Embed Size (px)
Citation preview

Anaerobic and Aerobic Activity in Isolated Muscle. 313
(4) Stella, * Journ. Physiol.,’ vol. 66, p. 19 (1928).(5) Eggleton, Eggleton and Hill, ‘ Roy. Soc. Proc.,’ B, vol. 103, p. 620 (1928).(6) Hill, ‘ Roy. Soc. Proc.,’ B, vol. 103, p. 117 (1928).(7) Hill, ‘ Roy. Soc. Proc.,’ B, vol. 104, p. 39 (1928).(8) Hill, * Roy. Soc. Proc.,’ B, Vol. 103, p. 183 (1928).(9) Hill, ‘ Roy. Soc. Proc.,’ B, vol. 103, p. 138 (1928).
6 1 2 . 7 4 4 . 2 i i . 0 8 5 .2
Anaerobic and Aerobic Activity in Isolated Muscle.By A. Y. H il l , F.R.S., and P. K u p a l o v .*
(Received June 27, 1929.)
(From the Department of Physiology and Biochemistry, University College, London.)
It is well known that a muscle excited anaerobically fails to contract any more when the concentration of lactic acid inside it rises to about 0 • 3 per cent. In the presence of oxygen the lactic acid disappears and the muscle recovers, but this does not prove that recovery is due solely to the removal of the acid; it might be argued that some other mechanism also needs restoration before the muscle can function normally again. Now it is possible, by suspending a sufficiently thin muscle in oxygen-free Ringer’s solution and exciting it sufficiently slowly, to allow diffusion to remove the lactic acid as it is formed. Under such conditions the muscle is found to liberate 2 to 3 times as much energy as it would if diffusion were not permitted. I t appears to continue working until all its carbohydrate is used up. Since, therefore, nothing is known to pass into the Ringer’s solution except lactic acid, we may, until some other substance is found, attribute the absence of fatigue to the absence of lactic acid in the muscle, and conclude that when diffusion is not allowed fatigue is due primarily to lactic acid.
In a recent paper (1, p. 51) the case was considered of a muscle excited by single shocks at regular intervals in oxygen-free Ringer’s solution, attaining a “ steady state ” in which outward diffusion of lactic acid balances new formation. If a plane sheet of muscle of thickness 26, exposed on both sides to Ringer’s solution, produce a gm. of lactic acid per c.c. per minute, and if
* Fellow of the Rockefeller Foundation.2 A 2
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from

314 A. V. Hill and P. Kupalov.
be the diffusion constant, the greatest concentration attained anywhere inside it, viz., in its middle plane, is (in gms. per c.c.) z.b2/2k.
Let us assume that the average single twitch, during the earlier stages of a prolonged series, yields about 5 X 10~6 gm. of lactic acid per gram,* and that n shocks are given per minute. The smallest frog’s sartorius which it is practicable to use weighs about 50 mgms. and is about 2 • 5 cm. long and 3 • 5 mm. wide. Its average thickness, therefore, is rather less than 0*6 m m .: let us take b — 0-03 cm. In the preceding paper the diffusion constant for lacticacid, in a surviving muscle at rest in oxygen-free Ringer’s solution, was found to be 1 • 65 X 10-5, which lies well between the extreme values given by Eggle- ton, Eggleton and Hill (2). Inserting these quantities the greatest concentration is found to be 1-36 n X 10~4 (in gm. per gm.) or 0-0136w per cent. Now it is probably inadvisable to allow the concentration of lactic acid anywhere inside the muscle to attain a value of more than (say) 0'08 per cent, if fatigue is to be completely avoided. Equating 0*0136n to 0*08 we find n = 6 ; thus in a muscle of the average size with which we have to deal an
interval of 10 seconds at least should be allowed between individual maximal twitches. We have made our experiments accordingly.
The sartorii of a frog were prepared separately, each with a portion of bone at the pelvic end. One was used for slow stimulation in oxygen-free Ringer’s solution, as considered above, the other for rapid stimulation (0*6 to 1 sec. between shocks) either in oxygen-free Ringer’s solution or in nitrogen. The Ringer’s solution contained no bicarbonate, its pH being adjusted by a known amount of phosphate. The muscles were mounted on carriers and were stimulated isometrically by just-maximal break induction shocks directly applied by silver electrodes. A tension record was made on a slowly moving drum by a lever of the usual type employed in this laboratory. The muscles on their carriers were inserted into narrow glass tubes, the tubes being filled either with Ringer’s solution or with nitrogen : in the former case the solution was kept oxygen-free by a slow stream of pure nitrogen bubbling through it from below : in the latter case a stream of pure moist nitrogen passed up slowly through the tube. The nitrogen was freed from oxygen by passing it over heated copper in a quartz tube.
From the tension record it is possible, with fair accuracy, to determine tlie lactic acid produced. Experiments on the “ isometric coefficient of lactic acid ” by Meyerhof, confirmed by experiments on heat-production, have shown that 1 gm.-cm. of tension-length developed in an isometric twitch
* Initial heat 0-0019 cal. per gram.
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from

corresponds to 10-8 gin. of lactic acid set free (see (3)). Thus the quantity 2T lX 10-8/M, where M is the mass of, the muscle in grams, 1 its length in centimetres, and T the tension in a twitch in grams weight, represents the lactic acid set free in the series, per gram of muscle : reckoned per 100 grams of muscle, the lactic acid produced is ST? X 10~6/M. I t has been shown (3) that the isometric coefficient does not change much in a long series : so far as any change is likely, the ratio of tension to lactic acid might be expected to diminish, so that the quantity of lactic acid calculated in this way would be rather too small. Actually the amount found by titration of the Ringer’s solution (see below) usually agrees well with the calculated quantity.
In a long series of contractions in Ringer’s solution allowance must be made for the lactic produced owing to the resting anaerobic metabolism. The following rates of lactic acid formation were assumed :—Temp. °C................................ 14 15 16 17 18 19 20Gms. of lactic acid per gram
of muscle per minute .... 2-6 X 10~6 2-85 3-12 3-42 3-75 4-1 4-5 X 10~6
These assume a Q10 of 2*5, and the evidence for the absolute value taken is given in (1), p. 51. The muscles were stimulated in every case until their response had practically disappeared.
An approximate estimate of the lactic acid which diffused out of the muscle, during the course of the stimulation, was made in some experiments as follows. The Ringer’s solution was buffered at pH 7-2 or 7*4 by sodium phosphate introduced to the extent of 20 or 40 mgms. per cent. P. The fluid in which the muscle had been working was titrated back to its original pH (as shown by an indicator) with dilute alkali, and the acid so determined was calculated as lactic acid.
The concentration of phosphate used in the Ringer’s solution (20 or 40 mgms. per cent. P) should be sufficient, according to Stella (4), to prevent the outward diffusion of phosphate even from a very fatigued muscle.
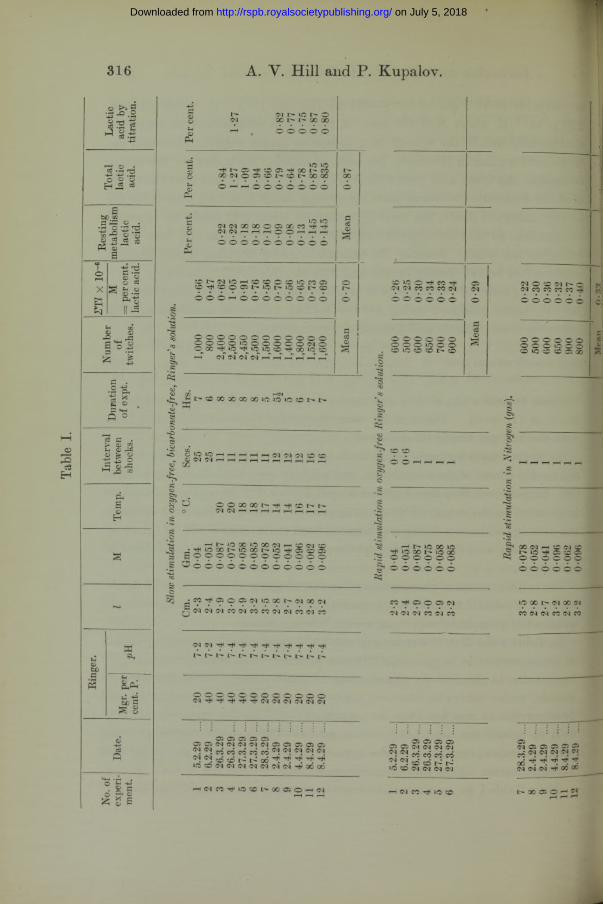
In Table I the first 12 entries represent 12 experiments on slow stimulation of single sartorii in oxygen-free Ringer’s solution. The next six entries give the results, in the first six experiments, of rapid stimulation of the opposite muscle, under otherwise identical conditions. In these the interval allowed between shocks was too short to permit any considerable quantity of lactic acid to diffuse out, and fatigue soon set in. The last six entries give the results of rapid stimulation of the opposite muscle in nitrogen gas, in the last six experiments. Here, also fatigue soon set in.
From Table I we see that, stimulated in nitrogen gas, or rapidly in Ringer’s
Anaerobic and Aerobic Activity in Isolated Muscle. 315
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from
1-4<s>
c3H
316
.3 js* oJ s l^ I J
3 1 2•+-> -+=> £ § §
bO 52I ® 2 ,i i i i
8 §a a.I I
$ ®
1 = 1
0 -e1 £
> § J3?h <D
i | - i
PhSH
bp
W
1(4J4 H be !
jg"§A
®'i -s
4 s l
A. Y. Hill and P. Kupalov.
^ I>(©OOGOOOXlf5«5»OCOI>l>
•I
| <®I .
Ifo OOOOOOh^^OhhO © WNh h h h h h h h-I
l s s i i i i i i i i i n2* r K • • • • ................................................
.§ 4 ^ 0 0 0 0 0 0 0 0 0 0 0 0 It
55Q C < I C < | ( M f O ( N f O f f O C < I C < ( C O ( M C O
l> !>• h* I> I>* l> I> !>■ I>- t'*
: i ! : : : : : : : ;
sslISSSs^sssS S S s s s ig ;? :^ :? - ?I 03 03 00 00
i—l (M « ^ U5 X I> X 05 O h ^r-H r“H i—H
Per c
ent.
1-2
7
0-8
20
-77
0-7
50
-87
0-8
0
*d i o iO§ t"- © rfi CO P tH X l> XO XWOOlCOhCOhXX 00 -g O'—i ^ O O O O O O O
Ph©
"§ 1© 19s CMCMOOOOOOOOCOt tHO WWh h h O O h h h
g
1£ O O O O O O O O O O
Ph
co.'l'* WWDHCOCOOCOiOW© X^XOC5l>lOt>lOCOl>X ©
O O O r H O O O O O O O O ©
O O O O O O O O O O O O0 0 0 0 * 0 0 0 0 0 0 0 3 0 10 00 no io lO CO ^ X lO CO»-h 03 03 03 03 r—r r—r f—r r—r i—r i—r
I
50 <-o> <-o> i
•§
I*00 „ H l> IQ X t o
^ lO X N lO Xo o o o o org o o o o o o §■< o o o o o o G$
M ^ 05 O P (N 03 03 03 CO 03 CO
o o o o® ^ (N ^ N
c| ©I JS f? *?I CO 50 1> l> 1 (M <Ml 03 CO lO CO
co o o ^ c o Tti03 03 CO CO CO 03
o03
03 O CD 03 t> o 03 CO CO CO CO -rji
O O © © © © © © © © © © ©
; |o o o o o o 0 0 0 * 0 0 0 CO *0 50 co co
© © © © © © 0 0 0 * 0 0 0 CO »o CO CO O 00
5
.§
*3
H
§■< 00 03 I—f CO 03 CD ^ lO Ol CO O O O O O O OO O © O © ©
lOXIXNXW CO 03 03 CO 03 CO
: : : : X M: : : r Mm
(M (N <N ^ XQO
i O H CM’ hhh
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from

Anaei'obic and Aerobic Activity Isolated Muscle. 317
solution, the muscle fails when it has attained an average concentration of lactic acid of about 0*3 per cent. ; stimulated, slowly, however, in Ringer’s solution so that lactic acid can escape, it can liberate more than twice as much, viz., 0*7 per cent., as the result of stimulation alone, or taking account of resting metabolism as well, nearly three times as much, viz., 0-87 per cent. It is difficult to resist the conclusion that lactic acid accumulation is the primary cause of failure in the former case. When failure finally occurs in the absence of lactic acid it seems likely that practically all the carbohydrate reserves of the muscle have been exhausted. There cannot have been much more than O’87 per cent, of available carbohydrate in the muscles of winter and spring frogs.
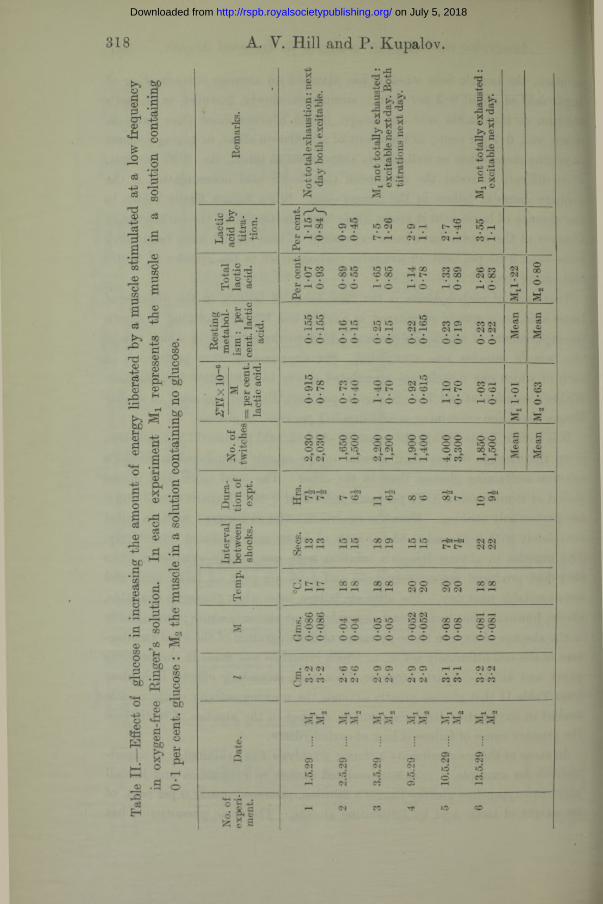
That lack of carbohydrate is the final cause of failure, in the absence of lactic acid, in a muscle stimulated in oxygen-free Ringer’s solution, is shown by experiments in which glucose was added to the solution in which one of the pair of muscles was stimulated. Six experiments were performed, in each of which one muscle was stimulated slowly to exhaustion in phosphate Ringer’s solution at pH 7*4 (20 mgms. per cent. P), and the other in similar Ringer’s solution containing in addition 0 • 1 per cent, of glucose.
It is seen in Table II that the quantity of lactic acid produced by stimulation is on the average 60 per cent, greater in the muscle stimulated in Ringer’s solution containing glucose than in the other. The total lactic acid in the period of stimulation is 1*22 per cent, instead of 0-80. Experiments 3 and 6 are particularly striking. Here the control muscles in Ringer’s solution failed completely with 0-85 and 0-83 per cent., of lactic acid produced respectively. The muscles in glucose-Ringer’s solution were not totally exhausted when they had produced respectively 1 • 65 and 1 • 26 per cent, of lactic acid, and were excitable next day. The ultimate failure, even when glucose is available, shows, however, that lack of carbohydrate is not the only cause : presumably in the absence of oxygen those processes of disintegration of the vital architecture of the cell ultimately set in, to which reference was made in the previous paper and in (5). Apart, however, from such secondary effects, it is clear from Table II that glucose greatly extends the working activity of the muscle soaked in oxygen-free Ringer’s solution, so that in the absence of lactic acid the primary cause of failure is lack of carbohydrate.
In some of these experiments a strong glycolysis was exhibited. In experiment 3, for example, a muscle stimulated almost to exhaustion and left all night in oxygen-free glucose-Ringer’s solution, produced nearly 6 per cent, of its weight of lactic acid by fermenting glucose. The same phenomenon has
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from
Tabl
e II
.—Ef
fect
of
gluc
ose
in i
ncre
asin
g th
e am
ount
of
ener
gy li
bera
ted
by a
mus
cle
stim
ulat
ed a
t a
low
fre
quen
cy
in o
xyge
n-fre
e R
inge
r’s s
olut
ion.
In
eac
h ex
perim
ent
Mi
repr
esen
ts
tbe
mus
cle
in
a so
lutio
n co
ntai
ning
0-
1 pe
r ce
nt, g
luco
se :
M2 t
be m
uscl
e in
a s
olut
ion
cont
aini
ng n
o gl
ucos
e.
318 A. V. Hill and P. Kupalov.
Rem
arks
.
Not
tota
lexh
aust
ion:
nex
t da
y bo
th e
xcita
ble.
Mx n
ot t
otal
ly e
xhau
sted
: ex
cita
ble
next
day
. Bot
h tit
ratio
ns n
ext
day.
not
tota
lly e
xhau
sted
: ex
cita
ble
next
day
.
Lac
tic
acid
by
titra
tio
n.
Per
cent
.1-
15
\0
-84
/
0-9
0-45
7-5
1-26
2-9
1-1
2-7
1-46
3-55
1-1
Tot
alla
ctic
acid
.
Per
cent
. 1-
07
0-93
0-89
0-55
1-65
0-85
1-14
0-78
1-33
0-89
1-26
0-83
cqcqpH
s"©3
Res
ting
met
abol
is
m :
per
cent
, lac
tic
acid
.
!
0-15
50-
155
0-16
0
15
0-25
0-15
0-22
0-16
5
0-23
0-19
0-23
0-22 Mea
n
Mea
n
to
©. X M
= p
er c
ent,
lact
ic a
cid.
0-91
50-
78 CO © l> rtf© © 1-
400-
70
0-92
0-61
5
1-10
0-70
1-03
0-61
10-llK
COCO©(N3
No.
of
twitc
hes
2.03
02.
030
1,65
01,
500
2,20
01,
200
1,90
01,
400
4,00
03,
300
1,85
01,
500
Mea
n
Mea
n
Dur
atio
n of
ex
pt.
s2 h!©M<m h!<mi> © H&l
H © pH
00 © ph]<m 00 t> 10 H
Inte
rval
betw
een
shoc
ks.
Secs
.13 13 pH pH
00 © rH H
lO lOrH pH
h!<mph|<m cq cq cq cq
Tem
p.
QO 00H pH
00 00pH rH
o o<M CM o oCq
00 00pH rH
§
Gm
s.0-
086
0-08
6
o o © o 0-
050-
05
0-05
20-
052
00 00 © ©© © 0-
081
0-08
1
Cm
.3-
23-
2
2-6
2-6
2-9
2-9
2-9
2-9 pH pH
CO CO 3-2
3-2
pH rH <N rH <M
3 3rH <N
S 3rH <N
S 3
Dat
e.
1.5.
29
....
2.5.
29
....
3.5.
29
....
9.5.
29
....
10.5
.29
....
13.5
.29
....
i
No.
of
expe
ri
men
t.
r—1 Cq CO lO ©
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from

Anaerobic and Aerobic Activity Isolated Muscle. 319
been observed in other cases in which muscles have been left without oxygen for long periods in Ringer's solution containing glucose. Lack of oxygen, however, is apparently not necessary to its occurrence : in experiments described below, up to 2 • 6 per cent, of lactic acid has been produced by muscles stimulated for 10 hours in oxygenated Ringer's solution containing glucose.
A few experiments were performed in which a comparison was made of cane- sugar and glucose. In contrast to the case when glucose was present no obvious increase appeared, in the energy set free by a muscle stimulated in oxygen-free Ringer’s solution, when cane-sugar was added.
Stimulation during a steady state in oxygenated Ringer's solution.—In a few experiments a muscle was stimulated in oxygenated Ringer’s solution for comparison with its companion stimulated in a similar but oxygen-free solution. Experiments were made by one of us in 1914 (7) on the total energy available in isolated muscles kept in oxygen or oxygenated Ringer’s solution : in these experiments the total tension developed in a long series of twitches was measured, and from ST/ the total heat was calculated in calories by multiplying 10~5. The best evidence now available (3) (8) gives a multiplier of 8 X 10-6. The highest value obtained in any single experiment was (with the new multiplier) 38 calories per gram, which corresponds to the oxidation of just 1 per cent, of glycogen. In oxygenated Ringer’s solution values of more than 24 calories per gram were “ practically always obtained,” which corresponds to the oxidation of 0*63 per cent, of glycogen. In two experiments, in oxygenated Ringer’s solution, we have foimd values for STZ/M of 3,110,000 and 5,550,000, corresponding to 25 and 44 calories respectively : these imply 0-65 and 1*15 per cent, of glycogen oxidised. The companion muscles in oxygen-free Ringer’s solution gave respectively 800,000 and 720,000 for ET//M, which imply respectively 0-8 and 0*72 per cent, of glycogen broken down to lactic acid.
In two more experiments in which oxygenated Ringer’s solution was employed, one muscle being in Ringer’s solution without glucose, the other in Ringer’s solution -{— 0 * 1 per cent, of glucose, the following values were obtained:—
Experiment of 14.5.29. 18° C.— With glucose,27TZ/M = 5,850,000 : heat liberated, 47 cals, per gram : carbohydrate oxidised, 1*27 per cent.
Without glucose, ETl jM = 4,350,000 : heat liberated, 35 cals, per gram : carbohydrate oxidised, 0 • 91 per cent.
By titration of the Ringer’s solution the muscle with glucose was found to have fermented 2 • 2 per cent, of its weight to lactic acid, the muscle without glucose practically none.
Experiment of 15.5.29. 18° C.— *-Withglucose, ETl/M = 5,000,000 : heat liberated, 40 cals, per gram : carbohydrate oxidised, 1 -04 per cent.
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from

Without glucose, 27T7/M = 4,120,000 : heat liberated, 33 cals, per gram : carbohydrate oxidised, 0 • 86 per cent.
By titration of the Ringer’s solution the muscle with glucose was found to have fermented 2*6 per cent, of its weight to lactic acid, the muscle without glucose practically none.
In these experiments the muscles in oxygenated Ringer’s solution gave 10,000 or more twitches : they were certainly in a “ steady state,” the oxygen supply being adequate to meet the demands of the tissue with a frequency of excitation of 17 to 23 per minute. I t was shown in (1), p. 45, that the greatest depth b to which oxygen can penetrate by diffusion in a plane sheet of tissue, during a steady state of oxygen consumption a c.c. per cubic centimetre of tissue per minute, is given by :
b = V 2
Here k is the diffusion constant of oxygen and its partial pressure outside. If the whole of the interior of a sartorius muscle, 0 • 06 cm. thick, exposed on both sides, is to be supplied with oxygen, the limiting case is when 0-03. Putting k = 1 -4 X 10~5 (Krogh) and y0 — 1 (for a fully oxygenated solution) we find, for the greatest allowable oxygen consumption, in c.c. per cubic centimetre of muscle per minute,
a — 2ky0/b2 — 0-031
or, per gram of muscle, 0 • 0295 c.c. per minute. In the average twitch discussed above, in which 5 X 10“ 6 gm. of lactic acid is liberated per gram of muscle the total heat (initial plus recovery) would be 2-1 X 385 X 5 X 10-6 = 4 X 10~3 calorie per gram, the oxygen consumption, therefore, about 8 X 10-4 c.c. per gram. Thus such a muscle should be able, in the limit, to stand a frequency of 0-0295/0-0008 = 37 shocks per minute. We have not worked at a frequency very close to the limiting one, but the fact that a muscle can stand a frequency of 23 shocks per minute for long periods, without perceptible fatigue, confirms the calculation in general.
That a thin isolated muscle, adequately supplied with oxygen by diffusion, and suspended in Ringer’s solution, can function so well for many hours, giving more than 10,000 twitches and developing a total tension per square centimetre of cross-section of over 6 tons,* is a sufficient commentary on the statement, frequently heard, that such a muscle is “ abnormal.” Abnormal it certainly becomes in the end, owing to the lack of certain factors initially
* Since Mass — density x length x cross-section = 1-05 X I X A, we have ZTZ/M = 0-95 S T I A. Thus if S T lfK = 5,850,000, ZT/A = 6,142,000 gms. wt. = 6,142 kilogms. weight = 6-14 metric tons per square centimetre.
320 A. V. Hill and P. Kupalov.
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from

Anaerobic and Aerobic A ctivity in Isolated Muscle. 321
present and usually supplied by the circulation ; a further study of the factors which would enable it to survive still longer and to develop a still greater total tension would be valuable in elucidating its mechanism. Undoubtedly, a muscle lasts longer, and develops more tension before final exhaustion, if it be given glucose : carbohydrate deficiency therefore is one limiting factor. Even with glucose, however, there is still a sharply defined lim it: the cause of this should be examined. May be, as Clark (9) found, for the case of the frog’s heart, the presence of lipoids normally supplied by blood plasma, is essential to activity, these lipoids being washed away by prolonged soaking, their absence leading to a hypodynamic condition. May be insulin, or some other hormone, is needed for continued activity. May be glucose is not fully available to the muscle, in its ordinary chemical form : this is suggested by the fact that a well-oxygenated muscle in glucose-Ringer’s solution may produce 2*6 per cent, of its weight of lactic acid during 10 hours of activity, while a companion muscle, without glucose, produces none. There is glucose in the blood, which is not normally broken down to lactic acid at such a rate. There are two known limiting factors to the performance of an isolated muscle suspended in oxygenated phosphate-Ringer’s solution, (a) the oxygen supply, and (6) the amount of carbohydrate present. Of these, (a) can be made unimportant by employing thin enough muscles in a fully oxygenated solution ; (b) can be, to some extent but not fully, avoided by adding glucose to the solution. The remaining factors, "which certainly exist, are still to be discovered.
In order to realise the performance of one of these isolated muscles it is interesting to compare the exercise taken by it with the case of man. An isolated muscle in oxygenated glucose-Ringer’s solution may show no sign of failure up to the time when it has oxidised 0-75 per cent, of carbohydrate. In a man of 73 kilos., walking at a steady speed of 5 kilometres (about 3 miles) per hour, the oxygen consumption in excess of basal is about 0-65 litre per minute (10). Let us assume that 20 kilos, of muscles are involved, and calculate how far he can go before they have utilised 0-75 per cent, of their weight of carbohydrate. The amount of carbohydrate available is 150 gm., which requires 112 litres of oxygen for its combusion. At 0 • 65 litre per minute this would last for 172 minutes, nearly 3 hours, in which time the subject would have walked 14-4 kilometres, about 9 miles. The muscles of a subject who can walk 9 miles without obvious signs of fatigue are not so “ abnormal ” that a physiologist need despair of investigating them.
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from

322 Anaerobic and Aerobic Activity in Isolated .
Summary.
1. In a thin frog’s sartorius muscle stimulated by single shocks at intervals of 10 seconds or more, in oxygen-free Ringer’s solution, the lactic acid produced can diffuse away and ordinary fatigue does not set in.
2. Under such conditions the total amount of energy set free, and lactic acid produced, before the muscle fails, is nearly three times as great as in a muscle suspended in nitrogen, where diffusion is impossible. Failure, therefore, in the latter case is due to accumulation of lactic acid : in the former, probably to exhaustion of the carbohydrate store.
3. That exhaustion of carbohydrate is the cause of ultimate failure in a muscle stimulated anaerobically, if lactic acid be removed by diffusion, is shown by the fact that the addition of 0 • 1 per cent, of glucose to the Ringer’s solution increases the amount of energy set free by 60 per cent. Cane sugar has no perceptible effect.
4. A thin sartorius muscle, in oxygenated Ringer’s solution, may be stimulated 23 or more times per minute for many hours and show no signs of fatigue. A steady state is set up in which recovery balances the break-down, the supply of oxygen by diffusion being adequate.
5. Ultimate failure is due, at any rate in part, to exhaustion of the carbohydrate store, as is shown by the fact that improvement results from glucose added to the solution.
6. In oxygenated Ringer’s solution an isolated muscle may give more than 10,000 twitches, may oxidise more than per cent, of carbohydrate, may develop a total tension of more than 6 tons per square centimetre of cross- section. These facts are considered in reference to the view, often expressed, that such muscles are “ abnormal.” The isolated muscle, without signs of failure or fatigue, may undertake an amount of work equivalent to a nine-mile walk by a man.
REFERENCES.
(1) Hill, ‘ Roy. Soc. Proc.,’ B, vol. 104, p. 39 (1928).(2) Eggleton, Eggleton and Hill, ‘ Roy. Soc. Proc.,’ B, vol. 103, p. 620 (1928).(3) Hill, ‘ Roy. Soc. Proc.,’ B, vol. 103, p. 163 (1928).(4) Stella, ‘ Journ. Physiol.,’ vol. 66, p. 19 (1928).(5) Hill, ‘ Roy. Soc. Proc.,’ B, vol. 103, p. 138 (1928).(6) Hill, ‘ Nature,’ May 11, p. 723 (1929).(7) Hill, ‘ Journ. Physiol.,’ vol. 48, p. xi (1914).(8) Hill, ‘ Roy. Soc. Proc.,’ B, vol. 103, p. 183 (1928).(9) Clark, ‘ Journ. Physiol.,’ vol. 47, p. 66 (1913).
(10) Hill, Long and Lupton, ‘ Roy. Soc. Proc.,’ B, vol. 97, p. 155 (1924).
on July 5, 2018http://rspb.royalsocietypublishing.org/Downloaded from





























