Embed Size (px)
Citation preview

Cretaceous Research (1992) 13, 517-537
Barremian-Aptian calcareous plankton biostratigraphy from the Gorgo Cerbara section (Marche, central Italy) and implications for plankton evolution
*Rodolfo Coccioni, tEfisabetta Erba and tlsabella Premoli-Silva
* Ist~2o di Geologia, Universi~z di Urbino, vkz S. Chiara 27, 61029 Urbino, Italy t Dipartimento di S cienze della Terra, Universit~ di M ilano, via M angiagalli 34, 20133 M ilano, Italy
Received 4 November 1991 and accepted 20 November 1991
The Barremian-Apdan boundary interval of the Gorgo Cerbara section (Marche, central Italy) was revisited in order to improve stratigraphic correlations and investigate plankton evolution. A very close sampling at cm scale was carried out in the upper Maiolica and basal Scisti a Fucoidi formations. The latter formation includes the Livello Selli, the landward sedimentary expression of oceanic anoxic subevent OAE la. The study of closely spaced samples revealed that (1) the first diversification among planktonic foraminifera, marked by the appearance of the genus GlobigerineUoides, occurred during the Barremian (this evem was dated by means of ammonites); thus the first occurrence (FO) of both Globigerinelloides duboisi and Globigerindloides blowi cannot be used to idendfy the Apdan; (2) the appearance of Rucinolithus irregularis is the biostratigraphic event which best approximates the Barremian-Apdan boundary; and (3) chron M0, being slighdy younger than the FO of R. irregularis, is very close to this boundary. Semiquamitative and quantitative analyses of planktonic foraminifera, calcareous nannofossils and radiolarians show that the three groups fluctuate in abundance and assemblage composition throughout the studied interval. An important change in plankton distribution patterns occurs within the G. Mow/ foraminiferal Zone and C. litterarius nannofossil Zone, where planktonic foraminifera, radiolarians and calcareous narmofossils start to show large-scale, higher frequency fluctuations in abundance; fluctuations in the three groups are out of phase. These new plankton distribution patterns are interrupted by the Livello Selli, which is preceded and followed by 'critical intervals'. The distribution and composition of all three planktonic groups lead to the interpretation of the 'critical intervals' as representing increased nutrient contents in the surface water and the Livello Selli as a very high fertility event. Among planktonic foraminifera the hedbergellids seem to indicate a more eutrophic habitat than the globigerinelloids. In calcareous narmofossil assemblages, the abrupt crisis affecting the nannoconids prior to deposition of the Liveflo Selli suggests that these nannofossfls are characteristic of more oligotrophic conditions, whereas Zygodiscus erectus is indicative of a more eutrophic environment. Based on our data, the early Aptian Livello Selli has the same oceanographic significance as the latest Cenomanian Livello Bonarelli, i.e., it represents a high fertility event on a global sole.
KEY WORDS: integrated calcareous plankton biostratigraphy; magnetostratigraphy; black shales; OAE la; palaeoceanography; Barremian-Aptian.
1. Introduction
This paper deals with a detailed biostratigraphic study, based on planktonic foraminifera and calcareous nannofossils, of the Barremian-Aptian interval from the Gorgo Cerbara section (Marche, central Italy) (Figures 1 and 2), a section already studied from the magnetostratigraphic point of view (Lowrie et al . , 1980; Lowrie & Alvarez, 1984). This study was undertaken in order to accomplish the goal of IGCP Project 262, concerning the improvement of stratigraphic correlation schemes in the Tethyan realm.
The Gorgo Cerbara section is located in the Umbria-Marche Basin, which contains a continuous pelagic sedimentary sequence of Early Jurassic-Oligocene age. This pelagic sequence consists of limestones and marly limestones, with subordinate marls and radiolarites, and rare, but characteristic, discrete black-shale layers rich in organic matter (see Cresta et al. , 1989, for an overview). All the
0195-6671/92/5/60519 + 21 $08.00/0 © 1992 Academic Press Limited
518 R. Coccioni et al .
E 2 ~
0 -
.o_ u
e -
E
u - i e -
ca
e~
. . . .
m e ~
.¢~ e -
0
0 r-
c5
d~
z~
o e~
Q
Q
0
Y=
e..
o . ,=
e~
o
o
o ,d
Barremian-Aptian calcareous plankton biostratigraphy 519
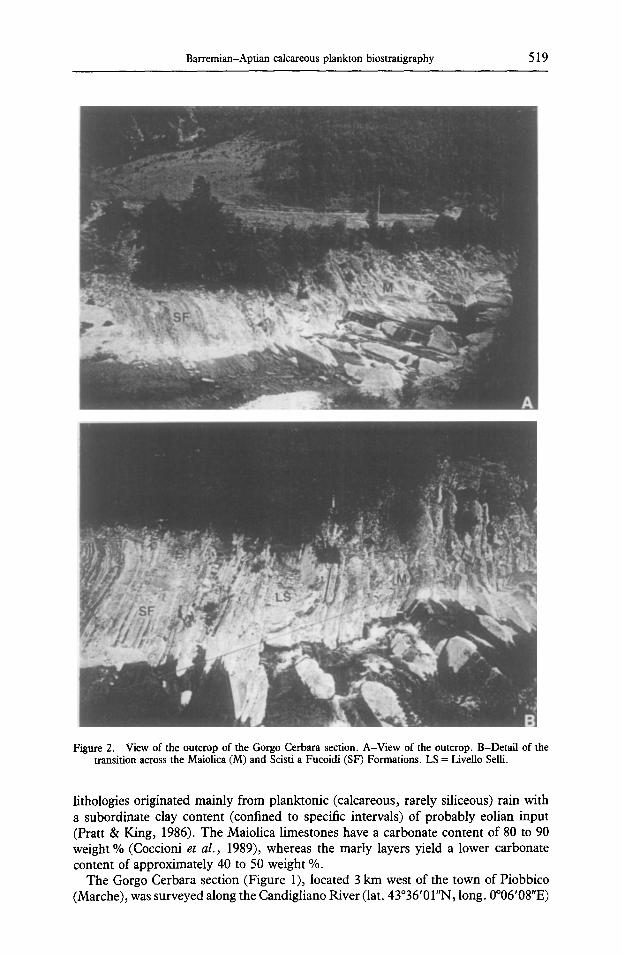
Figure 2. View of the outcrop of the Gorgo Cerbara section. A-View of the outcrop. B-Detail of the transition across the Maiolica (M) and Scisti a Fucoidi (SF) Formations. LS = Livello Selli.
lithologies originated mainly from planktonic (calcareous, rarely siliceous) rain with a subordinate clay content (confined to specific intervals) of probably eolian input (Pratt & King, 1986). The Maiolica limestones have a carbonate content of 80 to 90 weight % (Coccioni et al. , 1989), whereas the marly layers yield a lower carbonate content of approximately 40 to 50 weight %.
The Gorgo Cerbara section (Figure 1), located 3 km west of the town of Piobbico (Marche), was surveyed along the Candigliano River (lat. 43°36'01"N, long. 0°06'08"E)
520 R. Coccioni et al.
GORGO A CERBARA SECTION
] ~ a: ZONES ~ __ tu
ii ii e $ ! _ ~ o _== =a=
,
d - - / 3 o -
'~ I m r " 25 -
/ O Z
'< -- >" 20- | - - re"
.T" ~ M I
m ~ MI 10
MARLY LIMESTONE MARL a n d
a n d LIMESTONE CALCAREOUS MARL
_.=~ ~,o
.=
i i
~IANNOCONIDS (%)
40 80
NOT STUDIED
RAOIOLARIA (%)
l
ILL. 5
m m BLACK CHERT LAYER RADIOLARIAN SLUMPED SHALE and N O D U L E SANDY/SILT LAYER INTERVAL
MAJOR CHANGE
. . . _ A
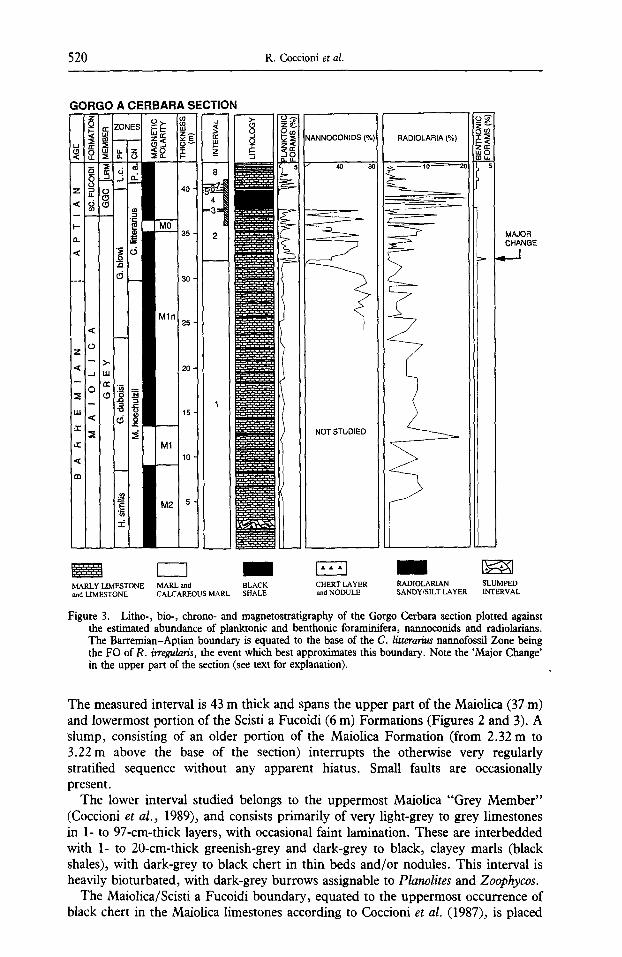
Figure 3. Litho-, bio-, chrono- and magnetostratigraphy of the Gorgo Cerbara section plotted against the estimated abundance of planktonic and benthonic foraminifera, narmoconids and radiolarians. The Barremian-Aptian boundary is equated to the base of the C. litterarius nannofossil Zone being the FO of R. irregularis, the event which best approximates this boundary. Note the 'Major Change' in the upper part of the section (see text for explanation).
The measured interval is 43 m thick and spans the upper part of the Maiolica (37 m) and lowermost portion of the Scisti a Fucoidi (6 m) Formations (Figures 2 and 3). A slump, consisting of an older portion of the Maiolica Formation (from 2.32 m to 3.22 m above the base of the section) interrupts the otherwise very regularly stratified sequence without any apparent hiatus. Small faults are occasionally present.
The lower interval studied belongs to the uppermost Maiolica "Grey Member" (Coccioni et al., 1989), and consists primarily of very light-grey to grey limestones in 1- to 97-cm-thick layers, with occasional faint lamination. These are interbedded with 1- to 20-cm-thick greenish-grey and dark-grey to black, clayey marls (black shales), with dark-grey to black chert in thin beds and/or nodules. This interval is heavily bioturbated, with dark-grey burrows assignable to Planolites and Zoophycos.
The Maiolica/Scisti a Fucoidi boundary, equated to the uppermost occurrence of black chert in the Maiolica limestones according to Coccioni et al. (1987), is placed

Barremian-Aptian calcareous plankton biostratigraphy 521
at 37 m above the base of the section. The measured portion of the Scisti a Fucoidi belongs to the "Greenish-grey cherty Member" (4.1 m thick) and "Lower reddish marly Member" (1.9 m thick). The former member can be subdivided into three parts: lower, middle and upper. The lower and upper parts, 0.78 m and 1.4 m thick respectively, are very similar and characterized primarily by bioturbated greenish- grey cherty limestones, marly limestones and subordinate marls in beds of 1- to 30-cm-thick. The middle part, corresponding to the Livello Selli (Coccioni et al., 1987, 1989), is 1.92 m thick and consists of laminated to bioturbated olive-grey, greenish-grey and dark-grey to black mudstones and shales, with 20 thin (1 to 3 cm) greyish-yellow, olive-grey and medium- to dark-grey radiolarian silty/sandy layers. The greenish-grey mudstones and radiolarian layers are typically bioturbated. Within the Livello Selli two intervals can be distinguished, a lower "green interval" (0.95 m thick) and an upper "black interval" (0.97 m thick) (Coccioni et al., 1987, 1989). Pyrite nodules occur mainly at the base of the black interval, and fish remains are recorded throughout the upper black interval. The studied portion of the "Lower reddish marly Member" of the Scisti a Fucoidi consists primarily of bioturbated, greenish-grey calcareous marls and marls, with subordinate marly limestones, in beds 1- to 30-cm-thick; chert is absent.
2. Materials and methods
The 465 samples studied were collected at 8 to 10 cm intervals in the lower part of the section, and at 5 cm intervals in the upper part. The measured section is marked on the outcrop with blue paint at metre intervals.
The study of foraminifera and radiolarians was primarily conducted in thin section, the predominant lithologies being hard, and subordinately on washed material from the more marly beds. Foraminiferal and radiolarian abundances were estimated directly by comparing the thin section content per square unit (>2 cm z) to the standard % diagrams of Baccelle & BoseUini (1965) (see also Tornaghi et al., 1989). Photomicrographs of selected planktonic foraminifera and microfacies are presented in Figures 4 to 6.
Calcareous nannofossils were studied on smear slides with an optical microscope at 1250x magnification. Slides were made from the same samples used for the foraminiferal study of the upper 20 m of the section. Quantitative data were collected by counting 300 specimens per sample in random traverses of smear slides prepared from raw material not subjected to concentration. Abundances of single species were normalized to 100 of the total floral abundance. Semiquantitative and qualitative investi- gations were performed to estimate the type and intensity of diagenetic modifications.
3. New biostratigraphic data
3.1. Planktonic foraminifera The interval studied spans the first important diversification of planktonic foramin- ifera. The major events (Figure 3) are: (1) the first occurrence (FO) of the genus Globigerinelloides, with G. duboisi appearing at 8.43 m above the base of the section (=abs); (2) the FO ofG. blowi at 23.28 m (abs) and (3) the FO ofLeupoldina cabri at 40.10 m (abs). Based on these events four biozones were identified (from bottom to top):
Hedbergella similis Zone (from bottom to 8.43 m) = Interval from the FO of the nominal species to the FO of Globigerinelloides duboisi.
522 R. Coccioni et al.
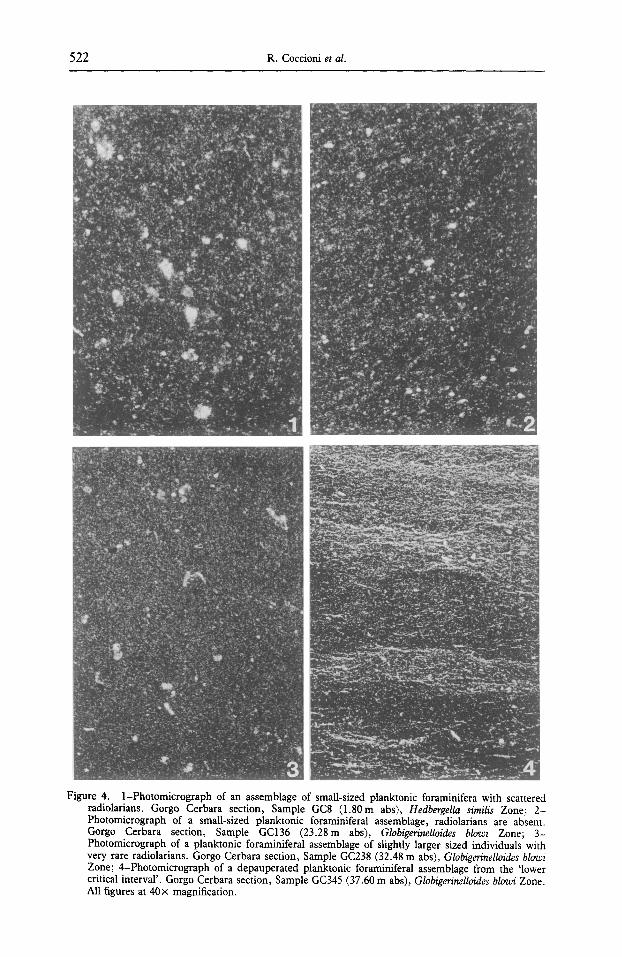
Figure 4. 1-Photomicrograph of an assemblage of small-sized planktonic foraminifera with scattered radiolarians. Gorgo Cerbara section, Sample GC8 (1.80m abs), Hedbergella similis Zone; 2- Photomicrograph of a small-sized planktonic foraminiferal assemblage, radiotarians are absent. Gorgo Cerbara section, Sample GC136 (23.28m abs), Globigerinelloides blowi Zone; 3- Photomicrograph of a planktonic foraminiferal assemblage of slightly larger sized individuals with very rare radiolarians. Gorgo Cerbara section, Sample GC238 (32.48 m abs), Globigerinelloides blowi Zone; 4-Photomicrograph of a depauperated planktonic foraminiferal assemblage from the 'lower critical interval'. Gorgo Cerbara section, Sample GC345 (37.60 m abs), Globigerinelloides blowi Zone. All figures at 40× magnification.
Barremian-Aptian calcareous plankton biostratigraphy 523
" ~ ] " 5 5 -- o .
I
s -
¢ • " 4 , - i? 4. - . ¢; •
(
- ~ ; - . " t . ~ ~
- - ° IL~ .
1
. I P
|
*
" . . . 4 o t
Figure 5. l-Photomicrograph of a depauperated planktonic foraminiferal assemblage from the 'upper critical interval'. Gorgo Cerbara section, Sample GC388 (39.78 m abs), Globigerinelloides blowi Zone; 2-Photomicrograph of a depauperated planktonic foraminiferal assemblage with scattered deformed radiolarians from the 'upper critical interval'. Gorgo Cerbara section, Sample GC388 (39.78 m abs), Globigerinelloides blowi Zone; 3-Photomicrograph of a depauperated planktonic foraminiferal assemblage with scattered deformed radiolarians from the 'upper critical interval'. Gorgo Cerbara section, Sample GC393 (39.94 m abs), Globigerinelloides blowi Zone; 4-Photomicrograph of a richer planktonic foraminiferal assemblage of 'normal' sized individuals with scattered radiolarians. Gorgo Cerbara section, Sample GC420 (41.10m abs), Leupoldina cabri Zone. All figures at 40x magnification.
524 R. Coccioni et al.
4
9" .~ ~ ' ~ ~#
L~
6
Figure 6. 1-Hedbergdla sigali Moullade, transverse section. Gorgo Cerbara section, Sample GC28 (5.50 m abs), Hedbergdla sirailis Zone; 2-Globigerindloides duboisi (Chevalier), axial section. Gorgo Cerbara section, Sample GC427 (41.34m abs), Leupoldina cabri Zone; 3-GlobigerineUoides duboisi (Chevalier), axial section. Gorgo Cerbara section, Sample GC255 (33.32 m abs), GlobigerineUoides b/ow/Zone; 4-Globigerinelloides blowi (BoUi), axial section. Gorgo Cerbara section, Sample G~27 (41.34 m abs), Leupoldina cabri Zone; 5-Globigerinelloidesferredemis (MouUade), axial section. Gorgo Cerbara section, Sample GC427 (41.34 m abs), Leupoldina cabri Zone; 6-GlobigerineUoides ferreolensis (Moullade), axial section. Gorgo Cerbara section, Sample GC395 (40.10 m abs), Leupddina cabri Zone. All figures at 160x magnification.
The zonal marker, although rare, is already present in the lowermost sample. Thus, the base of the zone lies in the underlying, unsampled interval still belonging to the Maiolica Formation. The planktonic foraminiferal fauna in this interval consists of very rare to rare, small-sized (<100#m) species such as the zonal marker, Hedbergella sigali, Globuligerina hoterivica, and Gubkinella graysonensis, a species now considered as a benthic form (Loeblich & Tappan, 1988).
Globigerinelloides duboisi Zone (8.43 m to 23.28 m; total thickness 14.85 m) = Interval from the FO of the nominal species to the FO of
GlobigerineUoides blowi. Besides the zonal marker and Globigerinelloides gottisi, which are rare and of small
size, the planktonic foraminiferal assemblage in this zone is still poor and consists of the same small-sized species as in the underlying zone. Rare, small-sized Hedbergella delrioensis appear close to the base of this interval.
Globigerinelloides blowi Zone (23.28 m to 40.10 m; total thickness 16.82 m) = Interval from the FO of the nominal species to the FO of
Leupoldina cabri. The planktonic foraminiferal assemblage of this interval is enriched by two new

Barremian-Aptian calcareous plankton biostratigraphy 525
species, the zonal marker and Hedbergella occulta, in association with the same species inherited from the previous zones. Planktonic foraminifera increase in abundance, but the size of specimens still remains small. The LiveUo Selli, included in this zone, is devoid of planktonic foraminifera.
Leupoldina cabn" Zone (40.1 m to top; measured thickness 2.90 m) = Total range zone.
The upper limit of this zone was not sampled. In this interval the planktonic foraminiferal assemblages display a marked increase in abundance associated with a clear increase in the size of the various species, especially Hedbergella delrioensis, which is represented by normal-sized specimens. Preservation also improves. The faunas include all the species inherited from the previous zone associated with very rare to rare Leupoldina cabri and Globigerinelloides ferreolensis. The latter species increases in size throughout this zone, with almost full-sized specimens occurring at the top of the sampled interval.
The succession of events and related biozones from the Gorgo Cerbara section are similar to those described by Tornaghi et al. (1989) from the nearby Piobbico core and from several other localities around the world. First order correlations between planktonic foraminifera and ammonite zonation, however, are seldom mentioned. Until now, this succession of events was mainly attributed to the Aptian (e.g. Sigal, 1977, 1979; Arthur & Premoli Silva, 1982; Erba & Quadrio, 1987; Sliter, 1989b). It is pointed out that Hedbergella delrioensis and Globigerinelloides ferreolensis appear earlier at Gorgo Cerbara than previously described, that is in the G. duboisi Zone and just above the base of the Leupoldina cabri Zone, respectively. Range charts with the detailed documentation of planktonic foraminiferal distribution will be included in a forthcoming paper (in preparation).
3.2. Calcareous nannofossils Calcareous nannofossils are abundant and moderately well preserved throughout the studied interval, with the exception of the Livello Selli which is barren of calcareous fossils. Three biostratigraphic events were recognized: the FO of Rucinolithus irregularis (29.90m abs), the FO of Parhabdolithus angustus (40.85 m abs) and the FO of Eprolithusfloralis (40.85 m abs). On the basis of these events three zones were identified (from bottom to top):
Micrantolithus hoschulzii Zine (from bottom to 29.90 m) = Interval from the last occurrence (LO) of Calcicalathina oblongata to the FO
of Rucinolithus irregularis and/or the FO of Chiastozygus litterarius and/or the LO of Nannoconus colomii (Thierstein, 1973).
The lower 29.90 m of the studied section were assigned to the M. hoschulzii Zone based on the absence of Cala'calathina oblongata, R. irregularis and C. litterarius (see also Bralower, 1987). The nannofossil assemblage is abundant, moderately diverse and moderately well preserved. Nannoconids are the dominant forms constituting up to 70% of the total nannoflora. The most abundant species are Nannoconus steinmannii, N. colomii, N. kampmeri, N. bucheri and N. wassallii. Other nannofossil species consistently recorded in this interval are Watznaueria barnesae, Parhab- dolithus asper, Lithraphidites carniolensis, Rucinolithus terebrodentarius and Cyclagelosphaer a margerelii.
Chiastozygus litterarius Zone (29.90 m to 40.85 m; total thickness 10.95 m) = Interval from the FO of Rucinolithus irregularis and/or the FO of Chiastozygus
litterarius and/or the LO of Nannoconus colomii to the FO of Parhabdolithus angustus and/or the FO of Eprolithus floralis (Thierstein, 1973).

526 R. Coccioni et al.
The FO of Rucinolithus irregularis was observed at 29.90 m and used to place the base of the zone. Nannofossil assemblages are very similar to those of the underlying interval, but wide fluctuations in the relative abundance of taxa were recorded. Nannoconids, in particular, display sharp changes in relative abundance, always related to the lithology (limestones are rich in nannoconids, marls are poor), and they disappear altogether above 37.52 m. N. steinmannii and N. colomii decrease in abundance from 32.00 m upward. Diversity (=number of species) is higher in this zone and species making their first appearance include FlabeUites oblongus, CoroUithion achylosus and Chiastozygus litterarius. In the upper portion of the zone nannofossil assemblages are relatively enriched with Zygodiscus spp.
Parhabdolithus angustus Zone (40.85 m to top; measured thickness 2.15 m) = Interval from the FO of Parhabdolithus angustus and/or the FO of Eprolithus
floralis to the FO of Prediscosphaera columnata (Thierstein, 1973) The FO of Parhabdolithus angustus was recorded at 40.85 m and the FO of
Eprolithusfloralis at 41.85 m. The base of the zone was equated to the former event; the upper limit was not sampled. In this interval the nannofossil assemblages display an increased diversity, while the total abundance and preservation remain similar to those recorded in the underlying zones. Nannoconids are absent in the lower part of the zone but reappear at 42.40 m; their relative abundance is low and N. truittii is the dominant species.
The sequence of nannofossil events and biozones is consistent with previous data documented by Monechi (1981), Erba (1988) and Erba et al. (1989a) from other sections of the Umbria-Marche basin. Minor discrepancies were noted with respect to the biostratigraphy provided by Bralower (1987), who also studied the Gorgo Cerbara section; these are possibly due to the much more detailed sampling of the present study. Bralower (1987) recorded the FO of R. irregularis at 35.10 m and the LO of N. steinmannii at 26.00 m in the present section. Based on these events Bralower (1987) distinguished the NC 5D and NC 5E subzones within the upper part of the M. hoschulzii Zone and correlated the FO of R. irregularis with the base of chron M0 as determined by Lowrie & Alvarez (1984). The new data show that the FO of R. irregularis occurs prior to M0, in agreement with other findings from the Pacific (Site 463, Tarduno et al., 1989), North Atlantic (Site 541, Moullade et al., 1988) and northern Italy (Cismon section, Channell et al., 1979; Pie'del Dosso and Capriolo sections, Channell & Erba, 1992). The LO of N. steinmannii is considered an unreliable event because it is masked by the general decrease of nannoconids, which display a crisis slightly earlier than chron M0. This species has been recorded in upper Aptian limestones from several land sections (Bralower, 1987; Erba & Quadrio, 1987) and deep-sea cores (e.g. Applegate & Bergen, 1988).
4. Correlation with ammonite zonation, magnetostratigraphy and chronostratigraphy
Hoedemaeker & Bulot (1990) presented an update of Early Cretaceous ammonite distribution and zonation, while Aguado et al. (1990, 1992) give a direct correlation between ammonite zonation and nannofossil and planktonic foraminiferal biostrati- graphic schemes. Accordingly, the appearance of Hedbergella similis and of the genus Globigerinelloides occur prior to the base of the Aptian and are associated with Barremian ammonites. Moreover, preliminary data on planktonic foraminifera from the Rio Argos section (Caravaca, S. Spain: personal observations), in which Hoedemaeker (pers. comm., 1991) established a very reliable ammonite zonation,

Barremian-Aptian calcareous plankton biostratigraphy 527
allows us to state that the FO of Globigerinelloides duboisi is in the uppermost ammonite zone of the lower Barremian (Holcodiscus caillaudianus Zone). G. blowi appears in the upper Barremian (Imerites giraudi ammonite Zone), and the FO of L. cabri is above the base of the Aptian, confirming Aguado et al.'s data. Consequently, the FO of Hedbergella similis, which predates the appearance of the genus Globigerinelloides, cannot be used to identify the base of the Aptian (Sigal, 1977, 1979; Lowrie et al., 1980; Arthur & Premoli Silva, 1982; Erba & Quadrio, 1987; Sliter, 1989b), nor can it be considered indicative of the upper Barremian (Harland et al., 1990). In addition, the FO ofG. blowi does not identify the base of the Aptian (Caron 1985).
Several reasons may account for such discrepancies: (a) the generally poor preservation, small size and scantiness of planktonic foraminifera at the beginning of their radiation; (b) most sections spanning the Barremian-Aptian interval are composed mainly of hard limestones, necessitating the study of their foraminifera in thin section; (c) the fact that planktonic foraminifera are unevenly distributed from layer to layer, so their real record can be obtained only with very close, detailed sampling and last, but not least, (d) the fact that calcareous plankton have only recently been investigated in detail from sections well dated by ammonites.
A first order correlation between planktonic foraminifera and magnetostratigraphy was not easy to obtain. The Gorgo Cerbara section has been the site of some of the first tentative efforts at such a correlation, but these have failed because of insufficient sampling density (Lowrie et al., 1980; Lowrie & Alvarez, 1984). At Site 641 in the North Atlantic, however, Moullade et al. (1988) recorded the occurrence of Globigerinelloides duboisi in the upper part of the M. hoschulzii Zone, prior to M0, supporting the older age of this event reported above.
Thierstein (1973) proposed three calcareous nannofossil events to determine the Barremian-Aptian boundary, namely the FO of R. irregularis, the FO of C. litterarius and the LO of N. colomii (=N. steinmannii). These biostratigraphic events were documented from the stratotypes and calibrated to both ammonite and planktonic foraminiferal biozones. Subsequent works on low latitude land sections have shown that C. litterarius is usually too rare to be considered a reliable marker and that N. colomii/N, steinmannii does not become extinct close to the FO of R. irregularis. The latter event, then, appears to be the best nannofossil event for approximating the Barremian-Aptian boundary. However, the precise correlation between the FO of R. irregularis and ammonite zonation is still under study.
In the Gorgo Cerbara section Lowrie et al. (1980) and Lowrie & Alvarez (1984) identified chrons M0 and M1. In the present section these chrons correspond to the intervals from 35.22m to 36.52m and from 9.1m to 13.5m, respectively. According to the present study (Figure 3), M1 occurs within, but close to the base of the G. duboisi Zone and M0 is included in the upper half of the G. blowi Zone. The FO of R. irregularis was observed 5.56 m below the base of chron M0, confirming previous correlations (Moullade etal., 1988; Channell & Erba, 1992). Since the FO of R. irregularis is the closest event to the Barremian-Aptian boundary, chron M0 is also very close to the boundary, as suggested by Bralower (1987) for totally different reasons.
5. Distribution of microfossils
5.1. Planktonic foraminifera The percentage abundance curve of planktonic foraminifera shows that they are not evenly distributed throughout the studied section (Figures 3 and 7). A number of

528 R. Coccioni et al.
< ...J IJJ
(D ,.,.I ,..J I l l _> . . I
F-
03
c~
O3
<z: C~ ..J O <
UPPER CRITI( iNTERVAL
N
£3
C) c~
LL
LOWER CRIT~ INTERVAL
SAMPLE I
,~ [ PLANKTONIC FORAMS (%) NANNOCONIDS (%]
5'0 100 0
i
RADIOLARIA (%)
40
7
200~BENTHONIC
6 ,
Figure 7. Detail of the Gorgo Cerbara section across the Livello Selli with detailed lithology, sample position, and 'lower' and 'upper critical intervals' plotted against the estimated abundance of planktonic and benthonic foraminifera, nannoconids, and radiolarians. Note the distribution of Zygodiscus erectus. Legend: 1-marl, clayey marl, marly clay, bioturbated (a) and laminated (b) clay and/or claystone and marlstone; 2-black shale; 3-greenish-grey marly limestone; 4-greyish-white limestone; 5-whitish-grey limestone and marly limestone; 6-radiolarian silty/sandy layer; 7-chert nodules and layers.
intervals can be distinguished (Figures 3 and 7) (from bottom to top): (1) A lowermost interval up to the middle of the G. blowi Zone (32.10 m), in
which planktonic foraminiferal abundance ranges between 1 and 2% with very rare horizons in which they are virtually absent.
(2) A second interval, still attributed to the G. blowi Zone, up to 37.52 m, is characterized by the constant presence and an overall increase in abundance of planktonic foraminifera (from 1% to about 5%). Fluctuations in abundance are closely spaced and larger than in the underlying interval.
(3) A third interval, 26-cm-thick (37.52 to 37.78 m), is characterized by a general decrease in abundance of planktonic foraminifera ranging from a maximum of 1% to almost zero, with close and ample fluctuations. In this interval, planktonic foraminiferal diversity decreases and the fauna consists almost exclusively of poorly preserved small-sized hedbergellids. This interval is called the 'lower critical interval'.
(4) A fourth interval, corresponding to the Livello SeUi (37.78 to 39.70 m), is devoid of foraminifera.

Barremian-Aptian calcareous plankton biostratigraphy 529
(5) A fifth interval, 27-cm-thick (39.70 to 39.97 m), is characterized by very low planktonic foraminiferal abundance (<0.5%). Assemblages are very similar to those found below the Livello Selli, consisting almost exclusively of small hedbergellids. This interval is called the 'upper critical interval'.
(6) A sixth interval, belonging to the uppermost part of the G. blowi Zone (39.97 to 40.10 m), is characterized by more common Globigerinelloides representatives in association with hedbergellids, which are still smaller than those from the second interval. Planktonic foraminifera overall are scarce (max. 1%).
(7) A seventh interval, belonging to the lower part of the L. cabri Zone (40.10 to 41.50m), is characterized by large fluctuations in abundance with discrete peaks ranging from about 0.5% to >4%. Moreover, the planktonic foraminifera are more diversified, although individuals are still of smaller size than those from interval 2.
(8) An eighth, uppermost interval, belonging to the uppermost portion of the L. cabri Zone studied here (41.50 to 43.00 m), is characterized by specimens that are equal to or even larger in size than those of interval 2, and by an increase in diversity. The abundance of planktonic foraminifera, however, never exceeds 2.5%.
The general trend in planktonic foraminifera from the Gorgo Cerbara section is an increase in abundance, diversity and mean size of the specimens from the H. similis Zone to the L. cabri Zone. A marked change (Figure 3) occurs within the G. blowi Zone where planktonic foraminifera display a significant increase in abundance and diversity. The general trend is interrupted by a very important event corresponding to the Livello Selli. The conditions leading to the deposition of this peculiar layer, however, initiated before and continued after the Livello Selli, as testified by the assemblages of the 'lower' and 'upper critical intervals', respectively.
5.2. Benthonic foraminifera Benthonic foraminifera are rare throughout the section. However, a definite pulse in the abundance of benthonic foraminifera occurs at the same level as that at which planktonic foraminifera markedly increase in abundance, at 32.10m. Benthonics were not recorded from the Livello Selli, but reappear in the 'upper critical interval', above which they show a second peak in abundance (Figures 3 and 7).
5.3. Radiolarians Radiolarians are generally abundant, except at the base of the section, but they are distributed unevenly throughout (Figure 3). In the lower portion their abundance fluctuates from 2% to 18% with positive peaks corresponding to a decrease in planktonic foraminifera and vice versa. The general trend in this interval is toward a decrease in radiolarian abundance reaching a minimum of 1% at 32.10 m, which coincides with the marked increase in planktonic foraminiferal abundance. Above this, radiolarians show a general increase in abundance but with ample, high frequency fluctuations, whose positive peaks correspond to negative peaks in planktonic foraminiferal abundance. The Livello-SeUi displays the maximum fluctuations in radiolarian abundance, from <1% up to 20%, with radiolarians concentrated in discrete layers (Figure 7). In the 'upper critical interval' radiolarians decrease in abundance with respect to the Livello Selli, but they still display wide abundance fluctuations, with one peak of 20% at 39.80 m. Correspond- ing with the new increase in abundance of planktonic foraminifera at 40.10 m (base of the L. cabri Zone) radiolarian abundance dramatically decreases (to 0.1-5%). Abundances similar to those of the Livello Selli are recorded only at the top of the interval studied.

530 R. Coccioni et al.
5.4. Calcareous nannofossils Quantitative analyses of calcareous nannofossils show wide fluctuations in assem- blage composition. Major changes are marked by the nannoconid group, which constitute the bulk of the nannoflora of the Majolica limestones. Investigation bed by bed provided a detailed picture of nannoconid abundance versus other nannofossils (mainly coccoliths) during the late Barremian and Aptian (Figures 3 and 7) and discrete intervals, similar to those identified for foraminifera, were distinguished as follows (from bottom to top):
(1) In the lowermost portion of the studied section (24.00 m to 32.10 m; upper part of the M. hoschulzii Zone and lower part of the C. litterarius Zone) nannoconids constitute between 55% and 75% of the nannofossil assemblages, which are characterized by low diversity.
(2) In the overlying interval (middle part of the C. litterarius Zone) Nannoconus spp. show high frequency fluctuations, constituting between 0% and 70% of the assemblages. The first drop in nannoconid abundance coincides with the first peak in planktonic foraminiferal abundance (32.10 m), which marks the base of the high frequency fluctuation interval where the nannoconid distribution pattern is not in phase with either planktonic foraminiferal or radiolarian curves.
(3) Nannoconids experienced a final crisis at 37.50m and the 'lower critical interval' (37.52 m to 37.78 m) is characterized by the absence of nannoconids. In this latter interval nannofloras are dominated by W. barnesae which accounts for up to 60% of the assemblages, with occasional peaks of 20% to 30% of R. terebrodentarius and A. infracretacea. Although W. barnease may be used as a preservation index owing to its resistance to dissolution/diagenesis (Thierstein, 1980; Rowth & Bowdler, 1981; Roth, 1984, 1987; Erba, 1986, 1992; Roth & Krumbach, 1986), the nannoconid crisis cannot be attributed to a dissolution event. In fact, the genus Nannoconus proved to be very resistant to diagenesis (Thierstein, 1976; Noel & Melguen, 1978) and dissolution-susceptible taxa such as Zygodiscus spp., virtually absent in most of the Gorgo Cerbara section, were recorded in the 'lower critical interval' where nannoconids are missing. The occurrence of Zygodiscus erectus (1% to 2%) and Discorhabdus rotatorius (1% to 3%) may indicate an increase in fertility (Roth & Krumbach, 1986; Erba, 1986; Premoli Silva et al., 1989a; Roth, 1989; Watkins, 1989; Erba, 1992).
(4) The Livello Selli (37.78 m to 39.70 m) is barren of calcareous nannofossils. (5) Nannofossil assemblages from the 'upper critical interval' (39.70m to
39.97 m) are very similar to those observed in the 'lower critical interval'. Again, W. barnesae is the dominant form, nannoconids are absent, R. terebrodentarius and A. infracretacea show abundance peaks of 20% to 30%, and Z. erectus is consistently present (1% to 2% of the assemblage).
(6, 7, and 8) Above the 'upper critical interval', there is an increase in diversity of nannofloras, characterized by P. asper, R. irregularis, R. terebrodentarius, L. carniolensis and A. infracretacea. Nannoconids reappear in the uppermost 0.60 m. Z. erectus was not observed above 40.84 m. The reappearance of nannoconids in the lower upper Aptian has already been noticed in several other Italian sections (Erba, 1989) and is correlatable with peaks documented by Mutterlose (1987, 1989, 1991) in the middle Aptian from NW Germany: they are interpreted as migrations of warm-water forms from Tethys during times of transgression.
6. Palaeoenvironmental remarks and interpretation
The distribution and composition of calcareous nannoflora and planktonic fauna

Barremian-Aptian calcareous plankton biostratigraphy 531
from the Gorgo Cerbara section show that the sampled section can be split into a number of eco-intervals, as follows:
(1) A lower segment characterized by a sparse planktonic foraminiferal fauna of low diversity and small-sized individuals, by few to common radiolarians which decrease in abundance upwards, and by dominant nannoconids,
(2) The base of interval 2 is marked by an abrupt change in planktonic content: planktonic foraminifera diversify and increase markedly in abundance, while both nannoconids and radiolarians start to show high frequency fluctuations in abun- dance. Fluctuations in radiolarian abundance are opposite to those of planktonic foraminifera, whereas nannoconids are out of phase with both groups.
(3) In the 'lower critical interval' planktonic foraminifera decrease in overall size and diversity becomes almost monogeneric (mainly hedbergellids). Nannoconids suddenly disappear and are replaced by very abundant W. barnesae, common R. terebrodentarius and A. infracretacea, and some Zygodiscus erectus (which first occurs slightly earlier). Radiolarians show a marked decrease in abundance. The scarcity of radiolarians in this interval is possibly due to preservation (Figure 7).
(4) The Livello Selli, which is alternately very rich in radiolarians and organic matter of marine origin up to 9% t.w. (Pratt & King, 1986; Coccioni et al., 1987, 1989), is devoid of calcareous plankton and benthos.
(5) The 'upper critical interval' mirrors the 'lower critical interval'. The planktonic foraminiferal fauna is almost monogeneric, poorly diversified, and consists of small-sized individuals. Benthonic foraminifera are very rare. Nan- nofloras are dominated by W. barnesae, with rare/few Z. erectus; nannoconids are totally absent. The radiolarian content is, however, much more abundant overall than in the 'lower critical interval'.
(6 and 7) Above the 'upper critical interval', planktonic foraminifera rediversify and become more abundant although the size of specimens remains small. Radiolarians become scarce, but Z. erectus is still present among the calcareous nannofossils.
(8) In the uppermost interval the planktonic foraminiferal fauna is more diverse, with larger specimens, while radiolarians fluctuate widely in abundance. The reappearance of nannoconids at the top of this interval corresponds to a trough in radiolarian abundance. Z. erectus was not recorded from this interval.
The major change in planktonic communities occurs very close to the Barremian/Aptian boundary and coincides with a marked change in sedimentary regime (more marly beds and frequent black shale layers). We wonder if this event is the first signal of the following Livello Selli, or if it is due to an independent cause. Major changes in nannofossil assemblage composition and in the rate of diversification close to the Barremian-Aptian boundary and prior to the deposition of the lower Aptian black shales have been noticed in both the Tethyan area (Erba & Larson, 1991) and the Boreal realm (Mutterlose, 1991). Erba & Larson (1991) pointed out the coincidence of changes in calcareous nannofloras with an episode of extremely high intraplate volcanism in the Pacific that was documented by Larson (1991a) and correlated with the widespread deposition of lower Aptian black shales of the OAE la subevent (Larson, 1991b). A possible link between volcanic episodes and black shale deposition was also hypothesized by Schlanger et al. (1987) who underlined the correlation between the intensity of mid-Cretaceous volcanism and the occurrence of carbonaceous sediments.
All the available data lead us to interpret the Livello Selli as an abrupt event of extremely high fertility during a period when oligotrophic conditions prevailed. This high fertility event is preceded and followed by transitional conditions more on the eutrophic side of the Trophic Resource Continuum (Figure 8; Hallock, 1987). The

532 R. Coccioni et al.
TROPHIC RESOURCE CONTINUUM
I I I I HIGHLY
OLIGOTROPHIC
OHGOTROPHIC MESOTROPHIC EUTROPHIC HIGHLY
EUTROPHIC
REDUCED MIXING
STRONG MIXING
MODERN PACIFIC
MODERN ATLANTIC
I==
LIFE-HISTORY k-mode intermediate STRATEGY specialists
EXAMPLES:
FORAMINIFERS
PLANKTONIC
r-mode opportunists
ticinellids Globigerinelloides hedbergellids heterohelicids
NANNOFOSSlLS nannoconids Zygodiscus erectus
SILICEOUS ORGANISMS RAD~LARIANS
I I I I I
DIATOMS
HIGHLY OLIGOTROPHIC MESOTROPHIC EUTROPHIC HIGHLY
OLIGOTROPHIC EUTROPHIC
Figure 8. A-A representation of the oceanic surface-water trophic-resource continuum (TRC), comparing the modern Atlantic and Pacific, and predicting how reduced oceanic mixing should expand the TRC while increased rates of oceanic mixing should reduce the TRC (adapted from HaUock, 1987). B-Lower Cretaceous life-history strategies and trophic resources; in the early history of planktonic foraminifera k-mode specialists, adapted to stable low-nutrient environments, are apparently missing, as are the r-mode opportunists. In fact, the ticineUids (k-mode specialists) first appear in the late Aptian and the heterohelicids (r-mode opportunists) (see Caron & Homewood, 1983) appear only in the late Albian along with a new ticineUid stock. In Barremian-Aptian time planktonic foraminifera seem to include only r-mode opportunists such as the hedbergellids, which were adapted to resource-richer, lower stability environments with eutrophic niches, while the globigerinelloids were apparently adapted to more mesotrophic conditions than hedbergellids. In contrast, calcareous nannofossils seem to occupy a much larger spectrum of the TRC: nannoconids appear adapted to oligotrophic to highly oligotrophic environments, while Zygodiscus erectus seems to prefer eutrophic conditions, partially overlapping with radiolarian's preferred environment.
conditions of maximum eutrophy are indicated by the C organic-rich black shales of the Livello Selli and very abundant radiolarians, which totally replaced the calcareous plankton (Erba et al., 1989b). The transitional conditions, below and above the Livello Selli, supported a depauperated planktonic foraminiferal fauna consisting almost totally of hedbergellids, which are considered the most eutrophic forms among Cretaceous planktonic foraminifera (Caron & Homewood, 1983; Leclde, 1987, 1989; Premoli Silva et al., 1989b; Tornaghi et al., 1989), and the nannofossil higher fertility index Zygodiscus erectus (Roth, 1981, 1989; Roth &

Barremian-Aptian calcareous plankton biostratigraphy 533
Krumbach, 1986; Premoli Silva et al., 1989a; Watkins, 1989; Erba, in press). Radiolarians are surprisingly scarce, presumably because of differential preservation. The 'normal' oligotrophic conditions are indicated by more diverse planktonic foraminiferal faunas, which include the globigerineUoids. Very abundant nan- noconids dominating the calcareous nannofloras seem to indicate extreme oli- gotrophy for the period investigated. These conclusions are consistent with those of Bussoon & NoEl (1991) who interpreted nannoconids as dinoflagellate cysts proliferating in nutrient-poor waters. In the latter conditions, radiolarians are present but are much less abundant than during the high fertility episode.
Carbon isotope analyses on bulk rock and/or organic matter of the Livello Selli from other coeval Italian sections and from sections outside Italy indicate a positive shift in a~3C of approximately 2%o; this is interpreted as a signal of 'heightened productivity' (Weissert et al., 1979; Weissert, 1989; Arthur et al., 1990; Weissert & Lini, 1991) supporting the high fertility hypothesis. A consequence of the high fertility event was the accumulation of organic matter of marine origin (Pratt & King, 1986). Moreover, layers rich in well preserved, entire fish have been noted in other Umbria-Marche sections of the Livello Selli (Coccioni et al., 1987, 1989). This finding supports the hypothesis that the oxygen content at the sea floor was very low, and that carbonates, whenever produced in the surface water, underwent strong dissolution at the sediment/water interface. Therefore, the Livello Selli is possibly the product of high fertility combined with poor oxygenation at the bottom. It is the local expression of what is known in the literature as OAE la (Oceanic Anoxic subevent la) (Arthur et al., 1990), a globally recognized event (Sliter, 1989a).
The Livello Selli is very similar to the LiveUo Bonarelli of latest Cenomanian age in fossil content, litho- and sedimentological characteristics (Coccioni et al., 1987, 1989), high total organic carbon, and in showing a positive shift in a13C (Schlanger et al., 1987; Weissert, 1989). The depositional sequences of the Livello Selli and the LiveUo Bonarelli are surprisingly similar in vertical trend. Both are preceded and followed by similar 'critical intervals' (Piergiovanni, 1989; Coccioni et al., 1991), although the Livello Bonarelli differs in that (1) the most oligotrophic planktonic foraminifera of the late Cenomanian, the rotaliporids, become extinct at the Livello Bonarelli, whereas no major extinctions occur within planktonic foraminifera at the Livello SeUi, and (2) calcareous nannofossil fertility indices did not undergo any drastic changes within the Bonarelli (Bralower, 1988). According to Arthur et al. (1990) the latest Cenomanian OAE2 event (=Livello Bonarelli) is interpreted as being due to an episode of increased productivity. This supports our interpretation that the Livello Selli is also due to a high fertility event probably accompanied by poor oxygenation at the bottom, which favoured the preservation of fish and of organic matter.
7. Conclusions
(a) The first diversification among planktonic foraminifera predates the Aptian; correlation with ammonites has shown that it occurred mainly during the Barremian.
(b) The biostratigraphic event that best approximates the Barremian-Aptian boundary is the FO of Rucinolithus irregularis.
(c) Chron M0 is slightly younger than the FO of R. irregularis confirming it to be very close to the Barremian-Aptian boundary.

534 R. Coccioni et al.
(d) Among planktonic foraminifera the general trend is toward an increase in diversity and abundance, and overall size of individuals.
(e) An important change in plankton distribution patterns occurs within the G. bIowi foraminiferal Zone and C. litterarius nannofossil Zone, where planktonic foraminifera, radiolarians, and calcareous nannofossils start to show large-scale, higher frequency fluctuations in abundance; fluctuations in the three groups are out of phase and may be cyclically induced.
(f) These new patterns are interrupted by the Selli Event (--OAE la of Arthur et al., 1990) which is preceded and followed by 'critical intervals'. The distribution and composition of planktonic foraminifera, calcareous nannoflora and radiolarians lead us to interpret the 'critical intervals' as periods of increased nutrient content in the surface water and the Livello Selli as a high fertility event.
(g) Among planktonic foraminifera the hedbergellids seem to indicate a more eutrophic habitat than the globigerinelloids.
(h) The abrupt crisis affecting the nannoconids in approaching the Livello Selli seems to indicate that nannoconids are characteristic of more oligotrophic condi- tions, whereas Zygodiscus erectus is related to a more eutrophic environment.
(i) Based on our data, the early Aptian Livello Selli has the same palaeoceano- graphic significance as the latest Cenomanian Livello BonareUi; that is, it represents a high fertility event on a global scale.
List of species
Planktonic foraminifera Globigerinelloides blowi (Bolli, 1959) Globigerinelloides duboisi (Chevalier, 1961) Globigerinelloides ferreolensis (Moullade, 1961) Globigerinelloides gottisi (Chevalier, 1961) Globigerinelloides hoterivica (Subbotina, 1953) Hedbergella delrioensis (Carsey, 1926) Hedbergella occulta Longoria, 1974 Hedbergella sigali Moullade, 1966 Hedbergella similis Longoria, 1974 Leupo/dina cabr/(Sigal, 1952) Calcareous nannofossils Assipetra infracretacea (Thierstein, 1973) Roth, 1973 Calcicalathina oblongata (Worsley, 1971) Thierstein, 1971 Chiastozygus litterarius (Gorka, 1957) Manivit, 1971 Corollithion achylosus (Stover, 1966) Thierstein, 1971 Cyclagelosphaera margerelii No~l, 1965 D/scorhabdus rotator/us (Bukry, 1969) Thierstein, 1973 Eprolithus floralis (Stradner, 1962) Stover, 1966 Fabellites oblongus (Bukry, 1969) Crux, 1982 Lithraphidites carniolensis Deflandre, 1966 Micrantholithus hoschuleii (Reinhardt, 1966) Thierstein, 1971 Nannoconus bucher/Brrnnimann, 1955 Nannoconus colomii (De Lapparent, 1931) Kampmer, 1938 N annoconus kampmeri Br~innimann, 1955 Nannoconus steinmannii Kampmer, 1931 N annoconus truittii Brrnnimann, 1955 Nannoconus wassallii Brrnnimann, 1955 Parhabdolithus angustus (Stradner, 1963) Stradner, Adamiker and Maresch, 1968 Prediscosphaera columnata (Stover, 1966) Manivit, 1971 Rucinolithus irregularis Thierstein in Roth and Thierstein, 1972 Rucinolithus terebrodentarius Applegate, Bralower, Covington and Wise, in Covington and Wise, 1987 Watznueria barnesae (Black in Black and Barnes, 1959) Perch-Nielsen, 1968 Zygodiscus erectus (Deflandre in Deflandre and Fert, 1954) Bukry and Bramlette, 1969

Barremian-Aptian calcareous plankton biostratigraphy 535
Acknowledgements
We warmly thank M. Caron and J. Mutterlose for their critical review of the manuscript, advice and support. Thanks are extended to M. B. Cita and D. Rutledge who also improved the paper with invaluable suggestions. The authors are deeply indebted to Dr Ph. Hoedemaker for providing the samples and ammonite zonation of the Rio Argos section (Spain).
The work was supported by Ministero Pubblica Istruzione, 40% to I.P.S. and 60% to R.C.
References
Aguado, R., Company, M., O'Dogherty, L., Sandoval, J. & Tavera, J. M. 1990. Biostratigraphical analysis of the pelagic Barremian-Aptian in the Betic Cordillera (Southern Spain): Preliminary data. IGCP 262--Tethyan Cretaceous Correlation, 2nd Pelagic and Flysch Facies Meeting, Krakow 1990, Abstracts, 9.
Aguado, R., Company, M., O'Dogherty, L., Sandoval, J. & Tavera, J. M. 1992. Biostratigraphical analysis of the pelagic Barremian-Aptian in the Betic Cordillera (Southern Spain): Preliminary data. Cretaceous Research 13, 445-452.
Applegate, J. L. & Bergen, J. A, 1988. Cretaceous calcareous nannofossil biostratigraphy of sediments recovered from the Galicia Margin, ODP Leg 103. In Proceedings of Ocean Drilling Program, Scientific Results (eds Boillot, G., Winterer, E. L. et al.) 103, 293-348 (College Station, TX Ocean Drilling Program).
Arthur, M. A. & Premoli Silva, I. 1982. Development of widespread organic carbon-rich strata in the Mediterranean Tethys. In Nature and origin of Cretaceous carbon-rich facies (eds Schlanger, S. O. & Cita, M. B.), pp. 7-54 (Academic Press, London).
Arthur, M. A., Jenkyns, H. C., Brumsack, H. J. & Schlanger, S. O. 1990. Stratigraphy, geochemistry, and paleoceanography of organic carbon-rich Cretaceous sequences. In Cretaceous resources, events and rhythms (eds Ginsburg, R. N. & Beaudoin, B.), pp. 75-119 (Kluwer Academic Publishers, Dordrecht).
Baccelle, L. & Bosellini, A. 1965. Diagrammi per la stima visiva della composizione percentuale nelle rocce sedimentarie. Annali dell' Univem~ di Ferrara IX, 1, 3, 59-62.
Bralower, T. J. 1987. Valanginian to Aptian calcareous narmofossil stratigraphy and correlation with the upper M-sequence magnetic anomalies. Marine Micropaleontology 11, 293-310.
Bralower, T. J. 1988. Calcareous nannofossil biostradgraphy and assemblages of the Cenomanian- Turonian boundary interval: implications for the origin and timing of oceanic anoxia. Paleoceanography 3, 275-316.
Busson, G. & N~I , D. 1991. Les nannoconidrs, indicateurs environnementaux des ocrans et mers ~picontinentales du Jurassique terminal et du Cr~tac6 inf~rieur. Oceanologica Acta 14, 333-356.
Caron, M. 1985. Cretaceous planktic foraminifera. In Plankton stratigraphy (eds Boili, H. M., Saunders, J. B. & Perch-Nielsen, K.), pp. 17-86 (Cambridge Earth Science Series, Cambridge University Press, Cambridge).
Caron, M. & Homewood, P. 1983. Evolution of early planktic foraminifers. Marine Micropaleontology 7, 453-462.
Channel.[, J. E. T., Lowrie, W. & Medizza, F. 1979. Middle and Early Cretaceous magnetic stratigraphy from the Cismon section, Northern Italy. Earth and Planetary Science Letters 42, 153-166.
Channell, J. E. T. & Erba, E. 1992. Early Cretaceous polarity chrons CM0 to CMll recorded in Northern Italian land sections near Brescia (Northern Italy). Earth and Planetary Science Letters, 108, 161-179.
Coccioni, R., Erba, E. & Premoli Silva, I. 1991. Litho- and biostratigraphy of the Livello Bonarelli close to the Cenomanian/Turonian boundary (Umbria-Marche Apennines, Italy) and possible paleoceano- graphic significance. Colloque International sur les Evknements de la limite C~nomanien-Turonien, Grenoble 1991, G~logie Alpine, M~moire H. S. 17, 25-26.
Coccioni, R., Franchi, R., Nesci, O., Wezel, C. F., Battistini, F. & Pallecchi, P. 1989. Stratigraphy and mineralogy of the Selli Level (early Aptian) at the base of the Marne a Fucoidi in the Umbrian-Marchean Apennines (Italy). In Cretaceous of the Western Tethys, 3rd International Symposium. Tiibingen 1987 (ed. Wiedmann, J.), pp. 563-584 (Schweizerbart, Stuttgart).
Coccioni, R., Nesci, O., Tramontana, M., Wezel, C. F. & Moretti, E. 1987. Descrizione di un livello guida "Radiolaritico-Bituminoso-Ittiolitico" alla base delle Marne a Fucoidi nelrAppennino Umbro-Marchigiano. Bollettino della Societfz Geologica Italiana 106, 183-192.
Cresta, S., Monechi, S. & Parisi, G., 1989. Stratigrafia del Mesozoico e Cenozoico nell'area Umbro-Marchigiana (Mesozoic-Cenozoic Stratigraphy in the Umbria-Marche area). Memorie descrittive della Carta Geologica d' Italia 39, 1-185.

536 R. Coccioni et al.
Erba, E. 1986. I Nannofossili calcarei nell'Aptiano-Albiano (Cretacico inferiore): biostratigrafia, paleoceangrafia e diagenesi degli Scisti a Fucoidi del Pozzo Piobbico (Marche). Unpublished PhD Thesis, Universit~ di Milano, 313 pp.
Erba, E. 1988. Aptian-Albian calcareous nannofossil biostratigraphy of the Scisti a Fucoidi cored at Piobbico (central Italy). Rivista Italiana di Paleontologia e Stratigrafia 94, 249-284.
Erba, E. 1989. Upper Jurassic to Lower Cretaceous Nanneconus distribution in some sections from Northern and Central Italy. Memorie di Scienze Geologiche 41, 255-261.
Erba, E. 1992. Calcareous nannofossil distribution in pelagic rhythmic sediments (Aptian-Albian Piobbico core, Central Italy). Rivista Italiana di Paleontologia e Stratigrafia 97, 455-484.
Erba, E. Middle Cretaceous calcareous nannofossils from the western tropical Pacific (ODP Leg 129): evidence for paleoequatorial crossings. Proceedings of Ocean Drilling Program, Scientific Results 129B (College Station, TX Ocean Drilling Program), in press.
Erba, E., Coccioni, R. & Premoli Silva, I. 1989a. Gli Scisti a Fucoidi nell'area Umbro-Marchigiana: Le sezioni della S. S. Apecchiese. Irf Stratigrafia del Mesozoico e Cenozoico hell'area Umbro-Marchigiana (Mesozoic-Cenozoic Stratigraphy in the Umbria-Marche area) (eds Cresta, S., Monechi, S. & Parisi, G.), M emorie descrittive della Carta Geologica d' Italia 39, 146-164.
Erba, E. & Larson, R. L. 1991. Narmofossils and superplumes. EOS, Abstract Volume for AGU Spring Meeting--Baltimore 1991, 301.
Erba, E., Premofi Silva, I., Pratt, L. M. & Tornaghi, M. E. 1989b. Aptian-Albian black shales from the "Scisti a Fucoidi" of Piobbico core (Marche, Italy). Abstracts of 28th International Geological Congress, Washington D.C. 1989 1, 455.
Erba, E. & Quadrio, B. 1987. Biostratigrafia a Nannofossili calcarei, Calpionellidi e Foraminiferi planctonici della Majolica (Titoniano supefiore-Aptiano) nelle Prealpi Bresciane (Italia Settentri- onale). Rivista Italiana di Paleontologia e Stratigrafia 93, 3-108.
Hallock, P. 1987. Fluctuations in the trophic resource continuum: A factor in global diversity cycles? Paleoceanography 2, 457-471.
Harland, W. B., Armstrong, R. L., Cox, A. V., Craig, L. E., Smith, A. G. & Smith, D. G. 1990. A geologic time scale, 263 pp. (Cambridge University Press, Cambridge).
Hoedemaeker, Ph. & Bulot, L. 1990. Preliminary ammonite zonation for the Lower Cretaceous of the Mediterranean region (Report of the Lower Cretaceous Cephalopod Team, Working Group of IGCP, Project 262). G~ologie Alpine 66, 123-127.
Larson, R. L. 1991a. Latest pulse of Earth: Evidence for a mid-Cretaceous superplume. Geology 19, 547-550.
Larson, R. L. 1991b. Geological consequences of superplumes. Geology 19, 963-966. Leckie, R. M. 1987. Paleoecology of mid-Cretaceous planktonic foraminifera: A comparison of open
ocean and epicontinental sea assemblages. Micropaleontology 33, 164-176. Leckie, R. M. 1989. An oceanographic model for the early evolutionary history of planktonic
foraminifera. Palaeogeography, Palaeoclimatology, Palaeoecology 73, 107-138. Loeblich, A. R. Jr & Tappan, H. 1988. Foraminiferal genera and their classification, 970 pp. (Van
Nostrand Reinhold Company, New York). Lowrie, W. & Alvarez, W. 1984. Lower Cretaceous magnetic stratigraphy in Umbrian pelagic limestone
sections. Earth and Planetary Science Letters 71, 315-328. Lowrie, W., Alvarez, W., Premoli Silva, I. & Monechi, S. 1980. Lower Cretaceous magnetic stratigraphy
in Umbrian pelagic carbonate rocks. Geophysical Journal of Royal Astronomical Society 60, 263-281. Monechi, S. 1981. Aptian-Cenomanian calcareous nannoplankton from some sections in the Umbrian
Apennine. Rivista Italiana di Paleontologia 'e Stratigrafia 87, 193-226. Moullade, M., Applegate, J. L., Bergen, J. A.~,Thurow, J., Doyle, P. S., Drugg, W. S., Habib, D.,
Masure, E., Ogg, J. & Taugourdeau-Lantz, J. 1988. Ocean Drilling Program Leg 103 biostratigraphic synthesis. In Proceedings of Ocean Drilling Program, Scientific Results (eds Boillot, G., Winterer, E. L. et al.) 103, 685-696 (College Station, TX Ocean Drilling Program).
Mutterlose, J. 1987. Calcareous nannofossils and belemnites as warmwater indicators from the NW German middle Aptian. Geologisches Jahrbuch 96, 293-313.
Mutterlose, J. 1989. Temperature-controlled migration of calcareous nannofloras in the northwest European Aptian. In Nannofossils and their applications (eds Crux, J. A. & van Heck, S. E.), pp. 122-142 (Ellis Horwood Ltd., Chichester).
Mutterlose, J. 1991. Das Verterilungs- und Migrations-Muster des kalldgen Nannoplanktons in der Unterkreide (Valangin-Apt) NW Deutschland,. Palaeontographica Abt. B 221, 27-152.
No~l, D. & Melguen, M. 1978. Nannofacies of Cape Basin and Walvis Ridge sediments, Lower Cretaceous to Pliocene (Leg 40). In Initial Reports ofDSDP (eds Bolli, H. M., Ryan, W. B. F. et al.) 40, 487-524 (U.S. Government Printing Office, Washington D.C.).
Piergiovanni, F. 1989. Eventi lito-biostratigrafici nella Scaglia Bianca umbro-marchigiana in connessione con l'episodio anossico del "Livello Bonarelli". Bollettino della Societ& Geologica Italiana 108, 289-314.
Pratt, L. M. & King, J. D. 1986. Variable marine productivity and high eolian input recorded by rhythmic black shales in mid-Cretaceous pelagic deposits from central Italy. Paleoceanography 1, 507-522.

Barremian-Aptian calcareous plankton biostratigraphy 537
Premoli Silva, I., Erba, E. & Tornaghi, M. E. 1989a. Paleoenvironmental signals and changes in surface fertility in mid Cretaceous C org-rich pelagic facies of the Fucoid Marls (central Italy). G~obios Mbnoire 11, 225-236.
Premoli Silva, I., Tornaghi, M. E. & Ripepe, M. 1989b. Planktonic foraminiferal distribution record productivity cycles: evidence from the Aptian-Albian Piobbico core (central Italy). Terra Nova 1, 443-448.
Roth, P. H. 1981. Mid-Cretaceous calcareous nannoplankton from the central Pacific: implications for paleoceanography. In Initial Reports of DSDP (eds Thiede, J., Vallier, T. L. et al.) 62, 471-489 (U.S. Government Printing Office, Washington D.C.).
Roth, P. H. 1984. Preservation of calcareous nannofossils and fine-grained carbonate particles in mid-Cretaceous sediments from the southern Angola Basin. In Initial Reports ofDSDP (eds Hay, W. W., Sibuet, J. C. et al.) 75, 651-655 (U.S. Government Printing Office, Washington D.C.).
Roth, P. H. 1987. Mesozoic palaeoceanography of the North Atlantic and Tethys Oceans. In North Atlantic palaeoceanography (eds Summerhayes, C. P. & Shackleton, N. J.), pp. 299-320 (Geological Society Special Publication 21).
Roth, P. H. 1989. Ocean circulation and calcareous nannoplankton evolution during the Jurassic and Cretaceous. Palaeogeography, Palaeoclimatology , Palaeoecology 74, 111-126.
Roth, P. H. & Bowdler, J. L. 1981. Middle Cretaceous calcareous nannoplankton biogeography and oceanography of the Atlantic ocean. Society of Economic Paleontologists and Mineralogists Special Publication 32, 517-546.
Roth, P. H. & Krumbach, K. R. 1986. Middle Cretaceous calcareous nannofossil biogeography and preservation in the Atlantic and Indian Oceans: implications for paleoceanography. Marine Mb:ropaleontology 10, 235-266.
Schlanger, S. O., Arthur, M. A., Jenkyns, H. C. & Scholle, P. A. 1987. The Cenomanian-Turonian Oceanic Anoxic Event, I. Stratigraphy and distribution of organic carbon-rich beds and the marine dl3C. In Marine petroleum source rocks (eds Brooks, J. & Fleet, A. J.), pp. 371-399 (Geological Society Special Publication 26).
Sigal, J. 1977. Essai de zonation du Cretac~ mediterran~en ~ l'aide des foraminiferes planctoniques. G~ologie Mdditerran~enne IV, 99-108.
Sigal, J. 1979. Chronostratigraphy and ecostratigraphy of Cretaceous Formations recovered on DSDP Leg 47B, Site 398. In Initial Reports o fDSDP (eds Sibuet, J. C., Ryan, W. B. F. et al.) 47B, 287-326 ~U.S. Government Printing Office, Washington D.C.).
Sliter, W. V. 1989a. Apt]an anoxia in the Pacific Basin. Geology 17, 909-910. Sliter, W. V. 1989b. Biostrat]graphic zonat]on for Cretaceous planktonic foraminifers examined in thin
section. Journal of Foraminiferal Research 19, 1-19. Tarduno, J. A., Sliter, W. V., Bralower, T. J., McWilliams, M., Premoli Silva, I. & Ogg, J. G. 1989.
M-sequence reversals recorded in DSDP sediment cores from the western Mid-Pacific Mountains and Magellan Rise. Geological Society of America Bulletin 101, 1306-1316.
Thierstein, H. R. 1973. Lower Cretaceous calcareous nannoplankton biostrafigraphy. Abhandlungen der Geologischen Bundesanstalt A 29, 1-52.
Thierstein, H. R. 1976. Mesozoic calcareous nannoplankton biostratigraphy of marine sediments. Mar/ne Micropaleontology 1, 325-362.
Thierstein, H. R. 1980. Selective dissolution of late Cretaceous and earliest Tertiary calcareous narmofossils: experimental evidence. Cretaceous Research 2, 2-12.
Tornaghi, M. E., Premoli Silva, I. & Ripepe, M. 1989. Lithostratigraphy and planktonic foraminiferal biostratigraphy of the Aptian-Albian "Scisti a Fucoidi" in the Piobbico core, Marche, Italy: background for cyclostratigraphy. Rivista Italiana di Paleontologia e Stratigrafia 95, 223-264.
Watkins, D. 1989. Narmoplankton productivity fluctuations and rhythmically-bedded pelagic carbonates of the Greenhorn Limestone (Upper Cretaceous). Palaeogeography, Palaeoclimatology, Palaeoecology 74, 75-86.
Weissert, H. 1989. C-isotope stratigraphy, a monitor of paleoenvironmental changes: a case study from the Early Cretaceous. Survey in Geophysics I0, 1-61.
Weissert, H. & Lini, A. 1991. Ice Age interludes during the time of Cretaceous greenhouse climate? In Controversies in modern geology (eds Muller, D. W., McKenzie, J. A. & Weissert, H.), pp. 173-191 (Academic Press, London).
Weissert, H., McKenzie, J. & Hochuli, P. 1979. Cyclic anoxic events in the Early Cretaceous Tethys Ocean. Geology 7, 147-151.





























