Embed Size (px)
Citation preview

Fakulteten för veterinärmedicin och husdjursvetenskap Institutionen för anatomi, fysiologi och biokemi
Arthrogryposis in Calves
Alexandra Odevall
Uppsala 2016
Veterinärprogrammet, examensarbete för kandidatexamen
Delnummer i serien: 2016:62


Arthrogryposis in Calves Artrogrypos hos kalv
Alexandra Odevall Handledare: Elisabeth Persson, institutionen för anatomi, fysiologi och biokemi
Examinator: Eva Tydén, institutionen för biomedicin och veterinär folkhälsovetenskap
Omfattning: 15 hp Nivå och fördjupning: grund nivå, G2E Kurstitel:Självständigt arbete i veterinärmedicin Kurskod: EX0700 Program: Veterinärprogrammet Utgivningsort: Uppsala Utgivningsår: 2016 Omslagsbild: Drawing by Alexandra Odevall, based on photos in Lee et al., 2007 Serienamn:Veterinärprogrammet, examensarbete för kandidatexamen Delnummer i serie: 2016:62 Elektronisk publicering: http://stud.epsilon.slu.se Nyckelord: Artrogrypos, kalv, nöt, medfödd missbildning, Crooked Calf Disease, lupiner, Lupinus, alkaloider, anagyrine Key words: Arthrogryposis, calf, bovine, congenital deformity, Crooked Calf Disease, lupines, Lupinus, alkaloids, anagyrine
Sveriges lantbruksuniversitet Swedish University of Agricultural Sciences
Fakulteten för veterinärmedicin och husdjursvetenskap Institutionen för anatomi, fysiologi och biokemi


TABLE OF CONTENTS
Summary 1
Sammanfattning 2
Introduction 3
Material and Methods 4
Literature Review 4Development of the Embryo 4Synepitheliochorial Placenta 6Development of the Fetus 6Arthrogryposis in Cattle 8Incidence of Arthrogryposis as a Part of Crooked Calf Disease 9Gross Pathology of Arthrogryposis 9Histology of Arthrogryposis 10Mechanisms of Arthrogryposis 10Etiology of Arthrogryposis 11Lupinus spp. as the Source of Arthrogryposis in Cattle 11
Discussion 13
References 16


1
SUMMARY
Arthrogryposis is a common congenital malformation in cattle and it affects the limbs through fibrosis of the joints leading to fixation. This is due to paralysis of the fetus because of alkaloid toxicosis in the pregnant dam during gestation day 40-70. When the fetus cannot move, the fibrosis develops in the joints and the extremities are fixated. The condition is more common in the forelimbs than the hind limbs and the hind limbs are usually not affected without forelimbs being affected. When arthrogryposis is accompanied by a combination of kyphosis, scoliosis, torticollis or cleft palate, it is most commonly referred to as Crooked Calf Disease. Arthrogryposis is accompanied by muscle atrophy and sometimes pale muscles and oedema interstitially. The organs appear normal but cavitation in the brainstem and pons medulla, hydranencephaly and a missing cerebellum has been reported. It has been reported that the neurons of the ventral horns are reduced in number and gliosis has been seen. Calves born with arthrogryposis have a hard time surviving as they cannot walk normally or feed as they cannot reach the udder. Also the symptoms worsen as the calves grow because of increased movement and pressure on the joints leading to secondary problems. A variety of lupine plants have been suggested to induce arthrogryposis in calves when ingested by the pregnant dam, especially during day 40-70 of gestation. Lupine induced arthrogryposis is mostly reported on in the United States where an incidence of up to 37% has been seen in grazing areas with an abundance of the most poisonous Lupinus plants, L. caudatus and L. sericeus. The teratogen is reported to most likely be the alkaloid toxin called anagyrine. This has been shown in several studies. Mechanisms of arthrogryposis includes the fetus being exposed to the alkaloid toxin anagyrine ingested by the dam through grazing on lupine abundant lands. The fetus needs to be exposed to the toxin for about 30 days for the congenital deformity to take form. The fetal muscles are not properly innervated because of acetylcholine receptor desensitization or underdevelopment of the innervating nerve leading to fibrosis of the joints and fixating them. The severity of arthrogryposis has been reported to be correlated to the severity of the toxicosis in the pregnant dam. It is dose dependent, although differences in effects at the same toxin level have been seen in different breeds and individuals with different body conditions. Also, the duration of the exposure on the fetus to the toxin correlates with the severity of the deformity. This is why sub-clinical toxicosis should be taken seriously. Viruses such as Akabane virus and Schmallenberg virus have also been suggested to induce arthrogryposis in cattle and are important differential diagnoses. Genetic and physiological differences as well as altered grazing periods have been suggested as means of lowering the incidence of arthrogryposis in calves and pros and cons are proposed and discussed.

2
SAMMANFATTNING
Artrogrypos är en av de vanligaste medfödda missbildningarna hos kalvar och fixerar lederna i benen genom fibros. Detta orsakas av att fostret blir paralyserat då den dräktiga kon får i sig en toxisk alkaloid under dag 40-70 av dräktigheten. När fostret inte kan röra på sina ben utvecklas fibros i lederna som då fixeras. Detta är mer vanligt i frambenen än bakbenen och bakbenen är oftast inte påverkade utan att frambenen också är det. När artrogrypos ses i samband med kyphos, skolios, torticollis eller gomspalt kallas det ofta för “Crooked Calf Disease”. Artrogrypos ses i samband med muskelatrofi och ibland blek muskelvävnad och interstitiella ödem. Organen är generellt opåverkade men ingröpning i hjärnstammen och pons medulla, hydranencefali och avsaknad av cerebellum har rapporterats. Det har också rapporterats att neuronen i de ventrala hornen är färre och glios har förekommit. Kalvar som föds med artrogrypos överlever ofta inte eftersom de inte kan gå normalt eller dia eftersom de inte når juvret. Symtomen blir även värre när kalven växer eftersom den ökade rörelsen och trycket på lederna ger sekundära skador. Flertalet olika lupinväxter har rapporterats inducera artrogrypos hos kalvar när de äts av den dräktiga kon, framför allt under dag 40-70 av dräktigheten. Lupininducerad artrogrypos har i huvudsak rapporterats om i USA där incidensen kan vara upp till 37% i områden där betesmarken har ett stort antal av de giftiga lupinerna Lupinus caudatus och Lupinus sericeus. Den teratogena substansen har rapporterats att till största sannolikhet vara alkaloidtoxinet anagyrine vilket flertalet studier visat. Mekanismen bakom artrogrypos innefattar att fostret utsätts för alkaloidtoxinet anagyrine genom att den dräktiga kon betat på lupinrika betesmarker. Fostret behöver utsättas för toxinet i cirka 30 dagar för att den medfödda missbildningen ska ta form. Fostrets muskler innerveras inte ordentligt på grund av att acetylkolinreceptorerna är desensitiserade eller av under-utvecklade nerver vilket ger fibros i lederna och slutligen fixering av dess. Hur allvarlig artrogryposen blir har rapporterats korrelera med hur allvarliga symtomen av förgiftningen i den dräktiga kon är. Den är beroende av dos även om skillnader i effekt vid samma nivå av toxinet har rapporterats för olika raser och kons kondition. Hur länge fostret utsätts för toxinet påverkar missbildningens allvarlighetsgrad varför subkliniska förgiftningar bör bli tagna på allvar. Virus så som Akabane virus och Schmallenberg virus har också rapporterats inducera artrogrypos hos nötkreatur och är därmed viktiga differentialdiagnoser. Beaktande av genetik och fysiologiska skillnader samt förändrade betesperioder har före-slagits för att minska incidensen av artrogrypos. För- och nackdelar diskuteras.

3
INTRODUCTION
Crooked Calf Disease was first reported in the 1950s and confirmed in the 1960s (Panter et al., 2007). It can be seen in sheep, cattle, horses and goats (Panter et al., 2007; Strake, 2015) but in this literature review focus will be on cattle. The plant genus Lupinus, when ingested by pregnant cows, can cause arthrogryposis in the calf (Shupe et al., 1967a) which sometimes is accompanied by kyphosis, scoliosis (Keeler et al., 1969), torticollis and cleft palate (Shupe et al., 1967a; b) and is then referred to as Crooked Calf Disease (Sprake, 2015). Arthrogryposis is one of the most common malformations of Crooked Calf Disease (Shupe et al., 1967b). Other names of arthrogryposis are Multiple Congenital Contractures, mostly used in human medicine (Swinyard et al., 1982) and Arthrogryposis Multiplex Congenita (Van Huffel et al., 1988) but I will refer to it as arthrogryposis in this study. The plant grows in early spring and flowers in early summer (Panter et al., 2007). Most species of lupines grow in the United States and they increase in abundance after disturbances such as fires, overgrazing and after rainy years. Lupines contain quinolizidine alkaloids and some piperidine alkaloids but this paper focuses on the quinolizidine alkaloid anagyrine. The etiology of arthrogryposis also includes genetics and infections by viruses of the family Bunyaviridae (Green et al., 2012; 2015; Sprake, 2015). In humans the direct cause of arthrogryposis is not known but different etiologies leading to fetal paralysis and arthrogryposis have been reported (Kowalczyk & Felus, 2014). The incidence of Crocked Calf Disease in cattle is estimated to be between 2.7 and 37.2% in Lupine abundant areas in the United States (Shupe et al., 1967b). It has been reported that the teratogen effect of Lupinus plants lies in the amount of the alkaloid anagyrine ingested by the pregnant cow and that the effect is dose dependent (Keeler, 1976; 1973a; 1984). Defects in the calves are also related to the signs of toxicity of the mother cow in Crooked Calf Disease (Keeler, 1976). Clinical signs of toxicosis include incoordination, twitching of the muscles, nervousness and fatigue (Keeler et al., 1969). The prognosis of survival of affected calves is poor (Sprake, 2015). Mechanisms of arthrogryposis development in fetuses include a reduction in number of motor neurons resulting in paresis (Van Huffel et al., 1988) and alkaloids being agonists to acetylcholine receptors leading to paralysis (Green et al., 2010; 2012). This gives a reduced locomotor capacity of the fetus and a fibrosis of the joints, leading to fixation i.e. arthrogryposis (Green et al., 2012). This literature review will address what has been reported on how Lupinus plants can cause arthrogryposis in bovine fetuses and discuss how it can be prevented.

4
MATERIAL AND METHODS
This literature study is based on searches in the databases PubMed and Web of Science. The key words arthrogryposis, crooked calf disease, lupines, toxicity, congenital deformities, teratogen and cattle were used in different combinations.
The description of bovine embryonic and fetal development is based on three books: McGeady et al., (2006), Hyttel et al., (2010) and Chapter 38 by Sprake in Large Animal Internal Medicine (Smith [ed], 2015).
LITERATURE REVIEW
Development of the Embryo
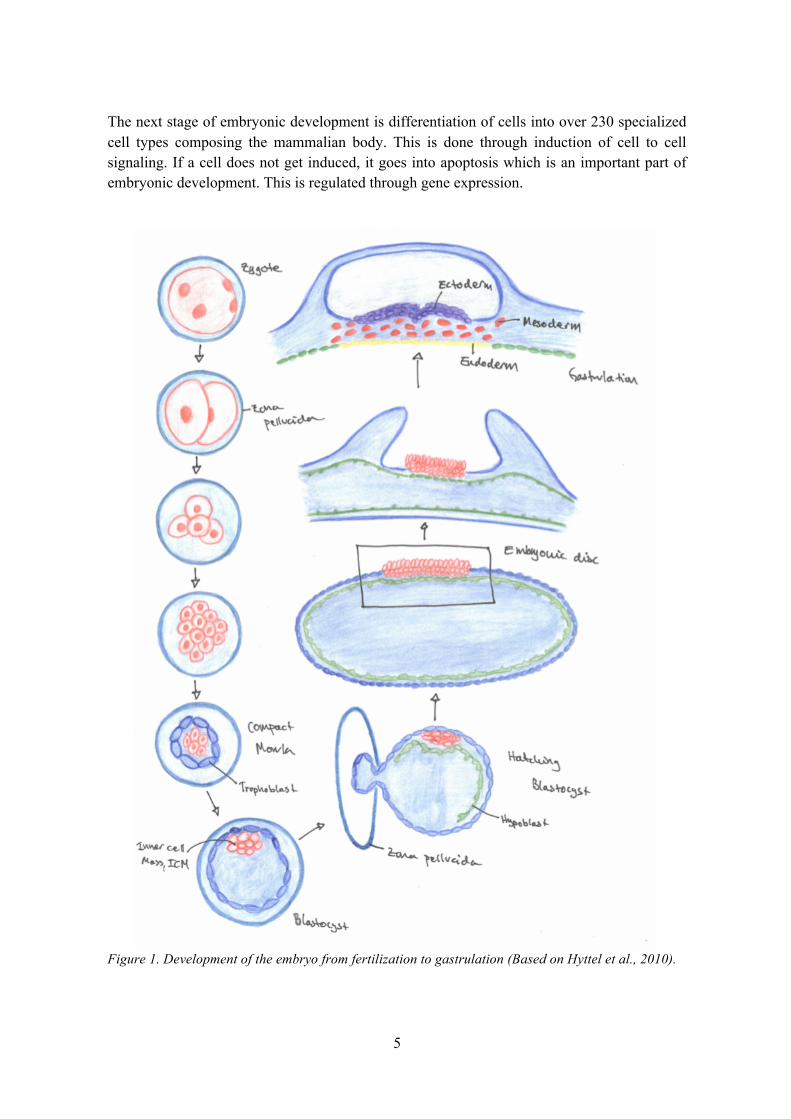
The embryonic period includes the development of major organ systems and starts when the spermatozoon fertilizes the oocyte and the zygote is formed. Here the haploid nucleus from the sperm and egg fuse into a diploid nucleus which then cleaves into two cells and then again into four cells without the cytoplasm actually increasing, and therefore the cell size decreases. These cleaved cells are called blastomeres and are surrounded by the oocyte’s zona pellucida. The process of cleavage continues a few more times until about 16-32 cells make up a small sphere which is called the morula.
The outermost cells of the morula align and adhere and are now called trophectoderm consisting of trophoblast cells, surrounding the inner cell mass, ICM/embryoblast, and the blastulation is initiated. In this stage of development, a fluid filled cavity, a blastocoel, forms within the embryo and the ICM cells are then located into one end of what is now called the blastocyst. The ICM will later form two layers of cells, the epiblast and the hypoblast. The hypoblast will in turn line the inner trophectoderm and create an enfolded space which will become the yolk sac and, dorsally, the primitive gut.
The blastocyst grows and hatches from the zona pellucida and the epiblast matures into the embryonic disc. The blastocyst is now oval in shape. The process from fertilization to oval blastocyst takes about two weeks in cattle. The blastocyst then elongates further and the embryonic disk will develop into the three somatic germ layers, ectoderm, mesoderm and endoderm. This process is called gastrulation (see Fig. 1). In this phase, cells of the embryonic disc gather together to form a streak called the primitive streak, with a cranial end, the primitive node, later giving rise to the notochord, the axial mesoderm. The epiblast dorsal to the notochord will differentiate into neuroectoderm and finally the central nervous system while the notochord itself is a base for the axial skeleton and will remain as nucleus pulposus of the intervertebral disks.
The mesoderm, originating from the primitive streak, will be found lateral to the notochord and there form arrangements called somitomeres in the posterior end which together with lateral plate mesoderm and neural crest cells develop into connective tissue, bone and cartilage of the face. In the anterior area, segmentation occurs, forming somites from which the dermis, skeletal muscle and vertebrae develop.
5
The next stage of embryonic development is differentiation of cells into over 230 specialized cell types composing the mammalian body. This is done through induction of cell to cell signaling. If a cell does not get induced, it goes into apoptosis which is an important part of embryonic development. This is regulated through gene expression.
Figure 1. Development of the embryo from fertilization to gastrulation (Based on Hyttel et al., 2010).

6
Synepitheliochorial Placenta
The bovine placenta is macroscopically described as cotyledonary, and consists of clusters of chorioallontoic villi called cotyledons that will be combined with 75-120 endometrial outgrowths, caruncles, to form placentomes. The cotyledons appose and attach to the caruncular areas, but do not invade more than having some giant trophoblast cells that pass over to the endometrial epithelium. This type of placenta is called synepitheliochorial.
Development of the Fetus
The neuroectodermal cells of the embryonic disk develop into a pseudostratified neuro-epithelium and form a setting called the neural plate. The primordium of the brain is formed through expansion of the cranial neural plate region. Caudally, it gives rise to the remaining neural tube. The lateral edges of the neural plate form the neural folds and a groove is formed in the midline, the neural groove. Folding of the neural plate now turns it into a V-shaped structure and proliferation leads to meeting and fusing, creating the neural tube. The ends of the neural tube are called the rostral and caudal neuropores, the last areas to fuse.
The neuroectodermal cells are exposed to different concentrations of signaling molecules depending on their position in the dorsal-ventral axis of the neural tube. Ventrally located cells differentiate into ventral neurons because of high exposure. As the neural tube fuses, neural crest cells are released and migrate to form sections of cells along each side of the neural tube and will eventually differentiate into, and form, the dorsal root ganglia, autonomic ganglia and glial cells of the peripheral nervous system.
The neural tube now consists of three layers. An inner ventricular layer giving rise to coating of the brain ventricles and the canal in the center of the spinal cord. The intermediate layer later develops into interneurons, that relay sensory impulses, and motor neurons. The gray matter of the spinal cord is also formed from the intermediate layer and grooves form inside the wall of the central neural canal expand through cell division and fuse. The third layer is the outer, marginal layer, containing neural processes, becoming the white matter of the spinal cord.
The central canal of the neural tube now becomes smaller in size and a medial groove on the ventral side of the spinal cord now develops and is called ventral fissure. Lateral horns are formed from dividing neuroblasts in the thoracic lumbar region and differentiate into motor neurons and form parts of the autonomic nervous system. Neural crest cells give rise to the dorsal root ganglia which is the sensory part of the peripheral nervous system. Microglia cells originate from mesodermal mesenchyme but not until vascularization is complete in the nervous system.
Axons grow out from each segment of the spinal cord into the vertebral canal. The part of the spinal cord located between the fifth and seventh cervical vertebrae increases in size and fills the vertebral canal. This is the result of an increase in neuron number when the developing thoracic limbs are to be stimulated by nerves. A similar process is seen in the area of the developing hind limbs.
7
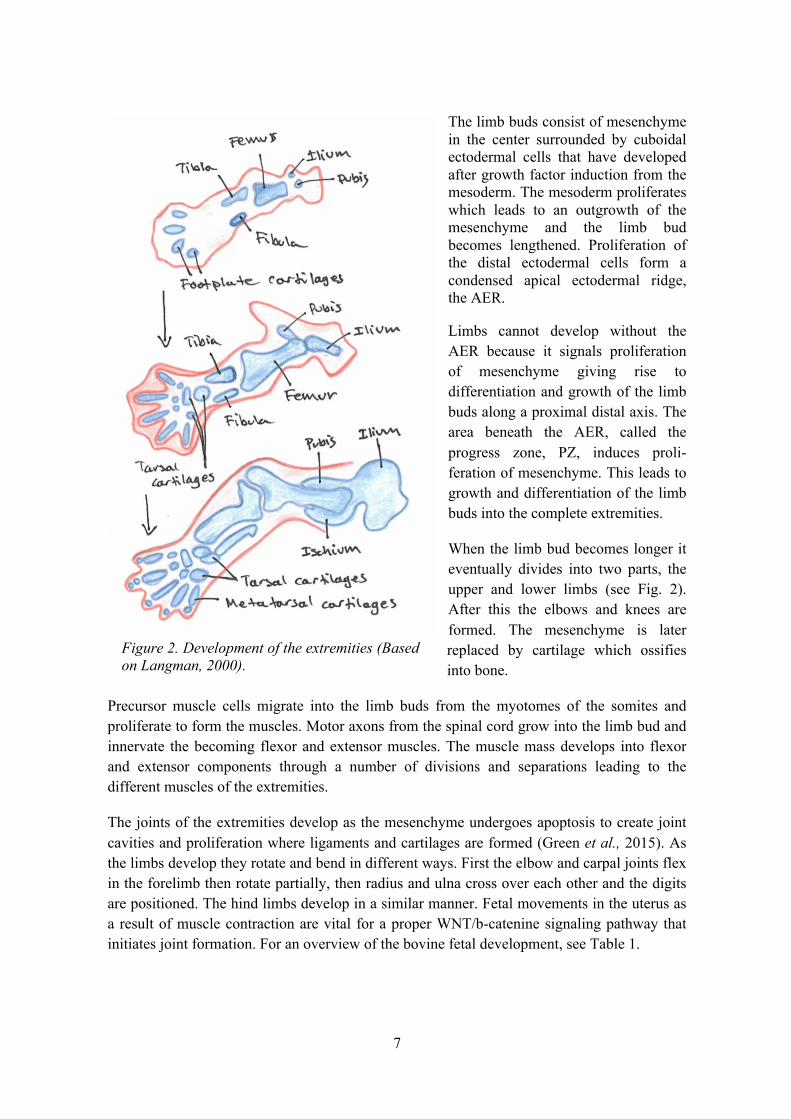
The limb buds consist of mesenchyme in the center surrounded by cuboidal ectodermal cells that have developed after growth factor induction from the mesoderm. The mesoderm proliferates which leads to an outgrowth of the mesenchyme and the limb bud becomes lengthened. Proliferation of the distal ectodermal cells form a condensed apical ectodermal ridge, the AER.
Limbs cannot develop without the AER because it signals proliferation of mesenchyme giving rise to differentiation and growth of the limb buds along a proximal distal axis. The area beneath the AER, called the progress zone, PZ, induces proli-feration of mesenchyme. This leads to growth and differentiation of the limb buds into the complete extremities.
When the limb bud becomes longer it eventually divides into two parts, the upper and lower limbs (see Fig. 2). After this the elbows and knees are formed. The mesenchyme is later replaced by cartilage which ossifies into bone.
Precursor muscle cells migrate into the limb buds from the myotomes of the somites and proliferate to form the muscles. Motor axons from the spinal cord grow into the limb bud and innervate the becoming flexor and extensor muscles. The muscle mass develops into flexor and extensor components through a number of divisions and separations leading to the different muscles of the extremities.
The joints of the extremities develop as the mesenchyme undergoes apoptosis to create joint cavities and proliferation where ligaments and cartilages are formed (Green et al., 2015). As the limbs develop they rotate and bend in different ways. First the elbow and carpal joints flex in the forelimb then rotate partially, then radius and ulna cross over each other and the digits are positioned. The hind limbs develop in a similar manner. Fetal movements in the uterus as a result of muscle contraction are vital for a proper WNT/b-catenine signaling pathway that initiates joint formation. For an overview of the bovine fetal development, see Table 1.
Figure 2. Development of the extremities (Based on Langman, 2000).

8
Table 1. Embryonic development of bovine fetuses with focus on the extremities at specific days of gestation (Hyttel et al., 2010)
Arthrogryposis in Cattle
Arthrogryposis is a common congenital malformation in cattle and it affects the forelimbs more often than the hind limbs (Hartley et al., 1974; Shupe et al., 1967a; Sprake, 2015; Andersson et al., 2008). It has been reported in many different breeds (Hartley et al., 1974) such as Hereford, Angus, Charolais, Shorthorn, Holstein-Friesian, Guernsey, Jersey, Ayshire, Brown Swiss and mixed breed (Greene et al., 1973). There do not seem to be any relation between occurrence of the defect and animals of a certain age or if the cow was inseminated or naturally bred (Hartley et al., 1974).
The toxin acts on the musculoskeletal system (Greene et al., 1973; Hartley et al., 1974) and it is a congenital deformation of the joints in fore and/or hind limbs defined as permanent joint contracture (Hartley et al., 1974; Greene et al., 1973; Shupe et al., 1967a). Arthrogryposis is not seen in the hind limbs unless the forelimbs also are affected (Hartley et al., 1974). The affected limbs can not be straightened (Hartley et al., 1974; Sprake, 2015) and may include incorrect articular positioning or rotational deformity (Sprake, 2015). The defect is mostly seen as bilateral in the fore limbs and less often in the hind limbs (Hartley et al., 1974).
Bovine congenital arthrogryposis due to poisoning is commonly induced during the summer months of the year i.e. around June through October (Hartley et al., 1974; Panter et al., 2007).
The prognosis of survival of affected calves is poor (Sprake, 2015) because most of them will not be able to follow the cow, nurse or even stand (Hartley et al., 1974; Shupe et al., 1967b).
Day Developmental stage 1 Fertilization 2-5 Morula 7-12 Blastocyst
16 Primitive Streak
19 Initiation of placentation 20 Neural groove and the first somite pairs are set 24 Forelimb buds (Motor axons grow into the limb bud and innervate) 26 Hind limb buds (Motor axons grow into the limb bud and innervate) 30 Paddle shaped forelimb buds 34 Forelimb digits 38 Hind limb digits 60 Hoof differentiation 83 Hoof confirmation 150 Hard Hoofs 159-230 Fur 279-290 Parturition

9
The condition of arthrogryposis worsens over time with the calf growing, due to secondary effects caused by increased stress of weight gain and increased moving around of the animal (Keeler et al., 1969; Shupe et al., 1967b). Also, the calves with severe defects of Crooked Calf Disease are forced to walk on their carpal joints due to the deformities (Keeler et al., 1976).
Incidence of Arthrogryposis as a Part of Crooked Calf Disease
In 1967 the incidence of Crocked Calf Disease was estimated to be between 2.7 and 37.2% in pregnant cows grazing in lupine abundant areas in the United States or being fed Lupinus sericeus. A survey taken over a 4-year period by four cattle associations showed that 2.7% of the calves in this area were born with malformations and Crooked Calf Disease only appeared in offspring from pregnant cows feeding in graze lands abundant in Lupinus. Exchange of the cattle and grazing lands was made and the disorder persisted in the calves born in the areas with lupines. This suggests that Crooked Calf Disease is caused by Lupinus containing teratogens rather than being a hereditary congenital malformation in cattle (Shupe et al., 1967b).
When examining calves born from dams fed either Lupinus caudatus or extract thereof, two out of four calves from cows fed with the plant itself had severe arthrogryposis and one out of three cows that had been fed the extract gave birth to one set of arthrogryposis in twin offspring (Keeler et al., 1969). The dams giving birth to calves with arthrogryposis showed clinical signs of toxicosis (Keeler et al., 1969; Lee et al., 2008). Clinical signs of toxicosis include incoordination, twitching of the muscles, nervousness and fatigue (Keeler et al., 1969). The study of 2008 was produced to find out if there was a difference in absorption and elimination of anagyrine in cattle with different body conditions. This will be discussed more closely in “Lupinus spp. as the Source of Arthrogryposis in Cattle”.
Gross Pathology of Arthrogryposis
Arthrogryposis is associated with neurogenic muscle atrophy (Hartley et al., 1974) which is observed in the extremities of calves with arthrogryposis (Hartley et al., 1974; Shupe et al., 1967b). This may be the result of no or decreased movement by the fetus (Green et al., 2012; 2015; Sprake, 2015) during pregnancy due to a reduced quantity of muscle nerves which leads to abnormal shortening of the muscle tissue. It was not specified if the actual muscle shortened or if this was a relative shortening do to lack of growth compared to other tissue. This makes the tissue unable to stretch and movement is hindered (Sprake, 2015). In the uncommon cases of muscle atrophy not being seen, the tissue is more pale than usual and with interstitial oedema (Hartley et al., 1974).
A close examination of one elbow joint in a calf showed that the positioning of the ulna against the articular surface of the distal humerus was incorrect and movement therefore prohibited. These deviations are permanent (Shupe et al., 1967b) and the ligaments of affected joints in four calves were not properly aligned when examined during necropsy (Keeler et al., 1969).

10
The internal organs of calves with congenital arthrogryposis appear normal when necropsied (Keeler et al., 1969) although rare cases of cavitation in the brainstem and pons medulla and a missing cerebellum have been seen. Associated hydranencephaly has also been reported (Hartley et al., 1974). One study of calves with Crooked Calf disease showed no primary myopathy nor gross lesion in the brain stem and spinal cord but the study included only eight calves and therefore it could not be ruled out that primary myopathy is a source of Crooked Calf disease. (Abbot et al., 1986).
Histology of Arthrogryposis
Different degrees of muscle atrophy in extremities with arthrogryposis have been reported and some normal muscle cells were seen within the atrophied muscle tissue. Interstitial oedema is not uncommon and in some cases the muscle tissue was substituted by adipose tissue (Hartley et al., 1974).
The cells of the ventral horn of the spinal cord are reported to be reduced in number (Hartley et al., 1974; Sprake, 2015) and with diffuse gliosis. In addition, perivascular cuffing in the brain, cord, leptomeninges, dorsal root ganglia and muscles, was seen. The remaining cells of the ventral horn were normal in structure except for a few showing a red cytoplasm and pyknotic cell nuclei, hinting anoxic necrosis. (Hartley et al., 1974).
The border of the white and gray matter was diffuse and reduced numbers of myelinated nerves were found in the ventral and lateral columns, where also an increased amount of fibrous tissue was seen (Hartley et al., 1974).
Mechanisms of Arthrogryposis
Absence of normal movement of muscles and tendons around the joints of the fetus leads to fibrosis of the joint capsule and periarticular ligaments and fixates it in the passive position of the bones (Witters et al., 2002). Segment C6 of the spinal cord delivers nerve fibers to some muscles in the forelimb. A noteworthy reduction of α-motor neurons in segment C6 was seen in calves with arthrogryposis when compared to calves born without deformities (Van Huffel et al., 1988). If the α-motorneurons are largely missing the function of the muscle tissue will also fail accordingly or not function properly. It was suggested that the decrease of this type of neuron in C6 results in paresis in the paired muscle of the shoulder area and elbow in the forelimbs which leads to persistent flexure by the M. triceps brachii giving rigidity to the affected joints. Alternatively, the dysfunction leads to extensor muscles in distal limb not being innervated properly and therefore paralyzed, giving arthrogryposis (Van Huffel et al., 1988). It was not shown that primary myopathy is the etiology of arthrogryposis (Abbot et al., 1986).

11
Etiology of Arthrogryposis
The etiology of arthrogryposis includes genetics, infection by viruses of the Bunyaviridae family (Akabane virus, Schmallenberg virus and even Bluetongue virus has been mentioned) and ingestion of toxic alkaloids of the cow during gestation (Green et al., 2012; 2015; Sprake, 2015). The genus Lupinus, when ingested during day 40-70 of the pregnancy, can cause arthrogryposis (Shupe et al., 1967a; Sprake, 2015).
In humans, the direct cause of arthrogryposis is not known but different etiologies leading to fetal paralysis and arthrogryposis have been reported (Kowalczyk & Felus, 2014). Among these are: motor center diseases, disorders of the peripheral nerves or neuromuscular junction, myogenic factors, diseases of surrounding tissue and abnormalities of the fetal environment such as maternal disease, mechanical factors or nutritional/vascular disorders.
Akabane virus is a virus of the genera Orthrobunya which has a strong teratogenic effect on fetuses of cattle, sheep and goats but is asymptomatic in the mother (Kirkland, 2015). It is vector borne (Culicoides) and crosses the placenta. The developmental deformities of the calf vary with gestation stage but the effect of the virus is the most damaging in the middle of the gestation period. The development of the central nervous system is usually the most susceptible and arthrogryposis and hydranencephaly are the malformations in the calf seen most often but these are usually secondary. When the dam is infected between day 105 and 170, arthrogryposis will be seen in the calf. The virus is common in Australia, Southeast Asia, the Middle East and some countries in Africa.
Another virus that has been reported to give arthrogryposis in bovine fetuses is the Schmallenberg virus, also an Orthrobunya virus (Wernike et al., 2015). It was discovered in Germany in 2011 and has since then spread quickly throughout Europe and has been identified in Sweden. The virus is vector borne (Culicoides) teratogen which affects ruminants and sheep more than cattle. It can lead to stillbirth, abortion or congenital deformations but the mother shows no symptoms or vague symptoms such as fever, diarrhea and a few days of reduced milk production. When infected around day 30 to 90 of pregnancy the fetal development is at highest risk and the results are varying degrees of malformations in the central nervous system and skeletal muscles, consistent with arthrogryposis but not all infected dams give birth to calves with deformities.
Lupines, Loco weed and lathyrus all give similar malformations (Keeler et al., 1969). One article suggests selenium or selenium combined with manganese deficiency as a teratogen leading to arthrogryposis (Leipold et al., 1973) but this was later disproved.
Lupinus spp. as the Source of Arthrogryposis in Cattle
Lupines are common in western United States (Panter et al., 2007; Lee et al., 2008) and most articles reporting arthrogryposis come from different parts of this area.
Epidemiologic evidence and feeding trials show that the teratogen effect of Lupinus plants lies in the amount of the alkaloid anagyrine ingested by the pregnant cow (Keeler 1976; Keeler, 1984) with about 30mg/kg inducing severe effects of Crooked Calf Disease (Keeler,

12
1984). For moderately deformed calves a dose of 6.5-11.9 mg/kg anagyrine is enough but even the small amount of about 1.44 g/kg can be enough (Keeler 1973a; 1984).
In the 1970s it was concluded that if an alkaloid toxin is responsible for congenital arthrogryposis it is most likely anagyrine ingested at day 40-70 of gestation, this was later proven in 2015 (Keeler 1973a; b; Keeler 1976; Keeler et al., 1976; Shupe et al., 1967a; Green et al., 2015;). Evidence also shows that even a small amount of anagyrine would give a toxic effect in the fetus (Keeler 1973a; Green et al., 2015). Some articles also show, through alkaloidal preparations and extracts, that Lupinus plants such as L. caudatus (Fig. 3) and L. sericeus (Fig. 4) rich in the alkaloid anagyrine does in fact produce calves with Crooked Calf Disease (Keeler 1976; Keeler et al., 1976; Green et al., 2015).
A longer feeding period of about 30 days was needed without interruption, for the deformity to have time to develop in the fetus (Keeler 1973a; Keeler 1976; Lee et al., 2008; Green et al., 2012).
The different doses of anagyrine ingested by pregnant cows give rise to different degrees of deformities. This shows that that Crooked Calf Disease induced by various Lupinus species containing the alkaloid anagyrine is dose dependent (Keeler 1976; Green et al., 2015). Defects in the calves are not automatically related to the signs of toxicity of the mother cow (Shupe et al., 1967a) although this was shown to be the case in one study where symptoms of toxicity in the mother cow was correlated to the severity of the deformities of the calves in Crooked Calf Disease, not specifying arthrogryposis (Keeler 1976; Panter et al., 2007).
Differences between bovine breeds regarding fetal movement due to lupine alkaloids have been shown in Angus vs Holstein (Green et al., 2015). Holsteins have a higher speed of elimination and lower serum concentrations than Angus when it comes to teratogenic alkaloids derived from Lupines. At the same time, it was shown that fetal activity in the Angus cattle was lower than in Holstein which also correlated with increased serum concentrations of anagyrine.
Lupinus sericeus and Lupinus caudatus seem to be more teratogenic (Keeler et al., 1969; 1976; Keeler 1984) than other Lupinus plants such as L. argeneteus and L. holosericeus. The plant concentration of anagyrine decreased as the plant matured (Keeler et al., 1969, 1976, Keeler et al., 1976) indicating that a dam in gestation day 40-70 is at greater risk feeding on younger Lupinus plants (Keeler 1984).
Figure 3. L. caudatus. Photo: Brent Miller 2011.

13
Alkaloid profiles of lupines may differ which can be due to the plants not belonging to the same species or that evolution has led to differentiation (Stephen et al., 2007). This was found in L. sulferus and L. leucophyllus. Another reason for this was suggested to be hybridization with other plant species but they all had the teratogen alkaloid anagyrine in common.
Differences in body condition of different individuals can also alter the character of effects by toxic alkaloids of L. leucophyllus when ingested (Lee et al., 2008). Cows with higher body condition have an almost three times higher mean serum concentration of anagyrine than cows with a low body condition. Absorption and elimination was also faster in cows with the higher body condition. This could in turn lead to an impact on the incidence of Crooked Calf Disease.
Teratogenic plant alkaloids can be agonists to the nicotinic acetylcholine receptor which is a ligand
gated cat ion channel that mediates the effect of acetylcholine in excitatory neurotransmission in the peripheral and central nervous system (Green et al., 2010; 2012). At first, the alkaloids stimulate the receptors but after a longer persistence the receptors become desensitized and the cat ions are inhibited leading to paralysis and eventually respiratory failure in the cow.
When ingested by a pregnant cow the alkaloid accumulates in the blood of the fetus since it is more acidic than the mothers. The fetus gets paralyzed, leading to arthrogryposis and other malformations such as cleft palate (Green et al., 2012). This was confirmed with ultrasound and cell based experiments.
The sub-clinical toxicosis of the dam should be considered as serious for the fetus because of the length of paralysis being critical to the development of arthrogryposis (Green et al., 2012).
DISCUSSION
Arthrogryposis in cattle can be due to viral infections and genetic causes but more commonly alkaloid poisoning from pregnant ruminant dams ingesting certain Lupinus plants. It is dose dependent and the severity of the symptoms of the dam corresponds to the severity of the arthrogryposis in the calves. Also, differences in sensitivity to this alkaloid toxicosis have been seen when comparing the bovine Holstein and Angus breeds and when comparing different body conditions.
Breed differences in sensitivity to the teratogenic alkaloid anagyrine found in toxic Lupinus plants suggest that a breeding program could result in certain breeds becoming less
Figure 4. L. sericeus. Photo: Tab Tanney 2011.

14
susceptible to lupine toxicosis. This in turn could lead to a lower incidence of arthrogryposis in calves in areas where these lupines are abundant. These cows could then graze the lands where lupines are abundant regardless of gestation day and the incidence of arthrogryposis would decrease and be kept at a low level or perhaps even be eliminated completely. Through these types breeding strategies cows can be genetically altered to withstand the toxin but this may take several generations.
Different body conditions of pregnant dams have also been shown to be relevant to the outcome of arthrogryposis in calves. Animals with lower body conditions have a lower serum concentration of anagyrine but also a slower absorption and elimination. This may reflect a lower ability to detoxify alkaloid toxins which in the case of pregnancy would increase the exposure period for the fetus. As the duration of exposure is an important factor, this could in turn have effects on the paralysis of the fetus and the outcome of the deformity. Before letting the pregnant cows out to graze in areas abundant in toxic lupine plants, the body condition should therefore be as strong as possible to avoid longer periods of sub-clinical toxicosis in the pregnant dam grazing in these areas. In this way the incidence could be lowered even further.
The condition of the cow may also influence it to graze on certain plants and the amount of time it grazes on this particular plant. Weak animals may eat plants they would not eat if their body condition was higher. If an animal has nothing else to eat or is starving it may lead to consumption of plants it would otherwise not eat. This could be important to take into account when letting cattle graze at lupine abundant areas.
Another way of avoiding a high incidence could be to change the time of grazing for pregnant cows to periods of the year when the lupines are not sprouting and early in growth, since these have been shown to be more toxic. This is in theory the easiest way to avoid arthrogryposis occurring but might not be possible to execute practically.
The differences in etiology of arthrogryposis in humans compared to the etiology in cows may be due to the relevance to the situation at hand. It would be irrelevant for research to be done on causes leading to arthrogryposis in calves that are not common in the bovine species. This might be the reason for the abundance in articles on grazing land and viruses as etiology. In cattle herds, calves born with this malformation are slaughtered and measures might be taken at an epidemiological level rather than at individual level which is the case in human babies.
The differences in day of gestation when anagyrine induces arthrogryposis and when one of the two viruses reported on inducing it differ with a couple of weeks. This could be due the size of the etiological substance and its possibility to pass the placenta. Also, the induction time could give rise to different degrees of severity of the deformity in the calf. It has been reported that arthrogryposis can be induced before and after the critical period of day 40-70 but only with moderate result.
The risk of arthrogryposis due to anagyrine poisoning in pregnant cows in Sweden seems to be minimal as it has not been reported on extensively. Jybe (1994) mentions lupines as a plant

15
of significant veterinary interests in Sweden but no information is given on the effects of it. In my opinion, it would rather be a Schmallenberg virus outbreak that would lead to arthrogryposis becoming a problem in Sweden today, as this virus has lead to outbreaks before. However, there is an awareness in Sweden about lupines as a risk for animal health as it is mentioned on the website of the National Veterinary Institute (Statens Veterinär-medicinska Anstalt, SVA, 2016)

16
REFERENCES
Abbott, L. C., Finnell, R. H., Chernoff, G. F., Parish, S. M. & Gay, C. C. (1986). Crooked calf disease: a histological and histochemical examination of eight affected calves. Veterinary Pathology, 23(6), pp 734–740.
Anderson, D. E., Desrochers, A. & St Jean, G. (2008). Management of tendon disorders in cattle. The Veterinary Clinics of North America. Food Animal Practice, 24(3), pp 551–566, viii.
Green, B. T., Lee, S. T., Panter, K. E. & Brown, D. R. (2012). Piperidine alkaloids: human and food animal teratogens. Food and Chemical Toxicology: An International Journal Published for the British Industrial Biological Research Association, 50(6), pp 2049–2055.
Greene, H. J., Leipold, H. W., Huston, K. & Guffy, M. M. (1973a). Bovine congenital defects: arthrogryposis and associated defects in calves. American Journal of Veterinary Research, 34(7), pp 887–891.
Green, B. T., Lee, S. T., Panter, K. E., Welch, K. D., Cook, D., Pfister, J. A. & Kem, W. R. (2010). Actions of piperidine alkaloid teratogens at fetal nicotinic acetylcholine receptors. Neurotoxicology and Teratology, 32(3), pp 383–390.
Greene, H. J., Leipold, H. W., Huston, K. & Guffy, M. M. (1973b). Bovine congenital defects: arthrogryposis and associated defects in calves. American Journal of Veterinary Research, 34(7), pp 887–891.
Green, B. T., Panter, K. E., Lee, S. T., Welch, K. D., Pfister, J. A., Gardner, D. R., Stegelmeier, B. L. & Davis, T. Z. (2015). Differences between Angus and Holstein cattle in the Lupinus leucophyllus induced inhibition of fetal activity. Toxicon: Official Journal of the International Society on Toxinology, 106, pp 1–6.
Hartley, W. J. & Wanner, R. A. (1974). Bovine congenital arthrogryposis in New South Wales. Australian Veterinary Journal, 50(5), pp 185–188.
Hyttel, P., Sinowatz, F., Vejlsted, M. & Betteridge, K. (2010). Essentials of domestic animal embryology. Edinburgh; New York: Saunders Elsevier. ISBN 978-0-7020-2899-1.
Jybe, K. (1994). En översikt över giftiga svenska växter av veterinärmedicinsk betydelse. En litteraturstudie. Sveriges lantbruksuniversitet, Inst. för farmakologi och toxikologi. Veterinärprogrammet (Fördjupningsarbete)
Keeler, R. F. (1973a). Lupin alkaloids from teratogenic and nonteratogenic lupins. I. Correlation of crooked calf disease incidence with alkaloid distribution determined by gas chromatography. Teratology, 7(1), pp 23–30.
Keeler, R. F. (1973b). Lupin alkaloids from teratogenic and nonteratogenic lupins. II. Identification of the major alkaloids by tandem gas chromatography-mass spectrometry in plants producing crooked calf disease. Teratology, 7(1), pp 31–35.
Keeler, R. F. (1976). Lupin alkaloids from teratogenic and nonteratogenic lupins. III. Identification of anagyrine as the probable teratogen by feeding trials. Journal of Toxicology and Environmental Health, 1(6), pp 887–898.
Keeler, R. F. (1984). Teratogens in plants. Journal of Animal Science, 58(4), pp 1029–1039.
Keeler, R. F., Binns, W., James, L. F. & Shupe, J. L. (1969). Preliminary investigation of the relationship between bovine congenital lathyrism induced by aminoacetonitrile and the lupine induced crooked calf disease. Canadian Journal of Comparative Medicine: Revue Canadienne De Médecine Comparée, 33(2), pp 89–92.
Keeler, R. F., Cronin, E. H. & Shupe, J. L. (1976). Lupin alkaloids from teratogenic and nonteratogenic lupins. IV. Concentration of total alkaloids, individual major alkaloids, and the teratogen anagyrine as a function of plant part and stage of growth and their relationship to crooked calf disease. Journal of Toxicology and Environmental Health, 1(6), pp. 899–908.

17
Keeler, R. F., James, L., Shupe, J. & Vankampen, K. (1977). Lupine-induced crooked calf disease and a management method to reduce incidence. Journal of Range Management, 30(2), pp. 97–102.
Kirkland, P. D. (2015). Akabane virus infection. Revue Scientifique Et Technique (International Office of Epizootics), 34(2), pp 403–410.
Kowalczyk, B. & Feluś, J. (2016). Arthrogryposis: an update on clinical aspects, etiology, and treatment strategies. Archives of medical science: AMS, 12(1), pp 10–24.
Langman, J. (2000). Langman’s medical embryology. 8. ed. / T.W. Sadler. Philadelphia: Lipppincott Williams & Wilkins. ISBN 978-0-683-30650-7.
Lee, S. T., Cook, D., Panter, K. E., Gardner, D. R., Ralphs, M. H., Motteram, E. S., Pfister, J. A. & Gay, C. C. (2007). Lupine induced ”crooked calf disease” in Washington and Oregon: identification of the alkaloid profiles in Lupinus sulfureus, Lupinus leucophyllus, and Lupinus sericeus. Journal of Agricultural and Food Chemistry, 55(26), pp 10649–10655.
Lee, S. T., Panter, K. E., Pfister, J. A., Gardner, D. R. & Welch, K. D. (2008). The effect of body condition on serum concentrations of two teratogenic alkaloids (anagyrine and ammodendrine) from lupines (Lupinus species) that cause crooked calf disease. Journal of Animal Science, 86(10), pp 2771–2778.
Leipold, H. W., Huston, K., Hulbert, L. C., Guffy, M. & Dennis, S. M. (1974). Congenital syndrome in Hereford calves with kyphoscoliosis, arthrogryposis and palatoschisis. The Cornell Veterinarian, 64(1), pp 123–135.
McGeady, T. A., Quinn, P.J., FitzPatrick E.S., Ryan M.T, (2006a). Foetal membranes. In: Veterinary embryology. Iowa: Blackwell Publishing, p. 74.
McGeady, T. A., Quinn, P.J., FitzPatrick E.S., Ryan M.T. (2006b). Muscular and skeletal systems. In: Veterinary embryology. Iowa: Blackwell Publishing, pp.184-185, 196-202.
McGeady, T. A., Quinn, P.J., FitzPatrick E.S., Ryan M.T. (2006c). Nervous System. In: Veterinary embryology. Iowa: Blackwell Publishing, 153-161.
Panter, K. E. & James, L. F. (1990). Natural plant toxicants in milk: a review. Journal of Animal Science, 68(3), pp 892–904.
Panter, K. E., Gardner, S. T., Lee, J. A, Pfisher, M. H., Ralphs, B. L., Stegelmeier, James, L. F. (2007). Important poisonous plants of the United States. In: Gupta, R. C. (ed) Veterinary Toxicology: Basic and Clinical Principles. Academic Press Elsevier, pp. 837-842.
Sinowatz, F. (2010). Musculo-skeletal system. In: Hyttel et al., Domestic Animal Embryology. Edinburgh; New York: Saunders Elsevier. ISBN 978-0-7020-2899-1. Chapter 16.
Shupe, J. L., Binns, W., James, L. F. & Keeler, R. F. (1967a). Lupine, a cause of crooked calf disease. Journal of the American Veterinary Medical Association, 151(2), pp 198–203.
Shupe, J. L., James, L. F. & Binnis, W. (1967b). Observations on crooked calf disease. Journal of the American Veterinary Medical Association, 151(2), pp 191–197.
Statens veterinärmedicinska anstalt, SVA (2016). Lupin. http://www.sva.se/djurhalsa/fodersakerhet/giftiga-vaxter-a-o-fodersakerhet/lupin [2016-03-20].
Sprake, P. (2015). Diseases of the bones, joints and connective tissues. In: Smith, B.P. (ed), Large Animal Internal Medicine. St. Louis: Elsevier Mosby, p. 1106.
Van Huffel, X., Weyns, A., Van Nassauw, L., Cockelbergh, D. & De Moor, A. (1988). Decreased number of alpha-motoneurons in the cervical intumescence of calves with arthrogryposis multiplex congenita of both thoracic limbs. Veterinary Research Communications, 12(2-3), pp 237–243.
Vejlsted, M. (2010a). Cellular and molecular mechanisms in embryonic development. In: Hyttel, P., Sinowatz, F., Vejlsted, M. & Betteridge, K. Domestic Animal Embryology. USA: Saunders Elsevier, 13-14, 16.

18
Vejlsted, M. (2010b). Comparative placentation. In: Hyttel, P., Sinowatz, F., Vejlsted, M. & Betteridge, K. Domestic Animal Embryology. USA: Saunders Elsevier, 108, 111-112.
Vejlsted, M. (2010c). Embryo cleavage and blastulation. In: Hyttel, P., Sinowatz, F., Vejlsted, M. & Betteridge, K. Domestic Animal Embryology. USA: Saunders Elsevier, 70-72.
Vejlsted, M. (2010d). Gastrulation, body folding and coelom formation. In: Hyttel, P., Sinowatz, F., Vejlsted, M. & Betteridge, K. Domestic Animal Embryology. USA: Saunders Elsevier, Chapter 7.
Wernike, K., Elbers, A. & Beer, M. (2015). Schmallenberg virus infection. Revue Scientifique Et Technique (International Office of Epizootics), 34(2), pp 363–373.
Witters, I., Moerman, P. & Fryns, J.-P. (2002). Fetal akinesia deformation sequence: a study of 30 consecutive in utero diagnoses. American Journal of Medical Genetics, 113(1), pp 23–28.





























