Embed Size (px)
Citation preview

J . Fish B i d . (1977) 11,175-184
Bimodal distribution of length of juvenile Atlantic salmon (Salmo salar L.) under artificial rearing conditions
J. E. THORPE Department of Agriculture and Fisheries, Freshwater Fisheries Laboratory,
Faskally, Pitlochry, Perthshire PH16 5LB
(Accepted 20 June 1976)
In 52 populations of juvenile Atlantic Salmon, Salmo sular L., from 14 pairings of wild parents, reared at the Almondbank smolt rearing station between 1973 and 1976 in circular radial-flow tanks, bimodality in the frequency distribution of length and weight has been characteristic by the first autumn of growth, and has become progressively clearer during the following winter.
The examination of 13 populations showed the modes did not correspond to separate sexes, from four others it was shown they did not correspond to immature fish and pre- cociously mature males. It is suggested that the two modes correspond to subpopulations smoltifying at ages ( t ) and ( r+ 1) years. The age-composition of the smolt-run originating from individual year-classes of salmon in four Scottish rivers was found to be consistent with stocks having size-group segregations, at the time of the first smolt emigration, similar to those found among Almondbank experimental material. It is inferred that bimodality of size distribution is the norm among sibling populations of juvenile Atlantic salmon.
I. INTRODUCTION
Since 1973 a series of experiments has been in progress at the Atlantic Salmon Smolt Rearing Unit, Almondbank, to evaluate the effects of a range of environmental variables on growth and survival of juvenile Atlantic Salmon, Salmo salar L. This paper reports on the consistent occurrence of bimodality in the distribution of length within the experimental populations, a feature also apparent in Norwegian experi- mental stocks (Knutsson & Grav, 1975) but not discussed by the authors.
11. MATERIALS AND METHOD§
Mature adult salmon were obtained from the River Almond in late October and early November by electrofishing at weirs during the upstream spawning migration. The growth history and age of these fish was determined from scale material, and selected pairings were made giving a range of crosses between parents of several combinations of freshwater and sea-age. Eggs from these pairings were incubated in separate baskets and the alevins hatching from them transferred to 2 m diameter radial-flow fibreglass tanks (Minaur, 1973) when they had reached an embryo: alevin dry weight ratio of approximately 80% (Marr, 1966). Initial stocking densities of 1500 and 700 individuals per tank were used in the main experiments in 1974 and 1975 under which conditions it was possible to achieve high survival rates during the critical first-feeding stage, and through the remainder of the first summer growing period, averaging 85 and 89.7 % for 16 populations in each year respectively. All of the fish were fed on a commercial dry diet. Growth in length and weight was monitored after 89, 155 and 319 days feeding in 1974 and after 161 and 209 days in 1975. On the first two occasions samples of 70 individuals per tank were used to determine the mean length and weight, on the assumption that such characteristics were normally distributed in these stocks, and from preliminary
175

J. E. THORPE 176
experiments to determine the necessary sample size to estimate these mean values to within 510% of their true value with 95 % confidence (Snedecor, 1950). Subsequently, measure- ments were made on every individual in each population. In the 1975 experiment the 16 tanks were arranged in a 4 x4 latin square design, in which the growth of two families of salmon progeny each reared under two hatchery regimes as alevins, was compared under two flow conditions (50 & 100 l/min.), with and without cover.
111. RESULTS
The sequences of length distributions in Fig. 1, for two populations of salmon from the 1974 experiment, show the development of bimodality in length over the first 10.5 months of feeding. It is also clear that samples of only 70 fish were inadequate to demonstrate the true form of the distributions, which only became apparent in the measurements from the total populations of fish at 319 days. At this stage, in family 8/73 the proportion in the upper mode was 72%, and 63 % in family 16/73. In the 1975 experiment the fish were not sampled until October, at which time every fish in each of the 16 tanks was measured and weighed. Bimodality was evident to a greater or lesser degree in every instance, and the proportions of fish in the modal groups in each population were calculated using arithmetic probability plots (Cassie, 1954). The proportions in the upper mode are shown in Table I, which also represents the spatial relationships (Latin square) of the 16 tanks used in the 1975 experiment. Analysis of variance of the angular transformations of these values (Table 11) shows there is a
-_ 12 Oct 74 I
-__-_ ~
15 n.837 21 Ma* 7 5 1
i;nnily ,o / 73
Fork lengfh (cm)
FIG. 1 . Development of bimodality in length frequency distribution of two groups of sibling juvenile Atlantic Salmon, Salmo saIar L.

B I M O D A L D I S T R I B U T I O N I N J U V E N I L E S A L M O N 177
TABLE I. Percentage of population in the upper mode of length distributions: 1975 experiment
Row 1 94.0 44.0 91.0 50.0 2 80.0 84.0 38.0 78.0 3 78.0 60.0 94.0 76.0 4 50.0 93.5 20.0 74.0
Mean values: family 3/74; 85.8 cover: 60.8 high flow: 75.8 family 2/74: 52.3 no cover: 77.3 low flow: 62.3
TABLE 11. Analysis of variance of angular transformation of data of Table I
1 Sum of Degrees of Mean Source of variation squares freedom squares P
Family Cover Flow Hatchery condition Rows Columns Error Total
1984.26 483.34 318.98 49.28
241.37 144-72 105.83
3327.78
1 1984.26 9.68 t0.001 1 483.34 4.78 <0.01 1 318.98 3.88 <0*02 1 49.28 1.53 Not significant 3 80.46 3.80 Not significant 3 48.24 2.28 Not significant 5 21.17
15
highly significant difference between families in the proportions occurring in the upper modal (rapid growth) group and significant differences also between the two major environmental treatment groups. The overall proportion of family 3/74 in the upper mode was 85.8 % and 52.3 % of family 2/14.
IV. DISCUSSION
The differences in size distribution between families of salmon indicate differences both in growth capacity and in degree of response to some environmental variants. Occurrence of wide variation in growth among individuals in salmonid populations is well-known but not the actual size distributions. Segregation of 2 growth groups among Almondbank stocks could be due to:
(a) sexual differences, (b) precocious maturation of males, (c) genetic effects, (d) environmental effects, (e) genetic x environmental effects, (f) social interactions.
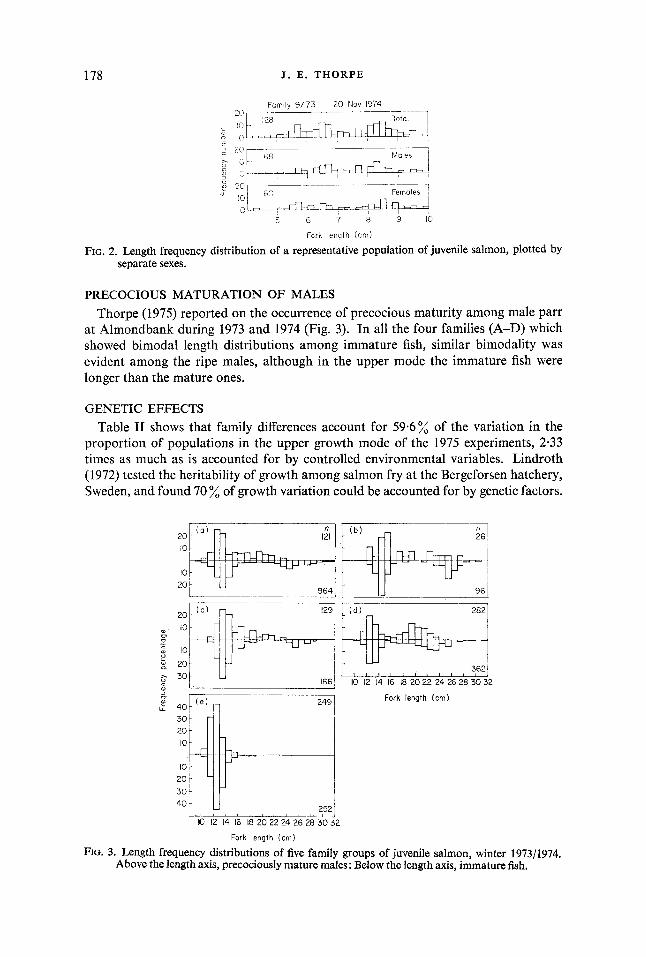
SEXUAL DIFFERENCES 13 bimodal populations were sacrificed in November 1974, and the length distribu-
tions plotted for each sex separately (Fig. 2). In each case both sexes showed similar distributions.
178 J . E . THORPE
Fomiiy 9 /73 20 Nov 19i4
a' U " -- iL 60 Females ,
'",- --- -,- d 5 6 7 8 9 1 0
Fork length (crn)
FIG. 2. Length frequency distribution of a representative population of juvenile salmon, plotted by separate sexes.
PRECOCIOUS MATURATION OF MALES Thorpe (1975) reported on the occurrence of precocious maturity among male parr
at Almondbank during 1973 and 1974 (Fig. 3). In all the four families (A-D) which showed bimodal length distributions among immature fish, similar bimodality was evident among the ripe males, although in the upper mode the immature fish were longer than the mature ones.
GENETIC EFFECTS Table I1 shows that family differences account for 59.6% of the variation in the
proportion of populations in the upper growth mode of the 1975 experiments, 2-33 times as much as is accounted for by controlled environmental variables. Lindroth (1972) tested the heritability of growth among salmon fry at the Bergeforsen hatchery, Sweden, and found 70 % of growth variation could be accounted for by genetic factors.
I FS 20 ID
10 20
Fork length (em)
10 12 14 16 18 20 22 24 26 28 30 32
Fork length (cm)
FIG. 3. Length frequency distributions of five family groups of juvenile salmon, winter 1973/1974. Above the length axis, precociously mature males: Below the length axis, immature fish.

B I M O D A L D I S T R I B U T I O N I N J U V E N I L E S A L M O N 179
However, he used overall mean weight in autumn as his index of size, and standard analysis of variance methods to distinguish sources of variation. If his stocks were in fact bimodally distributed with respect to weight, such methods were not strictly appropriate.
ENVIRONMENTAL EFFECTS Table I1 demonstrates rearing these two families of salmon under contrasted
conditions of overhead cover (none, versus 15 cm wide aluminium rings), and of flow rate, resulted in significantly different distributions of size within populations.
ENVIRONMENTAL x GENETIC EFFECTS When variance was analysed in families 2/74 and 3/74 separately, it was found that
whereas cover had the same significant effect on each, flow rate affected the size distribution within family 2/74 significantly, but not so in family 3/74. Thus in the 1975 experiment the origins of bimodality are not attributable to sexual or maturation differences. The proportions within each mode are affected by both parental origin and tank variables and although it is likely that bimodality is primarily of genetic origin, the numbers achieving relatively high growth-rate are modifiable environ- mentally.
SOCIAL INTERACTIONS It is conceivable that two size groups of fry could arise in the tanks as a result of
social interactions, dominant fish obtaining favourable feeding stations and showing more rapid growth than subordinate individuals, which would be displaced to less favourable stations. The radial-flow tank, as used here, is designed to minimise such social effects, since the radial dispersal of food particles ensures delivery of food to all parts of the tank floor. Indeed hierarchies do not appear to arise and aggressive behaviour is very infrequent. To test the hypothesis that the two modes in the length frequency distribution represented dominant and subordinate subpopulations the stock of one tank (2174 fish), showing an incipient bimodal distribution [Fig. 4(a)], was divided in October so that the large fish were separated from the small ones, and transferred to an identical tank. These two groups were remeasured after 7 months. Jointly they still showed a bimodal length distribution [Fig. 4(b)] : individually they showed skewed unimodal distributions [Fig. 4(c)], and not two bimodals as would have occurred if social factors determined this bimodality. The population densities in these two tanks were 950 and 1050 respectively which is 35-50 % higher than that in the 1975 tank experiments where bimodality occurred. Thus the lack of recurrence of bimodality in this test experiment is not likely to have been due to reduction of population density. Further incidental evidence supporting this finding comes from family 12/71, which was graded mechanically during the winter of 197213 and the fish >9 cm fork-length were tagged and released as smolts in the spring. The remaining small fish were grown on in the tanks for a further year, and when tagged during the winter of 1973/4 their length distribution was clearly unimodal [Fig. 3(e)]. Other families of the same year class, from whose populations no l-year-old smolts had been released, show bimodal distributions of length in the winter of 1973/4 [Fig. 3(a-d)]. The unimodal distribution of family 12/71 corresponds closely to the lower modal groups of (a-d) in Fig. 3.

180 J . E. THORPE
r -1
u 33 E2 37
Fork length (crn)
-5-74
Fork length (crn)
FIG. 4. Length frequency distributions of family 1/72. (a) On 1 October 1973: following measurement the population was graded into Iarge and smalI fish, corresponding to the shaded and un- shaded groups respectively. These subpopulations were reared on in separate identical tanks, and remeasured on 1 May 1974. (b) On 1 May 1974: the frequencies from the two separate subpopulations have been combined, to show the persistence of the bimodal distribution. (c) On 1 May 1974: the separate subpopulations show skewed unimodal length frequency distributions.
However, despite the attempt through tank design to eliminate social regulators of growth it would be unrealistic to assume the dominance relationships have been removed completely. Stress-induced suppression of growth in the smaller groups, as a consequence of increased levels of cortico-steroid hormones reducing appetite, might account in part for the bimodality phenomenon. Blood samples from representatives of each length mode of each of the 16 tank populations in the 1975 experiments, taken in January 1976, have shown higher levels of corticosteroids in fish from the lower size group (Simpson, personal communication). If such higher levels are a consequence of behavioural interactions with large fish, then the removal of those larger fish should remove a cause of growth suppression. In the case illustrated in Fig. 3(e) removal of the larger fish did not result in enhanced growth of the remainder, which remained as small as the lower modal groups of four other populations where the large fish had not been removed. Evidently the conditions are such that social behaviour may ensure the maintenance of distinct growth groups once established, but does not appear to be the primary cause of bimodality of size distribution.
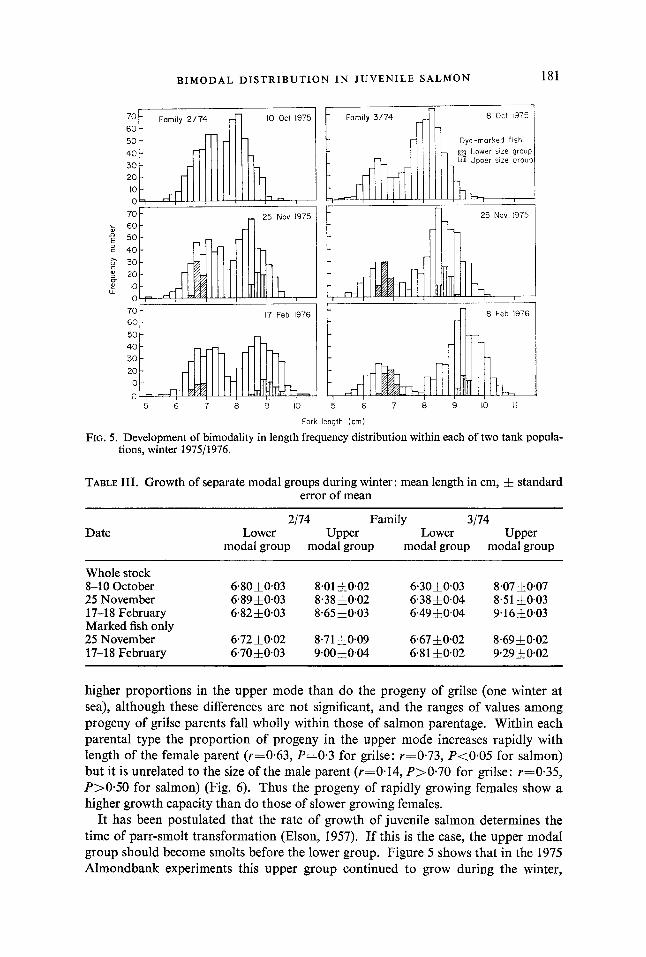
The development of distinct size groups becomes progressively clearer during the winter months. The fish from tanks in rows 1 and 4 of column 4, Table I, were measured again in November 1975, and 50 individuals from the median groups of each modal population were marked with a dyespot. The size distributions of these two tank stocks in October and November 1975, and February 1976, are shown in Fig. 5 , and data on mean lengths are given in Table 111. It is evident that:
(a) marked and unmarked fish grew in the same manner, (b) the slower growth groups grew little between October and February, (c) the faster growth groups have continued to grow during the winter. The discreteness of the marked groups confirms the discreteness of the two modal
groups, and their performance during the winter indicates real differences in growth pattern between them.
The proportion of the total population within a tank which belongs to the upper modal group varied between families (Table IV). When mean values are tabulated by type of parent (Table V) the progeny of salmon (two or more winters at sea) show
181 BIMODAL D I S T R I B U T I O N I N JUVENILE SALMON
Lower slze group Upper size grcup
I t n 25 Nov i975 I
5 6 7 8 9 10 5 6 7 8 9 10 I1
Fork length (c rn )
FIG. 5. Development of bimodality in length frequency distribution within each of two tank popula- tions, winter 197511976.
TABLE 111. Growth of separate modal groups during winter: mean length in cm, j~ standard error of mean
Date
Whole stock 8-10 October 25 November 17-18 February Marked fish only 25 November 17-18 February
2/74 Family 3/74 Lower upper Lower Upper
modal group modal group modal group modal group
6.8010.03 8.01 10.02 6.3030.03 8.0750.07 64910.03 8.3810.02 6.3810.04 8.51 50.03 6.82f0.03 8.653003 6.49 10.04 9.16 50.03
6.7250.02 8.71 f0.09 6.6710.02 8.69f0.02 6.7010.03 9-00f0.04 6.81 f0.02 9.2910-02
higher proportions in the upper mode than do the progeny of grilse (one winter at sea), although these differences are not significant, and the ranges of values among progeny of grilse parents fall wholly within those of salmon parentage. Within each parental type the proportion of progeny in the upper mode increases rapidly with length of the female parent (r=0-63, P=0*3 for grilse: r=0.73, P<0.05 for salmon) but it is unrelated to the size of the male parent (r=0.14, P>0.70 for grilse: r=0-35, P>0.50 for salmon) (Fig. 6). Thus the progeny of rapidly growing females show a higher growth capacity than do those of slower growing females.
It has been postulated that the rate of growth of juvenile salmon determines the time of parr-smolt transformation (Elson, 1957). If this is the case, the upper modal group should become smolts before the lower group. Figure 5 shows that in the 1975 Almondbank experiments this upper group continued to grow during the winter,

182 J . E . T H O R P E
TABLE IV. Proportion of each family in each length group
Parents % in length
group Length Egg diameter no* Age (years) Length Age (years)
River Sea (cm) River Sea (cm) (mm) Upper Lower
13/71 1/72 8 13 16 3/73 5 7 8 9 14 16 2/74 3
2 2 73.0 2 1 67.5 5.33 57 43 2 2 77.5 2 2 71.0 - 54 46 2 2 80.0 2 2 82.0 - 40 60 2 2 90.0 2 2 82.5 - 44 56 2 1 76.5 2 1 69.0 - 55 45 2 2 76.5 2 1 70-0 5.68 60 40 2 1 66.5 3 1 69.0 6.1 1 49 57 2 1 79.5 2 1 67.0 5.76 50 50 2 2 83.5 2 2 88.0 6.60 72 28 2 2 81.0 2 1 69.0 5.86 50 50 2 1 79.5 2 2 86.0 6.27 52 48 2 1 75.0 3 2 87.0 - 63 37 1 2 103.0 2 2 82.0 5.87 52 48 1 2 100.5 2 2 90.0 6.07 86 14
TABLE V. Mean percentage values in upper mode by type of pairing
Total Male parent Salmon Grilse
58.0 57-5 57.9 51.3 53.5 Female parent
Total 57.2 53.8
while the lower group stopped growing. In March 1976 the individuals of this upper group began to smoltify (silver appearance, loss of parr-marks, blackening of fins), while those of the lower group did not.
Since it is not possible to distinguish the progeny of individual pairings among stream populations, such bimodal size distributions have not been reported from the wild. However it is customary to find that emigration of smolts of each year-class takes place in a series of successive springs, rather than all in the same year (Koch, 1968). Data from smolt runs on four Scottish rivers are given in Table VI. In the North Esk, Tummel, and Almond rivers 934-99-7% of the total emigrants from a single year-class pass down the river in two successive springs : in the Girnock Burn 82.2 % of the emigrants move out in a single season’s migration (personal communication Shearer, Struthers and Buck). Whatever mechanism achieves segregation of growth groups within families among young salmon leading to smoltification in two successive springs, would give rise to such a migratory pattern in a population. Buck (per- sonal communication) has estimated annual survival of juvenile salmon between ages 2-3, 3-4, and 4-5, in the Girnock Burn, as 33%. From the data of Table VI, for every 1000 smolts emigrating from one year-class from this system, 99, 822, 76 and 3 left at ages 2, 3 ,4 and 5 respectively. The population at the time immediately before the 2-year-old smolts left was therefore:
N=99+822 (0.33) -l+76 (0*33)-2+3 (0*33)-3 =99+2491+698+83=3371 fish.

B I M O D A L D I S T R I B U T I O N I N J U V E N I L E S A L M O N
80
183
Male parents 0
-
"v I 0 .. $ x E l I
30
40 t 30 I I I I I
70 80 90 , 100 110
Fork length (cm)
FIG. 6. Relationships between the proportion of the population belonging to the upper mode of the progeny length frequency distribution, and the length of the parent fish. x , l-sea-winter fish; 0 2-sea-winter fish.
TABLE VI. Estimates of the proportions by age of those smolts originating from individual year-classes in several Scottish rivers
% of smolts emigrating at age 1 2 3 4 5 (from data of)
Authority River Year-classes
North Esk 1960-1971 5.0 62.8 30.6 1.6 - W. M. Shearer(pers.
Tummel 1965-1970 - 75.4 24.3 0.3 - G. Struthers(pers.
Almond 1968-1970 - 69.7 28.6 1.7 - G. Struthers (pers.
Girnock Burn 1966-1969 - 9.9 82.2 7.6 0.3 R. J. G. Buck (pers. comm.)
comm.)
comm.)
comm.)
Assuming further, the 99 fish are the most advanced individuals of the upper growth mode, then the two modes in this population would be made up of 2590 and 781 individuals respectively, that is 76.8 % in the upper and 23.2 % in the lower group respectively. By a similar argument the stocks of the other rivers would consist of bimodal size groups in proportion as follows (%).
Upper mode Lower mode North Esk 37.4 62.6 Tummel 49.7 50.3 Almond 40.5 59.5
Thus the migratory pattern in these four rivers is consistent with stocks which have size-group segregations similar to those found among Almondbank experimental

J. E . THORPE 184
material. An estimated 94 % of the adult females which enter the Girnock to spawn are two-sea-winter fish and older, ranging in length from 59-84 cm (mean 70.34~0-2 cm). This may account for the high proportion of progeny in the upper mode in the Girnock as compared to the other rivers which have a wider age- and size-range of female spawners.
The bimodal growth pattern among these tank populations of juvenile salmon, and its relationships to smoltification, is thus consistent with the migratory structure of natural populations. It is therefore inferred that bimodality rather than unimodality of size distribution is the norm among sibling populations of juvenile Atlantic Salmon. A recent paper by Chodorowski (1975) has shown the development of bimodal size distributions among juvenile perch (Perca fluviatilis L.), largemouth bass (Micro- pterus salmoides Lackppbde), pike-perch (Lucioperca lucioperca L.) and pike (Esox Zucius L.). However in each case this segregation has been related to difference in diet, the upper growth group consisting of fish which have transferred from a diet of smaller organisms (zooplankton, insect larvae) to one of larger organisms (insect larvae, fish). Such a mechanism cannot account for bimodality in the salmon experiments described here, since all fish were fed the same dry pelleted diet.
Further experiments are now being started with half-sibling groups to test the hypothesis that the expression of bimodality is influenced primarily by the female parent. This influence may be transmitted through egg quality, so that future batches of eggs will be monitored to establish their frequency distribution of dry weight and total fat content, and these distributions related to subsequent growth patterns. Mean egg diameter (Table IV) is not correlated with the subsequent proportion of the population in either size mode.
I am indebted to my colleagues, W. M. Shearer, R. J. G. Buck and G. Struthers, for allow- ing me to use their unpublished data; to Dr T. H. Simpson for the data on corticosteroid hormone levels in the 1975 Almondbank fish; to Dr R. I. G. Morgan, M. S. Miles and their staff at Almondbank for patience during the monitoring of tens of thousands of individual fishes; and to A. V. Holden and B. B. Parrish for their help in the preparation of this paper.
References
Cassie, R. M. (1954). Some uses of probability paper in the analysis of size frequency distributions. Austr. J. Mar. Fw. Res. 5, 513-522.
Chodorowski, A. (1975). Formation de populations bimodales chez les alevins de poissons carnassiers. Verh. Znternat. Verein Limnol. 19, 2546-2555.
Elson, P. F. (1957). The importance of size in the change from parr to smolt in Atlantic Salmon. Can. Fish Cult. 21, 1-6.
Knutsson, S. & Grav, T. (1975). Seawater adaptation in Atlantic Salmon (Salmo salar) at different experimental temperatures and photoperiods. ICES paper CM 1975/M20 25PP.
Koch, H. J. A. (1968). Migration. In Perspectives in Endocrinology (Eds Barrington, E. J. W. & Barker Jorgensen, C.) pp. 305-349. London: Academic Press.
Lindroth, A. (1972). Heritability estimates of growth in fish. Aquilo Ser. Zool. 13,77-80. Marr, D. H. A. (1966). Influence of temperature on the efficiency of growth of salmonid
Minaur, J. (1973). Smolt rearing at Almondbank, Perthsire. J. Znst. Fish. Mgmt 4,65-68. Snedecor, G. W. (1950). Statistical Methods. Ames, Iowa: Iowa State College Press. Thorpe, J. E. (1975). Early maturity in male Atlantic Salmon. Scott. Fisheries Bull. 42,
embryos. Nature, Lond. 212, 957-959.
15-17.


























![Vocabulario bimodal[1]](https://img.dokumen.tips/doc/110x75/55b317ebbb61ebef478b46c4/vocabulario-bimodal1.jpg)


