Embed Size (px)
Citation preview

Journ
alof
Cell
Scie
nce
ARNT controls the expression of epidermaldifferentiation genes through HDAC- and EGFR-dependent pathways
E. Douglas Robertson, Lynda Weir, Malgorzata Romanowska, Irene M. Leigh and Andrey A. Panteleyev*Centre for Oncology and Molecular Medicine, Division of Medical Sciences, College of Medicine, Dentistry and Nursing, Ninewells Hospital,University of Dundee, UK
*Author for correspondence ([email protected])
Accepted 22 March 2012Journal of Cell Science 125, 3320–3332� 2012. Published by The Company of Biologists Ltddoi: 10.1242/jcs.095125
SummaryPreviously we showed that spatial and developmental modulation of ARNT (HIF1b) expression in mouse epidermis is essential formaintenance of keratinocyte differentiation, proper formation of the barrier and normal desquamation. Here, using lentiviral suppression
or induction of ARNT in TERT-immortalized (N-TERT) and HaCaT cells we assessed the nature and mechanisms of ARNTinvolvement in control of differentiation in human epidermal keratinocytes. ARNT depletion did not affect the levels of basal keratinsK5 and K14, but significantly induced expression of several key differentiation markers (an effect abolished by EGF supplementation).
Furthermore, ARNT deficiency resulted in the downregulation of amphiregulin (AREG) – the most highly expressed EGFR ligand inhuman keratinocytes – whereas upregulation of ARNT showed the opposite. In ARNT-deficient monolayer cultures and 3D epidermalequivalents, the downregulation of AREG was concurrent with a decline of EGFR and ERK1/2 phosphorylation. TSA, a potent
suppressor of HDAC activity, abolished the effects of ARNT deficiency, implying a role for HDACs in ARNT-dependent modulation ofthe AREG–EGFR pathway and downstream epidermal genes. Total HDAC activity was significantly increased in ARNT-depleted cellsand decreased with ARNT overexpression. ARNT-dependent shifts in HDAC activity were specifically attributed to significant changesin the levels of HDAC1, HDAC2 and HDAC3 proteins (but not mRNA) in both monolayer and 3D cultures. Collectively, our results
suggest that ARNT controls AREG expression and the downstream EGFR–ERK pathway in keratinocytes, at least in part, bymodulating HDAC activity. This novel regulatory pathway targeting advanced stages of epidermal differentiation might have importantimplications for skin pathology such as psoriasis, atopic dermatitis and cancer.
Key words: Amphiregulin, Differentiation, EGFR, Epidermis, Epigenetics, ERK1/2, Filaggrin, Keratinocyte
IntroductionThe aryl hydrocarbon receptor nuclear translocator (ARNT or
HIF1b) is a basic helix-loop-helix protein that belongs to a
Period–ARNT–single-minded (PAS) protein family. PAS
proteins act as heterodimeric transcription factors activated by
environmental stress with ARNT being a master dimerization
partner in this protein family. Most characterised are the
heterodimeric complexes of ARNT with aryl hydrocarbon
receptor (AhR) and hypoxia-inducible factors (HIF-aproteins), formed upon AhR or HIF-a activation by
halogenated aromatic hydrocarbons or hypoxia, respectively
(Wenger and Gassmann, 1997; Beischlag et al., 2008). ARNT is
also essential for normal development and its knockout in mice
leads to early embryonic lethality due to abnormal angiogenesis,
cardial dysfunction, defective haematopoiesis and cranial
malformations (Kozak et al., 1997; Maltepe et al., 1997).
Possible roles for ARNT acting as a homodimer independent of
HIF1a and AhR have also been suggested (Sogawa et al., 1995;
Swanson and Yang, 1999).
Given the wide-ranging role of ARNT in adaptation to
environmental stress (Gu et al., 2000), its activity is likely to
be implicated in control of adaptive responses in normal skin and
in pathogenesis of certain environmentally mediated skin
disorders. Recently, we and others showed an essential role for
epidermal ARNT in mouse skin development, keratinocyte
differentiation, barrier function and dermal vascularisation
(Takagi et al., 2003; Geng et al., 2006; Wondimu et al., 2012).
In contrast to the existing view that ARNT is ubiquitously present
in mammalian tissues, we found that its expression in mouse and
human skin is subject to significant developmental and spatial
changes. In the epidermis, ARNT expression gradually declines
through the spinous layer and is totally excluded from the
granular layer suggesting a negative regulatory link between
ARNT activity and advanced steps of keratinocyte terminal
differentiation (Geng et al., 2006). We also showed that
alterations in the structure and function of the epidermis in
K14-ARNT KO mice are associated with elevated expression of a
number of differentiation markers mapped to the epidermal
differentiation complex on mouse chromosome 3. Abnormal
epidermal differentiation in these mice was also accompanied by
deregulation of certain ligands for the epidermal growth factor
receptor (EGFR) (Geng et al., 2006).
EGFR belongs to a family of ErbB tyrosine kinase receptors.
Upon activation by one of its ligands such as amphiregulin
3320 Research Article

Journ
alof
Cell
Scie
nce
(AREG), beta-cellulin (BTC), epidermal growth factor (EGF),epiregulin (EREG), epithelial mitogen (EPGN), heparin-binding
EGF-like growth factor (HBEGF), and transforming growthfactor alpha (TGFa) EGFR forms homo- or a heterodimer (withanother ErbB family member) leading to its autophosphorylation(Yarden and Schlessinger, 1987). This triggers many downstream
signalling cascades including the protein kinase C and Ras-activated ERK1/2 MAP kinase pathways (Roberts and Der, 2007;Getsios et al., 2009), with the later considered as a hallmark of
EGFR activity (Schlessinger, 2000). EGFR-mediated signaltransduction results in effects on cell proliferation, migrationand differentiation in various tissues and organs including the
skin (Yarden and Sliwkowski, 2001; Pastore et al., 2008). In theepidermis, dividing keratinocytes of the basal layer are typifiedby elevated activity of EGFR (Wang et al., 2006), whichmaintains their sustained proliferation (Hansen et al., 2000; Jost
et al., 2000). In turn, the inhibition of EGFR blocks proliferationand induces terminal differentiation of epidermal keratinocytes(Peus et al., 1997; Getsios et al., 2009) associated with induction
of differentiation markers such as K1 and K10 (Poumay andPittelkow, 1995). Furthermore, EGFR signalling is essential fornormal development of the epidermis and its appendages
(Luetteke et al., 1994; Miettinen et al., 1995; Sibilia andWagner, 1995; Hansen et al., 1997). The activation of EGFR isalso linked to pathogenesis of certain skin diseases such as
psoriasis (Nanney et al., 1986) and squamous cell carcinoma(Uribe and Gonzalez, 2011) while disruption of the EGFRpathway inhibits development of papillomas and carcinomasfrom immortalized epidermal keratinocytes (Woodworth et al.,
2000). In wound healing, EGFR plays a positive role bypromoting keratinocyte proliferation and migration (Repertingeret al., 2004).
The transition of epidermal keratinocytes through the definedstages of differentiation is associated with tightly regulatedexpressional shifts. Recently, evidence has begun to emerge
demonstrating a key role of epigenetic mechanisms (e.g. histonemodifications) in these regulations (Markova et al., 2007; Fryeet al., 2007; Sen et al., 2008; Ezhkova et al., 2009; LeBoeuf et al.,2010; Fessing et al., 2011). It was previously reported that in mouse
placenta, differentiation of trophoblast cells is controlled by ARNTthrough modulation of HDAC activity (Maltepe et al., 2005). Inaddition, class I HDAC1 and HDAC3, as well as class II HDAC4
and HDAC6, were shown to interact with HIF1a and enhanceHIF1a/ARNT-mediated transactivation (Qian et al., 2006; Kimet al., 2007). Then, it was shown that altered HDAC1 activity plays
a central role in Cyp1a1 expression driven by AhR – anotherconventional ARNT dimerization partner (Schnekenburger et al.,2007). Although these findings do not implicitly show direct
ARNT–HDAC interaction, they suggest that HDACs are mediatingdownstream effects of ARNT activity.
Here, using N-TERT and HaCaT human epidermal keratinocytecell lines with variable level of ARNT expression and 3D
epidermal equivalent models developed using these cells, weshow that ARNT deficiency positively regulates expression ofkeratinocyte differentiation markers through suppression of the
EGFR ligand AREG and concomitant downregulation of EGFRactivity and ERK1/2 phosphorylation. ARNT overexpressionshows opposite effects. This pathway is controlled through
modulation of HDAC activity, which appears to be the first stepin this novel ARNT-dependent regulatory cascade targetingadvanced stages of keratinocyte differentiation programme.
ResultsModulation of ARNT activity in N-TERT andHaCaT keratinocytesUsing transduction with lentiviral particles created from SigmaMission shRNA plasmids we developed two N-TERT
keratinocyte cell lines with stable suppression of ARNT (N-KO1 and N-KO2; Fig. 1A–D). 3D equivalents developed usingARNT-depleted keratinocytes (N-KO) showed an evident decline
in ARNT protein compared to control models (Fig. 1F).
Transduction of N-TERT keratinocytes with the lentiviral
vector pCDH1-MCS1-EF1-copGFP (al Yacoub et al., 2007)containing the full ORF fragment of ARNT cDNA resulted in asignificant increase of ARNT expression at both mRNA and
protein levels (Fig. 1C,E). This N-OXP cell line showed agradual loss of vector-retaining cells (up to 50% by day 44 ofculture; supplementary material Fig. S2). Nevertheless, 3Depidermal equivalents developed using N-OXP keratinocytes
showed at least twice the level of ARNT protein as compared tocontrol models even at the time of 3D culture completion (day 42post-transduction; Fig. 1E,G) thus confirming that gradual loss of
ARNT overexpressing cells does not impede the use of thissystem as a valid model. Of note, ARNT transduction resultednot only in a general increase of ARNT protein level in 3D
cultures but also in its ectopic appearance in the granular layer ofARNT-OXP epidermal equivalents (Fig. 1G).
In HaCaT keratinocytes, ARNT was effectively suppressed by
ARNT shRNA (H-KO1 and H-KO2 cell lines; supplementarymaterial Fig. S1A,B), or transiently upregulated with an ARNT/GFP-tagged expression vector (H-OXP1, H-OXP2; supplementary
material Fig. S1C,D).
Depletion of ARNT promotes keratinocyte differentiationWhile qPCR revealed no changes in the mRNA level of basal
keratins K5 and K14, differentiation-associated keratins K1 andK10, transglutaminase 1 (TGM1) and involucrin showed a 2.5- to3.5-fold increase in ARNT-depleted N-TERT (N-KO) cells.Transcription of late differentiation markers filaggrin and
loricrin was upregulated in these cells by 30 and 70 foldrespectively (Fig. 2A). Western blot (WB) analysis of N-KOkeratinocytes demonstrated a corresponding increase in involucrin,
TGM1 and K1/10 at the protein level (Fig. 2B, for quantificationsee supplementary material Fig. S3F). Filaggrin and loricrinproteins were not detected in N-KO cells cultured in monolayer,
even though their mRNA levels were significantly induced byARNT depletion. However, in 3D epidermal equivalents filaggrinprotein was easily detected. WB results revealed a prominent
increase of both filaggrin and K10 proteins in ARNT-depleted 3Dcultures while in ARNT-overexpressing epidermal equivalents wefound a substantial reduction of K10 and fully processed (40 kDa)filaggrin monomers (Fig. 2C, for quantification see supplementary
material Fig. S3G). Importantly, supplementation of culturemedium with EGF abolished the stimulatory effects of ARNTknockout on transcription of differentiation markers with the only
exception being K10 (Fig. 2D).
3D epidermal equivalents developed using ARNT-depleted
cells showed significant increase in the number of cornifying celllayers along with retention of nuclei (parakeratosis) as comparedto control equivalents (Fig. 2F, H&E, arrows). Additionally,
ARNT-deficient 3D culture models were characterized by areduced number of granule-containing cells and a loss of granularlayer contiguity (Fig. 2F, H&E).
ARNT in epidermal differentiation 3321

Journ
alof
Cell
Scie
nce
Immunohistochemistry (IHC) on paraffin sections also
revealed noticeable changes in the expression of several key
differentiation markers in N-KO epidermal equivalents (Fig. 2F).
In the control models, filaggrin was localized to the upper
suprabasal layers corresponding to the granular layer of the
normal epidermis. In N-KO models, filaggrin was increased in
the uppermost layers (Fig. 2F, Filaggrin) and also became
apparent in some immediate suprabasal cells suggesting
premature induction of its translation or processing
(supplementary material Fig. S2E, arrows). K10 was excluded
from the basal layer of the control epidermal equivalents and
showed low positivity in the uppermost layers being most
prominent in the spinous cells overlaying the basal layer (ADD
Fig. 2F, Keratin 10, control, arrows). In N-KO 3D models, the
overall level of K10 was markedly increased (confirmed by WB,
Fig. 2C) in all suprabasal layers including the uppermost
‘cornifying’ layers which were only weakly positive in controls
(Fig. 2F, Keratin 10, N-KO2).
Finally, involucrin expression in control cultures was confined
to the cytoplasm of suprabasal keratinocytes with most prominent
positivity in the granular layer (Fig. 2F, Involucrin, arrows).
ARNT-deficient 3D models showed significant increase of
involucrin positivity, which appeared even in the nuclei of
basal and spinous keratinocytes.
ARNT-overexpressing 3D cultures revealed no apparent
differences in morphology as compared to control epidermal
equivalents. At the same time, they appeared to be slightly thinner
than control models (supplementary material Fig. S2C,D), while
ARNT-deficient 3D epidermal equivalents were usually thicker
compared to controls (Fig. 2F). This effect seems to be related to a
negative correlation between the level of ARNT and the size of
keratinocytes rather than to variations in the number of cell layers
in corresponding 3D cultures.
ARNT deficiency has a negative effect upon
keratinocyte growth
As shown by direct cell counting (Neubauer hemocytometer), in
EGF-depleted medium, N-KO cells have a markedly reduced
growth rates as compared to the control. This effect of ARNT
deficiency was abolished when the medium was supplemented
Fig. 1. Modulation of ARNT level in N-TERT keratinocyte cell line and in 3D epidermal equivalents. (A) ARNT depletion in N-TERT cells by lentiviral
transduction as demonstrated by qPCR (n52). (B) ARNT depletion at protein level in ARNT-KO N-TERT cells shown by western blot (n52).
(C) Table summarising efficiency of ARNT depletion or overexpression at mRNA and protein levels in lentivirus-transduced ARNT-KO and ARNT-
overexpressing N-TERTs. (D) ARNT immunocytofluorescence in N-KO and control N-TERT keratinocytes. (E) Expression of ARNT in ARNT-OXP N-TERT
keratinocyte (N-OXP) cultures and corresponding 3D models. ARNT level (mRNA and protein) was assessed at different time points post-transduction (PT) as
detailed in supplementary material Fig. S2B. For both qPCR and WB, n52. (F) ARNT immunohistochemistry in control and ARNT-depleted 3D epidermal
equivalents (N-TERT cells). (G) ARNT immunohistochemistry in ARNT-overexpressing and control 3D epidermal equivalents.
Journal of Cell Science 125 (14)3322

Journ
alof
Cell
Scie
nce
with EGF (Fig. 2E). The negative effect of ARNT deficiency
upon keratinocyte growth was validated by WST-1 cell
proliferation assay, which showed approximately a 25% decline
of readings (irrespective of Ca2+ concentration) in N-KO
monolayer cultures (supplementary material Fig. S3A).
Depletion of ARNT results in downregulation of amphiregulin
and suppression of EGFR activity in human keratinocytes
In line with recent findings (Stoll et al., 2010a), our data showed
that AREG is the most highly transcribed EGFR ligand in normal
(control) N-TERT keratinocytes (about 70% of GAPDH
Fig. 2. Increased expression of differentiation markers in ARNT-depleted keratinocytes. (A) Transcription of differentiation markers (qPCR) in N-KO1 cells
compared with normal N-TERT keratinocytes (n52, *P,0.05, **P,0.01; EGF-depleted medium). (B) Protein level of involucrin, K1, K10 and TGM1 (WB) in
ARNT-depleted N-KO1 cells cultured in monolyer (EGF-depleted medium). Quantification is shown in supplementary material Fig. S3F. (C) K10 and filaggrin protein
levels (WB) in 3D epidermal equivalents developed using ARNT-deficient (N-KO) and ARNT-overexpressing (N-OXP) N-TERTs. The double band of <40 kDa
corresponds to fully processed filaggrin monomer. Quantification is shown in supplementary material Fig. S3G. (D) Transcription of differentiation markers (qPCR) in
ARNT-depleted N-KO1 cells cultured in EGF-supplemented medium (n52). (E) Proliferation rates of N-KO1 and N-KO2 cells grown in EGF-depleted and EGF-
supplemented medium (direct cell counting n53, *P,0.05, **P,0.01). (F) Histology (H&E) of ARNT-deficient and control 3D epidermal equivalents (N-TERTs)
and IHC for filaggrin, K10 and involucrin. Arrows are pointing to parakeratotic nuclei (H&E) and to granular layer cells highly positive for involucrin.
ARNT in epidermal differentiation 3323

Journ
alof
Cell
Scie
nce
expression level). TGFa message was detected at the level of
about 10% of GAPDH while other EGFR ligands showed no or
negligible expression in control cells (Fig. 3A). Comparing the
level of AREG in N-KO cells with that of the controls revealed a
significant downregulation of AREG mRNA – more than 50% in
N-KO2 (Fig. 3B, shaded bars). Genes coding for other EGFR
ligands showed insignificant changes (supplementary material
Fig. S3C). The high dependence of AREG transcription on the
Fig. 3. EGFR pathway is suppressed in ARNT-depleted keratinocytes. (A) Transcription of EGFR ligands (qPCR) in normal N-TERT keratinocytes (n53).
(B) qPCR analysis of AREG expression in control, N-KO1 and N-KO2 cells without TSA (n55, *P,0.05) and with TSA treatment (n53). (C) WB analysis of
AREG in the culture medium conditioned by N-KO cells compared with control (n52) and the level of pEGFR (Tyr1173) in N-KO1, N-KO2 and control cells
(n52; EGF-depleted medium). (D) AREG protein positivity (IHC) in ARNT-overexpressing (N-OXP) and control 3D epidermal equivalents. (E) IHC analysis of
ARNT-depleted (N-KO) and control 3D epidermal equivalents showing staining for AREG, pEGFR, ERK1/2 and pERK1/2. Arrows denote regions of high
positivity. Arrowheads point to positive nuclei. (F) Immunocytofluorescence showed that pEGFR-specific signal (red) occurs as distinct foci within the cytoplasm.
Cells were grown in EGF-depleted or EGF-supplemented medium. (G) Blind test counting of the number of foci per cell for 100 cells (**P,0.01) in N-KO and
control N-TERT keratinocytes. (H) AREG mRNA level in control and ARNT-overexpressing HaCaT cells (n54, **P,0.01). (I) WB analysis of phosphorylated
and non-phosphorylated ERK1/2 in N-KO1, N-KO2 and control cells (n52; EGF-depleted medium).
Journal of Cell Science 125 (14)3324

Journ
alof
Cell
Scie
nce
level of ARNT was further confirmed by a prominent increase of
AREG message in ARNT-overexpressing HaCaT keratinocytes
(Fig. 3H).
Significantly less AREG protein was secreted into the culture
medium by N-KO cells than by control keratinocytes (Fig. 3C).
Correspondingly, in ARNT-depleted epidermal equivalents,
AREG positivity (IHC) was fainter at the cell membrane and
eradicated from the nuclei as compared to controls where strong
nuclear and cell membrane positivity was observed (Fig. 3E,
AREG). Concomitantly with downregulation of AREG in
ARNT-depleted models, we found a prominent increase of
AREG positivity in ARNT-overexpressing 3D epidermal
equivalents (Fig. 3D).
EGFR phosphorylation at Tyr1173 (pEGFR) was significantly
reduced in the N-KO cells cultured in monolayer (Fig. 3C). This
was also confirmed by immunofluorescence: blind counting of
pEGFR-positive foci performed in duplicate revealed that the
number of foci per cell in N-KO keratinocytes was reduced by two
fold as compared to the controls. This result was not apparent when
the culture medium was supplemented with EGF (Fig. 3F,G). In
the control 3D epidermal equivalents, IHC showed prominent
pEGFR-positive signal at the plasma membranes of the spinous
cells and in the cytoplasm of the uppermost (‘granular’) cell layers
(Fig. 3E, pEGFR, arrows), whereas in N-KO 3D cultures pEGFR
positivity was not observed. While ARNT deficiency significantly
affected EGFR phosphorylation, it had no effect on EGFR
transcription (supplementary material Fig. S3B).
Induction of ERK1/2 phosphorylation is a classical
downstream effect of the EGFR-Ras signalling in many cell
types including epidermal keratinocytes (Roberts and Der, 2007;
Getsios et al., 2009). As shown by WB, ARNT depletion did not
affect the total level of ERK proteins but significantly reduced
ERK1/2 phosphorylation (Fig. 3I). Decrease of pERK1/2 in N-
KO cells was also confirmed by IHC in the epidermal equivalents
(Fig. 3E). In the control 3D cultures, pERK1/2 was localised to
the uppermost layers of flattening cells (same pattern as for
pEGFR, Fig. 3E, arrows) and to the nuclei of some basal
keratinocytes (Fig. 3E, arrowheads). In N-KO equivalents,
pERK1/2 positivity was lost or significantly reduced. No
apparent difference in the level of non-phosphorylated ERK1/2
proteins between control and N-KO models was observed
(Fig. 3E, ERK1/2).
Effects of ARNT depletion upon expression of downstreamtarget genes are modulated through HDAC activity
Suppression of HDAC activity using trichostatin-A (TSA), a
potent and specific inhibitor of class I and II HDACs, eradicated
the negative effects of ARNT deficiency on the level of AREG
transcription (Fig. 3B). Furthermore, TSA treatment eliminated
the stimulatory effects of ARNT-deficiency upon transcription
of keratinocyte differentiation markers as evidenced from the
qPCR analysis of TSA-treated normal and N-KO keratinocytes
when results were compared to corresponding (control and
ARNT-deficient) untreated cell cultures (Fig. 4A). At the same
time, comparison of all three manipulated cultures (normal
TSA-treated, N-KO, TSA-treated N-KO) against untreated
normal cells revealed that differentiation markers had
different responses to TSA (Fig. 4B). Transcription of
filaggrin, K10 and loricrin in N-TERT cells with a normal
level of ARNT was significantly reduced by TSA; TGM1 and
K1 mRNA showed a moderate increase while basal keratins K5
and K14 were not responsive to TSA treatment (as shown by
both methods of comparison). Combination of TSA treatment
with ARNT deficiency resulted in suppression of the stimulating
effects of ARNT knockdown for filaggrin, K10 and loricrin
mRNA while K5 and K14 again showed no response. Despite
both TSA treatment and ARNT deficiency have a positive effect
upon TGM1 and K1 transcription, combination of these two
factors resulted in abrogation of their individual stimulatory
effects.
Fig. 4. Changes in expression of differentiation markers in ARNT-depleted cells are dependent on HDAC activity. (A) Expression of differentiation
markers (qPCR) in N-KO1 cells without or with TSA treatment compared with untreated normal cells (dashed bar) or TSA-treated normal cells (white bars)
respectively (n52, EGF-depleted medium). (B) Same qPCR as in A, but results for N-KO1 cells with or without TSA treatment and for TSA-treated control
keratinocytes were plotted against untreated control cells (n52, EGF-depleted medium).
ARNT in epidermal differentiation 3325

Journ
alof
Cell
Scie
nce
In epidermal keratinocytes, the level of HDAC proteins and
the total HDAC activity are dependent on ARNT
An HDAC activity assay showed that in ARNT-depleted N-
TERT cells total HDAC activity was increased by 15–20%. In
ARNT-depleted HaCaT keratinocytes this increase reached 50%
(Fig. 5A,B). Correspondingly, ARNT overexpression in HaCaTs
resulted in a ,50% decrease of total HDAC activity (Fig. 5B).
To identify specific HDACs accountable for these changes we
first identified HDACs with a significant level of transcription in
control N-TERT keratinocytes (Fig. 5C). In these cells, class I
HDAC1 and HDAC3 appeared to be the most prominently
expressed histone deacetylases. Among the SIRT genes (coding
for class III HDACs), only SIRT2 showed a noticeable (but still
low) level of expression. In ARNT-depleted N-KO1 and N-KO2
cells cultured in monolayer, the mRNA level of all HDAC and
SIRT genes remained unchanged as compared to the controls
(supplementary material Fig. S3D,E). However, at the protein
level, depletion of ARNT resulted in a significant increase of
HDAC1, HDAC2 and HDAC3 (Fig. 5D). In ARNT-depleted
HaCaT cells the increase of HDAC1 and HDAC3 protein levels
was even more prominent (Fig. 5E) matching the changes in total
HDAC activity (Fig. 5A,B). Overexpression of ARNT in both N-
TERT and HaCaT keratinocytes cultured in monolayer led to a
prominent drop in HDAC1, HDAC2 and HDAC3 protein levels
(Fig. 5F,G).
Immunohistochemistry in ARNT-deficient 3D epidermal
equivalents revealed an increase in HDAC1 protein in the
keratinocyte nuclei as compared to controls (Fig. 6A). In
agreement with previous reports (Longworth and Laimins,
2006), HDAC3 protein in control 3D models was seen not only
in the nuclei but in the cytoplasm and at the plasma membrane as
well. In control cultures, the relatively low (compared to
HDAC1) nuclear HDAC3 positivity was mostly confined to the
lower cell layers whilst nuclei in the upper layers were HDAC3-
negative (Fig. 6A, HDAC3, arrowheads). In the N-KO 3D
models, high cytoplasmic positivity of HDAC3 was seen in all
cells including the uppermost layers, which were HDAC3
negative in controls (Fig. 6A, arrows). Nuclear HDAC3
staining increased in the lower layers and also appeared in
suprabasal keratinocytes (Fig. 6A). Increase of HDAC1, HDAC2
and HDAC3 protein levels in ARNT-deficient 3D cultures was
confirmed by WB (Fig. 6B). Concomitantly, in ARNT-
overexpressing 3D epidermal equivalents, the level of HDAC1
was significantly reduced (Fig. 6C).
DiscussionARNT is generally thought to be a ubiquitously expressed
protein. However, we have previously demonstrated strong
developmental and differentiation-associated regulation of
ARNT in mouse epidermis, which appeared to be essential for
epidermal development, cornification, barrier formation,
desquamation and epidermal–dermal communication (Geng
et al., 2006; Wondimu et al., 2012). Here, we elucidate a novel
ARNT-dependent mechanism that controls advanced stages of
differentiation in human epidermal keratinocytes. Given the
essential role of ARNT in the activity of HIF- and AhR-
dependent pathways implicated in adaptive response to hypoxia,
organic toxicants, ozone and UV (Wenger and Gassmann, 1997;
Sogawa et al., 1995; Afaq et al., 2009; Fritsche et al., 2007) our
studies also expose a potential mechanistic link between
environmental stress and the process of keratinocyte terminaldifferentiation in normal and diseased human skin.
According to our results, suppression of endogenous ARNT
significantly affects the process of differentiation in humankeratinocytes. Altered expression of key differentiation markersin N-KO cells is concomitant with morphological changes in
ARNT-depleted 3D epidermal equivalents i.e. increased numberof cornifying layers, diminished granular layer, and parakeratosis– all signs of deregulated differentiation.
At the same time, the epidermal markers reveal differentmodes of response to variations in the level of ARNT in vitro:basal keratins K5 and K14 showed no sensitivity at all;
suprabasal differentiation markers such as K1/10, TGM1 andinvolucrin were induced by ARNT deficiency at both mRNA(moderately) and protein levels; while filaggrin and loricrin –specific markers of the granular layer – were dramatically
induced at mRNA level (approximately 10-fold that of otherdifferentiation markers) but were not reliably detected at theprotein level in control and N-KO monolayer cultures. We
assume that monolayer culture may not provide the correctmicroenvironment for normal processing of these proteins – asituation analogous with b-casein expression in mammary
epithelial cells cultured in monolayer or in 3D conditions (Liet al., 1987). In line with this assumption, in 3D epidermalequivalents (which represent a more adequate model ofdifferentiating epidermis) filaggrin protein was easily detected
(Fig. 2C). The changes between control and ARNT-depleted or -overexpressing samples were proportional for different filaggrin-positive bands – 40 kDa, 74 kDa and 120 kDa – presumably
corresponding to filaggrin monomers, dimers and trimers (Daleet al., 1985; Sandilands et al., 2009). This fact, together withdramatic changes in filaggrin mRNA level (Fig. 2A) suggests
that ARNT controls (negatively) expression of profilaggrin generather than proteolytic processing of filaggrin protein. Ectopicappearance of ARNT positivity in the granular layer of N-OXP
3D epidermal equivalents and concomitant drop in total filaggrinprotein level in these models (Fig. 2C) further support thissuggestion.
Overall, the induction of differentiation markers in ARNT-
deficient keratinocytes and their suppression in ARNT-overexpressing 3D models (Fig. 2C) correspond to the absenceof ARNT protein in the upper layers of control 3D epidermal
equivalents (Fig. 1F; Control) and to the exclusion of this proteinfrom the granular layer of normal mouse and human epidermis(Geng et al., 2006). In general, the epidermal phenotype in
ARNT-deficient mice matches our findings in ARNT-KO 3Depidermal equivalents and monolayer cultures (profoundacceleration of differentiation in both in vivo and in vitro
systems). However, there are some discrepancies as well. In the
epidermis of ARNT-KO mouse newborns we observed somereduction of loricrin and filaggrin (Geng et al., 2006) whereasARNT-deficient 3D epidermal equivalents are characterized by
profound induction of these proteins. We believe that reductionof loricrin and filaggrin – two specific granular layer proteins – inthe ARNT-deficient mouse epidermis can be attributed to abrupt
cornification and the virtual absence of a granular layer in thesemice. Involucrin, which is normally expressed not only in thegranular but also in the spinous layer, did not show a decline in
ARNT-deficient mouse epidermis but rather is increased (Genget al., 2006) as we observed in the human system (3Dequivalents). The abrupt cornification characteristic of the
Journal of Cell Science 125 (14)3326

Journ
alof
Cell
Scie
nce
Fig. 5. Regulation of total HDAC activity and specific HDACs in ARNT-depleted and ARNT-overexpressing keratinocytes cultured in monolayer.
(A) Total HDAC activity in nuclear lysates in N-KO1 and N-KO2 cells compared with controls (n52, **P,0.01). (B) Total HDAC activity in ARNT-depleted
and ARNT overexpressing HaCaT cells (n52, *P,0.05). (C) Expression of individual HDACs and SIRT deacetylases (qPCR) in control N-TERT cells (n52).
(D) Protein levels of HDAC1, HDAC2 and HDAC3 in N-KO1 and N-KO2 cells compared with control N-TERTs (n54). (E) Protein levels of HDAC1 and
HDAC3 in H-KO1 and H-KO2 cells compared with control HaCaTs (n52). (F) Protein levels of both HDAC1 and HDAC3 are significantly reduced in ARNT-
overexpressing HaCaT cells (H-OXP1 and H-OXP2). (G) Protein levels of HDAC1 and HDAC2 are reduced in ARNT-overexpressing N-TERT cells
(monolayer cultures).
ARNT in epidermal differentiation 3327

Journ
alof
Cell
Scie
nce
ARNT-null mouse epidermis is not as prominent in 3D culture
and therefore the granular layer, while abnormal, is present in this
model. This allows us to perceive the effects of ARNT depletion
upon the expression of loricrin and filaggrin. Thus, the appearing
discrepancy in expression of loricrin and filaggrin between our
previous in vivo and current in vitro results is likely attributed to
the basic developmental and structural differences between intact
epidermis (in vivo) and in vitro experimental 3D models.
Activation of the EGFR pathway by ligand binding is known
to suppress differentiation and positively affect growth in
keratinocytes (Monzon et al., 1996; Peus et al., 1997; Jost et al.,
2000; Schneider et al., 2008). Here we found that effects of ARNT
depletion upon keratinocyte differentiation (positive) and growth
(negative) are abolished by supplementing cell culture medium
with EGF thus suggesting that these effects are instigated by
downregulation of certain EGFR ligand(s). The implication of the
EGFR pathway in ARNT control over the expression of
differentiation markers is further confirmed by decreased EGFR
and ERK1/2 phosphorylation in N-KO cells and its restoration in
EGF-supplemented medium. Decrease of EGFR phosphorylation
in ARNT-depleted keratinocytes is also supported by significantly
smaller number of pEGFR-positive cytoplasmic foci, which are
presumably representing late endosomes. Previously it was shown
that EGFR endocytosis is driven by its autophosphorylation
(Waterman and Yarden, 2001). Therefore, a reduced level of
EGFR internalization (Fig. 3F) corresponds with a decline of its
phosphorylation (Fig. 3C,E) in ARNT-deficient cells.
In line with previous reports designating AREG as a principal
EGFR ligand mediating autocrine growth in epidermal
keratinocytes (Piepkorn et al., 1994; Nylander et al., 1998;
Stoll et al., 2010a) we showed that AREG is the most abundant
EGFR ligand in normal N-TERT cells. In N-KO keratinocytes
AREG declined significantly at both mRNA and protein levels
and its nuclear positivity was completely lost (Fig. 3B,C,E)
whereas other conventional EGFR ligands showed no changes
(supplementary material Fig. S3C). These findings suggest
suppression of AREG as a likely mechanistic explanation for
downregulation of EGFR pathway in ARNT-depleted cells.
Recently, it was shown that knockdown of AREG in human
epidermal keratinocytes results in reduced ERK phosphorylation,
inhibition of growth, and induction of K1/10, involucrin, loricrin
and TGM1 (Stoll et al., 2010b) – exactly the same set of changes
we observed in ARNT-depleted N-TERTs. This strongly supports
our proposition that decreased EGFR and ERK phosphorylation
and subsequent induction of differentiation markers in N-KO
cells is mediated through downregulation of AREG, but not
through modulation of EGFR expression. This view is supported
by the lack of ARNT effects upon EGFR mRNA level
(supplementary material Fig. S3B).
Loss of nuclear AREG in ARNT-depleted keratinocytes
(Fig. 3E) is of particular interest here. It has been shown
previously that the C-terminal transmembrane fragment of
AREG can be internalized and accumulated in the nucleus to
block keratinocyte differentiation (Stoll et al., 2010b). Similarly,
internalization and nuclear accumulation of the C-terminal
transmembrane fragment of another conventional EGFR ligand,
HB-EGF, leads to silencing of transcription (Nanba et al., 2003;
Toki et al., 2005). Taking these findings into account, our results
identify ARNT as an upstream positive regulator of both
membrane (EGFR activation) and nuclear functions of AREG.
The fact that induction of K10 expression in N-KO cells is not
amended by EGF supplementation suggests that, in contrast to
other differentiation markers, transcription of K10 may be
specifically regulated by nuclear C-terminal domain of AREG
Fig. 6. Level of HDAC proteins in 3D epidermal
equivalent models developed using ARNT-depleted
and ARNT-overexpressing N-TERT keratinocytes.
(A) IHC analysis of 3D epidermal equivalents showing
staining for HDAC1 and HDAC3 in control and
ARNT-depleted models. While uppermost cell layers
in control epidermal equivalents are HDAC3-negative,
in N-KO models, they are highly positive (arrows). In
control cultures, HDAC3-negative nuclei are common
(arrowheads), while they are always HDAC3-positive
in N-KO equivalents. (B) HDAC1, HDAC2 and
HDAC3 protein levels (WB) in ARNT-deficient and
control 3D models. (C) HDAC1 protein level (WB) in
ARNT-overexpressing 3D model compared
with control.
Journal of Cell Science 125 (14)3328

Journ
alof
Cell
Scie
nce
through the EGFR-independent mechanism mentioned above
(Fig. 7).
It is increasingly apparent that HDAC activity is essential for
epidermal development and differentiation (Markova et al., 2007;
Ezhkova et al., 2009; LeBoeuf et al., 2010). ARNT controls
differentiation of trophoblast cells in the mouse placenta through
modulation of HDAC activity (Maltepe et al., 2005) and it is
very likely that similar mechanisms may control epidermal
differentiation. As shown by our results, TSA treatment abolishes
effects of ARNT deficiency upon expression of AREG and
differentiation markers thus strongly implicating HDAC activity
in ARNT control over AREG–EGFR pathway and keratinocyte
differentiation. However, we found that in normal N-TERT cells
different epidermal genes respond to TSA treatment differently and,
most interestingly, the mode of their response to TSA appeared to
match the changes in their expression in response to ARNT
depletion.
As evident from Table 1, the basal keratins K5 and K14
comprise a group of epidermal genes (Group 1), which, in
contrast to differentiation markers, are not responsive to ARNT
deficiency and also show no response to TSA. Previously, based
on the studies of differentiation in mouse trophoblast it was
suggested that HDAC inhibition phenocopies ARNT deficiency
(Maltepe et al., 2005). In normal keratinocytes, we found this to
be true for TGM1 and for K1 genes only (Group 2) which both
are induced by TSA and upregulated by ARNT deficiency.
Involucrin occupies an intermediate position between Group 1
and 2 genes as it is not responsive to TSA (as Group 1) but is
positively regulated by ARNT depletion (as Group 2 genes).
Filaggrin, K10 and loricrin genes (Group 3) are strongly induced
by ARNT depletion and are significantly suppressed by TSA
treatment in N-TERT keratinocytes. Moreover, TSA treatment of
N-KO cells not only eradicates stimulatory effect of ARNT
deficiency on Group 3 genes but significantly downregulates
their expression as compared to control. Thus, for genes of Group
3, which are negatively regulated by ARNT, HDAC activity
represents a key limiting factor. Also, there is an inverse
correlation between the changes in expression of Group 3 genes
and AREG in response to all kinds of manipulation (Table 1)
further suggesting that filaggrin, K10 and loricrin genes are
negatively regulated by ARNT through HDAC- and AREG-
mediated signalling. Interestingly, HDAC1- and HDAC2-
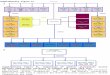
Fig. 7. A working model for the role of ARNT in the control of AREG–EGFR pathway and downstream differentiation markers. Our results suggest that
ARNT controls expression of differentiation markers through different mechanisms. First, ARNT regulates expression of filaggrin, loricrin and K10 (Group 3,
Table 1) through the modulation of the level of AREG. HDAC activity is essential for these effects. Nevertheless, the mechanisms by which HDAC1, HDAC2 and
HDAC3 are induced by ARNT deficiency and the mode of their interaction with ARNT to bring about regulation of downstream genes remain unclear (dashed
box 1). While being highly dependent on AREG, K10 is not responsive to EGF, suggesting an EGFR-independent regulation of K10 through the intracellular
C-terminal domain of AREG. This mechanism is hypothetical and requires further investigation (dashed box 2). Then, ARNT also has a negative effect upon
expression of involucrin, TGM-1 and K1 (Group 2, Table 1) but these genes show different response to TSA treatment compared with Group 3 and the
mechanisms of their regulation by ARNT remain to be elucidated (dashed box 3). Red arrows and T-bars show effects of experimental manipulation; blue arrows,
stimulatory activity; black T-bars, suppression.
Table 1. Effects of modulation of ARNT and HDAC activity
upon expression of epidermal differentiation markers
TSA Control cells TSA + ARNT-KO ARNT-KO
K5 < < <K14 < Q 2.06 <Involucrin < q 1.66 q 2.36TGM1 q 2.06 < q 2.86K1 q 1.56 q 1.66 q 2.56Flg Q 3.36 Q 2.06 q 30.06K10 Q 5.06 Q 33.36 q 3.76Loricrin Q 50.06 Q 100.06 q 68.46AREG q 1.56 q 1.36 Q 2.06
ARNT in epidermal differentiation 3329

Journ
alof
Cell
Scie
nce
knockout mice show an absence of K10 and loricrin expression in
the epidermis (LeBoeuf et al., 2010) thus supporting our
conclusion that effects of ARNT upon AREG, EGFR and
downstream differentiation markers are mediated by HDAC
activity. It was previously reported that TSA alters expression of
epidermal genes in a protein synthesis-dependent manner
(Markova et al., 2007) implying existence of an intermediate
protein regulator. Our data identify AREG as a putative
regulatory link between HDAC activity and expression of
filaggrin, loricrin and K10.
Thus, we have demonstrated that ARNT modulates
keratinocyte differentiation through HDAC- and EGFR-
dependent mechanisms. AREG appears to be the key
intermediate player linking together these two pathways. There
is evidence that PAS proteins such as AhR and HIF2a (which
both require dimerization with ARNT for their activity) control
the expression of AREG in urinal tract and cancer tissues (Choi
et al., 2006; Stiehl et al., 2011). These findings are in line with
the role of ARNT in control of AREG/EGFR pathway in the
epidermis revealed by our studies.
Importantly, while HDAC activity is essential for ARNT
effects upon AREG expression and keratinocyte differentiation,
HDACs themselves appeared to be dependent on ARNT level:
depletion of ARNT results in the prominent induction of total
HDAC activity and significant increase in HDAC1, HDAC2 and
HDAC3 protein levels. Concomitantly, upregulation of ARNT
leads to decrease of HDAC1 level (Fig. 6B,C).
All these interactions are shown on a schematic drawing
(Fig. 7) representing a putative model for ARNT control over
epidermal differentiation. This model suggests that ARNT, being
nuclear (and hence active) specifically in the lower layers of 3D
epidermal equivalents (Fig. 1F), is implicated in the deterrence
of differentiation in spinous keratinocytes. As keratinocytes
approach granular layer, ARNT is downregulated leading to an
increase in HDAC activity and suppression of AREG/EGFR
pathway thus triggering the expression of specific granular
layer proteins such as filaggrin and loricrin. This view also fits
prevalent expression of ARNT in lower layers of mouse and
human epidermis and its decline in the granular layer (Geng et al.,
2006). Our model, while instructive, still remains a subject for
further validation since the role of ARNT dimerization partners
(HIF-a proteins and AhR) in control of the ARNT-AREG-EGFR
pathway and mechanisms of ARNT control over activity of
specific HDACs are not yet clear, nor is the role of AREG in
control of K10 expression (Fig. 7, question marks). These issues
will be addressed in our future studies.
Materials and MethodsCell culture, modulation of ARNT expression and drug treatment
Human N-TERT keratinocytes immortalized with hTERT (a gift from J.
Rheinwald, Harvard Medical School, Boston, MA) were maintained in GIBCO
keratinocyte serum-free medium supplemented with BPE, Ca2+ and EGF (final
concentrations 25 mg/ml, 0.4 mM, 0.2 ng/ml, respectively) (Rheinwald and
Beckett, 1981). In experiments analysing the effect of EGF this supplement was
omitted from the media. To stably deplete ARNT in N-TERTs we used
transduction with lentiviral particles created from Sigma Mission shRNA
plasmids (SHGLY-NM_001668 and non-targeting control plasmid SHC002; seesupplementary material Table S1) as described previously (al Yacoub et al., 2007).
Successfully transduced N-TERT keratinocytes were selected for and further
maintained with 0.5 mg/ml puromycin at low Ca2+ (0.4 mM). Comparison of two
ARNT-depleted N-TERT cell lines (N-KO) with the non-targeting shRNA control
showed 60–80% decrease of ARNT mRNA (Fig. 1A,C) and 81% and 96%
decrease of ARNT protein for N-KO1 and N-KO2, respectively (Fig. 1B,C).
Immunocytochemistry demonstrated a high level of nuclear ARNT protein in
control N-TERTs and its visible decline in the nuclei of N-KO cells (Fig. 1D).
For viral overexpression of ARNT in N-TERT cells, ARNT cDNA ORF
fragment was cut out and purified from the OriGene expression vector (RG216724;
OriGene Technologies, Rockville, MD) by restriction enzyme digest with BamHIand NotI (R0136S and R0189S; NEB, Hitchin, UK). Lentiviral transfer vector
pCDH1-MCS1-EF1-copGFP (al Yacoub et al., 2007) was digested with BamHI
and NotI and the obtained ARNT fragment was ligated in. The ligation reactionwas transformed into Turbo Competent E. coli (C2984H; NEB). Resulting
ampicillin-resistant colonies were mini-prepped and checked for the correct
insertion by restriction digest and direct sequencing. These preparations (alongwith empty pCDH1-MCS1-EF1-copGFP vector for control purposes) were then
used to create lentiviral particles further utilized for transduction of N-TERT
keratinocytes (N-OXP) as described previously (al Yacoub et al., 2007). Efficiency
of transduction in N-OXP keratinocytes was monitored by FACS analysis of GFPfluorescence (supplementary material Fig. S2) using a BD Biosciences (Oxford,
UK) LSR Fortessa Flow Cytometer.
HaCaT keratinocytes (ECACC; Salisbury, UK) were transfected with ARNT
shRNA expressing vectors (Sigma; Dorset, UK) using Lipofectamine 2000 and
selected for stable transformants with puromycin. Three constructs targetingdifferent regions of the ARNT mRNA were used (supplementary material Table
S1). Cells transfected with non-targeting shRNA vector were used as a control.
The use of constructs 1 and 2 resulted in ,40 and 60% reduction of ARNT mRNA
(supplementary material Fig. S1A) and ,90% reduction of ARNT protein levelwhereas construct 3 gave only 20% reduction (supplementary material Fig. S1B).
For further studies we selected cell lines obtained using constructs 1 and 2 only (H-
KO 1 and 2). Transient over-expression of ARNT in HaCaT cells (H-OXP) usinglipofectamine-mediated transfection with an ARNT/GFP-tagged expression vector
(OriGene; Rockville, MD) resulted in ,200-5006 increase of ARNT mRNA level
(supplementary material Fig. S1C) and about 1.5- to 3.5-fold increase of total
ARNT protein (supplementary material Fig. S1D).
For suppression of total HDAC activity cells were grown in culture medium
containing 100 nM TSA for 48 hrs prior to taking lysates for protein and RNAisolation. Development of 3D epidermal equivalents using N-KO or N-OXP cell
lines resulted in formation of epidermis-like structures with significant depletion or
increase of ARNT levels, respectively, as was confirmed by IHC (Fig. 1F,G) andWB (Fig. 1E).
3D cell culture
N-TERT keratinocytes (26105) were resuspended in 400 ml PCT epidermalkeratinocyte medium (CnT-57; CELLnTEC, Bern, Switzerland) and added to
Millicell PCF 12 mm inserts with 0.4 um pore size (PIHP01250; Millipore,
Hertfordshire, UK) placed in a 60 mm Petri dish. Medium was added so that levelswere equal with that inside the insert. After formation of a confluent monolayer the
medium was replaced inside and outside the insert with 3D medium (CnT-02-
3DP1; CELLnTEC). The next day, the medium was aspirated from inside the
inserts. Inserts were left for 14 days with medium changes every three days.Membranes and attached cells were then removed from the inserts, fixed in
formalin overnight, processed through graded alcohols to xylene and embedded in
paraffin blocks. 4-mm-thick sections were cut for H&E and IHC staining using aLeica RM2135 rotary microtome.
RNA isolation and real-time PCR
RNA was isolated from cells grown to 70–80% confluency using Tri Reagent
(Sigma, Dorset, UK) according to the manufacturer’s protocol. RNA was diluted
1:10 and used in qPCR reactions with Invitrogen Superscript III Platinum SYBR
Green One-Step qRT-PCR. Reactions were run on a Bio-Rad mini-opticon real-time PCR machine as previously described (Weir et al., 2011a). Primers used are
shown in supplementary material Table S2.
Protein isolation and western blotting
Protein lysates were obtained from 70–80% confluent keratinocytes as reported
previously (Geng et al., 2006) and processed for WB (Weir et al., 2011a) usingappropriate antibodies (supplementary material Table S3).
Immunocytochemistry
Cells were cultured on 2 well permanox chamber slides (Lab-Tek; Scotts Valley,CA). For assessment of ARNT localisation cultures were fixed with acetone for
2 min at 220 C. For examination of EGFR phosphorylation, cells were fixed with
4% paraformaldehyde at RT, permeabilized with 0.1% Triton X-100 for 5 min andblocked for 30 min with 0.3% BSA in PBS blocking solution. After fixation and
blocking, cells were incubated with primary antibodies (supplementary material
Table S3) made up in either PBS or blocking solution. Washes were repeated
followed by incubation with secondary antibodies made up in PBS. Cells weremounted using ProLong Gold antifade reagent with DAPI (Invitrogen) and
examined using Zeis Axioskop2 microscope with fluorescence.
Journal of Cell Science 125 (14)3330

Journ
alof
Cell
Scie
nce
Immunohistochemistry
Deparaffinized sections of 3D epidermal equivalents were boiled in the microwavefor 363 min in 10 mM sodium citrate (pH 6.0) for antigen unmasking, incubatedwith primary antibodies (supplementary material Table S3) at RT for 1 hr withblocking and processed further using the Vectastain Elite ABC kit (pk-6200;Vector Laboratories, Peterborough, UK). Samples were developed using the DABperoxidase kit (pk-4100, Vector Laboratories) and counterstained withHaematoxylin. Samples treated with no primary antibodies were used as anegative control.
WST-1 assay and cell counting
The water-soluble tetrazolium salt (WST-1) assay, a standard method to assess cellviability and proliferation (K301-500; BioVision; distributed by CambridgeBioSciences, Cambridge, UK), was performed as described previously (Weiret al., 2011b). To induce keratinocyte differentiation in this experiment,ARNT-deficient N-TERT cells were exposed to high Ca2+ (1.4 mM) as comparedto normal cell culture conditions (0.4 mM). For direct counting, cells were releasedwith 0.05% trypsin and counted in triplicate using a Neubauer hemocytometer.Trypan Blue was used to exclude dead cells from the live cell count.
HDAC activity assay
HDAC activity was performed using a Fluorometric HDAC activity assay kit(ab1438; Abcam, Cambridge, UK). Equal amounts of cell lysate (10–50 mg) wereused and the assay was carried out as per the manufacturer’s description. Resultswere retrieved using a SpectraMax Gemini XS fluorescence plate reader(Molecular Devices, Berkshire, UK).
AcknowledgementsThe authors are grateful to Celine Pourreyron (Skin Cancer Group,University of Dundee) for her valuable help with some of thetechniques used in this study. We also acknowledge the FlowCytometry Core Facility at the College of Medicine, Dentistry andNursing at the University of Dundee for assistance with FACSanalysis.
FundingOur work was funded by Cancer Research UK [grant number C5314/A6695].
Supplementary material available online at
http://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.095125/-/DC1
ReferencesAfaq, F., Zaid, M. A., Pelle, E., Khan, N., Syed, D. N., Matsui, M. S., Maes, D. and
Mukhtar, H. (2009). Aryl hydrocarbon receptor is an ozone sensor in human skin. J.
Invest. Dermatol. 129, 2396-2403.
al Yacoub, N., Romanowska, M., Haritonova, N. and Foerster, J. (2007). Optimizedproduction and concentration of lentiviral vectors containing large inserts. J. Gene
Med. 9, 579-584.
Beischlag, T. V., Luis Morales, J., Hollingshead, B. D. and Perdew, G. H. (2008).The aryl hydrocarbon receptor complex and the control of gene expression. Crit. Rev.
Eukaryot. Gene Expr. 18, 207-250.
Choi, S. S., Miller, M. A. and Harper, P. A. (2006). In utero exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin induces amphiregulin gene expression in the developingmouse ureter. Toxicol. Sci. 94, 163-174.
Dale, B. A., Holbrook, K. A., Kimball, J. R., Hoff, M. and Sun, T. T. (1985).Expression of epidermal keratins and filaggrin during human fetal skin development.J. Cell Biol. 101, 1257-1269.
Ezhkova, E., Pasolli, H. A., Parker, J. S., Stokes, N., Su, I. H., Hannon, G.,
Tarakhovsky, A. and Fuchs, E. (2009). Ezh2 orchestrates gene expression for thestepwise differentiation of tissue-specific stem cells. Cell 136, 1122-1135.
Fessing, M. Y., Mardaryev, A. N., Gdula, M. R., Sharov, A. A., Sharova, T. Y.,
Rapisarda, V., Gordon, K. B., Smorodchenko, A. D., Poterlowicz, K., Ferone, G.
et al. (2011). p63 regulates Satb1 to control tissue-specific chromatin remodelingduring development of the epidermis. J. Cell Biol. 194, 825-839.
Fritsche, E., Schafer, C., Calles, C., Bernsmann, T., Bernshausen, T., Wurm, M.,
Hubenthal, U., Cline, J. E., Hajimiragha, H., Schroeder, P. et al. (2007).Lightening up the UV response by identification of the arylhydrocarbon receptor as acytoplasmatic target for ultraviolet B radiation. Proc. Natl. Acad. Sci. USA 104, 8851-8856.
Frye, M., Fisher, A. G. and Watt, F. M. (2007). Epidermal stem cells are defined byglobal histone modifications that are altered by Myc-induced differentiation. PLoS
ONE 2, e763.
Geng, S., Mezentsev, A., Kalachikov, S., Raith, K., Roop, D. R. and Panteleyev, A.A. (2006). Targeted ablation of Arnt in mouse epidermis results in profound defects indesquamation and epidermal barrier function. J. Cell Sci. 119, 4901-4912.
Getsios, S., Simpson, C. L., Kojima, S., Harmon, R., Sheu, L. J., Dusek, R. L.,
Cornwell, M. and Green, K. J. (2009). Desmoglein 1-dependent suppression ofEGFR signaling promotes epidermal differentiation and morphogenesis. J. Cell Biol.
185, 1243-1258.
Gu, Y. Z., Hogenesch, J. B. and Bradfield, C. A. (2000). The PAS superfamily:sensors of environmental and developmental signals. Annu. Rev. Pharmacol. Toxicol.
40, 519-561.
Hansen, L. A., Alexander, N., Hogan, M. E., Sundberg, J. P., Dlugosz, A.,Threadgill, D. W., Magnuson, T. and Yuspa, S. H. (1997). Genetically null micereveal a central role for epidermal growth factor receptor in the differentiation of thehair follicle and normal hair development. Am. J. Pathol. 150, 1959-1975.
Hansen, L. A., Woodson, R. L., 2nd, Holbus, S., Strain, K., Lo, Y. C. and Yuspa,
S. H. (2000). The epidermal growth factor receptor is required to maintain theproliferative population in the basal compartment of epidermal tumors. Cancer Res.
60, 3328-3332.
Jost, M., Kari, C. and Rodeck, U. (2000). The EGF receptor - an essential regulator ofmultiple epidermal functions. Eur. J. Dermatol. 10, 505-510.
Kim, S. H., Jeong, J. W., Park, J. A., Lee, J. W., Seo, J. H., Jung, B. K., Bae, M. K.and Kim, K. W. (2007). Regulation of the HIF-1alpha stability by histonedeacetylases. Oncol. Rep. 17, 647-651.
Kozak, K. R., Abbott, B. and Hankinson, O. (1997). ARNT-deficient mice andplacental differentiation. Dev. Biol. 191, 297-305.
LeBoeuf, M., Terrell, A., Trivedi, S., Sinha, S., Epstein, J. A., Olson, E. N.,Morrisey, E. E. and Millar, S. E. (2010). Hdac1 and Hdac2 act redundantly tocontrol p63 and p53 functions in epidermal progenitor cells. Dev. Cell 19, 807-818.
Li, M. L., Aggeler, J., Farson, D. A., Hatier, C., Hassell, J. and Bissell, M. J. (1987).Influence of a reconstituted basement membrane and its components on casein geneexpression and secretion in mouse mammary epithelial cells. Proc. Natl. Acad. Sci.
USA 84, 136-140.
Longworth, M. S. and Laimins, L. A. (2006). Histone deacetylase 3 localizes to theplasma membrane and is a substrate of Src. Oncogene 25, 4495-4500.
Luetteke, N. C., Phillips, H. K., Qiu, T. H., Copeland, N. G., Earp, H. S., Jenkins, N.A. and Lee, D. C. (1994). The mouse waved-2 phenotype results from a pointmutation in the EGF receptor tyrosine kinase. Genes Dev. 8, 399-413.
Maltepe, E., Schmidt, J. V., Baunoch, D., Bradfield, C. A. and Simon, M. C. (1997).Abnormal angiogenesis and responses to glucose and oxygen deprivation in micelacking the protein ARNT. Nature 386, 403-407.
Maltepe, E., Krampitz, G. W., Okazaki, K. M., Red-Horse, K., Mak, W., Simon, M.
C. and Fisher, S. J. (2005). Hypoxia-inducible factor-dependent histone deacetylaseactivity determines stem cell fate in the placenta. Development 132, 3393-3403.
Markova, N. G., Karaman-Jurukovska, N., Pinkas-Sarafova, A., Marekov, L. N.
and Simon, M. (2007). Inhibition of histone deacetylation promotes abnormalepidermal differentiation and specifically suppresses the expression of the latedifferentiation marker profilaggrin. J. Invest. Dermatol. 127, 1126-1139.
Miettinen, P. J., Berger, J. E., Meneses, J., Phung, Y., Pedersen, R. A., Werb, Z. and
Derynck, R. (1995). Epithelial immaturity and multiorgan failure in mice lackingepidermal growth factor receptor. Nature 376, 337-341.
Monzon, R. I., McWilliams, N. and Hudson, L. G. (1996). Suppression of cornifiedenvelope formation and type 1 transglutaminase by epidermal growth factor inneoplastic keratinocytes. Endocrinology 137, 1727-1734.
Nanba, D., Mammoto, A., Hashimoto, K. and Higashiyama, S. (2003). Proteolyticrelease of the carboxy-terminal fragment of proHB-EGF causes nuclear export ofPLZF. J. Cell Biol. 163, 489-502.
Nanney, L. B., Stoscheck, C. M., Magid, M. and King, L. E., Jr (1986). Altered[125I]epidermal growth factor binding and receptor distribution in psoriasis. J. Invest.
Dermatol. 86, 260-265.
Nylander, N., Smith, L. T., Underwood, R. A. and Piepkorn, M. (1998). Topographyof amphiregulin expression in cultured human keratinocytes: colocalization with theepidermal growth factor receptor and CD44. In Vitro Cell. Dev. Biol. Anim. 34, 182-188.
Pastore, S., Mascia, F., Mariani, V. and Girolomoni, G. (2008). The epidermalgrowth factor receptor system in skin repair and inflammation. J. Invest. Dermatol.
128, 1365-1374.
Peus, D., Hamacher, L. and Pittelkow, M. R. (1997). EGF-receptor tyrosine kinaseinhibition induces keratinocyte growth arrest and terminal differentiation. J. Invest.
Dermatol. 109, 751-756.
Piepkorn, M., Lo, C. and Plowman, G. (1994). Amphiregulin-dependent proliferationof cultured human keratinocytes: autocrine growth, the effects of exogenousrecombinant cytokine, and apparent requirement for heparin-like glycosaminogly-cans. J. Cell. Physiol. 159, 114-120.
Poumay, Y. and Pittelkow, M. R. (1995). Cell density and culture factors regulatekeratinocyte commitment to differentiation and expression of suprabasal K1/K10keratins. J. Invest. Dermatol. 104, 271-276.
Qian, D. Z., Kachhap, S. K., Collis, S. J., Verheul, H. M. W., Carducci, M. A.,Atadja, P. and Pili, R. (2006). Class II histone deacetylases are associated withVHL-independent regulation of hypoxia-inducible factor 1 alpha. Cancer Res. 66,8814-8821.
Repertinger, S. K., Campagnaro, E., Fuhrman, J., El-Abaseri, T., Yuspa, S. H. and
Hansen, L. A. (2004). EGFR enhances early healing after cutaneous incisionalwounding. J. Invest. Dermatol. 123, 982-989.
Rheinwald, J. G. and Beckett, M. A. (1981). Tumorigenic keratinocyte lines requiringanchorage and fibroblast support cultures from human squamous cell carcinomas.Cancer Res. 41, 1657-1663.
ARNT in epidermal differentiation 3331

Journ
alof
Cell
Scie
nce
Roberts, P. J. and Der, C. J. (2007). Targeting the Raf-MEK-ERK mitogen-activatedprotein kinase cascade for the treatment of cancer. Oncogene 26, 3291-3310.
Sandilands, A., Sutherland, C., Irvine, A. D. and McLean, W. H. (2009). Filaggrin inthe frontline: role in skin barrier function and disease. J. Cell Sci. 122, 1285-1294.
Schlessinger, J. (2000). Cell signaling by receptor tyrosine kinases. Cell 103, 211-225.Schneider, M. R., Werner, S., Paus, R. and Wolf, E. (2008). Beyond wavy hairs: the
epidermal growth factor receptor and its ligands in skin biology and pathology. Am. J.
Pathol. 173, 14-24.Schnekenburger, M., Peng, L. and Puga, A. (2007). HDAC1 bound to the Cyp1a1
promoter blocks histone acetylation associated with Ah receptor-mediated trans-activation. Biochim. Biophys. Acta 1769, 569-578.
Sen, G. L., Webster, D. E., Barragan, D. I., Chang, H. Y. and Khavari, P. A. (2008).Control of differentiation in a self-renewing mammalian tissue by the histonedemethylase JMJD3. Genes Dev. 22, 1865-1870.
Sibilia, M. and Wagner, E. F. (1995). Strain-dependent epithelial defects in micelacking the EGF receptor. Science 269, 234-238.
Sogawa, K., Nakano, R., Kobayashi, A., Kikuchi, Y., Ohe, N., Matsushita, N. and
Fujii-Kuriyama, Y. (1995). Possible function of Ah receptor nuclear translocator(Arnt) homodimer in transcriptional regulation. Proc. Natl. Acad. Sci. USA 92, 1936-1940.
Stiehl, D. P., Bordoli, M. R., Abreu-Rodriguez, I., Wollenick, K., Schraml, P.,
Gradin, K., Poellinger, L., Kristiansen, G. and Wenger, R. H. (2011). Non-canonical HIF-2alpha function drives autonomous breast cancer cell growth via anAREG-EGFR/ErbB4 autocrine loop. Oncogene. 31, 2283-2297.
Stoll, S. W., Johnson, J. L., Bhasin, A., Johnston, A., Gudjonsson, J. E., Rittie, L.and Elder, J. T. (2010a). Metalloproteinase-mediated, context-dependent function ofamphiregulin and HB-EGF in human keratinocytes and skin. J. Invest. Dermatol. 130,295-304.
Stoll, S. W., Johnson, J. L., Li, Y., Rittie, L. and Elder, J. T. (2010b). Amphiregulincarboxy-terminal domain is required for autocrine keratinocyte growth. J. Invest.
Dermatol. 130, 2031-2040.Swanson, H. I. and Yang, J. H. (1999). Specificity of DNA binding of the c-Myc/Max
and ARNT/ARNT dimers at the CACGTG recognition site. Nucleic Acids Res. 27,3205-3212.
Takagi, S., Tojo, H., Tomita, S., Sano, S., Itami, S., Hara, M., Inoue, S., Horie, K.,Kondoh, G., Hosokawa, K. et al. (2003). Alteration of the 4-sphingenine scaffolds of
ceramides in keratinocyte-specific Arnt-deficient mice affects skin barrier function.
J. Clin. Invest. 112, 1372-1382.
Toki, F., Nanba, D., Matsuura, N. and Higashiyama, S. (2005). Ectodomain shedding
of membrane-anchored heparin-binding EGF like growth factor and subcellular
localization of the C-terminal fragment in the cell cycle. J. Cell. Physiol. 202, 839-
848.
Uribe, P. and Gonzalez, S. (2011). Epidermal growth factor receptor (EGFR) and
squamous cell carcinoma of the skin: molecular bases for EGFR-targeted therapy.
Pathol. Res. Pract. 207, 337-342.
Wang, X., Bolotin, D., Chu, D. H., Polak, L., Williams, T. and Fuchs, E. (2006). AP-
2alpha: a regulator of EGF receptor signaling and proliferation in skin epidermis.
J. Cell Biol. 172, 409-421.
Waterman, H. and Yarden, Y. (2001). Molecular mechanisms underlying endocytosis
and sorting of ErbB receptor tyrosine kinases. FEBS Lett. 490, 142-152.
Weir, L., Robertson, D., Leigh, I. M., Vass, J. K. and Panteleyev, A. A. (2011a).
Hypoxia-mediated control of HIF/ARNT machinery in epidermal keratinocytes.
Biochim. Biophys. Acta 1813, 60-72.
Weir, L., Robertson, D., Leigh, I. M. and Panteleyev, A. A. (2011b). The reduction of
water-soluble tetrazolium salt reagent on the plasma membrane of epidermal
keratinocytes is oxygen dependent. Anal. Biochem. 414, 31-37.
Wenger, R. H. and Gassmann, M. (1997). Oxygen(es) and the hypoxia-inducible
factor-1. Biol. Chem. 378, 609-616.
Wondimu, A., Weir, L., Robertson, D., Mezentsev, A., Kalachikov, S. and
Panteleyev, A. A. (2012). Loss of Arnt (Hif1b) in mouse epidermis triggers dermal
angiogenesis, blood vessel dilation and clotting defects. Lab. Invest. 92, 110-124.
Woodworth, C. D., Gaiotti, D., Michael, E., Hansen, L. and Nees, M. (2000).
Targeted disruption of the epidermal growth factor receptor inhibits development of
papillomas and carcinomas from human papillomavirus-immortalized keratinocytes.
Cancer Res. 60, 4397-4402.
Yarden, Y. and Schlessinger, J. (1987). Self-phosphorylation of epidermal growth
factor receptor: evidence for a model of intermolecular allosteric activation.
Biochemistry 26, 1434-1442.
Yarden, Y. and Sliwkowski, M. X. (2001). Untangling the ErbB signalling network.
Nat. Rev. Mol. Cell Biol. 2, 127-137.
Journal of Cell Science 125 (14)3332