Embed Size (px)
Citation preview

Analysis of pFQ31, a 8551-bp cryptic plasmid from the symbioticnitrogen-¢xing actinomycete Frankia
Celine Lavire a;*, Dominique Louis b, Guy Perrie©re c, Jerome Briolay b,Philippe Normand a, Benoit Cournoyer a
a UMR CNRS 5557 Microbial Ecology, Universite Claude Bernard, Lyon 1, 69 622 Villeurbanne, Franceb DTAMB, Universite Claude Bernard, Lyon 1, 69 622 Villeurbanne, France
c UMR CNRS 5558 Biometrie et Biologie Evolutive, Universite Claude Bernard, Lyon 1, 69 622 Villeurbanne, France
Received 5 December 2000; received in revised form 6 February 2001; accepted 14 February 2001
Abstract
The actinomycete Frankia has never been transformed genetically. To favour the development of Frankia cloning vectors, we have fullysequenced the Frankia alni pFQ31 cryptic plasmid and performed analyses to characterise its coding and non-coding regions. This plasmid is8551 bp-long and contains 72% G+C. Computer-assisted analyses identified 18 open reading frames (ORFs). These ORFs show asynonymous codon usage different from the one of Frankia chromosomal genes, suggesting an evolutionary bias linked to the nature of thereplicon or a horizontal transfer. Three ORFs were found to encode genes likely to be involved in plasmid replication and stability: parFA(partition protein), ptrFA (transcriptional repressor of the GntR family) and repFA (initiation of replication). DNA signatures of areplication origin were identified in the ptrFA^repFA intergenic region. These structural motifs are similar to those observed among originsof iteron-containing plasmids replicating via a a mode. ß 2001 Federation of European Microbiological Societies. Published by ElsevierScience B.V. All rights reserved.
Keywords: Plasmid replication; Horizontal transfer; Frankia ; Actinomycete
1. Introduction
Bacterial genetic transformation can be obtained usingseveral strategies. One of them is through the developmentof vectors derived from native plasmids. In Frankia, plas-mids have been detected and divided into four groups: onefor the 8-kb plasmids, one for the 18-kb plasmids, one forthe 35-kb plasmids and one for the 55-kb plasmids [1].Frankia has not been genetically transformed so far, butsome attempts to build a cloning vector from a plasmid ofthe 8-kb group (pFQ31) have been made. Normand et al.[2] cloned an Escherichia coli^Streptomyces shuttle vectorinto pFQ31, leaving pFQ31 as the sole possible actinomy-cetal replicon. However, transformation of Streptomyceslividans using this construct could not be obtained. Someyears later, Cournoyer and Normand [3] also tried to
develop Frankia vectors including constructs based on theuse of the integrative properties of some actinomycetalvectors, and constructs that could make use of Frankiagenetic recombination. Nevertheless, transformation wasnot observed even if all these plasmids were shown toenter cells by electroporation [3,4]. Many reasons can ex-plain these failures to transform Frankia, such as problemswith heterologous genetic transcription, plasmid stabilityand protection systems against foreign DNA. Moreover,pFQ31 genetic manipulations were performed withoutknowledge of the location of its replication unit. Theabove reasons could have resulted in the deletion or dis-ruption of key positions. Here, we present the ¢rst reporton the characterisation of an indigenous Frankia plasmid(pFQ31). These data will favour the development of futureFrankia cloning vectors. This is also the ¢rst report show-ing that a Frankia plasmid can have codon biases di¡erentfrom the chromosomal genes, suggesting that plasmidsmight have been acquired by horizontal transfer. Sequenc-ing Frankia plasmids (and in the long run its genome) willimprove our understanding on Frankia and its interactionswith plants.
0378-1097 / 01 / $20.00 ß 2001 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.PII: S 0 3 7 8 - 1 0 9 7 ( 0 1 ) 0 0 0 9 5 - 7
* Corresponding author. Tel. : +33 (4) 72 43 13 79;Fax: +33 (4) 72 43 12 23; E-mail : [email protected]
FEMSLE 9857 26-3-01
FEMS Microbiology Letters 197 (2001) 111^116
www.fems-microbiology.org

2. Materials and methods
2.1. Strains and culture media
Frankia alni ArI3 [5] was grown in 0FTW medium [6].E. coli DH5K (Gibco BRL/Life Technologies, Cergy Pon-toise, France) was grown at 37³C in Luria^Bertani broth[7].
2.2. DNA manipulations and sequencing
All DNA manipulations were performed following stan-dard procedures [7]. Digestion of plasmid DNA with re-striction enzymes was carried out under the conditionsrecommended by the suppliers (Gibco BRL/Life Technol-ogies, Cergy Pontoise, France). The F. alni ArI3 plasmidextraction procedure was according to Simonet et al. [6].The complete nucleotide sequence of pFQ31 plasmid wasobtained after restriction enzyme digestions, subcloningsinto pBluescript (SK-, Stratagene Europe, The Nether-lands), and sequencing using the Dynamic and Dye termi-nator sequencing kit (Amersham Pharmacia Biotech, Or-say, France) on the automated £uorescence sequencer373A Perkin Elmer (Norwalk, CT, USA).
2.3. DNA sequence analysis
DNA sequence analyses were performed using the test-code, best-¢t, base composition, terminator and promoterprograms of the GCG (University of Wisconsin) and Sta-den (Cambridge) packages available on the INFOBIOGEN
server (http://www.infobiogen.fr/). Open reading frames(ORFs) were identi¢ed using FramePlot 2.3 (http://www.nih.go.jp/Vjun/cgi-bin/frameplot.pl). DNA and ami-no acid (aa) homology searches were done online usingBLASTN and BLASTX (http://www.ncbi.nlm.nih.gov/blast/).The between/within multivariate method available fromthe ADE-4 package (http://pbil.univ-lyon1.fr/ADE-4/Mac-Mul4.html) was used.
3. Results and discussion
3.1. General characteristics of pFQ31
pFQ31 (EMBL database accession number AJ297945)was found to have a length of 8551 bp and to have a totalG+C content of 72%. This is consistent with the totalG+C content of Frankia total DNA, estimated to bearound 66^75% [8]. The restriction endonuclease map ofpFQ31 plasmid, deduced from its sequence (Fig. 1), showssome di¡erences with the one presented by Normand et al.[2]. Two KpnI sites are observed here instead of one, andfour SphI sites instead of three. We have found one sitefor EcoRV and XhoI that were not detected experimen-tally by Normand et al. [2].
The testcode program was used to position coding andnon-coding regions on both strands of pFQ31 (Fig. 2).ORFs (longer than 70 bp) among the coding regionswere then identi¢ed using FramePlot 2.3 and all possiblestart codons. The 17 ORFs detected using this approachare shown in Fig. 2.
3.2. ORF analysis
For each ORF, the G+C content in the ¢rst, second andthird position of each codon was computed (respectively,GC1, GC2 and GC3). All representative Frankia chromo-somal genes available in the databases (see Fig. 3 for theiraccession numbers) were also analysed for their G+C con-tent at the three positions of each codon, to determine themost frequent situation encountered among Frankia andto identify pFQ31 ORFs that are likely to be coding se-quences. A unique G+C pro¢le is observed among theFrankia chromosomal genes characterised so far, GC3(91%)sGC1 (68%)vGC2 (49%), but pFQ31 G+C con-tent divides the ORFs into two groups; (a) ORFs withGC3sGC1sGC2: ORFs ArF, BF, EF, HF, PrF andQrF ; (b) ORFs with GC36GC1 or GC2: ORFs CrF,DrF, FrF, GF, JF, KrF, LrF, MF, NrF, OF and SrF. TheMann^Whitney non-parametric test shows that the di¡er-ences between these two groups are signi¢cant for GC1and GC3 (P = 0.024 and 0.001, respectively). The ¢rstgroup (a) of pFQ31 ORFs shows a pro¢le similar to theone observed for Frankia chromosomal genes. The average%G+C at each codon position is, respectively, 77, 56 and88%. These values, as the one observed for the chromoso-mal genes, are similar to the %G+C content predicted byMuto and Osawa for a genome of approximately 70%
Fig. 1. Physical map of pFQ31. Some restriction sites are shown. Bam-HI, HindIII, PstI, PvuII and XbaI do not cut pFQ31. Numbers indicatepositions in bp. See Fig. 2 for position of other ORFs. Arrows indicatethe location and orientation of the putative parFA (partitioning), ptrFA(transcriptional repressor), and repFA (replication initiation) genes. Theasterisk indicates BclI, the cloning site use to build pFQ199.
FEMSLE 9857 26-3-01
C. Lavire et al. / FEMS Microbiology Letters 197 (2001) 111^116112

G+C [9]. A sequence similar to a ribosome-binding site isobserved upstream of the start codon of each ORF of the¢rst group. A canonical 335/310 E. coli-type promoter isfound upstream of the start codon of ORFs ArF and QrF.In the second group (b) of ORFs, GC3 is lower than theGC1 and/or GC2. This is a situation di¡erent from theone observed for the Frankia chromosomal genes. No se-quence similar to a ribosome-binding site is observed up-stream of the start codon of any of these ORFs. Putativetranscriptional terminators are observed at the 5P-end ofORFs NrF and SrF, and two 335/310 E. coli-type pro-moters are observed upstream of ORFs CrF and OF pu-tative start codons.
3.3. Codon usage analysis
To test the di¡erences between the ORF predicted onthe plasmid and Frankia chromosomal sequences (acces-sion numbers are given in Fig. 3), we have ¢rst veri¢edthat the chromosomal genes available were likely to berepresentative of the chromosome and then performed abetween/within analysis. Our chromosomal genes data setis dominated by nif gene representatives. The representa-tivity of these genes for the study of the codon usage of amicroorganism was tested by using all gene sequencesavailable for Anabaena vulgaris (143 genes with 21 nif se-quences). A between/within analysis showed that there wasno signi¢cant di¡erence between the codon usage of nifgenes and the other chromosomal genes of this cyanobac-
teria. This suggested that we could use our data set tocompare codon usage between plasmid and chromosomalORFs. The between/within analysis was thus performed.This analysis was not performed on the two classes ofplasmid ORFs (described in Section 3.2) separately be-cause of the limited number of individuals for each class(respectively, 6 and 11). As the between/within analysistries to ¢nd the best linear combination of variables(here the codons) allowing separation of the di¡erentclasses, the grouping of the two categories of plasmidORFs has no incidence on the quality of the discrimina-tion. The ¢rst step of our between/within analysis con-sisted in the computation of a correspondence analysison a table in which the di¡erent groups to be discrimi-nated are ordered. A table containing the absolute fre-quencies for the 61 codons corresponding to amino acidswas used. The two groups were the 17 plasmid ORFs (#1)and the 26 chromosomal genes (#2). After that, the be-tween/within analysis itself was computed, using the sta-tistical triplet produced by the correspondence analysis.To estimate the validity of the discrimination betweenthe two groups, a jack-knife resampling test was per-formed. The distribution of the factor score for the twogroups on the only axis of the between/within analysis isgiven in Fig. 3a. The accuracy of the discrimination com-puted using the jack-knife test was found to be 90.7%,suggesting that the plasmid ORFs have a di¡erent synon-ymous codon usage than that of the chromosomal genes.The factor scores for the codons allowed us to identify the
Fig. 2. Analysis of pFQ31 coding regions and ORFs. The scaled line represents pFQ31 DNA sequence (8551 bp). Above this line, ORFs (identi¢ed bythe FramePlot program using ATG, GTG, CTG and TTG start codons) are positioned according to their coding strand (+ or 3) and frame (1, 2, or3). Each ORF was given a name. Large arrows indicate the most likely position of the start codon for each ORF and the small arrows indicate the oth-er potential start codons. Putative promoters (circles), ribosome-binding sites (triangles) and terminators (#) are shown. ORF ArF RBS is at 39 bpfrom its start codon, ORF BF RBS is at 35 bp from its start codon, ORF EF RBS is at 312 bp, ORF HF RBS is at 37 bp, ORF PrF RBS is at 323bp, and ORF QrF RBS is at 319 bp. Terminator coordinates are as follows: #1: 4847^4837, #2: 5435^5408. Below the line corresponding to pFQ31,coding (plain line) and non-coding (rectangles) regions (detected by the testcode program) are shown for both strands (+ and 3). Scale is given in bp.
FEMSLE 9857 26-3-01
C. Lavire et al. / FEMS Microbiology Letters 197 (2001) 111^116 113
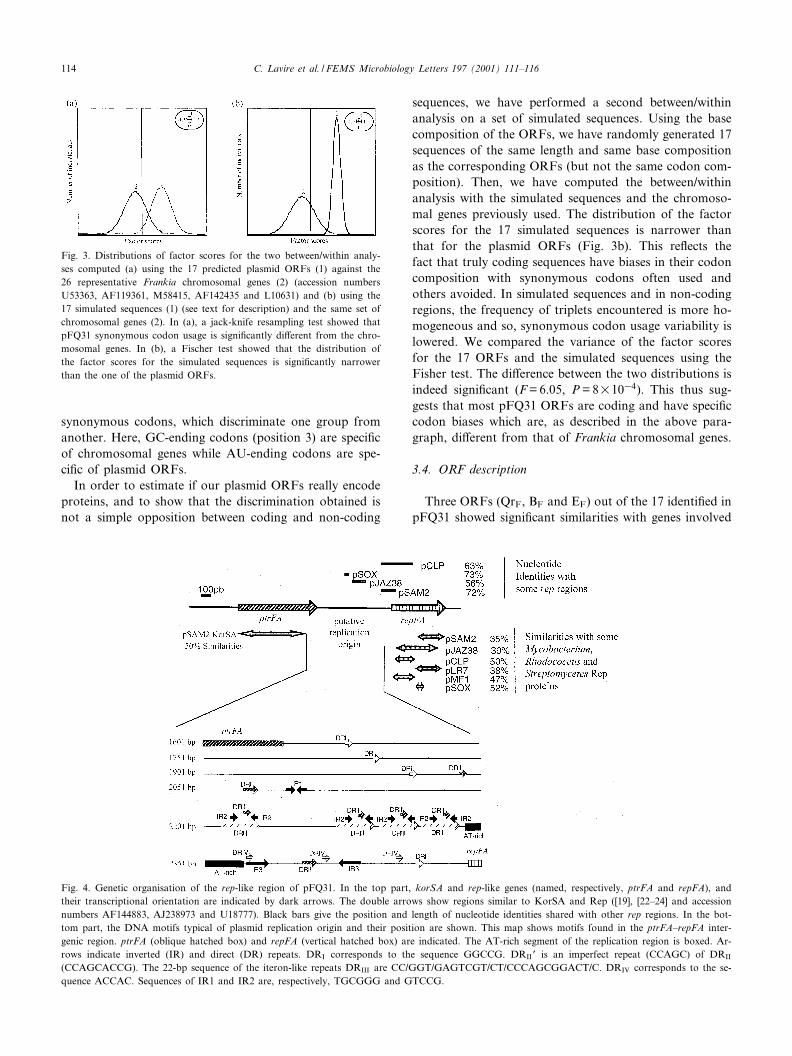
synonymous codons, which discriminate one group fromanother. Here, GC-ending codons (position 3) are speci¢cof chromosomal genes while AU-ending codons are spe-ci¢c of plasmid ORFs.
In order to estimate if our plasmid ORFs really encodeproteins, and to show that the discrimination obtained isnot a simple opposition between coding and non-coding
sequences, we have performed a second between/withinanalysis on a set of simulated sequences. Using the basecomposition of the ORFs, we have randomly generated 17sequences of the same length and same base compositionas the corresponding ORFs (but not the same codon com-position). Then, we have computed the between/withinanalysis with the simulated sequences and the chromoso-mal genes previously used. The distribution of the factorscores for the 17 simulated sequences is narrower thanthat for the plasmid ORFs (Fig. 3b). This re£ects thefact that truly coding sequences have biases in their codoncomposition with synonymous codons often used andothers avoided. In simulated sequences and in non-codingregions, the frequency of triplets encountered is more ho-mogeneous and so, synonymous codon usage variability islowered. We compared the variance of the factor scoresfor the 17 ORFs and the simulated sequences using theFisher test. The di¡erence between the two distributions isindeed signi¢cant (F = 6.05, P = 8U1034). This thus sug-gests that most pFQ31 ORFs are coding and have speci¢ccodon biases which are, as described in the above para-graph, di¡erent from that of Frankia chromosomal genes.
3.4. ORF description
Three ORFs (QrF, BF and EF) out of the 17 identi¢ed inpFQ31 showed signi¢cant similarities with genes involved
Fig. 3. Distributions of factor scores for the two between/within analy-ses computed (a) using the 17 predicted plasmid ORFs (1) against the26 representative Frankia chromosomal genes (2) (accession numbersU53363, AF119361, M58415, AF142435 and L10631) and (b) using the17 simulated sequences (1) (see text for description) and the same set ofchromosomal genes (2). In (a), a jack-knife resampling test showed thatpFQ31 synonymous codon usage is signi¢cantly di¡erent from the chro-mosomal genes. In (b), a Fischer test showed that the distribution ofthe factor scores for the simulated sequences is signi¢cantly narrowerthan the one of the plasmid ORFs.
Fig. 4. Genetic organisation of the rep-like region of pFQ31. In the top part, korSA and rep-like genes (named, respectively, ptrFA and repFA), andtheir transcriptional orientation are indicated by dark arrows. The double arrows show regions similar to KorSA and Rep ([19], [22^24] and accessionnumbers AF144883, AJ238973 and U18777). Black bars give the position and length of nucleotide identities shared with other rep regions. In the bot-tom part, the DNA motifs typical of plasmid replication origin and their position are shown. This map shows motifs found in the ptrFA^repFA inter-genic region. ptrFA (oblique hatched box) and repFA (vertical hatched box) are indicated. The AT-rich segment of the replication region is boxed. Ar-rows indicate inverted (IR) and direct (DR) repeats. DRI corresponds to the sequence GGCCG. DRIIP is an imperfect repeat (CCAGC) of DRII
(CCAGCACCG). The 22-bp sequence of the iteron-like repeats DRIII are CC/GGT/GAGTCGT/CT/CCCAGCGGACT/C. DRIV corresponds to the se-quence ACCAC. Sequences of IR1 and IR2 are, respectively, TGCGGG and GTCCG.
FEMSLE 9857 26-3-01
C. Lavire et al. / FEMS Microbiology Letters 197 (2001) 111^116114

in plasmid replication and stability. Signi¢cant similaritieswith the other 14 ORFs were not detected. The deducedaa sequence of ORF QrF (243 aa) shows similarities withpartitioning proteins: 47% similarity (29% identity) in a211-aa overlap to ParA of pRA2 of Pseudomonas alcali-genes (212 aa) [10] and 50% similarity (32% identity) in a188-aa overlap to ParA of pCLP of Mycobacterium cela-tum (214 aa) [11]. These proteins are thought to physicallyseparate DNA molecules and provide equal distribution ofgenetic material upon cell division. Such a partitioningmechanism allows stable inheritance of plasmids. ORFQrF was named parFA.
ORF BF encodes a putative protein of 294 aa. Thededuced aa sequence shows signi¢cant similarities withtranscriptional repressors of the GntR family involved ina kil^kor system. In such systems, the kil gene (kill phe-notype) speci¢es functions lethal to either the host or theplasmid, when unregulated by the kor (for kill override)gene. ORF BF shows 50% similarity (33% identity) in a254-aa overlap to KorSA of Streptomyces ambofaciensplasmid pSAM2 [12]. In Streptomyces plasmids, such aspIJ101 or pSAM2, the kil^kor system is associated withtransfer, the kill function being attributed to tra genes(transfer gene). pSAM2 KorSA indirectly controlspSAM2 replication, integration and excision via the regu-lation of the pra gene, which is di¡erent from the Korprotein of other Streptomyces plasmids that directly con-trol plasmid transfer [12^13]. No sequence similar to praor tra genes has been observed in pFQ31 nor sequencesresembling any of the kil genes described in the databases.Nevertheless, disruption of this ORF may lead to the ex-pression of a gene having a kill phenotype not described sofar. This would prevent successful transformation fromhappening, killing cells harbouring this plasmid. ORFBF was named ptrFA.
The deduced aa sequence of the ORF EF (202 aa) showssigni¢cant similarities with Rep proteins (Fig. 4) requiredfor initiation of replication. They are found in plasmidsreplicating via a or rolling circle (RC) mechanisms. ORFEF shows 50% similarity (41% identity) in a 71-aa overlapwith pCLP Rep of M. celatum [19], 39% similarity (34%identity) in a 153-aa overlap with pJAZ38 Rep of Myco-bacterium fortuitum [14], 52% similarity (41% identity) in a31-aa overlap with pSOX Rep of Rhodococcus sp. [15] and35% similarity (28.5% identity) in a 105-aa overlap withpSAM2 Rep of S. ambofaciens [16]. All of these proteinsseem to be longer than ORF EF ; pFQ31 ORF EF is 202aa-long whereas Mycobacterium and Rhodococcus Repwere suggested to be around 350 aa. However, forpSOX, only a 21.4-kDa Rep protein was observed exper-imentally, suggesting a length more in agreement with thedetected Frankia Rep [15]. We have thus performed thetestcode analysis on the pSOX rep region and con¢rmedthat the pSOX Rep protein is likely to be shorter (186 aa)than predicted earlier (350 aa). The pJAZ38 rep regionwas also re-analysed with the testcode program and,
again, we have found that its Rep protein is likely to beshorter than suggested earlier, matching the expected sizeof the Frankia Rep. On the basis of these similarities,pFQ31 ORF EF was named RepFA. The mechanisms ofreplication of Rhodococcus and Mycobacterium plasmidsshowing similarities to RepFA are not known but thatof pSAM2 is. pSAM2 replicates via a RC mechanismand the similarities observed between pSAM2 RepSAand RepFA could suggest a RC replication (RCR) forpFQ31. However, pSAM2 RepSA (459 aa) is twice aslong as RepFA, and the three motifs observed in RepSAas in other Rep proteins of RCR elements were not de-tected among RepFA. In fact, other properties describedbelow rather suggest that pFQ31 replicates via a a mode.
3.5. Origin of replication
The intergenic region between ptrFA and repFA showssigni¢cant similarities with the replication region of someplasmids (Fig. 4). It contains several structural motifs usu-ally found in the origin of replication of iteron-containingplasmids, replicating via a a mode of replication (Fig. 4).In such plasmids, repeated sequences of around 20 bp(called iterons) are located close to an AT-rich region.These iterons provide multiple binding sites for the Repprotein. Additional direct and/or indirect repeats thatcould serve as sites for regulation, and one or more sitesfor DnaA binding are also found in these origins (for re-view see [17]). The pFQ31 sequence from position 2342 to2374 has a total A+T content of 66.6%, which is muchhigher than the A+T content of the pFQ31 total sequence(38%). Four 22-bp direct iteron-like repeats (DRIII) areadjacent to this AT-rich region. Each iteron-like repeatcontains a 5-bp inverted repeat (IR2) and a 5-bp imperfectdirect repeat (DRIIP) of the 9-bp DRII. These DNA se-quence features are typical of Rep-binding sites. Someother direct and inverted repeats are also found in theptrFA^repFA intergenic region (Fig. 4) : one 6-bp invertedrepeat (IR1), ¢ve 5-bp direct repeats (DRI), three 5-bpdirect repeats (DRIV) and one 13-bp inverted repeat(IR3). These additional repeats could serve as binding sitesfor regulators or for the chromosomally encoded DnaAprotein. Boxes matching partially the consensus DnaA-binding site (TTG/ATCCACA [18]) are found insideDRIV and DRII.
3.6. A novel pFQ31 derivative, pFQ199
The present study identi¢ed many coding and non-cod-ing regions on pFQ31. These data will direct strategies forthe construction of pFQ31-based cloning vectors. We nowknow that EcoRV, EcoRI and XhoI are single cutters thatshould be avoided because they are likely to disrupt essen-tial parts of pFQ31. Normand et al. [2] used the SstIrestriction site of pFQ31 to build their cloning vector.Here, we show that this leads to the deletion of a 90-bp
FEMSLE 9857 26-3-01
C. Lavire et al. / FEMS Microbiology Letters 197 (2001) 111^116 115

region at the beginning of ptrFA. This deletion could ex-plain the absence of transformation with this pFQ31 de-rivative. Restriction enzymes that do not cut essential re-gions, such as BclI, BstBI or MstII, seem to representbetter choices for the insertion of genetic markers andfor the construction of pFQ31 cloning vectors.
We have thus attempted to build an actinomycetalpFQ31 vector using the BclI restriction site. A transfor-mation experiment using Frankia cells can take around6^12 months from the time that one gets putative trans-formants to the ¢nal tests about the maintenance of thecloning vector. S. lividans was thus used as a model actino-mycete to test the e¤cacy of our construct. pFQ31 wascut at its BclI site and was then ligated into pFQ188(a pKK232-8 derivative) which contains a CAT (chlor-amphenicol acetyltransferase) gene under the control ofFrankia rrnP2 promoter and RBS [19]. This construct,named pFQ199, could be cloned into E. coli by selectingfor Cm-r (chloramphenicol resistance). It was then ex-tracted and used to transform S. lividans TK24 protoplastsfollowing [20]. A pIJ487 [21] derivative with a CAT geneunder the control of rrnP2 (see [19]) was used to verify thetransformability of the S. lividans protoplasts. Neverthe-less, S. lividans could not be transformed using pFQ199.Several substitutions between Frankia and StreptomycesDnaA-like boxes could explain this failure to transformS. lividans. Frankia DnaA boxes might not be recognisedby the Streptomyces DnaA. We are now in the progress ofcloning Frankia dnaA to improve pFQ199. IncludingFrankia dnaA into a pFQ31 derivative could overcomeproblems related to DnaA-binding speci¢cities.
Acknowledgements
We thank F. Gourbie©re (UMR CNRS 5557) for usefuladvice on statistical analyses. We also thank the IFR41(Sciences et Methodes de l'Ecologie et de l'Evolution) forsupporting the collaboration between the two researchunits and the technical platform DTAMB involved inthis project.
References
[1] Simonet, P., Normand, P., Moiroud, A. and Lalonde, M. (1985)Restriction enzyme digestion patterns of Frankia plasmids. PlantSoil 87, 49^60.
[2] Normand, P., Downie, J.A., Johnston, A.W., Kieser, T. and Lalonde,M. (1985) Cloning of a multicopy plasmid from the actinorhizalnitrogen-¢xing bacterium Frankia sp. and determination of its restric-tion map. Gene 34, 367^370.
[3] Cournoyer, B. and Normand, P. (1992) Electropermeabilisation ofFrankia intact cells to plasmid DNA. Acta Oecol. 13, 369^378.
[4] Alegre, M.T., Cournoyer, B., Mesas, J.M., Guerineau, M., Normand,P. and Pernodet, J.L. (1994) Cloning of Frankia species putativetRNA(Pro) genes and their e¤cacy for pSAM2 site-speci¢c integra-
tion in Streptomyces lividans. Appl. Environ. Microbiol. 60, 4279^4283.
[5] Berry, A. and Torrey, J.G. (1979) Isolation and characterization invivo of an actinomycetous endophyte from Alnus rubra Bong. In:Symbiotic Nitrogen Fixation in the Management of Temperate For-ests (Gordon, J.C., Wheeler, C.T. and Perry, D.A., Eds.), pp. 69^83.Oregon State University, Corvallis, OR.
[6] Simonet, P., Cappelano, A., Navaro, E., Bardin, R. and Moiroud, A.(1984) An improved method for lysis of Frankia with achromopepti-dase allows detection of new plasmids. Can. J. Microbiol. 30, 1292^1295.
[7] Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular Clon-ing: a Laboratory Manual, 2nd edn. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY.
[8] Fernandez, M.P., Meugnier, H., Grimont, P.A.D. and Bardin, R.(1989) Deoxyribonucleic acid relatedness among members of the ge-nus Frankia. Int. J. Syst. Bacteriol. 39, 424^429.
[9] Muto, A. and Osawa, S. (1987) The guanine and cytosine content ofgenomic DNA and bacterial evolution. Proc. Natl. Acad. Sci. USA84, 166^169.
[10] Kwong, S.M., Yeo, C.C., Chuah, D. and Poh, C.L. (1998) Sequenceanalysis of plasmid pRA2 from Pseudomonas alcaligenes NCIB 9867(P25X) reveals a novel replication region. FEMS Microbiol. Lett.158, 159^165.
[11] Picardeau, M., Le Dantec, C. and Vincent, V. (2000) Analysis of theinternal replication region of a mycobacterial linear plasmid. Micro-biology 146, 305^313.
[12] Hage©ge, J., Pernodet, J.L., Sezonov, G., Gerbaud, C., Friedmann, A.and Guerineau, M. (1993) Transfer functions of the conjugative in-tegrating element pSAM2 from Streptomyces ambofaciens : character-ization of a kil^kor system associated with transfer. J. Bacteriol. 175,5529^5538.
[13] Sezonov, G., Possoz, C., Friedmann, A., Pernodet, J.L. and Gueri-neau, M. (2000) KorSA from the Streptomyces integrative elementpSAM2 is a central transcriptional repressor: target genes and bind-ing sites. J. Bacteriol. 182, 1243^1250.
[14] Gavigan, J.A., Ainsa, J.A., Perez, E., Otal, I. and Martin, C. (1997)Isolation by genetic labeling of a new mycobacterial plasmid,pJAZ38, from Mycobacterium fortuitum. J. Bacteriol. 179, 4115^4122.
[15] Denis-Larose, C., Bergeron, H., Labbe, D., Greer, C.W., Hawari, J.,Grossman, M.J., Sankey, B.M. and Lau, P.C. (1998) Characteriza-tion of the basic replicon of Rhodococcus plasmid pSOX and develop-ment of a Rhodococcus^Escherichia coli shuttle vector. Appl. Environ.Microbiol. 64, 4363^4367.
[16] Hagege, J., Boccard, F., Smokvina, T., Pernodet, J.L., Friedmann, A.and Guerineau, M. (1994) Identi¢cation of a gene encoding the rep-lication initiator protein of the Streptomyces integrating element,pSAM2. Plasmid 31, 166^183.
[17] Espinosa, M., Cohen, S., Couturier, M., Del Solar, G., Diaz-Orejas,R., Giraldo, R., Janniere, L., Miller, C., Osborn, M. and Thomas,C.M. (2000) Plasmid replication and copy number control. In: TheHorizontal Gene Pool, Bacterial Plasmids and Gene Spread (Thomas,C.M., Ed.), pp. 1^47. Harwood Academic Publishers, Amsterdam.
[18] Zakrezewska-Czerwinska, J. and Schrempf, H. (1992) Characteriza-tion of an autonomously replicating region from the Streptomyceslividans chromosome. J. Bacteriol. 174, 2688^2693.
[19] Cournoyer, B. and Normand, P. (1994) Gene expression in Frankia :characterization of promoters. Microbios 78, 229^236.
[20] Hopwood, D.A., Bibb, M.J., Chater, K.F., Kieser, T., Bruton, C.J.,Kieser, H.M., Lydiate, D.J., Smith, C.P., Ward, J.M. and Schrempf,H. (1985) Genetic Manipulation of Streptomyces : a Laboratory Man-ual. John Innes Foundation, Norwich.
[21] Ward, J.M., Janssen, G.R., Kieser, T., Bibb, M.J., Buttner, M.J. andBibb, M.J. (1986) Construction and characterization of a series ofmulti-copy promoter-probe vectors for Streptomyces using the amy-loglycoside phosphotransferase gene from Tn5 as indicator. Mol.Gen. Genet. 203, 468^475.
FEMSLE 9857 26-3-01
C. Lavire et al. / FEMS Microbiology Letters 197 (2001) 111^116116





























