la diferite niveluri. Primul punct de control este la nivelul receptorului,
care prin fosforilare se produce fenomenul de desensibilizare. Al doilea punct de
control este la nivelul proteinei G, prin activarea GTPazei intrinseci subunitatii alfa,
seproduce hidroliza GTP la GDP si fosfat anorganic. GDp ramane legat de subunitaea
alfa care astfel revine la forma de repaus. Al treilea punct de control este la nivelul
cAMP, in citoplasma existand fosfodiesteraze care il hidrolizeaza la AMP, astfel incat
T1/2 al acestui mesager second sa fie foarte scurt. Al patrulea punct de control este la
nivelul proteinelor fosforilate, in celula existand protein fosfataze care prin activare,
defosforileaza proteinele, influentand astfel functia lor.

In organism , in afara de subclasa Gs, mai exista si alte clase de proteine G.
Astfel proteina Gi (formata din αiGDP βγ) prin activare (αiGTP) determina inhibarea
activitatii AC, cu efecte opuse celor obtinute prin activarea proteinei Gs.
O alta clasa de proteine G, este reprezentata de proteinele Gq, care structural
sunt alcatuite tot din 3 lanturi αqGDP βγ. Prin activare se formeaza αqGTP care va
activa PLC. Aeasta enzima membranara va hidroliza un fosfolipid membranar PIP2 in
DAG si IP3. DAG este hidrofob si va ramane acorat in membrane si va active o clasa
de kinaze numite protein kinaze C (PKC). PKC va fosforila o serie de proteine
citoplasmatice, modificandule conformatile si respectiv functiile. Celalalt mesager
second, IP3 este hidrosolubil si va difuza in citoplasma, fixanduse pe reticulul
endoplasmatic, unde va determina deschiderea unor canale de Ca2+, cu cresterea
concentratiei acestui cation in celula. Ca2+ are diverse efecte biologice, fiind
considerat mesager secund.
Receptori intracelulari
Hormonii liposolubili (hormonii steroizi, vit.A, hormonii tiroidieni) au receptori
nucleari. Unii hormoni steroizi (aldosteron, estrogeni) au atat receptori nucleari cat si
membranari. Receptorii pentru glucocorticoizi (cei mai bine caracterizati), progesteron,
testosteron, se gasesc in repaus, in citoplasma cuplati cu proteime hsp (,,heat sheck
proteins”) in momentul fixarii ligandului de receptor, proteinele hsp se decupleaza si
formeaza ca niste ,,canale” la nivelul membranei nucleare, permitand complexului H
R sa treaca din ciitoplasma in nucleu (fig. 9). In nucleu complexul HR se fixeaza in
promotorul unei gene pe o secventa numita HRE (,,hormone response element”).
Aceasta secventa HRE este bine definita si pentru a activa transcriptia genei,
complexul HR dimerizeaza formand homodimeri.
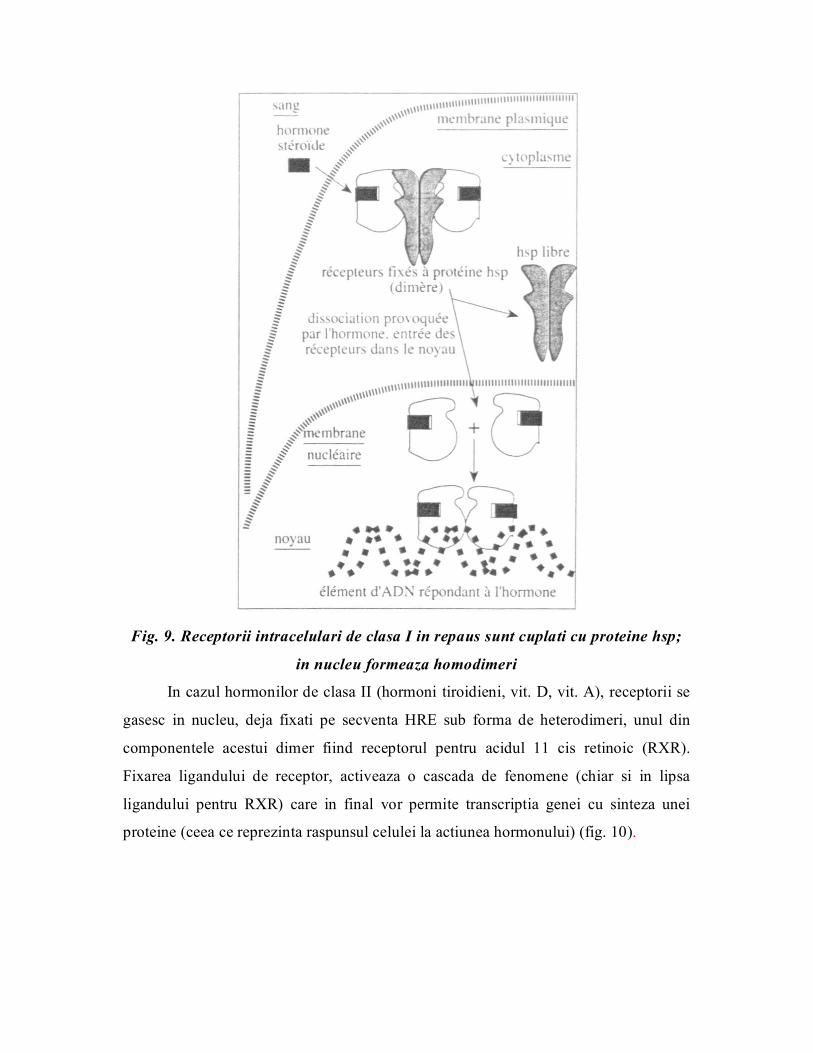
Fig. 9. Receptorii intracelulari de clasa I in repaus sunt cuplati cu proteine hsp;
in nucleu formeaza homodimeri
In cazul hormonilor de clasa II (hormoni tiroidieni, vit. D, vit. A), receptorii se
gasesc in nucleu, deja fixati pe secventa HRE sub forma de heterodimeri, unul din
componentele acestui dimer fiind receptorul pentru acidul 11 cis retinoic (RXR).
Fixarea ligandului de receptor, activeaza o cascada de fenomene (chiar si in lipsa
ligandului pentru RXR) care in final vor permite transcriptia genei cu sinteza unei
proteine (ceea ce reprezinta raspunsul celulei la actiunea hormonului) (fig. 10).

Fig. 10. Receptorii de clasa II formeaza heterodimeri cu RXR (H 2 )
In cazul receptorilor pentru glucocorticoizi, sa evidentiat faptul ca acesti
receptori se pot fixa nu numai pe ADN, ci si direct pe factorul de transcriptie ca de
exemplu AP1 si NFKB pentru a inhiba transcriptia unor factori implicati in inflamatie
(de ex: diverse interleukine, proteine de faza acuta, etc.) sau in imunitate (lantul usor al
imunoglobulinei) si prin aceste mecanisme se poate explica efectul antiinflamator al
glucocorticoizilor.
FACTORI DE CREŞTERE
1. INTRODUCERE Factorii de creştere sunt polipeptide care se
leagă de receptori celulari modulând creşterea,
diferenţierea şi funcţia celulei respective.
Aceasta este o definiţie largă care include
multe substanţe hormoni, diferite peptide,
citokinele care pot funcţiona ca factori de
creştere. Totuşi, prin convenţie, o serie de peptide
sunt cunoscute sub numele de factori de creştere
ACRONIME UTILIZATE
EGF ="Epidermal growth factor"
TGFa, TGFp="Transforming growth
factor"
FGF="fibroblast growth factor"
PDGF="plateletderived growth factor"
IGF="insulin like growth factor"
VEGF="vascular endothelial growt factor"
NGF="nerve growt factor"
ESF="Erythropoesis stimulating factor"

(Fc). Aceste peptide reglează diverse momente
din ciclul celular (figura. 1).
CSF="Colony stimulating factor"
IL=interleukine
Figura 1: Fazele ciclului celular.
Clasic sunt cunoscute patru faze ale ciclului celular: faza S, de replicare a ADN
şi faza M, de mitoză, separate prin două faze "gap" G] şi G2. Factorii de creştere
controlează atât trecerea din Go în Gi (şi invers) cât şi o serie de "puncte de control"
("chekpoints") pentru siguranţa evenimentelor ce au loc în timpul ciclului celular.
"Punctele de control" permit celulei săşi repare ADN, iar dacă leziunile ADN sunt
ireversibile, sunt activate o serie de mecanisme care declanşează fenomenul de
apoptoză.
Importanţa cunoaşterii unor factori de creştere rezultă din faptul că o serie de
afecţiuni includ în patogenia lor o proliferare anormală a unor celule: neoplaziile,
ateroscleroza, bolile reumatoide, ciroza hepatică etc.
Numărul factorilor de creştere descrişi până acum e relativ mare şi mereu se
descriu alţi factori. Cunoştinţele privind factorii de creştere au permis gruparea lor în
familii după asemănări structurale, după natura celulelor asupra cărora acţionează şi a
modului în care intervin în proliferarea şi diferenţierea celulară.

2. CLASIFICARE Unii factori de creştere au acţiuni asupra mai multor tipuri de celule, în timp ce alţii
acţionează numai pe un singur tip celular. Astfel, din prima categorie fac parte:
familia EGF ("Epidermal growth factor") cuprinde EGF, αTGF (α
"Transforming growth factor"), amfiregulina, "heparinbinding EGF like growth
factor", betacelulina, epiregulina şi neureguline;
familia FGF ("fibroblast growth factor") cuprinde FGF1 (FGF acid), FGF2 (FGF bazic) şi încă cel puţin 17 peptide înrudite;
familia PDGF ("plateletderived growth factor") cuprinde ca reprezetant
PDGF;
familia TGFβ ("Transforming growth factor (3") cuprinde trei izoforme
TGFβi, TGF(β2 şi TGF(β3, BMP ("bone morphogenic protein") şi activina;
familia IGF ("insulin like growt factor") cuprinde IGF I (somatomedina C)
şi IGF II;
familia VEGF ("vascular endothelial growth factor") cuprinde şase mebri:
VEGFA, VEGFB, VEGFC, VEGFD, VEGFE, PIGF. Pe lângă aceşti factori există
şi alţii care acţionează numai pe un anumit grup de celule;
NGF ("nerve growth factor") stimulează diferenţierea şi creşterea
terminaţiilor axonale;
eritropoietina (ESF "Erythropoesis stimulating factor") stimulează
eritropoieza;
familia CSF ("Colony stimulating factor") cuprinde factori de creştere care
stimulează proliferarea precursorilor leucocitari. Sau descris patru tipuri:
o "granulocytemacrophage CSF"
o "granulocyteCSF" "macrophageCSF"
o "multi CSF" (identic cu interleukina 3)
o interleukinele (IL) limfocitele produc o serie de proteine (altele decât
imunoglobuline) care acţionează autocrin, paracrin sau endocrin,
exercitând funcţii asupra componentelor sistemului imun.
3. STRUCTURĂ

Din punct de vedere strucutral factorii de creştere sunt formate dintrun singur
lanţ (EGF) sau din două lanţuri polipeptidice (PDGF, CSF etc).
Mai multe se cunosc despre factorii de creştere ce formează familia EGF
care toţi au un domeniu comun care este format din 4555 de aminoacizi şi care
include şase resturi de cisteină ce formează, datorită legăturilor SS, trei bucle. Acest
domeniu este bine conservat de la un factor la altul al familiei EGF şi are rol atât în
ataşarea factorului de creştere de receptorul specific cât şi în activarea acestuia.
Factorii IGF din punct de vedere structural se aseamănă cu proinsulina (vezi
cap. ) având omologie structurală de 52% cu insulina, între cei doi factori existând o
omologie structurală de 62%.
PDGF din punct de vedere structural este dimer, fiecare lanţ conţinând câte
aproximativ 112 aminoacizi. Există două tipuri de lanţuri A şi B care pot forma
molecula de PDGF. Din combinarea acestor două tipuri de lanţuri rezultă trei izoforme
pentru PDGF: AA, BB şi AB.
Factorii FGF (FGF1, FGF2) sunt proteine cu aproximativ 146 aminoacizi şi 18
kD care conţin un domeniu de fixare la heparansulfat/heparină esenţial pentru funcţia
lor.
Factorii VEGF sunt homodimeri, cu organizare structurală similară PDGF. S
au descris şi VEGP heterodimeri.
4. RECEPTORII PENTRU FACTORII DE CREŞTERE
Receptorii pentru factorii de creştere sunt proteine transmembranare care
străbat o singură dată membrana celulei. Prezintă un domeniu extracelular, Nterminal,
un domeniu transmembranar cu structură de tip ochelix şi un domeniu
intracitoplasmatic, Cterminal.
Domeniul Nterminal are rolul de a fixa ligantul (factorul de creştere);
domeniul Cterminal prezintă o funcţie enzimatică intrinsecă de tirozinkinază (TK)
care se activează în momentul fixării ligandului de receptor. Odată activată, TK
intrinsecă fosforilează resturi de tirozină din structura unor proteine adiacente
receptorului. Receptorul fosforilat poate transmite mai departe semnalul mitogen.
În funcţie de structura lor, receptorii pentru factorii de creştere se pot grupa în:
Receptori de clasa I pentru EGF cuprinde patru receptori:

receptor pentru EGF (sau Erb 1 )
proteina HER2 (sau Erb2/neu) la care nu sa descris încă un ligand
proteina HER 3 (sau Erb 3 )
proteina HER4 (sau Erb4).
Prezintă un domeniu extracelular relativ bine conservat printre membrii acestei
familii, prezentând două domenii bogate în cisteină, cu rol în fixarea ligandului
(figura.2). Domeniul TK intrinsec se repetă o singură dată în structura receptorului, iar
în proteina Erb 3 funcţia TK este scăzută, sau chiar anulată, deşi unii autori susţin că
Erb3 poate fixa ATP la nivelul TK citoplasmatice.
Figura 2: Structura EGFR
o Receptori din clasa a IIa
In această clasă intră: receptorul pentru insulina (IR), receptorul pentru (IGF r R),
receptorul pentru IGFn/Man6P care poate lega IGFn sau liganzi care conţin în anumite
situsuri resturi de manoz6fosfat şi doi receptori ai căror liganzi nu se cunosc încă:
"insulinreceptorrelated receptor" (IRR) şi "insulinIGF 1 hybrid receptor (IR/IGFIR).
Structural aceşti receptori sunt tetrameri, fiind formaţi din două subunităţi a şi
două subunităţi β (vezi cap. I). Subunităţile α au numai domeniu extracelular şi conţin
un segment bogat în resturi de cisteină cu rol în fixarea ligandului. Subunităţile β ale
receptorului sunt transmembranare, segmentul intracitoplasmatic conţine un domeniu
tirozinkinazic. Subunităţile a se leagă de subunităţile β prin legături disulfidice; de
asemenea şi între subunităţile β există legături SS.

Această familie de receptori prezintă o omologie structurală de peste 80% la
nivelul domeniului kinazic pe când, în domeniul extracelular identitatea aminoacizilor
este foarte mică de la un membru la altul al acestei familii
Figura 3: Structura IR
Receptorul pentru insulina Fig.3 este sintetizat sub forma unei singure proteine
(proreceptor) care derivă de la o singură genă localizată pe braţul scrurt al
cromozomului 19. Proreceptorul suferă un proces de proteoliză rezultând
heterotetramerul matur.
Există un grad înalt de omologie între IR şi IGF r R atât la nivelul genelor care îi
codifică cât şi la nivelul organizării structurale. în ciuda acestei omologii, IR este mai
puţin potent în transmisia semnalului mitogen decât IRF r R. Totuşi, dacă în mediu
există cantităţi crescute de IGF n , atunci IR poate contribui la mitogeneză.
(IGFn/Man6P)R este un receptor bifuncţional, care poate lega atât IGFn cât şi
liganzi care conţin manoz6fosfat în anumite situsuri. Acest receptor are un rol
important în traficul enzimelor lizozomale şi în degradarea IGFU, în schimb rolul său
în transducţia semnalului de creştere rămâne controversat.
Din punct de vedere structural este format dintrun lanţ polipeptidic care
străbate o singură dată membrana celulară.

În ultimii ani sa demonstrat existenţa in vivo a unor receptori pentru insulina şi
IGF atipici, capabili să lege unul sau mai mulţi liganzi cu afinitate relativ crescută.
Astfel IRR prezintă o omologie structurală de aproximativ 55% cu IR şi IGF r R. în
schimb domeniul C terminal intracitoplasmatic situat după domeniul tirozinkinazic
este mult mai scurt. Acest receptor nu fixează nici insulina, nici IGFi, IGFn,
proinsulina sau relaxina şi de aceea până în prezent IRR rămâne un receptor orfan,
căreia nu i sa descoperit încă un ligand.
Receptorul IR/IGFi este un receptor hibrid heterodimeric format dintrun "αβ
hemireceptor" provenit de la IR şi un "αβ hemireceptor" provenit de la IGFjR. Acest
receptor hibrid se pare că este prezent în multe ţesuturi, în unele ţesuturi fiind
receptorul exprimat în cantitatea cea mai mare. Totuşi, diferenţele de transmisie a
semnalului intracelular între IGFjR şi IR/IGFiR nau fost încă stabilite.
o Receptorii din clasa a IIIa sunt receptori pentru PDGF, CSF 1 ("colony
stimulating factor") şi SCF ("ster cell factor").
Pentru PDGF se cunosc două tipuri de receptori Ra şi RR fiind proteine cu 170
şi respectiv 180 kDa.
Extracelular, aceşti receptori conţin cinci domenii imunoglobulinlike, iar
intracelular conţin domenii tirozinkinazice. Genele pentru receptorii acestei clase se
găsesc localizate pe cromozomii 4 (pentru R« al PDGF şi SCF) şi 5 (pentru RR al
PDGF şi CSF1). Deoarece izoformele PDGF sunt dimeri, fixarea ligandului pe
receptor determină dimerizarea receptorului (figura.4).
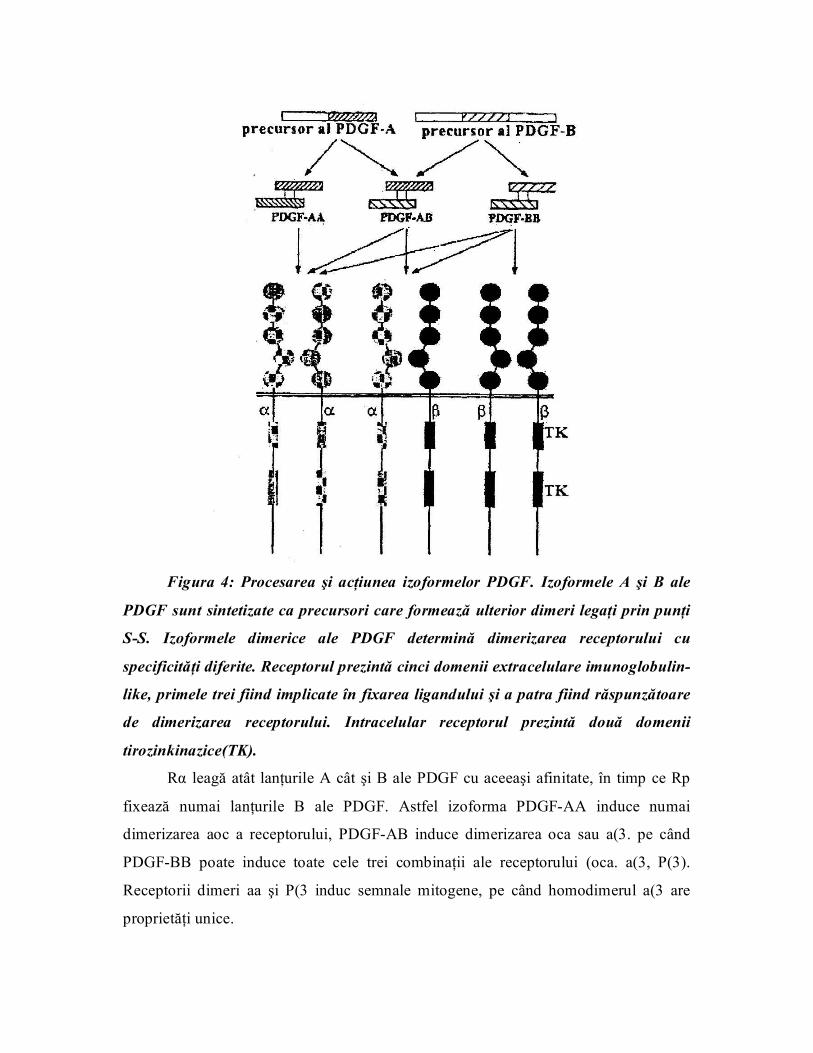
Figura 4: Procesarea şi acţiunea izoformelor PDGF. Izoformele A şi B ale
PDGF sunt sintetizate ca precursori care formează ulterior dimeri legaţi prin punţi
SS. Izoformele dimerice ale PDGF determină dimerizarea receptorului cu
specificităţi diferite. Receptorul prezintă cinci domenii extracelulare imunoglobulin
like, primele trei fiind implicate în fixarea ligandului şi a patra fiind răspunzătoare
de dimerizarea receptorului. Intracelular receptorul prezintă două domenii
tirozinkinazice(TK).
Rα leagă atât lanţurile A cât şi B ale PDGF cu aceeaşi afinitate, în timp ce Rp
fixează numai lanţurile B ale PDGF. Astfel izoforma PDGFAA induce numai
dimerizarea aoc a receptorului, PDGFAB induce dimerizarea oca sau a(3. pe când
PDGFBB poate induce toate cele trei combinaţii ale receptorului (oca. a(3, P(3).
Receptorii dimeri aa şi P(3 induc semnale mitogene, pe când homodimerul a(3 are
proprietăţi unice.

Receptorii din clasa a IVa conţin receptori pentru FGF (până în prezent
prezintă patru membri), care datorită "splicingului" alternativ pot exista în mai multe
variante.
Structural receptorul prezintă trei domenii inumoglobulinlike, iar intracelular
prezintă două domenii tirozinkinazice separate printrun domeniu neenzimatic (figura
5).
Pentru o potenţare a activităţii receptorului, FGF "se prezintă" receptorului după
ce sa fixat de heparansulfat sau heparină. Această prezentare implică crearea unui
complex ternar format din FGFheparan sulfat/heparinăreceptor FGF care determină
stimularea celulei.
Figura 5: Structura FGFreceptor
Receptorii din clasa a Va conţin receptori pentru VEGF, care se prezintă sub
trei forme: VEGF Rl, VEGF R2 şi VEGF R3. Structural cei trei receptori sunt
asemănători, prezentând în domeniul extracelular şapte domenii imunoglobulinlike;
domeniul intracitoplasmatic conţine segmentul tirozinkinazic (figura 6) care este
împărţit în două subdomenii de un segment cu funcţie neenzimatică, de lungime
variabilă.
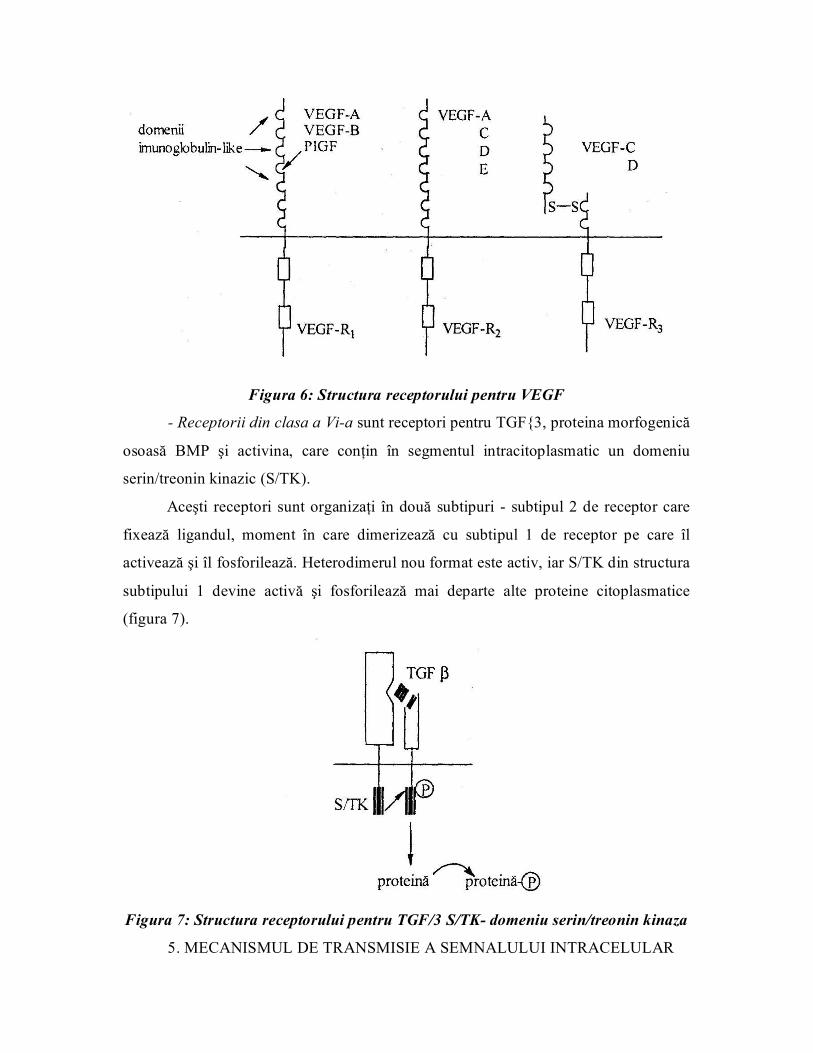
Figura 6: Structura receptorului pentru VEGF
Receptorii din clasa a Via sunt receptori pentru TGF3, proteina morfogenică
osoasă BMP şi activina, care conţin în segmentul intracitoplasmatic un domeniu
serin/treonin kinazic (S/TK).
Aceşti receptori sunt organizaţi în două subtipuri subtipul 2 de receptor care
fixează ligandul, moment în care dimerizează cu subtipul 1 de receptor pe care îl
activează şi îl fosforilează. Heterodimerul nou format este activ, iar S/TK din structura
subtipului 1 devine activă şi fosforilează mai departe alte proteine citoplasmatice
(figura 7).
Figura 7: Structura receptorului pentru TGF/3 S/TK domeniu serin/treonin kinaza
5. MECANISMUL DE TRANSMISIE A SEMNALULUI INTRACELULAR

Receptorii pentru factorii de creştere sunt proteine membranare, care în
momentul în care fixează un factor de creştere îşi modifică conformaţia şi dimerizează,
iar ulterior se activează domeniul kinazic intracitoplasmatic (TPK).
Iniţial sa descoperit că în urma legării factorilor de creştere de receptori,
aceştia formează hemodimeri. Recent sa demonstrat că în cadrul unei familii
receptorii pot forma şi heterodimeri. De exemplu, în familia receptorilor pentru EGF,
receptorul orfan HER2 (ErbE^) joacă un rol central, fiind "partenerul" preferat de
ceilalţi receptori în condiţiile formării unui heterodimer. Mai mult, în cazul HER 3 ,
receptorul HER2 este absolut necesar pentru funcţia sa tirozinkinazică, ştiind că HER3
are o activitate kinazică intrinsecă abolită.
Dimerizarea receptorului pentru PDGF şi TGF(3 a fost prezentată în figura 4,
respectiv figura 7.
In urma dimerizării receptorului are loc activarea domeniului tirozinkinazic
intracitoplasmatic care va determina fosforilarea receptorului: un domeniu tirozin
kinazic de pe un receptor din complex va fosforila celălalt receptor şi invers (figura 8).
Aceşti receptori fosforilaţi sunt activi şi transmit mai departe semnalul mitogen.
Figura 8: Autofosforilarea receptorului
Fc factor de creştere; TKdomeniu tirozin kinazic
Studii mai ample sau făcut asupra mecanismului de transmisie prin intermediul
receptorului pentru EGF fiind primul descris.
5.1. Mecanismul de transmisie intracelular pentru EGF

După fixarea EGF pe receptor, se produc modificări conformaţionale în
structura acestuia, ceea ce permite interacţiunea cu un al doilea receptor legat de
ligandul său. Dimerul rezultat activează domeniile tirozinkinazice citoplasmatice care
iniţiază fosforilarea unor resturi de tirozină din structura receptorului. Aceste
tirozine fosforilate reprezintă situri de recunoaştere pentru proteine care prezintă un
domeniu specific numit SH2. Domeniul SH2 este un fragment conservat de
aproximativ 100 aminoacizi care se poate fixa de resturi de tirozină fosforilată din
mediu.
Observaţii recente au arătat că moleculele care intervin în transducţia
(transmiterea) semnalului conţin diverse alte domenii care mediază interacţiunea dintre
diferite proteine care intervin în căile de semnalizare intracelulară. De exemplu, în
afară de domeniile SH2 şi domeniile PTB sunt implicate în recunoaşterea resturilor de
fosfotirozină. Mai mult, domeniile SH3 recunosc secvenţe bogate de prolină,
domeniile PH recunosc fosfolipide membranare şi domeniile PDZ recunosc carboxilul
terminal al resturilor de valină.
In cazul EGF, un număr crescut de proteine cu fragment SH 2 se pot fixa de
dimerul fosforilat; unele proteine au funcţie enzimatică: src, rasGAP, PLCy
(fosfolipaza y), PI3K (fosfotidilinozitol3kinaza), altele nu sunt enzime dar intervin în
transducţia semnalului mai departe şi se numesc proteine adaptor: Grt>2, Grby, shc,
Nck, Crk etc. Fixarea unei proteine sau a altei proteine de receptorul fosforilat este
dependentă de modalitatea de dimerizare (homo/heterodimerizare).
Odată ataşate prin domeniul SH2 de receptorul fosforilat, proteinele enzime cât
şi proteinele adaptor îşi modifică conformaţia şi transmit mai departe semnalele pe căi
diferite de la o proteină la alta.
Semnalul mitogen poate fi transmis pe mai multe căi, dar principala cale este
aşa numita cale a "MAPkinazelor". Această cale (figura 9) implică mai multe proteine
atât citoplasmatice cât şi nucleare.
Proteinele Grb 2 /SOS. Proteina Grt>2 este o proteină adaptor care conţine un
domeniu SH2 şi trei domenii SH3. Prin domeniul SH 2 se fixează de receptorul
fosforilat, iar de un domeniu SH3 fixează o altă proteină citoplasmatică SOS.

Formarea complexului Grb 2 /SOS determină activarea unei noi proteine numită
ras (p21 ras ). Proteina ras, din punct de vedere structural, este formată dintrun singur
lanţ polipeptidic şi are omologie structurală cu subunitatea a a proteinei trimere G. De
altfel, unii autori susţin că proteinele ras formează o familie de proteine G monomerice,
deoarece au capacitatea de a fixa nucleotide cu guanină; astfel, în repaus ras fixează
GDP, pe când în forma sa activă se găseşte cuplată cu GTP. Schimbul de nucleotide
GDP/GTP pe proteina ras este realizat de proteina SOS. In momentul formării
complexului GriVSOS este activat schimbul de nucleotide şi deci se activează proteina
ras. La fel ca şi subunitatea a a proteinelor G trimere şi proteina ras prezintă activitate
GTPazică, dar această funcţie este mai puţin activă.
Cuplată cu receptorul pentru EGF există o proteină GAP ("GTPase activated
protein") care are rol în activarea funcţiei GTPazice intrinseci a proteinei ras. Proteina
GAP se activează numai în momentul dimerizării şi fosforilării receptorului. Deci
stimularea receptorului pentru EGF intervine în modularea activităţii proteinei ras, atât
în sens stimulator cât şi inhibitor.
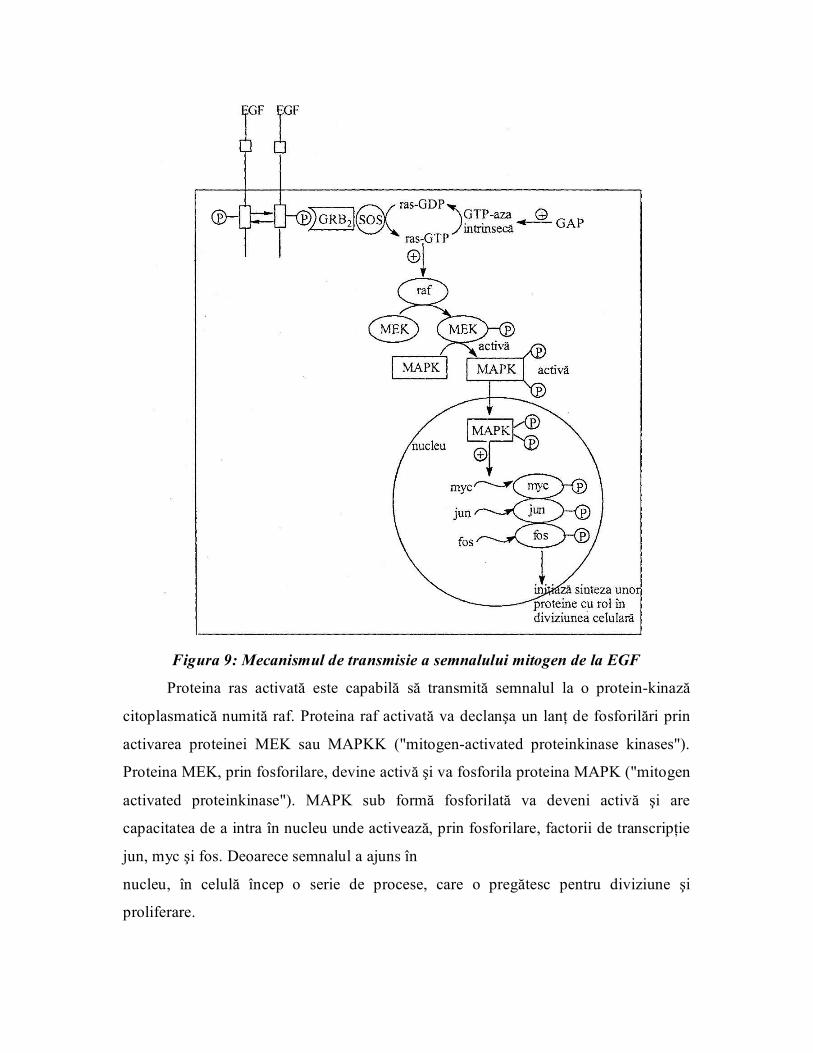
Figura 9: Mecanismul de transmisie a semnalului mitogen de la EGF
Proteina ras activată este capabilă să transmită semnalul la o proteinkinază
citoplasmatică numită raf. Proteina raf activată va declanşa un lanţ de fosforilări prin
activarea proteinei MEK sau MAPKK ("mitogenactivated proteinkinase kinases").
Proteina MEK, prin fosforilare, devine activă şi va fosforila proteina MAPK ("mitogen
activated proteinkinase"). MAPK sub formă fosforilată va deveni activă şi are
capacitatea de a intra în nucleu unde activează, prin fosforilare, factorii de transcripţie
jun, myc şi fos. Deoarece semnalul a ajuns în
nucleu, în celulă încep o serie de procese, care o pregătesc pentru diviziune şi
proliferare.

Etapele descrise reprezintă principala cale activată în momentul stimulării
receptorului pentru EGF de către ligandul său. Dar receptorul fosforilat poate să se
cupleze şi cu alte proteine (enzime sau proteine adaptoare) cu domeniu SH2, astfel
încât sunt şi alte căi modulate de fixarea EGF pe receptor. De exmplu, fosfolipaza C
Yi (PLC Yi) are domeniu SH2 şi se fixează de un rest de fosfotirozină de pe receptorul
pentru EGF. Ulterior PLCYi este fosforilată şi devine activă. Studii in vitro au
demonstrat că PLCYi nu este implicată în transmiterea semnalului mitogen de la
receptorul pentru EGF, dar se pare că intervine în mobilizarea calciului din reticulul
endoplasmatic în celula musculară netedă.
O altă proteinenzimă care este activată de receptorul pentru EGF fosforilat est
fosfatidilinozitol3 kinaza (PI3K). Această enzimă este formată din două subunităţi: o
subunitate reglatorie p85 şi o subunitate catalitică pi 10. Fosforilarea p85 pe un rest de
tirozină determină activarea enzimei care va cataliza fixarea unui fosfat în poziţia 3 a
fosfatidil inozitol 4,5 difosfatului [PI(4,5)P2] cu formarea fosfatidil inozitol 3,4,5
trifosfatului [PI(3,4,5)P3]. Acest compus funcţionează ca un mesager secund fiind
implicat în diferite căi intracelulare de răspuns: creşterea celulară, chemotaxia, efecte
antiapoptotice. Interesant, unii autori au arătat că proteina ras şi PI3K se pot asocia şi
se pot activa una pe cealaltă. Mai mult, formarea heterodimerilor influenţează diferit
timpul de înjumătăţire a MAPK şi deci a semnalului de creştere celulară. De exemplu,
"coexpresia" HER2' cu receptorul pentru EGF sau cu HER3 determină creşterea
afinităţii receptorului pentru EGF şi HER 3 pentru liganzii specifici şi se intensifică
activitatea MAPK, iar timpul de înjumătăţire se alungeşte. Astfel, este generată o largă
"reţea" de interacţiuni ligandreceptor, care în final va determina stimularea mai
multor semnale către nucleu.
5.2. Mecanismul de transmisie intracelular pentru PDGF
Autofosforilarea receptorilor pentru PDGF indusă de dimerizarea lor serveşte la
două funcţii importante. Pe de o parte, fosforilarea anumitor resturi de tirozină din
interiorul domeniului kinazic determină creşterea activităţii acestui domeniu, iar pe de
altă parte, autofosforilarea unor tirozine din afara domeniului kinazic crează situsuri de
recunoaştere pentru moleculele care intervin în transducţia semnalului (molecule cu
domeniul SH2 şi care pot fi enzime sau proteine adaptoare).

Receptorul pentru PDGF, ca şi receptorul pentru EGF, poate fixa enzime (ca
PI3K, PLCY, familia Src de tirozinkinaze, tirozinfosfataza SFfl^ sau proteina GAP
care activează funcţia GTPazică a proteinei ras) sau proteine adaptor (Grf>2, Grb 7 ,
Nck, Shc, Crk) (figura 10). Receptorul pentru PDGF fixează şi membri ai familiei de
proteine Stat. Aceste proteine Stat sunt factori de transcripţie care după fosforilare pe
resturi de tirozină dimerizează şi sunt translocate în nucleu unde influenţează
transcripţia unor gene specifice.
Figura 10 : Transmisia semnalului intracelular de la receptorul pentru PDGF
Astfel, fixarea proteinei adaptor Grb 2 (prin domeniul său SH2) de un rest de
tirozină fosforilată din structura receptorului pentru PDGF determină asocierea
acestuia cu proteina SOS. Complexul Grb 2 /SOS va activa proteina ras datorită
schimbului de nucleotide GDP/GTP realizat sub acţiunea proteinei SOS. Proteina ras
GTP activă va iniţia calea MAPkinazelor (la fel ca la receptorul pentru EGF) care în
final va determina stimularea unor factori de transcripţie nucleari implicaţi în creşterea
celulară.
Fixarea enzimei PI3K pe un rest de tirozină fosforilată din receptor va
determina fosforilarea subunităţii p85 cu activarea subunităţii catalitice pi 10. Această

enzimă joacă un rol central în transmisia semnalelor intracelulare de chemotaxie,
creştere celulară şi are efect antiapoptotic. Astfel PI3K poate interacţiona cu proteina
ras activânduse reciproc. Un alt rol al proteinei PI3K este acela că prin produsul său
de reacţie PI(3,4,5)P3 intervine în "ancorarea" enzimei PLCyîn membrană.
PLCy activată de receptorul fosforilat şi "ancorată" în membrana celulară va
transforma fosfatidilinozitol 4,5 difosfat (PIP2) în diacilglicerol (DAG) şi
inozitol 1,4.5 trifosfat (IP3). DAG activează o serie de proteinkinaze C (PKC), iar IP3
mobilizează Ca 2+ din rezervele intracelulare ale celulei. Totuşi, PLCy nu are rol de
transductor de primă importanţă în stimularea creşterii celulare sau motilităţii celulare;
totuşi, unii autori au arătat că anumite izoforme de PLCy pot influenţa aceste căi.
O altă enzimă care se poate fixa de receptorul pentru PDGF fosforilat este
proteina SHP2. Această proteină, din punct de vedere funcţional, este o tirozin
fosfatază care modulează atât în sens pozitiv cât şi în sens negativ transmisia
intracelualră a mesajului. SHP 2 este o tirozinfosfatază care prezintă două domenii SH2,
prin ambele domenii, enzima fixânduse la două resturi de tirozină fosforilată pentru
ca activarea enzimei să fie maximă. Având capacitatea de a defosforila receptorul
pentru PDGF cât şi substratele proteinkinazelor intrinseci receptorului, SHP2 este un
potenţial modulator negativ al transmisiei semnalului intracelular. Pe de altă parte,
SHP 2 poate fi şi modulator pozitiv al semnalizării intracelulare deoarece poate juca rol
de proteinăadaptor care este recunoscută de grt^/SOS şi care alterior activează
proteina ras.
Se cunosc şi alţi modulatori negativi în calea de semnalizare intracelulară
iniţiată de receptorul pentru PDGF. De exemplu, MAPkinaza, care este activată
printrun mecanism dependent de proteina ras, fosforilează proteina SOS şi o
inactivează (blocând astfel, activarea a noi proteine ras).
Un alt mecanisnm de feedback negativ este dependent de AMP C . Acest
mesager secund sintetizat în celulă în urma stimulării sintezei de prostaglandine şi a
activării ulterioare a adenilat ciclazei, activează proteinkinaza AAMP C dependentă
(PKA). Această serinăkinază fosforilează, şi astfel inactivează, mai multe proteine
implicate în calea de transmisie a semnalului intracelular. Alt mecanism inhibitor,
descris deja la EGF, este activarea proteinei GAP de către receptorul pentru PDGF

fosforilat. Această proteină, prin funcţia sa de activator al GTPazei intrinseci proteinei
ras, intervine în inactivarea acestei proteine.
Se poate spune că receptorul pentru PDGF fosforilat activează simultan atât
proteine implicate în transmisia semnalului intracelular, cât şi în blocarea acestuia,
acest control fiind necesar în buna desfăşurare a activităţii celulare. Orice dezechilibru
în această balanţă transmisia exagerată a semnalului stimulator şi/sau insuficienta
blocare a semnalului intracelular poate determina o creştere celulară exagerată cu
apariţia tumorilor.
5.3. Mecanismul de transmisie intracelular pentru FGF
Până în prezent se cunosc 19 peptide înrudite care formează familia FGF şi care
se pot fixa. cu afinitate mare, pe unul sau mai mulţi receptori pentru FGF. Pentru
activarea maximă a receptorului, ligandul trebuie să fie "prezentat" acestuia de către
heparan sulfat sau heparină. Această "prezentare" poate implica formarea unui
complex ternar receptorFGFheparan sulfat cu rol în dimerizarea receptorului,
activarea tirozinkinazei intrinseci şi autofosforilarea receptorului pe cel puţin şapte
resturi de tirozină.
Căile intracelulare de transmisie a semnalului sunt asemănătoare celor descrise
la EGF sau PDGF. Astfel, receptorul fosforilat poate fixa atât proteine adaptor shc,
FRS2, Crk, care în final vor activa proteina ras sau poate fixa enzime fosfolipaza y
(PLCy) sau src o tirozinkinază citoplasmatică care vor activa, proteina raf (figura
11).
Mecanismele de interacţiune între aceste proteine au fost descrise la EGF şi
PDGF.

Figura 11: Mecanismul de transmisie intracelulară a semnalului indus de FGF
5.4. Mecanismul de transmisie intracelular pentru VEGF
VEGF este implicat în angiogeneza normală cât şi patologică, arătânduse că o
serie de celule tumorale secretă VEGF care va stimula o proliferare anormală a vaselor
sanguine în tumora respectivă.
Din punct de vedere al mecanismului de transmisie intracelular, VEGF se
aseamănă cu FGF. Astfel fixarea VEGF de receptor determină
dimerizarea receptorului şi activarea tirozinkinazei intrinseci. Această enzimă va
fosforila receptorul cât şi proteine care se fixează de receptor deoarece au în
componenţa lor secvenţe SH2. Proteinele care transmit mai departe semnalul pot fi
enzime sau proteine adaptor care deşi nu au funcţie enzimatică pot fixa alte enzime pe
care le activează. Astfel, receptorul pentru VEGF activează un semnal mitogen atât
dependent de calea RAS/MAPkinazelor (mai ales în celulele endoteliale) cât şi un
semnal independent de ras. Acest mecanism este dependent de PLCy care legată de
receptorul fosforilat, se activează şi hidrolizează PIP2 în DAG şi IP3. DAG este
implicat în activarea proteinkinazei C care va activa proteina raf şi în final se activează

MAPkinaza. Se pare că această cale de activare a raf dependentă de PKC este
implicată în migrarea celulei endoteliale.
Şi receptorul pentru VEGF, ca şi receptorul pentru PDGF, activează o
fosfotirozin fosfatază SHP1 cu rol de modulator negativ al transmisiei celulare
(defosforilează proteine implicate în transducerea semnalului).
5.5 Mecanismul de transmisie intracelular pentru IGF
Mai multe date se cunosc despre transmisia intracelulară mediată de receptori
pentru IGFj. IGFi este un peptid sintetizat atât în ficat cât şi în alte ţesuturi, având
puternic efect mitogen.
Spre deosebire de ceilalţi receptori pentru factorii de creşterem care sunt
formaţi dintrun singur lanţ polipeptidic, receptorii pentru IGF sunt dimeri preformaţi.
Fixarea IGFi pe receptor determină o modificare conformaţională a receptorului cu
activarea domeniilor kinazice ale subunităţilor (3 şi autofosforilarea receptorului.
Domeniile kinazice ale receptorului fosforilează şi alte substrate, dintre care proteinele
IRS ('"insulin receptor substrate") şi shc sunt cele mai bine caracterizate (figura 12).
Astfel, proteinele IRS (se cunosc până în prezent 4 izoforme) fosforilate
recunosc şi activează PI3K care va genera fosfotidilinozitol 3,4,5P3 care ulterior va
stimula proliferarea generală şi va inhiba apoptoza.
Efecte proliferative şi antiapoptotice se vor obţine şi prin stimularea căii
ras/MAPkinazelor.
Datele cunoscute din literatură sugerează ipoteza că toate aceste căi de
semnalizare intracelulară interacţionează şi "comunică" una cu alta, existând la nivel
intracelular interacţiuni între receptorii tirozinkinazici şi alte tipuri de receptori (de
exemplu receptorul serpentină, receptorii pentru citokine) sau cu integrine.
MEK

Figura 12: Mecanismul molecular al semnalizării intracelulare
p58 şi pi00 reprezintă unitatea reglatorie respectiv catalitică a fosfatidil
inozitol3 kinaza (PI3K)
PI4,5P2fosfatidil inozitol 4,5 difosfat
PI3A5P3 fosfatidil inozitol 3,4,5 trifosfat
PI3,4P2 fosfatidil 3,4 difosfat
(+) stimulare; () inhibiţie
Şi alte căi de semnalizare intracelulară sunt activate în condiţiile activării
receptorului pentru IGF: de exemplu unele izoforme ale proteinkinazei C (PKC) sunt
reglate de IGF! care ulterior pot stimula calea MAPkinazelor. Ca şi receptorii pentru
PDGF şi receptorii pentru IGF activează o fosfoprotein fosfatază (SHPTP2) implicată
în defosforilarea receptorului şi a unor proteine intracelulare, modulând astfel negativ
aceste căi de transmisie ale semnalului.
6.MECANISME MOLECULARE IMPLICATE IN CREŞTEREA CELULARĂ
Importanţa factorilor de creştere în diviziunea celulară rezultă din faptul că sub
acţiunea acestora, celulele pot depăşi "punctele de control" ale ciclului celular şi trece

întro nouă fază. Fixarea unui factor de creştere pe receptorul său specific induce o
serie de modificări intracelulare care ulterior se transmit în nucleu.
Astfel se ştie că MAPkinaza fosforilată (activă) poate fi translocată în nucleu unde
intervine în fosforilarea unor proteine numite cicline. Aceste cicline se fixează de o
proteinkinază (cdk) pe care o activează. Complexul ciclinăcdk fosforilează o serie de
proteine care intervin în diviziunea celulară. Un substrat cunoscut al complexului cdk
ciclină este complexul format din două proteine: E2FRb. Prin fosforilarea acestui
complex, la nivelul proteinei Rb, E2F este eliberat şi acţionează ca factor de
transcripţie a unor proteine implicate în creşterea celulară (figura 13). Alte substrate
ale complexelor ciclinăcdk pot fi urmărite în tabele I şi II.
Figura 13: Un mecanism intranuclear al acţiunii unui factor de creştere asupra
unei celule; cdk — kinază dependentă de ciclină; (+) stimulare
Sa constatat că există mai multe tipuri de cicline (A, B, D, E) şi de cdk (2, 4, 6).
Pentru a putea depăşi un "punct de control" al ciclului celular, o ciclină se conbină cu o
anumită kinazâ (cdk) (figura 14).

Acest complex funcţionează până când celula reuşeşte să depăşească "punctul
de control"; odată depăşit, ciclina este degradată şi cdk se inactivează
Figura 14: Cuplarea ciclinelor cu kinazele specifice (cdk) în "punctele de control"
ale ciclului celular
Legătura exactă între calea de semnalizare intracitoplasmatică, iniţiată de
receptori pentru factori de creştere şi sinteza ciclinelor, nu se cunoaşte în totalitate, dar
sa observat că aceste proteine (cicline) se sintetizează în celulă numai dacă celula a
fost stimulată de un factor de creştere. Durata de viaţă a ciclinelor este scurtă, acestea
degradânduse după ce celula a trecut de "punctul de control" al ciclului celular.
Tabelul 1 Substrate ale complexului cdkciclină Ia trecerea Gi/S şi
în faza S a ciclului celular
1 Proteine implicate în ADN polimeraze
replicare Ribonucleotid reductaza
2 Proteine implicate în factori de transcripţie (p53, E2F)
transcripţie factori reglatori ai factorilor de transcripţie
(pllORb, pi07)
ARN polimeraza II
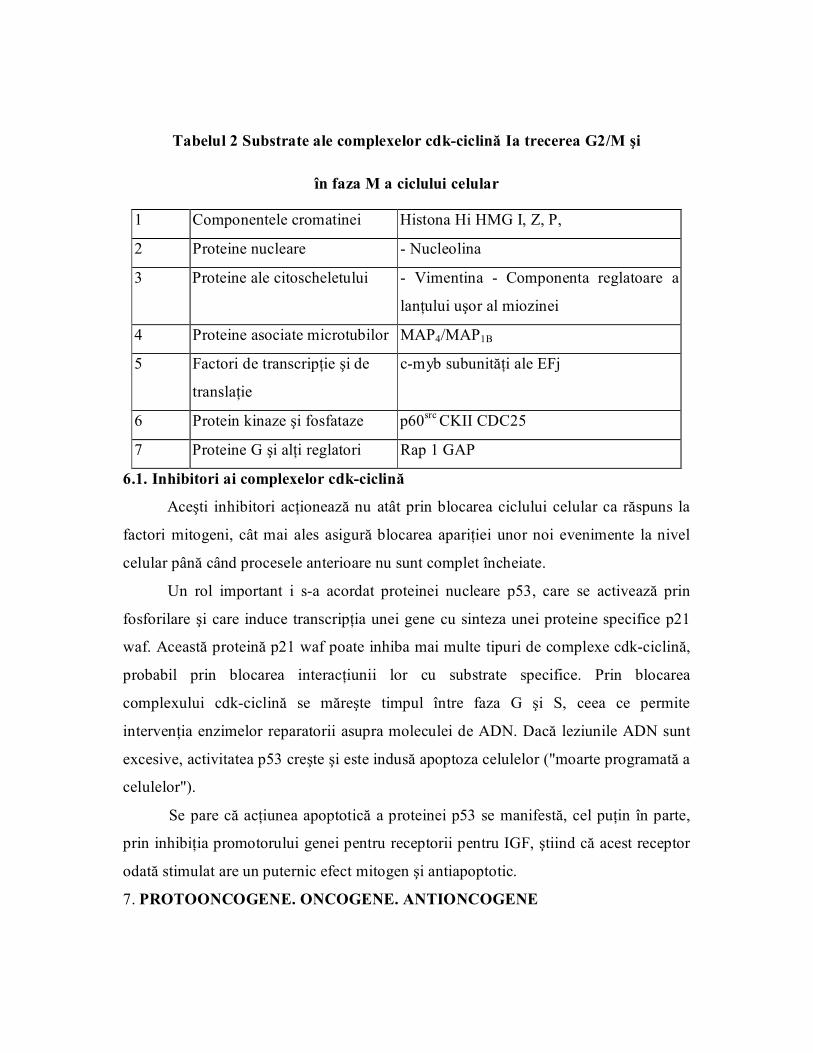
Tabelul 2 Substrate ale complexelor cdkciclină Ia trecerea G2/M şi
în faza M a ciclului celular
1 Componentele cromatinei Histona Hi HMG I, Z, P,
2 Proteine nucleare Nucleolina
3 Proteine ale citoscheletului Vimentina Componenta reglatoare a
lanţului uşor al miozinei
4 Proteine asociate microtubilor MAP4/MAP1B
5 Factori de transcripţie şi de
translaţie
cmyb subunităţi ale EFj
6 Protein kinaze şi fosfataze p60 src CKII CDC25
7 Proteine G şi alţi reglatori Rap 1 GAP
6.1. Inhibitori ai complexelor cdkciclină
Aceşti inhibitori acţionează nu atât prin blocarea ciclului celular ca răspuns la
factori mitogeni, cât mai ales asigură blocarea apariţiei unor noi evenimente la nivel
celular până când procesele anterioare nu sunt complet încheiate.
Un rol important i sa acordat proteinei nucleare p53, care se activează prin
fosforilare şi care induce transcripţia unei gene cu sinteza unei proteine specifice p21
waf. Această proteină p21 waf poate inhiba mai multe tipuri de complexe cdkciclină,
probabil prin blocarea interacţiunii lor cu substrate specifice. Prin blocarea
complexului cdkciclină se măreşte timpul între faza G şi S, ceea ce permite
intervenţia enzimelor reparatorii asupra moleculei de ADN. Dacă leziunile ADN sunt
excesive, activitatea p53 creşte şi este indusă apoptoza celulelor ("moarte programată a
celulelor").
Se pare că acţiunea apoptotică a proteinei p53 se manifestă, cel puţin în parte,
prin inhibiţia promotorului genei pentru receptorii pentru IGF, ştiind că acest receptor
odată stimulat are un puternic efect mitogen şi antiapoptotic.
7. PROTOONCOGENE. ONCOGENE. ANTIONCOGENE
Actual se ştie că proliferarea malignă a celulelor este o maladie genetică
datorită fie activării şi modificării unor protooncogene sau prin pierderea funcţiilor
unor gene supresoare de tumori.
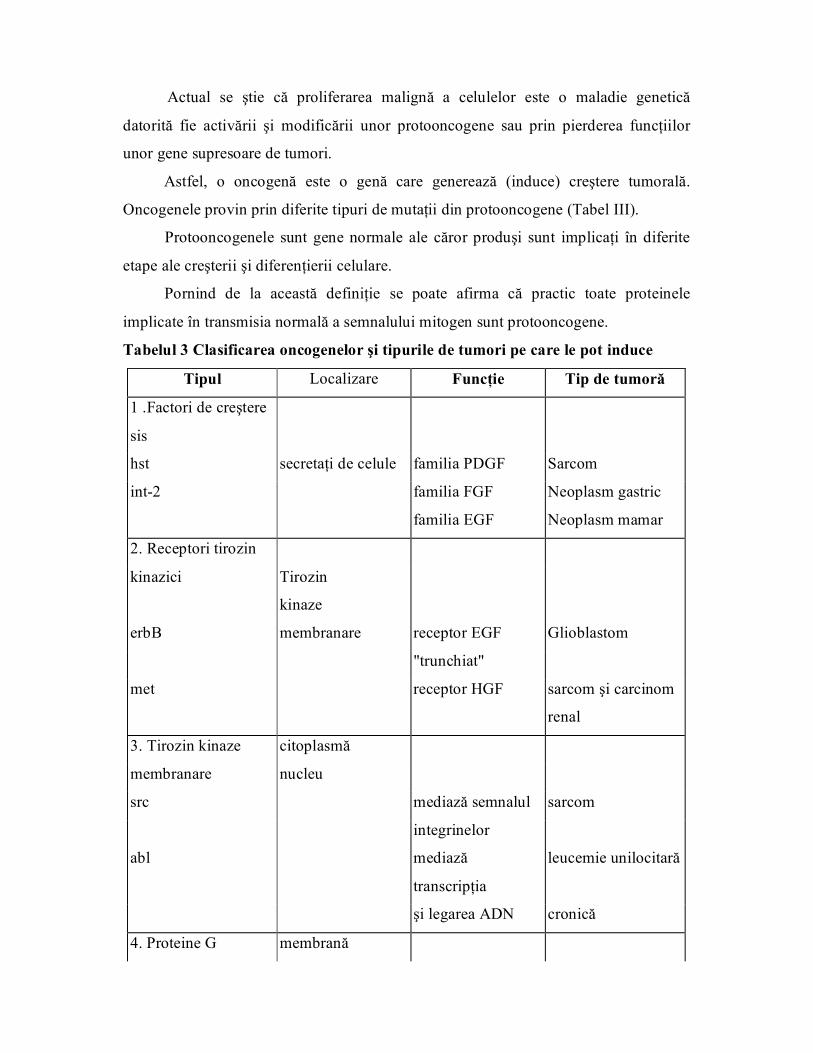
Astfel, o oncogenă este o genă care generează (induce) creştere tumorală.
Oncogenele provin prin diferite tipuri de mutaţii din protooncogene (Tabel III).
Protooncogenele sunt gene normale ale căror produşi sunt implicaţi în diferite
etape ale creşterii şi diferenţierii celulare.
Pornind de la această definiţie se poate afirma că practic toate proteinele
implicate în transmisia normală a semnalului mitogen sunt protooncogene.
Tabelul 3 Clasificarea oncogenelor şi tipurile de tumori pe care le pot induce
Tipul Localizare Funcţie Tip de tumoră
1 .Factori de creştere
sis
hst secretaţi de celule familia PDGF Sarcom
int2 familia FGF Neoplasm gastric
familia EGF Neoplasm mamar
2. Receptori tirozin
kinazici Tirozin
kinaze
erbB membranare receptor EGF Glioblastom
"trunchiat"
met receptor HGF sarcom şi carcinom
renal
3. Tirozin kinaze citoplasmă
membranare nucleu
src mediază semnalul sarcom
integrinelor
abl mediază
transcripţia
leucemie unilocitară
şi legarea ADN cronică
4. Proteine G membrană
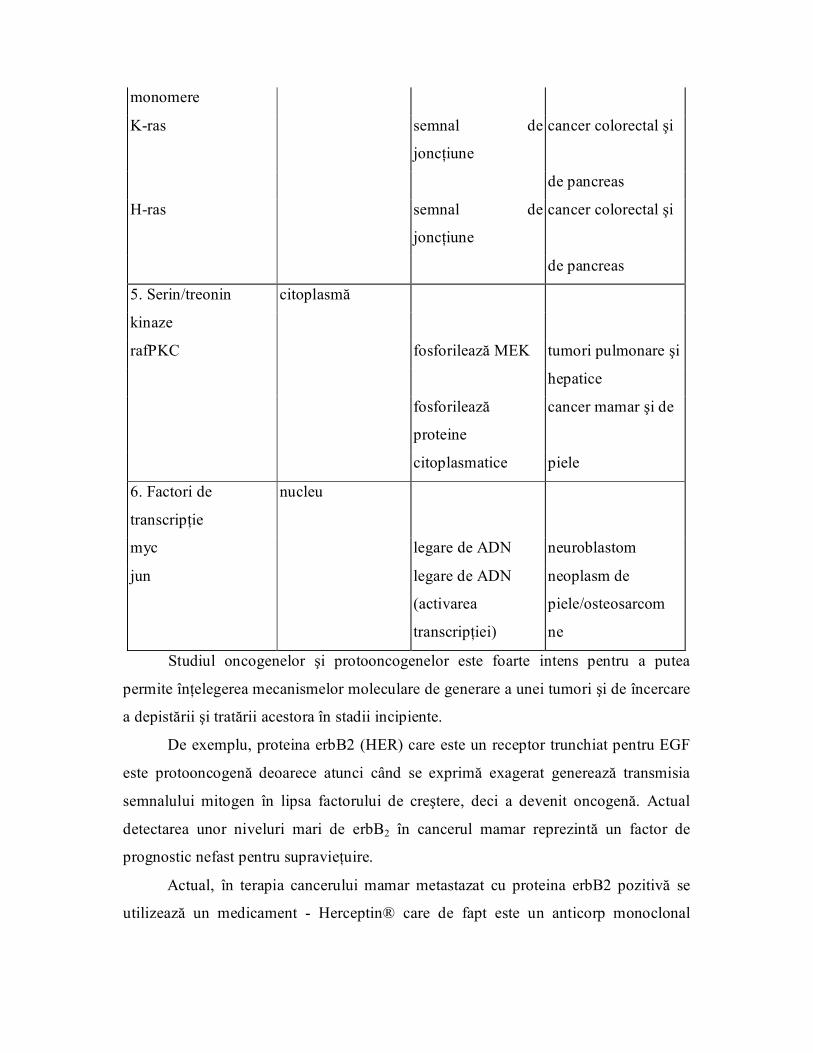
monomere
Kras semnal de
joncţiune
cancer colorectal şi
de pancreas
Hras semnal de
joncţiune
cancer colorectal şi
de pancreas
5. Serin/treonin citoplasmă
kinaze
rafPKC fosforilează MEK tumori pulmonare şi
hepatice
fosforilează
proteine
cancer mamar şi de
citoplasmatice piele
6. Factori de nucleu
transcripţie
myc legare de ADN neuroblastom
jun legare de ADN neoplasm de
(activarea piele/osteosarcom
transcripţiei) ne
Studiul oncogenelor şi protooncogenelor este foarte intens pentru a putea
permite înţelegerea mecanismelor moleculare de generare a unei tumori şi de încercare
a depistării şi tratării acestora în stadii incipiente.
De exemplu, proteina erbB2 (HER) care este un receptor trunchiat pentru EGF
este protooncogenă deoarece atunci când se exprimă exagerat generează transmisia
semnalului mitogen în lipsa factorului de creştere, deci a devenit oncogenă. Actual
detectarea unor niveluri mari de erbB 2 în cancerul mamar reprezintă un factor de
prognostic nefast pentru supravieţuire.
Actual, în terapia cancerului mamar metastazat cu proteina erbB2 pozitivă se
utilizează un medicament Herceptin® care de fapt este un anticorp monoclonal

umanizat care blochează proteina erbB2 de pe celula neoplazică, blocând astfel
multiplicarea acesteia.
Un alt receptor larg răspândit la nivelul diferitelor celule tumorale este
receptorul pentru IGFt datorită efectelor sale proliferative şi antiapoptotice. Din
această cauză se încearcă şi pentru receptorul pentru IGF1 găsirea unor anticorpi
blocanţi ai receptorului sau analogi structurali ai liganzilor care să blocheze receptorul.
O altă direcţie de cercetare în terapia cancerului este inhibiţia angiogenezei
(formarea de noi vase de sânge) în tumori, care este dependentă de VEGF şi FGF. într
adevăr, în multe tipuri de tumori sau găsit exprimaţi VEGFA şi FGF2. Aceşti factori,
prin stimularea formării de noi vase sanguine, permit expansiunea tumorală. Astfel,
noi strategii în terapia cancerului încearcă să utilizeze anticorpi monoclonali care să
blocheze receptorii pentru VEGF şi FGF sau droguri care să inhibe tirozinkinaza
intrinsecă acestor receptori.
în afara protooncogenelor şi oncogenelor, o atenţia deosebită se acordă şi
antioncogenelor gene care în mod normal intervin în inhibarea creşterii celulare.
Dintre antioncogene mai cunoscute sunt pRb şi p53 a căror funcţie a fost descrisă
anterior.
Antioncogenele prin nefucnţionalitate sau prin funcţionare deficitară duc la
creştere tumorală. Astfel, pRb deficitară sa descris în tumori pulmonare cu celule mici,
adenocarcinoame de prostată, tumori ale retinei, tumori osoase etc.
Deficitul de funcţionare a p53 sa descoperit, de asemenea, în multe tumori
hepatocelulare, ale colonului, creierului.
În concluzie, studiul proteinelor implicate în creşterea celulară are o importanţă
majoră deoarece numărul persoanelor noi cu cancer este în creştere iar mortalitatea
prin cancer ocupă locul al IIlea în lume după bolile cardiovasculare.
Mai mult, studiul acestor particule implicate în oncogeneză permite găsirea
unor markeri biologici specifici pentru depistarea precoce a cancerului.
Terapia cancerului în viitor se va adresa atât oncogenelor cât şi genelor
supresoare de tumori. Direcţiile acestei terapii sunt:
1) Terapia prin introducerea în organism a "genei normale" pentru a compensa gena
defectă sau lipsă ("gene therapy") în cazul nefuncţionării antioncogenelor;

2) Terapia prin anticorpi monoclonali "umanizaţi" care să inhibe eficient acţiunea
unei proteine produsă de o oncoproteină;
3) Terapia prin "antisense RNA", care are ca efect blocarea transcripţiei geneiţintă,
deci anularea sintezei produsului oncogenei.
























