Embed Size (px)
Citation preview

Benthic Recovery Project
SARF030
Kenny Black, Scottish Association for Marine Science
A REPORT COMMISSIONED BY SARF AND PREPARED BY

Published by the: Scottish Aquaculture Research Forum (SARF) This report is available at: http://www.sarf.org.uk Dissemination Statement This publication may be re-used free of charge in any format or medium. It may only be reused accurately and not in a misleading context. For material must be acknowledged as SARF copyright and use of it must give the title of the source publication. Where third party copyright material has been identified, further use of that material requires permission from the copyright holders concerned. Disclaimer The opinions expressed in this report do not necessarily reflect the views of SARF and SARF is not liable for the accuracy of the information provided or responsible for any use of the content. Suggested Citation Title: Benthic Recovery Project ISBN: 978-1-907266-42-3 First published: January 2012 © SARF 2010

SARF030 Final Report: Benthic Recovery Project
Kenny Black1
Chris Cromey
Thom Nickell1
1Scottish Association for Marine Science
Scottish Marine Institute
Oban, Argyll PA37 1QA
Scotland, UK
June 2011

1
1 Executive Summary
Our aspiration in this project was to provide a computer model able to predict the time of
benthic recovery for a fish farm on the basis of field information and modelling of organic
matter (OM) deposition on the seabed from the farming activity.
To better understand how recovery can be measured, we undertook a field campaign at 5
salmon farm sites on the west coast of Scotland. At these sites we sampled the benthos at a
known interval after the cessation of fish farming. For each of the sites we had access to the
benthic data that were collected by the farmers as part of their monitoring programme and to
husbandry data. We evaluated a standard suite of benthic indices to assess the recovery status
of each of the sites and also developed 2 new indices to aid this analysis: the Brooks
Recovery Index (BRI) and the Infaunal Trophic Recovery Index (IRI).
From our results we concluded that:
The studied sites fell into 2 categories: those that had high initial impacts but recovered
substantially within one year and those that had lower initial impacts but were further from
recovery after 2 years.
These results confirmed earlier work that showed that recovery processes have a site specific
component.
For modelling purposes, we based our approach on DEPOMOD. This model accommodates
the physical environment of the farm (depth, currents, resuspension) from observations and
uses an empirical relationship to predict characteristics (indices) of the benthic community
that would be expected based on the flux of material from the farm at around peak biomass.
The degradation rate of OM from fish farming has not hitherto received much attention. The
need to better understand this topic stimulated a separate degradation project at SAMS. This
work attempts to determine the rate constants for the degradation of fish feed and fish faeces.
This work has led to experimentally derived rate constants and an evaluation of the most
labile fractions of the OM in fish feed and faeces which have been used in the model.
Knowing something about the rate of decay of the OM that accumulates on the seabed, as
predicted by DEPOMOD, we can predict the oxygen demand from the sediment and

2
estimated estimate oxygen supply from the water column using a relationship based on near-
bed current speed from the literature. It is at times where demand exceeds the supply of
oxygen that we might expect the greatest stress on the macrobenthic community. We have
developed an index based on the work of Findlay and Watling (1997) in order to consider the
evolution of the ratio of oxygen demand to supply (Findlay Watling Index, FWI). Although
the FWI was a good predictor of impact during the operational phase of the study sites,
evaluation of this simple implementation of the FWI has not yielded good relationships
between observations and predictions of recovery order at the 5 study sites.
We subsequently developed a separate box model (named the RQ model) which attempts to
account for the role of sulphide formed through anaerobic degradation of OM in fish farm
sediments by including a first order equation for reoxidation of reduced chemical species
together with diffusive losses and burial of metal sulphides. The key hypothesis for this
model arose from the observed differences in the organic matter contents of sediments in the
mid-field i.e at around 50 m from the fish farms. We used this to scale the input of site
specific background OM. The resulting model proved able to account for the main features of
the observed order of recovery of the studied fish farms sites. It is suggested that background
levels of OM in marine sediments may be a good indicator of a site‟s potential to recover after
the cessation of fish farming. Further work will be required to test the validity of this
approach with field data and to refine the model parameterisation.

3
Table of Contents
1 Executive Summary........................................................................................................... 1
2 Introduction ....................................................................................................................... 5
2.1 Recovery of perturbed marine benthic systems ......................................................... 5
2.2 Recovery of benthic impacts from fish farming ........................................................ 7
3 Approach ......................................................................................................................... 12
4 Materials and Methods .................................................................................................... 15
4.1 Recovery study site characteristics .......................................................................... 15
4.2 Biological sampling method .................................................................................... 17
4.3 Chemical parameters ............................................................................................... 17
5 Results ............................................................................................................................. 18
5.1 Emamectin benzoate in sediment ............................................................................ 18
5.2 Organic matter ......................................................................................................... 19
5.3 Metals ...................................................................................................................... 20
6 Benthic biology ............................................................................................................... 22
6.1 Recovery .................................................................................................................. 22
6.2 Changes in benthic function .................................................................................... 28
6.3 Discussion of the benthic data ................................................................................. 33
7 Modelling approach 1 –the Findlay-Watling approach. .................................................. 35
7.1 Modelling approach ................................................................................................. 35
7.2 Method ..................................................................................................................... 36
7.3 Calculation of FWI using the method in Findlay and Watling (1997) and
comparison with observed ITI ............................................................................................. 42
7.3.1 Method ............................................................................................................. 42
7.3.2 Statistical testing of model performance ......................................................... 43
7.3.3 Results ............................................................................................................. 44
7.4 Hydrographic data summary ................................................................................... 44
7.5 Model validation ...................................................................................................... 45
7.5.1 (i) AutoDEPOMOD benthic module – ITI predictions and observations at time
of monitoring survey ....................................................................................................... 45
7.5.2 (ii) Findlay-Watling index (FWI) – suitability as indicator of impact and site
recovery 46
7.5.3 Dynamics of the FWI at the study sites ........................................................... 49
7.6 Discussion of modelling .......................................................................................... 51
8 Modelling approach 2 – the RQ model ........................................................................... 55
8.1 Introduction ............................................................................................................. 55
8.2 RQ model description .............................................................................................. 57

4
8.3 Model processes and parameters ............................................................................. 59
8.4 Results ..................................................................................................................... 66
8.5 Discussion and conclusions ..................................................................................... 73
9 Acknowledgements ......................................................................................................... 74
10 References ................................................................................................................... 75
11 Appendix 1: Summary diversity indices from peak biomass and recovery surveys at
studied sites. ............................................................................................................................ 79

5
2 Introduction
2.1 Recovery of perturbed marine benthic systems
While studies of the impacts of perturbations on marine benthic ecosystems are relatively
common, there has been much less work on processes that occur after the cessation of
perturbation. Such processes may affect the rate at which an impacted benthic habitat moves
towards conditions that are typical of ambient, un-impacted conditions. This process may be
called recovery, but it is important to appreciate that recovery to ambient un-impacted
conditions may not necessarily mean return to pre-impact conditions, as other factors, such as
perturbations caused by other anthropogenic or natural conditions or events, may mean that
such conditions no longer apply. Moreover, given the temporal dynamism and spatial
heterogeneity (or patchiness) of marine benthic ecosystems, which may be caused by natural
stochastic processes (e.g. changes in inter-annual recruitment success which may have
complex effects across food webs affecting community composition and structure) or more
systematic changes such as alterations to climatic regimes, the concept of recovery needs
careful consideration in order to avoid mistaking the consequences of unrelated processes for
the continuing effects of the original perturbation.
Studies on benthic ecosystems that have received attention with respect to recovery, rather
than simply impact, are shown in Table 1 sorted by perturbation type. There are a few lessons
that can be gleaned from this literature. Recovery is relatively fast when the benthic
community already contains species that are adapted to perturbation. For example, although
iceberg scouring can virtually wipe out a benthic assemblage, substantial recovery can take
place in as little as 12 months where the community is frequently assaulted by such
perturbations (Smale et al., 2008). This emphasises that the definition of local recovery may
be problematic where benthic communities face a high degree of natural or other
disturbances. Trawling impacts are likely to be the greatest global source of anthropogenic
disturbance to coastal seabed habitats and impacts may be severe and prolonged for slow
growing species such as sponges and soft corals (Kaiser et al., 2006). Again, the problem in
heavily trawled areas is to determine appropriate reference conditions. Tuck et al. (1998)
overcame this problem by considering impacts of trawling on a fjordic system that had been
un-fished for the previous 25 years. During experimental trawling, benthic biomass and
numbers of species increased but measures of diversity and evenness decreased. Eighteen
months after disturbance, it was still possible to distinguish benthic communities at trawled
and reference sites.

6
The recovery time of a perturbed benthic community depends on the nature, extent and
perhaps the timing of the impact, but some generic environmental and ecological factors are
likely to be play a role in most scenarios. These include: the habitat‟s history of previous
disturbance, the supply of organic matter and oxygen to the sediments, the biogeochemical
status of sediments, any changes caused to the substratum or any biogenic habitat feature, the
presence of any ecotoxic contaminants, the ambient hydrodynamics, and the timing and
availability of invertebrate larval supply. This great range contributes to the uncertainty of
predicting the time to recovery of benthic habitats and presents difficulties in determining the
most important causal factors that drive recovery, as these may vary during the recovery
process.

7
Table 1. Peer reviewed literature on benthic recovery
Perturbation (Author, year)
Aquaculture (Brooks and Mahnken, 2003)
(Brooks et al., 2004)
(Hall-Spencer et al., 2006)
(Holmer and Heilskov, 2008)
(Karakassis et al., 1999)
(Macleod et al., 2004)
(Macleod et al., 2007)
(Macleod et al., 2008)
(Morrisey et al., 2000)
(Pereira et al., 2004)
(Sanz-Lazaro and Marin, 2006)
Coral bleaching (Arthur et al., 2005)
(Arthur et al., 2006)
(Burt et al., 2008)
Dredging and aggregate extraction (Guerra-Garcia et al., 2003)
(Hilton and Hesp, 1996)
(Powilleit et al., 2006)
(Wilber et al., 2007)
Fishing (Hall and Harding, 1997)
(Hall-Spencer and Moore, 2000)
(Hiddink et al., 2006)
(Kaiser et al., 2006)
(Tuck et al., 1998)
Hydrocarbons (Lu and Wu, 2006)
(Readman et al., 1996)
(Zenetos et al., 2004)
Hypoxia (Lim et al., 2006)
Iceberg scour (Gutt et al., 1996)
(Smale et al., 2008)
(Teixido et al., 2004)
Industrial and urban effluents (Borja et al., 2006)
(Hall and Frid, 1995)
(Hall and Frid, 1997)
(Johnson and Frid, 1995)
(Lardicci et al., 1997)
(Lu and Wu, 2007)
(Miller et al., 1999)
(Shin et al., 2008)
(Ueda et al., 1994)
River diversion (Stora et al., 1995)
2.2 Recovery of benthic impacts from fish farming
There have been two previous investigations into the recovery of the benthos after the
cessation of fish farming in the UK. The first, a 3 year study completed in 1995 (Nickell et
al., 1995; Nickell et al., 1998), considered benthic recovery at 3 sites and concluded that a
simple numerical model which could be used to manage rotation of fish farm sites was not
possible from the data obtained, although a descriptive model based on indicator species and
numbers of species appeared to hold broadly for all three sites giving recovery to „normal‟
communities in around two years, even at the most heavily impacted (Table 2). There was no

8
obvious relationship between recovery times and ambient hydrography, and it appeared that
recovery was a complex process which had several drivers that might predominate at different
sites and seasons.
The second study (Pereira et al., 2004) of benthic recovery at a Scottish salmon farm was of a
shorter duration (15 months) and, at the most impacted station, recovery had not been
completed in that time. In contrast to the previous study, organic carbon was not found to be a
significant indicator of recovery, with different environmental variables of varying
importance at different stages in the process. The authors identified sedimentary oxygen
demand as the primary factor determining macrofaunal recolonisation.
Brooks and co-workers in Canada have probably made the most comprehensive series of
studies and observed a very wide range of recovery rates from a few weeks to 6+ years
(Brooks and Mahnken, 2003; Brooks et al., 2004). They proposed two very useful definitions
of recovery:
chemical – “defined as the reduction of accumulated organic matter with a
concomitant decrease in free sediment sulfide and an increase in sediment redox
potential under and adjacent to salmon farms to levels at which more than half the
reference area taxa can recruit and survive (free sulfides < 960 µM)”, and
biological – “defined as the restructuring of the infaunal community to include those
taxa whose individual abundance equals or exceeds 1% of the total invertebrate
abundance at a local reference station. Recruitment of rare species representing < 1%
of the reference abundance was not considered necessary for biological remediation
to be considered complete. As an example, if the mean reference station total
abundance was 8000 macrofauna m-2
, then all of those taxa with a mean abundance of
≥ 80 animals m-2
would be considered necessary for biological remediation to be
considered complete.”
Macleod and co-workers have studied recovery processes at salmon farms in Tasmania over
several years and reached some interesting conclusions:
1) macrobenthic recovery was slower than chemical recovery, so chemical methods
were not sufficient to define ecological recovery (Macleod et al., 2004)

9
2) recovery of macrobenthic community function (from analysis of life history
attributes of dominant fauna) is more rapid than return to community equivalence,
and may be a more useful measure of benthic recovery (Macleod et al., 2008)
3) macrobenthic recovery was faster at a more quiescent site than a more exposed site
attributed to the greater resilience of the species typically found at such sites and
differences in larval supply (Macleod et al., 2007).
In terms of the benthic community both Brook‟s definition of biological recovery and
Macleod and co-workers‟ idea of functional recovery appear attractive avenues for further
research.
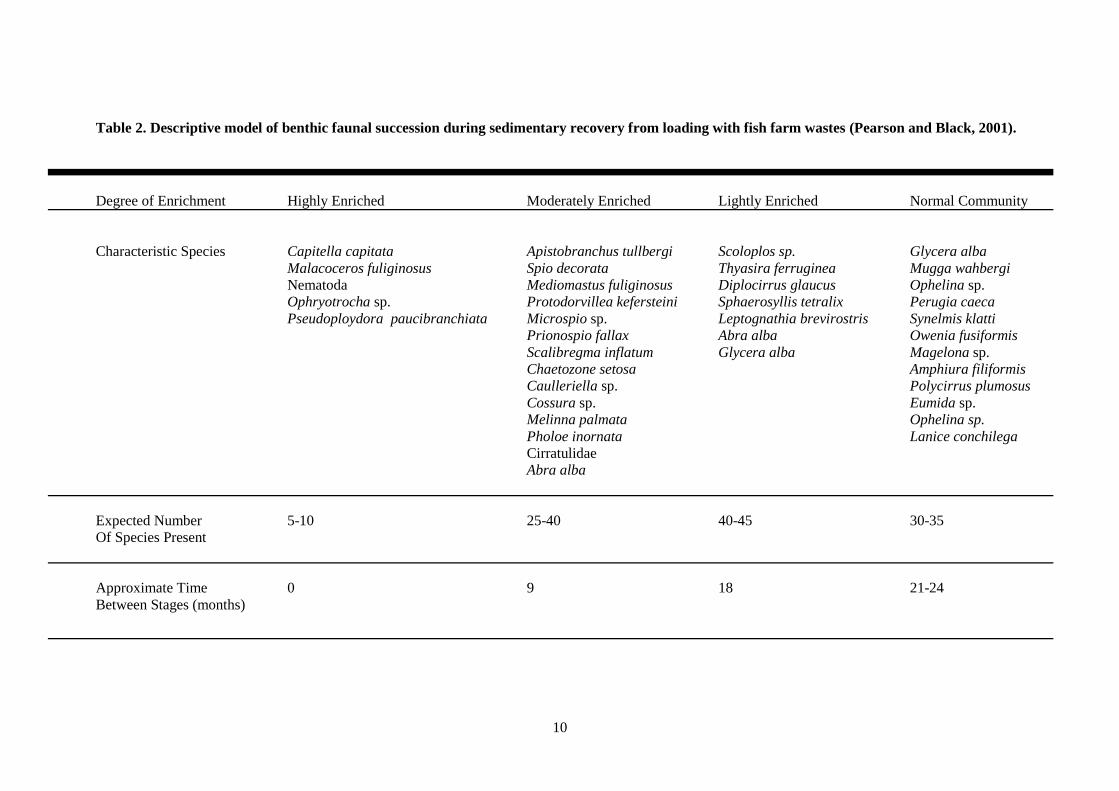
10
Table 2. Descriptive model of benthic faunal succession during sedimentary recovery from loading with fish farm wastes (Pearson and Black, 2001).
Degree of Enrichment Highly Enriched Moderately Enriched Lightly Enriched Normal Community
Characteristic Species Capitella capitata Apistobranchus tullbergi Scoloplos sp. Glycera alba
Malacoceros fuliginosus Spio decorata Thyasira ferruginea Mugga wahbergi
Nematoda Mediomastus fuliginosus Diplocirrus glaucus Ophelina sp.
Ophryotrocha sp. Protodorvillea kefersteini Sphaerosyllis tetralix Perugia caeca
Pseudoploydora paucibranchiata Microspio sp. Leptognathia brevirostris Synelmis klatti
Prionospio fallax Abra alba Owenia fusiformis
Scalibregma inflatum Glycera alba Magelona sp.
Chaetozone setosa Amphiura filiformis
Caulleriella sp. Polycirrus plumosus
Cossura sp. Eumida sp.
Melinna palmata Ophelina sp.
Pholoe inornata Lanice conchilega
Cirratulidae
Abra alba
Expected Number 5-10 25-40 40-45 30-35
Of Species Present
Approximate Time 0 9 18 21-24
Between Stages (months)

11
Karrakassis and co-workers, studying a farm in Greece (Karakassis et al., 1999) demonstrated
that recovery processess do not necessarily follow the linear model proposed in Table 2 as
seasonal effects can result in reversals owing to increased primary production stimulated by
sediment nutrient release. This non-linear pattern of recovery was also noted by Pereira et al.
(2004) (Figure 1).
Figure 1. Multi-dimensional scaling (MDS) plot of square-root transformed species-
abundance mean data at 3 recovering fish farm stations (A – least impacted, B, C – most
impacted) with respect to months post-cessation of farming (1, 2, 8, 14 and 15 months),
showing non-linearity of benthic community response (Pereira et al., 2004).
Since the earlier Scottish studies, salmon aquaculture in Scotland has changed significantly:
cages are bigger, average farm size has increased, more exposed sites have been developed
and the in-feed medicine Slice (active ingredient emamectin benzoate) has become widely
used. Although a recent study did not find a relationship between Slice in sediments and
community changes at active sites (Black et al., 2005) its potential to retard recovery has not
been studied. Copper is also widely used as an antifoulant and has been detected at very high
concentrations in fish farm sediments in Loch Craignish (Dean et al., 2007). Brooks and co-
workers argue that copper in enriched sediments is likely to be bound as sulphides and
therefore not bioavailable (Brooks and Mahnken, 2003; Brooks et al., 2004) but it has not
been shown whether recovering sediments (experiencing increased oxidation state) release
this copper back into pore waters with the potential to affect recolonisation.
More recent approaches to modelling inputs to the sea bed from cage farming have yielded an
improved understanding of effects on the macrofaunal community. The DEPOMOD model
has a benthic component (Cromey et al., 2002a; Cromey et al., 2002b) which at present
predicts likely benthic impact from modelled carbon flux to the sea bed. Morrisey et al.

12
(2000) had some success in predicting remineralisation of carbon/recovery rates in New
Zealand when using the Findlay-Watling oxygen supply model (Findlay and Watling, 1997);
they also noted the potential for increased recovery times due to the presence of heavy metals
in the sediment. Findlay and Watling‟s oxygen supply and demand model (Findlay and
Watling, 1997) was implimented in a non-dynamical mode with calculations of oxygen
supply based on short current records where the only data required are those relating to the 2-
hour period with minimum flow. Their hypothesis was that the benthic community is
structured by the balance between oxygen supply and demand at times of minimum oxygen
supply only. This is intuitively an attractive proposition, but their study had a number of
issues: 1) measurements of low-flow are highly sensitive to the threshold of the current meter
in very low flows; 2) their measurements of oxygen demand from the sediment were based on
measurments of organic matter flux using sediment traps, a technique prone to error owing to
the very steep contours of organic matter flux density around fish cages; 3) they did not take
into account resuspension processes that may remove carbon deposited on the bed before it
has time to cause an oxygen demand; and 4) they had only one study site (data point) at the
high end of the range of impacts typical of salmon farms. However, both the simplicity of
their model and the subsequent work by Morrisey et al. (2000) give strong grounds for
pursuing their approach as a basis for attempting to interpret and model benthic recovery
process around salmon farms.
3 Approach
Our aspiration in this project is to provide a computer model able to predict the time of
recovery for a site on the basis of information on how much organic material had
accumulated on the seabed from the farming activity and from simple environmental
conditions e.g. depth and current regime.
To better understand how recovery can be measured, we undertook a field campaign at 5
salmon farm sites on the west coast of Scotland (Section 5). At these sites we sampled the
benthos at a known interval after the cessation of fish farming. We sampled each site in the
autumn in order to reduce the variability in the results that might be associated with seasonal
effects, for instance caused by temperature, availability of larvae or timing of organic inputs
from natural primary production. For each of the sites we had access to the benthic data that
were collected by the farmers as part of their monitoring programme (Section 6). In all cases
this was collected in the second year of the farming cycle near peak biomass. In addition we
had access to the quantities of feed used by the farmers throughout the production cycle.

13
For modelling purposes, we based our approach on DEPOMOD. This model accommodates
the physical environment of the farm (depth, currents, resuspension) from observations and
uses an empirical relationship to predict characteristics (indices) of the benthic community
that would be expected based on the flux of material from the farm at around peak biomass.
However, the DEPOMOD programme has no validated temporal component. The link
between accumulation of organic matter (carbon in reduced form) to the bed and the benthos
is purely empirical with no consideration of the biogeochemical processes that underlie this
link (Section 8). These processes depend on the degradation of organic matter facilitated by
microbes which consumes oxygen and, when anaerobic, sulphate ion. Both the consumption
of oxygen and the generation of hydrogen sulphide from sulphate reduction generate
conditions that become increasingly hostile to benthic life forms. The rates at which oxygen is
consumed and sulphide is produced are dependent on several factors, but the dominant
processes are the degradation rate of organic mater under both aerobic and anaerobic
conditions (releasing sulphide) and the supply of oxygen from the water column (which itself
may be influenced by the irrigation of the sediments through animal burrows if these have not
been excluded).
For most organic matter, the rate at which it is oxidised does not depend strongly on whether
this takes place in the presence of oxygen or not although rate constants for aerobic oxidation
are generally higher than for anaerobic oxidation. However, for the organic matter that is very
recalcitrant, degradation is extremely slowly without oxygen (e.g. woody material is very
difficult for microbes to degrade without oxygen). Thus, only the least degradable carbon gets
permanently buried in anaerobic marine sediments, with the majority being degraded before it
can be buried.
The degradation rate of organic material from fish farming has not hitherto received much
attention. The need to better understand this topic has stimulated a separate degradation
project supervised by Prof. Ronnie Glud and Dr Henrick Stahl at SAMS and conducted by
Karl Attard, a student at SAMS (presented as an Annex to the present report, available from
SAMS on request). This work attempts to determine the rate constants for the degradation of
fish feed and fish faeces. In order to do this, we recognise that organic matter is a broad term
covering a wide range of organic components each of which will have distinct degradation
rates and be present in distinct amounts. By experimentally monitoring decay processes, we
have determined the rate constants and the amounts of different classes of material that can
used to approximate the decay characteristics of these materials in the marine system and use
this information in our models.

14
We followed 2 modelling approaches. The first used information on the rate of decay of the
material that accumulates on the seabed, as predicted by DEPOMOD, to predict the oxygen
demand from the sediment. But in order to be able to predict the impact of this oxygen
demand, we also estimated oxygen supply from the water column (Section 8). It is at times
where demand exceeds the supply of oxygen that we might expect the greatest stress on the
macrobenthic community. We have used the work of others in estimating the supply of
oxygen as a function of current speed at the farms. Very low near-bed currents will reduce the
supply of oxygen to the benthic boundary layer where transfer of oxygen becomes dependent
on molecular rather than turbulent processes. This approach did not produce a satisfactory
outcome for reasons that will be explained later.
The second approach involved the construction of a model in which the process of
reoxidation of sulphides produced during anaerobic oxidation of organic matter was included,
together with information on likely background levels of organic matter supply and this
approach provided some greater insights into factors that may be important in recovery rates.
Our first target was to determine whether we can correctly predict the order of recovery at the
sites sampled and the second target was to be able to predict the absolute time that a site night
take to reach recovery, using the study sites for calibrating the model.
15
4 Materials and Methods
4.1 Recovery study site characteristics
We have used 5 salmon farm sites to produce data that can be used for the project‟s purposes
and we have called the recovery sites in this report. The recovery sites had varying consented
biomasses and fallowing dates. These are presented in Table 3.
Table 3. Maximum biomass, maximum feed input rate and consented biomass.
Site Max biomass Max Feed input
Date Amount (t) Date Amount
(t month-1)
Consent
biomass (t)
Oban Bay May-07 494 Mar-07 84 500
Fishnish B Aug-06 1201 Jun-06 254 1300
Camasdoun Jan-06 1296 Sep-05 349 1300
Kingairloch Sep-06, May-07 1006 Sep-06 301 1000
Maol Ban Apr-06 826 Dec-05 120 1000
Dates of statutory (peak biomass) surveys and recovery surveys for the sites are presented in
Table 4.
Table 4. Dates of benthic sampling, removal of fish, recovery sampling and months of
recovery between removal of fish and recovery sampling.
Site Monitoring
Sampling
Fish removed Recovery
sampling
Months of
Recovery
Oban Bay July 2007 November 2007 September 2008 10
Fishnish B June 2006 November 2006 October 2007 11
Camasdoun April 2006 August 2006 August 2008 24
Kingairloch June 2007 January 2008 October 2008 9
Maol Ban April 2006 November 2006 November 2008 25
Slice had been used at each of the sites during the farming period prior to this study.
Concentrations of slice in the sediments as determined during Slice consent monitoring are
given in Table 5.
Current records used were those collected by the farmer and are those used by the farmers in
their discharge consents. Details of the instruments are given in Table 6. The ADCP and
electromagnetic types are sensitive to very low current speeds. The impeller types, as used at
Oban Bay and Fishnish B, are much less sensitive to low current speeds and can produce
artificially high numbers of zero current records.
Organic matter concentrations for each of the sites taken during the monitoring survey are
given in Table 7.
16
Table 5. Concentrations of Slice in sediments after the last treatment post cessation of
farming for each site (data courtesy of SEPA). ND = not detectable.
Station Site/Amount emamectin µg kg-1 sediment
Oban Bay Fishnish B Camasdoun Kingairloch Maol Ban
CE 1 3.24 3.42 5.72 4.07 ND
CE 2 7.83 4.07 6.33 4.05 1.07
CE 3 5.99 1.71 5.48 3.63 ND
25m 1 1.34 1.31
25m 2 1.22 ND
25m 3 2.07 1.19
100m 1 ND ND
100m 2 ND ND
100m 3 ND ND
150m 1 ND ND ND
150m 2 ND ND ND
150m 3 ND ND ND
End treatment 26/02/2007 16/02/2006 16/02/2006 13/03/2007 06/09/05
Survey 26/06/2007 05/06/2006 14/06/2006 05/06/2007 03/02/06
Table 6. Date of current records and type of meters used.
Site Date Meter Type
Kingairloch Dec-03 Aanderaa RDCP6001 ADCP
Maol Ban Nov-05 Teledyne RDI2 ADCP
Oban Bay Dec-09 Valeport BFM105D3 Impeller
Camasdoun Nov-07 Valeport Midas808-4004 Electromagnetic
Fishnish B Dec-08 Valeport BFM105D Impeller
Table 7. Organic matter from the monitoring reports for each site.
Site Method Station (0) % Station (25) % Station (50) % Ref1 % Ref2 %
Kingairloch OCa 0mW 3.8 25mW 2.4 50mW 1.3 REF1 0.9
Fishnish B LOI NW 0 17.7 NW 25 m 16.2 65 NW 2.8 Ref1B 2.1
Oban Bay LOI 0 NE 13.1 25 NE 12.0 50 NE 10.1 Ref 13.5 Ref2 10.3
Camasdoun LOI E0 22.9 25E 10.0 E50 4.8 Ref1 3.7 Ref2 8.8
Maol Ban OCa SW0 3.1 SW25 2.0 SW50 2.0 Ref1 3.1
a – For Maol Ban and Kingairloch, OM was reported as organic carbon. This has been converted here
to organic matter using the equation given in Craft et al. (1991), viz. Organic C = 0.40 [LOI] + 0.0025
[LOI]2
1 Aanderaa RDCP600: range 0 – 500 cm s-1; horizontal accuracy 0.5 cm s-1, vertical accuracy 1.0 cm s-1 2 RDI Workhorse Sentinel: range 0.1- 2000 cm s-1; accuracy 0.3% of the water velocity relative to the ADCP ±0.3
cm s-1
3 Valeport BFM105D: range 0.03 - 5.0 m s-1; accuracy <0.15 m s-1,±0.004 m/s ,>0.15 m s-1, ±1.5% reading,
resolution 0.001 m s-1 4 Valeport Midas 808-400: range 0 – 5 m s-1 ; accuracy ±1% reading; resolution 0.001 m s-1

17
Unfortunately, the very low levels of organic matter in sediments at Maol Ban and
Kingairloch (Table 7) are, based on our experience of sea loch sediments, not credible for an
active fish farm site in a relatively depositional environment.
Each of the farms used copper treated nets to reduce fouling. The dates of installation of
treated nets at the study sites are given in Table 8.
Table 8. Installation of nets with copper based anti-foulants.
Site Date of net installation Months to analysis during
recovery sampling
Kingairloch No copper treatment used N/A
Maol Ban No copper treatment used N/A
Oban Bay Dec 2006 21
Camasdoun 2005 >56
Fishnish B Dec 2005 22
4.2 Biological sampling method
Sediment was collected via triplicate 0.1 m-2
van Veen grabs, sieved on board over 1 mm
round mesh, and preserved in 10% buffered formosaline. Faunal identification to lowest
possible taxon was performed by Stirling Institute of Aquaculture. Biological data were
imported into Primer v6 (Clarke and Gorley, 2006) to compute diversity indices. AMBI
values were calculated using the AZTI Marine Biotic Index (AZTI-Tecnalia, www.azti.es)
v4.1. Infaunal Trophic Index (ITI) calculations were performed in Excel using SEPA 2004
corrections (available from SEPA on request).
4.3 Chemical parameters
All sediment chemical parameters were determined on surface to 4 cm depth triplicate
samples collected by sub-cores from grab. To determine organic matter content of sediment,
Loss on Ignition (LOI) was performed by measuring weight loss of freeze dried sediment
after combustion at 250 and 500 ˚C (Loh et al., 2002). Metal analysis was performed at
SAMS by ICP-MS. Metals included lithium, chromium, cobalt, nickel, zinc, copper,
cadmium, lead and uranium. Sediment emamectin benzoate concentrations were measured by
ERI Thurso, after the method detailed by Kim-Kang et al. (2001).
Data were first assessed for normality using the Shapiro-Wilks test as a macro in Excel;
appropriate transformations were applied to achieve normality before parametric analyses
were performed in Excel.

18
5 Results
5.1 Emamectin benzoate in sediment
Sediment Slice concentrations (active ingredient emamectin benzoate) concentrations were
measured on frozen samples by ERI Thurso, and the results are presented in Table 9. Note
that the near-field trigger concentration for emamectin benzoate in sediment is 7.63 µg kg-1
wet wt sed., and the far-field limit is 0.76 µg kg-1
wet wt sed. (SEPA, 1999).
Table 9. Emamectin benzoate (EB) concentrations in sediment (0-4 cm depth) at
recovery sites, by replicate. ND = not detectable; SD = standard deviation. Means
calculated on assumption that ND=0.
Site Sample Sample Date EB (ng g-1) Mean ±SD
Camasdoun E0 1 14/08/2008 1.19 1.89 0.87
E0 2 2.87
E0 3 1.62
E50 1 ND 0
E50 2 ND
E50 3 ND
Ref 1 1 ND 0
Ref 1 2 ND
Ref 1 3 ND
Oban Bay NE0 1 16/09/2008 1.39 0.91 0.79
NE0 2 1.35
NE0 3 ND
NE25 1 1.06 0.35 0.61
NE25 2 ND
NE25 3 ND
NE50 1 ND 0
NE50 2 ND
NE50 3 ND
Ref 1 ND 0
Ref 3 ND
Kingairloch Ref 1 09/10/2008 ND 0
Ref 2 ND
Ref 3 ND
W0 1 0.8 0.27 0.46
W0 2 ND
W0 3 ND
W25 1 1.19 0.76 0.66
W25 2 1.08
W25 3 ND
W50 1 ND 0
W50 2 ND
W50 3 ND
Maol Ban W0 1 13/11/2008 ND 0.52 0.91
W0 2 ND
W0 3 1.57
W25 1 ND 0
W25 2 ND
W25 3 ND
W50 1 ND 0
W50 2 ND
W50 3 ND
Fishnish B NW0 1 24/10/2007 ND 0
NW0 2 ND
NW0 3 ND
19
Examination of the concentrations of Slice at the cage edge stations at each site after the last
treatment before cessation of farming and from the post-cessation sampling for this project
(Table 10) shows that at most sites there is a decrease over the recovery period. Slice was not
detected at the cage edge stations at Fishnish B and Kingairloch despite these having been
fallowed for less than one year. The Slice concentration at Camasdoun is also much less than
during the production cycle but is still considerably above the detection limit after 24 months
of recovery. Oban Bay shows a greater reduction but this has been achieved in only 10
months. Maol Ban is exceptional in that Slice concentration is approximately unchanged, but
still low, after 25 months.
Table 10. Sediments concentrations of Slice before and after cessation of farming
Before cessation After cessation Months
since
cessation
Mean Stdev Mean Stdev
Oban Bay 5.69 2.31 0.91 0.79 10
Fishnish B 3.07 1.22 0 11
Camasdoun 5.84 0.44 1.89 0.87 24
Kingairloch 3.92 0.25 0 9
Maol Ban 0.36 0.62 0.52 0.91 25
Attempting to rank these sites in terms of their rate of removal of Slice is problematic but is
attempted thus:
Kingairloch ≈ FishnishB > Oban Bay > Camasdoun > Maol Ban
5.2 Organic matter
Organic matter from waste fish food and faeces contributes to the total organic load on the
benthic environment. To quantify this load, sediment samples were assessed for Total (TOM),
Refractory (ROM) and Labile (LOM) components by Loss On Ignition. Results from the
recovery study sites are presented in Table 11 below.
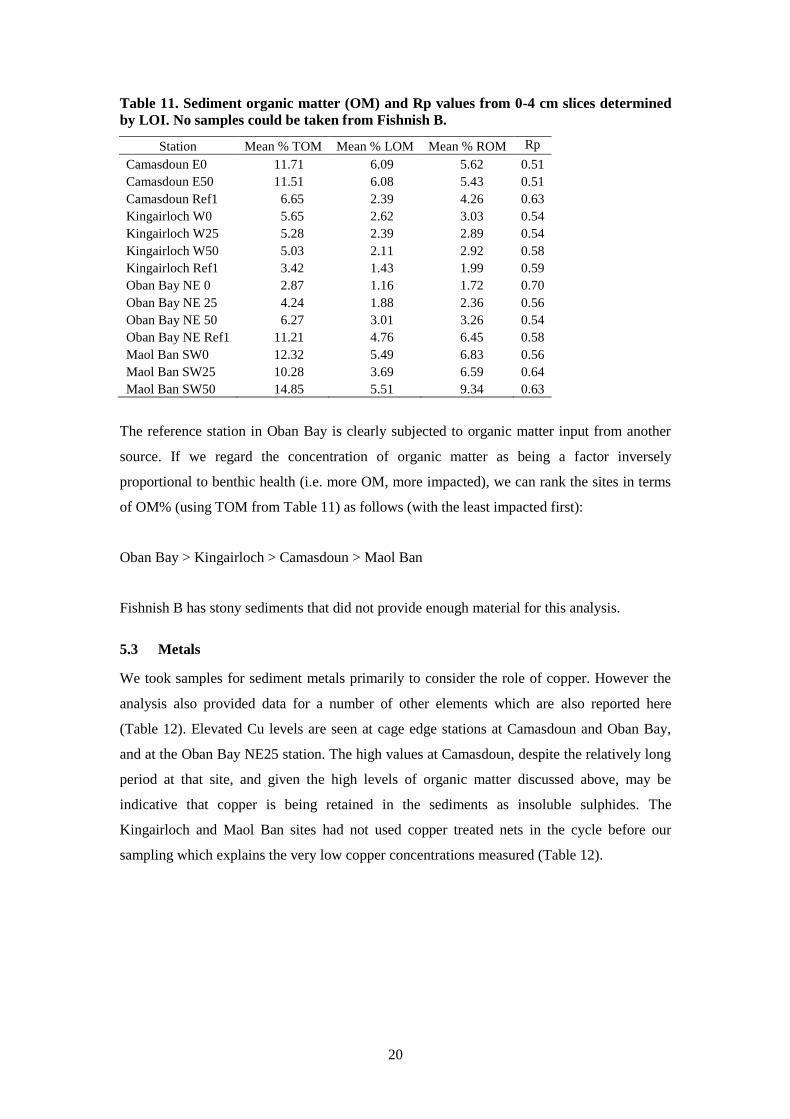
20
Table 11. Sediment organic matter (OM) and Rp values from 0-4 cm slices determined
by LOI. No samples could be taken from Fishnish B.
Station Mean % TOM Mean % LOM Mean % ROM Rp
Camasdoun E0 11.71 6.09 5.62 0.51
Camasdoun E50 11.51 6.08 5.43 0.51
Camasdoun Ref1 6.65 2.39 4.26 0.63
Kingairloch W0 5.65 2.62 3.03 0.54
Kingairloch W25 5.28 2.39 2.89 0.54
Kingairloch W50 5.03 2.11 2.92 0.58
Kingairloch Ref1 3.42 1.43 1.99 0.59
Oban Bay NE 0 2.87 1.16 1.72 0.70
Oban Bay NE 25 4.24 1.88 2.36 0.56
Oban Bay NE 50 6.27 3.01 3.26 0.54
Oban Bay NE Ref1 11.21 4.76 6.45 0.58
Maol Ban SW0 12.32 5.49 6.83 0.56
Maol Ban SW25 10.28 3.69 6.59 0.64
Maol Ban SW50 14.85 5.51 9.34 0.63
The reference station in Oban Bay is clearly subjected to organic matter input from another
source. If we regard the concentration of organic matter as being a factor inversely
proportional to benthic health (i.e. more OM, more impacted), we can rank the sites in terms
of OM% (using TOM from Table 11) as follows (with the least impacted first):
Oban Bay > Kingairloch > Camasdoun > Maol Ban
Fishnish B has stony sediments that did not provide enough material for this analysis.
5.3 Metals
We took samples for sediment metals primarily to consider the role of copper. However the
analysis also provided data for a number of other elements which are also reported here
(Table 12). Elevated Cu levels are seen at cage edge stations at Camasdoun and Oban Bay,
and at the Oban Bay NE25 station. The high values at Camasdoun, despite the relatively long
period at that site, and given the high levels of organic matter discussed above, may be
indicative that copper is being retained in the sediments as insoluble sulphides. The
Kingairloch and Maol Ban sites had not used copper treated nets in the cycle before our
sampling which explains the very low copper concentrations measured (Table 12).
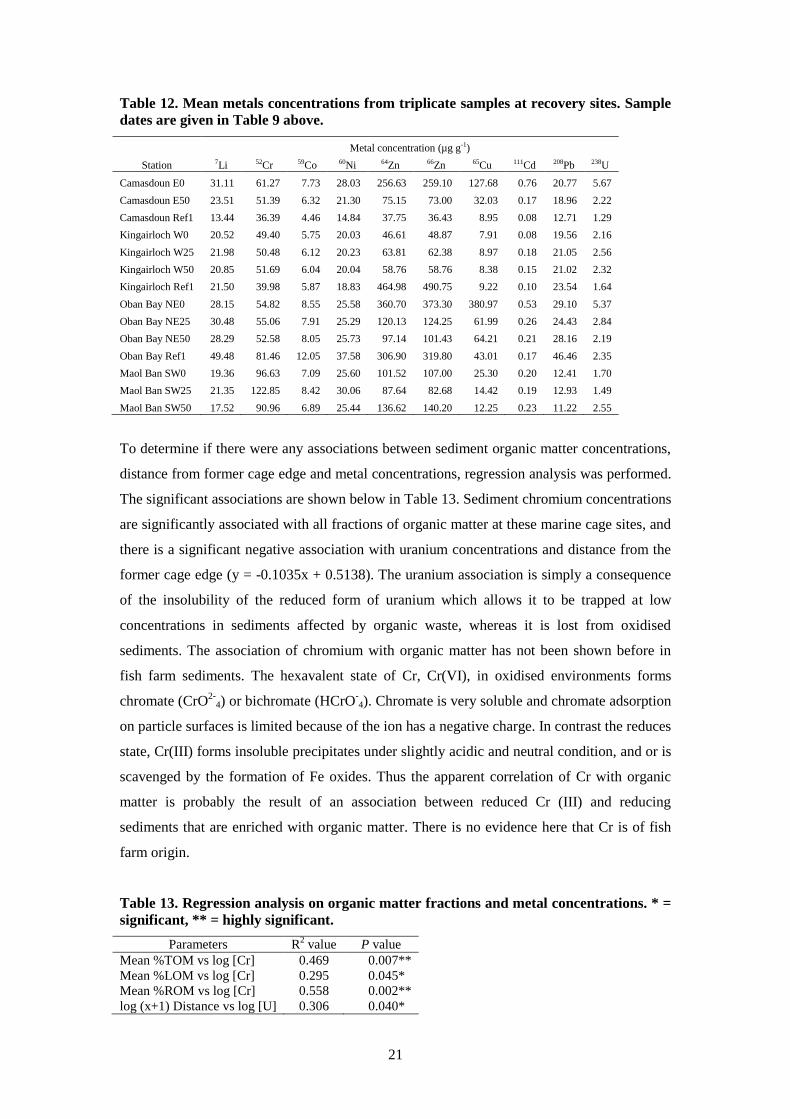
21
Table 12. Mean metals concentrations from triplicate samples at recovery sites. Sample
dates are given in Table 9 above.
Metal concentration (µg g-1)
Station 7Li 52Cr 59Co 60Ni 64Zn 66Zn 65Cu 111Cd 208Pb 238U
Camasdoun E0 31.11 61.27 7.73 28.03 256.63 259.10 127.68 0.76 20.77 5.67
Camasdoun E50 23.51 51.39 6.32 21.30 75.15 73.00 32.03 0.17 18.96 2.22
Camasdoun Ref1 13.44 36.39 4.46 14.84 37.75 36.43 8.95 0.08 12.71 1.29
Kingairloch W0 20.52 49.40 5.75 20.03 46.61 48.87 7.91 0.08 19.56 2.16
Kingairloch W25 21.98 50.48 6.12 20.23 63.81 62.38 8.97 0.18 21.05 2.56
Kingairloch W50 20.85 51.69 6.04 20.04 58.76 58.76 8.38 0.15 21.02 2.32
Kingairloch Ref1 21.50 39.98 5.87 18.83 464.98 490.75 9.22 0.10 23.54 1.64
Oban Bay NE0 28.15 54.82 8.55 25.58 360.70 373.30 380.97 0.53 29.10 5.37
Oban Bay NE25 30.48 55.06 7.91 25.29 120.13 124.25 61.99 0.26 24.43 2.84
Oban Bay NE50 28.29 52.58 8.05 25.73 97.14 101.43 64.21 0.21 28.16 2.19
Oban Bay Ref1 49.48 81.46 12.05 37.58 306.90 319.80 43.01 0.17 46.46 2.35
Maol Ban SW0 19.36 96.63 7.09 25.60 101.52 107.00 25.30 0.20 12.41 1.70
Maol Ban SW25 21.35 122.85 8.42 30.06 87.64 82.68 14.42 0.19 12.93 1.49
Maol Ban SW50 17.52 90.96 6.89 25.44 136.62 140.20 12.25 0.23 11.22 2.55
To determine if there were any associations between sediment organic matter concentrations,
distance from former cage edge and metal concentrations, regression analysis was performed.
The significant associations are shown below in Table 13. Sediment chromium concentrations
are significantly associated with all fractions of organic matter at these marine cage sites, and
there is a significant negative association with uranium concentrations and distance from the
former cage edge (y = -0.1035x + 0.5138). The uranium association is simply a consequence
of the insolubility of the reduced form of uranium which allows it to be trapped at low
concentrations in sediments affected by organic waste, whereas it is lost from oxidised
sediments. The association of chromium with organic matter has not been shown before in
fish farm sediments. The hexavalent state of Cr, Cr(VI), in oxidised environments forms
chromate (CrO2-
4) or bichromate (HCrO-4). Chromate is very soluble and chromate adsorption
on particle surfaces is limited because of the ion has a negative charge. In contrast the reduces
state, Cr(III) forms insoluble precipitates under slightly acidic and neutral condition, and or is
scavenged by the formation of Fe oxides. Thus the apparent correlation of Cr with organic
matter is probably the result of an association between reduced Cr (III) and reducing
sediments that are enriched with organic matter. There is no evidence here that Cr is of fish
farm origin.
Table 13. Regression analysis on organic matter fractions and metal concentrations. * =
significant, ** = highly significant.
Parameters R2 value P value
Mean %TOM vs log [Cr] 0.469 0.007**
Mean %LOM vs log [Cr] 0.295 0.045*
Mean %ROM vs log [Cr] 0.558 0.002**
log (x+1) Distance vs log [U] 0.306 0.040*

22
6 Benthic biology
6.1 Recovery
Several approaches were adopted to determine to what extent recovery had occurred at the
sites studied. A number of standard diversity indices for the benthic community at each
station were calculated, and the ratio of these indices to those at the reference station at each
site calculated. It was postulated that as the diversity index under consideration approached
the value of that at the reference station, the further advanced recovery would be, such that
the ratio of “diversity index at station x” : “diversity index at reference” would approach
unity.
The definition of recovery by Brooks and co-workers (Brooks and Mahnken, 2003; Brooks et
al., 2004) described previously (Section 2.2) was also emplyed, i.e. the taxa at the reference
station that contributed to 1% of the mean total abundance at that station were calculated.
For recovery to have occurred at a particular station, those same taxa must be present at that
station (but not necessarily in the same numbers as at the reference station). We have
developed this idea by taking the number of these “Brooks taxa” present at a particular station
and dividing this by the number of these taxa at the reference station to arrive at a ratio of
recovery, hereafter termed the Brooks Recovery Index (BRI) (Eq. 1).
mean total1%referenceat taxa
ref matchingstation at taxa(BRI)Index Recovery Brooks
(Eq. 1)
A summary of all the indices examined at each of the recovery stations is presented in Table
14 (peak biomass survey) and Table 15 (recovery survey). Increases in some indices will of
course indicate different things. For example, an increase in individual abundance relative to
the reference station is not necessarily indicative of improving (recovery) conditions, as
individual abundance generally reaches a maximum at an optimal enrichment level (the “peak
of opportunists” of Pearson and Rosenberg (Pearson and Rosenberg, 1978)). Likewise, an
increase in the AMBI index is indicative of impacted conditions, thus an increase in the
AMBI value at a particular station relative to the reference station value would also indicate
deteriorating or delayed recovery conditions. For these reasons the indices Abundance (A),
Abundance/Species (A/S), Abundance m-2
(A m-2
) and AMBI are presented as the inverse, i.e.
1
x, so that an increase in this inverse ratio is seen as an improvement in conditions (recovery)
relative to the reference station.
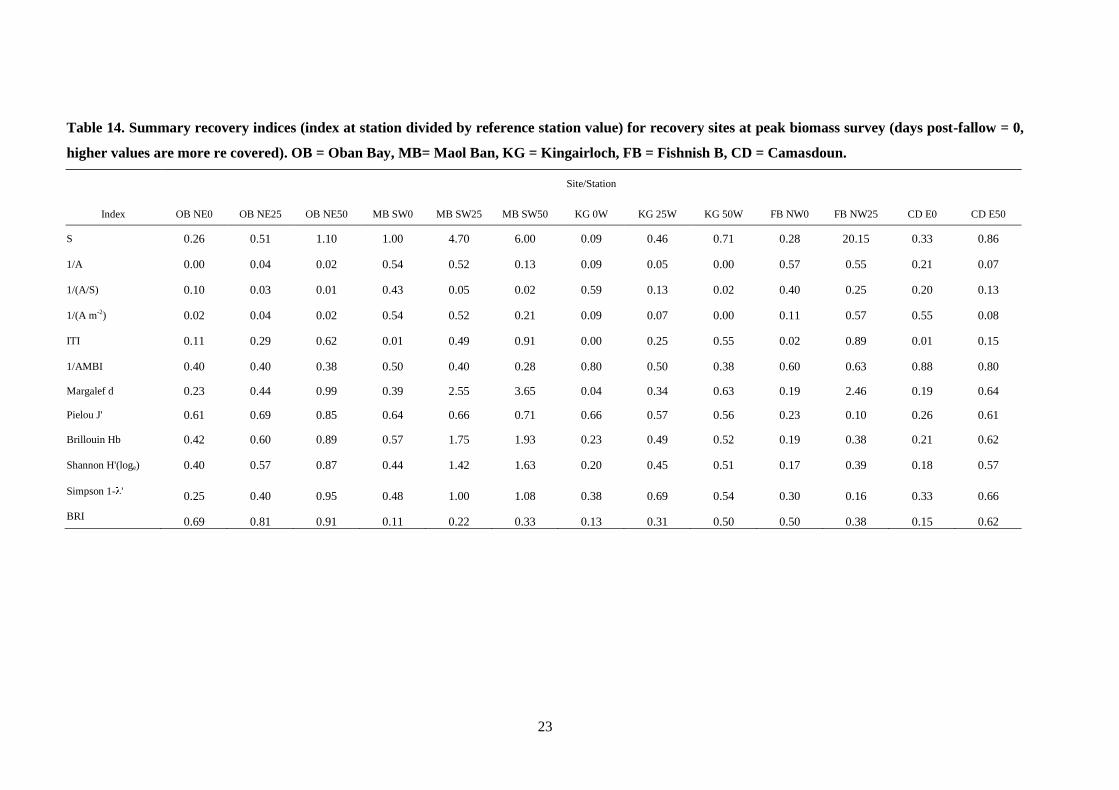
23
Table 14. Summary recovery indices (index at station divided by reference station value) for recovery sites at peak biomass survey (days post-fallow = 0,
higher values are more re covered). OB = Oban Bay, MB= Maol Ban, KG = Kingairloch, FB = Fishnish B, CD = Camasdoun.
Site/Station
Index OB NE0 OB NE25 OB NE50 MB SW0 MB SW25 MB SW50 KG 0W KG 25W KG 50W FB NW0 FB NW25 CD E0 CD E50
S 0.26 0.51 1.10 1.00 4.70 6.00 0.09 0.46 0.71 0.28 20.15 0.33 0.86
1/A 0.00 0.04 0.02 0.54 0.52 0.13 0.09 0.05 0.00 0.57 0.55 0.21 0.07
1/(A/S) 0.10 0.03 0.01 0.43 0.05 0.02 0.59 0.13 0.02 0.40 0.25 0.20 0.13
1/(A m-2) 0.02 0.04 0.02 0.54 0.52 0.21 0.09 0.07 0.00 0.11 0.57 0.55 0.08
ITI 0.11 0.29 0.62 0.01 0.49 0.91 0.00 0.25 0.55 0.02 0.89 0.01 0.15
1/AMBI 0.40 0.40 0.38 0.50 0.40 0.28 0.80 0.50 0.38 0.60 0.63 0.88 0.80
Margalef d 0.23 0.44 0.99 0.39 2.55 3.65 0.04 0.34 0.63 0.19 2.46 0.19 0.64
Pielou J' 0.61 0.69 0.85 0.64 0.66 0.71 0.66 0.57 0.56 0.23 0.10 0.26 0.61
Brillouin Hb 0.42 0.60 0.89 0.57 1.75 1.93 0.23 0.49 0.52 0.19 0.38 0.21 0.62
Shannon H'(loge) 0.40 0.57 0.87 0.44 1.42 1.63 0.20 0.45 0.51 0.17 0.39 0.18 0.57
Simpson 1- ' 0.25 0.40 0.95 0.48 1.00 1.08 0.38 0.69 0.54 0.30 0.16 0.33 0.66
BRI 0.69 0.81 0.91 0.11 0.22 0.33 0.13 0.31 0.50 0.50 0.38 0.15 0.62
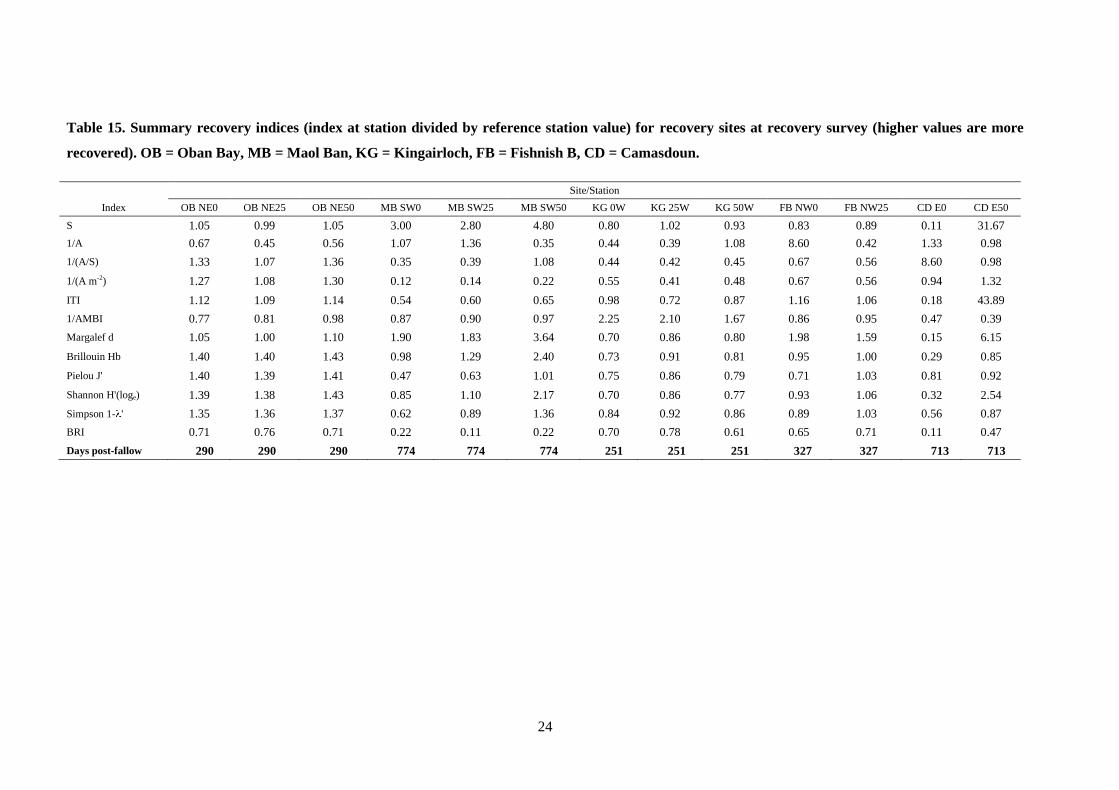
24
Table 15. Summary recovery indices (index at station divided by reference station value) for recovery sites at recovery survey (higher values are more
recovered). OB = Oban Bay, MB = Maol Ban, KG = Kingairloch, FB = Fishnish B, CD = Camasdoun.
Site/Station
Index OB NE0 OB NE25 OB NE50 MB SW0 MB SW25 MB SW50 KG 0W KG 25W KG 50W FB NW0 FB NW25 CD E0 CD E50
S 1.05 0.99 1.05 3.00 2.80 4.80 0.80 1.02 0.93 0.83 0.89 0.11 31.67
1/A 0.67 0.45 0.56 1.07 1.36 0.35 0.44 0.39 1.08 8.60 0.42 1.33 0.98
1/(A/S) 1.33 1.07 1.36 0.35 0.39 1.08 0.44 0.42 0.45 0.67 0.56 8.60 0.98
1/(A m-2) 1.27 1.08 1.30 0.12 0.14 0.22 0.55 0.41 0.48 0.67 0.56 0.94 1.32
ITI 1.12 1.09 1.14 0.54 0.60 0.65 0.98 0.72 0.87 1.16 1.06 0.18 43.89
1/AMBI 0.77 0.81 0.98 0.87 0.90 0.97 2.25 2.10 1.67 0.86 0.95 0.47 0.39
Margalef d 1.05 1.00 1.10 1.90 1.83 3.64 0.70 0.86 0.80 1.98 1.59 0.15 6.15
Brillouin Hb 1.40 1.40 1.43 0.98 1.29 2.40 0.73 0.91 0.81 0.95 1.00 0.29 0.85
Pielou J' 1.40 1.39 1.41 0.47 0.63 1.01 0.75 0.86 0.79 0.71 1.03 0.81 0.92
Shannon H'(loge) 1.39 1.38 1.43 0.85 1.10 2.17 0.70 0.86 0.77 0.93 1.06 0.32 2.54
Simpson 1- ' 1.35 1.36 1.37 0.62 0.89 1.36 0.84 0.92 0.86 0.89 1.03 0.56 0.87
BRI 0.71 0.76 0.71 0.22 0.11 0.22 0.70 0.78 0.61 0.65 0.71 0.11 0.47
Days post-fallow 290 290 290 774 774 774 251 251 251 327 327 713 713
25
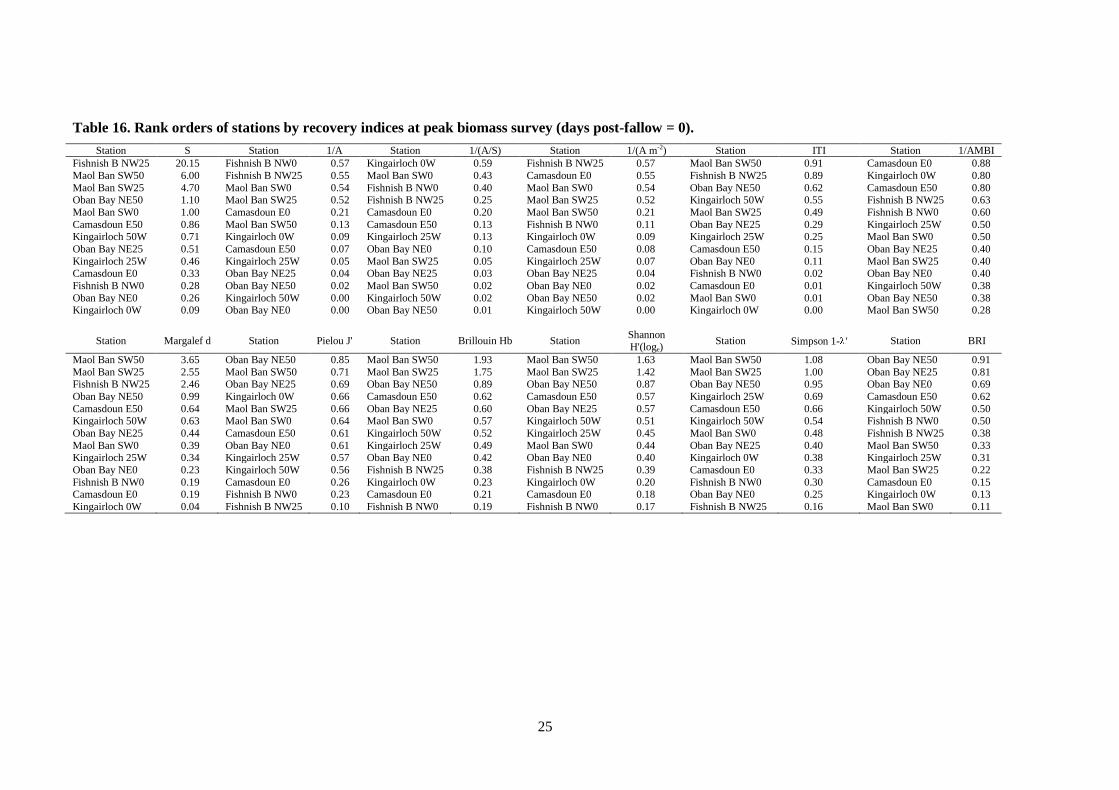
Table 16. Rank orders of stations by recovery indices at peak biomass survey (days post-fallow = 0).
Station S Station 1/A Station 1/(A/S) Station 1/(A m-2) Station ITI Station 1/AMBI
Fishnish B NW25 20.15 Fishnish B NW0 0.57 Kingairloch 0W 0.59 Fishnish B NW25 0.57 Maol Ban SW50 0.91 Camasdoun E0 0.88
Maol Ban SW50 6.00 Fishnish B NW25 0.55 Maol Ban SW0 0.43 Camasdoun E0 0.55 Fishnish B NW25 0.89 Kingairloch 0W 0.80
Maol Ban SW25 4.70 Maol Ban SW0 0.54 Fishnish B NW0 0.40 Maol Ban SW0 0.54 Oban Bay NE50 0.62 Camasdoun E50 0.80 Oban Bay NE50 1.10 Maol Ban SW25 0.52 Fishnish B NW25 0.25 Maol Ban SW25 0.52 Kingairloch 50W 0.55 Fishnish B NW25 0.63
Maol Ban SW0 1.00 Camasdoun E0 0.21 Camasdoun E0 0.20 Maol Ban SW50 0.21 Maol Ban SW25 0.49 Fishnish B NW0 0.60
Camasdoun E50 0.86 Maol Ban SW50 0.13 Camasdoun E50 0.13 Fishnish B NW0 0.11 Oban Bay NE25 0.29 Kingairloch 25W 0.50 Kingairloch 50W 0.71 Kingairloch 0W 0.09 Kingairloch 25W 0.13 Kingairloch 0W 0.09 Kingairloch 25W 0.25 Maol Ban SW0 0.50
Oban Bay NE25 0.51 Camasdoun E50 0.07 Oban Bay NE0 0.10 Camasdoun E50 0.08 Camasdoun E50 0.15 Oban Bay NE25 0.40
Kingairloch 25W 0.46 Kingairloch 25W 0.05 Maol Ban SW25 0.05 Kingairloch 25W 0.07 Oban Bay NE0 0.11 Maol Ban SW25 0.40 Camasdoun E0 0.33 Oban Bay NE25 0.04 Oban Bay NE25 0.03 Oban Bay NE25 0.04 Fishnish B NW0 0.02 Oban Bay NE0 0.40
Fishnish B NW0 0.28 Oban Bay NE50 0.02 Maol Ban SW50 0.02 Oban Bay NE0 0.02 Camasdoun E0 0.01 Kingairloch 50W 0.38
Oban Bay NE0 0.26 Kingairloch 50W 0.00 Kingairloch 50W 0.02 Oban Bay NE50 0.02 Maol Ban SW0 0.01 Oban Bay NE50 0.38 Kingairloch 0W 0.09 Oban Bay NE0 0.00 Oban Bay NE50 0.01 Kingairloch 50W 0.00 Kingairloch 0W 0.00 Maol Ban SW50 0.28
Station Margalef d Station Pielou J' Station Brillouin Hb Station Shannon H'(loge)
Station Simpson 1- ' Station BRI
Maol Ban SW50 3.65 Oban Bay NE50 0.85 Maol Ban SW50 1.93 Maol Ban SW50 1.63 Maol Ban SW50 1.08 Oban Bay NE50 0.91
Maol Ban SW25 2.55 Maol Ban SW50 0.71 Maol Ban SW25 1.75 Maol Ban SW25 1.42 Maol Ban SW25 1.00 Oban Bay NE25 0.81 Fishnish B NW25 2.46 Oban Bay NE25 0.69 Oban Bay NE50 0.89 Oban Bay NE50 0.87 Oban Bay NE50 0.95 Oban Bay NE0 0.69
Oban Bay NE50 0.99 Kingairloch 0W 0.66 Camasdoun E50 0.62 Camasdoun E50 0.57 Kingairloch 25W 0.69 Camasdoun E50 0.62
Camasdoun E50 0.64 Maol Ban SW25 0.66 Oban Bay NE25 0.60 Oban Bay NE25 0.57 Camasdoun E50 0.66 Kingairloch 50W 0.50 Kingairloch 50W 0.63 Maol Ban SW0 0.64 Maol Ban SW0 0.57 Kingairloch 50W 0.51 Kingairloch 50W 0.54 Fishnish B NW0 0.50
Oban Bay NE25 0.44 Camasdoun E50 0.61 Kingairloch 50W 0.52 Kingairloch 25W 0.45 Maol Ban SW0 0.48 Fishnish B NW25 0.38
Maol Ban SW0 0.39 Oban Bay NE0 0.61 Kingairloch 25W 0.49 Maol Ban SW0 0.44 Oban Bay NE25 0.40 Maol Ban SW50 0.33 Kingairloch 25W 0.34 Kingairloch 25W 0.57 Oban Bay NE0 0.42 Oban Bay NE0 0.40 Kingairloch 0W 0.38 Kingairloch 25W 0.31
Oban Bay NE0 0.23 Kingairloch 50W 0.56 Fishnish B NW25 0.38 Fishnish B NW25 0.39 Camasdoun E0 0.33 Maol Ban SW25 0.22
Fishnish B NW0 0.19 Camasdoun E0 0.26 Kingairloch 0W 0.23 Kingairloch 0W 0.20 Fishnish B NW0 0.30 Camasdoun E0 0.15 Camasdoun E0 0.19 Fishnish B NW0 0.23 Camasdoun E0 0.21 Camasdoun E0 0.18 Oban Bay NE0 0.25 Kingairloch 0W 0.13
Kingairloch 0W 0.04 Fishnish B NW25 0.10 Fishnish B NW0 0.19 Fishnish B NW0 0.17 Fishnish B NW25 0.16 Maol Ban SW0 0.11
26
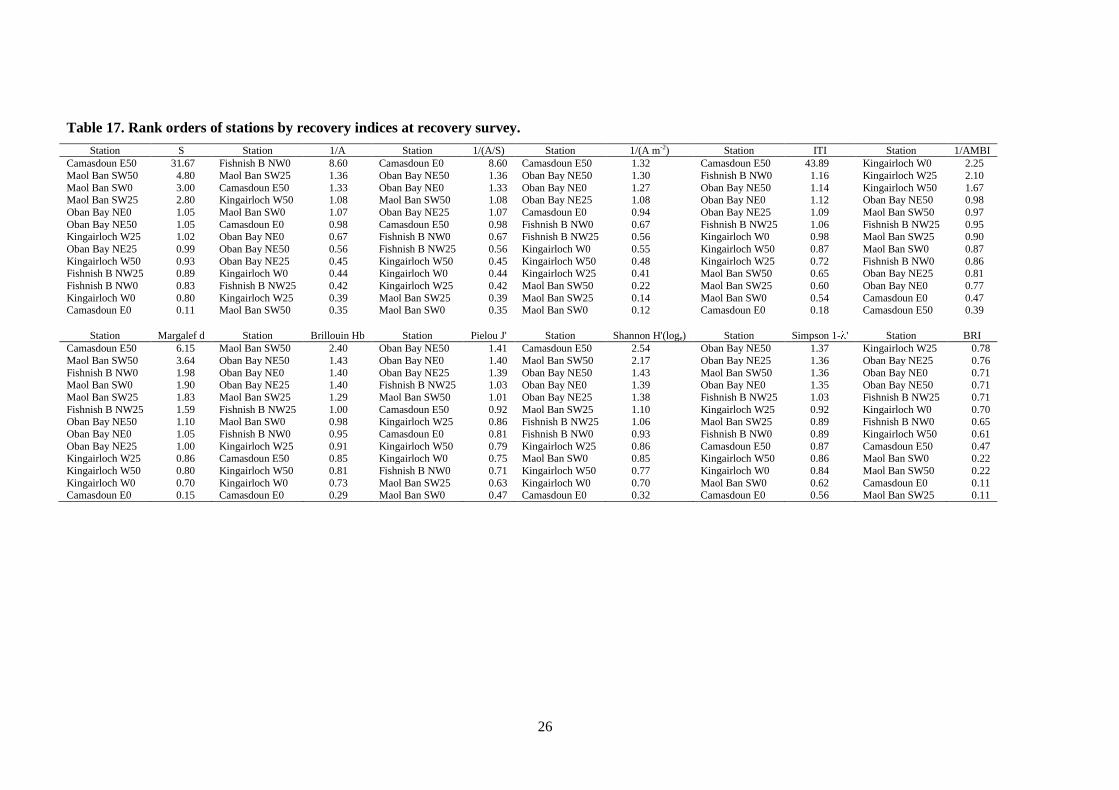
Table 17. Rank orders of stations by recovery indices at recovery survey.
Station S Station 1/A Station 1/(A/S) Station 1/(A m-2) Station ITI Station 1/AMBI
Camasdoun E50 31.67 Fishnish B NW0 8.60 Camasdoun E0 8.60 Camasdoun E50 1.32 Camasdoun E50 43.89 Kingairloch W0 2.25
Maol Ban SW50 4.80 Maol Ban SW25 1.36 Oban Bay NE50 1.36 Oban Bay NE50 1.30 Fishnish B NW0 1.16 Kingairloch W25 2.10
Maol Ban SW0 3.00 Camasdoun E50 1.33 Oban Bay NE0 1.33 Oban Bay NE0 1.27 Oban Bay NE50 1.14 Kingairloch W50 1.67 Maol Ban SW25 2.80 Kingairloch W50 1.08 Maol Ban SW50 1.08 Oban Bay NE25 1.08 Oban Bay NE0 1.12 Oban Bay NE50 0.98
Oban Bay NE0 1.05 Maol Ban SW0 1.07 Oban Bay NE25 1.07 Camasdoun E0 0.94 Oban Bay NE25 1.09 Maol Ban SW50 0.97
Oban Bay NE50 1.05 Camasdoun E0 0.98 Camasdoun E50 0.98 Fishnish B NW0 0.67 Fishnish B NW25 1.06 Fishnish B NW25 0.95 Kingairloch W25 1.02 Oban Bay NE0 0.67 Fishnish B NW0 0.67 Fishnish B NW25 0.56 Kingairloch W0 0.98 Maol Ban SW25 0.90
Oban Bay NE25 0.99 Oban Bay NE50 0.56 Fishnish B NW25 0.56 Kingairloch W0 0.55 Kingairloch W50 0.87 Maol Ban SW0 0.87
Kingairloch W50 0.93 Oban Bay NE25 0.45 Kingairloch W50 0.45 Kingairloch W50 0.48 Kingairloch W25 0.72 Fishnish B NW0 0.86 Fishnish B NW25 0.89 Kingairloch W0 0.44 Kingairloch W0 0.44 Kingairloch W25 0.41 Maol Ban SW50 0.65 Oban Bay NE25 0.81
Fishnish B NW0 0.83 Fishnish B NW25 0.42 Kingairloch W25 0.42 Maol Ban SW50 0.22 Maol Ban SW25 0.60 Oban Bay NE0 0.77
Kingairloch W0 0.80 Kingairloch W25 0.39 Maol Ban SW25 0.39 Maol Ban SW25 0.14 Maol Ban SW0 0.54 Camasdoun E0 0.47 Camasdoun E0 0.11 Maol Ban SW50 0.35 Maol Ban SW0 0.35 Maol Ban SW0 0.12 Camasdoun E0 0.18 Camasdoun E50 0.39
Station Margalef d Station Brillouin Hb Station Pielou J' Station Shannon H'(loge) Station Simpson 1- ' Station BRI
Camasdoun E50 6.15 Maol Ban SW50 2.40 Oban Bay NE50 1.41 Camasdoun E50 2.54 Oban Bay NE50 1.37 Kingairloch W25 0.78
Maol Ban SW50 3.64 Oban Bay NE50 1.43 Oban Bay NE0 1.40 Maol Ban SW50 2.17 Oban Bay NE25 1.36 Oban Bay NE25 0.76
Fishnish B NW0 1.98 Oban Bay NE0 1.40 Oban Bay NE25 1.39 Oban Bay NE50 1.43 Maol Ban SW50 1.36 Oban Bay NE0 0.71 Maol Ban SW0 1.90 Oban Bay NE25 1.40 Fishnish B NW25 1.03 Oban Bay NE0 1.39 Oban Bay NE0 1.35 Oban Bay NE50 0.71
Maol Ban SW25 1.83 Maol Ban SW25 1.29 Maol Ban SW50 1.01 Oban Bay NE25 1.38 Fishnish B NW25 1.03 Fishnish B NW25 0.71
Fishnish B NW25 1.59 Fishnish B NW25 1.00 Camasdoun E50 0.92 Maol Ban SW25 1.10 Kingairloch W25 0.92 Kingairloch W0 0.70 Oban Bay NE50 1.10 Maol Ban SW0 0.98 Kingairloch W25 0.86 Fishnish B NW25 1.06 Maol Ban SW25 0.89 Fishnish B NW0 0.65
Oban Bay NE0 1.05 Fishnish B NW0 0.95 Camasdoun E0 0.81 Fishnish B NW0 0.93 Fishnish B NW0 0.89 Kingairloch W50 0.61
Oban Bay NE25 1.00 Kingairloch W25 0.91 Kingairloch W50 0.79 Kingairloch W25 0.86 Camasdoun E50 0.87 Camasdoun E50 0.47 Kingairloch W25 0.86 Camasdoun E50 0.85 Kingairloch W0 0.75 Maol Ban SW0 0.85 Kingairloch W50 0.86 Maol Ban SW0 0.22
Kingairloch W50 0.80 Kingairloch W50 0.81 Fishnish B NW0 0.71 Kingairloch W50 0.77 Kingairloch W0 0.84 Maol Ban SW50 0.22
Kingairloch W0 0.70 Kingairloch W0 0.73 Maol Ban SW25 0.63 Kingairloch W0 0.70 Maol Ban SW0 0.62 Camasdoun E0 0.11 Camasdoun E0 0.15 Camasdoun E0 0.29 Maol Ban SW0 0.47 Camasdoun E0 0.32 Camasdoun E0 0.56 Maol Ban SW25 0.11

27
To appreciate how mixed the picture of recovery at each site is, the recovery indices at Oban
Bay may be examined. At the Oban Bay cage edge station, the recovery indices S, 1/(A/S),
1/(A m-2
), ITI, Margalef d, Brillouin Hb, Pielou J', Shannon H'(loge) and Simpson 1- ' all
yielded values higher than unity, suggesting that recovery had taken place 290 days post-
fallow (Table 15). In contrast the indices 1/A, 1/AMBI and the BRI all show values below
unity, indicating that recovery was not complete. This situation is repeated across all stations
and sites studied; wherever other recovery indices exceed unity and suggest recovery has
occurred, the BRI in all cases remains below unity.
To estimate the relative recovery status of the sites across all indices tested, the data were
ranked by each index and tabulated (Table 17). Taking the cage edge stations only into
account, these ranks were compared and the stations with the most top ranks tabulated (Table
18). This would suggest that the Oban Bay site had recovered the furthest taking all diversity
recovery indices into account, even though it had the second shortest recovery time since
fallowing. Furthermore, Camasdoun had the most bottom ranks despite having one of the
longest recovery times.
Table 18. Cage edge stations ranked on the basis of most top ranks and fewest bottom
ranks of benthic indices (from Table 17, recovery survey).
Station Top ranks Bottom ranks Days post-fallow
Oban Bay NE0 7 0 290
Fishnish B NW0 3 0 327
Kingairloch W0 1 1 251
Camasdoun E0 1 8 713
Maol Ban SW0 0 3 774
Using Table 18 to place the sites in order of recovery, we obtain:
Oban Bay (most recovered) > Fishnish B > Kingairloch > Camasdoun > Maol Ban
It is also worth considering how the starting points affect the recovery process by analysing
the relative states of impact at the peak biomass survey. From consideration of the highest and
lowest ranked cage edge stations from the various indicators in Table 16, it appears (perhaps
counter-intuitively) that the farms with the least impacted peak biomass stations were in fact
those which had also showed the lowest degree of recovery and vice versa (Table 19). Thus
the order of impact at peak biomass is as follows (with least impacted first):
Maol Ban < Camasdoun < Oban Bay < Kingairloch ≈ Fishnish B.

28
Table 19. Cage edge stations ranked on the basis of most top ranks and fewest bottom
ranks of benthic indices (from Table 16, peak biomass survey).
Station Top ranks Bottom ranks
Maol Ban SW0 5 1
Camasdoun E0 2 0
Oban Bay NE0 2 5
Fishnish B NW0 1 3
Kingairloch W0 1 3
6.2 Changes in benthic function
Macleod and co workers (2008) suggested that functional recovery of the benthic community
was more important than community equivalence:
Ultimately, when assessing recovery, it is the functional recovery that may represent
the critical reference point. Where no further impacts on the environment are
expected, functional recovery may be considered to represent the point at which a
system has demonstrated its capacity to recover based on the biological
characteristics of the community (Macleod et al., 2008).
Following this direction, one hypothesis of recovery would be that, as a community
experiences a decrease in the amount of food (carbon) during recovery, feeding groups
present would shift from sub-surface deposit feeders to filter/suspension feeders normally
excluded from highly depositional environments, and to surface detritus feeders. This
hypothesis was tested by examining the proportion of the fauna at each recovery station that
was placed into the separate ITI feeding groups.
The Infaunal Trophic Index (Word, 1979) classifies benthos into 4 groups:
group 1 are suspension feeders
group 2 are surface detritus feeders
group 3 are surface deposit feeders
group4 are sub–surface deposit feeders
In this section, we examine whether the functionality of the benthos, as inferred from ITI
grouping, can assist in determining the recovery status of the studied fish farms.
Data from Camasdoun are displayed in Table 20. As would be expected, the Camasdoun cage
edge station (E0) at peak biomass (2006) is made up overwhelmingly of Group 4 taxa, with a
tiny proportion of Group 1 taxa. The recovery sample in 2008 shows a remarkable drop in
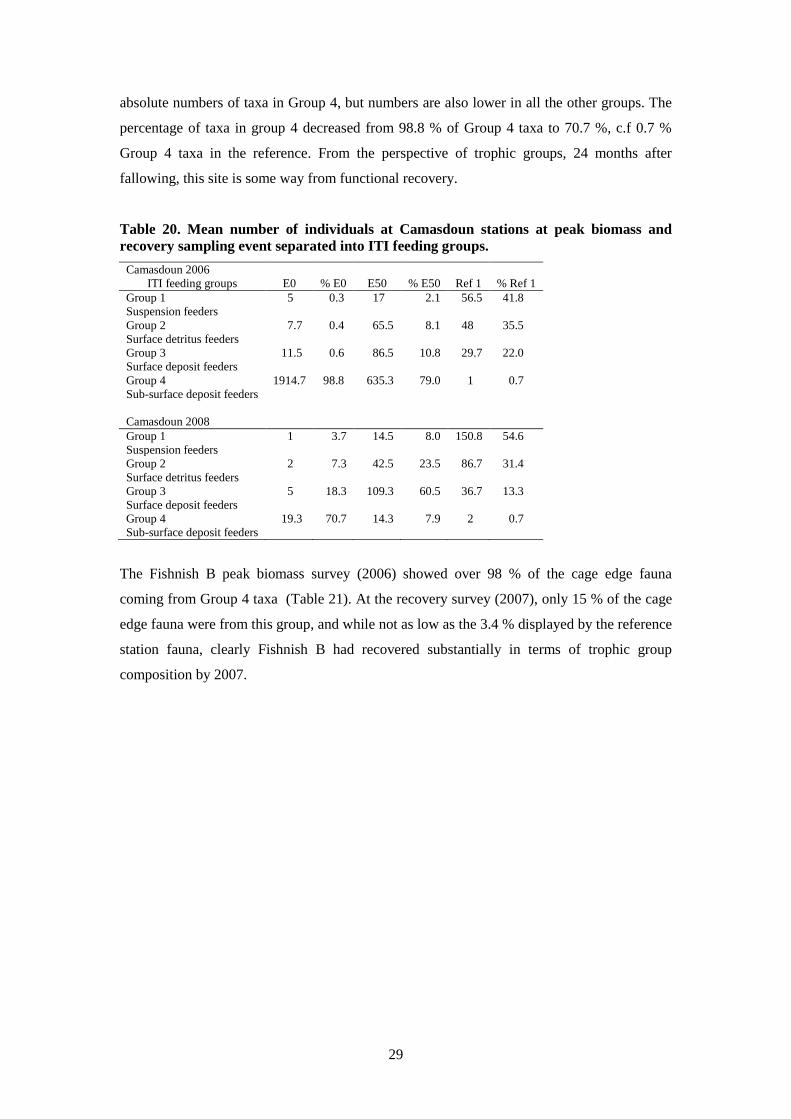
29
absolute numbers of taxa in Group 4, but numbers are also lower in all the other groups. The
percentage of taxa in group 4 decreased from 98.8 % of Group 4 taxa to 70.7 %, c.f 0.7 %
Group 4 taxa in the reference. From the perspective of trophic groups, 24 months after
fallowing, this site is some way from functional recovery.
Table 20. Mean number of individuals at Camasdoun stations at peak biomass and
recovery sampling event separated into ITI feeding groups.
Camasdoun 2006
ITI feeding groups E0 % E0 E50 % E50 Ref 1 % Ref 1
Group 1 5 0.3 17 2.1 56.5 41.8
Suspension feeders
Group 2 7.7 0.4 65.5 8.1 48 35.5
Surface detritus feeders
Group 3 11.5 0.6 86.5 10.8 29.7 22.0
Surface deposit feeders
Group 4 1914.7 98.8 635.3 79.0 1 0.7
Sub-surface deposit feeders
Camasdoun 2008
Group 1 1 3.7 14.5 8.0 150.8 54.6
Suspension feeders
Group 2 2 7.3 42.5 23.5 86.7 31.4
Surface detritus feeders
Group 3 5 18.3 109.3 60.5 36.7 13.3
Surface deposit feeders
Group 4 19.3 70.7 14.3 7.9 2 0.7
Sub-surface deposit feeders
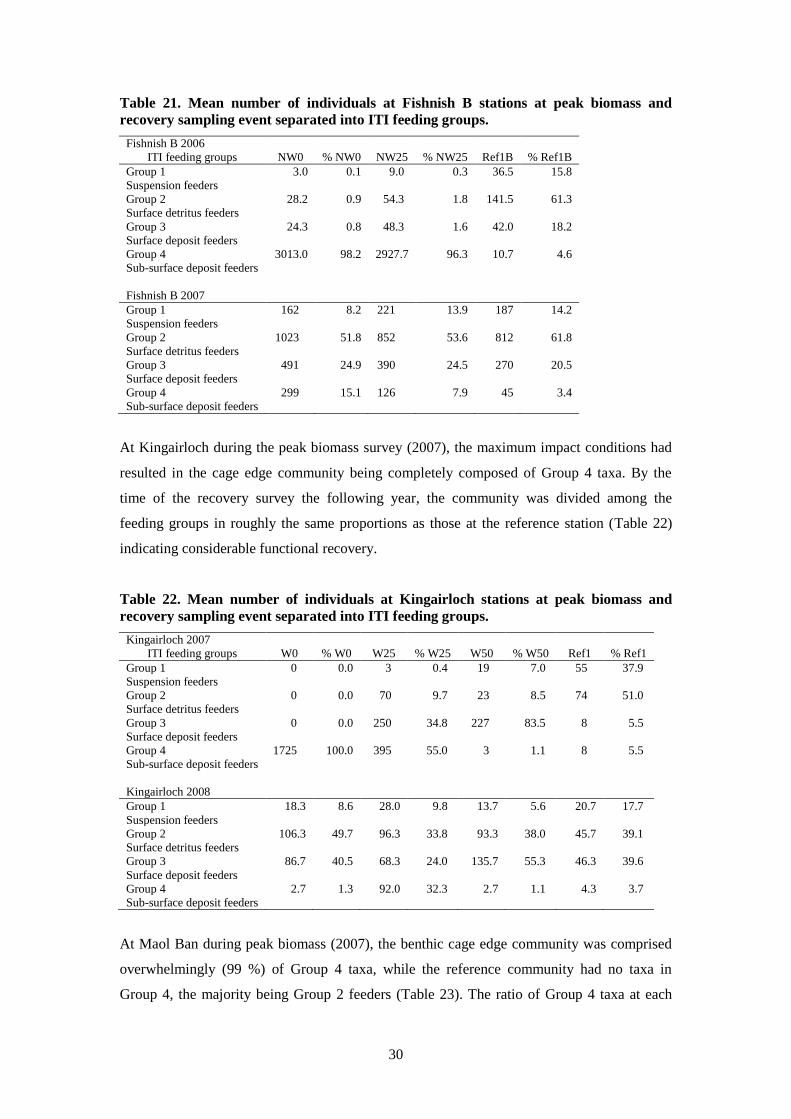
The Fishnish B peak biomass survey (2006) showed over 98 % of the cage edge fauna
coming from Group 4 taxa (Table 21). At the recovery survey (2007), only 15 % of the cage
edge fauna were from this group, and while not as low as the 3.4 % displayed by the reference
station fauna, clearly Fishnish B had recovered substantially in terms of trophic group
composition by 2007.
30
Table 21. Mean number of individuals at Fishnish B stations at peak biomass and
recovery sampling event separated into ITI feeding groups.
Fishnish B 2006
ITI feeding groups NW0 % NW0 NW25 % NW25 Ref1B % Ref1B
Group 1 3.0 0.1 9.0 0.3 36.5 15.8
Suspension feeders
Group 2 28.2 0.9 54.3 1.8 141.5 61.3
Surface detritus feeders
Group 3 24.3 0.8 48.3 1.6 42.0 18.2
Surface deposit feeders
Group 4 3013.0 98.2 2927.7 96.3 10.7 4.6
Sub-surface deposit feeders
Fishnish B 2007
Group 1 162 8.2 221 13.9 187 14.2
Suspension feeders
Group 2 1023 51.8 852 53.6 812 61.8
Surface detritus feeders
Group 3 491 24.9 390 24.5 270 20.5
Surface deposit feeders
Group 4 299 15.1 126 7.9 45 3.4
Sub-surface deposit feeders
At Kingairloch during the peak biomass survey (2007), the maximum impact conditions had
resulted in the cage edge community being completely composed of Group 4 taxa. By the
time of the recovery survey the following year, the community was divided among the
feeding groups in roughly the same proportions as those at the reference station (Table 22)
indicating considerable functional recovery.
Table 22. Mean number of individuals at Kingairloch stations at peak biomass and
recovery sampling event separated into ITI feeding groups.
Kingairloch 2007
ITI feeding groups W0 % W0 W25 % W25 W50 % W50 Ref1 % Ref1
Group 1 0 0.0 3 0.4 19 7.0 55 37.9
Suspension feeders
Group 2 0 0.0 70 9.7 23 8.5 74 51.0
Surface detritus feeders
Group 3 0 0.0 250 34.8 227 83.5 8 5.5
Surface deposit feeders
Group 4 1725 100.0 395 55.0 3 1.1 8 5.5
Sub-surface deposit feeders
Kingairloch 2008
Group 1 18.3 8.6 28.0 9.8 13.7 5.6 20.7 17.7
Suspension feeders
Group 2 106.3 49.7 96.3 33.8 93.3 38.0 45.7 39.1
Surface detritus feeders
Group 3 86.7 40.5 68.3 24.0 135.7 55.3 46.3 39.6
Surface deposit feeders
Group 4 2.7 1.3 92.0 32.3 2.7 1.1 4.3 3.7
Sub-surface deposit feeders
At Maol Ban during peak biomass (2007), the benthic cage edge community was comprised
overwhelmingly (99 %) of Group 4 taxa, while the reference community had no taxa in
Group 4, the majority being Group 2 feeders (Table 23). The ratio of Group 4 taxa at each
31
station to the reference station value is thus undefined. By the recovery survey in 2008, the
former cage edge station had less than 2 % of taxa in Group 4. As no reference station could
be taken in 2008, direct comparisons can not be made, however if the previous survey‟s
reference station is used as a guide, then Maol Ban still had some way to go in terms of
trophic shift before recovery was completed: Group 4 dominance had changed to Group 3
dominance (88 %) whereas the reference site was previously dominated (88 %) by Group 2
animals.
Table 23. Mean number of individuals at Maol Ban stations at peak biomass and
recovery sampling event separated into ITI feeding groups.
Maol Ban 2006
ITI feeding groups SW0 % SW0 SW25 % SW25 SW50 % SW50 Ref1 % Ref1
Group 1 3 0.3 27.5 6.9 17.5 7.4 1 5.9
Suspension feeders
Group 2 0 0.0 112 27.9 170.5 72.4 15 88.2
Surface detritus feeders
Group 3 5.5 0.6 94 23.4 37 15.7 1 5.9
Surface deposit feeders
Group 4 860.5 99.0 167.5 41.8 10.5 4.5 0 0.0
Sub-surface deposit feeders
Maol Ban 2008
Group 1 2.3 1.6 1.0 0.8 5.7 7.5
Suspension feeders
Group 2 11.7 8.0 24.3 19.8 19.3 25.5
Surface detritus feeders
Group 3 129 88.5 96.0 78.0 42.3 56.0
Surface deposit feeders
Group 4 2.7 1.9 1.7 1.4 8.3 11.0
Sub-surface deposit feeders
Table 24 shows the Oban Bay cage edge station to be heavily dominated (90 %) at peak
biomass (2007) by Group 4 taxa, with the reference station showing a majority of the fauna
falling into Groups 3 and 2. By the recovery survey (2008), the Group 4 taxa at the cage edge
had fallen from 90 to 14 % of the fauna with an increase in Group 1 species. While the
reference station was dominated (72 %) by Group 3 animals, the NE0 recovery station
showed a balance between groups 2 and 3 (~ 40 %)
All cage edge stations showed an increase in the proportion of taxa in Group 1 at the recovery
survey relative to the maximum impact (peak biomass) survey. This should also be seen as a
general indication of recovery of the community, as these taxa are the most susceptible to
disturbance from organic enrichment.
32
Table 24. Mean number of individuals at Oban Bay stations at peak biomass and
recovery sampling event separated into ITI feeding groups.
Oban Bay 2007
ITI feeding groups NE0 % NE0 NE25 % NE25 NE50 % NE50 Ref1 % Ref1
Group 1 0.3 0.1 0.7 0.1 7.3 1.2 9.3 2.1
Suspension feeders
Group 2 11.0 2.1 25.3 4.5 112.7 18.0 163.3 36.9
Surface detritus feeders
Group 3 38.7 7.4 167.0 29.9 275.3 44.0 247.3 55.8
Surface deposit feeders
Group 4 471.0 90.4 365.7 65.5 231.0 36.9 23.0 5.2
Sub-surface deposit feeders
Oban Bay 2008
Group 1 9.0 5.2 3.7 1.8 5.0 3.0 6.3 2.9
Suspension feeders
Group 2 72.0 41.6 98.0 48.3 69.0 41.2 48.3 22.0
Surface detritus feeders
Group 3 68.0 39.3 67.0 33.0 87.0 52.0 158.0 72.0
Surface deposit feeders
Group 4 24.0 13.9 34.3 16.9 6.3 3.8 6.7 3.1
Sub-surface deposit feeders
As with the other benthic indices, this analysis of ITI trophic groups is difficult to synthesise
as there are several changes between stations and references and with different groups over
the variable times of the recovery period. In order to simplify this, we have developed a new
index (tentatively called the ITI Recovery Index, IRI) which takes into account the similarity
of the percentage composition of the benthic recovery stations and the appropriate reference
station on the assumption that the reference conditions represent an undisturbed functional
optimum.
The IRI index is calculated as follows:
For each recovery station in the preceding tables (Table 20-Table 24), the % of each trophic
group is compared to the % of that group in the reference station and the 2 numbers are
divided such that the larger number is always the numerator and so the quotient is always
greater than or equal to unity. This is repeated for all four trophic groups and the sum of these
is divided by 4. Thus for identical trophic group proportions between any station and its
reference will yield an IRI index of 1. The larger the value of the index, the more different the
station is from its reference in terms of the proportions of the 4 groups.
In order to compute the IRI index for stations where there is an absence of one or more
trophic groups, 1 % is added to all trophic group percentages within both the station and
reference. For Maol Ban, where no reference station was sampled during the recovery survey,
the monitoring reference composition was used instead. This method then assigns a value to
each station at both sampling points and these can be compared rather easily. Table 25 shows
33
the value of this new index for all the stations in this study ranked in order of the most
recovered.
Table 25. New recovery index (IRI) based on the degree of similarity of the proportion
of benthos in the ITI trophic groups between an impacted station and associated
reference conditions. This data has been ranked with the most recovered station (lower
numbers) at the top. The near cage stations are given in bold italic.
Site Station Monitoring Recovery
Oban Bay NE50 3.0 1.4
Fishnish B NW25 29.8 1.4
Kingairloch W0 28.2 1.6
Kingairloch W50 6.6 1.9
Fishnish B NW0 67.6 2.1
Oban Bay NE0 15.9 2.5
Oban Bay NE25 10.9 2.9
Kingairloch W25 11.7 2.9
Maol Ban NW50 2.6 5.9
Camasdoun E50 34.8 6.0
Maol Ban NW25 12.6 6.5
Maol Ban NW0 49.7 7.1
Camasdoun E0 101.5 30.4
Table 25 can be divided into three groups: those which are relatively similar to references
(indexes of 1 – 3), those that are relatively dissimilar to their reference (index =5-8) and one
outlier where the index is very high (30) indicating a considerable dissimilarity. This yields an
order of recovery with respect to the cage edge stations of: Kingairloch ≈ Fishnish B ≈ Oban
Bay > Maol Ban > Camasdoun. This is similar to the order achieved by consideration of the
ranking of the other indices (Table 18).
6.3 Discussion of the benthic data
The individual indices of recovery at each of the sites show a variety of behaviours. We have
attempted to aggregate and rank these to determine general trends. There is no index available
that gives the “right” answer. This is because benthic communities are in a continuous state of
flux caused by natural as well as anthropogenic drivers as well as purely stochastic processes.
This is the case at both the impacted stations and at reference stations. Thus for each index an
arbitrary criterion is required to indicate various states such as “recovered”, “substantially
recovered” or “far from recovered”. Never-the-less, the use of such a broad suite of indices
does give an intuitively reasonable sense of both of the relative states of recovery and the
closeness to being recovered.
From this consideration, it is clear that Oban Bay, Kingairloch and Fishnish B are much
nearer recovery than Maol Ban and Camasdoun despite the former having less than half the

34
recovery period than the latter. Most interestingly, when we rank the sites by the values of the
indices at the peak biomass survey, to test the hypothesis that sites with a higher impact take
longer to recover (and vice versa) we find that the opposite appears to be the case.
Thus we have two seemingly counter-intuitive conclusions from analysis of these data:
1. site recovery seemed to be less complete at sites which were sampled ca. 2 years after
cessation of farming compared to sites sampled after only one year.
2. site recovery appears greater at the sites with highest monitoring survey benthic impacts
and lower at those sites where the monitoring survey benthic impacts were lower.
As these two conclusions refer to the same sites a composite can be made:
3. The studied sites fell into 2 categories: those that had high initial impacts but recovered
substantially within one year and those that had lower initial impacts but were further from
recovery after 2 years.
The sediment copper levels encountered in the present study (381 µg g-1
at Oban Bay; 128 µg
g-1
at Camasdoun) compare with the maximum of 805 µg g-1
found by Dean et al. (2007) in
their study of a Scottish fish farm site. These authors attributed the high concentrations to the
use of copper-based antifoulants and possibly from feed; Brooks and co-workers (Brooks and
Mahnken, 2003; Brooks et al., 2004) observed these metal concentrations decline to reference
values during recovery in their studies of Canadian Pacific salmon farming. Background
values of copper were calculated by Taylor (1964) at 55 µg g-1
; this compares with the 44 µg
g-1
found by Nickell and Anderson (1997) in their study of distillery effluent in Loch
Harport.
Finally, and this will be important in the second modelling approach, concentrations of
organic matter were much higher in sediments at Camasdoun and Maol Ban cage edge
stations than at the other 3 sites.

35
7 Modelling approach 1 –the Findlay-Watling approach.
7.1 Modelling approach
To determine performance of the existing DEPOMOD model for each of the study sites, the
model was used to predict flux and Infaunal Trophic Index (ITI) at each station at the time of
the monitoring survey for each of the five study sites. This allowed verification of benthic
impact predictions for each site by comparing predicted with observed ITI for each station.
The Findlay-Watling Index (Findlay and Watling, 1997; Morrisey et al., 2000) (denoted FWI
hereafter) was then developed in the model and FWI predictions obtained throughout most of
the production cycle and recovery period. The FWI is the ratio between oxygen supply to the
sediments from near bottom waters and oxygen demand arising from degradation of fish farm
wastes in the sediments. Where oxygen supply is less than demand, FWI is less than 1 and
this causes an oxygen stress on the benthic community and high impact. A FWI of around 1
was found to indicate moderate impact. Oxygen demand was determined from a relationship
between observed carbon flux to the sediments and associated oxygen demand (Findlay and
Watling, 1997; Morrisey et al., 2000). Both of these studies determined oxygen supply from
Fickian diffusion associated with current velocity, temperature and oxygen concentration in
near-bed flow. From current velocity data, the minimum 2 hour average was used as exposure
to reduced oxygen and elevated hydrogen sulphide over 2 hours was hypothesised to results
in permanent damage to gill tissue and thus significant changes to the benthic community.
Thus, Findlay and Watling (1997) and Morrissey et al. (2000) determined only 1 value of the
FWI for each station, based on maximum carbon flux and the minimum 2 hour average of
near bed current.
In our study, we modified the method for calculating oxygen demand and supply, but retained
the principle of the FWI as the ratio of oxygen supply to demand. Oxygen demand was
determined using a relationship between oxygen and carbon degradation, determined using
the first order G-model within DEPOMOD (Westrich and Berner, 1984). Recently measured
fractions and decay rates of salmonid feed and faeces were used (Attard, 2010). In our initial
modelling, we found the minimum 2 hour average to be inappropriate for estimating oxygen
supply, as this statistic is dependent on the details of the current meter deployment
(instrument type (accuracy and minimum threshold), sampling frequency and height above
the bed of deployment). In our initial modelling, we found the minimum 2 hour average to be
inappropriate for estimating oxygen supply, as this statistic is dependent on the details of the
current meter deployment (instrument type - accuracy and minimum threshold, sampling

36
frequency and height above the bed of deployment). In particular, this statistic is influenced
by the high incidence of zeros in a current record obtained with a high threshold current
meter. We overcame this by developing a method which used hourly current data for
calculcation of oxygen supply, rather than a single statistic which was biased by a high
number of zeros in the record. Oxygen supply is calculated from near bed current speed
(Findlay and Watling, 1997), but current is usually measured at different heights above the
bed between sites. Thus, we fitted a logarithmic profile to the measured near-bed current to
determine velocity at 1 cm above the bed, before using the relationships described in Findlay
and Watling (1997). These improvements over the Findlay and Watling method helped to
overcome the sensitivity of the calculation of oxygen supply to instrument type and height
above the bed of deployment. A further improvement we made was to determine the FWI for
each hour throughout most of the production cycle and post fallow period, so that trends in
the FWI could be correlated with benthic impact at the time of monitoring survey, and rate of
recovery for each site.
Thus, the objectives of the modelling were to:
1. verify predictions of benthic impact with observations using ITI - the standard data set of
information collected for each site as part of the regulatory process was used
2. determine whether predictions of FWI correlate with observed benthic impact measured at
the time of the monitoring survey
3. determine if trends in the FWI during both the production cycle and the fallow period
correlate with the rate of benthic recovery of sites after cessation of farming.
We undertook modifications to the DEPOMOD code, included new model parameters in the
G-model and used benthic faunal data from the project surveys. However, to assist with
further application of the model, we largely used site-specific data and model parameters used
in the regulatory process. Revised resuspension model parameters were used (vr ≈ 15 cm s-1
),
primarily influencing predictions at the dispersive site Fishnish B.
7.2 Method
Site specific model input data
Information supplied by the operators for each study was mostly in the form of pre-prepared
data sets used in consent modelling. e.g. current meter data (surface, mid-water and near-bed),

37
bathymetry and cage layouts. For each site, operators supplied monthly biomass and feed
input data for the duration of the production cycle of interest.
Default model input data
Mainly the standard set of model parameters used in regulatory modelling was used as
described in SEPA (2005). The exceptions were the resuspension model settings and water
content of the feed (this was set to 4 % when the G-model was in use following the
experiments of Attard (2010)). All model runs were undertaken with resuspension model
switched on and the revised set of resuspension model parameters (vr ≈ 15 cm s-1
). This
threshold for erosion is higher than the threshold currently used in regulatory modelling, and
this primarily affected Fishnish B (Table 30) by eroding less from the deposition footprint.
Model validation
(i) AutoDEPOMOD benthic module – ITI predictions and observations at time of monitoring
survey
To predict impact at the time of the monitoring survey, benthic fauna monitoring reports
submitted to SEPA were used to determine actual cage layouts and sampling station positions
which were input to AutoDEPOMOD. Three month average feed input was specified by
switching AutoDistribute off and adjusting SFR appropriately. Perform Single Run function
was used so that a single run was undertaken using the feed input at the time of monitoring
survey. No empty cages were modelled.
Predictions of flux and ITI were obtained from the AutoDEPOMOD mapping module
function for the actual sampling station positions at the time of monitoring survey.
Comparisons were then made with observations of ITI taken from benthic fauna monitoring
reports using an average ITI of duplicates where appropriate.
(ii) Findlay-Watling index (FWI) – suitability as indicator of impact and site recovery
To determine whether a relationship existed between FWI and observed benthic impact
and/or rate of recovery, DEPOMOD was set up to undertake lengthy runs for the production
cycle and fallow period. For the month of the monitoring survey, a monthly average for FWI
was determined for each station and compared with observed ITI. Thus, in the model, the
value of FWI would be influenced by the quantity and lability of waste material on the bed
and associated oxygen demand, and so was expected to be a useful indicator of impact.

38
The same model set up was used as previously described in terms of depth, hydrography and
bathymetry. However, a feed water content of 4 % was used as this was determined by Attard
(2010) during experiments on the degradation of feed.
A time series file was created with monthly feed input spread evenly across the number of
cages, and carbon in the feed and faeces was modelled (31 % and 12 % respectively – Attard
(2010)). This file contained information on the mass of carbon in feed and faeces and feed
input discharged every hour, a similar method used to model in-feed chemotheraputants.
The first order decay G-model (Westrich and Berner, 1984) was used to determine the mass
of carbon degraded in the waste feed and faeces every hour for each station throughout the
modelled period:
Gt G1ek1t G2e
k2 tGNR (Eq. 2)
where Gt = carbon at time t (year), G1, G2 and GNR are highly degradable, less degradable and
non-degradable fractions at time t = 0 (k1 and k2 are rate constants for highly and less
degradable carbon respectively - Table 26).
The Arrhenius equation was used with near-bed water temperature measurements for Loch
Creran (5.9 to 9.5 °C) to adjust rate constants dependent on water temperature throughout the
annual cycle for all sites:
k A e
E A
RT (Eq. 3)
where k = rate constant, R = gas constant (= 8.31 J K-1
mol-1
), EA = activation energy (≈ 50
000 J mol-1
assumed constant), T = near-bed water temperature (Kelvin; 273 K = 0 °C) and A
= frequency factor (or pre-exponential or steric factor). Decreasing near-bottom water
temperature from 10 °C to 8 °C resulted in a reduction in the rate constant of 14 % using this
relationship, compared to a reduction of 13 % and 4 % for food and faeces respectively
observed by Attard (2010). Therefore, although useful for feed, the rate constants for faeces
were reduced with temperature more than observed by Attard (2010).
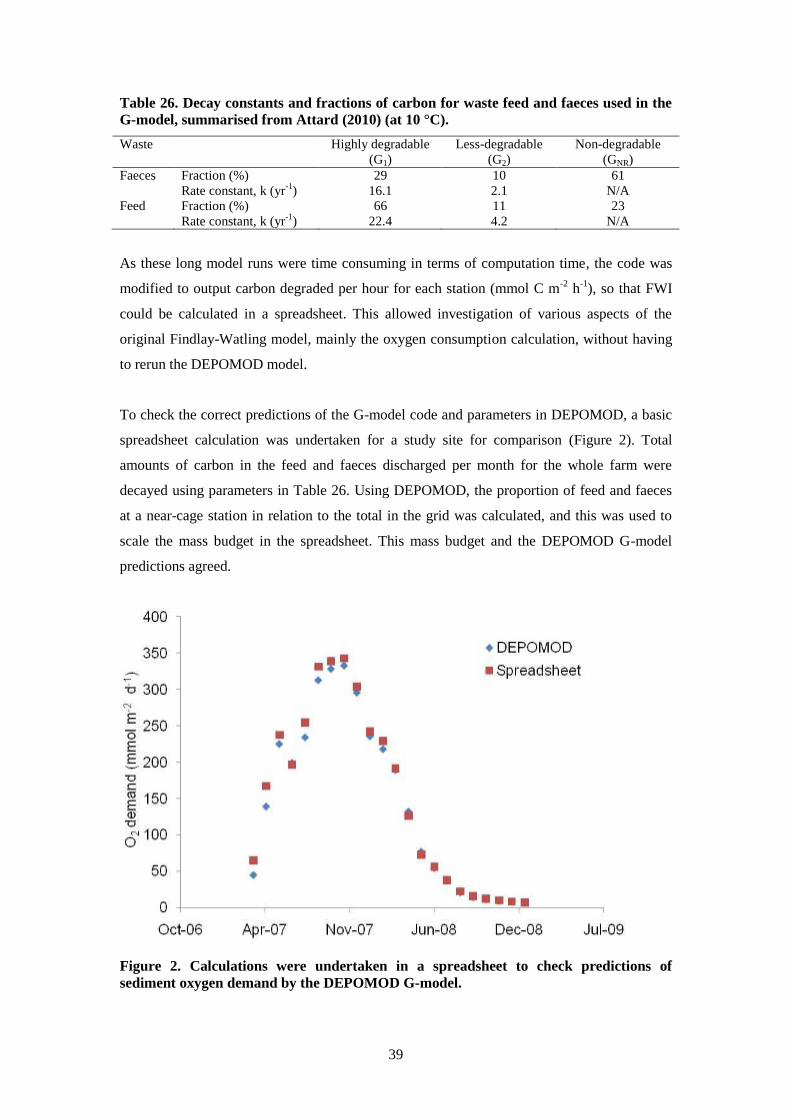
39
Table 26. Decay constants and fractions of carbon for waste feed and faeces used in the
G-model, summarised from Attard (2010) (at 10 °C).
Waste Highly degradable
(G1)
Less-degradable
(G2)
Non-degradable
(GNR)
Faeces
Fraction (%)
Rate constant, k (yr-1
)
29
16.1
10
2.1
61
N/A
Feed
Fraction (%)
Rate constant, k (yr-1
)
66
22.4
11
4.2
23
N/A
As these long model runs were time consuming in terms of computation time, the code was
modified to output carbon degraded per hour for each station (mmol C m-2
h-1
), so that FWI
could be calculated in a spreadsheet. This allowed investigation of various aspects of the
original Findlay-Watling model, mainly the oxygen consumption calculation, without having
to rerun the DEPOMOD model.
To check the correct predictions of the G-model code and parameters in DEPOMOD, a basic
spreadsheet calculation was undertaken for a study site for comparison (Figure 2). Total
amounts of carbon in the feed and faeces discharged per month for the whole farm were
decayed using parameters in Table 26. Using DEPOMOD, the proportion of feed and faeces
at a near-cage station in relation to the total in the grid was calculated, and this was used to
scale the mass budget in the spreadsheet. This mass budget and the DEPOMOD G-model
predictions agreed.
Figure 2. Calculations were undertaken in a spreadsheet to check predictions of
sediment oxygen demand by the DEPOMOD G-model.
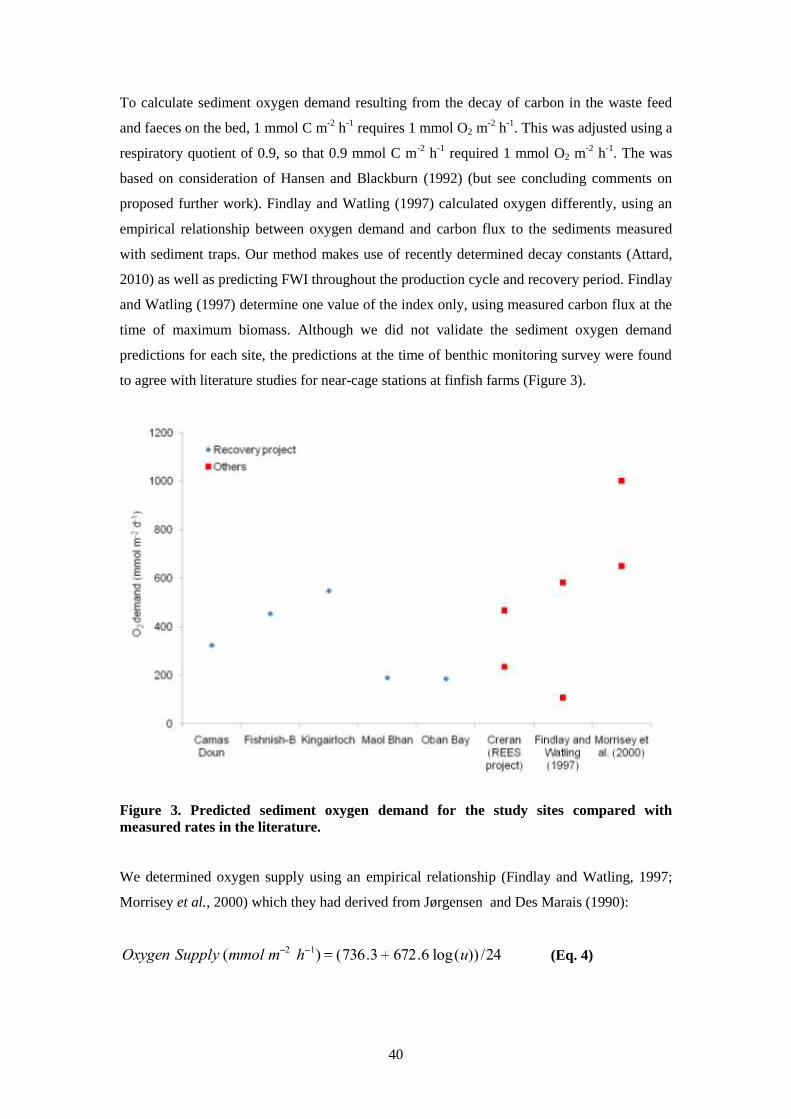
40
To calculate sediment oxygen demand resulting from the decay of carbon in the waste feed
and faeces on the bed, 1 mmol C m-2
h-1
requires 1 mmol O2 m-2
h-1
. This was adjusted using a
respiratory quotient of 0.9, so that 0.9 mmol C m-2
h-1
required 1 mmol O2 m-2
h-1
. The was
based on consideration of Hansen and Blackburn (1992) (but see concluding comments on
proposed further work). Findlay and Watling (1997) calculated oxygen differently, using an
empirical relationship between oxygen demand and carbon flux to the sediments measured
with sediment traps. Our method makes use of recently determined decay constants (Attard,
2010) as well as predicting FWI throughout the production cycle and recovery period. Findlay
and Watling (1997) determine one value of the index only, using measured carbon flux at the
time of maximum biomass. Although we did not validate the sediment oxygen demand
predictions for each site, the predictions at the time of benthic monitoring survey were found
to agree with literature studies for near-cage stations at finfish farms (Figure 3).
Figure 3. Predicted sediment oxygen demand for the study sites compared with
measured rates in the literature.
We determined oxygen supply using an empirical relationship (Findlay and Watling, 1997;
Morrisey et al., 2000) which they had derived from Jørgensen and Des Marais (1990):
Oxygen Supply (mmol m 2 h 1) (736.3 672.6 log(u)) /24 (Eq. 4)

41
where u is near-bed current velocity. However, we standardised near-bed current by fitting a
logarithmic profile and calculating near bed velocity at a height of 0.01 m, as this height was
consistent with the data presented in the experiments by Jørgensen and Des Marais (1990) on
which Findlay and Watling (1997) depended:5
U *u(z,t)
ln (z zo) (Eq. 5)
where U* = shear velocity, u(z,t) = current velocity at height above the bed z and time t, κ =
von Kàrmàn constant (= 0.4) and zo is bed roughness (= 0.0002 m (Soulsby, 1983)). Using
this equation, u* was calculated for each hour and then u (z=0.01 m, t) was calculated by
rearranging Equation 5. Thus, a time series of near-bed current velocity at z = 0.01 m for each
site was used to calculate oxygen supply for each hour. Bed roughness was not varied
between sites.
Thus, using predictions of oxygen demand and supply the Findlay-Watling index was
calculated on an hourly basis:
FWIOxygen supply
Oxygen demand (Eq. 6)
Our method predicted oxygen supply and demand for each hour, resulting in an hourly value
of FWI throughout the modelled period. Although we do not expect the model to predict
accurately the balance between oxygen supply and demand on such a high temporal scale, it
does allow averaging of FWI for a particular month. This approach is in contrast to Findlay
and Watling‟s (1997) study, where a minimum 2 hour average current velocity is used to
determine oxygen supply resulting in one value of FWI only for each site.
To validate predictions of FWI as an indicator of impact, FWI was calculated for the month of
the benthic monitoring survey for each station and compared with observed ITI. To
investigate FWI as an indicator of potential for recovery the following were investigated for
cage-edge stations:
5 It appears that Findlay and Watling (and therefore subsequently Morrisey et al. (2000)) used current speeds at 1
m above the bed while using a relationship given by Jørgensen and Des Marais (1990) that had measured diffusive
oxygen fluxes in the mm scale diffusive boundary layer using currents measured at 1 cm above the bed. In the
present study we have driven the oxygen supply model using the relationships given in the Jørgensen and Des
Marais (1990) study.

42
(i) monthly averages of FWI throughout the production cycle and fallow period, as this may
indicate the general balance of demand and supply for a site.
(ii) the number of consecutive 2 hour periods where FWI was less than 1, as this may also
indicate prolonged periods of oxygen stress and may be related to potential for recovery.
(iii) the rate at which the FWI increases after cessation of farming. i.e. if FWI increases
rapidly then this indicates that oxygen supply far exceeds demand and may be a contributing
factor to the potential for a site to recover.
7.3 Calculation of FWI using the method in Findlay and Watling (1997) and
comparison with observed ITI
7.3.1 Method
Morrisey et al. (2000) stated a limitation of FWI as calculated in Findlay and Watling (1997)
to be over reliance on a single value of the minimum 2 hour average near-bed current velocity
calculated for each site. We also note, that the relationship between oxygen demand and
carbon flux results in zero oxygen demand for low carbon flux (< 130 g C m-2
yr-1
; < 800 g
solid m-2
yr-1
). This means that using the method in Findlay and Watling (1997) would result
in zero oxygen demand at some of our intermediate stations. In addition, for two of our sites
the minimum 2 hour average is less than 0.1 cm s-1
, resulting in zero oxygen supply from the
Findlay and Watling (1997) relationship. Therefore, to test these limitations, we predicted
FWI using the method of Findlay and Watling (1997) to determine if the index correlated
with observed ITI at the time of monitoring survey.
Predictions of solids flux at each station were taken from AutoDEPOMOD as previously
described. As AutoDEPOMOD predicts solids and not carbon, to calculate carbon flux from
solids flux a factor of 0.152 was applied. This takes account of the expected proportions of
feed and faeces at each station, and also the carbon fractions of feed and faeces. This
approach however, does not vary proportions of feed and faeces between near-cage and far-
field stations.
Carbon flux was converted to mmol m-2
d-1
and oxygen demand calculated from Findlay and
Watling (1997) (oxygen demand = 1.1*carbon flux – 32.6). FWI was calculated as previously
described and compared with observed ITI for each station at the time of monitoring survey.

43
7.3.2 Statistical testing of model performance
Model performance was statistically tested following the method proposed by Portilla and
Tett (2007) and applied by Jusup et al. (2009), where model predictions were placed on the
x-axis and observed values on the y-axis. The primary reason for plotting predicted on the x-
axis, is that predictions for each scenario are under the sole control of the modeller and are
likely to show less variation than observations (Portilla and Tett, 2007).
Models were assessed using the linear regression of Yi = β0 + β1Xi for observed (Yi) and
predicted (Xi) values, where β0 is the intercept and β1 the slope. A Student‟s t-test was
undertaken for r2, β0, β1 (α=0.05, n-2 degrees of freedom) to test if r
2, β0 and β1 were
significantly different from 1, 0 and 1 respectively (Mesplé et al., 1996). Model performance
was classified according to the outcome of these tests (Oreskes et al., 1994) (Table 27).
Table 27. Classification of model performance according to the linear regression
between observed and predicted values.
Slope (β1) Intercept (β0) Linear correlation coefficient (r2) Classification
not significantly different
from 1
not significantly
different from 0
significantly > 0 Excellent
Only one of the above
criteria are satisfied
significantly > 0 Good
significantly different from 1 significantly different
from 0
significantly > 0 Fair
n/a n/a not significantly different from 0 Poor
Efficiency (E), which is a measure of goodness of fit, was also used to assess model
performance and the agreement between observations and predictions (Mayer and Butler,
1993):
E 1( ˆ Y i Yi)
2
i 1
n
(Yi Y )2
i 1
n (Eq. 7)
where Yi observed, ˆ Y modelled and Y observed mean for n comparisons.
The higher the value of E, the higher the agreement between predictions and observations.
Mayer and Butler (1993) caution against using models where E < 0.
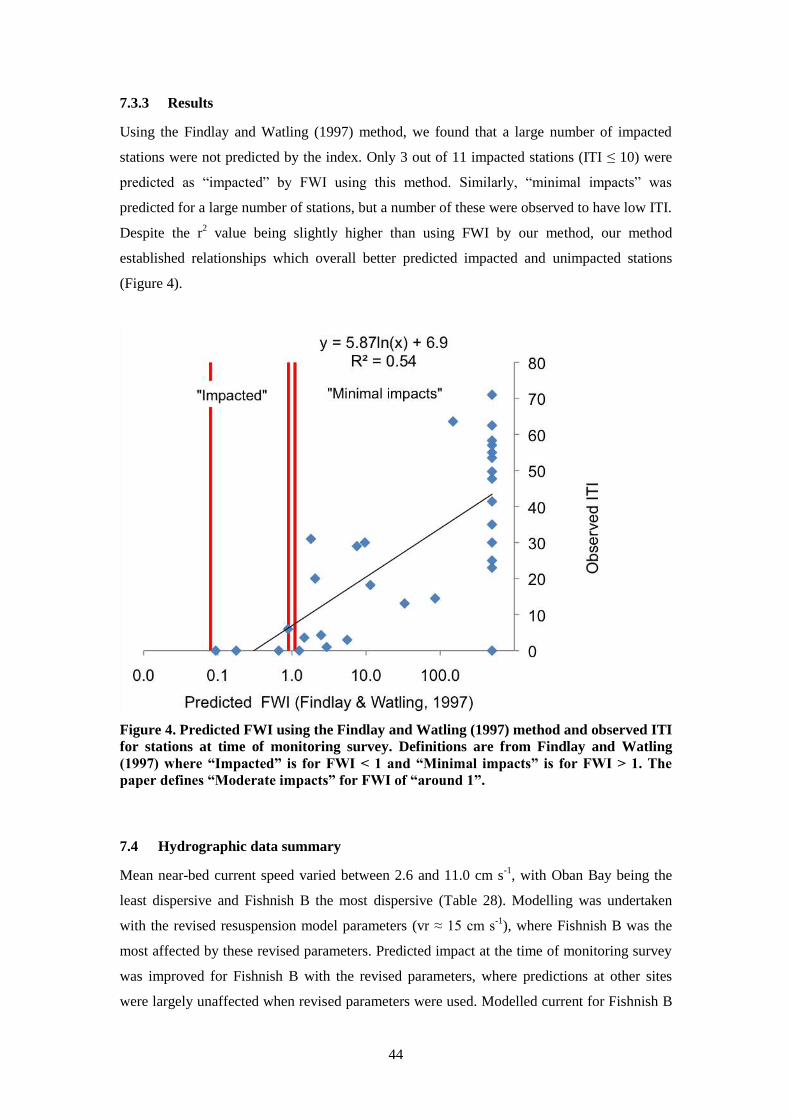
44
7.3.3 Results
Using the Findlay and Watling (1997) method, we found that a large number of impacted
stations were not predicted by the index. Only 3 out of 11 impacted stations (ITI ≤ 10) were
predicted as “impacted” by FWI using this method. Similarly, “minimal impacts” was
predicted for a large number of stations, but a number of these were observed to have low ITI.
Despite the r2 value being slightly higher than using FWI by our method, our method
established relationships which overall better predicted impacted and unimpacted stations
(Figure 4).
Figure 4. Predicted FWI using the Findlay and Watling (1997) method and observed ITI
for stations at time of monitoring survey. Definitions are from Findlay and Watling
(1997) where “Impacted” is for FWI < 1 and “Minimal impacts” is for FWI > 1. The
paper defines “Moderate impacts” for FWI of “around 1”.
7.4 Hydrographic data summary
Mean near-bed current speed varied between 2.6 and 11.0 cm s-1
, with Oban Bay being the
least dispersive and Fishnish B the most dispersive (Table 28). Modelling was undertaken
with the revised resuspension model parameters (vr ≈ 15 cm s-1
), where Fishnish B was the
most affected by these revised parameters. Predicted impact at the time of monitoring survey
was improved for Fishnish B with the revised parameters, where predictions at other sites
were largely unaffected when revised parameters were used. Modelled current for Fishnish B
45
was much higher than observed – these predictions have not been calibrated as the focus of
the hydrographic model calibration undertaken in a previous project was Fiunary, north of
Fishnish B in the Sound of Mull (Black et al., 2009).
Table 28. Hydrographic data summary for study sites and rank on mean near-bed
current speed.
Near-bed current (cm s-1)
Site
Highest
rank on
mean (= most
dispersive)
Min Mean Max <=3 cm s-1 <=9.5 cm
s-1 <=15 cm s-1
Min. 2
houra
Depth 0 m
station (m)
Fishnish B 4 0.1 11.0 31.9 21.9 48.6 69.4 0.08 25
Kingairloch 3 0.1 5.1 17.8 36.1 86.9 99.4 0.53 22
Maol Ban 2 0.1 4.1 14.1 36.9 96.1 100.0 0.29 26
Camasdoun 1 0.3 3.8 13.4 46.7 96.9 100.0 0.24 28
Oban Bay 0 0.0 2.6 14.1 70.6 97.8 100.0 0.00 30
Fishnish B (mod.)b - 1.3 22.8 61.6 1.9 22.2 36.1 3.30 -
Notes: a. Minimum 2 hour average current velocity (used by Findlay and Watling (1997);
b. Using
modelled current from SSPO Sound of Mull project
7.5 Model validation
7.5.1 (i) AutoDEPOMOD benthic module – ITI predictions and observations at time
of monitoring survey
Model performance for predicting ITI was defined as „Excellent‟ using the Portilla and Tett
(2007) classification (Table 27). The outliers were 2 stations at Fishnish B where the model
underpredicted impact (a dispersive site) and 2 stations at Maol Ban where the model
overpredicted impact (a depositional site) (Figure 5). Better predictions were obtained for
Oban Bay, Camasdoun and Kingairloch. These results indicate the overall model performance
at the five study sites is acceptable (Table 29) and allows some confidence in the predictions
of the waste flux to the sea bed which is required for prediction and validation of FWI.

46
Figure 5. Model performance – observed and predicted ITI for the benthic monitoring
survey for (a) individual sites and (b) a trend line fitted to the whole data set. Reference
stations are not shown.
7.5.2 (ii) Findlay-Watling index (FWI) – suitability as indicator of impact and site
recovery
Predicted oxygen supply to the sediments dependent on near-bed current speed showed
significant differences between sites (Figure 6). Oban Bay was predicted to have prolonged
periods of low oxygen supply, whereas Fishnish B had the highest average oxygen supply,
although still exhibiting periods of low oxygen supply associated with low current. NB: For
both of these sites the discharge consent current record was measured using impeller type
current meters with relatively high current thresholds (Table 6).
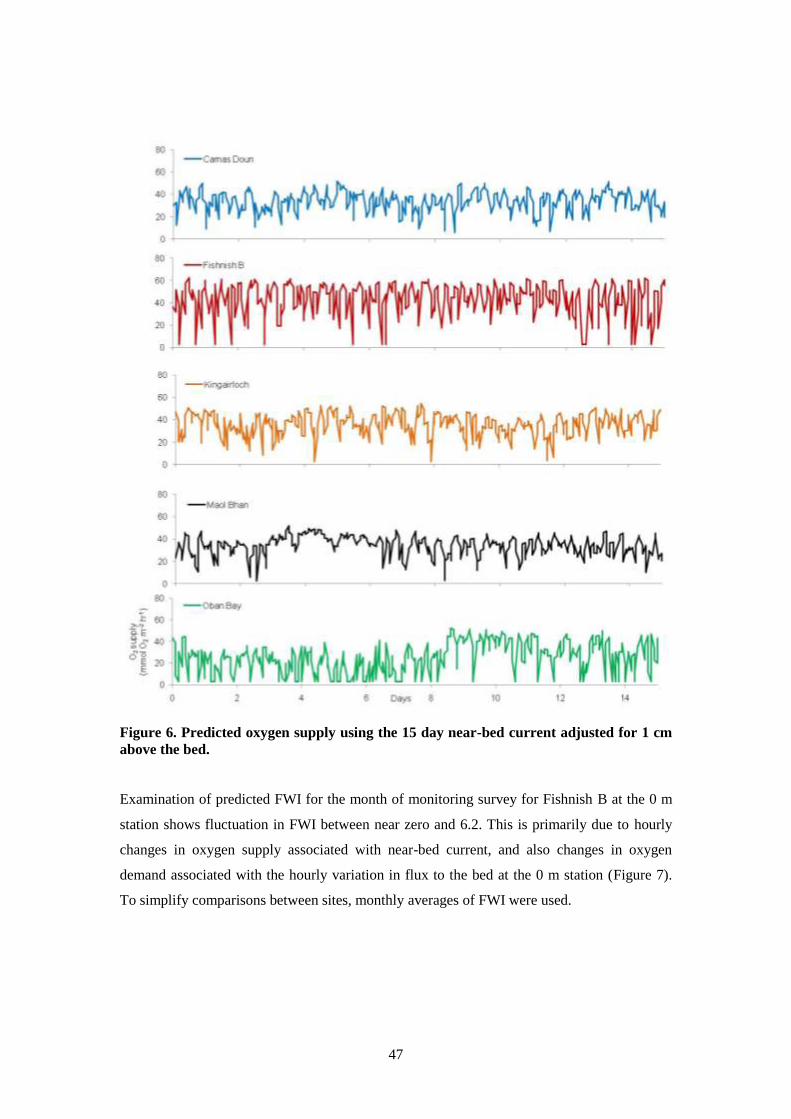
47
Figure 6. Predicted oxygen supply using the 15 day near-bed current adjusted for 1 cm
above the bed.
Examination of predicted FWI for the month of monitoring survey for Fishnish B at the 0 m
station shows fluctuation in FWI between near zero and 6.2. This is primarily due to hourly
changes in oxygen supply associated with near-bed current, and also changes in oxygen
demand associated with the hourly variation in flux to the bed at the 0 m station (Figure 7).
To simplify comparisons between sites, monthly averages of FWI were used.
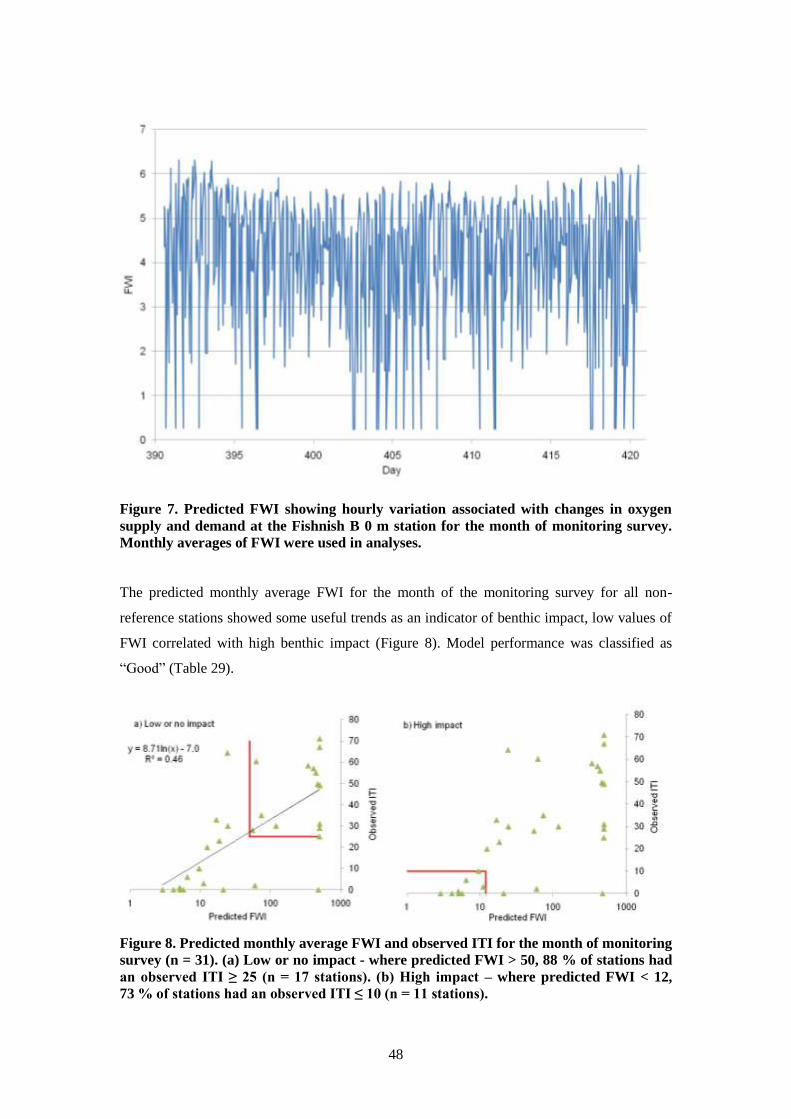
48
Figure 7. Predicted FWI showing hourly variation associated with changes in oxygen
supply and demand at the Fishnish B 0 m station for the month of monitoring survey.
Monthly averages of FWI were used in analyses.
The predicted monthly average FWI for the month of the monitoring survey for all non-
reference stations showed some useful trends as an indicator of benthic impact, low values of
FWI correlated with high benthic impact (Figure 8). Model performance was classified as
“Good” (Table 29).
Figure 8. Predicted monthly average FWI and observed ITI for the month of monitoring
survey (n = 31). (a) Low or no impact - where predicted FWI > 50, 88 % of stations had
an observed ITI ≥ 25 (n = 17 stations). (b) High impact – where predicted FWI < 12,
73 % of stations had an observed ITI ≤ 10 (n = 11 stations).
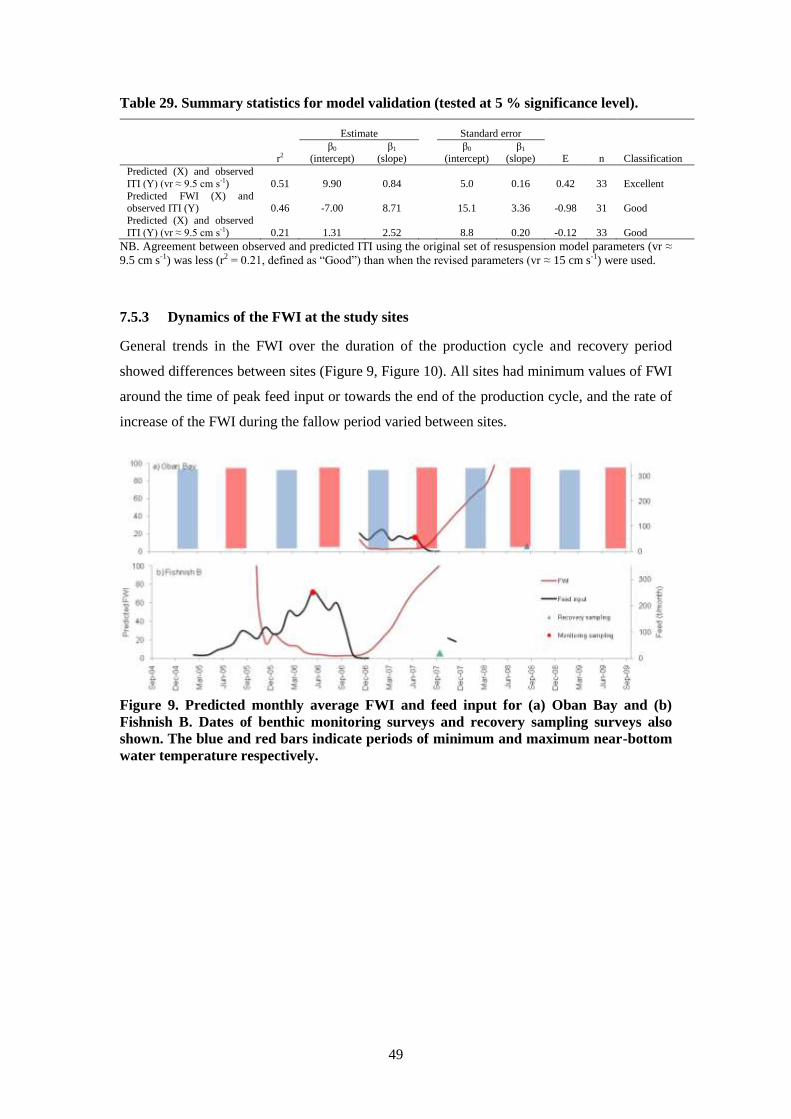
49
Table 29. Summary statistics for model validation (tested at 5 % significance level).
Estimate
Standard error
r2 β0
(intercept) β1
(slope) β0
(intercept) β1
(slope) E n Classification
Predicted (X) and observed
ITI (Y) (vr ≈ 9.5 cm s-1) 0.51 9.90 0.84
5.0 0.16 0.42 33 Excellent
Predicted FWI (X) and observed ITI (Y) 0.46 -7.00 8.71
15.1 3.36 -0.98 31 Good
Predicted (X) and observed
ITI (Y) (vr ≈ 9.5 cm s-1) 0.21 1.31 2.52
8.8 0.20 -0.12 33 Good
NB. Agreement between observed and predicted ITI using the original set of resuspension model parameters (vr ≈
9.5 cm s-1) was less (r2 = 0.21, defined as “Good”) than when the revised parameters (vr ≈ 15 cm s-1) were used.
7.5.3 Dynamics of the FWI at the study sites
General trends in the FWI over the duration of the production cycle and recovery period
showed differences between sites (Figure 9, Figure 10). All sites had minimum values of FWI
around the time of peak feed input or towards the end of the production cycle, and the rate of
increase of the FWI during the fallow period varied between sites.
Figure 9. Predicted monthly average FWI and feed input for (a) Oban Bay and (b)
Fishnish B. Dates of benthic monitoring surveys and recovery sampling surveys also
shown. The blue and red bars indicate periods of minimum and maximum near-bottom
water temperature respectively.
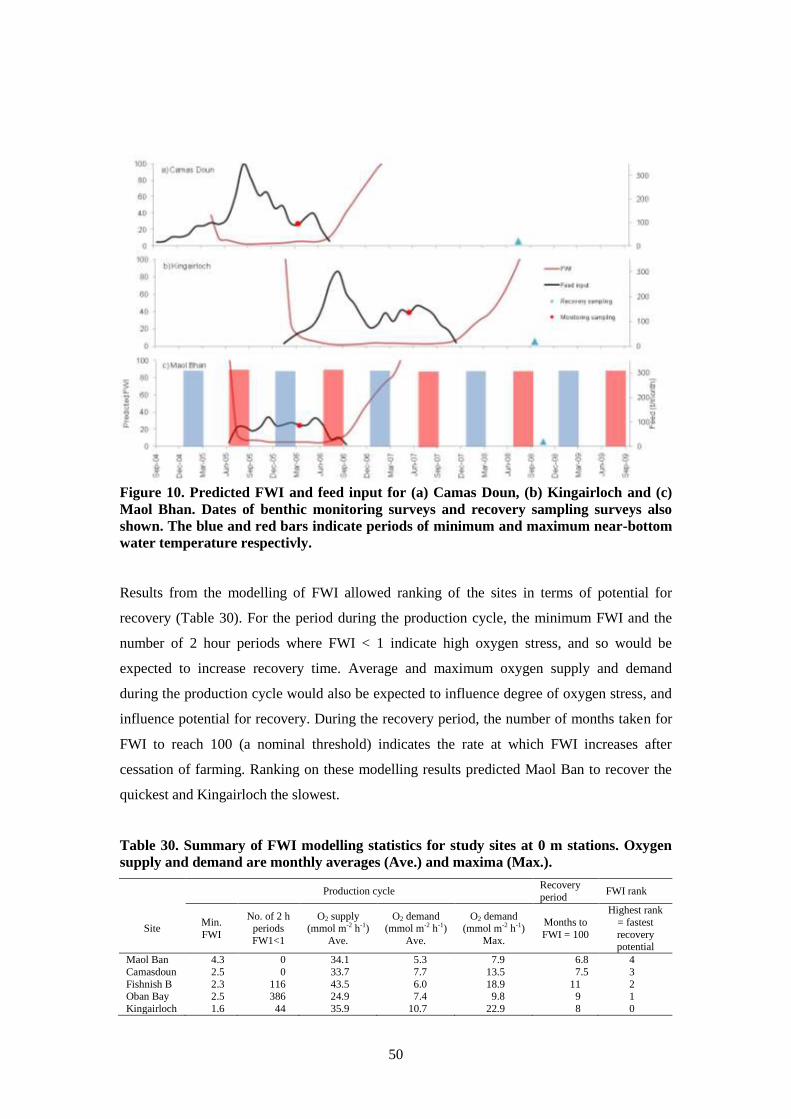
50
Figure 10. Predicted FWI and feed input for (a) Camas Doun, (b) Kingairloch and (c)
Maol Bhan. Dates of benthic monitoring surveys and recovery sampling surveys also
shown. The blue and red bars indicate periods of minimum and maximum near-bottom
water temperature respectivly.
Results from the modelling of FWI allowed ranking of the sites in terms of potential for
recovery (Table 30). For the period during the production cycle, the minimum FWI and the
number of 2 hour periods where FWI < 1 indicate high oxygen stress, and so would be
expected to increase recovery time. Average and maximum oxygen supply and demand
during the production cycle would also be expected to influence degree of oxygen stress, and
influence potential for recovery. During the recovery period, the number of months taken for
FWI to reach 100 (a nominal threshold) indicates the rate at which FWI increases after
cessation of farming. Ranking on these modelling results predicted Maol Ban to recover the
quickest and Kingairloch the slowest.
Table 30. Summary of FWI modelling statistics for study sites at 0 m stations. Oxygen
supply and demand are monthly averages (Ave.) and maxima (Max.).
Production cycle Recovery
period FWI rank
Site Min.
FWI
No. of 2 h periods
FW1<1
O2 supply (mmol m-2 h-1)
Ave.
O2 demand (mmol m-2 h-1)
Ave.
O2 demand (mmol m-2 h-1)
Max.
Months to
FWI = 100
Highest rank
= fastest
recovery potential
Maol Ban 4.3 0 34.1 5.3 7.9 6.8 4
Camasdoun 2.5 0 33.7 7.7 13.5 7.5 3
Fishnish B 2.3 116 43.5 6.0 18.9 11 2 Oban Bay 2.5 386 24.9 7.4 9.8 9 1
Kingairloch 1.6 44 35.9 10.7 22.9 8 0
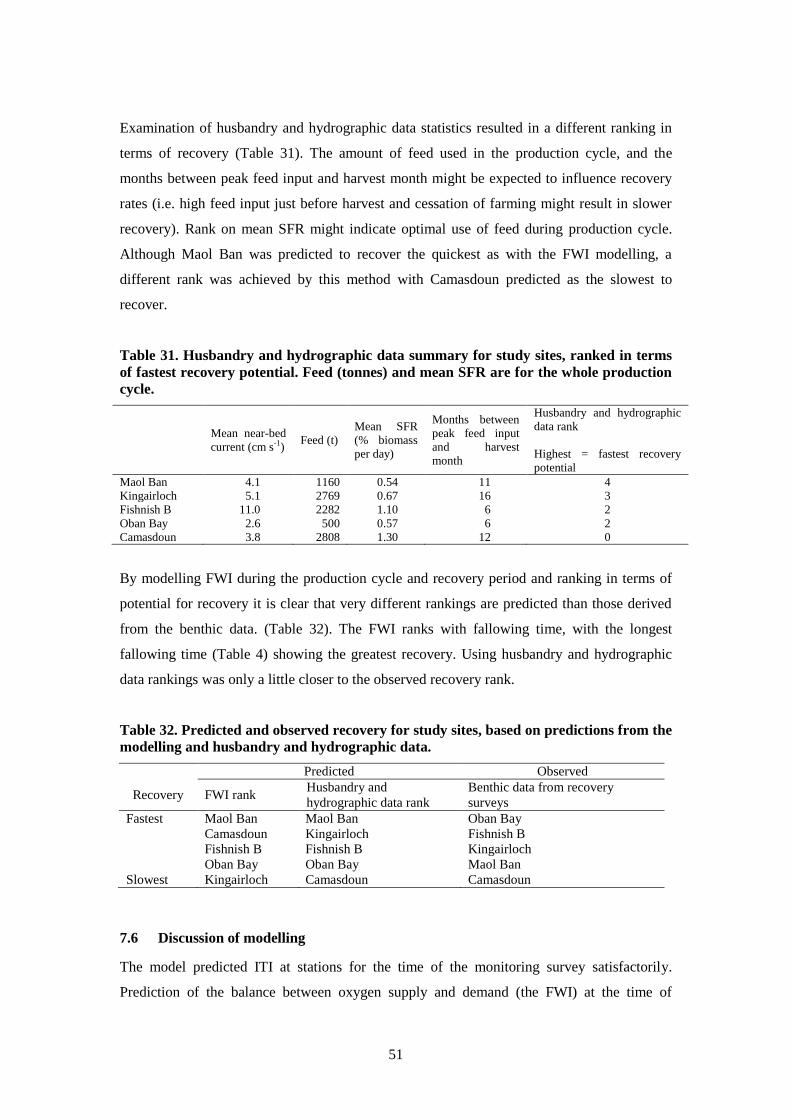
51
Examination of husbandry and hydrographic data statistics resulted in a different ranking in
terms of recovery (Table 31). The amount of feed used in the production cycle, and the
months between peak feed input and harvest month might be expected to influence recovery
rates (i.e. high feed input just before harvest and cessation of farming might result in slower
recovery). Rank on mean SFR might indicate optimal use of feed during production cycle.
Although Maol Ban was predicted to recover the quickest as with the FWI modelling, a
different rank was achieved by this method with Camasdoun predicted as the slowest to
recover.
Table 31. Husbandry and hydrographic data summary for study sites, ranked in terms
of fastest recovery potential. Feed (tonnes) and mean SFR are for the whole production
cycle.
Mean near-bed
current (cm s-1) Feed (t)
Mean SFR
(% biomass
per day)
Months between
peak feed input
and harvest
month
Husbandry and hydrographic
data rank
Highest = fastest recovery
potential
Maol Ban 4.1 1160 0.54 11 4
Kingairloch 5.1 2769 0.67 16 3
Fishnish B 11.0 2282 1.10 6 2
Oban Bay 2.6 500 0.57 6 2
Camasdoun 3.8 2808 1.30 12 0
By modelling FWI during the production cycle and recovery period and ranking in terms of
potential for recovery it is clear that very different rankings are predicted than those derived
from the benthic data. (Table 32). The FWI ranks with fallowing time, with the longest
fallowing time (Table 4) showing the greatest recovery. Using husbandry and hydrographic
data rankings was only a little closer to the observed recovery rank.
Table 32. Predicted and observed recovery for study sites, based on predictions from the
modelling and husbandry and hydrographic data.
Predicted Observed
Recovery FWI rank Husbandry and
hydrographic data rank
Benthic data from recovery
surveys
Fastest Maol Ban Maol Ban Oban Bay
Camasdoun Kingairloch Fishnish B
Fishnish B Fishnish B Kingairloch
Oban Bay Oban Bay Maol Ban
Slowest Kingairloch Camasdoun Camasdoun
7.6 Discussion of modelling
The model predicted ITI at stations for the time of the monitoring survey satisfactorily.
Prediction of the balance between oxygen supply and demand (the FWI) at the time of

52
monitoring survey for each of the stations also correlated with observed ITI, indicating the
FWI has some potential for prediction of impact during the farming period. In particular,
some general relationships were established for low FWI (high impact) and ITI. In addition,
the predicted sediment oxygen demand for 0 m stations was similar to literature values
obtained from field measurements at active fish farms. These findings indicate that prediction
of FWI for a particular time in the production cycle is useful for predicting impact and
provided better predictive capability than using a single value of FWI based on minimum 2
hour current value (as undertaken by Findlay and Watling (1997)).
However, FWI was unable to predict the potential for recovery of sites. Use of husbandry and
hydrographic data statistics was also not useful in ranking the sites.
Apart from the sensitivity of predictions of dispersion to hydrographic data used for each site,
modelling of oxygen supply was highly sensitive to hydrographic data. In particular, where
low current was measured by current meters this results in very low predictions of oxygen
supply and associated low values of FWI. This influences results significantly, as prolonged
periods of low oxygen supply are predicted, when in fact, these may be an artefact of the
senstitivity of the current meter in use i.e. a high threshold current meter may record zero
current for quiescent periods. The Findlay and Watling (1997) oxygen supply equation is
sensitive to low current values and therefore instrument type influences this type of modelling
significantly. By standardising the measured current to a height of 1 cm above the bed, we
minimised the influence of measurements at different heights above the bed to some extent.
However, we could not completely overcome the influence of current meter type – this is
likely to be a factor in the poor performance of the FWI rankings.
Modelling of Fishnish B using modelled current from a hydrodynamic model resulted in
predictions of very high values of FWI > 100 throughout the production cycle (minimal
oxygen stress). This was due to the predictions of high oxygen supply associated with the
high near-bed currents (mean of 22.8 cm s-1
as opposed to 11.0 cm s-1
for observed), as well
as dispersion of the wastes. These modelled currents were not calibrated for Fishnish B, but
they indicate the influence of near bed current on this type of modelling.
As the model could not predict the differences between sites in terms of speed of recovery, it
could also not predict the time required for recovery. To investigate oxygen demand in the
recovery period, decay constants were reduced by 20% and the model run for Fishnish B,
keeping relative fractions of the labile and refractory components constant. Where lower
decay constants were used, a lower oxygen demand was predicted during the production cycle
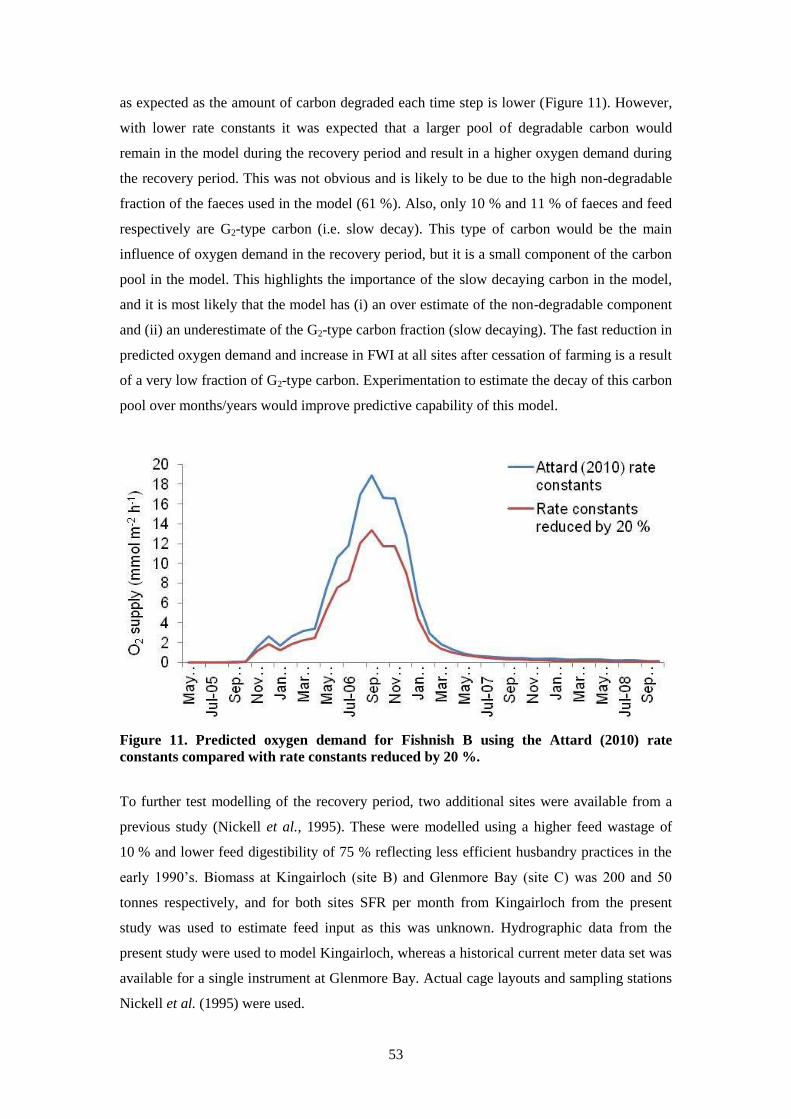
53
as expected as the amount of carbon degraded each time step is lower (Figure 11). However,
with lower rate constants it was expected that a larger pool of degradable carbon would
remain in the model during the recovery period and result in a higher oxygen demand during
the recovery period. This was not obvious and is likely to be due to the high non-degradable
fraction of the faeces used in the model (61 %). Also, only 10 % and 11 % of faeces and feed
respectively are G2-type carbon (i.e. slow decay). This type of carbon would be the main
influence of oxygen demand in the recovery period, but it is a small component of the carbon
pool in the model. This highlights the importance of the slow decaying carbon in the model,
and it is most likely that the model has (i) an over estimate of the non-degradable component
and (ii) an underestimate of the G2-type carbon fraction (slow decaying). The fast reduction in
predicted oxygen demand and increase in FWI at all sites after cessation of farming is a result
of a very low fraction of G2-type carbon. Experimentation to estimate the decay of this carbon
pool over months/years would improve predictive capability of this model.
Figure 11. Predicted oxygen demand for Fishnish B using the Attard (2010) rate
constants compared with rate constants reduced by 20 %.
To further test modelling of the recovery period, two additional sites were available from a
previous study (Nickell et al., 1995). These were modelled using a higher feed wastage of
10 % and lower feed digestibility of 75 % reflecting less efficient husbandry practices in the
early 1990‟s. Biomass at Kingairloch (site B) and Glenmore Bay (site C) was 200 and 50
tonnes respectively, and for both sites SFR per month from Kingairloch from the present
study was used to estimate feed input as this was unknown. Hydrographic data from the
present study were used to model Kingairloch, whereas a historical current meter data set was
available for a single instrument at Glenmore Bay. Actual cage layouts and sampling stations
Nickell et al. (1995) were used.
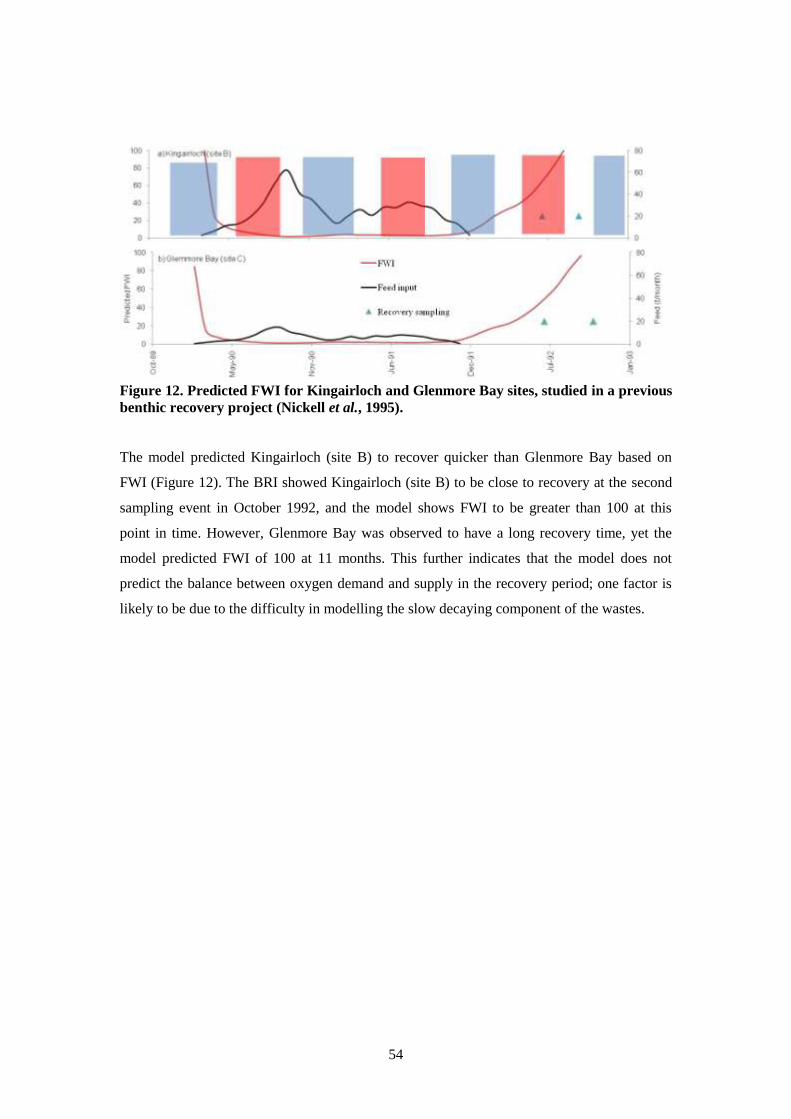
54
Figure 12. Predicted FWI for Kingairloch and Glenmore Bay sites, studied in a previous
benthic recovery project (Nickell et al., 1995).
The model predicted Kingairloch (site B) to recover quicker than Glenmore Bay based on
FWI (Figure 12). The BRI showed Kingairloch (site B) to be close to recovery at the second
sampling event in October 1992, and the model shows FWI to be greater than 100 at this
point in time. However, Glenmore Bay was observed to have a long recovery time, yet the
model predicted FWI of 100 at 11 months. This further indicates that the model does not
predict the balance between oxygen demand and supply in the recovery period; one factor is
likely to be due to the difficulty in modelling the slow decaying component of the wastes.

55
8 Modelling approach 2 – the RQ model
8.1 Introduction
The ultimate objective is to produce a model that includes only the first order processes that
are sufficient to make useful predictions. It is not intended to produce a biogeochemical
model that attempts a full description of all relevant processes - such a model would be
complex, data intensive, computing power intensive and ultimately unfit for use by industry
or regulatory agents.
The benthic results of this project are at first sight counter intuitive (Section 6.3) and reveal
that recovery is a complex process that is not simply a function of the degree of impact at the
end of the farming period and that the rate of recovery varies considerably between sites. This
is in a sense confirmed by the modelling carried out (section 7), as this has been built on the
assumption that the Findlay-Watling index (FWI) on its own would correlate with recovery
time, which it clearly does not. The FWI worked well in published studies at active sites
(Findlay and Watling, 1997; Morrisey et al., 2000) and also in this study during the period of
maximum biomass (Section 7.5). What is clear from our work is that these simple
assumptions are insufficient during the recovery process and that further work is necessary to
incorporate more complex processes.
In order to address the issue of the present model‟s inadequacies, our first thoughts were that
we need to look again at the currents - especially for the 2 sites that used impeller current
meters. DEPOMOD is very sensitive to the accuracy of hydrodynamical parameters, and this
is likely to be especially important when considering oxygen supply. For this reason these
new measurements are justified but this is most likely to be a second order improvement and
unlikely to make a large qualitative difference to the present results.
Our next thought was that the problem lay in the rate constants/carbon proportions for
degradation so we have explored whether revised parameters would under any likely
circumstances be able to change the predicted order of recovery between the sites. We
concluded that this is not likely to make a qualitative difference but think that this should be
done to provide greater confidence in the quantitative aspects. However, we would wish to do
this using a sediment system rather than a pure carbon system used in the present associated
study (Attard, 2010). This would allow study of the storage of reduced products during the
production phase for re-oxidation during the recovery phase, more of which later.

56
Our further thinking has come from an analysis of the key benthic conclusion:
The studied sites fell into 2 categories: those that had high initial impacts but recovered
substantially within one year and those that had lower initial impacts but were further from
recovery after 2 years."
Considering the LOI results it is clear that the sites that take longest to recover have much
higher levels of organic matter (OM) both at impacted and control stations compared to the
three sites which recovered most quickly. This leads us back to consideration of Macleod et
al.‟s paper (2007) on the differences of recovery rate between sites and their dependence on
the ambient environment. The following is a quotation form the abstract of this paper:
At the Stringers Cove site the primary ecological functions of the background community
were restored, but this was not the case at Creeses Mistake. These differences in recovery
response were a direct reflection of background environmental conditions. Stringers Cove
sediments had naturally high organic carbon content and as a result there was greater
similarity in the ecological function of the unimpacted and impacted conditions at this
location than at Creeses Mistake, where, under unimpacted conditions, the sediments had a
very low organic content.
The paper goes on to hypothesise that sites that have high organic carbon content might be
expected to recover more quickly than sites with low organic carbon content but, as that study
considered only short term process (3 months), and the proposed mechanisms for this
hypothesis related to larval supply for re-colonising sediments - rather than relating to the
biogeochemical status of the sediments, this mechanism is probably not the most important on
the timescales of most interest to the present project (1-3 years).
The model described above (section 7) takes local chemical and biological aspects into
account through the consideration of recovery (biologically) with respect to local controls
rather than through any direct measurement of the carbon richness of the system. What if sites
with high background levels of OM from natural sources have a) a greater resilience to further
OM input (say from a fish farm) and b) a longer recovery time from the same level of
additional OM input than a site that has a lower background OM supply?
Can we test this hypothesis by adapting the model? Essentially we need an input for OM
background at the beginning of the process. We then need to generate an additional base level
OM demand based on some function of background status using information from the

57
literature on the degradation rate of marine primary production as well as information on the
degradation rates of terrestrial detritus and some estimates of OM supply.
But to allow the maintenance of an impact that extends well beyond the farming period (as
observed) we need to generate a greater oxygen demand from our model than is taking place
currently. We can hypothesise that our original assumption that oxygen demand can be
estimated from carbon degradation was not supported by the model outcomes. Thus we need
to include processes where sulphide concentration increases in sediment and its subsequent
re-oxidation to sulphate causes an oxygen demand long after the original organic material has
been degraded.
8.2 RQ model description
This model was originally constructed in MS Excel and was later developed as stand-alone
computer software written in Delphi. At the outset it was called the RQ model – for
Respiratory Quotient6– as initial work used the RQ as an important model parameter.
The concept of the model is visualised in figure 13 although all of the compartments and
processes described are not modelled (but could be in a future version if thought necessary).
The model is constructed as a simple box model as a first approximation but could later be
elaborated into a more dynamical model with feedbacks between processes. Our initial
assumption is that while these feedbacks are likely to be very important in obtaining good
numerical fits between predictions and observations, they may be less important in estimating
the rank of sites in terms of recovery, given the scale of the OM input from fish farms
compared to most natural systems.
This model is not designed as a comprehensive biogeochemical model as we believe that at
present this would not be a practical option as a regulatory tool. However, we also hypothesis
that the fish farm sedimentary system may be so extreme (in terms of OM flux and
6 RQ = CO2 produced/ O2 consumed. Metabolism of different types of organic matter yield different values of RQ
as a consequence of their stoichiometry. In the context of marine sediments, where the production of carbon
dioxide (dissolved inorganic carbon) and the demand for oxygen may be decoupled for long periods through the
production of sulphide intermediates, the apparent RQ of the sediment system may vary in time where organic
matter supply is non-steady state. For example, we would expect RQ to be greater than unity under high OM
fluxes as much of the oxidising power comes from sulphate and the sulphides are to some extent stored in
sediments, whereas under later low OM fluxes (e.g. during recovery) we might expect an RQ of less than unity as
respiration is diminished but oxygen demand is still caused by the later oxidation of reduced intermediates like
sulphide.
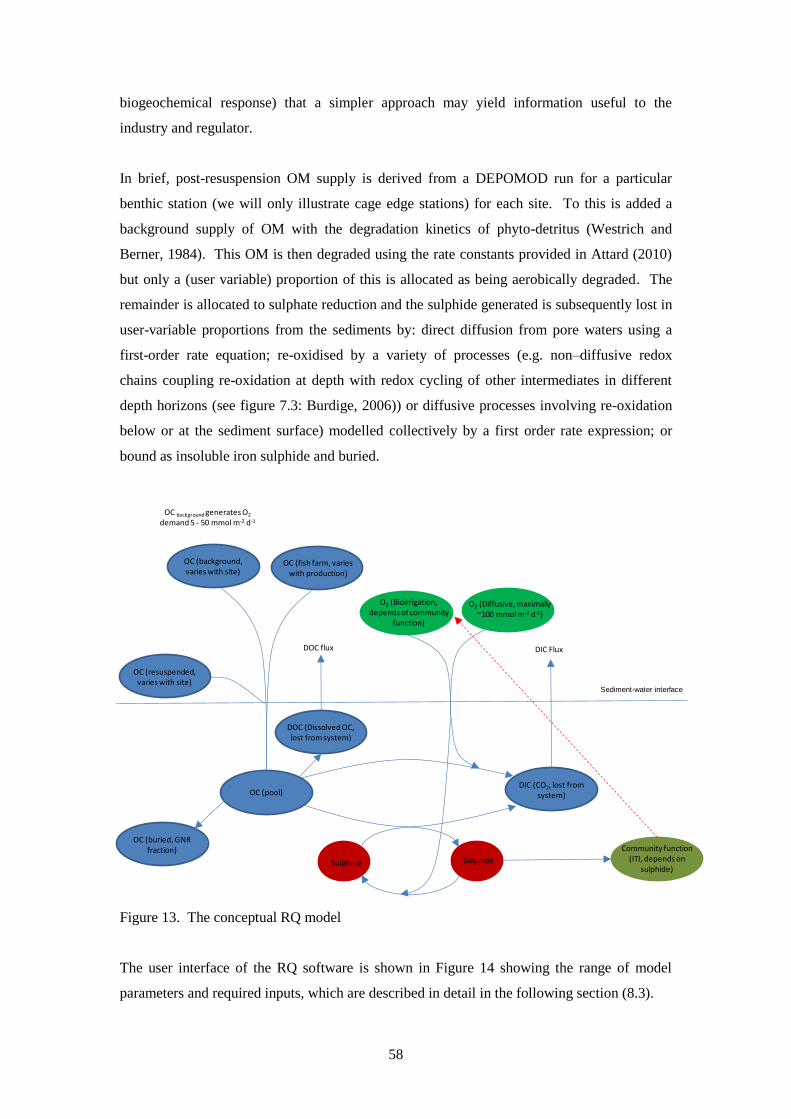
58
biogeochemical response) that a simpler approach may yield information useful to the
industry and regulator.
In brief, post-resuspension OM supply is derived from a DEPOMOD run for a particular
benthic station (we will only illustrate cage edge stations) for each site. To this is added a
background supply of OM with the degradation kinetics of phyto-detritus (Westrich and
Berner, 1984). This OM is then degraded using the rate constants provided in Attard (2010)
but only a (user variable) proportion of this is allocated as being aerobically degraded. The
remainder is allocated to sulphate reduction and the sulphide generated is subsequently lost in
user-variable proportions from the sediments by: direct diffusion from pore waters using a
first-order rate equation; re-oxidised by a variety of processes (e.g. non–diffusive redox
chains coupling re-oxidation at depth with redox cycling of other intermediates in different
depth horizons (see figure 7.3: Burdige, 2006)) or diffusive processes involving re-oxidation
below or at the sediment surface) modelled collectively by a first order rate expression; or
bound as insoluble iron sulphide and buried.
Sediment-water interface
OC (background, varies with site)
OC (fish farm, varies with production)
OC (pool)
OC (buried, GNR fraction)
OC (resuspended, varies with site)
DIC (CO2, lost from system)
O2 (Diffusive, maximally ~100 mmol m-2 d-1)
O2 (Bioirrigation, depends of community
function)
DOC (Dissolved OC, lost from system)
DOC flux
Sulphate Sulphide
DIC Flux
OC background generates O2
demand 5 - 50 mmol m-2 d-1
Community function (ITI, depends on
sulphide)
Figure 13. The conceptual RQ model
The user interface of the RQ software is shown in Figure 14 showing the range of model
parameters and required inputs, which are described in detail in the following section (8.3).

59
Figure 14 The RQ model user interface
8.3 Model processes and parameters
The architecture of the model allows the creation of an initialisation file (*.ini) that captures
the range of parameter values set through the interface allowing for stepwise varying of
individual parameters without tedious re-entry of unvaried parameter values. In practice, we
have developed site specific *.ini files for each of the five sites (section 4.1) for further
convenience.
Farm box
In this box, a menu allows the direct input of site-specific monthly feed inputs or biomass
data with specific feeding rates - we had access to monthly feed inputs and used these
throughout.
Feed box
This box allows the specification of percentages of feed water content, feed digestibility and
wastage for which we used values of 4, 85 and 3% respectively in all simulations. A
digestibility of 85% has been used in previous work (Cromey et al, 2002a) and comes from
manufacturer‟s information. Factors related to differences in feed formulation between the
sites together with differences in feeding rate and disease status may all affect digestibility but
we had no information on this and did not test the model sensitivity to this parameter.

60
Similarly the proportion of waste feed likely varies between sites and seasons but is very
difficult to estimate and so a constant value was chosen – the authors accept that this value is
likely to be important, as feed contains more reactive organic matter than faeces, but there is
presently no good method to estimate this over a farm cycle unless farms are using
sophisticated feeders with waste feed trapping and recycling. In the present comparative
exercise, the difference in impact between farms is more important than the absolute values
obtained and thus differences in feed wastage rates between farms is more important than the
absolute value (3%) chosen to represent an average. The water content was measured from
feed samples (Attard, 2010).
Background sedimentation
As mentioned above, background sedimentation may influence the recovery rate of sites.
Also, where background rates are high, the natural benthic assemblage may be more tolerant
of organic enrichment from the farm. In addition, measures of recovery which compare
reference values with impact stations, i.e. relative indices, may yield lower impact values that
would be obtained from consideration of impacted stations in isolation. Natural organic
matter accumulation rates (i.e. sedimentation minus resuspension) are hard to measure and
relatively few measurements have been made in Scottish fjordic systems. We attempted to
rank the sites in terms of sediment accumulation by using organic matter values for stations
close to the farms which were unlikely to experience a large farm input i.e. the 50m stations
(65m in the case of Fishnish B, Table 7). We did not use the reference values as these were
clearly unrepresentative of the farm location in at least one site (Oban Bay). There are few
published data on primary production in Scottish sea lochs. A study in Loch Linnhe has
shown annually integrated carbon fixation rates of between 78 and 199 g m-2
yr-1
(Rees et al.,
1995) which, assuming a carbon content of around 25% (Platt and Irwin, 1973) yields an OM
flux rate of 312 to 796 g m-2
yr-1
. Accordingly, we used a value of 800 g m-2
yr-1
as an
estimate of the supply rate of background OM to the site with the most OM rich 50 m station
and scaled the other sites linearly (Table 33). As the % OM in sediments reflects both the
total carbon supply rate and the non-reactive carbon content, this method of estimation of
relative supply depends on the quality of supply being approximately equal between sites, on
which we have no information, although it is probably that sites near river inputs have
considerable contributions of terrestrial as well as marine OM. At present there is no option to
vary background OM by month although some data exist that would enable this refinement to
be implemented (Rees et al., 1995).
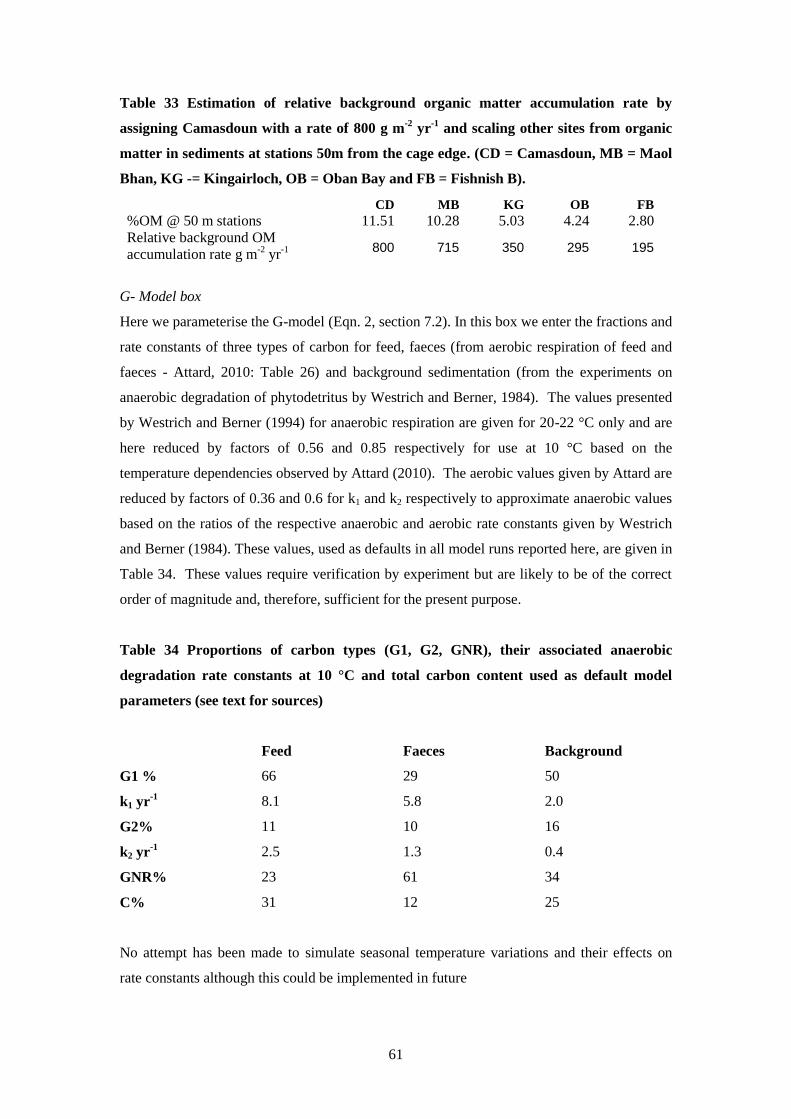
61
Table 33 Estimation of relative background organic matter accumulation rate by
assigning Camasdoun with a rate of 800 g m-2
yr-1
and scaling other sites from organic
matter in sediments at stations 50m from the cage edge. (CD = Camasdoun, MB = Maol
Bhan, KG -= Kingairloch, OB = Oban Bay and FB = Fishnish B).
CD MB KG OB FB
%OM @ 50 m stations 11.51 10.28 5.03 4.24 2.80
Relative background OM
accumulation rate g m-2
yr-1
800 715 350 295 195
G- Model box
Here we parameterise the G-model (Eqn. 2, section 7.2). In this box we enter the fractions and
rate constants of three types of carbon for feed, faeces (from aerobic respiration of feed and
faeces - Attard, 2010: Table 26) and background sedimentation (from the experiments on
anaerobic degradation of phytodetritus by Westrich and Berner, 1984). The values presented
by Westrich and Berner (1994) for anaerobic respiration are given for 20-22 °C only and are
here reduced by factors of 0.56 and 0.85 respectively for use at 10 °C based on the
temperature dependencies observed by Attard (2010). The aerobic values given by Attard are
reduced by factors of 0.36 and 0.6 for k1 and k2 respectively to approximate anaerobic values
based on the ratios of the respective anaerobic and aerobic rate constants given by Westrich
and Berner (1984). These values, used as defaults in all model runs reported here, are given in
Table 34. These values require verification by experiment but are likely to be of the correct
order of magnitude and, therefore, sufficient for the present purpose.
Table 34 Proportions of carbon types (G1, G2, GNR), their associated anaerobic
degradation rate constants at 10 °C and total carbon content used as default model
parameters (see text for sources)
Feed Faeces Background
G1 % 66 29 50
k1 yr-1
8.1 5.8 2.0
G2% 11 10 16
k2 yr-1
2.5 1.3 0.4
GNR% 23 61 34
C% 31 12 25
No attempt has been made to simulate seasonal temperature variations and their effects on
rate constants although this could be implemented in future
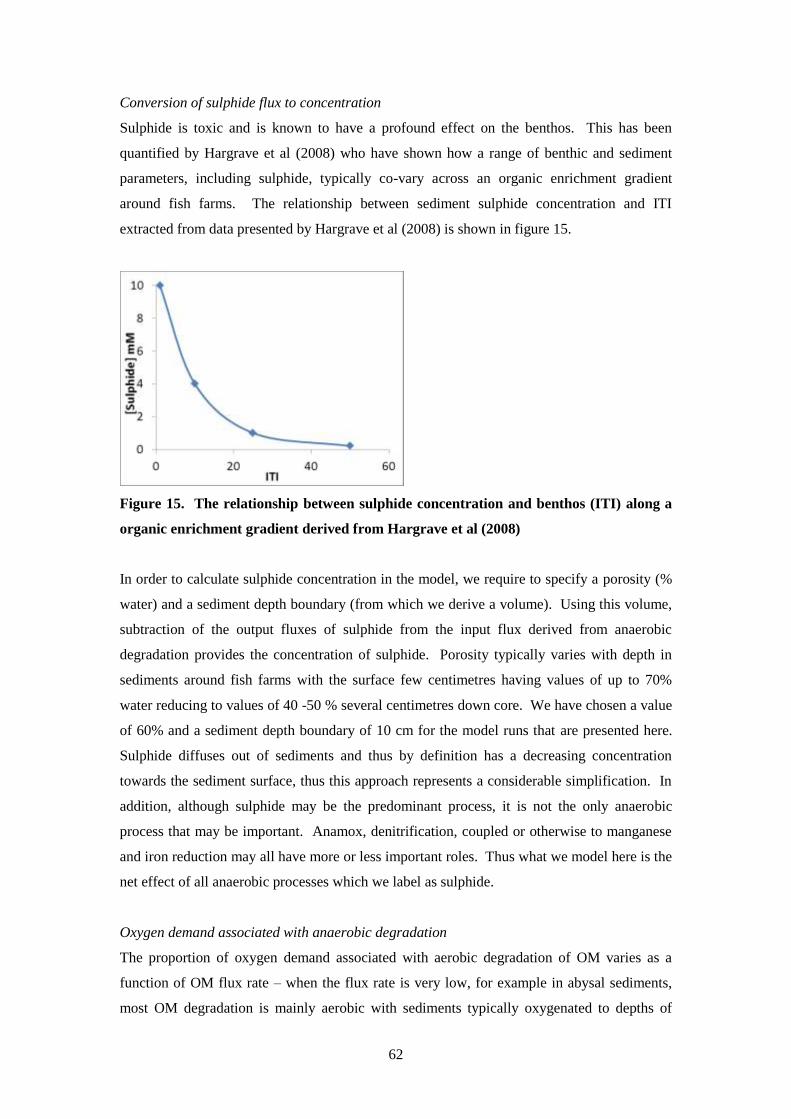
62
Conversion of sulphide flux to concentration
Sulphide is toxic and is known to have a profound effect on the benthos. This has been
quantified by Hargrave et al (2008) who have shown how a range of benthic and sediment
parameters, including sulphide, typically co-vary across an organic enrichment gradient
around fish farms. The relationship between sediment sulphide concentration and ITI
extracted from data presented by Hargrave et al (2008) is shown in figure 15.
Figure 15. The relationship between sulphide concentration and benthos (ITI) along a
organic enrichment gradient derived from Hargrave et al (2008)
In order to calculate sulphide concentration in the model, we require to specify a porosity (%
water) and a sediment depth boundary (from which we derive a volume). Using this volume,
subtraction of the output fluxes of sulphide from the input flux derived from anaerobic
degradation provides the concentration of sulphide. Porosity typically varies with depth in
sediments around fish farms with the surface few centimetres having values of up to 70%
water reducing to values of 40 -50 % several centimetres down core. We have chosen a value
of 60% and a sediment depth boundary of 10 cm for the model runs that are presented here.
Sulphide diffuses out of sediments and thus by definition has a decreasing concentration
towards the sediment surface, thus this approach represents a considerable simplification. In
addition, although sulphide may be the predominant process, it is not the only anaerobic
process that may be important. Anamox, denitrification, coupled or otherwise to manganese
and iron reduction may all have more or less important roles. Thus what we model here is the
net effect of all anaerobic processes which we label as sulphide.
Oxygen demand associated with anaerobic degradation
The proportion of oxygen demand associated with aerobic degradation of OM varies as a
function of OM flux rate – when the flux rate is very low, for example in abysal sediments,
most OM degradation is mainly aerobic with sediments typically oxygenated to depths of

63
several centimetres. In contrast, in coastal system with higher flux rates of OM, a large
proportion of the OM is metabolised anaerobically of which the majority is by sulphate
reduction (Holmer and Kristensen, 1992; Jorgensen, 1977). However, as we are dealing
primarily with highly reducing fish farm sediments, we have decided to simply allocate a
small proportion (10%) of the OM flux to aerobic degradation. This allows computational
simplicity but a more dynamic approach could be implemented in the future if warranted,
where the proportion of the OM oxidised by oxygen is dependent on the oxygen demand
caused by both aerobic and anaerobic processes. This would allow the balance between
aerobic and anaerobic processes to vary according to the changing state of the seabed during
the farming and recovery cycle.
The proportion allocated to aerobic oxidation is degraded over time using the multi-G model
with the parameterisation described earlier. At present, the model does not allow for different
rate constants between the aerobic fraction and the anaerobic fraction but this could be added
in a later version. The consequence of this is likely to be relatively minor as the proportion of
the carbon that is degraded aerobically is small and the difference in rate constants is only a
factor of 2-3. The aerobically degraded fraction causes an appropriate oxygen demand. The
larger anaerobic fraction is degraded using the same rate constants to produce a sulphide pool
which is then reoxidised according to the first order rate constant referred to as the diffusion
coefficient as it is likely to correspond to the net effect of several diffusion limited processes.
We have used a value of 2.8 month-1
in the results given here as this yielded plausible oxygen
demand and sulphide concentration results. This constant should be evaluated in a future
study.
Time
The time input box allows variation of the time (month) where feeding is started in the model
and the total duration of the model run. In order to display the information with respect to
recovery time, we adjusted the feeding start month for each farm so that they all ceased
feeding on the same month. We set this feeding end time at 80 months as this provided
sufficient time for the model to evolve to a steady state before feed input comenced and we
typically allowed the model to run for a total of 180 months (15 years).
Convert to mmol/m2/d
The model works on a whole farm scale. However, results for particular stations allow
comparison with benthic and sediment field data and the check box instructs this. At an early
stage in the modelling process we decided that to include the RQ model within DEPOMOD
for all grid points would result in enormous computational times that would both hamper

64
development and make the model practically unusable. We solve this by selecting stations for
which we have information from DEPOMOD and run the RQ model in stand-alone mode
(run times < 1s).
By calculating a station specific factor, we can estimate the flux at the near cage station each
month using the husbandry data specified. The factor can be calculated from DEPOMOD
predictions as follows:
Factor = Cflux (g C m-2
month-1
) / Cdischarged (g C month-1
)
where Cflux is the DEPOMOD prediction of flux at a station and Cdischarged is the amount
of carbon (waste feed and faeces) discharged from the farm each month calculated in a
spreadsheet. As Cflux is directly proportional to Cdischarged, Factor is a constant. Factor is
different for each station, as Cflux varies between stations. We have concentrated our effort
on cage edge stations for each of the five study sites (Table 5).
Table 35 Factors used to convert whole grid calculations to cage edge stations in mmol
m-2
d-1
Site Factor
Oban Bay 0.000120
Camasdoun 0.000029
Fishnish B 0.000024
Kingairloch 0.00053
Maol Bhan 0.000050
Loss of sulphide by diffusion
Sulphide is lost from sediments by diffusion. However, a considerable proportion of this may
be reoxidised at the sediment surface (creating an oxygen demand) and recycled as sulphate
back into the sediments. In this box we allow a first order loss of sulphide from the system
which does not contribute to oxygen demand. Similar to the issues relating to the balance
between aerobic and anaerobic metabolism discussed earlier, the amount of sulphide lost from
the system is a function of the sediment sulphide concentration as when the concentration is
small most sulphide is likely to be deoxidised at the sediment surface and recycled, but when
the it high the flux rate will become higher than the rate of reoxidation at the surface and
more of it will be able to escape to the water column. However, determining and modelling
this function is beyond the scope of this project and so the present approach is recognised as a
potentially large compromise which is likely to be more important during the recovery phase

65
when sulphide concentrations decrease. This may cause an underestimate of the oxygen
demand from sulphide reoxidation during this phase. The model is parameterised as a simple
first order rate equation with a rate constant and a reaction constant. At –present we have left
the reaction constant as unity in all runs but have changed the rate constant – on which we
presently have no information – in various model runs. Increasing the rate constant
effectively reduces the amount of sulphide available for re-oxidation and therefore the
modelled sediment sulphide concentration and consequently also reduces the oxygen demand
from the sediment. We have used a rate constant of 0.14 month-1
for the results presented in
this report.
DOC production from sediment
Under conditions of high OM flux, dissolved organic carbon builds up in sediments as the
products of initial degradation reactions on larger molecules at a rate faster than the rate at
which they are further degraded in subsequent degradation steps, leading to a flux of DOC
from the sediments. Similar to the argument above for sulphide diffusion, the rate of this
process is likely to be lower during low flux periods during the recovery phase. A recent
Faroes study (A Nordi et al., In press) has shown that over a period of several months
approximately 14% of the carbon flux from a fish farm was lost from the sediments as DOC.
We have used this figure for waste from the farm but have used a figure of 0% for the
background carbon supply on the basis that this is likely to make an important contribution to
the model outputs only when the total carbon flux is small and when DOC losses are likely to
be proportionally small.
Iron sulphide from sulphide
The sulphur cycle in sediments ultimately leads to the burial of metal sulphides, particularly
of iron (FeS2, pyrite). The sulphur cycle is complex with intermediate species such as FeS
which can either further react to pyrite or be reoxidised. According to Jorgensen (1977) about
11% of the total sulphate reduction can be “lost” to FeS and FeS2 and we have used this figure
in the model. However, Jorgensen was working in coastal sediments rather than under a fish
farm and it is most likely that this is not a good estimate. Iron sulphide formation may in
some circumstances be limited by the availability of highly reactive iron in the sediment and
these may apply in a fish farm sediments where sulphide is being cycled very rapidly. Until
further observational information is available we assume an 11% loss of sulphur to iron
sulphides and do not allow these to be reoxidised.
Operating the model
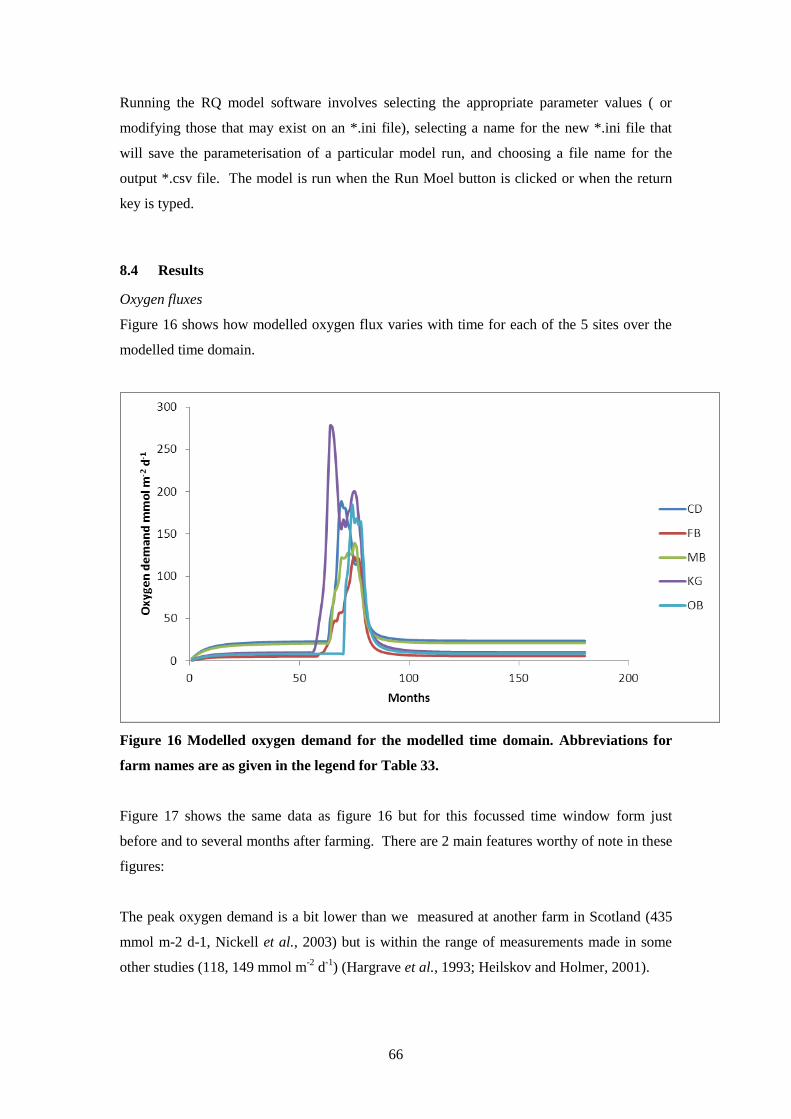
66
Running the RQ model software involves selecting the appropriate parameter values ( or
modifying those that may exist on an *.ini file), selecting a name for the new *.ini file that
will save the parameterisation of a particular model run, and choosing a file name for the
output *.csv file. The model is run when the Run Moel button is clicked or when the return
key is typed.
8.4 Results
Oxygen fluxes
Figure 16 shows how modelled oxygen flux varies with time for each of the 5 sites over the
modelled time domain.
Figure 16 Modelled oxygen demand for the modelled time domain. Abbreviations for
farm names are as given in the legend for Table 33.
Figure 17 shows the same data as figure 16 but for this focussed time window form just
before and to several months after farming. There are 2 main features worthy of note in these
figures:
The peak oxygen demand is a bit lower than we measured at another farm in Scotland (435
mmol m-2 d-1, Nickell et al., 2003) but is within the range of measurements made in some
other studies (118, 149 mmol m-2
d-1
) (Hargrave et al., 1993; Heilskov and Holmer, 2001).
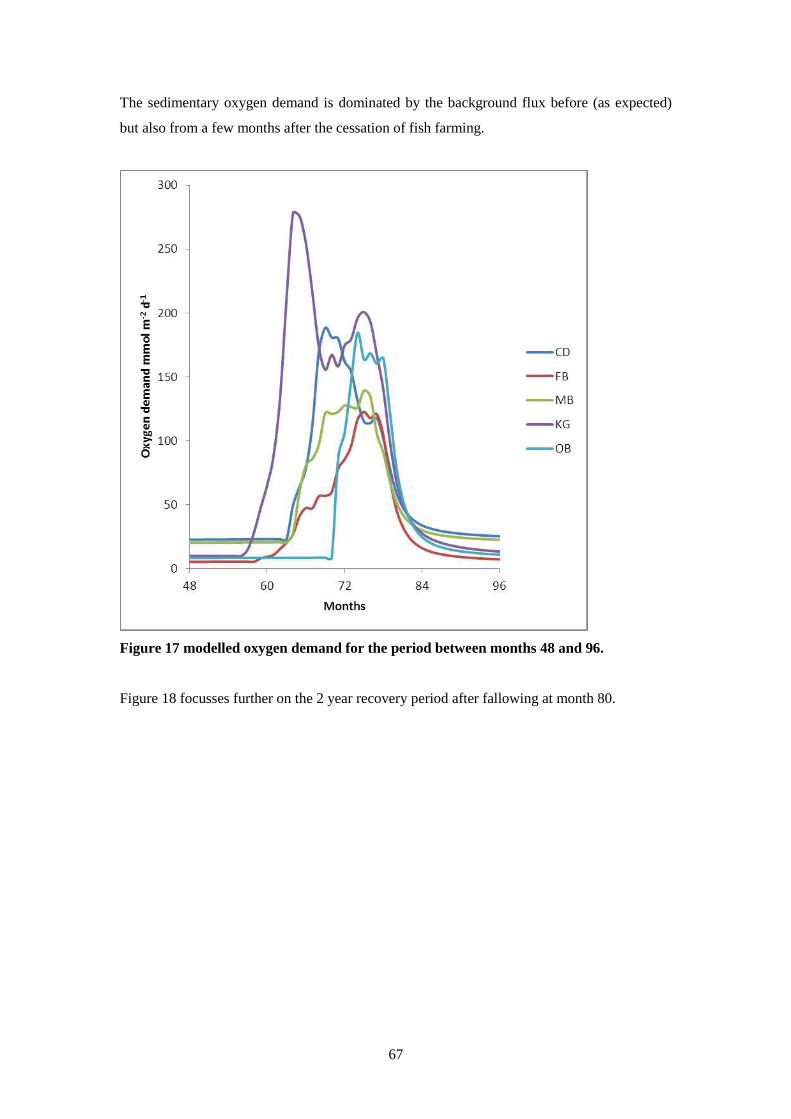
67
The sedimentary oxygen demand is dominated by the background flux before (as expected)
but also from a few months after the cessation of fish farming.
Figure 17 modelled oxygen demand for the period between months 48 and 96.
Figure 18 focusses further on the 2 year recovery period after fallowing at month 80.
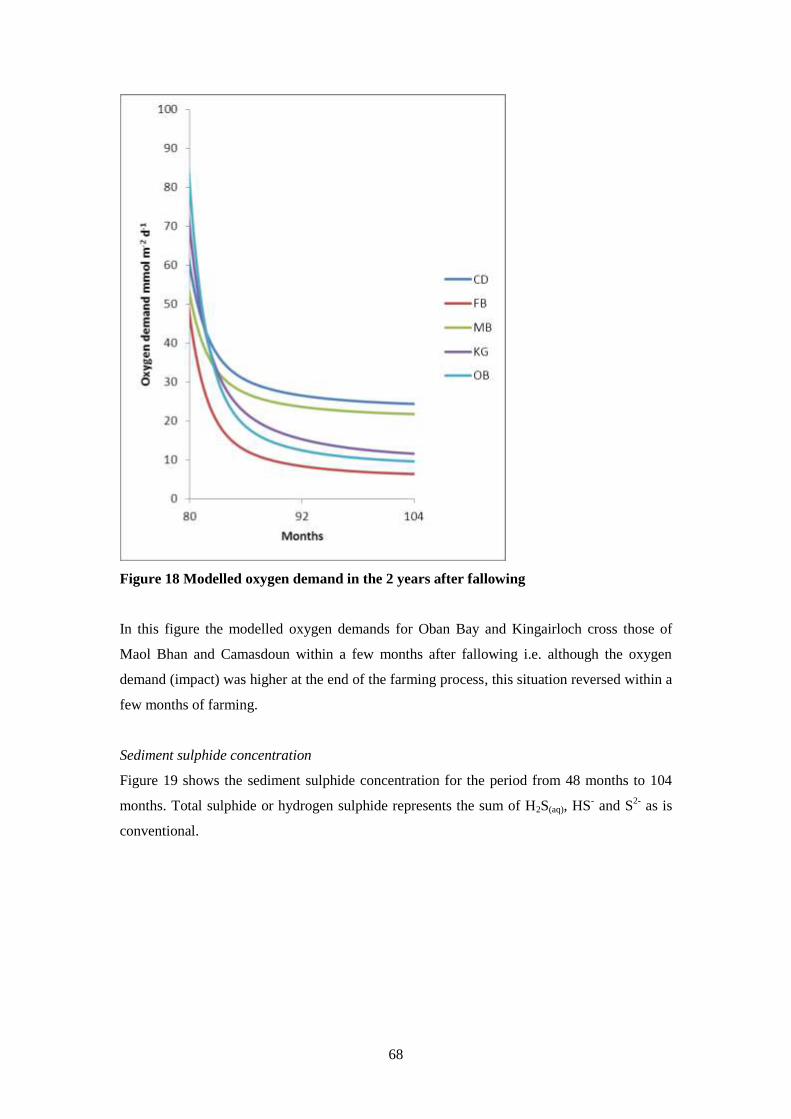
68
Figure 18 Modelled oxygen demand in the 2 years after fallowing
In this figure the modelled oxygen demands for Oban Bay and Kingairloch cross those of
Maol Bhan and Camasdoun within a few months after fallowing i.e. although the oxygen
demand (impact) was higher at the end of the farming process, this situation reversed within a
few months of farming.
Sediment sulphide concentration
Figure 19 shows the sediment sulphide concentration for the period from 48 months to 104
months. Total sulphide or hydrogen sulphide represents the sum of H2S(aq), HS- and S
2- as is
conventional.
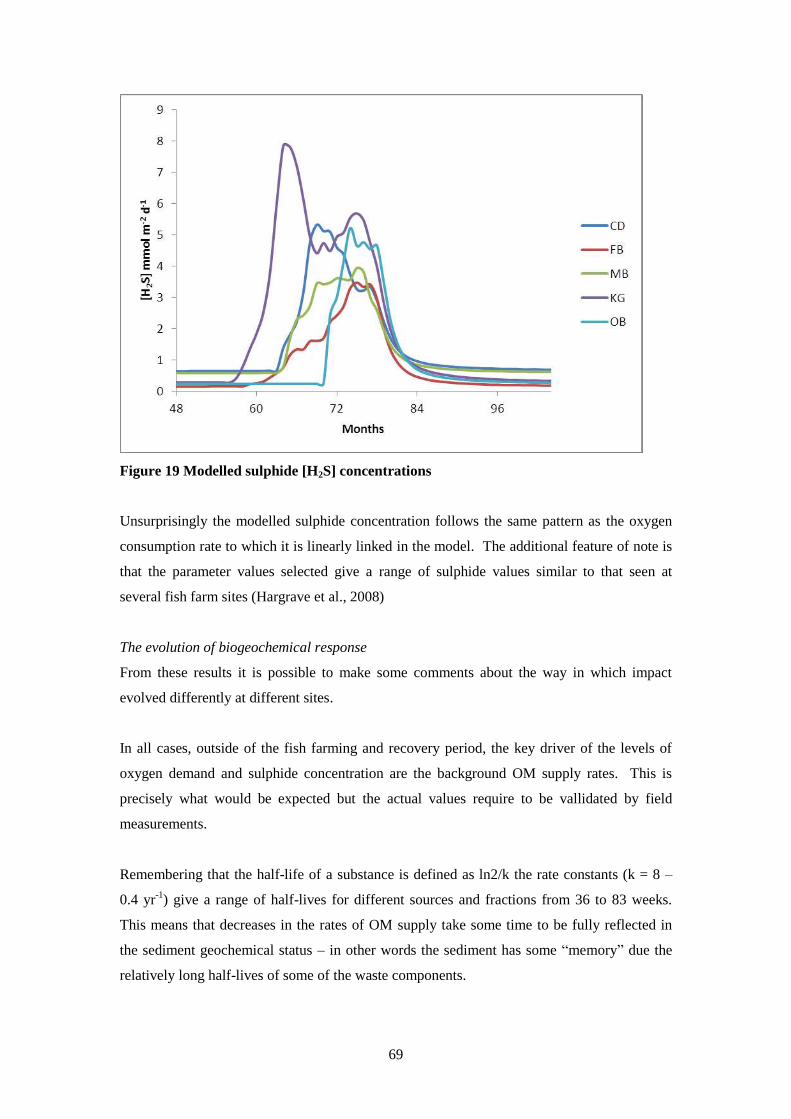
69
Figure 19 Modelled sulphide [H2S] concentrations
Unsurprisingly the modelled sulphide concentration follows the same pattern as the oxygen
consumption rate to which it is linearly linked in the model. The additional feature of note is
that the parameter values selected give a range of sulphide values similar to that seen at
several fish farm sites (Hargrave et al., 2008)
The evolution of biogeochemical response
From these results it is possible to make some comments about the way in which impact
evolved differently at different sites.
In all cases, outside of the fish farming and recovery period, the key driver of the levels of
oxygen demand and sulphide concentration are the background OM supply rates. This is
precisely what would be expected but the actual values require to be vallidated by field
measurements.
Remembering that the half-life of a substance is defined as ln2/k the rate constants (k = 8 –
0.4 yr-1
) give a range of half-lives for different sources and fractions from 36 to 83 weeks.
This means that decreases in the rates of OM supply take some time to be fully reflected in
the sediment geochemical status – in other words the sediment has some “memory” due the
relatively long half-lives of some of the waste components.
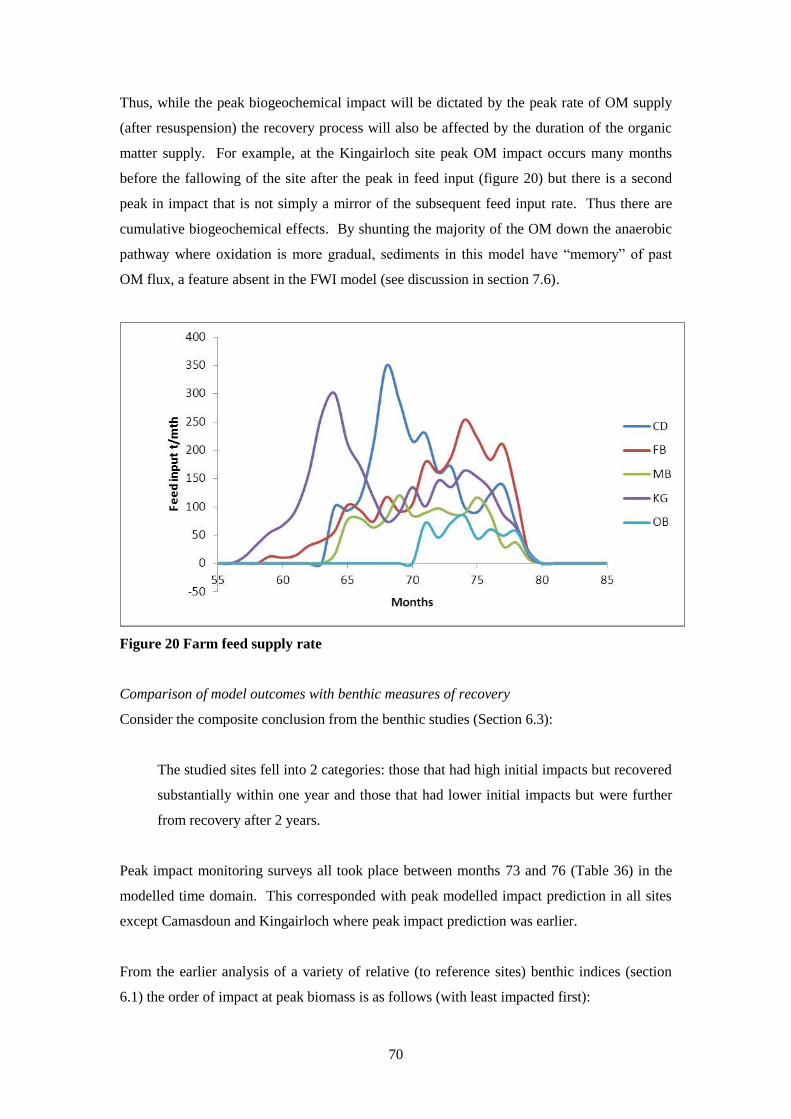
70
Thus, while the peak biogeochemical impact will be dictated by the peak rate of OM supply
(after resuspension) the recovery process will also be affected by the duration of the organic
matter supply. For example, at the Kingairloch site peak OM impact occurs many months
before the fallowing of the site after the peak in feed input (figure 20) but there is a second
peak in impact that is not simply a mirror of the subsequent feed input rate. Thus there are
cumulative biogeochemical effects. By shunting the majority of the OM down the anaerobic
pathway where oxidation is more gradual, sediments in this model have “memory” of past
OM flux, a feature absent in the FWI model (see discussion in section 7.6).
Figure 20 Farm feed supply rate
Comparison of model outcomes with benthic measures of recovery
Consider the composite conclusion from the benthic studies (Section 6.3):
The studied sites fell into 2 categories: those that had high initial impacts but recovered
substantially within one year and those that had lower initial impacts but were further
from recovery after 2 years.
Peak impact monitoring surveys all took place between months 73 and 76 (Table 36) in the
modelled time domain. This corresponded with peak modelled impact prediction in all sites
except Camasdoun and Kingairloch where peak impact prediction was earlier.
From the earlier analysis of a variety of relative (to reference sites) benthic indices (section
6.1) the order of impact at peak biomass is as follows (with least impacted first):
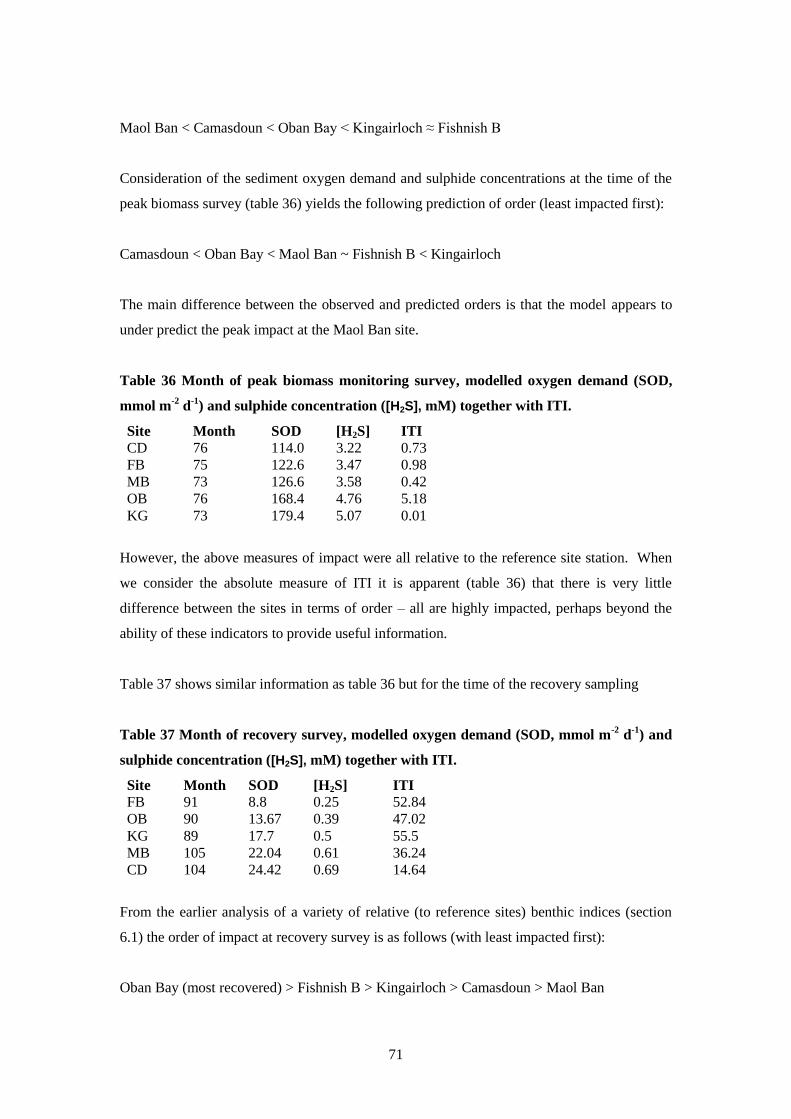
71
Maol Ban < Camasdoun < Oban Bay < Kingairloch ≈ Fishnish B
Consideration of the sediment oxygen demand and sulphide concentrations at the time of the
peak biomass survey (table 36) yields the following prediction of order (least impacted first):
Camasdoun < Oban Bay < Maol Ban ~ Fishnish B < Kingairloch
The main difference between the observed and predicted orders is that the model appears to
under predict the peak impact at the Maol Ban site.
Table 36 Month of peak biomass monitoring survey, modelled oxygen demand (SOD,
mmol m-2
d-1
) and sulphide concentration ([H2S], mM) together with ITI.
Site Month SOD [H2S] ITI
CD 76 114.0 3.22 0.73
FB 75 122.6 3.47 0.98
MB 73 126.6 3.58 0.42
OB 76 168.4 4.76 5.18
KG 73 179.4 5.07 0.01
However, the above measures of impact were all relative to the reference site station. When
we consider the absolute measure of ITI it is apparent (table 36) that there is very little
difference between the sites in terms of order – all are highly impacted, perhaps beyond the
ability of these indicators to provide useful information.
Table 37 shows similar information as table 36 but for the time of the recovery sampling
Table 37 Month of recovery survey, modelled oxygen demand (SOD, mmol m-2
d-1
) and
sulphide concentration ([H2S], mM) together with ITI.
Site Month SOD [H2S] ITI
FB 91 8.8 0.25 52.84
OB 90 13.67 0.39 47.02
KG 89 17.7 0.5 55.5
MB 105 22.04 0.61 36.24
CD 104 24.42 0.69 14.64
From the earlier analysis of a variety of relative (to reference sites) benthic indices (section
6.1) the order of impact at recovery survey is as follows (with least impacted first):
Oban Bay (most recovered) > Fishnish B > Kingairloch > Camasdoun > Maol Ban
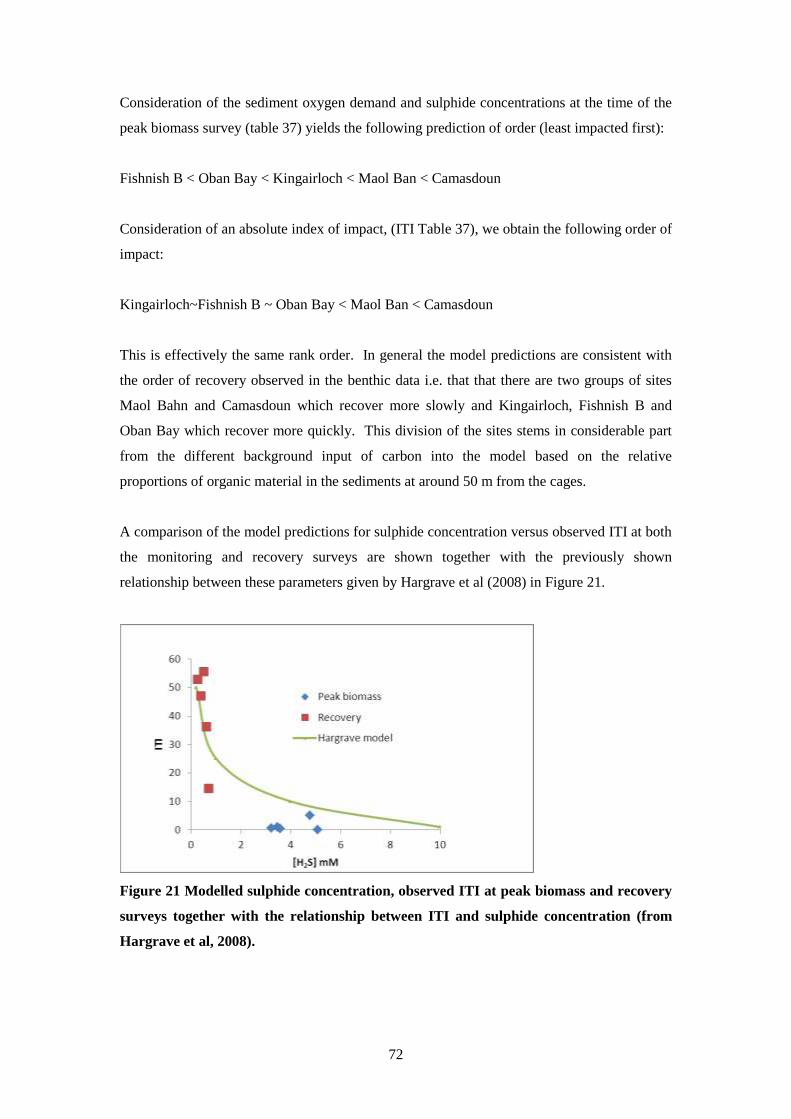
72
Consideration of the sediment oxygen demand and sulphide concentrations at the time of the
peak biomass survey (table 37) yields the following prediction of order (least impacted first):
Fishnish B < Oban Bay < Kingairloch < Maol Ban < Camasdoun
Consideration of an absolute index of impact, (ITI Table 37), we obtain the following order of
impact:
Kingairloch~Fishnish B ~ Oban Bay < Maol Ban < Camasdoun
This is effectively the same rank order. In general the model predictions are consistent with
the order of recovery observed in the benthic data i.e. that that there are two groups of sites
Maol Bahn and Camasdoun which recover more slowly and Kingairloch, Fishnish B and
Oban Bay which recover more quickly. This division of the sites stems in considerable part
from the different background input of carbon into the model based on the relative
proportions of organic material in the sediments at around 50 m from the cages.
A comparison of the model predictions for sulphide concentration versus observed ITI at both
the monitoring and recovery surveys are shown together with the previously shown
relationship between these parameters given by Hargrave et al (2008) in Figure 21.
Figure 21 Modelled sulphide concentration, observed ITI at peak biomass and recovery
surveys together with the relationship between ITI and sulphide concentration (from
Hargrave et al, 2008).

73
This presents a reasonable fit between the model predictions and the Hargrave model. It may
be possible to improve this fit by varying model parameters but it would be best to first have
access to the data that Hargrave used to produce this relationship or to collect new data before
optimising the parameterisation.
8.5 Discussion and conclusions
We have produced a simple box model (RQ) that can account for the major features of the
benthic data obtained at the five study site. This model is more appropriate for considering
recovery processes than the FWI approach attempted earlier (section 7). This is because
outside of the period of high oxygen demand when farm biomass is high, oxygen supply
issues are unlikely to be the factor which determines community structure. In considering
future work aimed at trying to improve our understanding of the processes that operate
throughout the faming cycle, it may be useful to combine the Findlay-Watling approach with
the RQ model.
The RQ model could be made more sophisticated and take into account seasonal effects by
including the facility to vary the input of OM by month and to introduce temperature
dependency in the rate constants. In addition we could allow different rate constants for
aerobic and anaerobic degradation. In addition, the degradation rate of so called non-reactive
components should be further investigated.
The RQ model should be tested with other datasets. It is possible that some of the
assumptions made are too drastic and a more sophisticated numerical approach is required.
Further work on the sulphur cycle would be extremely useful as this undoubtedly is a key
process during recovery.
In terms of practical application, the main conclusion from this work is that the background
input of organic carbon is a site specific factor important to recovery processes. We have
used the amount of organic material in sediments as a proxy for the background supply rate.
This has some merit as the organic content is likely to be largely made up of unreactive
carbon fractions but the accumulation of these unreactive fractions is a function of
background OM supply but further validation is required.
In simple terms this work supports the idea that recovery rate will be strongly influenced by
background OM supply, and that the higher the level of natural OM in sediments the longer
the likely recovery time.

74
9 Acknowledgements
We are very grateful to Sally Davies of Scottish Seafarms and to Chris Read of Marine
Harvest for supporting this project with data from the farms studied and for facilitating access
to the sites. We are very grateful to the fish farmers on the 5 sites studied for accommodating
our fieldwork and providing sampling vessels and other logistical support. We also gratefully
acknowledge the contributions of our sub-contractors, Dr Kenneth Boyd at ERI, North
Highland College, Thurso and Dr Richard Corner at the University of Stirling.

75
10 References
A Nordi, G., Glud, R.N., Gaard, E. and Simonsen, K. In press. Environmental impacts of coastal fish
farming: carbon and nitrogen budgets for trout farming in Kaldbaksfjørður (Faroe Islands).
Marine Ecology Progress Series.
Arthur, R., Done, T.J. and Marsh, H. 2005. Benthic recovery four years after an El Nino-induced coral
mass mortality in the Lakshadweep atolls. Current Science. 89. 694-699.
Arthur, R., Done, T.J., Marsh, H. and Harriott, V. 2006. Local processes strongly influence post-
bleaching benthic recovery in the Lakshadweep Islands. Coral Reefs. 25. 427-440.
Attard, K.M., 2010. The degradation kinetics of organic materials in the marine environment - A case
study using materials associated with sites of finfish aquaculture,The degradation kinetics of
organic materials in the marine environment - A case study using materials associated with
sites of finfish aquaculture. UHI Millenium Institute, Scottish Association for Marine Science,
pp. 30.
Black, K.D., Cromey, C.J., Dale, A. and Nickell, T.D., 2009 Modelling benthic effects of large salmon
cage farms in Scotland. Report to the Scottish Salmon Producers Organisation. 97 pp.
Black, K.D., Blackstock, J., Cromey, C.J., Duncan, J., Gee, M., Gillibrand, P., Needham, H., Nickell,
T.D., Pearson, T.H., Powell, H., Sammes, P., Somerfield, P., Walsham, P., Webster, L. and
Willis, K., 2005 The ecological effects of sea lice treatment agents. 286 pp.
http://www.sams.ac.uk/research/departments/ecology/past-projects/researchproject.2007-03-
09.9693635791/
Borja, A., Muxika, I. and Franco, J. 2006. Long-term recovery of soft-bottom benthos following urban
and industrial sewage treatment in the Nervion estuary - (southern Bay of Biscay). Marine
Ecology Progress Series. 313. 43-55.
Brooks, K.M. and Mahnken, C.V.W. 2003. Interactions of Atlantic salmon in the Pacific northwest
environment II. Organic wastes. Fisheries Research. 62. 255-293.
Brooks, K.M., Stierns, A.R. and Backman, C. 2004. Seven year remediation study at the Carrie Bay
Atlantic salmon (Salmo salar) farm in the Broughton Archipelago, British Columbia, Canada.
Aquaculture. 239. 81-123.
Burdige, D.J., 2006. Geochemistry of marine sediments. Princeton University Press, Woodstock, 609
pages pp.
Burt, J., Bartholomew, A. and Usseglio, P. 2008. Recovery of corals a decade after a bleaching event in
Dubai, United Arab Emirates. Marine Biology. 154. 27-36.
Clarke, K.R. and Gorley, R.N., 2006. PRIMER v6: User manual/tutorial. PRIMER-E, Plymouth, 192
pp.
Craft, C.B., Seneca, E.D. and Broome, S.W. 1991. LOSS ON IGNITION AND KJELDAHL
DIGESTION FOR ESTIMATING ORGANIC-CARBON AND TOTAL NITROGEN IN
ESTUARINE MARSH SOILS - CALIBRATION WITH DRY COMBUSTION. Estuaries.
14. 175-179.
Cromey, C.J., Nickell, T.D. and Black, K.D. 2002a. DEPOMOD - modelling the deposition and
biological effects of waste solids from marine cage farms. Aquaculture. 214. 211-239.
Cromey, C.J., Nickell, T.D., Black, K.D., Provost, P.G. and Griffiths, C.R. 2002b. Validation of a fish
farm waste resuspension model by use of a particulate tracer discharged from a point source in
a coastal environment. Estuaries. 25. 916-929.
Dean, R.J., Shimmield, T.M. and Black, K.D. 2007. Copper, zinc and cadmium in marine cage fish
farm sediments: an extensive survey. Environmental Pollution. 145. 84-95.
Findlay, R.H. and Watling, L. 1997. Prediction of benthic impact for salmon net-pens based on the
balance of benthic oxygen supply and demand. Marine Ecology Progress Series. 155. 147-
157.
Guerra-Garcia, J.M., Corzo, J. and Garcia-Gomez, J.C. 2003. Short-term benthic recolonization after
dredging in the harbour of Ceuta, North Africa. Marine Ecology-Pubblicazioni Della Stazione
Zoologica Di Napoli I. 24. 217-229.
Gutt, J., Starmans, A. and Dieckmann, G. 1996. Impact of iceberg scouring on polar benthic habitats.
Marine Ecology Progress Series. 137. 311-316.
Hall-Spencer, J., White, N., Gillespie, E., Gillham, K. and Foggo, A. 2006. Impact of fish farms on
maerl beds in strongly tidal areas. Marine Ecology Progress Series. 326. 1-9.
Hall-Spencer, J.M. and Moore, P.G. 2000. Scallop dredging has profound, long-term impacts on maerl
habitats. ICES Journal of Marine Science. 57. 1407-1415.
Hall, J.A. and Frid, C.L.J. 1995. Responses of estuarine benthic macrofauna in copper-contaminated
sediments to remediation of sediment quality. Marine Pollution Bulletin. 30. 694-700.

76
Hall, J.A. and Frid, C.L.J. 1997. Estuarine sediment remediation: Effects on benthic biodiversity.
Estuarine, Coastal and Shelf Science. 44. 55-61.
Hall, S.J. and Harding, M.J.C. 1997. Physical disturbance and marine benthic communities: The effects
of mechanical harvesting of cockles on non-target benthic infauna. Journal of Applied
Ecology. 34. 497-517.
Hansen, L.S. and Blackburn, T.H. 1992. EFFECT OF ALGAL BLOOM DEPOSITION ON
SEDIMENT RESPIRATION AND FLUXES. Marine Biology. 112. 147-152.
Hargrave, B.T., Holmer, M. and Newcombe, C.P. 2008. Towards a classification of organic enrichment
in marine sediments based on biogeochemical indicators. Marine Pollution Bulletin. 56. 810-
824.
Hargrave, B.T., Duplisea, D.E., Pfeiffer, E. and Wildish, D.J. 1993. Seasonal-changes in benthic fluxes
of dissolved-oxygen and ammonium associated with marine cultured Atlantic salmon. Marine
Ecology Progress Series. 96. 249-257.
Heilskov, A.C. and Holmer, M. 2001. Effects of benthic fauna on organic matter mineralization in fish-
farm sediments: importance of size and abundance. ICES Journal of Marine Science. 58. 427-
434.
Hiddink, J.G., Jennings, S. and Kaiser, M.J. 2006. Indicators of the ecological impact of bottom-trawl
disturbance on seabed communities. Ecosystems. 9. 1190-1199.
Hilton, M.J. and Hesp, P. 1996. Determining the Limits of Beach-Nearshore Sand Systems and the
Impact of Offshore Coastal Sand Mining. Journal of Coastal Research. 12. 496-519.
Holmer, M. and Kristensen, E. 1992. Impact of marine fish cage farming on metabolism and sulphate
reduction of underlying sediments. Marine Ecology Progress Series. 80. 191-201.
Holmer, M. and Heilskov, A.C. 2008. Distribution and bioturbation effects of the tropical alpheid
shrimp Alpheus macellarius in sediments impacted by milkfish farming. Estuarine, Coastal
and Shelf Science. 76. 657-667.
Johnson, L.J. and Frid, C.L.J. 1995. The recovery of benthic communities along the County Durham
coast after cessation of colliery spoil dumping. Marine Pollution Bulletin. 30. 215-220.
Jorgensen, B.B. 1977. Sulfur cycle of a coastal marine sediment (Limfjorden, Denmark). Limnology
and Oceanography. 22. 814-832.
Jørgensen, B.B. and Des Marais, D.J. 1990. The diffusive boundary layer of sediments: oxygen
microgradients over a microbial mat. Limnology and Oceanography. 35. 1343-1355.
Jusup, M., Klanjscek, J., Petricioli, D. and Legovic, T. 2009. Predicting aquaculture-derived benthic
organic enrichment: Model validation. Ecological Modelling. 220. 2407-2414.
Kaiser, M.J., Clarke, K.R., Hinz, H., Austen, M.C.V., Somerfield, P.J. and Karakassis, I. 2006. Global
analysis of response and recovery of benthic biota to fishing. Marine Ecology Progress Series.
311. 1-14.
Karakassis, I., Hatziyanni, E., Tsapakis, M. and Plaiti, W. 1999. Benthic recovery following cessation
of fish farming: a series of successes and catastrophes. Marine Ecology Progress Series. 184.
205-218.
Kim-Kang, H., Crouch, L.S., Bova, A., Robinson, R.A. and Wu, J. 2001. Determination of emamectin
residues in the tissues of Atlantic salmon (Salmo salar L.) using HPLC with fluorescence
detection. Journal of Agricultural and Food Chemistry. 49. 5294-5302.
Lardicci, C., Rossi, F. and Castelli, A. 1997. Analysis of macrozoobenthic community structure after
severe dystrophic crises in a Mediterranean coastal lagoon. Marine Pollution Bulletin. 34.
536-547.
Lim, H.S., Diaz, R.J., Hong, J.S. and Schaffner, L.C. 2006. Hypoxia and benthic community recovery
in Korean coastal waters. Marine Pollution Bulletin. 52. 1517-1526.
Loh, P.S., Reeves, A.D., Overnell, J., Harvey, S.M. and Miller, A.E.J. 2002. Assessment of terrigenous
organic carbon input to the total organic carbon in sediments from Scottish transitional waters
(sea lochs): methodology and preliminary results. Hydrology and Earth System Sciences. 6.
959-970.
Lu, L. and Wu, R.S.S. 2006. A field experimental study on recolonization and succession of
macrobenthic infauna in defaunated sediment contaminated with petroleum hydrocarbons.
Estuarine, Coastal and Shelf Science. 68. 627-634.
Lu, L. and Wu, R.S.S. 2007. A field experimental study on recolonization and succession of subtidal
macrobenthic community in sediment contaminated with industrial wastes. Marine Pollution
Bulletin. 54. 195-205.
Macleod, C.K., Crawford, C.M. and Moltschaniwskyj, N.A. 2004. Assessment of long term change in
sediment condition after organic enrichment: defining recovery. Marine Pollution Bulletin. 49.
79-88.

77
Macleod, C.K., Moltschaniwskyj, N.A. and Crawford, C.M. 2008. Ecological and functional changes
associated with long-term recovery from organic enrichment. Marine Ecology Progress Series.
365. 17-24.
Macleod, C.K., Moltschaniwskyj, N.A., Crawford, C.M. and Forbes, S.E. 2007. Biological recovery
from organic enrichment: some systems cope better than others. Marine Ecology Progress
Series. 342. 41-53.
Mayer, D.G. and Butler, D.G. 1993. Statistical validation. Ecological Modelling. 68. 21-32.
Mesplé, F., Troussellier, M., Casellas, C. and Legendre, P. 1996. Evaluation of simple statistical
criteria to qualify a simulation. Ecological Modelling. 88. 9-18.
Miller, K.L., Fernandes, T.F. and Read, P.A. 1999. The recovery of populations of dogwhelks suffering
from imposex in the Firth of Forth 1987-1997/98. Environmental Pollution. 106. 183-192.
Morrisey, D.J., Gibbs, M.M., Pickmere, S.E. and Cole, R.G. 2000. Predicting impacts and recovery of
marine-farm sites in Stewart Island, New Zealand, from the Findlay-Watling model.
Aquaculture. 185. 257-271.
Nickell, L.A., Black, K.D., Hughes, D.J., Overnell, J., Brand, T., Nickell, T.D., Breuer, E. and Harvey,
S.M. 2003. Bioturbation, sediment fluxes and benthic community structure around a salmon
cage farm in Loch Creran, Scotland. Journal of Experimental Marine Biology and Ecology.
285. 221-233.
Nickell, T.D. and Anderson, S.J., 1997 Environmental assessment of Loch Harport. Report prepared
for United Distillers. SAMS Internal Report No. 228. 49 pp.
Nickell, T.D., Black, K.D., Provost, P.G., Davies, I.M. and Pearson, T.H., 1995 Final report to the
Department of Trade and Industry/Scottish Salmon Growers Association on the Benthic
Recovery Programme, Progress Report No. 7. 73 pp.
Nickell, T.D., Black, K.D., Provost, P.G., Davies, I.M. and Pearson, T.H. 1998. The recovery of the
sea-bed after the cessation of fish farming: benthos and biogeochemistry. ICES CM. 1998
V:1.
Oreskes, N., Shrader-Frechette, K. and Belitz, K. 1994. Verification, validation, and confirmation of
numerical models in the earth sciences. Science. 263. 641-646.
Pearson, T.H. and Rosenberg, R. 1978. Macrobenthic succession in relation to organic enrichment and
pollution of the marine environment. Oceanography and Marine Biology Annual Review. 16.
229-311.
Pearson, T.H. and Black, K.D., 2001. The environmental impact of marine fish cage culture. In: Black,
K.D. (Ed.), Environmental Impacts of Aquaculture. Sheffield Academic Press, Sheffield, pp.
1-31.
Pereira, P.M.F., Black, K.D., McLusky, D.S. and Nickell, T.D. 2004. Recovery of sediments after
cessation of marine fish farm production. Aquaculture. 235. 315-330.
Platt, T. and Irwin, B. 1973. CALORIC CONTENT OF PHYTOPLANKTON. Limnology and
Oceanography. 18. 306-310.
Portilla, E. and Tett, P., 2007. Assessing goodness of fit for ECASA models (ECASA internal
paper),Assessing goodness of fit for ECASA models (ECASA internal paper). Napier
University, Edinburgh.
Powilleit, M., Kleine, J. and Leuchs, H. 2006. Impacts of experimental dredged material disposal on a
shallow, sublittoral macrofauna community in Mecklenburg Bay (western Baltic Sea). Marine
Pollution Bulletin. 52. 386-396.
Readman, J.W., Bartocci, J., Tolosa, I., Fowler, S.W., Oregioni, B. and Abdulraheem, M.Y. 1996.
Recovery of the coastal marine environment in the Gulf following the 1991 war-related oil
spills. Marine Pollution Bulletin. 32. 493-498.
Rees, A.P., Owens, N.J.P., Heath, M.R., Plummer, D.H. and Bellerby, R.S. 1995. Seasonal nitrogen
assimilation and carbon fixation in a fjordic sea loch. Journal of Plankton Research. 17. 1307-
1324.
Sanz-Lazaro, C. and Marin, A. 2006. Benthic recovery during open sea fish farming abatement in
Western Mediterranean, Spain. Marine Environmental Research. 62. 374-387.
SEPA, 1999. Emamectin benzoate use in marine fish farms: An environmental risk assessment. SEPA
Board Paper 65/99,Emamectin benzoate use in marine fish farms: An environmental risk
assessment. SEPA Board Paper 65/99, Stirling.
SEPA, 2005. Regulation and monitoring of marine cage fish farming in Scotland. Annex H. Methods
for modelling in-feed anti-parasitics and benthic effects,Regulation and monitoring of marine
cage fish farming in Scotland. Annex H. Methods for modelling in-feed anti-parasitics and
benthic effects Issue No. 2.3, 18 May 2005.

78
Shin, P.K.S., Lam, N.W.Y., Wu, R.S.S., Qian, P.Y. and Cheung, S.G. 2008. Spatio-temporal changes
of marine macrobenthic community in sub-tropical waters upon recovery from eutrophication.
I. Sediment quality and community structure. Marine Pollution Bulletin. 56. 282-296.
Smale, D.A., Barnes, D.K.A., Fraser, K.P.P. and Peck, L.S. 2008. Benthic community response to
iceberg scouring at an intensely disturbed shallow water site at Adelaide Island, Antarctica.
Marine Ecology Progress Series. 355. 85-94.
Soulsby, R.L., 1983. The bottom boundary layer of shelf seas. In: Johns, B. (Ed.), Physical
oceanography of coastal and shelf seas. Elsevier, London, pp. 189–266.
Stora, G., Arnoux, A. and Galas, M. 1995. Time and spatial dynamics of Mediterranean lagoon
macrobenthos during an exceptionally prolonged interruption of freshwater inputs.
Hydrobiologia. 301. 123-132.
Taylor, S.R. 1964. Abundance of chemical elements in the continental crust: a new table. Geochimica
et Cosmochimica Acta. 28. 1273-1285.
Teixido, N., Garrabou, J., Gutt, J. and Arntz, W.E. 2004. Recovery in Antarctic benthos after iceberg
disturbance: trends in benthic composition, abundance and growth forms. Marine Ecology
Progress Series. 278. 1-16.
Tuck, I.D., Hall, S.J., Robertson, M.R., Armstrong, E. and Basford, D.J. 1998. Effects of physical
trawling disturbance in a previously unfished sheltered Scottish sea loch. Marine Ecology
Progress Series. 162. 227-242.
Ueda, N., Tsutsumi, H., Yamada, M., Takeuchi, R. and Kido, K. 1994. Recovery of the marine bottom
environment of a Japanese bay. Marine Pollution Bulletin. 28. 676-682.
Westrich, J.T. and Berner, R.A. 1984. The role of sedimentary organic matter in bacterial sulphate
reduction: The G model tested. Limnology and Oceanography. 29. 236-249.
Wilber, D.H., Clarke, D.G. and Rees, S.I. 2007. Responses of benthic macroinvertebrates to thin-layer
disposal of dredged material in Mississippi Sound, USA. Marine Pollution Bulletin. 54. 42-52.
Word, J.Q., 1979 The Infaunal Trophic Index. Annual Report 1978. 19-39 pp.
Zenetos, A., Hatzianestis, J., Lantzouni, M., Simboura, M., Sklivagou, E. and Arvanitakis, G. 2004.
The Eurobulker oil spill: mid-term changes of some ecosystem indicators. Marine Pollution
Bulletin. 48. 122-131.
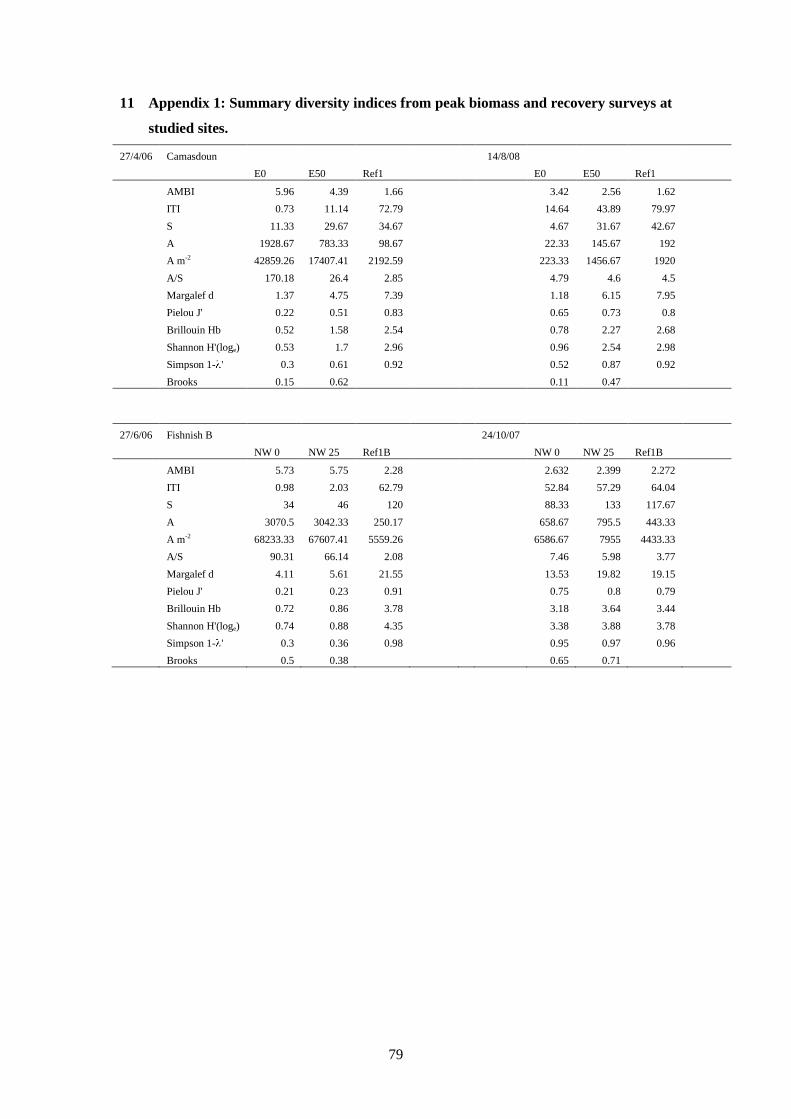
79
11 Appendix 1: Summary diversity indices from peak biomass and recovery surveys at
studied sites.
27/4/06 Camasdoun 14/8/08
E0 E50 Ref1 E0 E50 Ref1
AMBI 5.96 4.39 1.66 3.42 2.56 1.62
ITI 0.73 11.14 72.79 14.64 43.89 79.97
S 11.33 29.67 34.67 4.67 31.67 42.67
A 1928.67 783.33 98.67 22.33 145.67 192
A m-2 42859.26 17407.41 2192.59 223.33 1456.67 1920
A/S 170.18 26.4 2.85 4.79 4.6 4.5
Margalef d 1.37 4.75 7.39 1.18 6.15 7.95
Pielou J' 0.22 0.51 0.83 0.65 0.73 0.8
Brillouin Hb 0.52 1.58 2.54 0.78 2.27 2.68
Shannon H'(loge) 0.53 1.7 2.96 0.96 2.54 2.98
Simpson 1- ' 0.3 0.61 0.92 0.52 0.87 0.92
Brooks 0.15 0.62 0.11 0.47
27/6/06 Fishnish B 24/10/07
NW 0 NW 25 Ref1B NW 0 NW 25 Ref1B
AMBI 5.73 5.75 2.28 2.632 2.399 2.272
ITI 0.98 2.03 62.79 52.84 57.29 64.04
S 34 46 120 88.33 133 117.67
A 3070.5 3042.33 250.17 658.67 795.5 443.33
A m-2 68233.33 67607.41 5559.26 6586.67 7955 4433.33
A/S 90.31 66.14 2.08 7.46 5.98 3.77
Margalef d 4.11 5.61 21.55 13.53 19.82 19.15
Pielou J' 0.21 0.23 0.91 0.75 0.8 0.79
Brillouin Hb 0.72 0.86 3.78 3.18 3.64 3.44
Shannon H'(loge) 0.74 0.88 4.35 3.38 3.88 3.78
Simpson 1- ' 0.3 0.36 0.98 0.95 0.97 0.96
Brooks 0.5 0.38 0.65 0.71
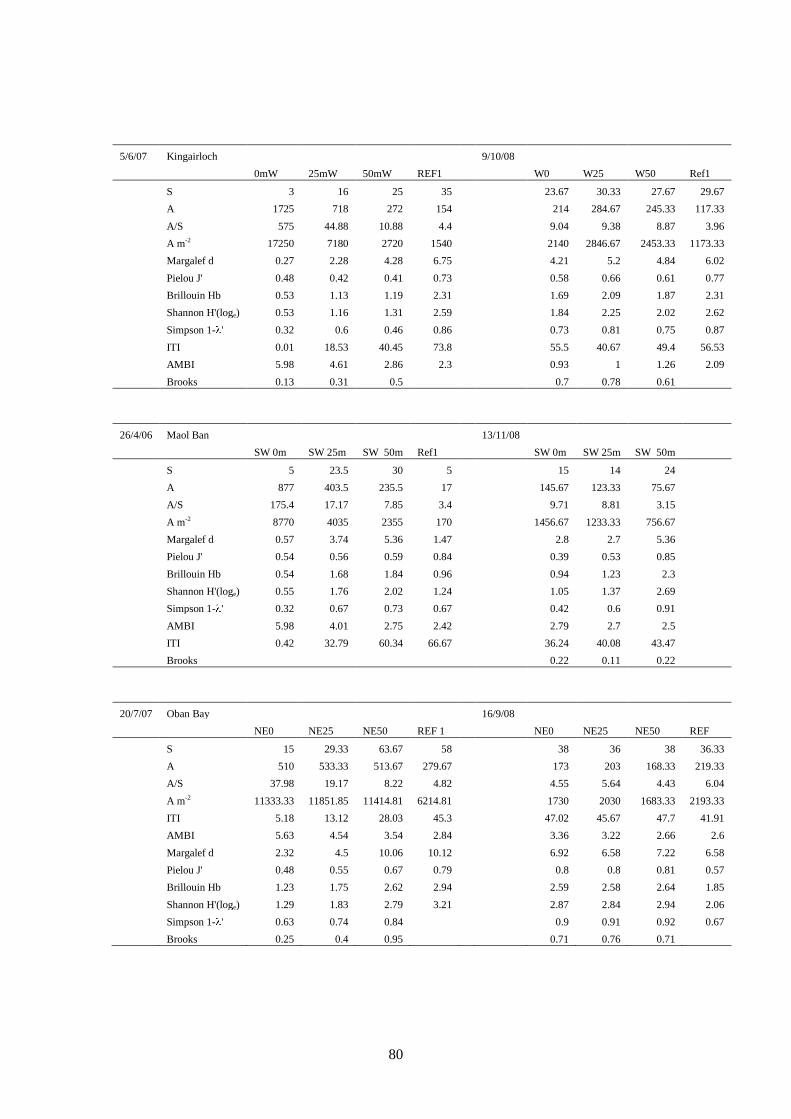
80
5/6/07 Kingairloch 9/10/08
0mW 25mW 50mW REF1 W0 W25 W50 Ref1
S 3 16 25 35 23.67 30.33 27.67 29.67
A 1725 718 272 154 214 284.67 245.33 117.33
A/S 575 44.88 10.88 4.4 9.04 9.38 8.87 3.96
A m-2 17250 7180 2720 1540 2140 2846.67 2453.33 1173.33
Margalef d 0.27 2.28 4.28 6.75 4.21 5.2 4.84 6.02
Pielou J' 0.48 0.42 0.41 0.73 0.58 0.66 0.61 0.77
Brillouin Hb 0.53 1.13 1.19 2.31 1.69 2.09 1.87 2.31
Shannon H'(loge) 0.53 1.16 1.31 2.59 1.84 2.25 2.02 2.62
Simpson 1- ' 0.32 0.6 0.46 0.86 0.73 0.81 0.75 0.87
ITI 0.01 18.53 40.45 73.8 55.5 40.67 49.4 56.53
AMBI 5.98 4.61 2.86 2.3 0.93 1 1.26 2.09
Brooks 0.13 0.31 0.5 0.7 0.78 0.61
26/4/06 Maol Ban 13/11/08
SW 0m SW 25m SW 50m Ref1 SW 0m SW 25m SW 50m
S 5 23.5 30 5 15 14 24
A 877 403.5 235.5 17 145.67 123.33 75.67
A/S 175.4 17.17 7.85 3.4 9.71 8.81 3.15
A m-2 8770 4035 2355 170 1456.67 1233.33 756.67
Margalef d 0.57 3.74 5.36 1.47 2.8 2.7 5.36
Pielou J' 0.54 0.56 0.59 0.84 0.39 0.53 0.85
Brillouin Hb 0.54 1.68 1.84 0.96 0.94 1.23 2.3
Shannon H'(loge) 0.55 1.76 2.02 1.24 1.05 1.37 2.69
Simpson 1- ' 0.32 0.67 0.73 0.67 0.42 0.6 0.91
AMBI 5.98 4.01 2.75 2.42 2.79 2.7 2.5
ITI 0.42 32.79 60.34 66.67 36.24 40.08 43.47
Brooks 0.22 0.11 0.22
20/7/07 Oban Bay 16/9/08
NE0 NE25 NE50 REF 1 NE0 NE25 NE50 REF
S 15 29.33 63.67 58 38 36 38 36.33
A 510 533.33 513.67 279.67 173 203 168.33 219.33
A/S 37.98 19.17 8.22 4.82 4.55 5.64 4.43 6.04
A m-2 11333.33 11851.85 11414.81 6214.81 1730 2030 1683.33 2193.33
ITI 5.18 13.12 28.03 45.3 47.02 45.67 47.7 41.91
AMBI 5.63 4.54 3.54 2.84 3.36 3.22 2.66 2.6
Margalef d 2.32 4.5 10.06 10.12 6.92 6.58 7.22 6.58
Pielou J' 0.48 0.55 0.67 0.79 0.8 0.8 0.81 0.57
Brillouin Hb 1.23 1.75 2.62 2.94 2.59 2.58 2.64 1.85
Shannon H'(loge) 1.29 1.83 2.79 3.21 2.87 2.84 2.94 2.06
Simpson 1- ' 0.63 0.74 0.84 0.9 0.91 0.92 0.67
Brooks 0.25 0.4 0.95 0.71 0.76 0.71






























