Embed Size (px)
Citation preview

743
Adenosine-Sensitive Afterdepolarizationsand Triggered Activit in Guinea Pig
Ventricular Myocytes
Yejia Song, Sheryl Thedford, Bruce B. Lerman, and Luiz Belardinelli
This study examines the cellular basis and specificity of the effects of adenosine on early afterdepolar-izations (EADs), delayed afterdepolarizations (DADs), and triggered activity (TA) induced by variousdrugs with different mechanisms of action. Membrane potential and currents were measured in isolatedguinea pig ventricular myocytes. Adenosine (10-100 ,uM) significantly (p<O.OS) reduced the amplitude ofDADs and suppressed TA induced by isoproterenol (10-50 nM) and forskolin (1 ,uM) but not thoseinduced by dibutyryl cAMP (1 ,M), ouabain (1-5 ,M), and 7.2 mM (Ca2+]0. Adenosine also abolishedEADs and TA induced by isoproterenol. In contrast, adenosine failed to abolish EADs and TA induced byquinidine (3 ,M) or those that occurred spontaneously (i.e., in the absence of drugs). Transient inwardcurrent (ITi) was induced on repolarization after 2-second-long single depolarizing voltage steps or after12-second-long trains of 300-msec depolarizing pulses. Concomitant with the attenuation of DADs,adenosine suppressed 'Ti caused by isoproterenol and forskolin but not those induced by ouabain,dibutyryl cAMP, and elevated [Ca2+]. The amplitude of 'Ti was dependent on the magnitude of theactivating voltage step, but the suppression of IT; by adenosine was not. The selective A,-adenosinereceptor antagonist N-0861 (9-methyladenine derivative) antagonized the effects of adenosine onafterdepolarizations, IT;, and TA. In myocytes from guinea pigs treated with pertussis toxin, adenosinefailed to attenuate DADs and ITi or abolish TA induced by isoproterenol or forskolin. In parallelexperiments, isoproterenol (10 nM) raised cellular cAMP from 5.7±0.2 to 8.1±0.1 pmol and the selectiveA, receptor agonist cyclopentyladenosine (5 AtM) reduced it to 6.5+±0.2 pmol (p<0.05). Thus, adenosinespecifically attenuates afterdepolarizations and abolishes TA by suppressing ITi; that are associated withstimulation of adenylate cyclase via a pertussis toxin-sensitive A, receptor-mediated action. In conclu-sion, the response of TA to adenosine may identify a mechanism of afterdepolarizations related tostimulation of adenylate cyclase. (Circulation Research 1992;70:743-753)KEY WoRDs * action potentials * voltage clamp * adenosine receptor * triggered rhythms *
arrhythmias * catecholamines * calcium overload
T here has been increasing recognition of the roleplayed by triggered activity caused by eitherearly afterdepolarizations (EADs) or delayed
afterdepolarizations (DADs) in the genesis of clinicalarrhythmias.12 EADs that arise at membrane potentialsnear the action potential plateau appear to be due to time-and voltage-dependent reactivation of the L-type calciumcurrent.34 On the other hand, DADs have been attributedto an oscillatory membrane current occurring on repolar-ization, hereinafter referred to as transient inward current(ITi).5 This current appears to be either a nonspecificcation current or a current generated by electrogenicNa-Ca exchange, and it is attributed to intracellular cal-
From the Departments of Medicine and Pharmacology, Univer-sity of Florida, Gainesville, Fla., and the Department of Medicine,Division of Cardiology, New York Hospital, Cornell UniversityMedical College, New York.
Supported in part by a grant-in-aid from the Suncoast Chapterof the American Heart Association and National Institutes ofHealth grants HL-35272 and HL-44747. B.B.L. is an EstablishedInvestigator of the American Heart Association.Address for correspondence: Luiz Belardinelli, MD, Depart-
ment of Medicine, Box J-277 JHMHC, University of Florida,Gainesville, FL 32610.
Received August 26, 1991; accepted December 4, 1991.
cium overload and spontaneous oscillatory release ofcalcium from the sarcoplasmic reticulum.6,7We previously proposed that the nucleoside adeno-
sine may identify a subset of ventricular tachyarrhyth-mias that are catecholamine dependent (i.e., cAMPmediated).8 This was based on clinical and basic obser-vations.8-10 For instance, adenosine effectively termi-nates exercise- or isoproterenol-induced ventriculartachycardias in patients with no structural heart disease,but it is ineffective in terminating reentrant ventriculartachycardia.8 In ventricular myocytes adenosine attenu-ates the increase in peak calcium inward current (Ic)caused by isoproterenol and forskolin.10,1' This effect isassociated with a decrease in cellular cAMP,10 at least inpart caused by inhibition of adenylate cyclase,12-14 and isthe basis for the anti-13-adrenergic action of adenosine.These findings have led us to suggest that attenuation ofisoproterenol-induced DADs and of triggered activityby adenosine is due to a decrease in cellular calciumoverload.9 However, none of the above studies demon-strated the linkage between attenuation by adenosine ofisoproterenol-induced ITi and the decrease in amplitudeof DADs and termination of triggered activity. Like-wise, the characterization of the effects of adenosine on
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from

744 Circulation Research Vol 70, No 4 April 1992
afterdepolarizations (EADs and DADs) and triggeredactivity resulting from drugs that alter the intracellularCa2' load of myocytes via different mechanism(s) hasnot been investigated.
Thus, in the present study, we sought to 1) examinethe effects of adenosine on EADs, DADs, and resultingtriggered activity induced by various drugs with differ-ent mechanisms of action; 2) determine whether aden-osine depresses ITi; and 3) establish the role of aninhibitory guanine nucleotide binding protein (Gi pro-tein) as mediator of the effects of adenosine by usingpertussis toxin (PTX). Finally, because adenosine inhi-bition of adenylate cyclase can be mediated by activa-tion of either the cell surface extracellular A,-adenosinereceptor14 or the intracellular purine (P) site,15 the site(intracellular or extracellular) mediating the action ofadenosine on afterdepolarizations and triggered activitywas also investigated.
Materials and MethodsChemicals
Adenosine, N6-cyclopentyladenosine (CPA), 8-cyclo-pentyltheophylline (CPT), ouabain, forskolin, dideoxy-adenosine (DDA), and bovine serum albumin werepurchased from Sigma Chemical Co., St. Louis, Mo.Dispase and N6,2'-0-dibutyryladenosine 3 ',5 '-cyclicmonophosphate (DBcAMP) were purchased fromBoehringer Mannheim, Indianapolis, Ind. Collagenasewas purchased from Worthington Biochemical Co.,Freehold, N.J., and trypsin from Serva Fine Biochemi-cals, Heidelberg, FRG. PTX was purchased from Cal-biochem Corp., San Diego, Calif., and (+)-N6-endonor-bornan-2-yl-9-methyladenine (N-0861) was a gift fromWhitby Research Inc., Richmond, Va.
SolutionsKrebs-Henseleit (K-H) solution contained (mM)
NaCl 127, KCl 4.6, CaCI2 2, MgSO4 1.1, sodium pyruvate2, glucose 10, creatine 10, taurine 20, ribose 5, adenine0.01, allopurinol 0.1, and HEPES 5, adjusted with NaOHto pH 7.4. The composition of the Ca24-free solution wasthe same as that of the K-H solution except for thedeletion of CaCl2. The enzyme solution contained 0.4mg/ml collagenase (type II), 0.04 mg/ml dispase, 0.04mg/ml trypsin, and 2 mg/ml albumin, in Ca2+-free solu-tion. The conventional micropipettes were filled with 3 MKCl, whereas the "patch electrodes" were filled with asolution (i.e., internal pipette solution) containing (mM)KCl 10, potassium aspartate 130, Na2ATP 4, MgCl2 1,and HEPES 1, and pH was adjusted to 7.2 with KOH.Note that EGTA, which is a normal constituent ofinternal pipette solutions, had to be omitted because 'Ticould not be reproducibly induced by isoproterenol orouabain when EGTA was present.
Isolation ProcedureSingle ventricular myocytes were prepared from adult
Hartley guinea pigs of either sex (weighing 300-500 g).In a separate series of studies, single ventricular myo-cytes were isolated from guinea pigs injected with PTX(150 ,ug/kg) via the jugular vein 72 hours before theexperiments. Briefly, the hearts from control (untreat-ed) and PTX-treated guinea pigs were perfused with the
a rate of 8 ml/min: 1) K-H solution for 10 minutesfollowed by 2) Ca24-free solution for 10 minutes and 3)enzyme solution for another 20 minutes. After perfu-sion with the enzyme solution, the ventricles were
removed, minced, and gently shaken for 10-20 minutes.The cells were harvested from the suspension, filtered,and stored at room temperature in K-H solution con-taining 0.1 mM Ca24.
Electrophysiological TechniquesIsolated ventricular myocytes were transferred into a
recording chamber that was mounted on the stage of aninverted microscope coupled to a video camera system.Cells were continuously superfused with K-H solution ata rate of about 2 ml/min. The bath temperature in allexperiments was maintained at 34.5+0.5°C. Transmem-brane action potentials and currents were measuredwith conventional single microelectrodes filled with 3 MKCI and resistances of 30-50 MfQ by using an AxoclampModel 2A (Axon Instruments, Burlingame, Calif.). Tofacilitate the development of DADs and 1Ti, 15-second-long drives at a rate of 2 Hz and 12-second-long trainsof 300-msec depolarizing voltage-clamp steps from -80to +40 mV were applied, respectively. The actionpotentials and current tracings were directly recordedon paper with a two-channel Gould (model 2200S) chartrecorder. Action potential durations were measured at50% (APD50) and 90% (APD9o) of repolarization. Theamplitude of DAD was taken as the difference betweenthe maximum diastolic potential and the peak of thefirst DAD after the stimulation train. ITi was measuredfrom the holding current to the peak inward currentafter the last depolarizing clamp pulse.
In experiments designed to investigate the effects ofadenosine on the IT, induced by isoproterenol andouabain, the whole-cell voltage-clamp method, as de-scribed by Hamill et al,16 was used. The patch electrodeshad resistances of 2-3 Mfl when filled with the internalpipette solution described above. The junctional poten-tial was corrected by zeroing the current before themicroelectrode contacted the surface of the myocyte.When the cell membrane was ruptured, the holdingpotential was set at the zero current level (restingmembrane potential). Recordings were obtained involtage-clamp mode by using an Axopatch Model 1C(Axon). To inactivate the fast sodium current, theholding potential was set at -40 mV. To construct thecurrent-voltage relations for 1Ti' 2-second-long depolar-izing voltage-clamp pulses from holding potential topotentials ranging between -30 and +80 mV were
applied at a rate of 0.1 Hz. Membrane currents weredisplayed on a storage oscilloscope (model 5113, Tek-tronix, Beaverton, Ore.) and also digitized at the sam-pling frequency of 40 kHz on-line using a 25-MHzNorthgate Computer (model Slimline 386, Minneapolis,Minn.) and PCLAMP software version 5.5 from AxonInstruments, Foster City, Calif.
cAMP AssayGuinea pig ventricular myocytes, isolated by the proce-
dure described above, were allowed to settle at room
temperature in K-H solution before drug incubation. Withthe supematant removed, the cell suspension was concen-
following solutions gassed with 100% 02 at 35°C and at trated to about 1.6X 1W rod-shaped cells per milliliter and
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from

Song et al Effect of Adenosine on Afterdepolarizations 745
then aliquoted (400 ,ul). At a constant temperature of35°C, the cells were pretreated with CPT and/or CPA orvehicle, as indicated, for 2 minutes and then incubatedwith isoproterenol for another 2 minutes. At the end ofthe incubation, 592 jdu of 85 mM HCl was added to eachsample to stop the reaction and lyse the cells. A 500-glaliquot of each sample was acetylated and analyzed forcAMP content by using the automated radioimmunoassaymethod of Brooker et al.17
Data AnalysisValues are given as mean+SEM. The paired Stu-
dent's t test and Duncan's multiple range test were usedfor the statistical analysis. Differences between valueswere considered statistically significant atp<0.05.
ResultsGeneral
Ventricular myocytes superfused with the normalK-H solution (4.6 mM K' and 2.0 mM Ca2') at34.5 +0.5°C had a resting membrane potential of -84+ 3mV. On stimulation (0.25 Hz), transmembrane actionpotentials were elicited. The action potential ampli-tudes were 130±3 mV and had overshoots of +40±2mV. APD50 and APD%0 were 204±10 and 236±10 msec(n= 20), respectively.
Afterdepolarizations and triggered activity, whichwere rarely observed during control conditions (i.e.,absence of drugs) could be reproducibly induced byisoproterenol, forskolin, ouabain, DBcAMP, and quini-dine and by raising the extracellular calcium concentra-tion from 2.0 to 7.2 mM. The afterdepolarizations andtriggered activity caused by the various drugs sometimesappeared during stimulation of the myocytes at a rate of0.25 Hz but were more easily elicited with overdrivestimulation (15-second-long drive at 2 Hz). Coincidentwith the appearance of DADs, in the same myocytes, 'Ticould be induced with 12-second-long trains of 300-msec depolarizing pulses from a holding potential of-80 to +40 mV (see "Materials and Methods").
Afterdepolarizations and Triggered Activity Inducedby Isoproterenol and Forskolin
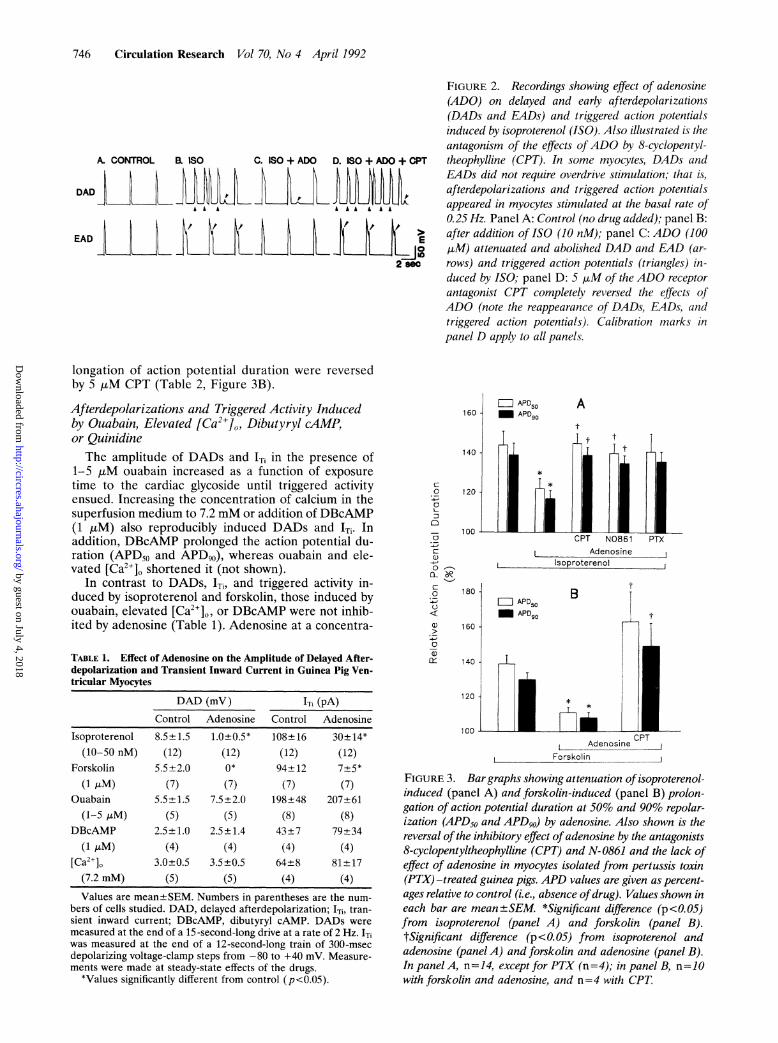
Isoproterenol. In the presence of isoproterenol (10-50nM), overdrive stimulation reproducibly elicited DADs(Figures 1 and 2, Table 1), which sometimes developedinto triggered activity. Adenosine attenuated the iso-proterenol-induced DADs and 1Ti (Figure 1, Table 1)and terminated triggered activity. Adenosine (10-100,uM) significantly reduced the amplitude of DADs andITi by 87% and 72%, respectively (Table 1). In somemyocytes, isoproterenol elicited DADs, EADs, andtriggered activity without requiring overdrive stimula-tion (Figure 2). Adenosine also attenuated and abol-ished these afterdepolarizations and triggered activity(Figure 2C). Consistent with its anti-f3-adrenergic ef-fects on the afterdepolarizations, 1Ti, and triggeredactivity, in the same myocytes, adenosine significantlyreduced isoproterenol-induced prolongation of APD50and APD,1 by 50% and 56%, respectively (Figure 3A).As shown in Table 2 and Figures 1 and 2, the inhibitoryeffects of adenosine on isoproterenol-induced DADsand 'Ti were significantly antagonized by the A,-adeno-sine selective non-xanthine receptor blocker N-0861
A. CONTROL. s4 C m V
B. ISO
C. ISO + ADO
WU149D. ISO + ADO + N0861
0.2 sec i sec 2 e
FIGURE 1. Effect of adenosine (ADO) on delayed afterde-polarizations and transient inward current induced by isopro-terenol (ISO). Also illustrated is the antagonism of the effectsofADO by the selective A,-ADO receptor antagonist N- 0861.Each panel illustrates a single action potential recorded at abasal stimulation rate of 0.25 Hz, followed by the last threeaction potentials (middle column) and membrane currentrecordings (far right column) of a 15-second-long overdrivestimulation (stim) applied at a frequency of 2 Hz and a12-second-long train of300-msec depolarizing pulses, respec-tively. Only the lowerpart of the action potentials (middle) isdepicted. Delayed afterdepolarizations (straight arrows) andtransient inward currents (curved arrows) were recorded atthe end of overdrive stimulation and depolarizing voltage-clamp pulses, respectively. Panel A: Control (without drugadded); panel B: after 10 nMISO (closed triangle points to asingle triggered action potential); panel C: ISO plus 100 ,uMADO; panel D: in the presence ofISO, ADO, and 10 ,uM oftheA1-ADO antagonist N-0861. ADO abolished ISO-induceddelayed afterdepolarization and transient inward current andN-0861 reversed this effect ofADO. Note that the duration ofthe action potentials showed corresponding changes. Allrecordings (action potentials and currents) are from a singleventricular myocyte. Calibration marks in panelD apply to allpanels.
(Figure iD) and by the xanthine derivative CPT (Figure2D). Likewise, inhibition by adenosine of the isopro-terenol-induced prolongation of the APD50 and APD90was antagonized 100% and 96%, respectively, by 5 ,MCPT or 10 MuM N-0861 (Figure 3A). Not shown, neitherN-0861 nor CPT alone or in combination with isopro-terenol caused any significant detectable change inaction potential configuration. That is, these com-pounds only antagonized the effects of adenosine.
Forskolin. As in the case of isoproterenol, adenosinesignificantly diminished the amplitude of DADs and 'Ticaused by 1 ,M forskolin by 98% and 93%, respec-tively (Table 1). In the same myocytes, adenosineshortened forskolin-induced prolongation of APD50and APD90 by as much as 72% and 73%, respectively(Figure 3B). Similar to isoproterenol, the effects ofadenosine on forskolin-induced DADs, 1Ti, and pro-
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from
746 Circulation Research Vol 70, No 4 April 1992
A. CONTROL ISO C. ISO + ADO D. ISO + ADO + CPT
DADLA A A>A
EAD 1E
2 86c
longation of action potential duration were reversedby 5 ,AM CPT (Table 2, Figure 3B).
Afterdepolarizations and Triggered Activity Inducedby Ouabain, Elevated [Ca2"J0, Dibutyryl cAMP,or QuinidineThe amplitude of DADs and 1Ti in the presence of
1-5 ptM ouabain increased as a function of exposuretime to the cardiac glycoside until triggered activityensued. Increasing the concentration of calcium in thesuperfusion medium to 7.2 mM or addition of DBcAMP(1 ,uM) also reproducibly induced DADs and 1Ti. Inaddition, DBcAMP prolonged the action potential du-ration (APD5O and APD90), whereas ouabain and ele-vated [Ca2']0 shortened it (not shown).
In contrast to DADs, 1Ti, and triggered activity in-duced by isoproterenol and forskolin, those induced byouabain, elevated [Ca2']0, or DBcAMP were not inhib-ited by adenosine (Table 1). Adenosine at a concentra-
TABLE 1. Effect of Adenosine on the Amplitude of Delayed After-depolarization and Transient Inward Current in Guinea Pig Ven-tricular Myocytes
DAD (mV) ITi (pA)Control Adenosine Control Adenosine
Isoproterenol 8.5-+-1.5 1.0+0.5* 108+--16 30±14*
(10-50 nM) (12) (12) (12) (12)Forskolin 5.5+2.0 0* 94±12 7+5*
(1 gM) (7) (7) (7) (7)Ouabain 5.5±1.5 7.5±2.0 198±48 207±61
(1-5 gM) (5) (5) (8) (8)DBcAMP 2.5-+-1.0 2.5±+ 1.4 43±7 79-+-34
(1 gM) (4) (4) (4) (4)[Ca2+ I 3.0±0.5 3.5±0.5 64+8 81+17
(7.2 mM) (5) (5) (4) (4)
Values are mean±SEM. Numbers in parentheses are the num-bers of cells studied. DAD, delayed afterdepolarization; 1Ti, tran-sient inward current; DBcAMP, dibutyryl cAMP. DADs weremeasured at the end of a 15-second-long drive at a rate of 2 Hz. 'Tiwas measured at the end of a 12-second-long train of 300-msecdepolarizing voltage-clamp steps from -80 to +40 mV. Measure-ments were made at steady-state effects of the drugs.
*Values significantly different from control (p<0.05).
FIGURE 2. Recordings showing effect of adenosine(ADO) on delayed and early afterdepolarizations(DADs and EADs) and triggered action potentialsinduced by isoproterenol (1SO). Also illustrated is theantagonism of the effects ofADO by 8-cyclopentyl-theophylline (CPT). In some myocytes, DADs andEADs did not require overdrive stimulation, that is,afterdepolarizations and triggered action potentialsappeared in myocytes stimulated at the basal rate of0.25 Hz. Panel A: Control (no drug added); panel B:after addition of ISO (10 nM); panel C: ADO (100,uM) attenuated and abolished DAD and EAD (ar-rows) and triggered action potentials (triangles) in-duced by ISO; panel D: 5 ,uM of the ADO receptorantagonist CPT completely reversed the effects ofADO (note the reappearance of DADs, EADs, andtriggered action potentials). Calibration marks inpanel D apply to all panels.
c0
00U
CQ)0-40no\'1-:
>
C-
180 -
160 -
140
120
100
L AdenosinelIsoproterenol
t
= APDs I3EIJP50 tM APD9
CPTAdenosine
Forskolin
FIGURE 3. Bargraphs showing attenuation of isoproterenol-induced (panel A) and forskolin-induced (panel B) prolon-gation of action potential duration at 50% and 90% repolar-ization (APD50 and APD90) by adenosine. Also shown is thereversal ofthe inhibitory effect ofadenosine by the antagonists8-cyclopentyltheophylline (CPT) and N-0861 and the lack ofeffect of adenosine in myocytes isolated from pertussis toxin(PTX) -treated guinea pigs. APD values are given as percent-ages relative to control (i.e., absence ofdrug). Values shown ineach bar are mean±SEM. *Significant difference (p<0.05)from isoproterenol (panel A) and forskolin (panel B).tSignificant difference (p<0.05) from isoproterenol andadenosine (panel A) and forskolin and adenosine (panel B).In panel A, n=14, except for PTX (n=4); in panel B, n=10with forskolin and adenosine, and n-4 with CPT.
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from

Song et al Effect of Adenosine on Afterdepolarizations 747
TABLE 2. Reversal of the Effects ofAdenosine on the Amplitude ofDelayed Afterpolarization and Transient Inward Current by Aden-osine Receptor Antagonists in Guinea Pig Myocytes
DAD (mV) IT (pA)
Isoproterenol Forskolin Isoproterenol Forskolin
Control 8.4±2.0 3.0±0.7 108±16 88±10
(6) (4) (12) (4)Adenosine 1.0±0.7* 0.2±0.2* 30±14* 13±7*
(6) (4) (12) (4)Adenosine 6.5 ± 1.7t 5.8±1.7t 97±16t 123 +28t+CPT (6) (4) (12) (4)
Adenosine 7.3±1.7t ... 151±32t+N-0861 (6) (5)
Values are mean±SEM. Numbers in parentheses are the num-bers of cells studied. DAD, delayed afterdepolarization; 1Ti, tran-sient inward current; CPT, cyclopentyltheophylline; N-0861, N6-endonorbornyl-9-methyladenine. DADs were measured at the endof a 15-second-long drive at a rate of 2 Hz. 1Ti was measured at theend of a 12-second-long train of 300-msec depolarizing voltage-clamp steps from -80 to +40 mV. Measurements were made atsteady-state effects of the drugs. The concentrations of the drugswere as follows: isoproterenol, 10 nM; forskolin, 1 ,uM; adenosine,100 ,LM; CPT, 5 ^M; and N-0861, 10 gM.
*Values significantly different from control (p<0.05). tValuessignificantly different from isoproterenol + adenosine andforskolin+adenosine.
tion as high as 100 ,uM did not reduce the amplitude ofDADs, 1Ti, and triggered activity elicited by ouabain,elevated [Ca2+]0, or DBcAMP (Table 1), nor did itreverse the changes in action potential configurationcaused by these interventions (not shown). DADs, 1Ti,and triggered activity induced by ouabain, DBcAMP,and 7.2 mM [Ca2+]l were attenuated and/or abolishedafter washout of the drugs.
Quinidine (2-3 ,M) markedly increased the actionpotential duration and often induced EADs that even-tually led to triggered activity (Figure 4). These EADsand triggered activity, unlike those induced by isopro-terenol, were not attenuated by adenosine (100 ,uM) butwere abolished by increasing the concentration of mag-nesium in the superfusion medium to 5 mM (Figure4D).As indicated above, in the course of this study,
spontaneous (i.e., in the absence of drugs) occurrenceof DADs, EADs, and triggered activity was relativelyinfrequent. Not shown, spontaneously occurring EADsthat arise from the action potential plateau from anotherwise normal and rod-shaped myocyte with a rest-ing potential of -80 mV were not attenuated by aden-osine (100 ,uM). Similarly, not shown, in other myocyteswith normal resting potential that developed spontane-ous DADs and/or triggered activity, adenosine (100,uM) and its A1 receptor-selective analogue CPA (5,uM) failed to attenuate or abolish the DADs andtriggered activity, respectively.
Dependence of Transient Inward Current Induced byIsoproterenol and Ouabain on the Amplitude of theVoltage-Clamp StepTo further elucidate the underlying electrophysiolog-
ical basis of adenosine's effect on DADs, the influenceof the depolarizing clamp step voltage on the amplitudeOf 'Ti induced by isoproterenol and ouabain in the
A. CONTROL
B. QUINIDINE
C. QUINIDINE + ADO
D. QUINIDINE + 5mMEMgJ0
0.2 SC 2 secFIGURE 4. Lack of effect of adenosine (ADO) on earlyafterdepolarizations induced by quinidine. Action potentialswere recordedfrom cells stimulated at a basal rate of 0.25 Hzthroughout the experiment. Panels A-C: Actionpotentials arefrom a single ventricular myocyte. Arrows point to earlyafterdepolarizations. Quinidine-induced early afterdepolar-izations were not attenuated by ADO (panel C) but wereabolished by raising the concentration of magnesium (panelD). Calibration marks in panel D apply to all panels.
absence and presence of adenosine was determined.Figure 5 shows membrane currents elicited by a 2-sec-ond-long depolarizing clamp pulse to +20 mV from aholding potential of -40 mV. During control conditions(i.e., absence of drugs), 1Ti was not observed (Figure 5,top tracing). In the presence of 10 nM isoproterenol(within 2-3 minutes of application), the amplitude of Icincreased and 1Tis were observed after the depolarizingclamp step (Figure 5, middle tracing). The addition of100 ,uM adenosine, still in the presence of isoprotere-nol, rapidly abolished 'Ti and reduced the amplitude ofICa to baseline (Figure 5, bottom tracing). The relationbetween the voltage of the depolarizing step and themagnitude of 'Ti induced by isoproterenol (Figure 6A)and ouabain (Figure 6B) is sigmoidal, with an activationthreshold between -20 and -10 mV, and maximalcurrent was observed at voltages greater than +40 mV.Adenosine (100 ,M), independent of membrane poten-tial, abolished ITi induced by isoproterenol (Figure 6A),whereas the amplitude of ouabain-induced 'Tis wasunaffected by adenosine (Figure 6B).
Mechanism ofAction ofAdenosine on DelayedAfterdepolarizations, Triggered Activity, andTransient Inward Current Caused by IsoproterenolThe mechanism underlying the functional antago-
nism between adenosine and catecholamines is at leastin part due to inhibition of the cAMP-producing en-zyme adenylate cyclase.12-14 This inhibitory effect ofadenosine is presumably mediated by the extracellularA1-adenosine receptor but could also involve the inhib-itory intracellular P site.15 Thus, studies were performedto determine the effects of the adenosine P site agonistDDA on isoproterenol-induced afterdepolarizations,triggered activity, and 'Ti. In addition, to determine
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from

748 Circulation Research Vol 70, No 4 April 1992
CONTROL
0.5 :]0
-0.5
-1
0-
aE
Q0
c:
(DL-
.0c11)
0 0.5 1 1.5 2 2.5 3
1 - ISO (1 o-8M)0
-0.5 l-1 h . ,
0 0.5 1 1.5 2 2.5 3
ISO (108M) + ADO (10-4M)0.5
0-
-0.5
-1
0 0.5 1 1.5 2 2.5 3TIME (Sec)
FIGURE 5. Effect of adenosine (ADO) on isoproterenol(ISO)-induced transient inward current in a guinea pigventricular myocyte. Whole-cell membrane currents were
elicited by a 2-second-long depolarizing voltage-clamp pulseto +20 mVfrom a holding potential of -40 mV Top panel:Control tracing. Middle panel: Tracing 3 minutes afteraddition ofISO. Note the appearance of the transient inwardcurrent (indicated by the large arrow) after return to holdingpotential. ISO also increased the peak inward current (smallarrow). Bottom panel: Tracing showing ADO completelyabolished the transient inward current and attenuated theincrease in the peak inward current caused by ISO. Allmembrane current recordings are from the same myocyte.
whether the electrophysiological antagonism of cate-cholamines by adenosine is associated with changes incellular cAMP, the effects of the A1-adenosine selectiveagonist CPA on isoproterenol-induced cAMP accumu-lation in ventricular myocytes were investigated. Finally,to establish the involvement of an inhibitory G proteinmediating the effects of adenosine, experiments werecarried out in myocytes isolated from guinea pigs pre-treated with PTX.
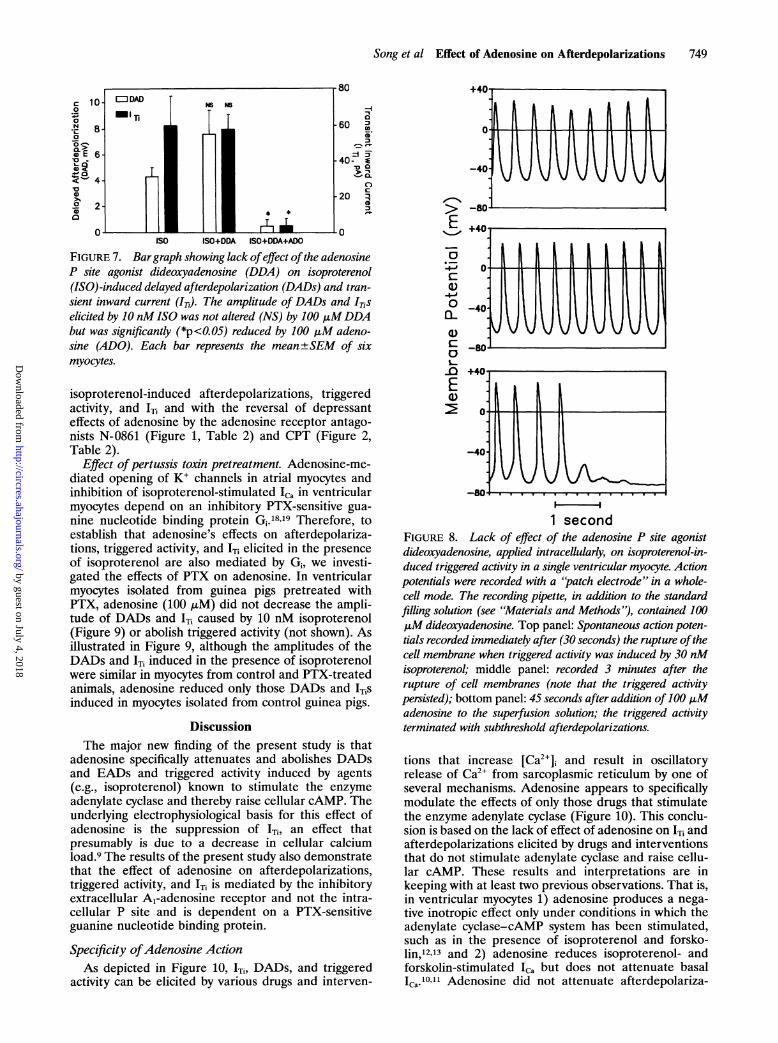
Role of the intracellular P site. Stimulation of theintracellular P site is another mechanism whereby aden-osine can modulate adenylate cyclase.15 In a separateseries of experiments, the effects of the P site selectiveagonist DDA, applied extracellularly and intracellu-larly, on isoproterenol-induced afterdepolarizations and'Ti were investigated. As summarized in Figure 7, DADsand 'Ti (not shown) caused by 10 nM isoproterenol werenot attenuated by 100 ,M DDA applied extracellularlybut were abolished by 100 gM adenosine. The potentialrole of the intracellular P site in mediating the actions ofadenosine was further examined by intracellular appli-cation of DDA (Figure 8). In these experiments (n=3),the recording microelectrodes (patch electrodes) werefilled with a solution containing 100 ,uM DDA. In the
Q-E
01)
13.-
E
S -4
0.8
0.6
0.4
0.2
0.0
1.2
1.0
0.8
0.6
-40 -20 0 20 40 60 80
04 '
0.2 [_________________0.0[_____________
-40 -20 0 20 40 60 80
Membrane Potential (mV)FIGURE 6. Graphs showing effect of adenosine (ADO) on therelation between the magnitude of the depolarizing voltage-clamp pulses and the amplitude ofthe transient inward current(Ip) induced by isoproterenol (ISO, panel A) or ouabain (panelB). Amplitude of Ir, is plotted against the membrane potentiaL.The cells were depolarized by a series of2-second-long voltage-clamp pulses in the range of -30 to +80 mVfrom a holdingpotential of -40 mV Panel A: ADO (o, 100 ,M) completelyabolished Il- induced byISO (0, 10 nM);panelB:ADO (o, 100pgM) had no effect on IT induced by ouabain (0, 2 j.tM). Eachdata point is the mean±SEM of three orfour myocytes. Curveswere drawn on a computer using a linear curve-fitting program.Not shown, durng control (i.e., absence of drug), IT, was notobserved in any of the cells in this series.
example illustrated in Figure 8, triggered activity in-duced in the presence of isoproterenol was not slowedor terminated by internal perfusion of the myocyte withDDA, but it was abolished by extracellular applicationof adenosine (10 ,M).
Changes in cellular cAMP. The effect of the A1-adenosine agonist CPA and of the adenosine receptorantagonist CPT on isoproterenol-stimulated cAMP ac-cumulation in ventricular myocytes was also investi-gated. The control (basal) cAMP level was 5.69+0.18pmol. Isoproterenol (10 nM) alone caused a significantincrease in cellular cAMP to 8.14+0.11 pmol. Theaddition of 5 ,uM CPA, in the continued presence ofisoproterenol, significantly reduced the cellular cAMPaccumulation to 6.55+0.19 pmol. CPA alone had nosignificant effect on the basal levels of cAMP. Theattenuation by CPA of isoproterenol-induced increasein cellular cAMP accumulation was completely antago-nized by the adenosine antagonist CPT. That is, in thepresence of isoproterenol plus CPA, the addition of 10,uM CPT caused a significant increase in cAMP, from6.55+0.19 to 8.90+0.20 pmol. These data on cAMPlevels are consistent with the effects of adenosine on
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from
Song et al Effect of Adenosine on Afterdepolarizations 749
-IA0a
-60 CC.
-40- *0a>
CL
-20 ia
ISO ISO+DDA ISO+DDA+ADO
FIGURE 7. Bargraph showing lack ofeffect ofthe adenosineP site agonist dideoxyadenosine (DDA) on isoproterenol(ISO)-induced delayed afterdepolarization (DADs) and tran-sient inward current (IT). The amplitude of DADs and ITiselicited by 10 nMISO was not altered (NS) by 100 jiMDDAbut was significantly (*p<0.05) reduced by 100 jiM adeno-sine (ADO). Each bar represents the mean+±SEM of sixmyocytes.
isoproterenol-induced afterdepolarizations, triggeredactivity, and 'Ti and with the reversal of depressanteffects of adenosine by the adenosine receptor antago-nists N-0861 (Figure 1, Table 2) and CPT (Figure 2,Table 2).
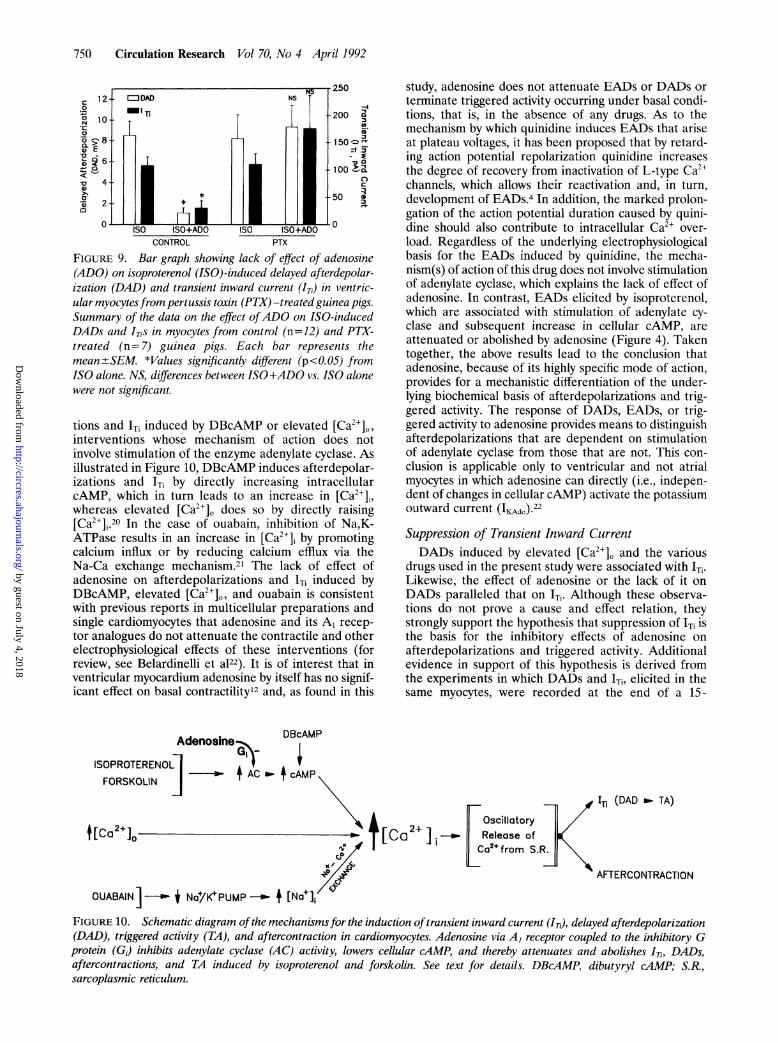
Effect ofpertussis toxin pretreatment. Adenosine-me-diated opening of K' channels in atrial myocytes andinhibition of isoproterenol-stimulated Ic, in ventricularmyocytes depend on an inhibitory PTX-sensitive gua-nine nucleotide binding protein Gi.18,9 Therefore, toestablish that adenosine's effects on afterdepolariza-tions, triggered activity, and ITi elicited in the presenceof isoproterenol are also mediated by Gi, we investi-gated the effects of PTX on adenosine. In ventricularmyocytes isolated from guinea pigs pretreated withPTX, adenosine (100 ,uM) did not decrease the ampli-tude of DADs and 'Ti caused by 10 nM isoproterenol(Figure 9) or abolish triggered activity (not shown). Asillustrated in Figure 9, although the amplitudes of theDADs and 'Ti induced in the presence of isoproterenolwere similar in myocytes from control and PTX-treatedanimals, adenosine reduced only those DADs and 1TiSinduced in myocytes isolated from control guinea pigs.
DiscussionThe major new finding of the present study is that
adenosine specifically attenuates and abolishes DADsand EADs and triggered activity induced by agents(e.g., isoproterenol) known to stimulate the enzymeadenylate cyclase and thereby raise cellular cAMP. Theunderlying electrophysiological basis for this effect ofadenosine is the suppression of 1Ti, an effect thatpresumably is due to a decrease in cellular calciumload.9 The results of the present study also demonstratethat the effect of adenosine on afterdepolarizations,triggered activity, and 'Ti is mediated by the inhibitoryextracellular A1-adenosine receptor and not the intra-cellular P site and is dependent on a PTX-sensitiveguanine nucleotide binding protein.
Specificity ofAdenosine ActionAs depicted in Figure 10, 1Ti, DADs, and triggered
activity can be elicited by various drugs and interven-
E
C}* _
4-,
c:~-4-,i0Ia-0
c:
L.-0E:2
10
-4
-50'
+401
-40.
-80+40j
-40
1 secondFIGURE 8. Lack of effect of the adenosine P site agonistdideoxyadenosine, applied intracellularly, on isoproterenol-in-duced triggered activity in a single ventricular myocyte. Actionpotentials were recorded with a 'patch electrode" in a whole-cell mode. The recording pipette, in addition to the standardfilling solution (see "Materials and Methods"), contained 100,uM dideoxyadenosine. Top panel: Spontaneous action poten-tials recorded immediately after (30 seconds) the rupture ofthecell membrane when triggered activity was induced by 30 nMisoproterenol; middle panel: recorded 3 minutes after therupture of cell membranes (note that the triggered activitypersisted); bottom panel: 45 seconds after addition of 100 jiMadenosine to the superfusion solution; the triggered activityterminated with subthreshold afterdepolarizations.
tions that increase [Ca2"]i and result in oscillatoryrelease of Ca2' from sarcoplasmic reticulum by one ofseveral mechanisms. Adenosine appears to specificallymodulate the effects of only those drugs that stimulatethe enzyme adenylate cyclase (Figure 10). This conclu-sion is based on the lack of effect of adenosine on ITi andafterdepolarizations elicited by drugs and interventionsthat do not stimulate adenylate cyclase and raise cellu-lar cAMP. These results and interpretations are inkeeping with at least two previous observations. That is,in ventricular myocytes 1) adenosine produces a nega-tive inotropic effect only under conditions in which theadenylate cyclase-cAMP system has been stimulated,such as in the presence of isoproterenol and forsko-lin,12'3 and 2) adenosine reduces isoproterenol- andforskolin-stimulated Ic but does not attenuate basallCa.10'11 Adenosine did not attenuate afterdepolariza-
YUJUII
* 40.
-B . . . . . . .. . . l . . . . .
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from
750 Circulation Research Vol 70, No 4 April 1992
10
20010 10--- 3~~~~~10~
CL>~~~~~~~~~~~~~~~~~~~~~~C
V 4- ca * ~~~~~~~~~~~500
ii2*0
-
ISO ISO+ADO ISO ISO+ADOCONTROL PTX
FIGURE 9. Bar graph showing lack of effect of adenosine(ADO) on isoproterenol (ISO)-induced delayed afterdepolar-ization (DAD) and transient inward current (IT,) in ventric-ular myocytesfrom pertussis toxin (PTX) -treated guinea pigs.Summary of the data on the effect ofADO on ISO-inducedDADs and ITis in myocytes from control (n=12) and PTX-treated (n=7) guinea pigs. Each bar represents themean ±+SEM. *Values significantly different (p<O.05) fromISO alone. NS, differences between ISO+ADO vs. ISO alonewere not significant.
tions and 'Ti induced by DBcAMP or elevated [Ca']j0,interventions whose mechanism of action does notinvolve stimulation of the enzyme adenylate cyclase. Asillustrated in Figure 10, DBcAMP induces afterdepolar-izations and 'Ti by directly increasing intracellularcAMP, which in turn leads to an increase in [Ca2+]Lwhereas elevated [Ca2+]0 does so by directly raising[Ca2'].20 In the case of ouabain, inhibition of Na,K-ATPase results in an increase in [Ca21]i by promotingcalcium influx or by reducing calcium efflux via theNa-Ca exchange mechanism.21 The lack of effect ofadenosine on afterdepolarizations and 'Ti induced byDBcAMP, elevated [Ca2+']0, and ouabain is consistentwith previous reports in multicellular preparations andsingle cardiomyocytes that adenosine and its A, recep-tor analogues do not attenuate the contractile and otherelectrophysiological effects of these interventions (forreview, see Belardinelli et a122). It is of interest that inventricular myocardium adenosine by itself has no signif-icant effect on basal contractility'2 and, as found in this
study, adenosine does not attenuate EADs or DADs orterminate triggered activity occurring under basal condi-tions, that is, in the absence of any drugs. As to themechanism by which quinidine induces EADs that ariseat plateau voltages, it has been proposed that by retard-ing action potential repolarization quinidine increasesthe degree of recovery from inactivation of L-type Ca2'channels, which allows their reactivation and, in turn,development of EADs.4 In addition, the marked prolon-gation of the action potential duration caused by quini-dine should also contribute to intracellular Ca'+ over-load. Regardless of the underlying electrophysiologicalbasis for the EADs induced by quinidine, the mecha-nism(s) of action of this drug does not involve stimulationof adenylate cyclase, which explains the lack of effect ofadenosine. In contrast, EADs elicited by isoproterenol,which are associated with stimulation of adenylate cy-clase and subsequent increase in cellular cAMP, areattenuated or abolished by adenosine (Figure 4). Takentogether, the above results lead to the conclusion thatadenosine, because of its highly specific mode of action,provides for a mechanistic differentiation of the under-lying biochemical basis of afterdepolarizations and trig-gered activity. The response of DADs, EADs, or trig-gered activity to adenosine provides means to distinguishafterdepolarizations that are dependent on stimulationof adenylate cyclase from those that are not. This con-clusion is applicable only to ventricular and not atrialmyocytes in which adenosine can directly (i.e., indepen-dent of changes in cellular cAMP) activate the potassiumoutward current (IKXdo).22Suppression of Transient Inward CurrentDADs induced by elevated [Ca2+],, and the various
drugs used in the present study were associated with ITi.Likewise, the effect of adenosine or the lack of it onDADs paralleled that on 1Ti. Although these observa-tions do not prove a cause and effect relation, theystrongly support the hypothesis that suppression of 1Ti isthe basis for the inhibitory effects of adenosine onafterdepolarizations and triggered activity. Additionalevidence in support of this hypothesis is derived fromthe experiments in which DADs and ITi, elicited in thesame myocytes, were recorded at the end of a 15-
DBcAMP
ISOPROTERENOL I vFORSKOLIN _ AC o. + cAMP
t[Ca2+io0
OUABAIN]-K $ Na+/K+PUMP -- + [Na+];
[Ca2+ _
I- (DAD _ TA)
hAFTERCONTRACTION
FIGURE 10. Schematic diagram ofthe mechanismsfor the induction oftransient inward current (ITi), delayed afterdepolarization(DAD), triggered activity (TA), and aftercontraction in cardiomyocytes. Adenosine via A1 receptor coupled to the inhibitory Gprotein (G) inhibits adenylate cyclase (AC) activity, lowers cellular cAMP, and thereby attenuates and abolishes ITi, DADs,aftercontractions, and TA induced by isoproterenol and forskolin. See text for details. DBcAMP, dibutyryl cAMP; S.R.,sarcoplasmic reticulum.
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from

Song et al Effect of Adenosine on Afterdepolarizations 751
second-long train of action potentials (overdrive stimu-lation) and depolarizing voltage-clamp steps, respec-tively. Because DADs and 'Ti were recorded undersimilar experimental conditions and in the same myo-cytes, a more direct comparison between them shouldbe possible. Moreover, the depolarizing voltage-clampsteps from -80 to +40 mV and 300 msec in duration arein the same range as the resting potential, overshoot,and action potential duration (APD9o) recorded duringisoproterenol, forskolin, and DBcAMP. The relation of'Ti to adenosine's inhibition of isoproterenol-inducedDADs is also supported by the results of the experi-ments that investigated the influence of the activatingdepolarizing step on the amplitude of ITi (Figure 6). Theeffect of adenosine on 'Ti induced in the presence ofisoproterenol (Figure 6A) or lack of it in the case ofouabain (Figure 6B) was independent of the activatingvoltage-clamp step. The dependence of the magnitudeof isoproterenol- and ouabain-induced ITi on the ampli-tude of the depolarizing clamp step is similar to resultsdescribed for 'Ti induced by strophanthidin23 and morerecently rose bengal24 in ventricular myocytes fromguinea pig and rabbit hearts, respectively.
Detection and quantification of 'Ti may be affected byactivation of other ion currents. However, the closelyparallel responses of DADs and 'Ti to adenosine are toocoincidental to simply be attributed to changes in somecontaminating current(s). Therefore, suppression of 'Ti isthe most plausible explanation for the attenuation andabolition of DADs and triggered activity by adenosine.
Cellular calcium overload followed by spontaneousoscillatory release of calcium from sarcoplasmic reticu-lum appears to induce a nonspecific cation currentand/or a Na-Ca exchange current referred to as ITi.6'7 Inthis context, adenosine, by attenuating the increase inICa and the prolongation of action potential caused byisoproterenol and forskolin, is expected to renormalizethe excess cellular calcium load produced by theseagents. Hence, suppression of 'Ti by adenosine mayreflect a reduction in cellular calcium overload andthereby a decrease in the spontaneous oscillatory re-lease of calcium from the sarcoplasmic reticulum. In thepresent study, increased cellular calcium influx to raise[Ca2"], may have occurred via different mechanismsdepending on the drug used to elicit 'Ti (Figure 10). Inthe case of isoproterenol, as shown in Figure 5, en-hanced cellular calcium influx occurred via an increasein ICa and prolongation of the action potential. Similarly,forskolin and DBcAMP have also been shown to in-crease 'Ca and prolong the action potential duration inventricular myocytes.10"' Ouabain, increased cellularcalcium influx, and/or decreased calcium efflux mostlikely occurred via an increase in Na-Ca exchangecaused by a rise in intracellular sodium.5,21'25 Thus, thestimulation of Ilc, Na-Ca exchange, and the prolonga-tion of the action potential may alone, or combined,create a state of cellular calcium overload, which in turnleads to activation of 1Ti. In the present study, calciumoverload was presumably further exacerbated by the15-second-long overdrive stimulation or depolarizingvoltage-clamp steps. As for quinidine, as discussedabove, prolongation of the action potential repolariza-tion should accelerate L-type ICa recovery4 and hencelead to increased cellular calcium influx.
The evidence for increased cellular calcium load withthe drugs used in the present study is based on theobservation (not shown) that DADs were associatedwith aftercontractions (detected with a cell-edge track-ing device). In addition, we previously showed thatDADs and aftercontractions induced with isoproterenolare both attenuated by adenosine (Figure 8 of Refer-ence 9). However, this is only indirect evidence andfurther experiments in which intracellular calcium ismeasured directly will be required to conclusively dem-onstrate that adenosine reduces cellular calcium load.
Mechanism ofAdenosine ActionModulation of adenylate cyclase activity and cellular
cAMP levels by adenosine is mediated by extracellularadenosine receptors (A, and A2 subtypes) and via theintracellular P site.26 Activation of the A,-adenosinereceptor and the P site inhibits, whereas activation ofthe A2-adenosine receptor stimulates, adenylate cyclaseactivity with corresponding changes in cellular cAMP.26A1-adenosine receptor. In contrast to activation of the
potassium outward current, IKAdo, which is independentof cAMP, attenuation by adenosine of isoproterenol-and forskolin-stimulated 'Ca is due to inhibition ofadenylate cyclase and is accompanied by a concomitantreduction in cellular cAMP levels.22 In the presentstudy, these previous observations were not only con-firmed but also extended to include adenosine's effecton ITi and resulting DADs. Furthermore, the resultslead to the conclusion that inhibition by adenosine of 'Tiand DADs elicited in the presence of isoproterenol andforskolin is mediated by the cell surface A1 receptorsubtype, which is coupled to an inhibitory PTX-sensi-tive guanine nucleotide binding protein, most likely Gi.Evidence implicating the extracellular A1-adenosine re-ceptor as the mediator of the effects of adenosine in thisstudy is the complete reversal of adenosine's effect bythe antagonists CPT and N-0861. The xanthine CPT isa potent (pA2 value of 7.5)27 and competitive adenosineantagonist that is slightly selective for A, receptors (i.e.,with an affinity AJ/A2 of 140).28 In comparison, in theheart, the newly developed nonxanthine adenosine an-tagonist N-0861, which is also a competitive antagonist,is less potent (pA2 value of 6.2)29 than CPT but is devoidof functional A2 receptor antagonism. That is, pharma-cologically, N-0861 behaves as a pure A1-adenosineantagonist, which does not attenuate the coronary va-sodilation (A2 receptor-mediated effect) caused byadenosine.The failure of adenosine to attenuate and abolish
isoproterenol- and forskolin-induced 'Ti, DADs, andtriggered activity in myocytes obtained from PTX-treated guinea pigs indicates that adenosine's effects aremediated by a receptor coupled to the inhibitory Giprotein. Consistent with an A1-adenosine receptor-effector coupling mechanism that includes Gi, the selec-tive A1-adenosine receptor agonist CPA significantlyreduced accumulation of cellular cAMP caused byisoproterenol. This effect of CPA on cAMP, like theelectrophysiological effects of adenosine, was com-pletely reversed by the adenosine antagonist CPT.
Thus, similar to other previously described cardiacelectrophysiological effects of adenosine,22 the effect ofadenosine on 'Ti, afterdepolarizations, and triggered
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from

752 Circulation Research Vol 70, No 4 April 1992
activity induced by isoproterenol and forskolin is medi-ated by A1-adenosine receptors coupled to Gi. However,unlike the activation of the potassium outward current,IKAdo, in supraventricular tissues, which is also mediatedby A1-adenosine receptors coupled to Gi but is indepen-dent of inhibition of adenylate cyclase,22 the effect ofadenosine on 1Ti, afterdepolarizations, and triggeredactivity in ventricular myocytes is dependent on inhibi-tion of accumulation of the second messenger cAMP.
Role of the intracellular P site. Like the extracellularA,-adenosine receptor, the intracellular P site, which is adistinct binding domain located in the catalytic subunit ofadenylate cyclase, inhibits the stimulated form of theenzyme and lowers cAMP levels.15 In isolated myocardialmembranes prepared from rat hearts, LaMonica et al13found that 10-5 M of the P site agonist DDA caused a 19%inhibition of adenylate cyclase activity previously stimu-lated with 10 6 M isoproterenol. The observation that thiseffect of DDA on adenylate cyclase was not antagonizedby either isobutylmethylxanthine or 8-phenyltheophyllinewas interpreted to indicate that the P site exists in myo-cardial membranes.13 However, the importance of this Psite in modulation of functions dependent on adenylatecyclase activity in intact cardiac cells has not yet beenadequately established. That is, in heart muscle, thephysiological role of the P site remains to be demon-strated. In the present study, DDA, a P site agonist, addedto either the superfusion medium or the internal recordingpipette solution (i.e., intracellularly) failed to inhibit iso-proterenol-induced DADs, 'Ti, and triggered activity (Fig-ures 7 and 8). Similarly, DDA did not attenuate theprolongation of the ventricular action potential caused byisoproterenol. Furthermore, the finding that both adeno-sine antagonists, that is, the A,-adenosine selectiveN-0861 and nonselective CPT, completely reversed theeffects of adenosine indicates that the observed actions ofadenosine were entirely mediated by the extracellularA,-adenosine receptor and not the intracellular P site.
Clinical ImplicationsThe results of the present study clearly establish that
in ventricular myocytes adenosine is highly specific forDADs, EADs, and resulting triggered activity caused byagents known to stimulate adenylate cyclase and in-crease cellular cAMP. These results are consistent withclinical findings that show that adenosine is effective interminating only exercise-induced ventricular tachycar-dia in patients without structural heart disease.8,30Adenosine has no effect on ventricular tachycardiacaused by reentry (associated with previous myocardialinfarction) whether or not catecholamine facilitated.8 Inconclusion, the response of triggered activity to adeno-sine may identify a mechanism of afterdepolarizationsrelated to stimulation of adenylate cyclase.
Note added in proof While this manuscript was inpress, the recent report of Fenton et a131 came to ourattention. These investigators presented evidence in sup-port of the idea that in rat ventricular myocytes theanti-f3-adrenergic action of adenosine is in part due toattenuation of isoproterenol-induced changes in intracel-lular [Ca2+]. Furthermore, isoproterenol-induced sponta-neous high frequency Ca2+ transients were observed afterantagonism of R-PIA, an A1-adenosine receptor agonist,
with a receptor antagonist. These results are consistentwith our hypothesis that the suppression of isoprotere-nol- and forskolin-induced 'Ti and triggered activity byadenosine is due to a reduction in cellular calcium and adecrease in the spontaneous oscillatory release of cal-cium from the sarcoplasmic reticulum.
AcknowledgmentsWe thank Dr. John Shryock for helpful discussion of this
project, Dr. Sheng-Nan Wu for electrophysiological study ofdideoxyadenosine, and Mrs. Kathleen Duvall for excellentsecretarial assistance.
References1. Zipes DP: Cardiac electrophysiology: Promises and contributions.JAm Coll Cardiol 1989;13:1329-1352
2. Wit AL, Rosen MR: Afterdepolarizations and triggered activity, inFozzard HA, Haber E, Jennings RB, Kate AM, Morgan HE (eds):The Heart and Circulatory System. New York, Raven Press, Pub-lishers, 1986, pp 1449-1490
3. January CT, Riddle JM: Early afterdepolarizations: Mechanism ofinduction and block: A role for L-tvpe Ca2 current. Circ Res1989;64:977-990
4. January CT, Shorofsky S: Early afterdepolarizations: Newerinsights into cellular mechanisms. J Cardiovasc Electrophysiol 1990;1:161-169
5. Lederer WJ, Tsien RW: Transient inward current underlyingarrhythmogenic effects of cardiotonic steroids in Purkinje fibers.JPhysiol (Lond) 1976;263:73-100
6. Matsuda H, Noma A, Kurachi Y, Irisawa H: Transient depolar-ization and spontaneous voltage fluctuations in isolated single cellsfrom guinea pig ventricles. Circ Res 1982;51:142-151
7. Kort AA, Lakatta EG: Calcium-dependent mechanical oscillationsoccur spontaneously in unstimulated mammalian cardiac cells. CircRes 1984;54:396-404
8. Lerman BB, Belardinelli L, West GA, Berne RM, DiMarco JP:Adenosine-sensitive ventricular tachycardia: Evidence suggestingcyclic AMP-mediated triggered activity. Circulation 1986;74:270-280
9. Belardinelli L, Isenberg G: Actions of adenosine and isoproterenol onisolated mammalian ventricular myocytes. Circ Res 1983;53:287-297
10. West GA, Isenberg G, Belardinelli L: Antagonism of forskolineffects by adenosine in isolated hearts and ventricular myocytes.Am J Physiol 1986;250:H769-H777
11. Isenberg G, Belardinelli L: Ionic basis for the antagonism betweenadenosine and isoproterenol in isolated mammalian ventricularmyocytes. Circ Res 1984;55:309-325
12. Schrader J, Baumann G, Gerlach E: Adenosine as inhibitor ofmyocardial effects of catecholamines. Pflugers Arch 1977;372:29-35
13. LaMonica DA, Frohloff N, Dobson JG: Adenosine inhibition ofcatecholamine-stimulated cardiac membrane adenylate cyclase.Am J Physiol 1985;248:H737-H744
14. Linden J, Hollen CE, Patel A: The mechanism by which adenosineand cholinergic agents reduce contractility in rat myocardium. CircRes 1985;56:728-735
15. Johnson RA, Yeung S-M, Bushfield B, Stubner D, Shoshani 1:Physiological and biochemical aspects of 'P"-site-mediated inhibi-tion of adenylyl cyclase, in Jacobson KA, Daly JW, Managaniello V(eds): Purines in Cellular Signaling: Targets for New Drugs. NewYork, Springer-Verlay New York, Inc, 1990, pp 158-165
16. Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ: Improvedpatch-clamp techniques for high-resolution current recording fromcells and cell-free membrane patches. Pflugers Arch 1981;391:85-100
17. Brooker GW, Terasaki L, Price MG: Gammaflo: A completelyautomated radioimmunoassay system. Science 1976;194:270-276
18. Kurachi Y, Nakajima T, Sugimoto T: On the mechanism ofactivation of muscarinic K' channels by adenosine in isolated atrialcells: Involvement of GTP-binding proteins. Pflugers Arch 1986;407:264-274
19. Isenberg G, Cerbai E, Lockner U: Ionic channels and adenosine inisolated heart cells, in Becker B, Gerlach E (eds): Topics andPerspectives ofAdenosine Research. Berlin, Springer-Verlag, 1987,pp 324-334
20. Kass RS, Tsien RW: Fluctuations in membrane current driven byintracellular calcium in cardiac Purkinje fibers. Biophys J 1982;38:259-269
21. Eisner DA, Lederer WJ: Na-Ca exchange: Stoichiometry andelectrogenicity. Am JPhysiol 1985;248:C189-C202
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from

Song et al Effect of Adenosine on Afterdepolarizations 753
22. Belardinelli L, Linden J, Berne RM: The cardiac effects ofadenosine. Prog Cardiovasc Dis 1989;32:73-97
23. Fedida D, Noble D, Rankin AC, Spindler AJ: The arrhythmogenictransient inward current 'Ti and related contraction in isolated guinea-pig ventricular myocytes. J Physiol (Lond) 1987;392:523-542
24. Matsuura H, Shattock MJ: Membrane potential fluctuations andtransient inward currents induced by reactive oxygen intermedi-ates in isolated rabbit ventricular cells. Circ Res 1991;68:319-329
25. Vassalle M, Lee CO: The relationship among intracellular sodiumactivity, calcium, and strophanthidin inotropy in canine cardiacPurkinje fibers. J Gen Physiol 1984;83:287-307
26. Londos C, Wolff J: Two distinct adenosine-sensitive sites onadenylate cyclase. Proc Natl Acad Sci U S A 1977;74:5482-5486
27. Dennis D, Boykin M, Jacobson KA, Belardinelli L: A novelirreversible adenosine antagonist attenuates the negative dromo-
tropic effect of adenosine in guinea pig heart (abstract). FASEB J1990;4:A1008
28. Bruns RF, Lu GF, Pugsley TA: Adenosine receptor subtypes:Binding studies, in Becker B, Gerlach E (eds): Topics and Perspec-tives of Adenosine Research. Berlin, Springer-Verlag, 1987, pp59-73
29. Shryock JC, Travagli H, Belardinelli L: Evaluation of N-0861,(±+)N-endonorbornan-2-yl-9-methyladenine, as an A1 subtype-selective adenosine receptor antagonist in the guinea pig heart.J Pharmacol Exp Ther (in press)
30. Griffith MJ, Ward DE, Linker NJ, Camm AJ: Adenosine in thediagnosis of broad complex tachycardia. Lancet 1988;1:672-675
31. Fenton RAE, Moore EDW, Fay FS, Dobson JG Jr: Adenosinereduces the Ca2' transients of isoproterenol-stimulated rat ventric-ular myocytes. Am J Physiol 1991;261:C1107-C1114
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from

Y Song, S Thedford, B B Lerman and L Belardinellimyocytes.
Adenosine-sensitive afterdepolarizations and triggered activity in guinea pig ventricular
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1992 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.70.4.7431992;70:743-753Circ Res.
http://circres.ahajournals.org/content/70/4/743World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 4, 2018http://circres.ahajournals.org/
Dow
nloaded from





























